
Submitted:
21 March 2025
Posted:
21 March 2025
You are already at the latest version
Abstract
Xiangwanxian 13 (XWX13), a fragrant indica rice variety with high yield and good grain quality, is generally welcomed by farmers in China. However, the fact that XWX13 is highly susceptible to rice blast and its high grain cadmium (Cd) accumulation in Cd polluted paddy fields has been limiting its commercial utilization. Therefore, a genomics-assisted selection approach was employed to improve its blast resistance and lower grain Cd accumulation. In this study, four major genes, namely Pi1, Pi2, OsHMA3 and OsNramp5, were precisely introduced into XWX13 with the KASP marker-assisted selection followed by evaluation of genomic background with the genome-wide RICE 1K SNP array. Two preferable improved lines iXWX13-1 (stacking Pi1+Pi2+OsHMA3) and iXWX13-2 (stacking Pi1+Pi2+OsNramp5) were obtained with a genomic background recovery rates of 94.44% and 94.63%, respectively. Resistance spectrum assay showed that the two improved lines at the seedling stage exhibited high-level resistance to 39 Magnaporthe oryzae isolates, with more than 97.44% blast resistance frequency. Also, an enhanced resistance was observed for leaf and panicle neck blast in the two improved lines than that of the original XWX13 in natural blast nurseries. For grain Cd content, a multi-site field trial showed that a trace amount of grain Cd accumulation (0.009 to 0.077 mg/kg) was detected in the iXWX13-2, 90.98%-98.87% lower than that of XWX13. Moreover, the high similarity was observed in the yield, main agronomic traits and grain quality when comparing the improved lines to the original XWX13. This work was the first successful practice to apply molecular marker assisted backcrossing with genomics-based background selection to improve the blast resistance and lower grain Cd accumulation of XWX13 without penalty of the yield, main agronomic traits and grain quality. This proposed strategy would pave the way to integration of multiple desirable genes into the elite varieties in crop genetic improvement.
Keywords:
rice
; genomics-assisted improvement
; blast resistance
; low cadmium accumulation
; yield and agronomic traits
1. Introduction
Rice (Oryza sativa L.) is one of the most important crops with food supply for more than half of the world's population [1]. To ensure food security, efforts have still been made for a sustainable increase in rice yield in the last decade [2]. However, high-yield cultivation of rice varieties has caused excessive use of pesticides due to lack of resistance to pests and diseases, which increased input costs and damaged the ecological environment [3,4,5]. In addition, industrial emissions, agro-chemical use, and anthropogenic activities have resulted in excessive heavy metals such as cadmium (Cd) in rice paddies, posing a serious threat to food safety [6,7,8]. Therefore, it is necessary to develop rice varieties with superior resistance and tolerance to major biotic and abiotic stresses.
Rice blast, caused by pathogenic fungus Magnaporthe oryzae (M. oryzae), is one of the major diseases of rice resulting in yield loss from 30% to 50% in large rice planting areas under favorable conditions [3]. Great efforts have been devoted to evaluating and characterizing blast resistance of rice germplasm resources. To date, more than 100 blast resistance (R) genes have been identified and mapped on rice genome, 38 of them have been cloned and characterized [9,10]. Most of the genes are distributed in the gene clusters on chromosomes 6, 11 and 12, and very few of them were introgressed into popular rice varieties [9]. The high variable nature of the pathogen M. oryzae leads to frequent emergence of new virulent races resulting in loss of resistance within 3-5 years of rice cultivation. Among the blast resistance genes, Pi1 is an allele at the Pik locus mapped at the end of chromosome 11, conferring dominant and durable resistance against geographically diverse M. oryzae isolates [11,12]. Likewise, gene Pi2 is an allele at the Piz locus located on chromosome 6 close to the centromere, and also confers dominant and broad-spectrum resistance to various M. oryzae isolates [13,14,15,16]. Currently, several attempts had been made to deploy Pi1 and Pi2 in rice breeding [17,18,19,20], along with enhanced resistance to rice blast under high pressure in epidemic fields.
Among abiotic stresses, Cd contamination of soil poses a serious threat to sustainable agriculture and food safety. As the main source of Cd intake in human dietary [21], rice generally poses a higher risk of Cd exposure compared to other crops [22,23]. To reduce the effects of Cd toxicity on human health, a maximum Cd concentration of 0.4 mg/kg in polished rice is allowed internationally, and even a stricter limit of 0.2 mg/kg was established in China [24,25]. However, cases of excessive Cd accumulation in rice grain have occurred frequently in the past decade [26,27,28,29]. Some measures have been made to reduce the risk of excessive Cd accumulation in rice, such as soil-Cd remediation, fertilizer control, irrigation management, and growing low-Cd-accumulating varieties [25]. Studies have shown that genetic improvement and development of low-Cd rice varieties are cost-effective in agricultural practice [22,30]. Currently, the physiological and genetic mechanisms of Cd accumulation in rice have been well studied, some quantitative trait loci (QTL) and genes associated with variation in grain Cd accumulation have been identified [31,32], and four Cd-QTL hotspots in genomic regions have been mapped on chromosomes 2, 3, 7, and 8 [33,34,35,36,37,38,39,40,41,42,43,44,45,46,47]. With a series of the associated QTLs and genes being found, genetic improvements in rice specifically targeting at reducing grain Cd accumulation have become a top priority for rice breeders. Among the known cases, the natural resistance associated macrophage protein gene OsNramp5, located on Chromosome 7 and encoding a key transporter for Cd uptake in rice, is an ideal target gene to be manipulated to reduce Cd uptake [48,49,50,51]. Additionally, OsHMA3, encoding the P-type heavy metal ATPase, is another target gene that has been manipulated to sequester Cd into vacuoles of root cells, which subsequently reduced Cd transportation [52,53]. The use of OsNramp5 and OsHMA3 to generate low-Cd rice varieties were more effective when compared with genetic modification of genes such as OsNramp1, OsZIP5, OsZIP9, OsZIP7, and OsLCT1 [47].
Molecular breeding can not only improve target traits of recipient materials, but also develop new varieties with excellent performance by pyramiding different traits, which is more accurate and efficient than traditional breeding. Marker-assisted selection (MAS) is regarded as a highly efficient breeding method because of the rapid and precise selection of the desired genes [54]. Application of MAS has significantly improved the rice breeding process, although with some disadvantages such as linkage drag and unknown genetic background effects [19]. Furthermore, genomic technology has been now widely used for improving breeding efficiency by foreground and background selection with MAS [55]. Based on large numbers of informative markers, foreground selection chooses individuals carrying the target genes [56], while background selection is used to improve the genomic recovery of the recipient parent [57]. The kompetitive allele specific PCR (KASP) markers have considerably enhanced crop improvement and variety development with more efficient and precise genotyping [58]. In addition, single nucleotide polymorphism (SNP) arrays based on the Affymetrix and Illumina technologies have been developed and used for background selection applied in rice [59], maize [60], and other crops [61,62,63]. Recently, new SNP panels have also been developed by using Genotyping by Pinpoint Sequencing of multiplex PCR products (mGPS) and Genotyping by Pinpoint Sequencing of captured targets (cGPS) technologies (https://www.higentec.com). To date, a series of rice SNP arrays with different marker numbers and genomic coverage have been developed and used for MAS [64], such as the 44K SNP chip, RICE6K array, 50K SNP chip, Rice700K SNP array, Cornell 7K Array Infinium Rice (C7AIR), Rice3K56 SNP array, and so on [65,66,67,68,69,70]. Therefore, all these advancements in genomic technologies can provide more powerful selection of plant traits, and facilitate comprehending and improving various crops.
Xiangwanxian 13 (XWX13) is a fragrant indica rice variety with a late-maturing characteristic in China. It is highly welcomed by farmers due to the high yield and good grain quality, and has been commercially cultivated over 600,000 hectares in total. However, XWX13 is highly susceptible to rice blast, and the grain Cd concentration in high pollution paddy fields still exceeded the Cd-safe standard of 0.2 mg/kg though it was ever an emergency variety for low Cd [71], indicating a production risk and limit for large-scale commercial utilization. In the present study, an efficient genomic breeding strategy was adopted for enhancing blast resistance and lowering grain Cd accumulation in XWX13. Four major genes, namely Pi1, Pi2, OsHMA3, and OsNramp5, were precisely introduced into XWX13 from three donor parents with aid of genomics-assisted selection. Our results provide a successful breeding practice in genomics-assisted rapid and directional improvement of XWX13 for rice blast resistance and low Cd accumulation.
2. Materials and Methods
2.1. Plant Materials
Rice plant materials used in this study include the original parent Xiangwanxian 13 (XWX13, a high-quality indica rice cultivar with fragrance), blast resistance gene donor HZ02411 (Pi1+Pi2), low-Cd accumulation gene donors HZ02415 (OsHMA3) and HZ02416 (OsNramp5). The rice variety CO39 was used as susceptible control, while Gumei4 was used as resistant control.
2.2. Population Development and Breeding Selection Procedure
The improved XWX13 lines (iXWX13-1 and iXWX13-2) were developed by marker-assisted backcrossing as described in Figure 1. The late-maturing indica rice cultivar XWX13 was crossed with each of three different donors, and the foreground SNP markers were used to genotype XWX13, the corresponding donors, and the F1 plants. The F1 plants were then backcrossed with the recurrent parent XWX13. The BC1F1 plants were genotyped for foreground and background selections, and three plants with the target genes and the highest background recovery rate were selected to further backcross with XWX13. The same genotyping and selections were done for the BC2F1 and BC3F1 populations. To pyramid genes for different traits, the BC3F1 plants with the target genes and the highest background recovery rate were used for multiple hybridization, and double-cross F1 population was generated. Based on continuous self-pollination, foreground and background selections, and field phenotypic screening, two improved lines iXWX13-1 (Pi1+Pi2+OsHMA3) and iXWX13-2 (Pi1+Pi2+OsNramp5) were successfully developed.
2.3. Marker-Assisted Selection and Background Analysis
SNP markers tightly linked to the genes, Pi1, Pi2, OsHMA3, and OsNramp5, were used to track the target genes in foreground selection. And 120 polymorphic SNP markers evenly distributed on 12 chromosomes were used for background selection. KASP genotyping was performed as described by He et al. [20].
The whole-genome SNP array RICE 1K mGPS was further used for detecting the genetic background between the recurrent parent XWX13 and the improved lines, iXWX13-1 and iXWX13-2. The SNP array RICE 1K mGPS was developed by Huazhi Biotechnology Co., Ltd., and comprises 5,456 loci in 1,048 intervals uniformly distributed on the rice genome. According to the similar methods previously reported [20], SNP array genotyping was performed and the background recovery rate was calculated.
2.4. Evaluation of Rice Blast Resistance
XWX13 and the improved lines were tested for resistance spectrum at the seedling stage during 2022-2023 in Huazhi Biotechnology Co., Ltd., Changsha, China. The 39 M. oryzae isolates were used for spraying inoculation in the greenhouse according to the method described by Liu et al. [72]. Disease reaction was assessed one week after inoculation following a standard 0-9 rating evaluation system [73,74]. The rice varieties CO39 and Gumei4 were used as the susceptible and resistant control, respectively. Disease evaluation was performed in three replications. Resistance frequency was defined as ratio of resistance-showing M. oryzae isolates to the total number of inoculated M. oryzae isolates on the rice genotype (line), and a higher frequency represents a broader resistance spectrum.
2.5. Evaluation of Yield, Main Agronomic Traits and Grain Quality
XWX13 and the improved lines were planted in multi-site trials in 2022. Five test sites were set up, namely Changsha and Yueyang in Hunan, Yichun and Shangrao in Jiangxi, and Hezhou in Guangxi. In the field layout, each variety or line was planted in a plot of 13.34 m2. The field cultivation management was carried out according to the method of China National Rice Variety Regional Trial. The heading date was recorded for each plot. At maturity stage, ten individual plants in the middle of the central row in each plot were sampled for measurements of main agronomic traits including plant height (PH, cm), panicle length (PL, cm), spikelets per panicle (SPP), filled-grain percentage (FGP, %), and 1000-grain weight (GW, g). The plot yield was measured for each variety or line at each site.
Bulked harvested seeds from each plot were used for grain quality analysis, including brown rice percentage (BRP, %), milled rice percentage (MRP, %), head rice percentage (HRP, %), chalky rice percentage (CRP, %), chalkiness degree (CD, %), rice grain length (RGL, mm), grain length/width ratio (L/W), alkali spreading value (ASV), amylose content (AC, %), gel consistency (GC, mm).
The single factor ANOVA in the software SPSS Statistics 19 was used to test the significance of variation for yield, main agronomic traits and grain quality.
2.6. Determination of the Grain Cd Concentration
The grain Cd concentration of XWX13 and the improved lines in multi-site trials were determined by using NX-100FA food heavy metal detector (NCS Testing Technology Co., Ltd., Suzhou, Jiangsu, China). Bulked harvested seeds from each plot were randomly sampled three times, and measured for the grain Cd concentration following the instrument manual. The values of grain Cd concentration given are mean of three replications. Differences of the grain Cd concentration between XWX13 and the improved lines were statistically tested by the single factor ANOVA in SPSS Statistics 19.
3. Results
3.1. Development of the Improved Lines
Following the breeding procedure shown in Figure 1, the four resistance genes were respectively transferred into the XWX13 background from HZ02411 (Pi1+Pi2), HZ02415 (OsHMA3), and HZ02416 (OsNramp5). Three single-cross BC3F1 populations were developed by marker-assisted foreground and background selections, and then two double-cross F1 populations were generated by multiple hybridization. After four generations of selfing screening and identification, two improved lines iXWX13-1 (Pi1+Pi2+OsHMA3) and iXWX13-2 (Pi1+Pi2+OsNramp5) were obtained.
The two improved lines, iXWX13-1 and iXWX13-2, and the original recipient parent XWX13 were further genotyped for genetic background analysis using SNP array RICE 1K mGPS. It was found that the genetic background recovery rates of iXWX13-1 and iXWX13-2 were 94.44% and 94.63%, indicating a high genomic similarity with XWX13 (Figure 2 and Figure 3).
3.2. Blast Resistance of the Improved Lines
The two improved lines, iXWX13-1 and iXWX13-2, the recurrent parent (XWX13), CO39 (susceptible control), and Gumei4 (resistant control) were scored at the seedling stage by spraying inoculation with 39 M. oryzae isolates in the greenhouse (Table 1), and the typical symptoms of two strains on five varieties or lines were shown in Figure 4. The two improved lines, iXWX13-1 and iXWX13-2, and the resistant control Gumei4 exhibited broad-spectrum resistance to rice blast, with a high resistance frequency of 97.44%, 100%, and 92.31%, respectively, whereas the recipient variety XWX13 and the susceptible control CO39 showed a low resistance frequency of 17.95% (Table 1). The results verified that the enhanced blast resistance is occurring in the two improved lines (possessing Pi1+Pi2).
The increased resistance of the two improved lines (iXWX13-1and iXWX13-2) were also observed under two natural blast nurseries in comparison to the recurrent parent (XWX13). In detail, XWX13 showed 7-8 level of leaf blast (S-HS) and 15-18% of panicle neck blast infection rate (MS) in the blast nursery of Taojiang County in 2022, and thus was susceptible to rice blast (Table 2, Figure 5). While the two improved lines (possessing Pi1+Pi2) exhibited a higher level resistance reaction, both the iXWX13-1 and iXWX13-2 showed grade 2 of leaf blast (R) and 4-5% of panicle neck blast infection rate (R). In the same year, iXWX13-1 and iXWX13-2 demonstrated resistance to leaf blast (0-1 grade) in the blast nursery of Dawei Mountain, whereas XWX13 showed grade 7 of leaf blast (S) (Table 2). In short, the resistance to leaf and panicle neck blast under natural conditions were also effectively enhanced in the two improved lines (possessing Pi1+Pi2).
3.3. Yield, Agronomic Traits and Grain Quality of the Improved Lines
To test whether the traits of the two improved lines,iXWX13-1 and iXWX13-2, were identical to that of the original recipient parent XWX13, the main agronomic traits of the lines were observed in a multi-site trial in 2022. Minor differences were found for a few traits, including days to maturity (DTM), plant height (PH) and 1000-grain weight (GW) in overall. For example, the DTM was slightly longer (123 d vs. 120.4 d) for the two lines than XWX13, and the GW of iXWX13-1and iXWX13-2 were subtle higher (0.6-0.8 g) than that of XWX13 together with a decrease in plant height (2-3 cm) of iXWX13-1and iXWX13-2 (Table 3). In general, no significant difference was observed between the two improved lines and XWX13 for most of the agronomic traits. These results indicated that the main agronomic traits of the improved lines are highly similar to that of XWX13.
The grain yield of these tested lines in the multi-site trial was investigated. The average yields of XWX13, iXWX13-1, and iXWX13-2 from five test sites were 7.50 t/ha, 7.37 t/ha, and 7.51 t/ha, respectively (Table 3). Non-significant differences were found between XWX13 and the improved lines. In term of each test site, the yield of iXWX13-2 was slightly higher than that of iXWX13-1, but the difference was not statistically significant as indicated by the single factor ANOVA test (Table 3).
To profile grain quality, rice grains from the five test sites were investigated for ten important rice quality traits with significant difference only in grain shape (Table 4). For iXWX13-1 and iXWX13-2, grains were shorter (0.2-0.3 mm) than that of XWX13, thus resulting in relatively round grains. The other eight rice quality traits, BRP, MRP, HRP, CRP, CD, ASV, AC, and GC, showed statistically non-significant differences between XWX13 and the improved lines, revealing highly similar grain quality.
3.4. Grain Cd Accumulation in the Improved Lines
To evaluate the grain Cd content under Cd-polluted conditions, the grain Cd concentration of all the lines in the multi-site trial were examined and showed that Cd was hardly accumulated into the grains of iXWX13-2. Across five test sites, the grain Cd concentration in iXWX13-2 ranged from 0.009 to 0.077 mg/kg, significantly lower than that in XWX13 (0.618-1.052 mg/kg) and iXWX13-1 (0.569-0.864 mg/kg) (Figure 6). Compared with the original recipient parent XWX13, the grain Cd concentration in iXWX13-2 decreased by 90.98-98.87%. Regarding to grain Cd accumulation between XWX13 and iXWX13-1, significant differences were only detected in two sites, Shangrao and Hezhou, where the grain Cd concentration in iXWX13-1 were both relatively lower than that in XWX13 (Figure 6).
Of all tested samples, only the grain Cd concentration in iXWX13-2 (possessing OsNramp5) in each plot was obviously lower than the limit value of 0.2 mg/kg in China National Standard for Food Safety (GB 2762-2022), revealing that iXWX13-2 is a stable Cd-safe improved line and superior to iXWX13-1 (possessing OsHMA3). Therefore, iXWX13-2 (stacking Pi1+Pi2+OsNramp5) demonstrated the enhanced blast resistance and low Cd accumulation without penalty of the yield, main agronomic traits and grain quality.
4. Discussion
Improvement Achieved by Genomic Marker-Assisted Selection
The phenotype-based selection in conventional breeding has played a great role in developing elite rice varieties in the past decades, although with a long breeding cycle and low efficiency [75,76]. Alternatively, genomic marker-assisted breeding is an innovative approach that utilizes modern molecular tools and genomic information to improve the accuracy and efficiency of plant breeding [58], and it has been successfully applied to improve biotic or abiotic stresses tolerance in rice varieties [20,47,77,78,79,80]. Also high-throughput sequencing and genotyping platforms have accelerated the process of genomic marker-assisted breeding [58]. In this study, an efficient genomic breeding strategy for the integration of blast resistance and low Cd accumulation was adopted by the genomic marker-assisted foreground and background selection. Four major genes, namely Pi1, Pi2, OsHMA3, and OsNramp5 from three donors, were precisely introduced into the genetic background of a good quality fragrant rice variety XWX13 by using genomic marker-assisted selection. Eventually two preferable improved lines iXWX13-1 (Pi1+Pi2+OsHMA3) and iXWX13-2 (Pi1+Pi2+OsNramp5) were obtained with XWX13 background recovery rates of 94.44% and 94.63%. Compared to the traditional 6-8 year breeding cycle, our strategy shortens the cycle up to 2-3 years as reported in the previous studies [19,20]. Therefore, this study demonstrated a successful breeding practice in genomics-assisted rapid improvement of XWX13 for blast resistance and low Cd accumulation.
Pyramiding Genes to Improve Blast Resistance and Cd Accumulation
Blast resistance is easily lost after a few years of cultivation due to variation and evolution of pathogen populations. For instance, Tang et al. [81] found elite indica rice varieties from South China with Pi1 were not resistant to blast, while those with Pi2 exhibited 83.3% resistance frequencies. Similarly, it was found that multi-gene pyramiding lines showed significantly higher resistance levels than monogenic lines [82]. So there is an urgent need for pyramiding multiple genes in breeding programs [83,84], and pyramiding of multiple resistance genes has proven to be an effective way for developing rice varieties with durable resistance to M. oryzae [20,55]. As found in our study, the two improved lines iXWX13-1 and iXWX13-2 carrying Pi1+Pi2 exhibited more than 97.44% blast resistance frequency under artificial inoculation at the seedling stage, indicating an obviously increased resistance spectrum compared with that of the original recipient variety XWX13. Furthermore, the two improved lines iXWX13-1 and iXWX13-2 showed a grade 2 of leaf blast (R) and 4-5% of panicle neck blast infection rate (R) in blast nursery in Taojiang County, and thus possessed the enhanced resistance to leaf and panicle neck blast. The increased resistance to leaf blast were also observed in blast nursery in Dawei Mountain. These results revealed that it is feasible to pyramid multiple broad-spectrum R genes into elite rice varieties, enhancing their resistance and durability against blast disease.
The grain Cd accumulation poses serious safety concern for the rice industry. To date, various approaches, including water management and administration of contaminated soil additives, have been used to alleviate Cd accumulation in rice grain [85,86]. Correspondingly, some low-Cd rice varieties have been identified and developed through mass selection, marker-assisted selection, and genetic manipulation [39,47]. Hu et al. [80] confirmed that gene OsNramp5 was the most efficient in lowering Cd accumulation, and first generated low Cd accumulation restorer (R) lines by editing OsNramp5, OsLCD, and OsLCT1 in both the japonica and indica rice. In the present study, two improved lines iXWX13-1 (Pi1+Pi2+OsHMA3) and iXWX13-2 (Pi1+Pi2+OsNramp5) were obtained by genomic marker-assisted selection. The evaluation of Cd accumulation in a multi-site trial showed that the grain Cd concentration in iXWX13-2 carrying OsNramp5 decreased by 90.98-98.87% compared with that in the original recipient variety XWX13, which was far below the threshold value of 0.2 mg/kg in China (GB 2762-2022), demonstrating a non-transgenic Cd-safe improved line, and also reconfirmed the stable efficiency of OsNramp5 in lowering Cd concentration [47,80]. Here, statistically significant differences of the grain Cd concentration between XWX13 and iXWX13-1 were observed in two test sites (Shangrao and Hezhou), but not in the other three sites (Changsha, Yueyang, and Yichun), and both of them exceeded the safe threshold value of 0.2 mg/kg, indicating that the improved line iXWX13-1 carrying OsHMA3 had no obvious effect on lowering Cd accumulation. The gene OsHMA3 is not as effective in lowering Cd accumulation as previously reported [47,87,88]. As a typical quantitative trait, the variations of grain Cd accumulation in rice are determined by both the genotypes and the environmental factors that determine Cd phyto-availability in soil [28,89,90], which make it difficult to accurately evaluate the genetic effects of grain-Cd variation [91]. It is usually accepted that there is natural variation in Cd accumulation among different rice varieties [92]. Previous studies have indicated that the indica rice varieties generally accumulate more Cd comparing to the japonica rice varieties [93]. However, a consistent conclusion has not been obtained by comparing Cd accumulation in the same genetic background with different Cd-related genes. In our study, the two improved lines iXWX13-2 and iXWX13-1 have the similar genomic background of XWX13 (Figure 2 and Figure 3), and thus OsNramp5 is obviously more effective in lowering grain Cd accumulation than OsHMA3 in the same genetic background (Figure 6). The performance of the two genes in other genetic backgrounds needs to be investigated in the upcoming further studies.
Improvement of Traits Without Penalty of the Yield, Main Agronomic Traits and Grain Quality
Plants face the constant challenge of allocating limited resources to immunity and growth to ensure survival and reproduction, therefore, one major aim of crop breeding is to balance biotic resistance and abiotic tolerance with yield [94]. The genomics-assisted breeding strategy can provide more powerful selection of quantitative traits, and modify tradeoffs effectively. In this study, the characterization of the two improved lines iXWX13-1 and iXWX13-2 showed that their yield and main agronomic traits were almost identical to those of the recurrent parent XWX13, and the average values for each trait in a multi-site trail were not significantly different. In term of rice quality, eight traits (brown rice percentage, milled rice percentage, head rice percentage, chalky rice percentage, chalkiness degree, alkali spreading value, amylose content, and gel consistency) showed no statistically significant difference between XWX13 and the improved lines. In view of the potential tradeoff between resistance and yield, multiple traits should be paid attentions when screening the target plants selected based on the genomic marker-assisted foreground and background selections [95]. The successful cases for how to select superior recombinants in the breeding programs have been previously reported [79,96]. In general, when the donor line is a wild relative of the crop, linkage drag is a very common problem which can strongly limit the use of a particular gene in breeding [97]. Our study also provide an optimized strategy for overcoming linkage drag in crop breeding by using commercial variety harboring favorable genes as an improved donor instead of the wild donor, which would help in further improving the background of the pyramided lines and shorting the breeding cycle.
5. Conclusions
Two improved good-quality rice lines iXWX13-1 (Pi1+Pi2+OsHMA3) and iXWX13-2 (Pi1+Pi2+OsNramp5) were developed by using genomic breeding strategy, which is composed of genomics-assisted foreground selection and background selection, and phenotypic screening. The improved line iXWX13-1 can be used as a blast-resistant version, and the improved line iXWX13-2 can be used as a blast-resistant and low Cd accumulation version to replace XWX13. The present study is the first successful practice to apply genomic marker-assisted backcrossing and background selection to improve the blast resistance and to lower the grain Cd accumulation in XWX13 without penalty of the yield, main agronomic traits and grain quality. This proposed strategy would pave the way to integration of multiple desirable genes into the background of elite varieties in crop genetic improvement.
Author Contributions
Conceptualization, Z.X., Z.H. and J.P.; methodology, Y.H. and H.C.; validation, Z.X., Z.H. and Y.W.; formal analysis, Z.X., J.C. and C.Y.; investigation, Y.H., H.C., L.L. and H.Y.; resources, Z.L. and Z.H.; data curation, Z.X., Z.H. and Y.W.; writing – original draft preparation, Z.X. and Z.H.; writing – review & editing, Z.X., Z.H., J.C., C.Y. and J.P.; supervision, Z.H., Y.W. and B.T.; project administration, Z.H., Y.W. and B.T.; funding acquisition, Z.H., L.Z., Y.W. and B.T.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Biological Breeding-National Science and Technology Major Project (2023ZD04076), Hunan Science and Technology Program (2017NK2022, 2023NK2001), Sichuan Science and Technology Program (2024YFHZ0191), and Suzhou Science and Technology Program (SNG2021003).
Data Availability Statement
The original contributions presented in this study are included in the article, and further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar]
- Yuan, L. Development of hybrid rice to ensure food security. Rice Sci. 2014, 21, 1–2. [Google Scholar]
- Skamnioti, P.; Gurr, S.J. Against the grain: safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar]
- Yang, L.; Zhang, W. Genetic and biochemical mechanisms of rice resistance to planthopper. Plant Cell Rep. 2016, 35, 1559–1572. [Google Scholar]
- Yu, S.; Ali, J.; Zhang, C.; Li, Z.; Zhang, Q.F. Genomic breeding of Green Super Rice varieties and their deployment in Asia and Africa. Theor. Appl. Genet. 2020, 133, 1427–1442. [Google Scholar] [CrossRef] [PubMed]
- Kubier, A.; Wilkin, R.T.; Pichler, T. Cadmium in soils and groundwater: A review. Appl. Geochem. 2019, 108, 1–16. [Google Scholar] [CrossRef]
- Wang, P.; Chen, H.; Kopittke, P.; Zhao, F. Cadmium contamination in agricultural soils of China and the impact on food safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar]
- Ma, J.; Shen, R.; Shao, J. Transport of cadmium from soil to grain in cereal crops: A review. Pedosphere 2021, 31, 3–10. [Google Scholar] [CrossRef]
- Xiao, N.; Wu, Y.; Li, A. Strategy for use of rice blast resistance genes in rice molecular breeding. Rice Sci. 2020, 27, 263–277. [Google Scholar]
- Devanna, B.N.; Jain, P.; Solanke, A.U.; Das, A.; Thakur, S.; Singh, P.K.; Kumari, M.; Dubey, H.; Jaswal, R.; Pawar, D.; Kapoor, R.; Singh, J.; Arora, K.; Saklani, B.K.; AnilKumar, C.; Maganti, S.M.; Somah, H.; Deshmukh, R.; Rathour, R.; Sharma, T.R. Understanding the dynamics of blast resistance in rice-magnaporthe oryzae interactions. J. Fungi. 2022, 8, 584. [Google Scholar]
- Yu, Z.H.; Mackill, D.J.; Bonman, J.M.; McCouch, S.R.; Guiderdoni, E.; Notteghem, J.L.; Tanksley, S.D. Molecular mapping of genes for resistance to rice blast (Pyricularia grisea Sacc.). Theor. Appl. Genet. 1996, 93, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Hua, L.; Wu, J.; Chen, C.; Wu, W.; He, X.; Lin, F.; Wang, L.; Ashikawa, I.; Matsumoto, T.; Wang, L.; Pan, Q. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast. Theor. Appl. Genet. 2012, 125, 1047–1055. [Google Scholar]
- Liu, G.; Lu, G.; Zeng, L.; Wang, G. Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physically linked on rice chromosome 6. Mol. Genet. Genom. 2002, 267, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Hashimoto, N.; Daigen, M.; Ashikawa, I. Development of PCR-based SNP markers for rice blast resistance genes at the Piz locus. Theor. Appl. Genet. 2004, 108, 1212–1220. [Google Scholar] [CrossRef]
- Deng, Y.; Zhu, X.; Shen, Y.; He, Z. Genetic characterization and fine mapping of the blast resistance locus Pigm(t) tightly linked to Pi2 and Pi9 in a broad-spectrum resistant Chinese variety. Theor. Appl. Genet. 2006, 113, 705–713. [Google Scholar]
- Zhu, X.; Chen, S.; Yang, J.; Zhou, S.; Zeng, L.; Han, J.; Su, J.; Wang, L.; Pan, Q. The identification of Pi50(t), a new member of the rice blast resistance Pi2/Pi9 multigene family. Theor. Appl. Genet. 2012, 124, 1295–1304. [Google Scholar] [PubMed]
- Jiang, H.; Feng, Y.; Bao, L.; Li, X.; Gao, G.; Zhang, Q.; Xiao, J.; Xu, C.; He, Y. Improving blast resistance of Jin23B and its hybrid rice by marker-assisted gene pyramiding. Mol. Breed. 2012, 30, 1679–1688. [Google Scholar]
- Mi, J.; Yang, D.; Chen, Y.; Jiang, J.; Mou, H.; Huang, J.; Ouyang, Y.; Mou, T. Accelerated molecular breeding of a novel P/TGMS line with broad-spectrum resistance to rice blast and bacterial blight in two-line hybrid rice. Rice 2018, 11, 11. [Google Scholar]
- Yang, D.; Tang, J.; Yang, D.; Chen, Y.; Ali, J.; Mou, T. Improving rice blast resistance of Feng39S through molecular marker-assisted backcrossing. Rice 2019, 12, 70. [Google Scholar] [CrossRef]
- He, Z.; Xin, Y.; Wang, C.; Yang, H.; Xu, Z.; Cheng, J.; Li, Z.; Ye, C.; Yin, H.; Xie, Z.; Jiang, N.; Huang, J.; Xiao, J.; Tian, B.; Liang, Y.; Zhao, K.; Peng, J. Genomics-assisted improvement of super high-yield hybrid rice variety "Super 1000" for resistance to bacterial blight and blast diseases. Front Plant Sci. 2022, 13, 881214. [Google Scholar] [CrossRef]
- Chaney, R.L.; Reeves, P.G.; Ryan, J.A.; Simmons, R.W.; Welch, R.M.; Angle, J.S. An improved understanding of soil Cd risk to humans and low cost methods to remediate soil Cd risks. BioMetals 2004, 17, 549–553. [Google Scholar] [CrossRef]
- Clemens, S.; Aarts, M.M.; Thomine, S.; Verbruggen, N. Plant science: the key to preventing slow cadmium poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [PubMed]
- Chaney, R.L. How does contamination of rice soils with Cd and Zn cause high incidence of human Cd disease in subsistence rice farmers. Curr. Pollut. Rep. 2015, 1, 13–22. [Google Scholar] [CrossRef]
- FAO/WHO. Evaluation of Certain Food Additives and Contaminants, Fifty-fifth Report of the Joint FAO/WHO Expert Committee on Food Additives, WHO Technical Report Series. 2001, No. 901.
- Zhao, F.; Ma, Y.; Zhu, Y.; Tang, Z.; McGrath, S.P. Soil contamination in China: current status and mitigation strategies. Environ. Sci. Technol. 2015, 49, 750–759. [Google Scholar]
- Hu, H.; Jin, Q.; Kavan, P. A study of heavy metal pollution in China: Current status, pollution-control policies and countermeasures. Sustainability 2014, 6, 5820–5838. [Google Scholar] [CrossRef]
- Wang, M.; Chen, W.; Peng, C. Risk assessment of Cd polluted paddy soils in the industrial and township areas in Hunan, Southern China. Chemosphere 2016, 144, 346–351. [Google Scholar]
- Mclaughlin, M.J.; Smolders, E.; Zhao, F.; Grant, C.; Montalvo, D. Managing cadmium in agricultural systems. Adv. Agron. 2021, 166, 1–129. [Google Scholar]
- Shi, Z.; Carey, M.; Meharg, C.; Williams, P.N.; Signes-Pastor, A.J.; Triwardhani, E.A.; Pandiangan, F.I.; Campbell, K.; Elliott, C.; Marwa, E.M.; Xiao, J.; Farias, J.G.; Nicoloso, F.T.; De Silva, P.M.C.S.; Lu, Y.; Norton, G.; Adoako, E.; Green, A.J.; Moreno-Jimenez, E.; Zhu, Y.; Carbonell-Barrachina, A.A.; Haris, P.; Lawgali, Y.F.; Sommella, A.; Pigna, M.; Brabet, C.; Montet, D.; Njira, K.; Watts, M.; Hossain, M.; Islam, M.; Tapia, Y.; Oporto, C.; Meharg, A.A. Rice Grain Cadmium Concentrations in the Global Supply-Chain. Expo. Health 2020, 12, 869–876. [Google Scholar]
- Chen, C.Y.; Tang, W.B. A perspective on the selection and breeding of low-Cd rice. Res. Agric. Mod. 2018, 39, 1044–1051. [Google Scholar]
- Clemens, S.; Ma, J.F. Toxic heavy metal and metalloid accumulation in crop plants and foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar]
- Zhao, F.; Tang, Z.; Song, J.; Huang, X.; Wang, P. Toxic metals and metalloids: Uptake, transport, detoxification, phytoremediation, and crop improvement for safer food. Mol. Plant 2022, 15, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Xu, W.; Xie, J.; Gao, Y.; Wu, L.; Sun, L.; Feng, L.; Chen, X.; Zhang, T.; Dai, C.; Li, T.; Lin, X.; Zhang, Z.; Wang, X.; Li, F.; Zhu, X.; Li, J.; Li, Z.; Chen, C.; Ma, M.; Zhang, H.; He, Z. Variation of a major facilitator superfamily gene contributes to differential cadmium accumulation between rice subspecies. Nat. Commun. 2019, 10, 2562. [Google Scholar] [CrossRef]
- Luo, J.; Huang, J.; Zeng, D.; Peng, J.; Zhang, G.; Ma, H.; Guan, Y.; Yi, H.; Fu, Y.; Han, B.; Lin, H.; Qian, Q.; Gong, J. A defensin-like protein drives cadmium efflux and allocation in rice. Nat. Commun. 2018, 9, 645. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Mao, D.; Sun, L.; Wang, R.; Tan, L.; Zhu, Y.; Huang, H.; Peng, C.; Zhao, Y.; Wang, J.; Huang, D.; Chen, C. CF1 reduces grain-cadmium levels in rice (Oryza sativa). Plant J. 2022, 110, 1305–1318. [Google Scholar] [CrossRef]
- Uraguchi, S.; Fujiwara, T. Rice breaks ground for cadmium-free cereals. Curr. Opin. Plant Biol. 2013, 16, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Pinson, S.R.; Tarpley, L.; Huang, X.Y.; Lahner, B.; Yakubova, E.; Baxter, I.; Guerinot, M.L.; Salt, D.E. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 2014, 127, 137–165. [Google Scholar] [CrossRef]
- Sun, L.; Xu, X.; Jiang, Y.; Zhu, Q.; Yang, F.; Zhou, J.; Yang, Y.; Huang, Z.; Li, A.; Chen, L.; Tang, W.; Zhang, G.; Wang, J.; Xiao, G.; Huang, D.; Chen, C. Genetic diversity, rather than cultivar type, determines relative grain Cd accumulation in hybrid rice. Front. Plant Sci. 2016, 7, 1407. [Google Scholar]
- Sun, L.; Wang, R.; Tang, W.; Chen, Y.; Zhou, J.; Ma, H.; Si, L.; Deng, H.; Han, L.; Chen, Y.; Tan, Y.; Zhu, Y.; Lin, D.; Zhu, Q.; Wang, J.; Huang, D.; Chen, C. Robust identification of low-Cd rice varieties by boosting the genotypic effect of grain Cd accumulation in combination with marker-assisted selection. J. Hazard. Mater. 2022, 424, 127703. [Google Scholar]
- Yang, M.; Lu, K.; Zhao, F.; Xie, W.; Ramakrishna, P.; Wang, G.; Du, Q.; Liang, L.; Sun, C.; Zhao, H.; Zhang, Z.; Liu, Z.; Tian, J.; Huang, X.; Wang, W.; Dong, H.; Hu, J.; Ming, L.; Xing, Y.; Wang, G.; Xiao, J.; Salt, D.; Lian, X. Genome-wide association studies reveal the genetic basis of ionomic variation in rice. Plant Cell 2018, 30, 2720–2740. [Google Scholar]
- Zhao, J.; Yang, W.; Zhang, S.; Yang, T.; Liu, Q.; Dong, J.; Fu, H.; Mao, X.; Liu, B. Genome-wide association study and candidate gene analysis of rice cadmium accumulation in grain in a diverse rice collection. Rice 2018, 11, 61. [Google Scholar] [CrossRef]
- Liu, X.; Chen, S.; Chen, M.; Zheng, G.; Peng, Y.; Shi, X.; Qin, P.; Xu, X.; Teng, S. Association study reveals genetic loci responsible for arsenic, cadmium and lead accumulation in rice grain in contaminated farmlands. Front. Plant Sci. 2019, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Sui, F.; Zhao, D.; Zhu, H.; Gong, Y.; Tang, Z.; Huang, X.; Zhang, G.; Zhao, F. Map-based cloning of a new total loss-of-function allele of OsHMA3 causes high cadmium accumulation in rice grain. J. Exp. Bot. 2019, 70, 2857–2871. [Google Scholar] [CrossRef]
- Tan, Y.; Sun, L.; Song, Q.; Mao, D.; Zhou, J.; Jiang, Y.; Wang, J.; Fan, T.; Zhu, Q.; Huang, D.; Xiao, H.; Chen, C. Genetic architecture of subspecies divergence in trace mineral accumulation and elemental correlations in the rice grain. Theor. Appl. Genet. 2020, 133, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhou, J.; Wang, J.; Sun, L. The genetic architecture for phenotypic plasticity of the rice grain ionome. Front. Plant Sci. 2020, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Zhu, Y.; Li, S.; Deng, H.; Wang, J.; Tang, W.; Sun, L. Genetic control diversity drives differences between cadmium distribution and tolerance in rice. Front. Plant Sci. 2021, 12, 638095. [Google Scholar] [CrossRef]
- Sun, L.; Tan, Y.; Chen, C. The road toward Cd-safe rice: from mass selection to marker-assisted selection and genetic manipulation. The Crop Journal 2023, 4, 1059–1071. [Google Scholar] [CrossRef]
- Ishikawa, S.; Ishimaru, Y.; Igura, M.; Kuramata, M.; Abe, T.; Senoura, T.; Hase, Y.; Arao, T.; Nishizawa, N.K.; Nakanishi, H. Ion-beam irradiation, gene identification, and marker-assisted breeding in the development of low-cadmium rice. Proc. Natl. Acad. Sci. U. S. A. 2012, 109, 19166–19171. [Google Scholar] [CrossRef]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; Pan, Y.; Hu, Y.; Peng, Y.; Fu, X.; Li, H.; Xia, S.; Zhao, B. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef]
- Wang, T.; Li, Y.; Fu, Y.; Xie, H.; Song, S.; Qiu, M.; Wen, J.; Chen, M.; Chen, G.; Tian, Y.; Li, C.; Yuan, D.; Wang, J.; Li, L. Mutation at different sites of metal transporter gene OsNramp5 affects Cd accumulation and related agronomic traits in rice (Oryza sativa L.). Front. Plant Sci. 2019, 10, 1081. [Google Scholar] [CrossRef]
- Chang, J.D.; Huang, S.; Konishi, N.; Wang, P.; Chen, J.; Huang, X.; Ma, J.; Zhao, F.; Dietz, K. Overexpression of the manganese/cadmium transporter OsNRAMP5 reduces cadmium accumulation in rice grain. J. Exp. Bot. 2020, 71, 5705–5715. [Google Scholar] [CrossRef]
- Sasaki, A.; Yamaji, N.; Ma, J. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice. J. Exp. Bot. 2014, 65, 6013–6021. [Google Scholar]
- Lu, C.; Zhang, L.; Tang, Z.; Huang, X.; Ma, J.; Zhao, F. Producing cadmium-free indica rice by overexpressing OsHMA3. Environ. Int. 2019, 126, 619–626. [Google Scholar] [PubMed]
- Tanksley, S.; Young, N.; Paterson, N.; Bonierbale, M. RFLP mapping in plant breeding: new tools for an old science. Nat. Biotechnol. 1989, 7, 257–264. [Google Scholar]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar]
- Bai, J.; Zhang, Q.; Jia, X. Comparison of different foreground and background selection methods in marker-assisted introgression. Acta Genet. Sin. 2006, 33, 1073–1080. [Google Scholar] [PubMed]
- Xu, Y.; Lu, Y.; Xie, C.; Gao, S.; Wan, J.; Prasanna, B.M. Whole-genome strategies for marker-assisted plant breeding. Mol. Breed. 2012, 29, 833–854. [Google Scholar]
- Mangal, V.; Verma, L.K.; Singh, S.K.; Saxena, K.; Roy, A.; Karn, A.; Rohit, R.; Kashyap, S.; Bhatt, A.; Sood, S. Triumphs of genomic-assisted breeding in crop improvement. Heliyon 2024, 10, e35513. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Morales, K.Y.; Singh, N.; Perez, F.A.; Ignacio, J.C.; Thapa, R.; Arbelaez, J.D.; Tabien, R.E.; Famoso, A.; Wang, D.R.; Septiningsih, E.M.; Shi, Y.; Kretzschmar, T.; McCouch, S.R.; Thomson, M.J. An improved 7K SNP array, the C7AIR, provides a wealth of validated SNP markers for rice breeding and genetics studies. PLoS ONE 2020, 15, e0232479. [Google Scholar]
- Tian, H.; Yang, Y.; Yi, H.; Xu, L.; He, H.; Fan, Y.; Wang, L.; Ge, J.; Liu, Y.; Wang, F.; Zhao, J. New resources for genetic studies in maize (Zea mays L.): a genome-wide Maize6H-60K single nucleotide polymorphism array and its application. Plant J. 2020, 105, 1113–1122. [Google Scholar]
- Rasheed, A.; Hao, Y.; Xia, X.; Khan, A.; Xu, Y.; Varshney, R.K.; He, H. Crop Breeding Chips and Genotyping Platforms: Progress, Challenges, and Perspectives. Mol. Plant. 2017, 10, 1047–1064. [Google Scholar] [CrossRef]
- Bayer, M.M.; Rapazote-Flores, P.; Ganal, M.; Hedley, P.E.; Macaulay, M.; Plieske, J.; Ramsay, L.; Russell, J.; Shaw, P.D.; Thomas, W.; Waugh, R. Development and Evaluation of a Barley 50k iSelect SNP Array. Front. Plant Sci. 2017, 8, 1792. [Google Scholar]
- Sun, C.; Dong, Z.; Zhao, L.; Ren, Y.; Zhang, N.; Chen, F. The Wheat 660K SNP array demonstrates great potential for marker-assisted selection in polyploid wheat. Plant Biotechnol J. 2020, 18, 1635. [Google Scholar] [CrossRef]
- Yu, S.; Ali, J.; Zhou, S.; Ren, G.; Xie, H.; Xu, J.; Yu, X.; Zhou, F.; Peng, S.; Ma, L.; Yuan, D.; Li, Z.; Chen, D.; Zheng, R.; Zhao, Z.; Chu, C.; You, A.; Wei, Y.; Zhu, S.; Gu, Q.; He, G.; Li, S.; Liu, G.; Liu, C.; Zhang, C.; Xiao, J.; Luo, L.; Li, Z.; Zhang, Q. From Green Super Rice to green agriculture: reaping the promise of functional genomics research. Mol. Plant 2022, 15, 9–26. [Google Scholar]
- Tung, C.W.; Zhao, K.Y.; Wright, M.H.; Ali, M.L.; Jung, J.; Kimball, J.; Tyagi, W.; Thomson, M.J.; McNally, K.; Leung, H.; Kim, H.; Ahn, S.N.; Reynolds, A.; Scheffler, B.; Eizenga, G.; McClung, A.; Bustamante, C.; McCouch, S.R. Development of a research platform for dissecting phenotype-genotype associations in rice (Oryza spp.). Rice 2010, 3, 205–217. [Google Scholar]
- Yu, H.; Xie, W.; Li, J.; Zhou, F.; Zhang, Q. A whole genome SNP array, RICE6K, for genomic breeding in rice. Plant Biotechnol. J. 2014, 12, 28–37. [Google Scholar] [CrossRef]
- Singh, N.; Jayaswal, P.K.; Panda, K.; Mandal, P.; Kumar, V.; Singh, B.; Mishra, S.; Singh, Y.; Singh, R.; Rai, V.; Gupta, A.; Sharma, T.R.; Singh, N.K. Single-copy gene based 50 K SNP chip for genetic studies and molecular breeding in rice. Sci. Rep. 2015, 5, 11600. [Google Scholar] [CrossRef] [PubMed]
- McCouch, S.; Wright, M.; Tung, C.W.; Maron, L.; McNally, K.; Fitzgerald, M.; Singh, N.; DeClerck, G.; Agosto, P.F.; Korniliev, P.; Greenberg, A.J.; Naredo, M.E.; Mercado, S.M.; Harrington, S.E.; Shi, Y.; Branchini, D.A.; Kuser-Falcão, P.R.; Leung, H.; Ebana, K.; Yano, M.; Eizenga, G.; McClung, A.; Mezey, J. Open access resources for genome wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar]
- Morales, K.Y.; Singh, N.; Perez, F.A.; Ignacio, J.C.; Thapa, R.; Arbelaez, J.D.; Tabien, R.E.; Famoso, A.; Wang, D.R.; Septiningsih, E.M.; Shi, Y.; Kretzschmar, T.; McCouch, S.R.; Thomson, M.J. An improved 7K SNP array, the C7AIR, provides a wealth of validated SNP markers for rice breeding and genetics studies. PLoS ONE 2020, 15, e0232479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, M.; Liang, L.; Xiang, J.; Zhang, F.; Zhang, C.; L, Yi.; Liang, J.; Zheng, T.; Zhang, F.; Li, H.; Fu, B.; Shi, Y.; Xu, J.; Tian, B.; Li, Z.; Wang, W. Rice3K56 is a high-quality SNP array for genome-based genetic studies and breeding in rice (Oryza sativa L.). The Crop Journal 2023, 11, 800–807. [Google Scholar]
- Zhang, Y.; Fang, B.; Teng, Z.; Chen, G.; Liu, Y.; Ling, W.; Xiang, S.; Bai, L. Screening and verification of rice varieties with low cadmium accumulation. Hunan Agric. Sci. 2017, 12, 19–25. [Google Scholar]
- Liu, M.; Zhang, S.; Hu, J.; Sun, W.; Padilla, J.; He, Y.; Li, Y.; Yin, Z.; Liu, X.; Wang, W.; Shen, D.; Li, D.; Zhang, H.; Zheng, X.; Cui, Z.; Wang, G.; Wang, P.; Zhou, B.; Zhang, Z. Phosphorylation-guarded light-harvesting complex II contributes to broad-spectrum blast resistance in rice. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 17572–17577. [Google Scholar] [CrossRef] [PubMed]
- International Rice Research Institute (IRRI), Standard Evaluation System for Rice. Manila. Philippines. 1996, 4th ed, pp. 17-18.
- Latif, M.A.; Badsha, M.A.; Tajul, M.I.; Kabir, M.S.; Rafii, M.Y.; Mia, M. Identification of genotypes resistant to blast.; bacterial leaf blight.; sheath blight and tungro and efficacy of seed treating fungicides against blast disease of rice. Sci. Res. Essays 2011, 6, 2804–2811. [Google Scholar]
- Wing, R.A.; Purugganan, M.D.; Zhang, Q. The rice genome revolution: from an ancient grain to Green Super Rice. Nat. Rev. Genet. 2018, 19, 505–517. [Google Scholar]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar]
- Zhou, X.; Jiang, G.; Yang, L.; Qiu, L.; He, P.; Nong, C.; Wang, Y.; He, Y.; Xing, Y. Gene diagnosis and targeted breeding for blast-resistant Kongyu 131 without changing regional adaptability. J. Genet. Genomics 2018, 45, 539–547. [Google Scholar]
- Wang, H.; Gao, Y.; Mao, F.; Xiong, L.; Mou, T. Directional upgrading of brown planthopper resistance in an elite rice cultivar by precise introgression of two resistance genes using genomics-based breeding. Plant Sci. 2019, 288, 110211. [Google Scholar] [PubMed]
- Yang, D.; Xiong, L.; Mou, T.; Mi, J. Improving the resistance of the rice PTGMS line Feng39S by pyramiding blast, bacterial blight, and brown planthopper resistance genes. The Crop Journal 2022, 10, 1187–1197. [Google Scholar] [CrossRef]
- Hu, S.; Zhou, L.; Wang, J.; Mawia, A.M.; Hui, S.; Xu, B.; Jiao, G.; Sheng, Z.; Shao, G.; Wei, X.; Wang, L.; Xie, L.; Zhao, F.; Tang, S.; Hu, P. Production of grains with ultra-low heavy metal accumulation by pyramiding novel Alleles of OsNramp5 and OsLsi2 in two-line hybrid rice. Plant Biotechnol. J. 2024, 22, 2921–2931. [Google Scholar]
- Tang, L.; Song, J.; Cui, Y.; Fan, H.; Wang, J. Detection and evaluation of blast resistance genes in backbone indica rice varieties from South China. Plants 2024, 13, 2134. [Google Scholar] [CrossRef]
- Wu, Y.; Xiao, N.; Chen, Y.; Yu, L.; Pan, C.; Li, Y.; Zhang, X.; Huang, N.; Ji, H.; Dai, Z.; Chen, X.; Li, A. Comprehensive evaluation of resistance effects of pyramiding lines with different broad-spectrum resistance genes against Magnaporthe oryzae in rice (Oryza sativa L.). Rice 2019, 12, 11. [Google Scholar]
- Hittalmani, S.; Parco, A.; Mew, T.V.; Zeigler, R.S.; Huang, N. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor. Appl. Genet. 2000, 100, 1121–1128. [Google Scholar]
- Singh, V.K.; Singh, A.; Singh, S.P.; Ellur, R.K.; Choudhary, V.; Sarkel, S.; Singh, D.; Krishnan, S.G.; Nagarajan, M.; Vinod, K.K. Incorporation of blast resistance into “PRR78”, an elite Basmati rice restorer line, through marker assisted backcross breeding, Field Crops Res. 2012, 128, 8-16.
- Zeng, P.; Wei, B.; Zhou, H.; Gu, J.; Liao, B. Co-application of water management and foliar spraying silicon to reduce cadmium and arsenic uptake in rice: A two-year field experiment. Sci. Total Environ. 2022, 818, 151801. [Google Scholar] [PubMed]
- Zhao, F.J.; Wang, P. Arsenic and cadmium accumulation in rice and mitigation strategies. Plant and Soil 2020, 446, 1–21. [Google Scholar]
- Hao, X.; Wu, C.; Wang, R.; Tian, L.; Song, T.; Tan, H.; Peng, Y.; Zeng, M.; Chen, L.; Liang, M.; Li, D. Association between sequence variants in cadmium-related genes and the cadmium accumulation trait in thermo-sensitive genic male sterile rice. Breed. Sci. 2019, 69, 455–463. [Google Scholar]
- Zhang, J.; Zhu, Y.; Yu, L.; Yang, M.; Zou, X.; Yin, C.; Lin, Y. Research Advances in Cadmium Uptake, Transport and Resistance in Rice (Oryza sativa L.). Cells 2022, 11, 569. [Google Scholar] [CrossRef]
- Yu, H.; Liu, C.; Zhu, J.; Li, F.; Deng, D.; Wang, Q.; Liu, C. Cadmium availability in rice paddy fields from a mining area: The effects of soil properties highlighting iron fractions and pH value. Environ. Pollut. 2016, 209, 38–45. [Google Scholar] [PubMed]
- Zhang, M.; Pinson, S.R.M.; Tarpley, L.; Huang, X.Y.; Lahner, B.; Yakubova, E.; Baxter, I.; Guerinot, M.L.; Salt, D.E. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 2014, 127, 137–165. [Google Scholar]
- Wang, X.; Whalley, W.; Miller, A.; White, P.; Zhang, F.; Shen, J. Sustainable Cropping Requires Adaptation to a Heterogeneous Rhizosphere. Trends Plant Sci. 2020, 25, 1194–1202. [Google Scholar]
- He, J.; Zhu, C.; Ren, Y.; Yan, Y.; Jiang, D. Genotypic variation in grain cadmium concentration of lowland rice. J. Plant Nutr. Soil Sci. 2006, 169, 711–716. [Google Scholar] [CrossRef]
- Hui, Y.; Wang, J.; Wei, F.; Yuan, J.; Yang, Z. Cadmium accumulation in different rice cultivars and screening for pollution-safe cultivars of rice. Sci. Total Environ. 2006, 370, 302–309. [Google Scholar]
- Deng, Y.; He, Z. The seesaw action: balancing plant immunity and growth. Sci. Bull. 2024, 69, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Nelson, R.; Wiesner-Hanks, T.; Wisser, R.; Balint-Kurti, P. Navigating complexity to breed disease-resistant crops. Nat. Rev. Genet. 2018, 19, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Ellur, R.K.; Khanna, A.; S, G.K.; Bhowmick, P.K.; Vinod, K.K.; Nagarajan, M.; Mondal, K.K.; Singh, N.K.; Singh, K.; Prabhu, K.V.; Singh, A.K. Marker-aided incorporation of Xa38, a novel bacterial blight resistance gene, in PB1121 and comparison of its resistance spectrum with xa13 + Xa21. Sci. Rep. 2016, 6, 29188. [Google Scholar] [CrossRef] [PubMed]
- Mores, A.; Borrelli, G.M.; aidò, L.G.; Petruzzino, G.; Pecchioni, N.; Amoroso, L.G.; Desiderio, F.; Mazzucotelli, E.; Mastrangelo, A.M.; Marone, D. Genomic approaches to identify molecular bases of crop resistance to diseases and to develop future breeding strategies. Int. J. Mol. Sci. 2021, 22, 5423. [Google Scholar] [CrossRef]
Figure 1.
The scheme for development of the improved lines, iXWX13-1 and iXWX13-2.
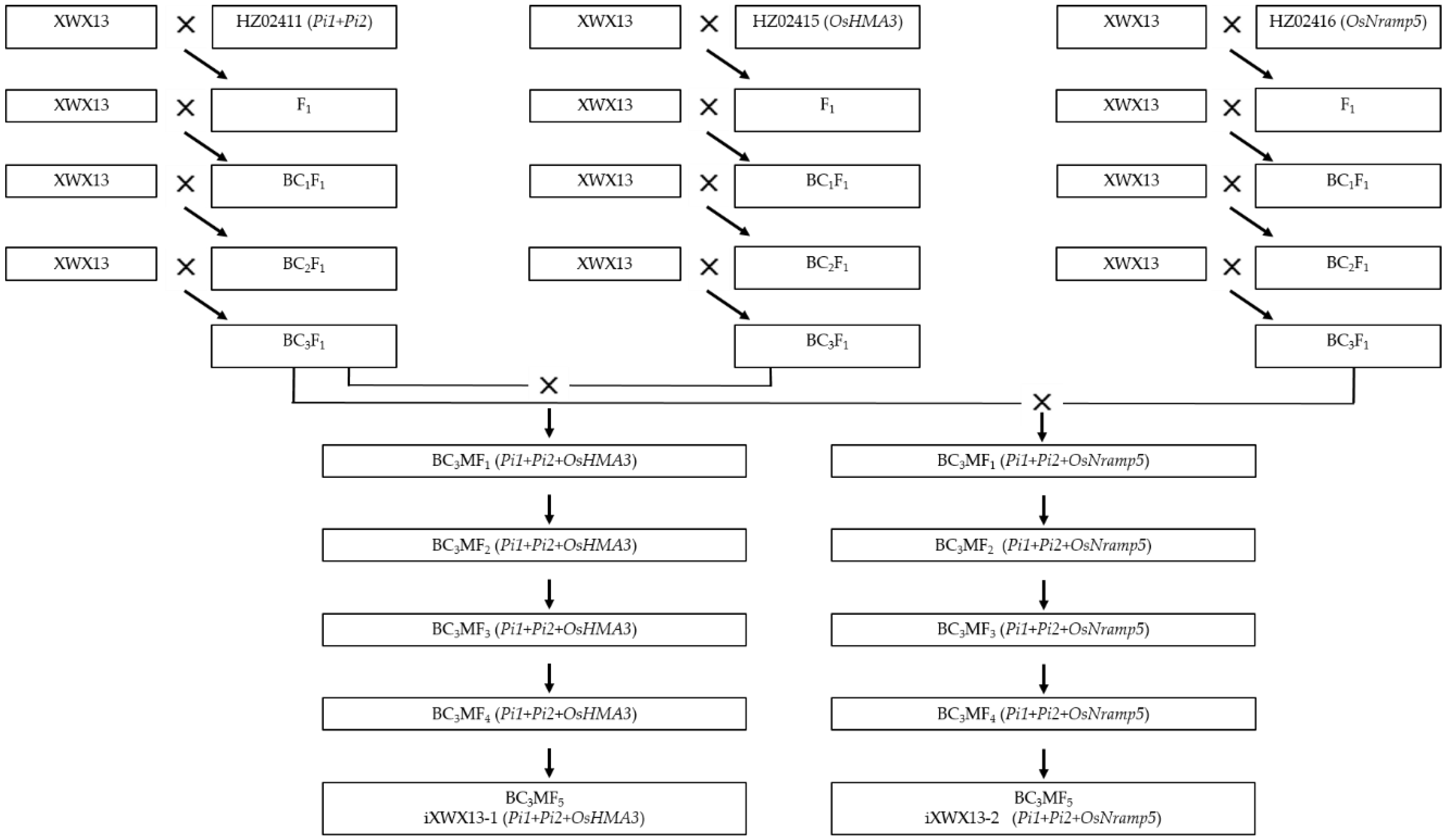
Figure 2.
Genetic background analysis for the improved line iXWX13-1 (Pi1+Pi2+OsHMA3) by the rice 1K mGPS. Twelve chromosomes of rice are labeled from 1 to 12, and the triangle symbols indicate the positions of the target genes. The blue lines indicate the different and homozygous SNP loci between iXWX13-1 and the recurrent parent XWX13.
Figure 2.
Genetic background analysis for the improved line iXWX13-1 (Pi1+Pi2+OsHMA3) by the rice 1K mGPS. Twelve chromosomes of rice are labeled from 1 to 12, and the triangle symbols indicate the positions of the target genes. The blue lines indicate the different and homozygous SNP loci between iXWX13-1 and the recurrent parent XWX13.
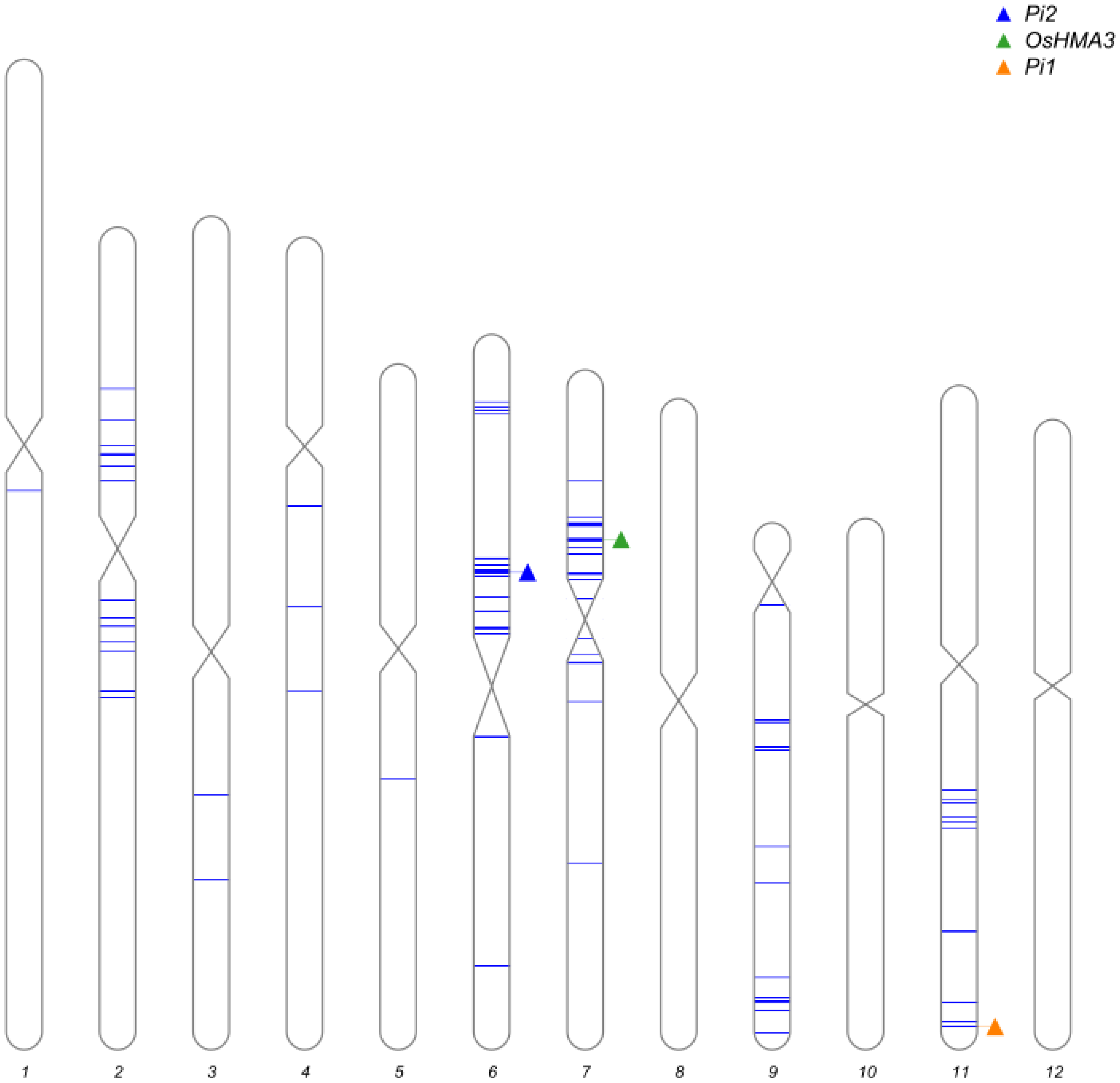
Figure 3.
Genetic background analysis for the improved line iXWX13-2 (Pi1+Pi2+ OsNramp5) by the rice 1K mGPS. Twelve chromosomes of rice are labeled from 1 to 12, and the triangle symbols indicate the positions of the target genes. The green lines indicate the different and homozygous SNP loci between iXWX13-2 and the recurrent parent XWX13.
Figure 3.
Genetic background analysis for the improved line iXWX13-2 (Pi1+Pi2+ OsNramp5) by the rice 1K mGPS. Twelve chromosomes of rice are labeled from 1 to 12, and the triangle symbols indicate the positions of the target genes. The green lines indicate the different and homozygous SNP loci between iXWX13-2 and the recurrent parent XWX13.
Figure 4.
The typical symptoms of seedling blast resistance of XWX13 and the improved lines. HR, highly resistant; S, susceptible; HS, highly susceptible. (A) The blast isolate 21DWS-2 originated from Dawei Mountain, Liuyang City, Hunan, China in 2021. (B) The blast isolate 20CHTN1-2 originated from Chunhua Town, Changsha County, Hunan, China in 2020.
Figure 4.
The typical symptoms of seedling blast resistance of XWX13 and the improved lines. HR, highly resistant; S, susceptible; HS, highly susceptible. (A) The blast isolate 21DWS-2 originated from Dawei Mountain, Liuyang City, Hunan, China in 2021. (B) The blast isolate 20CHTN1-2 originated from Chunhua Town, Changsha County, Hunan, China in 2020.

Figure 5.
The performance for leaf blast of XWX13 and the improved lines in the field of Taojiang County, Yiyang City, Hunan, China in 2022.
Figure 5.
The performance for leaf blast of XWX13 and the improved lines in the field of Taojiang County, Yiyang City, Hunan, China in 2022.

Figure 6.
Comparisons of the grain Cd concentration in a multi-site trial. The values of grain Cd concentration given are mean of three replications. The letter a, b, and c indicate the Student-Newman-Keuls significance at 5% probability level in each test site based on the single factor ANOVA test.
Figure 6.
Comparisons of the grain Cd concentration in a multi-site trial. The values of grain Cd concentration given are mean of three replications. The letter a, b, and c indicate the Student-Newman-Keuls significance at 5% probability level in each test site based on the single factor ANOVA test.
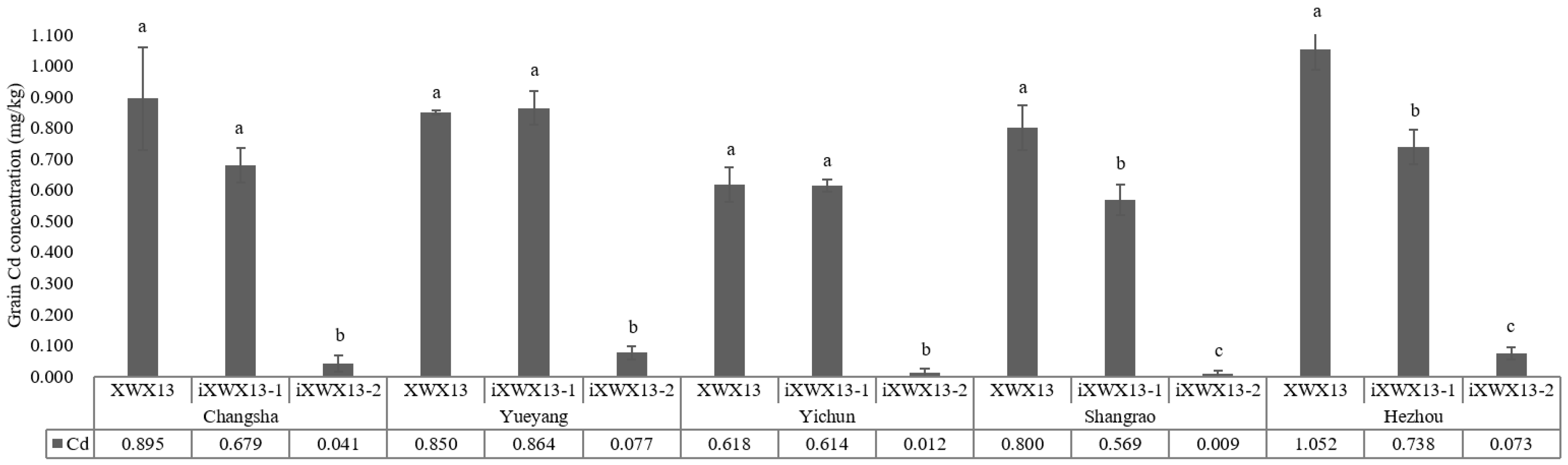

Table 1.
The lesion scores of rice lines at seedling stage against 39 M. oryzae isolates.
| Code of isolates | Place of origin | XWX13 | iXWX13-1 | iXWX13-2 | CO39 | Gumei4 |
|---|---|---|---|---|---|---|
| 110-2 | Hunan, China | 5 | 0 | 0 | 5 | 0 |
| 236-1 | Hunan, China | 0 | 0 | 0 | 5 | 0 |
| 2016CH-2 | Hunan, China | 8 | 0 | 0 | 5 | 0 |
| 21TJ-9 | Hunan, China | 6 | 0 | 0 | 3 | 0 |
| 21TJ-18 | Hunan, China | 8 | 0 | 0 | 4 | 0 |
| 21TJ-23 | Hunan, China | 9 | 0 | 0 | 3 | 0 |
| 21DWS-2 | Hunan, China | 9 | 0 | 0 | 7 | 0 |
| 21DWS-4 | Hunan, China | 9 | 0 | 0 | 5 | 0 |
| 21DWS-19 | Hunan, China | 9 | 0 | 0 | 6 | 0 |
| 20JYR900-3 | Hunan, China | 6 | 0 | 0 | 4 | 0 |
| 20JYR900-18 | Hunan, China | 7 | 0 | 0 | 3 | 0 |
| 20JYR900-23 | Hunan, China | 7 | 0 | 0 | 4 | 0 |
| 20CHR900-5 | Hunan, China | 6 | 0 | 0 | 4 | 0 |
| 20CHTN1-2 | Hunan, China | 8 | 0 | 0 | 7 | 0 |
| 20CHTN1-28 | Hunan, China | 8 | 0 | 0 | 5 | 0 |
| E2007046A2 | Hubei, China | 7 | 0 | 2 | 8 | 6 |
| E2007038A3 | Hubei, China | 7 | 0 | 0 | 4 | 0 |
| 19-765-1-2 | Zhejiang, China | 4 | 0 | 0 | 6 | 0 |
| 19-763-7-2 | Zhejiang, China | 7 | 0 | 0 | 5 | 6 |
| RB7 | Guangdong, China | 5 | 0 | 0 | 7 | 0 |
| CHL1743 | Guangdong, China | 9 | 0 | 0 | 8 | 0 |
| M2006123A3 | Fujian, China | 8 | 0 | 0 | 8 | 0 |
| M2006123A1 | Fujian, China | 9 | 0 | 0 | 8 | 0 |
| C8-3-1 | Sichuan, China | 5 | 0 | 0 | 3 | 0 |
| 20DT-1 | Sichuan, China | 6 | 0 | 0 | 6 | 0 |
| 20DT-6 | Sichuan, China | 7 | 0 | 0 | 5 | 0 |
| 20DT-14 | Sichuan, China | 8 | 0 | 0 | 6 | 0 |
| 12-1 | Sichuan, China | 4 | 0 | 0 | 6 | 0 |
| 19-9-6-2 | Sichuan, China | 4 | 0 | 0 | 2 | 0 |
| CH-11391 | Yunnan, China | 3 | 3 | 0 | 5 | 0 |
| CH105a | Yunnan, China | 3 | 0 | 0 | 5 | 0 |
| 2016ZY-8 | Unknown | 5 | 0 | 0 | 5 | 0 |
| 95097AZC13 | Unknown | 5 | 0 | 0 | 3 | 0 |
| chnos60-2-3 | Unknown | 5 | 0 | 0 | 3 | 0 |
| ROR1 | South Korea | 2 | 0 | 0 | 6 | 0 |
| P06-6 | Philippines | 2 | 0 | 0 | 5 | 0 |
| Guy11 | France | 8 | 4 | 0 | 5 | 0 |
| ES6 | Spain | 3 | 0 | 0 | 5 | 0 |
| IC-17 | America | 3 | 0 | 0 | 4 | 4 |
| Number of resistant isolates | 7 | 38 | 39 | 7 | 36 | |
| Number of susceptible isolates | 32 | 1 | 0 | 32 | 3 | |
| Resistance frequency | 17.95% | 97.44% | 100.00% | 17.95% | 92.31% | |
The lesion scores of 0-3 were classified as resistant, and 4-9 were classified as susceptible against the isolates.
Table 2.
Evaluation of leaf and panicle neck blast resistance of XWX13 and the improved lines in the blast epidemic fields in 2022.
Table 2.
Evaluation of leaf and panicle neck blast resistance of XWX13 and the improved lines in the blast epidemic fields in 2022.
| Entries | Dawei Mountain, Liuyang City | Taojiang County, Yiyang City | |
|---|---|---|---|
| Leaf blast score | Leaf blast score | Panicle neck blast infection rate | |
| CO39 | 7 | 7 | 50% |
| Fengliangyou4 | 7 | 7 | 40% |
| XWX13 | 7 | 8 | 18% |
| iXWX13-1 | 0 | 2 | 4% |
| XWX13 | 7 | 7 | 15% |
| iXWX13-2 | 1 | 2 | 5% |
| Gumei4 | 2 | 2 | 6% |
Table 3.
Yield and main agronomic traits of XWX13 and the improved lines in a multi-site trial in 2022.
Table 3.
Yield and main agronomic traits of XWX13 and the improved lines in a multi-site trial in 2022.
| Sites | Entries | YD | DTM | PH | PL | SPP | FGP | GW |
|---|---|---|---|---|---|---|---|---|
| Changsha | XWX13 | 6.70 | 116.0 | 116.2 | 23.5 | 128.2 | 81.1 | 29.6 |
| iXWX13-1 | 6.53 | 116.0 | 115.9 | 23.8 | 120.3 | 77.8 | 32.9 | |
| iXWX13-2 | 6.72 | 116.0 | 115.2 | 23.6 | 125.3 | 79.6 | 31.8 | |
| Yueyang | XWX13 | 8.19 | 115.0 | 116.5 | 24.1 | 120.3 | 84.1 | 33.2 |
| iXWX13-1 | 8.15 | 115.0 | 115.3 | 24.3 | 125.3 | 82.6 | 33.2 | |
| iXWX13-2 | 8.25 | 115.0 | 114.8 | 24.4 | 124.8 | 82.4 | 33.1 | |
| Yichun | XWX13 | 7.99 | 125.0 | 133.0 | 23.8 | 144.9 | 82.1 | 28.3 |
| iXWX13-1 | 7.89 | 130.0 | 128.4 | 21.9 | 140.7 | 74.2 | 27.6 | |
| iXWX13-2 | 8.05 | 132.0 | 128.8 | 22.8 | 142.5 | 76.4 | 28.1 | |
| Shangrao | XWX13 | 7.13 | 128.0 | 95.5 | 21.2 | 152.8 | 76.3 | 25.1 |
| iXWX13-1 | 6.93 | 136.0 | 91.5 | 22.4 | 141.1 | 76.6 | 27.2 | |
| iXWX13-2 | 7.15 | 134.0 | 91.2 | 21.8 | 150.4 | 77.4 | 26.8 | |
| Hezhou | XWX13 | 7.49 | 118.0 | 115.2 | 23.9 | 118.5 | 81.9 | 33.5 |
| iXWX13-1 | 7.34 | 118.0 | 112.6 | 24.1 | 127.4 | 79.3 | 33.0 | |
| iXWX13-2 | 7.37 | 118.0 | 111.6 | 23.6 | 122.4 | 79.1 | 33.3 | |
| Mean | XWX13 | 7.50 | 120.4 | 115.3 | 23.3 | 132.9 | 81.1 | 30.0 |
| iXWX13-1 | 7.37 | 123.0 | 112.7 | 23.3 | 131.0 | 78.1 | 30.8 | |
| iXWX13-2 | 7.51 | 123.0 | 112.3 | 23.2 | 133.1 | 79.0 | 30.6 | |
| F | 0.076 | 0.163 | 0.071 | 0.005 | 0.044 | 1.515 | 0.097 | |
| p-value | 0.927 | 0.851 | 0.931 | 0.995 | 0.957 | 0.259 | 0.909 | |
| Student-Newman-Keuls (p=0.05) | 0.936 | 0.875 | 0.935 | 0.996 | 0.962 | 0.247 | 0.910 | |
YD, Yield (t/ha); DTM, Days to maturity (d); PH, Plant height (cm); PL, Panicle length (cm); SPP, Spikelets per panicle; FGP, Filled-grain percentage (%); GW, 1000-grain weight (g); The single factor ANOVA test was performed to examine the significance of variation in yield and agronomic traits by the software SPSS Statistics 19.
Table 4.
Grain quality of XWX13 and the improved lines in the multi-site trial.
| Sites | Entries | BRP | MRP | HRP | CRP | CD | RGL | L/W | ASV | AC | GC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Changsha | XWX13 | 80.5 | 70.4 | 55.1 | 11.4 | 2.7 | 6.9 | 3.3 | 2.5 | 14.6 | 48.8 |
| iXWX13-1 | 80.7 | 70.9 | 54.0 | 14.5 | 2.5 | 6.7 | 3.2 | 2.2 | 14.7 | 46.0 | |
| iXWX13-2 | 80.7 | 70.8 | 54.7 | 12.0 | 2.6 | 6.8 | 3.2 | 2.3 | 14.6 | 47.0 | |
| Yueyang | XWX13 | 79.0 | 71.8 | 65.3 | 8.3 | 2.2 | 6.9 | 3.5 | 3.6 | 14.2 | 44.8 |
| iXWX13-1 | 79.5 | 71.5 | 62.0 | 8.4 | 2.0 | 6.4 | 3.3 | 3.6 | 14.5 | 55.5 | |
| iXWX13-2 | 79.2 | 71.7 | 64.3 | 9.0 | 2.1 | 6.7 | 3.2 | 3.5 | 14.4 | 50.0 | |
| Yichun | XWX13 | 79.0 | 70.6 | 63.0 | 7.2 | 2.3 | 6.8 | 3.4 | 2.2 | 14.0 | 51.5 |
| iXWX13-1 | 79.3 | 70.9 | 63.9 | 4.4 | 1.3 | 6.6 | 3.2 | 2.0 | 16.0 | 55.0 | |
| iXWX13-2 | 79.4 | 70.8 | 63.6 | 5.0 | 1.5 | 6.6 | 3.3 | 2.1 | 15.5 | 54.0 | |
| Shangrao | XWX13 | 80.9 | 72.1 | 60.7 | 21.6 | 5.5 | 7.0 | 3.4 | 2.8 | 14.6 | 44.0 |
| iXWX13-1 | 80.9 | 71.5 | 56.1 | 22.9 | 4.8 | 6.6 | 3.2 | 3.3 | 16.5 | 45.0 | |
| iXWX13-2 | 80.5 | 71.8 | 58.5 | 22.0 | 4.9 | 6.7 | 3.2 | 3.1 | 16.3 | 44.0 | |
| Hezhou | XWX13 | 80.0 | 70.3 | 49.6 | 11.4 | 2.6 | 6.8 | 3.2 | 1.4 | 14.9 | 60.8 |
| iXWX13-1 | 80.3 | 70.7 | 41.3 | 16.8 | 3.3 | 6.6 | 3.1 | 1.3 | 16.3 | 65.8 | |
| iXWX13-2 | 80.4 | 70.9 | 44.8 | 13.0 | 2.8 | 6.6 | 3.1 | 1.3 | 16.2 | 64.0 | |
| Mean | XWX13 | 79.88 | 71.04 | 58.74 | 11.98 | 3.06 | 6.88 a | 3.36 a | 2.50 | 14.46 | 49.98 |
| iXWX13-1 | 80.14 | 71.10 | 55.46 | 13.40 | 2.78 | 6.58 b | 3.20 b | 2.48 | 15.60 | 53.46 | |
| iXWX13-2 | 80.04 | 71.20 | 57.18 | 12.20 | 2.78 | 6.68 b | 3.20 b | 2.46 | 15.40 | 51.80 | |
| F | 0.149 | 0.088 | 0.221 | 0.070 | 0.073 | 13.462 | 5.565 | 0.003 | 3.135 | 0.256 | |
| p-value | 0.863 | 0.916 | 0.805 | 0.932 | 0.930 | 0.001* | 0.019* | 0.997 | 0.080 | 0.778 | |
BRP, Brown rice percentage (%); MRP, Milled rice percentage (%); HRP, Head rice percentage (%); CRP, Chalky rice percentage (%); CD, Chalkiness degree (%); RGL, Rice grain length (mm); L/W, Grain length/width ratio; ASV, Alkali spreading value; AC, Amylose content (%); GC, Gel consistency (mm). The letter a, b, and asterisk symbol indicate significance of difference at 5% probability level based on the single factor ANOVA test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



