
Submitted:
27 March 2025
Posted:
28 March 2025
You are already at the latest version
Abstract
Improving wheat drought stress tolerance is a critical and challenging task, and more research is necessary since many parts of the world depend on this crop for food and feed. Our current work is focused on the influence of probiotic microorganisms in combination with calcium salts on the physiological and biochemical metabolic pathways that wheat uses when exposed to drought stress and on the analysis of gene expression levels that contribute to wheat drought tolerance. The research was conducted in the laboratory under controlled conditions, simulating a prolonged drought. Seedlings were treated with different microorganisms (Bacillus subtilis, Lactobacillus paracasei, and some yeast) in 105 CFU/ml concentrations for seed priming and later in the same concentration for seedling spraying. 70 g/m2 CaCO3 or 100 g/m2 CaCl2 was added to the soil before the sowing seeds. Almost all tested treatments improved plant growth and positively affected prolonged drought resistance in winter wheat. Bacillus subtilis, in combination with calcium salts, had the greatest effect on maintaining the relative leaf water content (RWC). Proline, MDA, and H2O2 tests proved a significant positive impact of the treatments on the plant's response at the biochemical level and growth parameters close to those of irrigated plants, along with lower levels of drought-induced gene expression. This study showed that tested microorganisms in combination with calcium salts can activate plants' defense reactions in response to drought. The practical significance of this study is that these ecological measures can be useful under field conditions.
Keywords:
drought stress
; Triticum aestivum
; PGPM
; Bacillus subtilis
; calcium salts
1. Introduction
Drought is the most critical abiotic limiting factor for plant cultivation in many parts of the world [1,2,3,4,5]. Drought affects plants' morphological, physiological, biochemical, and molecular processes, resulting in growth inhibition [6,7,8]. A bioinformatic analysis of European crop losses over recent decades revealed that drought waves are responsible for greater yield loss in cereals (9% and 7.3%) compared to non-cereal crops (3.8% and 3.1%) [9]. Many studies have explored the genetic and molecular mechanisms behind drought resistance in cereals such as wheat, barley, maize, sorghum, and rice [10,11].
Wheat is the leading food for over a third of the world’s population [12]. Wheat production in recent years (2018-2025) is lower than it used to be in the largest producing countries such as Canada, Iran, France, Germany, and Greece due to drought [13,14,15]. According to the European Drought Observatory (EDO), a drought warning was in effect for nearly half of the EU territory in 2022, with a red alert for 15%. By mid-January 2025, the Combined Drought Indicator (CDI) showed warning drought conditions in southern Italy, the eastern Baltic Sea region, eastern Poland, Belarus, central-eastern Ukraine, Greece, part of the Balkans, Cyprus, Malta, and other Mediterranean islands, as well as Ireland, northern UK, and more than a half of Türkiye [16]. Therefore, developing new pathways to help plants overcome drought stress is essential.
Nutrient management and organic amendments can reduce the harmful effects of drought stress on crops [17]. As an essential plant nutrient, calcium plays a vital role in plant growth and development. Drought stress is known to alter plant mineral status and metabolic processes. Calcium (here and later Ca) is involved in plant signaling responses to drought [18] and thereby positively affects plant adaptation to drought stress [19]. Ca2+ can help plants adapt to drought quickly by adjusting their stomatal opening/closing, optimizing their gas exchange, and improving their photosynthetic efficiency [20]. Probiotic microorganisms increase plants' uptake of essential elements from the soil. For example, microbial biostimulants enhance plant growth under stress conditions by facilitating the uptake of insoluble elements and essential minerals [21]. Probiotic bacteria are also thought to induce drought tolerance in plants by altering root morphology, activating the antioxidant system, and promoting the expression of genes associated with abiotic stress tolerance [22]. Plant growth-promoting microorganisms (PGPMs) use different mechanisms that directly or indirectly help in crop plants' growth and development, increasing crop productivity [23]. Bacillus subtilis is one of the most attractive probiotics for developing natural plant protection products because it is generally considered safe for food use [24]. B. subtilis strains produce cytokinin that interferes with drought-induced suppression of shoot growth, thereby increasing plant growth during periods of drought [25].
Several drought tolerance indicators have been identified as reliable drought resilience measures, including relative water content (RWC), water use efficiency, stomatal conductance, photosynthesis, and drought-responsive genes. The RWC is considered a measure of plants' water status and the dehydration index. It allows you to assess plants' physiological states and shows their ability to retain water in unfavorable conditions [26,27]. Malondialdehyde (MDA) is commonly used to assess the status of redox and osmotic adaptation, which is important for plant adaptation to environmental stresses. MDA is a good marker of oxidative stress because ROS breaks down polyunsaturated lipids and forms MDA. The overall increase in membrane lipid peroxidation is proportional to the intensity of drought stress and may result from spontaneous reactions of ROS with organic molecules contained in membranes [28]. Among ROS species, hydrogen peroxide (H2O2) is an especially important one involved in plant responses to different environmental stressors. During stress, peroxide concentration increases because it works as a signaling molecule. However, increasing H2O2 concentration over time can harm plants (oxidative stress) [29]. In stressful conditions, plants accumulate metabolites, especially some amino acids such as proline. Proline plays the role of the osmolyte for plants; during drought stress, it can take the role of a signaling molecule, a metal chelator, and an antioxidant defense molecule. It is often used as a stress marker [30,31]. Plants respond to stressful environments by altering gene expression and protein production [6]. It is known that some genes that code late embryonic protein (LEA) are affected by drought [32,33]. LEA proteins are generally predicted to maintain intrinsically disordered proteins in a fully hydrated state, which can then fold under water-deficient conditions to adopt α-helical structures. Ordered LEAs can act as molecular chaperones to bind enzymes, membranes, water or ions, and reactive oxygen species (ROS) and, therefore, play a critical role in protein/membrane stabilization and maintaining cellular environmental homeostasis under drought stress [33].
The experiments were modeled and conducted with the microorganisms – Bacillus subtilis, Lactobacillus paracasei, Zygosaccharomyces bailii, Geotrichum silvicola. This study aimed to investigate how the effects of different microorganisms and Ca could modify the drought tolerance of wheat at the molecular, biochemical, and morphological levels. Further work focused on determining which of the selected microorganisms help the plant overcome drought stress, especially in the presence of added Ca in the soil. The objectives of this study were to (1) explore whether chosen microorganisms and Ca salts improve shoot growth under drought stress, (2) to study the changes in biochemical and molecular responses of treated winter wheat to drought, (3) to investigate which microorganisms work better in combination with Ca.
2. Materials and Methods
2.1. Plant Material, Growth Conditions, and Treatments
Wheat seeds (Triticum aestivum L. cv. ‘Skagen’) were sown in plastic cylindric pots (10 × 13 cm), 20 seeds per pot, in a peat moss substrate (pH 5.5 – 6.5). Plants were germinated and grown under controlled conditions of a constant temperature of 22 ± 1 °C, a photoperiod of 16/8 h, and a fluorescent light photon flux of 60 µmol m−2 s−1 at soil level (Figure 1). Soil moisture was maintained at 60 - 70%.
The following treatments were used for the drought stress control studies:
- a)
- Ca was added to the soil in the form of CaCO3 at a rate of 1.3 g per pot, based on 70 g m−2, and in the form of CaCl2 prediluted in water at a rate of 1 g in 100 ml per pot.
- b)
- The following microorganisms - Bacillus subtilis, Lactobacillus paracasei, Zygosaccharomyces bailii, Geotrichum silvicola - were used for seed priming in 105 CFU/ml concentrations (microorganisms were grown in their specific liquid media (Nutrient, MRS, YPD) to the log phase and then diluted with distilled water to obtain the required concentration) and later in the same concentration for seedlings spraying in the 1–2 leaf stage (BBCH-scale 1–2) [34]
2.2. Experimental Design and Drought Conditions
Treatments were added according to the scheme (Table 1). Each treatment was carried out in 6 pots, divided into two groups: 3 for simulating drought and 3 for rational watering. Drought was initiated immediately after plants were sprayed with microorganisms at the 1-2 leaf stage as irrigation termination. As a result, the soil gradually dried, reaching 15 - 20 % humidity. Soil humidity of watered plants was kept at a 60 - 70% level. Soil moisture was measured using a soil moisture meter (Aicevoos, AS-PH3, China).
2.3. Sampling
Plant samples were taken for analysis when soil moisture in the drought treatments was 15 - 20%. The watered plants were sampled simultaneously (soil moisture 60 - 70%). Shoots of wheat seedlings were sampled for morphometrical measurements: 10 plants from each pot. For biochemical and molecular analysis, three independent replicates were carried out using the third leaves of wheat plants. For MDA, H2O2, proline assays, and RNA isolation, the samples were immediately frozen in liquid nitrogen and stored in a low-temperature freezer (Skadi Green line, International Labmate Ltd., St Albans, UK) at −80 °C until the analysis.
2.4. Morphometrical Measurements
Shoot length and biomass were measured using a ruler and balances (Kern EWJ).
2.5. RWC
RWC was determined according to Weng et al. [35]. Fresh wheat leaves were collected and weighed as fresh weight (FW). After that, the leaves were left in the water for 24 hours and weighed again to obtain a saturated weight (SW). The dry weight (DW) was obtained by drying the leaves in a drying chamber and weighted. RWC has been calculated according to the formula [35]:
RWC = [(FW − DW) / (SW − DW)] × 100%,
2.6. Assessment of Biochemical Parameters
2.6.1. H2O2
For analysis of H2O2 and MDA, leaf material (0.5 g) was homogenized using 5% trichloracetic acid (TCA) (Sigma-Aldrich, St. Louis, MO, USA). The homogenates were centrifuged at 10000 × g for 15 min (centrifuge MPW-351 R). H2O2 content in leaves was determined according to Velikova et al. [36]. The supernatant was mixed with 10 mM, pH 7.0 potassium phosphate buffer (Alfa Aesar) and 1 M potassium iodide (Alfa Aesar) in a ratio of 1:1:2. The reaction solution was incubated for 30 min at 25° C in the dark. The absorbance of the supernatant was measured at 390 nm by a spectrophotometer (Analytik Jena Specord 210 Plus, Analytik Jena, Jena, Germany). The amount of H2O2 was calculated using a standard curve. The results are expressed in µmol g−1 FW.
2.6.2. Lipid Peroxidation According to MDA
The method of Hodges et al. [37], with slight modifications, has been used to estimate MDA. The supernatant was added to 20% TCA containing 0.5% thiobarbituric acid (TBA) (Alfa Aesar, Haverhill, MA, USA). The homogenate was incubated in a heater (Blockthermostat BT 200) at 95 °C for 30 minutes and then cooled on the ice. The optical density was measured at 532 and 660 nm by a spectrophotometer (Analytik Jena Specord 210 Plus, Analytik Jena, Jena, Germany). The results were expressed in µmol g−1 FW [37].
2.6.3. Proline
Carillo & Gibon’s proline extraction and determination method was used to determine the free proline content [38]. Leaf material (0.5g) was homogenized using 40% ethanol and left overnight at +4 °C. The homogenates were centrifuged at 14000 × g for 5 min (centrifuge MPW-351 R), and the supernatant was added to the reaction mix (1% ninhydrin, 60% acetic acid, 20% ethanol). The homogenate was incubated in a heater (Blockthermostat BT 200) at 95 °C for 20 min. The optical density was measured at 520 nm by a spectrophotometer (Analytik Jena Specord 210 Plus, Analytik Jena, Jena, Germany). The corresponding content of the proline was determined using the standard curve. The calculations were made using the equation given in the method [38]. The results were expressed in µmol g−1 FW.
2.7. Molecular Techniques
2.7.1. RNA Extraction and Reverse Transcription
Total RNA was extracted from 200 mg of plant leaf material using the PureLink RNA Mini Kit (Ambion, Waltham, MA, USA) following the manufacturer's recommendations and using Heraeus Fresco 21 Centrifuge (Thermo Scientific, Waltham, MA, USA). To avoid contamination with genomic DNA, extracted total RNA was treated with RapidOut DNA Removal Kit (Thermo Scientific). The concentration and purity of treated RNA were evaluated with the spectrophotometer NanoPhotometer P330 (IMPLEN, Westlake Village, CA, USA). DNase-treated RNA samples were reverse-transcribed using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Waltham, MA, USA) following the manufacturer's recommendations. The obtained cDNA was stored at −20 °C.
2.7.2. Real-Time Quantitative PCR
Real-time quantitative PCR was carried out using SYBR® Green Universal Master Mix kit (Applied Biosystems) following the manufacturer’s recommendations and employing QuantStudioTM 5 real-time PCR system (Applied Biosystems). Two microliters of cDNA (the equivalent of 25 ng of total RNA) were used as a template for PCR. Cycling conditions comprised one cycle at 95 °C for 2 min and 40 cycles at 95 °C for 15 s followed by 60 °C for 1 min. After the PCR run, a melting curve was generated and analyzed each time with the Quant Studio Design & Analysis Software v.1.5.2 (Applied Biosystems). Gene expression was calculated using the 2−ΔΔCt method [39].
2.7.3. Primers
The sequences of primers used in the work were based on the sequences of LEA protein genes and were taken from the publication of Ali-Benali et al. [40]. The wheat mitochondrial 26S ribosomal RNA gene was used as a housekeeping gene [40]. The primer concentrations in all cases were 200 nM except for the Td27e, where the concentration was 900 nM. The sequences of all primers used in the work are listed in Table 2.
2.8. Statistical Analysis
The results are presented as mean ± standard deviation (SD) of three independent experiments with at least three replicates. The data were analyzed using a two-way analysis of variance with replication (two-way ANOVA with replication). The significant differences between treatment means were determined using analysis of variance and mean separation at a 5% significance level (p ˂ 0.05).
3. Results
3.1. Impact of PGPMs and Ca Salts on Morphometric Parameters of Wheat Seedlings Exposed to Prolonged Drought Stress
The shoot length and biomass of wheat were significantly higher when the plants were treated with probiotic microorganisms and grown in soil supplemented with Ca salts, especially in the ones with the combination of B. subtilis and CaCO3. This was seen in both drought-stressed and normally watered plants (Table 3).
3.2. Effect of Used PGPMs and Ca Salts on RWC of Wheat Seedlings Exposed to Prolonged Drought
Wheat plants treated with PGPMs and Ca salts retained higher water content in their leaves after exposure to prolonged drought (Figure 2). The ones with a combination of B. subtilis and Ca salts had the best retention of leaf water content at ~ 80% (moderate stress). In comparison, the RWC of Control wheat leaves was 47% (high-stress level).
3.3. Effect of PGPMs and Ca Salts on Biochemical Responses of Wheat Plants Exposed to Prolonged Drought
3.3.1. Hydrogen Peroxide (H2O2)
The levels of H2O2 increased in drought-affected plants (Figure 3). However, H2O2 content was found to be significantly lower in all variants treated with PGPMs and Ca salts in combinations. The ones treated with B. subtilis alone or with Ca salts had the lowest H2O2 content, 0.86 - 0.96 µmol g−1 FW, compared to 3.85 µmol g−1 FW for the Control. The difference is also significant when comparing plants only treated with Bacillus subtilis, Lactobacillus paracasei, or with Ca salts.
3.3.2. Malondialdehyde (MDA)
MDA levels that indicate the lipid peroxidation levels showed that although drought stress significantly increased MDA content in wheat leaves, the amount of MDA was considerably lower in treated plants, especially in plants with a combination of B. subtilis and Ca salts (18 - 22 μmol g−1 FW) compared to the Control plants (72.4 μmol g−1 FW). The difference is also significant when comparing plants only treated with B. subtilis (31.013 μmol/ g−1 FW) or with Ca salts (47 - 49,4 μmol g−1 FW). Combinations of Lactobacillus paracasei and CaCl2 (28.47 μmol g−1 FW) and Zygosaccharomyces bailii and CaCl2 (21.96 μmol g−1 FW) were also very effective in reducing MDA levels (Figure 4).
3.3.3. Free Proline
The amount of free endogenous proline in Control plants exposed to drought stress was 8.88 µmol g−1 FW. In contrast, in plants treated with PGPMs and Ca salts, it ranged from 1.97 to 6.1 µmol g−1 FW (except Lactobacillus paracasei + CaCO3, which was closer to control plants), the lowest in the samples treated with the combinations of Bacillus subtilis and CaCl2 and Zygoaccharomyces bailii and CaCl2 (Figure 5).
3.4. Effect of PGPMs and Ca Salts Application on Late Embryogenesis Abundant (lea) Genes Expression Levels of Wheat Plant Exposed to Prolonged Drought
Quantitative results of RT-qPCR analysis showed that among the three studied genes, the Td27e gene was more highly expressed in drought stress. The level of Td27e expression in the Control sample was significantly higher than in the all treated ones (Figure 6a). Td29b gene expression levels were markedly lower in all treated variants compared with Control (Figure 6b). The lowest levels of expression of the Td11 gene were recorded in the samples of plants treated with the combinations of Bacillus subtilis and Ca salts and Geotrichum silvicola and Ca salts (Figure 6c).
4. Discussion
According to the Population Division of the United Nations, the world human population will reach about 9.5 billion by 2050. Agriculture is the primary source of food production and must solve this problem by producing enough food for the global population [41].
This work is a logical continuation of the previous work [42]. Since a positive reaction of plants under drought conditions to the combination of commercial plant probiotics and CaCO3 was shown, it was important to understand which PGPMs were at work here and what role Ca played. Therefore, experiments were modeled and conducted with the ingredient microorganisms (or related to them): Bacillus subtilis, Lactobacillus paracasei, Zygosaccharomyces bailii, Geotrichum silvicola. In addition to CaCO3, CaCl2 was added to the experimental model as a more easily absorbed form of Ca to compare which works better in combination with microorganisms. A series of experiments were performed to explore how the application of probiotic microorganisms and Ca affects the response of wheat to drought stress at physiological, biochemical, and molecular levels. A test experiment was designed and performed to exclude the possibility that minerals, sugars, or other components of microbial media have a more significant positive effect on plants than microorganisms. The results showed that microorganisms are essential for obtaining the previously observed positive impact. Further work focused on determining which of the selected microorganisms help the plant overcome drought stress, especially in the presence of added Ca in the soil.
Much research has been done to understand how plants respond to drought and how to optimize that response [28,29,31,43]. Thanks to previous research, we were able to select some of the most significant markers that show the level of stress in plants. For example, the RWC results showed that our treatments stimulated plants' ability to retain water and subsequently maintain their physiological state close to that of watered control plants. Proline, MDA, and H2O2 tests, in turn, proved a significant positive impact of the treatments on the plant's response at the biochemical level. Some of the PGPMs used with Ca salts, maintained concentrations of these stress indicators at almost half those of untreated controls. The MDA concentration difference is significant when comparing plants only treated with B. subtilis or in combination with Ca salts, which indicates that exactly the combination of them is the most beneficial for the plant. The same is visible with H2O2 test results: when comparing plants only treated with Bacillus subtilis, Lactobacillus paracasei, or in combination with Ca salts and for proline test with the combinations of Bacillus subtilis and CaCl2 and Zygoaccharomyces bailii and CaCl2. The three genes that we used for our studies have a major role in wheat response to drought stress in the early stage of growth. Td27e and Td11 code for group 2 LEA proteins, also named dehydrins. Dehydrins are produced by plants in response to cold and drought stress and are well-known as stress proteins [40,44]. Td27e gene encode YSK2-type dehydrin, Td11 gene a SK3-type dehydrin. Td29b gene codes for a group 4 LEA protein and plays an important role in the early stages of seed development by protecting cells against damage caused by desiccation [40]. In this study, these genes were used as stress indicators to see the level of changes at the molecular level in plants and the effects of treatments at that level. As a result, their expression levels proved to be significantly less when plants were treated with chosen PGPMs and Ca salts.
Several studies have been conducted on using plant growth-promoting microorganisms under abiotic stress, especially from the Bacillus genus, and they have shown great potential for organic farming [5,45,46]. Previous research has already shown that B. subtilis plays an important role in wheat's response to adverse environmental factors [47]. Based on our experiments, we can confirm that B. subtilis helped winter wheat plants maintain growth even under prolonged drought conditions and maintained their biochemical and molecular homeostasis close to that of plants watered rationally. Moreover, we see that in combination with Ca salts, the positive effect of B. subtilis increases significantly.
In contrast to the Bacillus genus, several studies have been conducted in animal models that have linked the genus Lactobacillus and the efficiency of Ca absorption in their presence [48,49,50]. An interesting research connected the use of Ca supplements with L. paracasei colonization in the gut [51]. These studies and our results allow us to hypothesize that using Lactobacillus species in combination with Ca may also positively affect plants, but further research is necessary to clarify the essence of this combination.
Some other studies have linked yeast (Zygosaccharomyces genus) use, to improve yield growth under abiotic stress conditions [52,53]. Our results showed great potential for using Zygosaccharomyces bailli in combination with Ca salts to alleviate the impact of drought on winter wheat plants. Geotrichum silvicola, which isn’t a well-investigated yeast yet, also showed this potential.
The results of our research allow us to propose several hypotheses of the connection of the use of PGPMs in combination with Ca for further investigation of possible pathways related to their effect on plants during drought. The current winter wheat study data showed that using these treatments can activate the defense responses of plants that can compensate for the negative impact of drought. This again proves the results of previous research [54,55] that plant growth-promoting microorganisms have big potential to be used in agriculture.
5. Conclusions
All treatments have a generally positive effect in drought conditions. Exogenous application of our PGPMs and Ca salts improved the prolonged drought tolerance of winter wheat. B. subtilis, in combination with Ca salts, had the most significant impact on maintaining the relative water content of the leaves and keeping plant growth parameters close to those of irrigated plants. This treatment also showed the best results for maintaining plants' biochemical and molecular homeostasis. The results of this study in winter wheat showed that using probiotic microorganisms in combination with Ca salts can activate defense reactions that can compensate for the harmful effects of drought stress. PGPMs and Ca salts used in this study are suitable for organic farming since they won't harm the environment.
Author Contributions
Conceptualization, M.Z. and V.Š.; methodology, S.J., A.P., R.M. and V.Š.; formal analysis, V.Š. and M.Z.; investigation, M.Z.; data curation, M.Z. and V.Š.; writing—original draft preparation M.Z.; writing—review and editing, S.J., V.G., R.M; visualization, V.Š. and M.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Nature Research Centre R & D II program.
Data Availability Statement
The data supporting the reported results can be found in the Nature Research Centre's archive of scientific reports.
Acknowledgments
We thank the staff of the Laboratory of Plant Physiology of the Nature Research Centre for their support and assistance and the Laboratory of Biodeterioration Research for providing the necessary microorganisms.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
References
- Fathi, A.; Tari, D.B. Effect of drought stress and its mechanism in plants. Int. J. Life Sci. 2016, 10, 1–6. [Google Scholar]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; Ihsan, M.Z.; Alharby, H.; Wu, C.; Wang, D.; Huang, J. Crop production under drought and heat stress: plant responses and management options. Front Plant Sci. 2017, 8, 1147. [Google Scholar]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res. 2021, 242, 126626. [Google Scholar] [CrossRef]
- Todorova, D.; Sergiev, I.; Katerova, Z.; Shopova, E.; Dimitrova, L.; Brankova, L. Assessment of the biochemical responses of wheat seedlings to soil drought after application of selective herbicide. Plants 2021, 10, 733. [Google Scholar] [CrossRef]
- Chattaraj, S.; Samantaray, A.; Ganguly, A.; Thatoi, H. Employing plant growth-promoting rhizobacteria for abiotic stress mitigation in plants: with a focus on drought stress. Discov. Appl. Sci. 2025, 7, 68. [Google Scholar]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 12, 1–4. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants (Basel) 2021, 10, 259. [Google Scholar]
- Oguz, M.C.; Aycan, M.; Oguz, E.; Poyraz, I.; Yildiz, M. Drought stress tolerance in plants: interplay of molecular, biochemical and physiological responses in important development stages. Physiologia 2022, 2, 180–197. [Google Scholar] [CrossRef]
- Brás, T.A.; Seixas, J.; Carvalhais, N.; Jägermeyr, J. Severity of drought and heatwave crop losses tripled over the last five decades in Europe. Environ. Res. Lett. 2021, 16, 065012. [Google Scholar]
- Geng, L.; Li, M.; Zhang, G.; Ye, L. Barley: a potential cereal for producing healthy and functional foods. Food Qual. Saf. 2022, 6, fyac012. [Google Scholar]
- Batîr Rusu, D.C.; Murariu, D.; Gheorghita, R.; Graur, M. Some nutritional value aspects of barley and oat and their impact in human nutrition and healthy life. Plants 2024, 13, 2764. [Google Scholar] [CrossRef] [PubMed]
- Sultan, M.A.R.F.; Hui, L.; Yang, L.Y.; Xian, Z.H. Assessment of drought tolerance of some Triticum L. species through physiological indices. Czech J. Genet. Plant Breed. 2012, 48, 178–184. [Google Scholar]
- US Wheat Associates 2020. Available online: https://www.uswheat.org/wheatletter/2020-global-wheat-harvest-update/ (accessed on 12 January 2025).
- USDA1. World Agricultural Supply and Demand Estimates 2021. Available online: World Agricultural Supply and Demand Estimates (WASDE) | USDA Foreign Agricultural Service (accessed on 20 January 2025).
- Eurostat. Available online: https://ec.europa.eu/eurostat/web/agriculture/publications (accessed on 15 February 2025).
- European Drought Observatory. Available online: https://joint-research-centre.ec.europa.eu/european-and-global-drought-observatories/current-drought-situation-europe_en (accessed on 15 February 2025).
- Farooq, S.; Ul-Allah, S.; Hussain, M. Drought sress in crop plants and its management. In Disaster Risk Reduction in Agriculture; Ahmed, M., Ahmad, S., Eds.; Publisher Springer: Singapore, 2023; pp. 21–42. [Google Scholar]
- Cousson, A. Involvement of phospholipase C-independent calcium-mediated abscisic acid signaling during Arabidopsis response to drought. Biol. Plant 2009, 53, 53–62. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Réthoré, E.; Pluchon, S.; Ali, N.; Billiot, B.; Yvin, J.C. Calcium application enhances drought stress tolerance in sugar beet and promotes plant biomass and beetroot sucrose concentration. Int. J. Mol. Sci. 2019, 20, 3777. [Google Scholar] [CrossRef]
- Feng, D.; Wang, X.; Gao, J.; Zhang, C.; Liu, H.; Liu, P.; Sun, X. Exogenous calcium: its mechanisms and research advances involved in plant stress tolerance. Front. Plant Sci. 2023, 14, 1143963. [Google Scholar]
- Glick, B.R. Plant growth-promoting bacteria: mechanisms and applications. Scientifica (Cairo), 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Saberi Riseh, R.; Fathi, F.; Moradzadeh-Eskandari, M. The effect of some probiotic bacteria in induction of drought tolerance in cucumber plants. Microbiol. Metabolites Biotechnol. 2018, 2, 113–127. [Google Scholar]
- Kumar, M.; Shamshad Ahmad, P.; Singh, R.P. Plant growth promoting microbes: diverse roles for sustainable and ecofriendly agriculture. Energy Nexus, 2022, 7, 100133. [Google Scholar]
- Lubyanova, A.R.; Allagulova, C.R.; Lastochkina, O.V. The effects of seed pretreatment with endophytic bacteria Bacillus subtilis on the water balance of spring and winter wheat seedlings under short-time water deficit. Plants 2023, 12, 2684. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Potential use of Bacillus spp. as an effective biostimulant against abiotic stresses in crops—A review. Curr. Res. Biotechnol. 2023, 5, 100128. [Google Scholar] [CrossRef]
- Soltys-Kalina, D.; Plich, J.; Strzelczyk-Żyta, D.; Śliwka, J.; Marczewski, W. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of 'Katahdin'-derived potato cultivars. Breed Sci. 2016, 66, 328–331. [Google Scholar] [PubMed]
- González-Espíndola, L.Á.; Pedroza-Sandoval, A.; Trejo-Calzada, R.; Jacobo-Salcedo, M.d.R.; García de los Santos, G.; Quezada-Rivera, J.J. Relative water content, chlorophyll index, and photosynthetic pigments on Lotus corniculatus L. in Response to Water Deficit. Plants 2024, 13, 961. [Google Scholar] [CrossRef]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar]
- Belkadhi, A.; De Haro, A.; Soengas, P.; Obregon, S.; Cartea, M.E.; Chaibi, W.; Djebali, W. Salicylic acid increases tolerance to oxidative stress induced by hydrogen peroxide accumulation in leaves of cadmium-exposed flax (Linum usitatissimum L.). J. Plant Interact. 2014, 9, 647–654. [Google Scholar]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: a review. Plant Signal Behav. 2012, 7, 1456–1466. [Google Scholar]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. 2013, 19 (9), 998–1011. Antioxid Redox Signal. 2013, 19, 998–1011. [Google Scholar]
- Pnueli, L.; Hallak-Herr, E.; Rozenberg, M.; Cohen, M.; Goloubinoff, P.; Kaplan, A.; Mittler, R. Molecular and biochemical mechanisms associated with dormancy and drought tolerance in the desert legume Retama raetam. Plant J. 2002, 31, 319–330. [Google Scholar]
- Li, Z.; Chi, H.; Liu, C.; Zhang, T.; Han, L.; Li, L.; Pei, X.; Long, Y. Genome-wide identification and functional characterization of LEA genes during seed development process in linseed flax (Linum usitatissimum L.). BMC Plant Biol. 2021, 21, 193. [Google Scholar]
- Weber, E.; Bleiholder, H.; Lancashire, P.D.; Langelüddecke, R.; Stauss, R.; Van der Boom, T.; Witzen-Berger, A. Growth stages of mono- and dicotyledonous plants. In BBCH Monograph., 1st ed.; Meier, U., Ed.; Julius Kühn-Institut: Quedlinburg, Germany, 2018; pp. 18–21. [Google Scholar]
- Weng, M.; Cui, L.; Liu, F.; Zhang, M.; Shan, L.; Yang, S.; Deng, X.-P. Effects of drought stress on antioxidant enzymes in seedlings of different wheat genotypes. Pak. J. Bot. 2015, 47, 49–56. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar]
- Hodges, D.; DeLong, J.; Forney, C.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar]
- Carillo, P.; Gibon, Y. Extraction and Determination of Proline. Available online: https://www.researchgate.net/publication/211353600_PROTOCOL_Extraction_and_determination_of_prolineproline (accessed on 10 July 2016).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time Quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]
- Ali-Benali, M.A.; Alary, R.; Joudrier, P.; Gautier, M.-F. Comparative expression of five LEA genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochim. Biophys. Acta, Gene Struct. Expression 2005, 1730, 56–65. [Google Scholar] [CrossRef]
- Menendez, E.; Garcia-Fraile, P. Plant probiotic bacteria: solutions to feed the world. AIMS Microbiol. 2017, 3, 502–524. [Google Scholar] [PubMed]
- Mockevičiūtė, R.; Jurkonienė, S.; Šveikauskas, V.; Zareyan, M.; Jankovska-Bortkevič, E.; Jankauskienė, J.; Kozeko, L.; Gavelienė, V. Probiotics, proline and calcium induced protective responses of Triticum aestivum under drought stress. Plants 2023, 12, 1301. [Google Scholar] [CrossRef]
- Dikilitas, M.; Şimşek, E.; Roychoudhury, A. Role of proline and glycine betaine in overcoming abiotic stresses. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress: Biochemical and Molecular Perspectives, 1st ed.; Roychoudhury, A., Tripathi, D.K., Eds.; John Wiley & Sons Ltd,, 2020; pp. 1–23. [Google Scholar]
- Yang, Y.; He, M.; Zhu, Z.; Li, S.; Xu, Y.; Zhang, C.; Singer, S.; Wang, Y. Identification of the dehydrin gene family from grapevine species and analysis of their responsiveness of various forms of abiotic and biotic stress. BMC Plant Biol. 2012, 12, 140. [Google Scholar]
- Lalarukh, I.; Al-Dhumri, S.A.; Al-Ani, L.K.T.; Hussain, R.; Al Mutairi, K.A.; Mansoora, N.; Amjad, S.F.; Abbas, M.H.H.; Abdelhafez, A.A.; Poczai, P.; Meena, K.R.; Galal, T.M. A Combined use of Rhizobacteria and Moringa leaf extract mitigates the adverse effects of drought stress in wheat (Triticum aestivum L.). Front Microbiol. 2022, 13, 813415. [Google Scholar] [CrossRef]
- Yadav, V.K.; Yadav, R.C.; Choudhary, P.; Sharma, S.K.; Bhagat, N. Mitigation of drought stress in wheat (Triticum aestivum L.) by inoculation of drought tolerant Bacillus paramycoides DT-85 and Bacillus paranthracis DT-97. J. Appl. Biol. Biotechnol. 2022, 10, 59–69. [Google Scholar]
- Xie, J.; Bai, L.; Zhou, W. Research progress of Bacillus subtilis in wheat growth promotion and disease control. Int. J. Biol. Life Sci. 2024, 7, 68–72. [Google Scholar] [CrossRef]
- Collins, F.L.; Rios-Arce, N.D.; Schepper, J.D.; Parameswaran, N.; McCabe, L.R. The potential of probiotics as a therapy for osteoporosis. Microbiol Spectr. 2017, 5, 10.1128. [Google Scholar] [CrossRef] [PubMed]
- Raveschot, C.; Coutte, F.; Frémont, M.; Vaeremans, M.; Dugersuren, J.; Demberel, S.; Drider, D.; Dhulster, P.; Flahaut, C.; Cudennec, B. Probiotic Lactobacillus strains from Mongolia improve calcium transport and uptake by intestinal cells in vitro. Food Res. Int. 2020, 133, 109201. [Google Scholar] [CrossRef]
- Harahap, I.A.; Suliburska, J. Probiotics and isoflavones as a promising therapeutic for calcium status and bone health: a narrative review. Foods 2021, 10, 2685. [Google Scholar] [CrossRef]
- Trautvetter, U.; Ditscheida, B.; Kiehntop, M.; Jahreis, G. A combination of calcium phosphate and probiotics beneficially influences intestinal lactobacilli and cholesterol metabolism in humans. Clin Nutr. 2012, 31, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Bilek, F.N.; Rezki, M.A.; Grondin, C.; Yahia, N.; Bekki, A. Plant growth promoting characteristics and stress tolerance of yeasts isolated from algerian agricultural soils. South Asian J. Exp. Biol. 2020, 10, 413–426. [Google Scholar]
- Pérez-Moncada, U.A.; Santander, C.; Ruiz, A.; Vidal, C.; Santos, C.; Cornejo, P. Design of microbial consortia based on Arbuscular Mycorrhizal fungi, yeasts, and bacteria to improve the biochemical, nutritional, and physiological status of strawberry plants growing under water deficits. Plants 2024, 13, 1556. [Google Scholar] [CrossRef]
- Menendez, E.; Garcia-Fraile, P. Plant probiotic bacteria: solutions to feed the world. AIMS Microbiol. 2017, 3, 502–524. [Google Scholar] [CrossRef]
- Souza Vandenberghe, L.P.; Garcia, L.M.B.; Rodrigues, C.; Camara, M.C.; de Melo Pereira, G.V.; de Oliveira, J.; Soccol, C.R. Potential applications of plant probiotic microorganisms in agriculture and forestry. AIMS Microbiol. 2017, 3, 629–648. [Google Scholar] [CrossRef]
Figure 1.
Growth conditions.

Figure 2.
Impact of PGPMs and Ca salts on RWC of winter wheat leaves after drought stress. Error bars represent the standard deviation of the mean. Means with different letters in the same color columns are significantly different (p < 0.05).
Figure 2.
Impact of PGPMs and Ca salts on RWC of winter wheat leaves after drought stress. Error bars represent the standard deviation of the mean. Means with different letters in the same color columns are significantly different (p < 0.05).
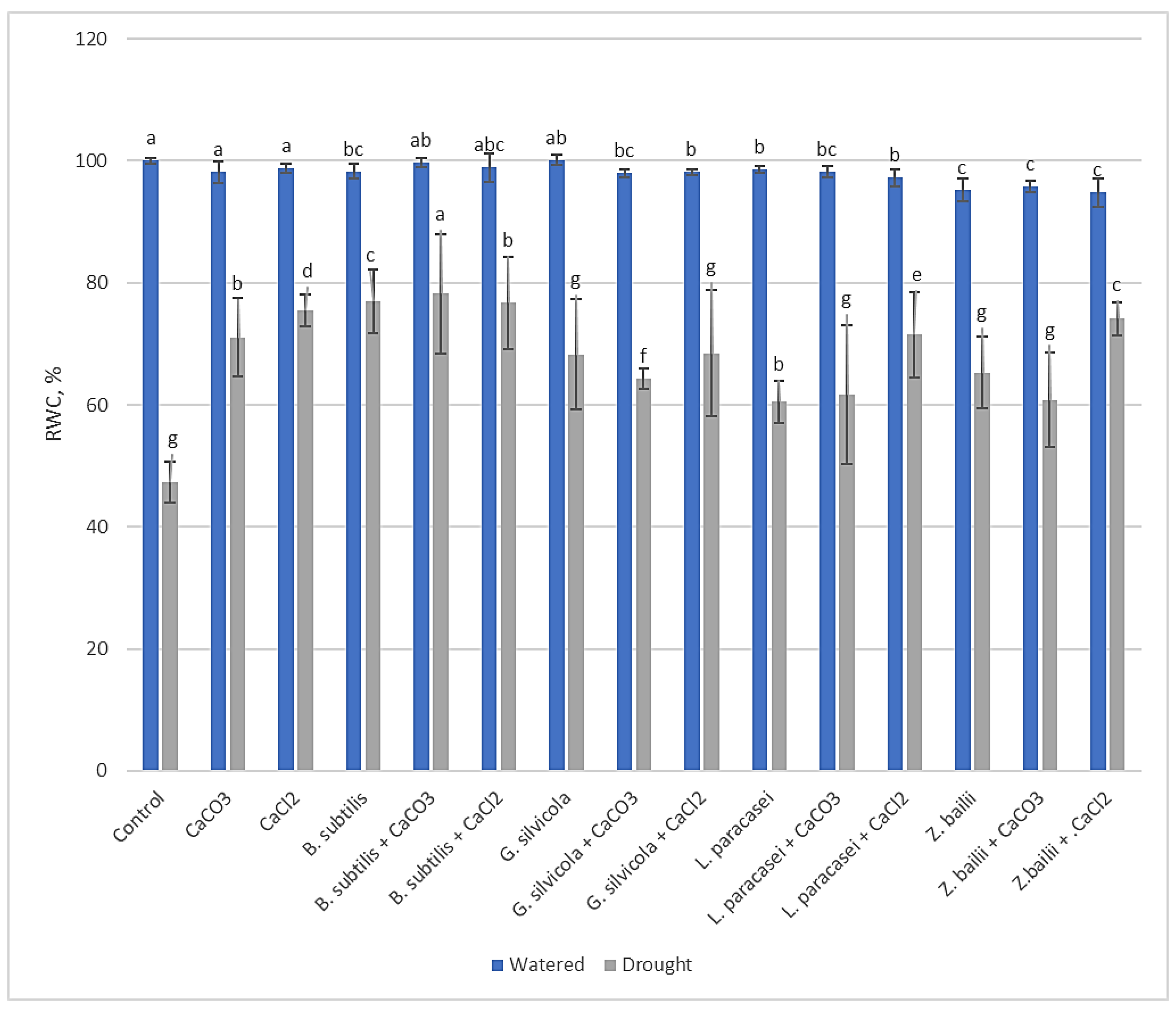
Figure 3.
The effect of PGPMs and Ca salts application and prolonged drought stress on winter wheat H2O2 content. Error bars represent the standard deviation of the mean. Different lowercase letters in the same color columns indicate statistically significant differences (p < 0.05).
Figure 3.
The effect of PGPMs and Ca salts application and prolonged drought stress on winter wheat H2O2 content. Error bars represent the standard deviation of the mean. Different lowercase letters in the same color columns indicate statistically significant differences (p < 0.05).
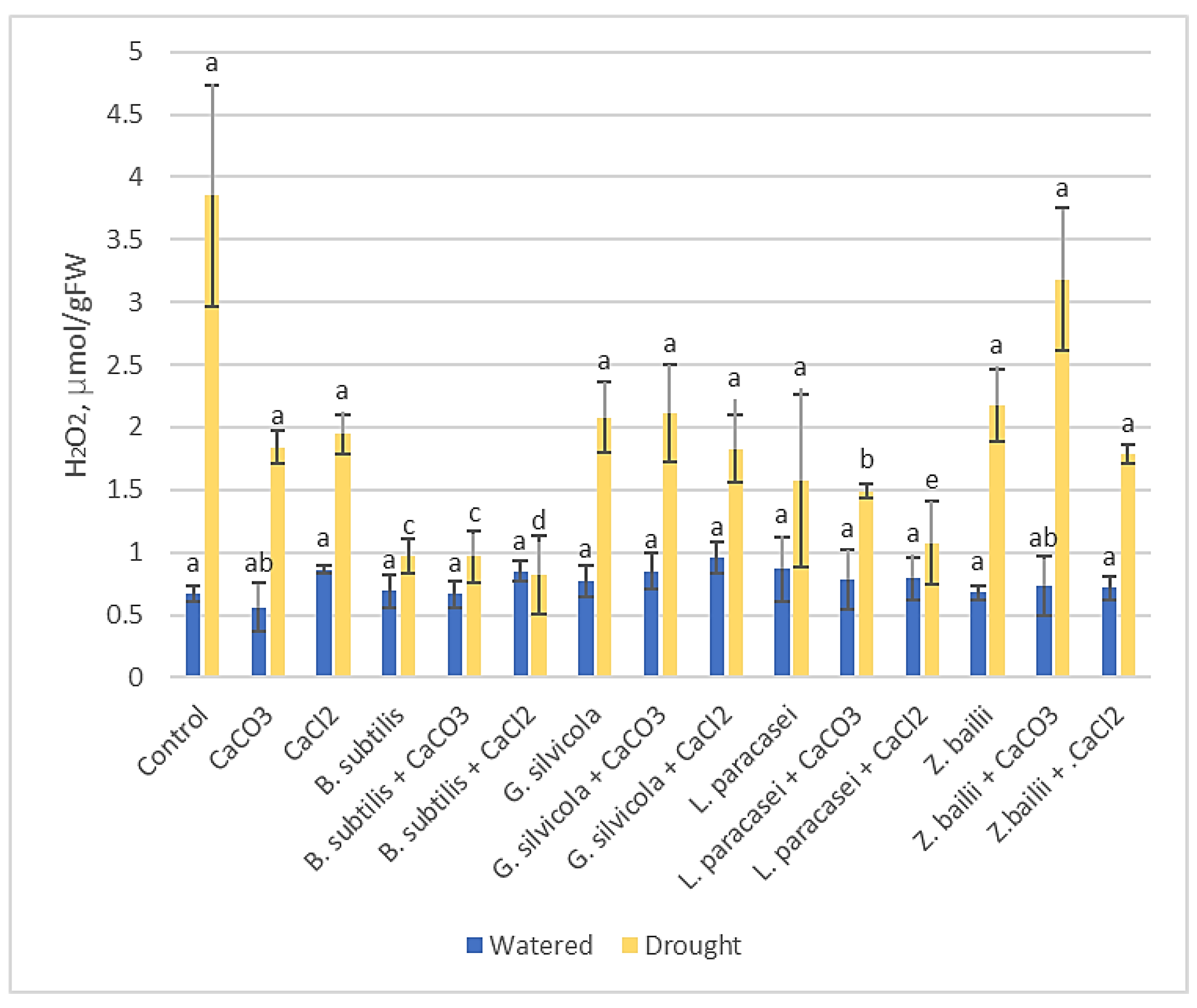
Figure 4.
The effect of PGPMs and Ca salts application and prolonged drought stress on winter wheat MDA content. Error bars represent the standard deviation of the mean. Different lowercase letters in the same color columns indicate statistically significant differences (p < 0.05).
Figure 4.
The effect of PGPMs and Ca salts application and prolonged drought stress on winter wheat MDA content. Error bars represent the standard deviation of the mean. Different lowercase letters in the same color columns indicate statistically significant differences (p < 0.05).
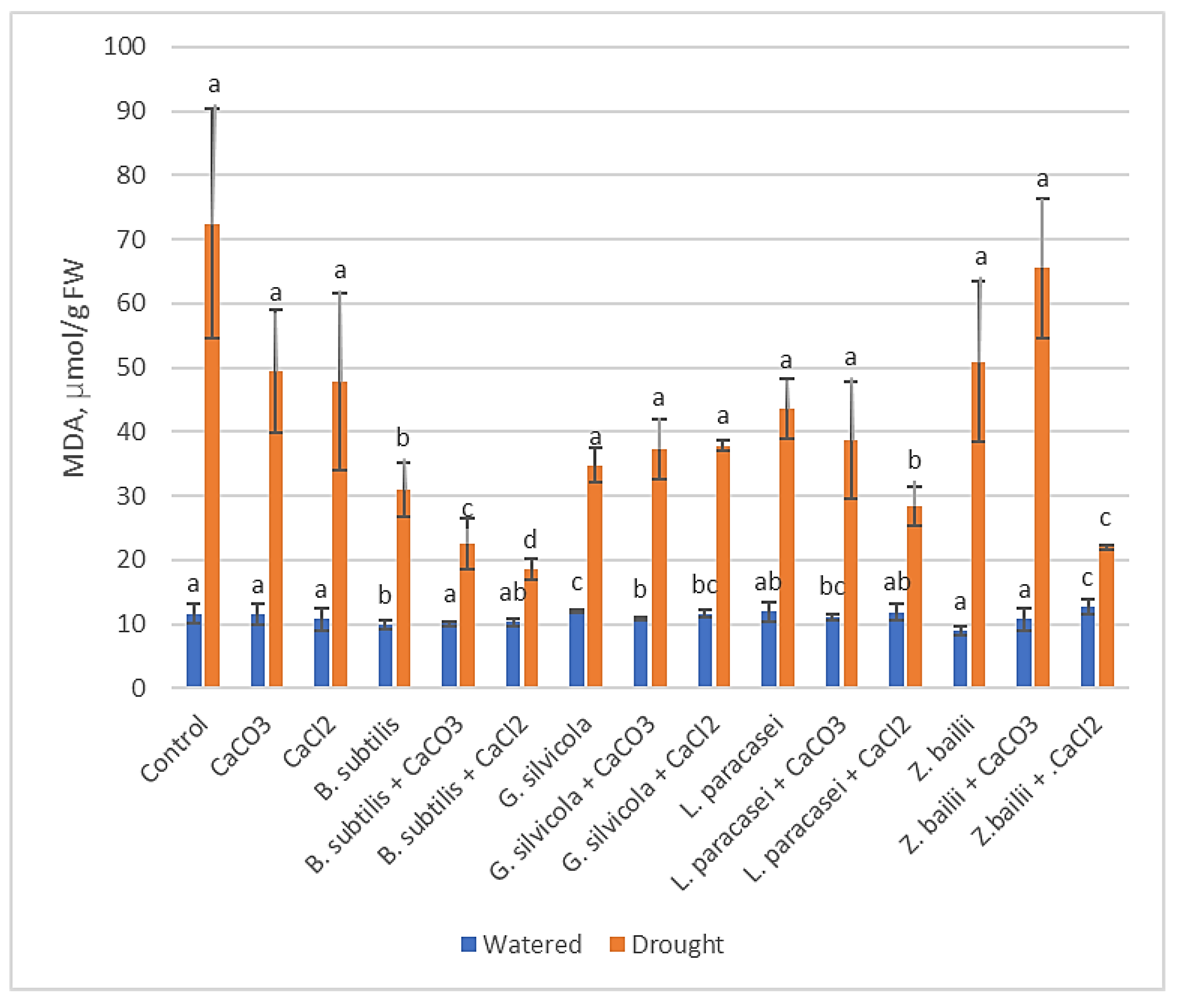
Figure 5.
The effect of PGPMs and Ca salts application and prolonged drought stress on winter wheat proline accumulation. Error bars represent the standard deviation of the mean. Different lowercase letters in the same color columns indicate statistically significant differences (p < 0.05).
Figure 5.
The effect of PGPMs and Ca salts application and prolonged drought stress on winter wheat proline accumulation. Error bars represent the standard deviation of the mean. Different lowercase letters in the same color columns indicate statistically significant differences (p < 0.05).
Figure 6.
The effects of PGPMs and Ca salts application and prolonged drought stress on winter wheat lea genes (a - Td27e, b - Td29b, c - Td11) expression level. Different lowercase letters in different gene fold changes indicate statistically significant differences (p < 0.05).
Figure 6.
The effects of PGPMs and Ca salts application and prolonged drought stress on winter wheat lea genes (a - Td27e, b - Td29b, c - Td11) expression level. Different lowercase letters in different gene fold changes indicate statistically significant differences (p < 0.05).
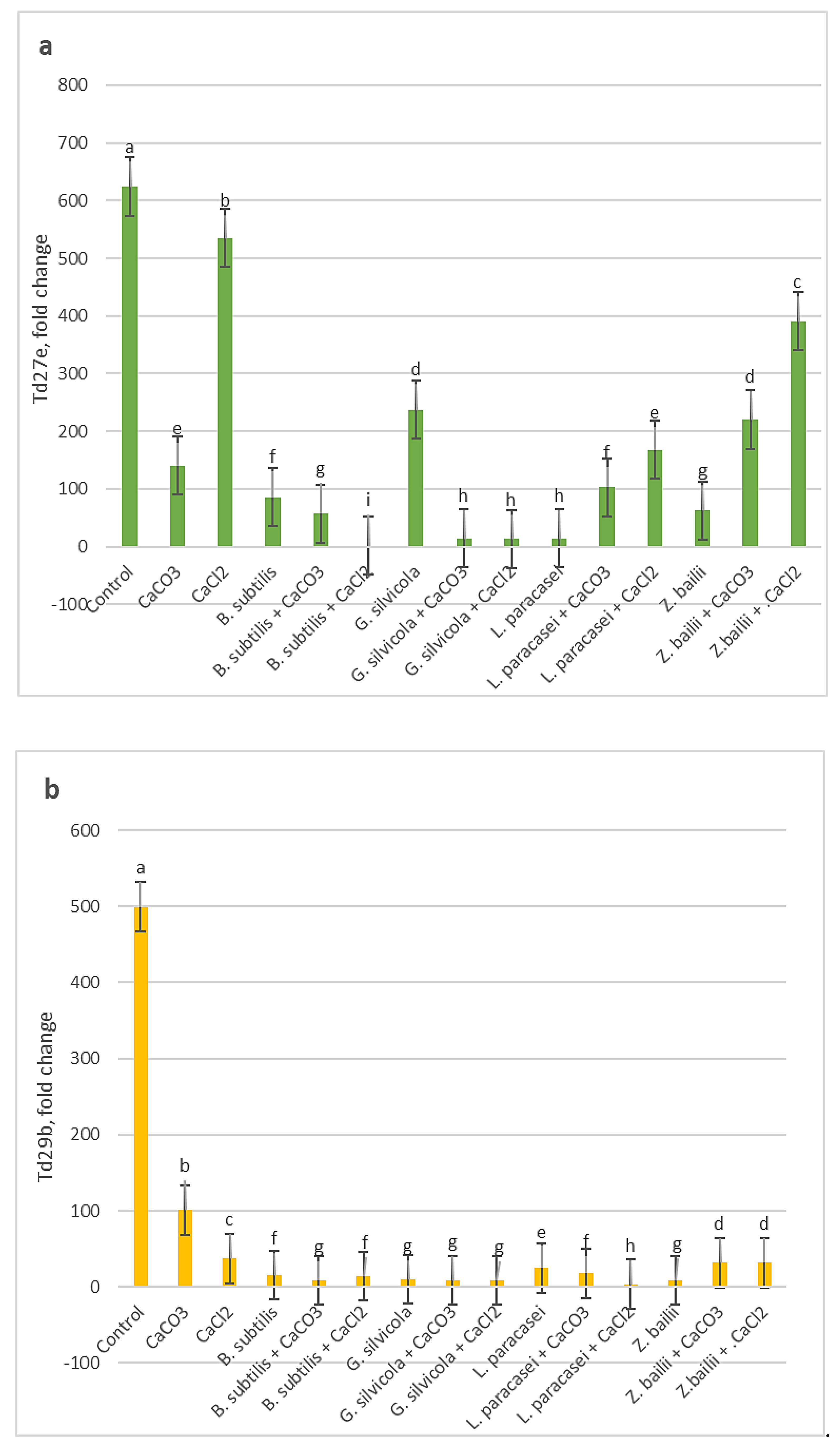
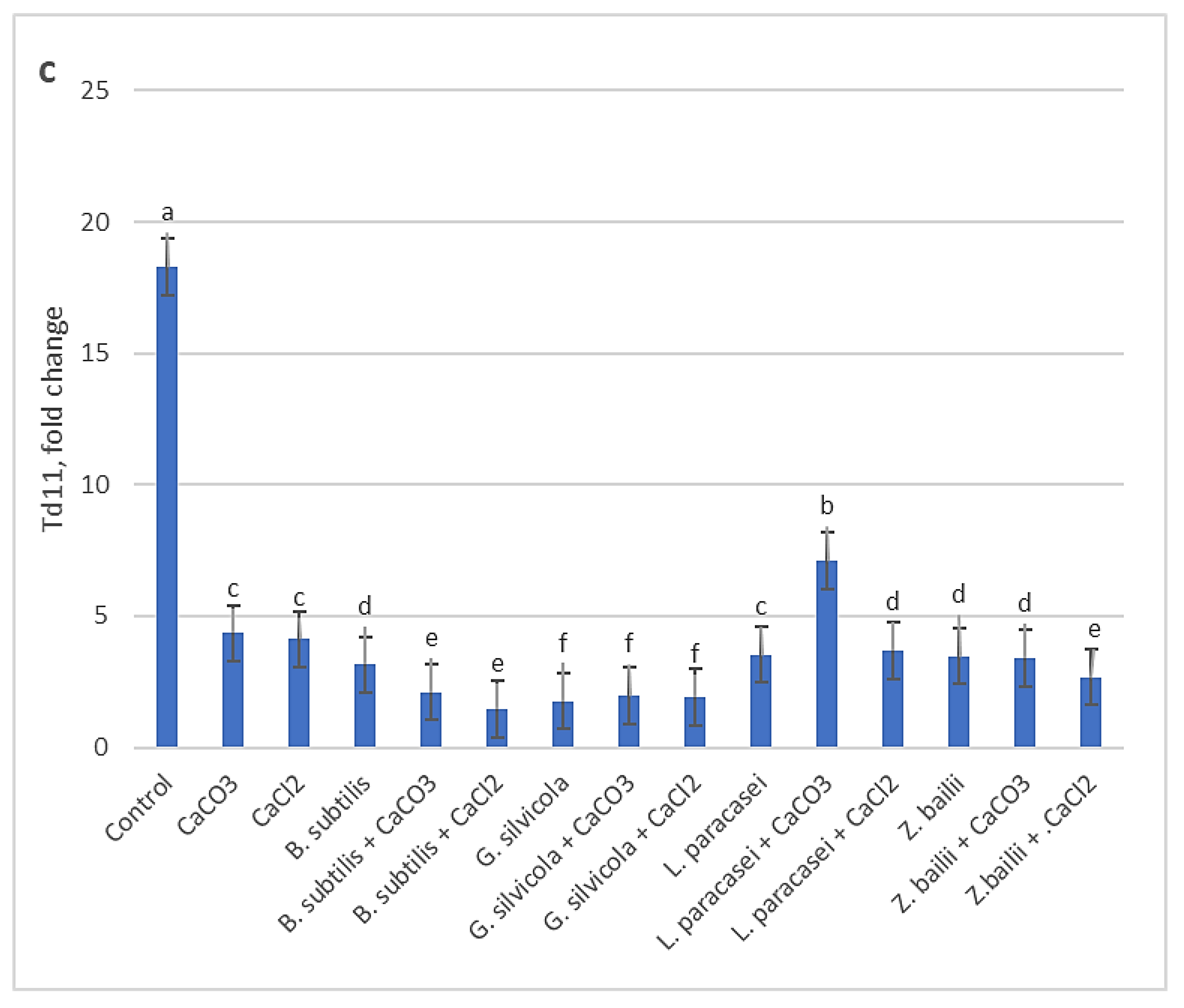

Table 1.
Experimental design.
| Treatment | CaCO3 | CaCl2 | Microorganisms |
|---|---|---|---|
| Control | - | - | - |
| CaCO3 | + | - | - |
| CaCl2 | - | + | - |
| Bacillus subtilis | - | - | + |
| B. subtilis + CaCO3 | + | - | + |
| B. subtilis + CaCl2 | - | + | + |
| Geotrichum silvicola | - | - | + |
| G. silvicola + CaCO3 | + | - | + |
| G. silvicola + CaCl2 | - | + | + |
| Lactobacillus paracasei | - | - | + |
| L. paracasei + CaCO3 | + | - | + |
| L. paracasei + CaCl2 | - | + | + |
| Zygosaccharomyces bailii | - | - | + |
| Z. bailii + CaCO3 | + | - | + |
| Z. bailii + CaCl2 | - | + | + |
Table 2.
The sequences of primers used in the work.
| Gene | LEA Proteins Group | Primer Pairs | Primer Sequences (5′-3′) |
|---|---|---|---|
| 26S | F/R | CCGGTTGTTATGCCAATAGCA/GCGGCGCAGCAGTTCT | |
| Td11 | 2 | F/R | AGGTGATCGATGACAACGGTG/ACCCTCGGTGTCCTTGTGG |
| Td29b | 4 | F/R | CGCACCCAGCTAGTAAGTTCG/CCCAGCCCAGTAATAACCCAT |
| Td27e | 2 | F/R | CAGCACTGAGCCGACGG/ACGTGGAACTAGAAGGCATTGAC |
Table 3.
Effect of PGPMs and Ca salts on morphometric parameters of winter wheat seedlings (per plant). Different letters in columns designate statistically significant differences at p < 0.05.
Table 3.
Effect of PGPMs and Ca salts on morphometric parameters of winter wheat seedlings (per plant). Different letters in columns designate statistically significant differences at p < 0.05.
| Average shoot length, cm | Average shoot biomass, g | |||
|---|---|---|---|---|
| Watered | Drought | Watered | Drought | |
| Control | 24.2 d | 15.95 f | 0.288 e | 0.039 f |
| CaCO3 | 26.95 d | 17.05 e | 0.3159 c | 0.036 f |
| CaCl2 | 24.55 c | 17.75 e | 0.267 f | 0.0335 f |
| Bacillus subtilis | 27.95 a | 20.65 b | 0.333 b | 0.1278 b |
| B. subtilis + CaCO3 | 27.55 a | 21.65 a | 0.345 a | 0.1164 c |
| B. subtilis + CaCl2 | 27.85 a | 19.73 c | 0.3197 c | 0.1218 b |
| Geotrichum silvicola | 26.6 b | 20.5 b | 0.29 e | 0.107 d |
| G. silvicola + CaCO3 | 26.9 b | 19.56 c | 0.297 d | 0.109 d |
| G. silvicola + CaCl2 | 27.1 ab | 18.45 d | 0.316 c | 0.112 c |
| Lactobacillus paracasei | 25.3 c | 18.55 d | 0.269 f | 0.068 e |
| L. paracasei + CaCO3 | 26.55 b | 18.85 d | 0.306 d | 0.057 e |
| L. paracasei + CaCl2 | 25.8 d | 20.45 b | 0.253 f | 0.134 a |
| Zygosaccharomyces bailii | 25.1 c | 18.65 d | 0.2846 e | 0.072 e |
| Z. bailii + CaCO3 | 26.45 b | 16.4 f | 0.309 d | 0.035 f |
| Z. bailii + CaCl2 | 27.4 a | 20.5 b | 0.289 e | 0.1454 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



