
Submitted:
20 March 2025
Posted:
21 March 2025
You are already at the latest version
Abstract
Tylosema esculentum and Tylosema fassoglense are highly nutritious, drought tolerant perennial legumes that typically grow in nutrient poor soil in hot environments. Despite marama beans being the ideal crop for domestication, the legume remains unexploited. Marama beans are prone to infestation by insect pests although limited information exists in literature on the pests of the legumes. Insect pests of marama beans that could pose a threat to yield were identified in this study. Active and passive insect collection methods were undertaken on marama plants growing in the wild in the North West and Limpopo provinces of South Africa. Sampled insects were identified manually and by molecular methods. Identified insects were from the orders Lepidoptera, Coleoptera, Hemiptera, Orthoptera, Isoptera, Odonata, Mantodea, Diptera and Blattodea. Coleopteran, Orthopteran and Hemipteran adults caused damage to leaves and flowers while Lepidopteran larvae caused damage to the leaves, pods and seeds. Identified insects have the potential to limit yield of marama beans, therefore this study presents an opportunity for the determination of abundance of specific pests, the extent of damage as it relates to productivity to establish relevant pest management practices.
Keywords:
Tylosema esculentum
; Tylosema fassoglense
; Marama bean
; Insect pests
; pollinators
; food security
1. Introduction
Cereals such as Sorghum bicolor, Zea mays and Triticum aestivum as well as legumes such as Phaseolus vulgaris, Glycine max and Pisum sativum are examples of crops grown in South Africa that serve as essential food sources and animal feed [1,2,3]. Although these food grains play a significant role in assisting households to become food secure, there is an increasing need for additional sources of nutrients. Indigenous crops, such as marama beans, have the potential to supplement these long-established crops. The marama bean is a perennial legume that belongs to the family Fabaceae of the subfamily Cerdidiodeae. Commonly known as the Gemsbok bean, Gemsbokboontjies, Muraki, Braaiboontjies, Marumama, Tamani berry, and Murama [4,5], the genus Tylosema comprises four recognized species that exhibit morphological variations both between and within populations [6]. These are T. esculentum (Burch.) A. Schreib., T. fassoglense (Schweinf). Torre & Hillc., T. argenteum (Chiov.) Brenan and T. humifusum (Pic. Serm & Roti Mich.) Brenan [6,7]. Classification also includes T. angolense that was located in the southern parts of Angola [6]. Tylosema species are indigenous to Southern Africa and are widely distributed across the Kalahari Desert, Botswana (Kgalagadi), Eastern and North eastern Namibia, the Northern part of South Africa (Northwest, Northern Cape & Limpopo), Angola, Zambia and Mozambique [7,8]. T. fassoglense is located in East African countries that include Angola while T. esculentum is restricted to Namibia, Botswana and the Northern parts of South Africa [6].
T. esculentum (Burch.) A. Schreib. and T. fassoglense (Schweinf). Torre & Hillc. are characterized by large tubers which can grow to weights of 12 kg or even up to 80 kg at maturity [8,9,10]. Tylosema species possess trailing, climbing or running stems, which can grow up to 6m long in T. fassoglense and 3m long in T. esculentum [8,11]. T. esculentum leaves are bilobed for half of the leaf length while those of fassoglense are bilobed for one tenth to a third of the leaf length, which typically ranges from 8 cm to 15 cm in both species. Yellow flowers that eventually fade to pink in fassoglense begin to emerge in October to February or until March for esculentum.
Marama bean pods produce one to two seeds per pod that vary in colour and shape. Seeds are round and brownish-black or flattened and chestnut-brown in esculentum and fassoglense respectively [11,12,13]. Seeds are rich in protein, lipids, vitamins, carbohydrates, iron, zinc, calcium, copper, magnesium, potassium, phosphate and sodium [8,11,14,15], with mature seeds exhibiting a higher nutrient content than immature edible seeds [16]. T. fassoglense seeds have been shown to consist of a higher protein composition than those of T. esculentum, which have a protein content similar to that of soybean [17].
The tuber consists of carbohydrates, proteins and 90% water [9]. Marama bean seeds are consumed raw, boiled or roasted and can be used to produce marama milk, oil, flour and butter [9]. Stems and tubers are often used as animal feed and feed for game [18,19,20,21].
Despite their economic potential, marama beans remain largely unexploited. Moreover, their domestication and commercialization could enhance crop diversity by supplementing traditional food grains [5]. Furthermore, increased levels of subsistence and commercial production of traditional crops in addition to marama beans can potentially reduce the risks associated with food insecurity for poor households in rural and urban areas [22,23]. However, infestation by insect pests often compromises productivity of crops. The effect of insect infestations on productivity, nutrient composition and yield of maize, sorghum, peas, common bean and soybean has been widely studied. On the contrary, no evidence exists in liter[11–13ature on the effect of insect pest infestations on marama beans, in addition to the identification of insects that are associated with the indigenous plant. Hence, the aim of this study was to identify insect pests that could potentially reduce productivity and yield of T. esculentum and T. fassoglense.
2. Materials and Methods
2.1. Description of Study Sites
The study was conducted in the Limpopo and the Northwest provinces of South Africa. Study sites in Limpopo and the Northwest provinces were natural habitats of marama bean in Tzaneen (-23.87302, 30.09474) and Lephalale (-23.81556, 27.86202) as well as Tosca (-25.84633, 23.81300) and Pomfret (-25.84703, 24.13538) respectively. The elevations of Tosca and Pomfret are 1127.17 m and 1158 m above sea level. The sites have a subtropical steppe climate with an average annual precipitation of 37.16 mm. Similarly, study sites in the Limpopo province are classified as Subtropical steppe and are situated at an altitude of 1020.61 meters above sea level. The average annual precipitation in Limpopo is approximately 55.48 mm.
2.2. Sampling of Insects
A combination of active and passive collection methods as described by Southwood and Henderson [24] and Woodcock [25] were used to sample insects from all sites. Sweep nets with 89 cm long handles, 41 cm diameter net ring and 91 cm net depth were used to actively sweep entire fields for all insects while some were hand-picked and captured by beating from plant leaves, stems and flowers. Malaise traps were used to capture flying insects and 10 cm wide (brim), 12 cm long, 500 ml disposable plastic cups were used as pitfall traps to capture ground dwelling arthropods. The plastic cups were half-filled with 40 parts water and 60 parts propylene glycol to preserve as well as prevent insects from escaping. Traps were buried in the ground such that the brim would be at the same level as the surface of the ground. Two pitfall traps were set for each marama bean plant at both sites while single malaise traps were set in Tosca and Pomfret (NW).
Traps (pitfall and MT) were removed three days after setting. Pitfall traps can be removed at anytime between two days and four weeks [24,25] however, at these study sites, traps were removed after three days because the most accessible marama bean populations were located near the road so traps were prone to damage by large animals and passers-by when left for prolonged periods.
The average distance between marama bean plants was 40 m in both provinces. Marama bean populations were visited during the months of January, March, May and July in each Province. Collected insects from all sites were placed in glass vials consisting of propylene glycol and taken to the Plant Biotechnology laboratory at the University of the Western Cape (-22.9314799, 18.6244442) in Cape Town, South Africa where they were washed and frozen for molecular identification. Manual identification for larger and easily identifiable insects was done in the field before transportation to the laboratory.
2.3. Identification of Sampled Insects
2.3.1. Visual Identification Using Morphological Characteristics
Insects were identified visually by using morphological characteristics as described by Picker[26], Brier[27], Marshall[28] and Scholtz and Holm[29]. Insects were placed in plastic petri-dishes and visualised using a Novax-Holland Model 65.550 dissecting microscope. Identification was done up to species level although some were identifiable only up to family level.
2.3.2. Molecular Identification
2.3.2.1. Extraction of Genomic DNA
Approximately 50 mg insect tissue consisting of either a leg, thorax, abdomen or a wing were grinded in liquid nitrogen using a motar and pestle. The resulting powder was used for DNA isolation using the Zymo research Quick-DNA Tissue/Insect Mini-prep KitTM (catalog D6016, Zymo research corp; California, USA). For this DNA isolation, the tissue powder was transferred to a 2.0 mm ZR BashingBead™ Lysis tube following the catalog D6016 protocol. Afterwhich, 750 µl of BashingBead™ Lysis Buffer was added to the tube and the tube was tightly closed. The tube was then placed in a vortex mixer for 5 minutes to homogenise the mixture. The tube containing the homogenate was then centrifuged in a microcentrifuge at 10,000 x g for 1 minute. Thereafter, 400 µl of the supernatant was transferred to a Zymo-Spin™ III-F Filter which was placed in a 2.0mm collection tube and centrifuged at 8,000 x g for 1 minute. After centrifugation, 1,200 µl of Genomic Lysis Buffer was added to the filtrate in the collection tube and mixed well. 800 µl of the mixture with the genomic lysis buffer was then transferred to a Zymo-Spin™ IICR Column that was placed in a collection tube and centrifuged at 10,000 x g for 1 minute. The flow through from the collection tube was discarded and an additional 800 µl of the genomic lysis buffer mixture was transferred to the Zymo-Spin™ IICR Column and re-centrifuged at 10 000 x g for 1 minute. The flow through was discarded again. The DNA in the Zymo-Spin™ IICR Column was washed by adding 200 µl DNA Pre-Wash Buffer and centrifuging at 10,000 x g for 1 minute. An additional wash step involved the addition of 500 µl g-DNA Wash Buffer to the Zymo-Spin™ IICR Column that was then centrifuged at 10,000 x g for 1 minute. Thereafter, the Zymo-Spin™ IICR Column was transferred to a clean 1.5 ml microcentrifuge tube and 100 µl of DNA Elution Buffer was added directly to the column matrix. The tube, with the column was centrifuged at 10,000 x g for 30 seconds to elute the DNA. Extracted DNA was quantified using a Nanodrop spectrophotometer and gel electrophoresis. Quantifying by gel electrophoresis was achieved by using 1% gels consisting of 1X TAE buffer (40mM Tris-base, 20mM Acetic acid and 1mM EDTA) with a pH of 8.6 and 1 g of agarose powder. Gels were placed in gel running chambers that were filled with TAE buffer and wells loaded a 6 µl sample consisting of 5µl extracted insect DNA and 1 µl of 6X DNA loading dye. The gels were run at 80 V for 60 minutes until the dye was at the bottom of the gel. The gels were then viewed and documented using a BIO-RAD ChemiDocTM imaging system.
2.3.2.2. Genomic DNA Amplification
PCR reactions were set up by adding 2.5 µl DNA to a 22.5 µl Mastermix consisting of 8.5 µl Nuclease-free water, 12 µl amplicon, 1 µl reverse (HCO2198: 5'-TAA ACT TCA GGG TGA CCA AAA AAT CA-3') and 1 µl forward (LCO1490: 5'-GGT CAA CAA ATC ATA AAG ATA TTG G-3') primers [30] to a final volume of 25 µl in PCR tubes. The tubes were centrifuged and placed in a thermocycler. Thermocycling conditions involved an initial denaturation step at 95°C for 1 minute, followed by 10 cycles of 95°C for 2 minutes, 45°C for 1 minute and 72°C for 2 minutes. The next step included 35 cycles of 93°C for 45 seconds, followed by 58°C for 30 seconds and 72°C for 1 minute and 25 seconds. This was followed by a 10-minute cycle at 72°C and concluded with a 4°C infinite step. PCR products were visualised through agarose gel electrophoresis.
2.3.2.3. Purification and Sequencing of PCR Products
DNA fragments were excised from the agarose gel using a scalpel and weighed. The excised fragments were then transferred to 1.5 ml microcentrifuge tubes in which ADB (Agarose Dissolving Buffer) three times the volume of the excised agarose gel fragments was added. The samples were incubated at 55 °C for 20 minutes and mixed by vortexing. The completely dissolved and melted agarose solution was transferred to a Zymo-Spin™ Column in a Collection Tube and centrifuged for 1 minute. After which the flowthrough was discarded. 200 µl of DNA Wash Buffer as added to the column which was then centrifuged for 30 seconds to purify the sample. The flow through was discarded and the sample washed again. 6 µl DNA Elution Buffer consisting of 10 mM Tris, pH 8.5 and 0.1mM EDTA (Zymo research corp, USA) was added directly to the column matrix. The column was placed into a 1.5 ml tube and centrifuged for 1 minute to elute DNA.
PCR products were sent to the Central Analytics Facility (CAF) DNA sequencing unit at Stellenbosch University, Cape Town, for sequencing. Sequences were edited and analysed using the BioEdit (Biological Sequence Alignment Editor) software. Following editing and alignment, sequences were checked for similarities against information available in NCBI (National centre for Biotechnology Information) databases using a BLAST (Basic Local Alignment Search Tool) nucleotide search.
3. Results
3.1. Insect Pests Associated with Tylosema fassoglense
Insects associated with T. fassoglense were identified using molecular methods, while others were identified morphologically. Sampled insects were identified to the species level and their feeding guilds were determined, as shown in Table 1.
Those that were un-identifiable were designated as Undetermined and assigned numbers while their feeding guilds were assigned based on the taxonomic family under which they were identified.
A total of seven orders, seventeen families and twenty-two species were identified. Identified insects were from the orders Coleoptera, Orthoptera, Hemiptera, Lepidoptera, Isoptera, Blattodea and Mantodea. The order Lepidoptera contained the highest number of families and species, whereas Mantodea and Blattodea were the least represented, each with a single family and species.
Two families and two species each represented the orders Isoptera and Hemiptera. Four families and five species represented the order Coleoptera. Orthoptera were represented by two families and three species although two species under the family Acrididae remained unidentified. Five families and eight species including an unidentified caterpillar represented Lepidoptera. A majority of identified insects were herbivorous pests.
Identified species from the order Blattodea were cockroaches (Deropeltis erythrocephala Fabricius, 1781), while those from the order Isoptera were the harvester termites Microhodotermes viator Latreille, 1804 and Amitermes hastatus, Haviland, 1898. Insects from the order Coleoptera were leaf beetles (Diabrotica undecimpunctata howardi Barber), net winged beetles (Lycus sp Guérin-Méneville, 1835), flower chafers (Protaetia brevitarsis Lewis, 1879 & Maladera drescheri Moser, 1913) and blister beetles (Zonitis sayi Wickham, 1905). The order Mantodea was represented by mantids (Hoplocoryphella grandis Brancsik, 1895) while insects from the order Orthoptera were gaudy grasshoppers (Zonocerus elegans Thunberg, 1815) and two unidentified species designated as Undetermined 2 & Undetermined 3 as shown in Figure 2.
Hemiptera were burrowing bugs (Sehirus cinctus Palisot, 1811) and stinkbugs (Nezara viridula Linnaeus, 1758). Lepidopteran insects were monkey moths and caterpillars (Tantaliana tantalus or Jana tantalus Herrich Schäffer, 1854), owlet moths and caterpillars (Proteuxoa sp Hampson (1903) and Agrotis ipsilon Hufnagel, 1766), lappet moths (Macrothylacia rubi Linnaeus, 1758 and Janomima mariana White, 1843), geometer moths, brush-footed butterfly larvae (Cynthia or Vanessa cardui Linnaeus, 1758), hawkmoths (Theretra oldenlandiae Fabricius, 1775) and a species designated as Undetermined
3.2. Insects Associated with Tylosema esculentum
Six orders, twenty-five families and nineteen species that include seven unidentifiable insects were sampled on T. esculentum. T. esculentum showed greater variation in feeding guilds, taxonomic families and species when compared to T. fassoglense. Identified orders were similar to those that were identified for T. fassoglense, with an addition of Odonata although Mantodea were excluded. The order Coleoptera consisted of more families and species, followed by Orthoptera and Lepidoptera. The least represented orders were Blattodea and Odonata that consisted of a single species each, although the species under Blattidae was unidentifiable and designated as Undetermined 4. Identified insect species are listed in Table 2.
Insects from the order Coleoptera were ground beetles (Anthia cinctipennis Lequien, 1832 and Passalidius fortipes Boheman, 1860), darkling beetles (Gonopus tibialis Fabricius, 1798, Psammodes bertolonii Guerin-Meneville, 1844 and P. striatus Fabricius, 1775), hide beetles (Ormogus aperulatus Harold, 1872), scarab beetles (Sisyphus sp Latreille, 1807), pleasing fungus beetles (Amblyopus sp Lewis 1889), Leaf beetles (Altica sp Geofrroy 1762 and Lema rufotestacia Clarke, 1866), blister beetles (Zonitis sayi Wickham, 1905) and shining flower beetles (Olibrus sp Erichson, 1845).
Hemiptera were froghoppers (Locris arithmetica Walker, 1851), jewel bugs (Solenostethium liligerum) and stink bugs (Nezara viridula Linnaeus, 1758). Insects from the order Orthoptera were identified as skimmers, armoured ground crickets (Acanthoplus discoidalis Walker, 1869; Figure 4), gaudy grasshoppers (Zonocerus elegans
Thunberg, 1815), short-horned grasshoppers (Locustana pardalina Walker, 1870) and bush crickets (Tettigonia viridissima Linnaeus, 1758) as shown in Figure 3 below. Lepidopteran insects were tiger moths and caterpillars (Achaea janata Linnaeus, 1758 and Utetheisa pulchella Linnaeus, 1758), owlet moths and caterpillars (Proteuxoa sp Hampson(1903), Helicoverpa armigera Hübner (1808), Cyligramma latona Cramer (1779) and Agrotis ipsilon Hufnagel, 1766), lappet moths (Macrothylacia rubi Linnaeus, 1758) and hawkmoths (Theretra oldenlandiae Fabricius, 1775) and unidentified caterpillars designated as Undetermined 6, 7 and 8 . Insects that were associated with T. esculentum were mostly herbivorous and omnivorous pests with few exclusive predators.
4. Discussion
4.1. Insects Associated with T. fassoglense
Insects from the orders Blattodea, Isoptera, Coleoptera, Hemiptera, Lepidoptera, Mantodea and Orthoptera were sampled form Tylosema fasssoglense in the Limpopo Province of South Africa. Sampled insects were predominately pests except for Mantodea that were identified as predators. Insects from the order Lepidoptera were the second represented order after Coleoptera. Lepidopteran pests represented a variety of feeding guilds including pod and seed borers as well as leaf suckers and flower feeders. Similarly, Coleoptera were designated as leaf and flower feeders while Isoptera feed on roots, stems, and seeds. Hemiptera were a combination of leaf sap, pod and seed feeders while Orthopteran pests are typically leaf feeders. Although various feeding guilds were represented, most sampled insects were observed on leaves, while some were suspended in pitfall traps at the time of removal. Selected Lepidoptera and Coleoptera were however, observed in pods, within seeds and on flowers. Generally, insects from the order Isoptera are not recognised as major pests of crops, although sighted causing damage to trees since they have an affinity for wood. M. viator and A. hastatus that were sampled in T. fassoglense pods as shown in Figure 2A were rarely reported as pests in literature until recently. Harvester termites were shown to build mounds on agricultural land thereby reducing productivity of cultivated crops [31]. Similarly, another study highlighted how M. viator build mounds in nutrient poor soil and attack drought-stricken crops [32]. A. hastatus on the other hand was shown to have caused observable damage in sugarcane fields in Brazil in addition to being responsible for crop losses that ranged from 3 to 100% in Africa and Asia [32,33,34,35,36]. Additionally, the effect of termite infestation on maize was shown in Ethiopia [37], highlighting the affinity of termites for maize, although also arguing that a maize, soybean intercropping system could possibly control the occurrence of these pests on maize. Similarly, percentage loss in maize in Zambia is reported to range between 15-25% [38]. Interestingly, sightings of termites in legumes is not widely reported, thus this study is among the few to report termites in legumes and the first to document termites on T. fassoglense. Although a few researchers to report on global sightings of termites attacking roots of soybean and other crops at seedling stage [39]. The occurrence of termites in mature marama bean pods that still consisted of intact seeds in this study is justified given the woody nature of mature T. fassoglense pods.
The bush cockroach, D. erythrocephala, although sampled from pitfall traps that were setup on T. fassoglense is not reported in literature as a pest of crops, however, it has been shown to occur in gardens. Like termites, D. erythrocephala is reportedly prevalent in areas that have an abundance of wood and is regarded as a household pest that hides in crevices rather than an agricultural pest [40]. Although, damage that could have been caused by the insect on T. fassoglense is unclear, sampling in T. fassoglense could largely be attributed to the fact that the vegetation that surrounded marama bean plants consisted mostly of shrubs, in addition to mature marama bean pods being woody. Similarly, the Mantid, H. grandis was found on marama bean leaves that had characteristic brown patches. H. grandis is a predator that is widely distributed in Medagascar, Tanzania, the former Transvaal in South Africa (between the Limpopo province and the Vaalriver) and Zimbabwe [41]. Although the predatory status of this insect is poorly reported in literature, it is said to be a generalist predator of a variety of insects that include flies, moths, crickets, and grasshoppers [42]. Given its feeding behaviour as a generalist predator, its presence in T. fassoglense may be because of the abundance of the preferred diet. H. grandis, as shown in Table 1 occurred together with Lepidoptera and Orthoptera. On the contrary, Coleopteran pests such as Z. sayi and D. undecimpunctata are foliage and root feeders respectively. Z sayi is a blister beetle that is distributed in America, although the genus Zonitis is globally distributed [43]. Z. sayi infestations are reported on alfafa, pigweed, amaranth, goathead, peanuts and soybean in the USA [44]. For this study, the insect was found sucking on marama bean leaves, causing notable damage to the leaf. Some Z. sayi were sampled while feeding, consequently, leaves appeared translucent at the feeding site. Records of Z. sayi on crops or in field studies in South Africa are limited. However, D. undecimpunctata as a pest is reported extensively in literature [45,46,47]. Both the larval and adult form of the insect cause damage to specific host plants. The larval form of D. undecimpunctata is known as the corn rootworm that feeds on the roots of cucurbits, maize, peanuts and beans while the adult beetle that is recognised as a generalist feeder is called the spotted cucumber beetle [48]. D. undecimpunctata adults lay eggs on soil near the base of host plant seedlings. Emerging larvae then feed on the roots until development is complete. Pupation then proceeds in the soil until adults emerge to feed on foliage. Characteristic damage is normally observed on leaves and stems [49]. The likeliness of D. undecimpunctata to spread over a wide area was highlighted [50,51] which suggests that upon the introduction of marama bean in agricultural fields, D. undecimpunctata could prevail. Burrower bugs, Sehirus cinctus are generally not considered pests of food grains, consequently, there is limited information in literature on these bugs on crops. S. cinctus in this study were sampled form pitfall traps and on leaves of T. fassoglense although no damage was observed on the leaves at the position where S. cinctus was observed. In the USA, S. cinctus was sampled on peanuts where no significant damage nor economical loss resulted from the infestation [52,53]. Infestations on soybean were reported in America [54] while no records of infestations exist in South Africa. Generally, Cydnidae are reported as generalist feeders although legumes, grasses and conifers are preferred [55], hence their presence on marama beans. The other Hemipteran sampled on T. fassoglense, N. viridula, contrary to S. cinctus is considered a major pest of a variety of food plants. N. viridula was also sampled on soybean in a separate study, further highlighting the preference of Cydnidae and Pentatomidae for leguminous food grains. This suggests that the introduction of marama bean in trial sites normally used for soybean and maize cultivation, N. viridula would be a potential pest for both soybean and Tylosema species, given their affinity for legumes.
Lepidopteran larvae sampled on T. fassoglense were observed causing damage to pods and seeds. The larvae of V. cardui were removed from leaves during feeding as shown in Figure 2 G. The marama bean leaf the insect was feeding on had been extensively defoliated. V. cardui larvae are known to infest a variety of host plants, although a recent report from Brazil showed infestation in soybean that led to significant damage on all plants in the plot [56]. V. cardui adults feed on flowers, with the most preferred flowers in South Africa being those of Ditrichia viscosa, Verbesina encelioides, and Medicago sativa. Infestations have also been reported on maize, alfafa and sunflower [57]. The developmental cycle involves adult females laying eggs on leaves. Emerging larvae feed on host plant foliage as observed in T. fassoglense. Adult V.cardui butterflies were found to migrate between the North West Province of South Africa and Europe [58]. Migration to the North West emphasises the possibility of V. cardui on T. esculentum, although in this study, it was sampled in T. fassoglense in the Limpopo province. Significant damage on a mature pod and seed that was caused by the herringbone caterpillar was also observed. The pod appeared healthy; however, a small hole was observed on the pod and upon opening, the herringbone caterpillar emerged from a hole on the seed. Species of the herringbone caterpillar, Proteuxoa sp, are not mentioned in literature as pests in agricultural fields. Consequently, sightings are reported in dry habitats inclusive of forest edges and grassland in Australia and New Zealand, although also suspected to be herbivorous [59] since grass blades were damaged in grassland habitats in Australia [60]. Proteuxoa species on T. fassoglense pods and damage caused to seeds show the potential of the insect as a major pest that could lead to significant economic loss in agricultural crops. The black cut worm A. ipsilon is a serious pest of vegetables, fruit, cereals, pulses, and legumes. Infestations in fields have been reported across the globe [61]. The larvae are reported as the most destructive life form that completely defoliates crops through feeding on leaves, stems, and young roots of seedlings [61,62] while adult moths attack flowers. Their developmental cycle involves the laying of eggs on host plant leaves by moth adults. Low lying plants with broad leaves are the most preferred for oviposition [61,62]. A. ipsilon is a major threat to marama bean given that the plant is a trailer or climber with broad leaves. All parts of the marama bean plant can be attacked by A. ipsilon especially since both life forms, the caterpillar and the adult moth were sampled on T. fassoglense. No mention is made in literature of Jana tantalus or Tantaliana tantalus and Janomima mariana on crops. However, Grewia villosa and Panicum maximum are listed as host plants of J. mariana in the Limpopo province [58] while evidence on the host plants of Jana tantatus is lacking. Specific Eupterodidae and Lasiocampidae are not recorded although they were sampled from T. fassoglense that was surrounded by grass as shown in Figure 2 D. Information on the origins and distribution of the two species is lacking although thought to be endemic to South Africa [63,64,65,66]. The pest status of both insects is unkown. The nature of interaction between J. mariana and listed host plants is also not discussed. Similarly, J. mariana and J. tantalus sampled on marama beans were not observed causing damage to the plant. The other Lasiocampid, M. rubi is reported as a major pest of Blackberries in Europe [67,68]. In Finland, M. rubi was observed on stems and roots of glasses and clover although no damage was observed [69]. Similarly, the insect was not observed actively causing damage to T. fassoglense. No record exists on M. rubi on agricultural crops in Africa, although observed on T. fassoglense in this study. It is possible that it might not be a pest of crops although studies on the ecology and feeding behaviour of M. rubi would have to be undertaken to confirm this. At this stage, there is no information in literature on such.
4.2. Insect Pests Associated with T. esculentum
Insect species from the orders Blattodea, Hemiptera, Lepidoptera, Odonata and Orthoptera were sampled on T. esculentum. Sampled species were predominately herbivorous pests, followed by predators. The occurrence of listed predators with their prey such as Sisyphus sp and Geometridae suggests that these insects visit T. esculentum for predation on other insects. Similarly, identified P. striatus have been shown to feed on other insects as well as nuts and legumes [70]. In addition to the identified omnivorous insects, carnivorous A. cinctipennis and P. fortipes were sampled from T. esculentum. Anthia species have been shown to occur on vegetation with their prey. The occurrence of Anthia sp with its prey that include insects from the orders Coleoptera, Isoptera, lepidoptera and Orthoptera as observed on T. esculentum in this study, is in agreement with the work of Mawdsley [71] although Anthia species were for their study sampled on grassland. There is no record of A. cinctipennis and P. fortipes on food crops, although both species are widespread with observations recorded in the Eastern Cape province as well. Although they are unable to fly, occurrence on crops that consist of a variety of their prey is possible. Insect pests that were observed causing damage to leaves, pods and seeds of T. esculentum were Z. sayi, A. janata and H. armigera. However, other pests that include L. arithmetica, S. lilligerum, C. latona, H. grandis, Libellulidae, A discoidalis, Z. elegans, L. pardalina, T. viridissima, U. pulchella and Altica sp were sampled on leaves, flowers and pods although not observed causing damage. The damage that was caused by Z. sayi to T. esculentum leaves was similar to that observed in T. fassoglense. The occurrence of Z. sayi on T. esculentum and T. fassoglense in the North West and Limpopo provinces respectively indicates that Z. sayi has affinity for marama beans although also observed in other plants in the USA. Similarly, H. armigera, although more widespread when compared to Z. sayi, has been reported causing damage to several food crops. Damage caused to T. esculentum pods and seeds, as observed in this study, was quite extensive which indicates that H. armigera is a serious pest of marama beans, although not observed on T. fassoglense. It is highly possible that larvae could attack other Tylosema species. Pods that had been infested with H. armigera spotted characteristic holes. Upon opening, H. armigera caterpillars emerged from one of the seeds while the other seed was relatively healthy looking. The caterpillar appeared to have fed on the endosperm. Similarly, sampled A. Janata were observed on stems and pods of young marama plants. No damage was observed on stems upon sampling although these caterpillars were observed scraping the surface of green T. esculentum pods which suggests that the damage to marama beans could be like that caused by H. armigera. A. Janata and H. armigera adults are strong fliers and could consequently move between crops. The lifecycle of A. Janata is like that of H. Armigera although A. Janata has been shown to prefer oilseed plants such as the castor plant [72,73]. Although this study presents the first record of A. Janata larvae and adults on a food crop in South Africa, the castor semilooper has been observed on soybean by a few researchers [74,75,76]. One of the other adult moths, U. pulchella, was observed on T. esculentum flowers as shown in Figure 3 D. The larval form of this month was not found on marama beans. U. pulchella adults are reported as pests of various plants although Heliotropium species are preferred [77]. Occurrence of U. pulchella on food crops is not shown in literature. Similarly, the moth C. latona is reported as a pest of fruit in South Africa [78], occurrence on T. esculentum could indicate that it has the potential to infest food crops although no evidence of infestation on other crops besides marama beans is shown. On the contrary, Geometridae are generally important pests with infestations on crop plants reported in China and India [79] although species could not be identified for the study. In South Africa and Nigeria, Locris sp and S. lilligerum that were observed on T. esculentum leaves were observed causing significant damage to all parts of the sorghum plant in another study [80,81]. However, the pest status of L. arithmetica which was observed on marama beans is not extensively reported in literature. Sampled Orthoptera and Libellulidae have the potential to cause severe economic loss to marama beans should the environment favour the development of gregarious forms. Ormogus sp are recognised as necrophagous scavengers [70,82] although Baena [83] showed this species as a predator of turtle eggs. No record in literature shows Ormogus species as pests or pollinators of crops. Occurrence on T. esculentum is likely for feeding on the carrion of insects that had been killed by other insect predators.
5. Conclusions
Generally, pests were the most diverse from all insect orders that were identified in marama beans although predators were also sampled. The evidence of common pests and predators is indicative of the possibility of cross infestation between T. esculentum and T. fassoglense pests. This further highlights the polyphagous nature of identified species. As much as commonality existed, the biology of some of the identified pests is known and control of their prevalence in host plants widely studied and implemented, although some may argue that the damage caused to marama beans by specific pests was quite significant and could lead to severe economic loss on marama and significant damages in other crops. On the contrary, no new control methods would need to be established given the lack of variation in feeding guilds. One may then conclude that the introduction of marama may not be detrimental to yield and productivity of maize and soybeans. Instead, the diversity of insects would increase especially since insect diversity has decreased because of climate change as reported in literature [84,85,86,87,88]. In addition to climate change negatively affecting diversity of insects, tillage practices in agrosystems as well as the intensification of agriculture may also reduce the diversity of pests, pollinators and their natural enemies [85,89]. We are inclined to agree with Outhwaite and co-workers [85] as well as Raven and Wagner [89] based on the observations of this study. The lack of disturbance as it relates to agricultural practices in marama bean could have largely contributed to the diversity of insect pests and the extent of damage to plant parts. Furthermore, the presence of listed natural enemies that were shown to occur with their prey are possible agents for the development of novel biological control strategies of pests in food crop fields. The effect of damage by insects identified in this study to yield and nutritional composition of marama beans is another avenue worth exploring. This study provides insight into the implications of marama bean introduction to the traditional subsistence agrosystem that is largely dominated by cereals such as maize, sorghum and pearl millet and pulses that include common beans, peas and more recently soybean, globally and in South Africa as well as avenues for the development of specific IPM strategies for marama beans should it be domesticated and commercialised.
Author Contributions
Conceptualization of the study, development of methods and data curation were undertaken by; A.N.M, N.L, I.ME, A.E.E.A and M.O.A. Resources, project administration and acquisition of funds; N.L.; writing—original draft preparation, A.N.M; writing—review and editing, A.N.M; N.L and I.M.E.; supervision, N.L; I.M.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The National Research Foundation (NRF) Centre of excellence for food security at the University of the Western Cape
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors would like to acknowledge and thank Jodene Foster and Adriaan Engelbrecht of the University of the Western Cape for their assistance and support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Burhan, K.; Ertek, A.; Bekir, A. Mineral nutrient content of sweet corn under deficit irrigation. Journal of Agricultural Sciences 2016, 22, 54–61. [Google Scholar]
- Messina, V. Nutritional and health benefits of dried beans. Am. J. Clin. Nutr. 2014, 100, 437S–442S. [Google Scholar] [CrossRef]
- Pragya, S.; Rita, S.R. Finger millet for food and nutritional security. African Journal of Food Science 2012. 6, 77–84.
- Omotayo, A.O.; Aremu, A.O. Marama bean [Tylosema esculentum (Burch.) A. Schreib.]: an indigenous plant with potential for food, nutrition, and economic sustainability. Food Funct. 2021, 12, 2389–2403. [Google Scholar] [CrossRef]
- Cullis, C.; Lawlor, D.W.; Chimwamurombe, P.; Bbebe, N.; Kunert, K.; Vorster, J. Development of marama bean, an orphan legume, as a crop. Food Energy Secur. 2019, 8. [Google Scholar] [CrossRef]
- Castro, S.; Silveira, P.; Coutinho, A.P.; Figueiredo, E. Systematic studies in Tylosema (Leguminosae). Bot. J. Linn. Soc. 2005, 147, 99–115. [Google Scholar] [CrossRef]
- Chimwamurombe, P.; Khulbe, R. Domestication, in Biology and breeding of food legumes. CABI Wallingford UK, 2011. p. 19-34.
- Jackson, J.C.; Duodu, K.G.; Holse, M.; de Faria, M.D.L.; Jordaan, D.; Chingwaru, W.; Hansen, A.; Cencic, A.; Kandawa-Schultz, M.; Mpotokwane, S.M.; et al. The morama bean (Tylosema esculentum): a potential crop for southern Africa. Advances in Food and Nutrition Research 2010, 61, 187–246; [Google Scholar] [PubMed]
- Hamunyela, M.H.; Nepolo, E.; Emmambux, M.N. Proximate and starch composition of marama (Tylosema esculentum) storage roots during an annual growth period. South African Journal of Science 2020, 116, 1–6. [Google Scholar]
- Powell, A.M. Marama Bean (Tylosema esculentum, Fabaceae) seed crop in Texas. Econ. Bot. 1987, 41, 216–220. [Google Scholar] [CrossRef]
- Holse, M.; Husted, S.; Hansen, Å. Chemical composition of marama bean (Tylosema esculentum)—A wild African bean with unexploited potential. J. Food Compos. Anal. 2010, 23, 648–657. [Google Scholar] [CrossRef]
- Li, J.; Cullis, C. The Multipartite Mitochondrial Genome of Marama (Tylosema esculentum). Front. Plant Sci. 2021, 12, 787443. [Google Scholar] [CrossRef]
- Brink, M. Belay, and J. De Wet, Plant resources of tropical Africa 1: Cereals and pulses. PROTA Foundation Wageningen, The Netherlands, 2006. 2006. [Google Scholar]
- Cullis, C.; Chimwamurombe, P.; Kunert, K.; Vorster, J. Perspective on the present state and future usefulness of marama bean (Tylosema esculentum). Food Energy Secur. 2023, 12, e422. [Google Scholar] [CrossRef]
- Mataranyika, P.N.; Chimwamurombe, P.M.; Fuyane, B.; Chigayo, K.; Lusilao, J. Comparative Nutritional Analysis of Tylosema esculentum (Marama Bean) Germplasm Collection in Namibia. J. Food Nutr. Sci. 2020, 8, 74. [Google Scholar] [CrossRef]
- Mosele, M.M.; Hansen, Å.S.; Engelsen, S.B.; Diaz, J.; Sørensen, I.; Ulvskov, P.; Willats, W.G.; Blennow, A.; Harholt, J. Characterisation of the arabinose-rich carbohydrate composition of immature and mature marama beans (Tylosema esculentum). Phytochemistry 2011, 72, 1466–1472. [Google Scholar] [CrossRef]
- Amonsou, E.O.; Taylor, J.R.; Beukes, M.; Minnaar, A. Composition of marama bean protein. Food Chem. 2012, 130, 638–643. [Google Scholar] [CrossRef]
- Schultz, F.; Anywar, G.; Wack, B.; Quave, C.L.; Garbe, L.-A. Ethnobotanical study of selected medicinal plants traditionally used in the rural Greater Mpigi region of Uganda. J. Ethnopharmacol. 2020, 256, 112742. [Google Scholar] [CrossRef]
- Kigen, G. , et al. Ethnopharmacological survey of the medicinal plants used in Tindiret, Nandi County, Kenya. African Journal of Traditional, Complementary and Alternative Medicines 2016, 13, 156–168. [Google Scholar]
- Otieno, V.O. Conditions for Optimum Germination of Sprawling Bauhinia Seed (Tylosema Fassoglense)(Kotschy Ex Schweinf.) Torre & Hillc. university of Nairobi, 2020.
- Kama-Kama, F.; Midiwo, J.; Nganga, J.; Maina, N.; Schiek, E.; Omosa, L.K.; Osanjo, G.; Naessens, J. Selected ethno-medicinal plants from Kenya with in vitro activity against major African livestock pathogens belonging to the “ Mycoplasma mycoides cluster”. J. Ethnopharmacol. 2016, 192, 524–534. [Google Scholar] [CrossRef]
- Mandisvika, G.; Chirisa, I.; Bandauko, E. Post-Harvest Issues: Rethinking Technology for Value-Addition in Food Security and Food Sovereignty in Zimbabwe. Adv. Food Technol. Nutr. Sci. Open J. 2015, 1, S29–S37. [Google Scholar] [CrossRef]
- Nukenine, E. Stored product protection in Africa: Past, present and future. [CrossRef]
- Southwood, T. and P. Henderson, Ecological methods / T.R.E. Southwood, P. A. Henderson. SERBIULA (sistema Librum 2.0), 2000.
- Woodcock, B.A. Pitfall trapping in ecological studies. Insect sampling in forest ecosystems, 2005. 37–57.
- Picker, M. , Field guide to insects of South Africa: Penguin Random House South Africa, 2012.
- Hugh Brier, J.W. , Kate Charleston, An identification guide for pest and beneficial insects in summer pulses, soybeans, peanuts and chickpeas, in Good bug Bad bug, H.B.a.T. Grundy, Editor. DEEDI (Primary Industries): Australia, 2012: p. 45.
- Marshall, S.A. , Insects: their natural history and diversity: with a photographic guide to insects of eastern North America. Firefly Books Buffalo, NY, 2006.
- Scholtz, C.H. and E. Holm, Insects of southern Africa: Butterworth, 1985.
- Mathenge, C.W.; Riegler, M.; Beattie, G.A.C.; Spooner-Hart, R.N.; Holford, P. Genetic variation amongst biotypes of Dactylopius tomentosus. Insect Sci. 2015, 22, 360–374. [Google Scholar] [CrossRef]
- Ware, J. and W. Megan, Termites, social cockroaches. 2022.
- Picker, M.D.; Hoffman, M.T.; Leverton, B. Density of Microhodotermes viator (Hodotermitidae) mounds in southern Africa in relation to rainfall and vegetative productivity gradients. J. Zoöl. 2006, 271, 37–44. [Google Scholar] [CrossRef]
- Edde, P.A. 5 - Arthropod pests of sugarcane (Saccharum officinarum L.), in Field Crop Arthropod Pests of Economic Importance, P.A. Edde, Editor., Academic Press. 2022, p. 276-347.
- Jouquet, P.; Chaudhary, E.; Kumar, A.R.V. Sustainable use of termite activity in agro-ecosystems with reference to earthworms. A review. A review. Agron. Sustain. Dev. 2018, 38, 3. [Google Scholar] [CrossRef]
- Macedo, N. , et al., Chapter 5 - Management of Pests and Nematodes, in Sugarcane, F. Santos, A. Borém, and C. Caldas, Editors., Academic Press: San Diego. 2015, p. 89-113.
- Miranda, C.S.; Vasconcellos, A.; Bandeira, A.G. Termites in sugar cane in Northeast Brazil: ecological aspects and pest status. Neotropical Èntomol. 2004, 33, 237–241. [Google Scholar] [CrossRef]
- Demissie, G.; Mendesil, E.; Diro, D.; Tefera, T. Effect of crop diversification and mulching on termite damage to maize in western Ethiopia. Crop. Prot. 2019, 124, 104723. [Google Scholar] [CrossRef]
- Mutsamba, E.F.; Nyagumbo, I.; Mafongoya, P. Termite prevalence and crop lodging under conservation agriculture in sub-humid Zimbabwe. Crop. Prot. 2016, 82, 60–64. [Google Scholar] [CrossRef]
- Rana, A.; Chandel, R.S.; Verma, K.S.; Joshi, M.J. Termites in Important Crops and Their Management. Indian J. Èntomol. 2021, 83, 486–504. [Google Scholar] [CrossRef]
- Robinson, William H. Urban insects and arachnids: a handbook of urban entomology. Cambridge University Press, 2005.
- Patel, S., G. Singh, and R. Singh, A checklist of global distribution of Liturgusidae and Thespidae (Mantodea: Dictyoptera). Journal of Entomology and Zoology Studies, 2016. 4(6): p.
- Symondson, W., K. Sunderland, and M. Greenstone, Can generalist predators be effective biocontrol agents? Annual review of entomology, 2002. 47(1): p. 561-594.
- Arnold, D.C. , Blister beetles of Oklahoma. 1976.
- Pierce, J.B. , Blister Beetles in Alfalfa. NM State University Cooperative Extension Service, 2014.
- Eben, A. , Ecology and evolutionary history of Diabrotica beetles—overview and update. Insects, 2022. 13(2), p.156.
- Walsh, G.C.; Avila, C.J.; Cabrera, N.; Nava, D.E.; de Sene Pinto, A.; Weber, D.C. Biology and Management of Pest Diabrotica Species in South America. Insects 2020, 11, 421. [Google Scholar] [CrossRef]
- Olaitan, A.F.; Abiodun, T.A.; Foluke, A.O.; Oluwaseyi, O.E. Comparative assessment of insect pests population densities of three selected cucurbit crops. Acta Fytotech. et Zootech. 2018, 20, 78–83. [Google Scholar] [CrossRef]
- Haber, A.I.; Pasteur, K.; Guzman, F.; Boyle, S.M.; Kuhar, T.P.; Weber, D.C. Spotted cucumber beetle (Diabrotica undecimpunctata howardi) is attracted to vittatalactone, the pheromone of striped cucumber beetle (Acalymma vittatum). J. Pest Sci. 2023, 96, 21–27. [Google Scholar] [CrossRef]
- Health, E.P.o.P. , et al., Pest categorisation of non-EU Tephritidae. EFSA Journal, 2020. 18(1): p. e05931.
- Jin, Z.; Zhao, H.; Xian, X.; Li, M.; Qi, Y.; Guo, J.; Yang, N.; Lü, Z.; Liu, W. Early warning and management of invasive crop pests under global warming: estimating the global geographical distribution patterns and ecological niche overlap of three Diabrotica beetles. Environ. Sci. Pollut. Res. 2024, 31, 13575–13590. [Google Scholar] [CrossRef]
- A Marchioro, C.; Krechemer, F.S. Potential global distribution of Diabrotica species and the risks for agricultural production. Pest Manag. Sci. 2018, 74, 2100–2109. [Google Scholar] [CrossRef]
- Chapin, J.W. and J.S. Thomas, Burrower bugs (Heteroptera: Cydnidae) in peanut: seasonal species abundance, tillage effects, grade reduction effects, insecticide efficacy, and management. Journal of economic entomology, 2003. 96(4): p. 1142-1152.
- Aigner, B.L., M. S. Crossley, and M.R. Abney, Biology and management of peanut burrower bug (Hemiptera: Cydnidae) in Southeast US Peanut. Journal of Integrated Pest Management, 2021. 12(1): p. 29.
- Monnig, N.; Bradley, K.W. Impact of fall and early spring herbicide applications on insect populations and soil conditions in no-till soybean. Crop. Prot. 2008, 27, 143–150. [Google Scholar] [CrossRef]
- Schaefer, C.W. , The food plants of some'primitive'Pentatomoidea (Hemiptera: Heteroptera). 1988. [Google Scholar]
- Fernanda-Ruiz-Cisneros, M. , et al., First Record of Feeding by Vanessa cardui Caterpillars1 on Bean Plants and Their Parasitism by Lespesia melalophae2. Southwestern Entomologist, 2020. 45(3): p. 629-638.
- Arpaia, S.; Baldacchino, F.; Bosi, S.; Burgio, G.; Errico, S.; Magarelli, R.A.; Masetti, A.; Santorsola, S. Evaluation of the potential exposure of butterflies to genetically modified maize pollen in protected areas in Italy. Insect Sci. 2018, 25, 549–561. [Google Scholar] [CrossRef]
- Stefanescu, C.; Puig-Montserrat, X.; Samraoui, B.; Izquierdo, R.; Ubach, A.; Arrizabalaga, A. Back to Africa: autumn migration of the painted lady butterfly Vanessa cardui is timed to coincide with an increase in resource availability. Ecol. Èntomol. 2017, 42, 737–747. [Google Scholar] [CrossRef]
- Hoare, R.J. , Noctuinae (Insecta: Lepidoptera: Noctuidae). Part 1, Austramathes, Cosmodes, Proteuxoa, Physetica. Fauna of New Zealand, 2017. 73.
- Forbes, R.J.; Watson, S.J.; O’connor, E.; Wescott, W.; Steinbauer, M.J. Diversity and abundance of Lepidoptera and Coleoptera in multiple-species reforestation plantings to offset emissions of carbon dioxide. Aust. For. 2019, 82, 89–106. [Google Scholar] [CrossRef]
- Rodingpuia, C.; Lalthanzara, H. An insight into black cutworm (Agrotis ipsilon): A glimpse on globally important crop pest. Sci. Vis. 2021, 21, 36–42. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, H.-A.; Kim, G.-H.; Baek, S. Effects of host plant on the development and reproduction of Agrotis ipsilon (Lepidoptera: Noctuidae) on horticultural crops. Heliyon 2023, 9, e17836. [Google Scholar] [CrossRef]
- Häuser CL, Steiner A, Bartsch D, Holstein J. On the identity of an enigmatic Delias from New Britain, Papua New Guinea: Delias mayrhoferi Bang-Haas, 1939 and Delias shunichii Morita, 1996, syn. n. Lepidoptera: Pieridae). Nachrichten Entomologischen Vereins Apollo NF. 2009;30(3):121-4.
- Bouyer, T. Démembrement et réorganisation des genres africains Jana Herrich-Schäffer, 1854 et Hoplojana Aurivillius, 1901 (Lepidoptera, Eupterotidae). Lambillionea. 2011;111(3):211-8.
- Municipality ML, District E. Biodiversity assessment. 2021.
- Staude, H.S.; Maclean, M.; Mecenero, S.; Pretorius, R.J.; Oberprieler, R.G.; Van Noort, S.; Sharp, A.; Sharp, I.; Balona, J.; Bradley, S.; et al. Noctuoidea: Erebidae: Lymantriinae, Rivulinae, Scoliopteryginae, Thiacidinae, Tinoliinae, Toxocampinae, undetermined subfamily. Metamorphosis 2022, 31, 200–219. [Google Scholar] [CrossRef]
- Pierce, W.W. A manual of dangerous insects likely to be introduced in the United States through importations. Ed. by W. Dwight Pierce, entomologist, southern field crop insect investigations. 2011.
- Tan, J.L.; Trandem, N.; Fránová, J.; Hamborg, Z.; Blystad, D.-R.; Zemek, R. Known and Potential Invertebrate Vectors of Raspberry Viruses. Viruses 2022, 14, 571. [Google Scholar] [CrossRef]
- Vappula, Niilo A. "Pests of cultivated plants in Finland." (1962).
- Gayo, L. Influence of afforestation on coleopterans abundance and diversity at the University of Dodoma, Tanzania. Environ. Sustain. Indic. 2022, 16. [Google Scholar] [CrossRef]
- Mawdsley, J.R. , et al., The genus Anthia Weber in the Republic of South Africa, Identification, distribution, biogeography, and behavior (Coleoptera, Carabidae). Zookeys, 2011(143): p. 47-81.
- Bhut, J.; Khanpara, D.; Bharadiya, A.; Madariya, R. Bio-Efficacy of Chemical Insecticides Against Defoliators Spodoptera litura and Achaea janata in Castor. J. Phytopharm. 2022, 11, 368–370. [Google Scholar] [CrossRef]
- Jeyasankar, A. , et al., Feeding and growth inhibition activities of Tragiain volucrata Linn (Euphorbiaceae) on Achaea janata (Linn.)(Noctuidae: Lepidoptera) and Pericallia ricini (Fab.)(Lepidoptera: Arctidae). Open Access Library Journal, 2014. 1(4): p. 1-7.
- Phartale, N.; Bhede, B.; Patait, D.; Kadam, T.; Gyananath, G. Pest control potential of four predatory spiders from soybean fields. J. Èntomol. Res. 2019, 43, 367. [Google Scholar] [CrossRef]
- Bhamare, V. , et al., Impact of abiotic factors on population dynamics of lepidopteran insect-pests infesting sole soybean and soybean intercropped with pigeonpea. Journal of Entomology and Zoology studies, 2018. 6(5): p. 430-436.
- Dhurgude, S. , et al., Key mortality factors of semilooper Achae janata (Linn) infesting soybean. Journal of Entomological Research, 2014. 38(2): p. 125-128.
- Bolu, H. , et al., A new host record for Exorista xanthaspis (Wiedemann, 1830) (Diptera: Tachinidae) from Turkey. Journal of the Entomological Research Society, 2019. 21(3): p. 373-378.
- Kriegler, P. , Notes on the occurrence of fruit-sucking moths on deciduous fruits in the winter rainfall region. South African Journal of Agricultural Science, 1958. 1(3): p. 245-247.
- Roy, S.; Barooah, A.K.; Ahmed, K.Z.; Baruah, R.D.; Prasad, A.K.; Mukhopadhyay, A. Impact of climate change on tea pest status in northeast India and effective plans for mitigation. Ecol. Front. 2020, 40, 432–442. [Google Scholar] [CrossRef]
- Jayi, O. and F.A. Oboite, Importance of spittle bugs, Locris rubens (Erichson) and Poophilus costalis (Walker) on sorghum in West and Central Africa, with emphasis on Nigeria. Annals of Applied Biology, 2000. 136(1): p. 9-14.
- Kruger, M.; Berg, J.v.D.; Du Plessis, H. Diversity and seasonal abundance of sorghum panicle-feeding Hemiptera in South Africa. Crop. Prot. 2008, 27, 444–451. [Google Scholar] [CrossRef]
- Waseem, M.; Kumar, S.; Sultana, R. A New Species of the Sub-Genus Afromorgus (Trogidae: Scarabaeoidea) from Cholistan Desert, Pakistan. Pak. J. Zoöl. 2023, 57. [Google Scholar] [CrossRef]
- Baena, M.L.; Escobar, F.; Halffter, G.; García–Chávez, J.H. Distribution and Feeding Behavior of Omorgus suberosus (Coleoptera: Trogidae) in Lepidochelys olivacea Turtle Nests. PLOS ONE 2015, 10, e0139538. [Google Scholar] [CrossRef]
- Harvey, J.A.; Tougeron, K.; Gols, R.; Heinen, R.; Abarca, M.; Abram, P.K.; Basset, Y.; Berg, M.; Boggs, C.; Brodeur, J.; et al. Scientists' warning on climate change and insects. Ecol. Monogr. 2023, 93. [Google Scholar] [CrossRef]
- Outhwaite, C.L.; McCann, P.; Newbold, T. Agriculture and climate change are reshaping insect biodiversity worldwide. Nature 2022, 605, 97–102. [Google Scholar] [CrossRef]
- Peace, N. , Impact of climate change on insects, pest, diseases and animal biodiversity. International Journal of Environmental Sciences & Natural Resources, 2020. 23(5): p. 151-153.
- Wilson, R.J.; Fox, R. Insect responses to global change offer signposts for biodiversity and conservation. Ecol. Èntomol. 2021; 46, 699–717. [Google Scholar] [CrossRef]
- Pareek, A. , et al., Impact of climate change on insect pests and their management strategies. Climate change and sustainable agriculture, 2017: p. 253-286.
- Raven, P.H.; Wagner, D.L. Agricultural intensification and climate change are rapidly decreasing insect biodiversity. Proc. Natl. Acad. Sci. 2021, 118, e2002548117. [Google Scholar] [CrossRef]
Figure 1.
A map of South Africa that shows study sites for insect sampling on T. esculentum and T. fassoglense.
Figure 1.
A map of South Africa that shows study sites for insect sampling on T. esculentum and T. fassoglense.
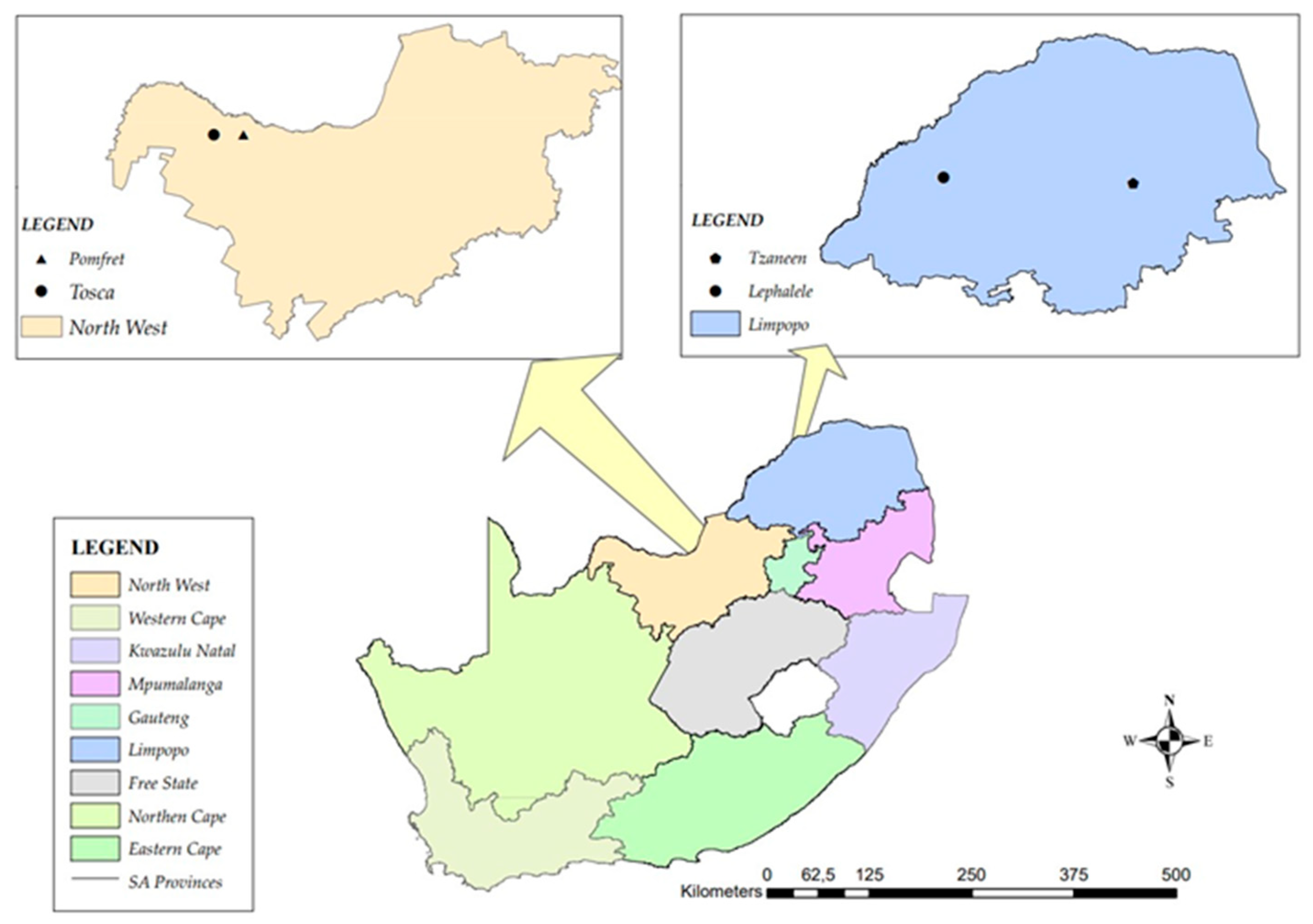
Figure 2.
Insect pests associated with T. fassoglense. Microhodotermes viator Latreille (Isoptera-Hodotermidae)-A, Zonitis sayi Wickham (Coleoptera: Meloidae)-B, Lycus sp Guérin-Méneville (Coleoptera: Lycidae)-C, Janomima mariana White (Lepidoptera: Lasiocampidae)-D, Undetermined 1 (Lepidoptera)-E, Proteuxoa sp Hampson (Lepidoptera: Noctuidea)-F, Vanessa cardui Linnaeus (Lepidoptera: Nymphalidae)-G, Zonocerus elegans Thunberg (Orthoptera: Pyrgomorphidae)-H, Hoplocoryphella grandis Brancsik (Mantidae: Thespidae)-I and Undetermined 2 & Undetermined 3 (Orthoptera: Acrididae)-J & K.
Figure 2.
Insect pests associated with T. fassoglense. Microhodotermes viator Latreille (Isoptera-Hodotermidae)-A, Zonitis sayi Wickham (Coleoptera: Meloidae)-B, Lycus sp Guérin-Méneville (Coleoptera: Lycidae)-C, Janomima mariana White (Lepidoptera: Lasiocampidae)-D, Undetermined 1 (Lepidoptera)-E, Proteuxoa sp Hampson (Lepidoptera: Noctuidea)-F, Vanessa cardui Linnaeus (Lepidoptera: Nymphalidae)-G, Zonocerus elegans Thunberg (Orthoptera: Pyrgomorphidae)-H, Hoplocoryphella grandis Brancsik (Mantidae: Thespidae)-I and Undetermined 2 & Undetermined 3 (Orthoptera: Acrididae)-J & K.

Figure 3.
Insect pests associated with T. esculentum. Altica sp Geofrroy (Coleoptera: Chrysomelidae)-A, Locris arithmetica Walker (Hemiptera: Cercopidae)-B, Achaea Janata Linnaeus (Lepidoptera: Erebidae) adult and larva-C, Utetheisa pulchella Linnaeus (lepidoptera: Noctuidea)-D, Helicoverpa armigera Hübner (Lepidoptera: Noctuidea) larva-E.
Figure 3.
Insect pests associated with T. esculentum. Altica sp Geofrroy (Coleoptera: Chrysomelidae)-A, Locris arithmetica Walker (Hemiptera: Cercopidae)-B, Achaea Janata Linnaeus (Lepidoptera: Erebidae) adult and larva-C, Utetheisa pulchella Linnaeus (lepidoptera: Noctuidea)-D, Helicoverpa armigera Hübner (Lepidoptera: Noctuidea) larva-E.

Figure 4.
Insect pests associated with T. esculentum. Tettigonia viridissima Linnaeus (Orthoptera: Tettigoniidae) nymph-F and adult-G, Undetermined 6 (Lepidoptera)-H and Acanthoplus discoidalis Walker (Orthoptera: Bradyporidae)-I. Undetermined 7-J, Undetermined 8-K.
Figure 4.
Insect pests associated with T. esculentum. Tettigonia viridissima Linnaeus (Orthoptera: Tettigoniidae) nymph-F and adult-G, Undetermined 6 (Lepidoptera)-H and Acanthoplus discoidalis Walker (Orthoptera: Bradyporidae)-I. Undetermined 7-J, Undetermined 8-K.


Table 1.
This table shows insect pests that were sampled from T. fassoglense at Lephalale and Tzaneen in the Limpopo province of South Africa.
Table 1.
This table shows insect pests that were sampled from T. fassoglense at Lephalale and Tzaneen in the Limpopo province of South Africa.
| Order | Family | Species | Feeding guild |
| Blattodea | Blattidae | Deropeltis erythrocephala | Omnivorous scavengers |
| Isoptera | Hodotermidae Termidae |
Microhodotermes viator Amitermes hastatus |
Detritivores |
| Coleoptera | Chrysomelidae | Diabrotica undecimpunctata | Herbivorous pests |
| Meloidea | Zonitis sayi | ||
| Lycidae Scarabaeidae |
Lycus sp Proatetia brevitarsis Maladera drescheri |
||
| Hemiptera | Cydnidae | Sehirus cinctus | Herbivorous pests |
| Pentatomidae | Nezara viridula | ||
| Eupterodidae | Tantaliana tantalus | Unknown | |
| Lepidoptera | Noctuidae | Proteuxoa sp | Herbivorous pest |
| Agrotis ipsilon | Herbivorous pest | ||
| Lasiocampidae | Macrothylacia rubi | Herbivorous pest | |
| Janomima mariana | Unknown | ||
| Nymphalidae | Vanessa cardui | Pest | |
| Sphingidae | Theretra oldenlandiae | Herbivorous pest | |
| Undetermined 1 | Pests | ||
| Mantodea | Thespidae | Hoplocoryphella grandis | Carnivorous predator |
| Orthoptera | Pyrgomorphidae | Zonocerus elegans | Herbivorous pests |
| Acrididae | Undetermined 2 & Undetermined 3 | Herbivorous pests | |
Table 2.
This table shows insect pests that were sampled from T. esculentum at Tosca and Pomfret in the North West province of South Africa.
Table 2.
This table shows insect pests that were sampled from T. esculentum at Tosca and Pomfret in the North West province of South Africa.
| Order | Family | Species | Feeding guild | |
| Blattodea | Blattidae | Undetermined 4 | Omnivorous scavengers | |
| Coleoptera | Chrysomelidae |
Altica sp Lema rufotestacea |
Herbivorous pests | |
| Meloidea | Zonitis sayi | |||
| Erotylidae | Amblyopus sp | Unknown | ||
| Scarabaeidae | Sisyphus sp | Coprophagous | ||
| Trogidae | Omorgus asperulatus | Necrophagous | ||
| Tenebrionidae |
Gonopus tibialis Psammodes bertelonii Psammodes striatus |
Herbivorous pest | ||
| Unknown | ||||
| Herbivorous pest | ||||
| Carabidae |
Anthia cinctipennis Passalidius fortipes |
Predators | ||
| Phalacridae | Olibrus sp | Herbivorous pests | ||
| Hemiptera | Cercopidae | Locris arithmetica | Herbivorous pests | |
| Scutelleridae | Solenostethium liligerum | |||
| Lepidoptera | Erebidae |
Achaea Janata Utetheisa pulchella |
Herbivorous pests | |
| Noctuidae |
Helicoverpa armigera Cyligramma latona Undetermined 5 |
|||
| Geometridae |
Undetermined 6 Undetermined 7 & 8 |
|||
| Odonata | Libellulidae | Undetermined 9 | Carnivorous pest | |
| Orthoptera | Pyrgomorphidae Acrididae Bradyporidae |
Zonocerus elegans Undetermined 10 Locustana pardalina Acanthoplusdiscoidalis |
Herbivorous pests | |
| Haematophagous | ||||
| Omnivorous pests | ||||
| Tettigoniidae | Tettigonia viridissima | Herbivorous pests | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



