Submitted:
23 October 2024
Posted:
24 October 2024
You are already at the latest version
Abstract
Rice blast is the most harmful disease caused by Magnaporthe oryzae, leading to yield loss. The aim of the work is to improve resistance of rice to blast by introducing and pyramiding genes using conventional breeding technique and modern MAS analysis. Pyramiding is the combination of several resistance genes in one genotype, in this case it improves the resistance of rice to pathogens. MAS analysis revealed that 3 out of 4 studied domestic high-yielding rice cultivars (Bakanasski, Aisaule and Aru) contain 1 blast resistance gene (Pi-2). It was discovered 7 pyramidal lines with the introduction of a combination of 2 blast resistance genes (Pi-1 and Pi-ta) into 6 hybrids and 3 genes (Pi-1, Pi-33 and Pi-ta) into 1 line. Phytopathological test of local cultivars and pyramidal lines of rice showed that 3 out of 7 pyramidal lines have moderate resistance: 2 lines (F2 Bakanasski/7667 (var. vulgaris Koern.) and F2 Aisaule/7664 (var. italica Alef.)), containing 2 resistance genes (Pi-1 and Pi-ta) and 1 line – F2 Aisaule/7689 (var. zeravschanica Brsch.), carrying 3 genes (Pi-1, Pi-33 and Pi-ta), while the local cultivars Bakanasski, Aisaule and Aru are susceptible. As a result, the blast resistance of rice was improved by creating pyramidal lines. Pyramided lines can be used for germplasm exchange and in rice breeding programs to improve blast resistance in rice.
Keywords:
Rice
; hybrid
; Magnaporthe oryzae
; resistance
; Marker Assisted Selection (MAS)
; gene pyramiding
; phytopathology
1. Introduction
Rice blast is one of the most dangerous diseases [1,2], and the causative agent of this disease is the parasitic fungus Magnaporthe oryzae. Defeat of rice by pathogen in the early stages of development leads to a decrease in seed germination, sparseness of crops and death of seedlings. The greatest harm from infection occurs during the heading and flowering, which is associated with the formation of underdeveloped or feeble seeds, which significantly reduces the quality of the grain [3]. Blast reduces rice production in the world, causing crop losses from 15-40% and reaching up to 80-100% in epiphytotic years [4].
Global climate change affects the evolution of pathogen biotypes, which challenges breeders and forces them to increase rice productivity by creating and introducing new cultivars resistant to changing races or isolates of Magnaporthe oryzae [5]. Currently, more than 100 blast resistance genes have been discovered [5,6,7,8,9,10,11,12,13,14,15]. Most of them with broad spectrum resistance to pathogen [5,6,16,17,18,19], there are some major genes with a wide spectrum of resistance, such as: Pi-z5 (Pi-2) [20], Pi-9 (t) [21], Pigm [22,23,24], Pi-ta [25], Pi-1 and Pi-33 [7,8,9,10,11,12,13,14,26], which serve as indicators of blast resistance gene clusters, which is important when choosing parental pairs for hybridization. Most blast resistance genes are dominant, some of them are quantitative [27], and most of them, such as Pita, Pi-1, Pi25, Pigm and Pia, encode proteins that recognize the signal and inhibit the growth of the phytopathogen, maintaining plant immunity [23,28,29,30]. For example, the Pita gene encodes a cytoplasmic membrane receptor protein that leads to resistance. [31]. And the Pigm gene encodes two proteins, PigmR and PigmS, which regulate resistance and yield of rice plants [23]. Recently, the Pigm-1 gene has been found, encoding the Pigm-1 protein, which is an allelic variant of PigmR [32]. Moreover, many resistance genes are clustered on chromosomes 6, 9, 11, and 12 [33,34]. Disease resistance is controlled by one or two [35,36], three [37] or more pairs of genes [38]. Typically, rice has one or two dominant resistance genes that are effective against one race of the fungus [39].
Determination of resistance genes in genetically diverse rice material is important to identify new sources of blast resistance. DNA markers are used to identify resistance genes [26]. The most effective way to combat blast is to grow resistant rice cultivars. However, after a few years, the cultivar loses resistance due to the high variability of rice pathogens [40,41].
The selection of disease-resistant cultivars based on conventional breeding methods is complicated due to the difficulty of determining the presence of the desired allele of a particular gene. The use of molecular markers closely linked to genes that provide resistance to the pathogen greatly facilitates breeding work [42]. Therefore, the main factor in rice breeding for blast resistance is the selection of resistance donors based on the identification of genes that control this trait in rice and the creation of new cultivars with a high level of resistance [43]. Success in combat with rice blast has been achieved by introducing and pyramiding major blast resistance genes using MAS [6,7,8,9,10,11,12,13,14]. Most resistance genes provide rice immunity only to certain races of the pathogen. It is known that such genes, when pyramided in one genotype, can provide stable resistance to the disease.
Pyramiding of multiple resistance genes in a single genotype is the most efficient strategy in breeding for resistance to a variable population of Magnaporthe oryzae. Cultivars that combine combinations of 3-5 resistance genes are more resistant, as they show an increase and expansion of the spectrum of resistance to blast. Thus, the introduction of genes by combining (pyramiding) several blast resistance genes in one genotype is the most promising direction of rice selection [7,8,9,10,11,12,13,14,43]. Screening cultivars for the presence of genes for resistance to Magnaporthe oryzae, as well as the introduction and pyramid of resistance genes to the pathogen, contribute to a significant reduction in the time for rice improvement. Introduction of resistance genes into local cultivars is a preferred strategy in a rice breeding program for disease prevention. The aim of the work is to improve the resistance of rice to Magnaporthe oryzae based on the introduction and pyramid of the major blast resistance genes using MAS.
2. Results
2.1. Screening of Parental Rice Genotypes for Blast Resistance Genes Pi-1 and Pi-2
According to the electropherogram, the presence of the Pi-1 gene was identified in 4 samples 04636 (well 5), 03-27 (well 6), 04468 (well 7), 04470 (well 8) and they are heterozygous, 7668 (well 19) homozygous for Pi-1 because it repeated the DNA profile of the positive control C101-Lac (Figure 1A). Several samples (03-27 (well 6), 04468 (well 7), 04470 (well 8) and 7662 (well 14) homozygous for the Pi-2 gene were found, since they are identical in profile to the DNA of the differentiator cultivar C101-A51, the carrier Pi-2 gene (Figure 1A). The sizes of PCR products corresponding to the dominant genes Pi-1 and Pi-2 are 133 bp and 325 bp, respectively.
In Figure 1B shows that samples 7679 (well 5), 7683 (well 6), 7684 (well 7), 7689 (well 8) and 7695 (well 10) turned out to be homozygous for the Rm224 and Rm144 loci, which indicates the presence of a dominant rice blast resistance gene Pi-1. The following samples 7702 (well 12) and 7712 (well 13) are heterozygous for this gene. At the Rm527 and SSR140 loci, samples 7684 (well 7), 7690 (well 9), Bakansski (well 16), Aisaule (well 18) and Aru (well 19) showed DNA profiles like the positive control C101-A51 for the Pi-2 gene. In the remaining samples, the presence of genes in their genotypes in recessive states was revealed.
2.2. Screening of Parental Genotypes for the Presence of Pi-33 and Pi-ta Genes
The electropherogram of Figure 2A shows that samples 7662 (well 16) and 7663 (well 17) were identified as carriers of a homozygous dominant allele of the resistance gene Pi-33, since these genotypes repeat the resistance gene carrier profile of cv. C101-Lac. The heterozygous state of the Pi-ta gene was found in genotypes 03-27 (well 8) and 04468 (well 9). Also genotypes 25-14 (well 12) and 7664 (well 18) were homozygous for this locus (Figure 2A), because these genotypes match the DNA pattern of the positive control IR36. The sizes of PCR products corresponding to the dominant genes Pi-33 and Pi-ta are 198 bp and 400 bp, and for recessive genes are 300 bp and 600 bp, respectively. According to the results (Figure 2B), the Pi-33 resistance gene was found in a homozygous state in genotypes 7668 (well 6), 7679 (well 7), 7684 (well 9), 7686 (well 10), 7689 (well 11) and 7690 (well 12) compared to C101-Lac. The locus co-segregating with the dominant resistance gene Pi-ta was found in genotypes 7686 (well 10) and 7701 (well 15), (Figure 2B), while in genotypes 7667 (well 5) the Pi-ta gene is in a heterozygous state. The genotype 7686 (well 10) repeats the bands of two positive controls IR36 and C101-Lac, since it carries both Pi-33 and Pi-ta genes. Figure 2C shows that local rice cultivars did not have Pi-33 and Pi-ta resistance genes in their genotypes.
According to electropherogram data from 4 analyzed local high-yielding cultivars used as maternal lines in crossings, it turned out that three cultivars Bakanasski, Aisaule and Aru contain the Pi-2 gene. It should be noted that all 4 local genotypes do not contain the other 3 studied genes (Pi-1, Pi-33 and Pi-ta). Therefore, further experiments were conducted to introduce and pyramid these genes into local cultivars using hybridization and MAS techniques to improve rice blast resistance.
2.3. Screening of Rice Hybrids for the Presence of Pi-1 and Pi-2 Genes
Figure 2C shows that local rice cultivars do not have Pi-33 and Pi-ta resistance genes in their genotypes shows that 4 hybrids, 7824/Aisaule var. italica (well 11), 7824/Aisaule var. subuzbekistanica (well 12), 7698/Bakanasski var. subvulgaris (well 14), 7698/Bakanasski var. italica (well 15) are heterozygotes for Pi-2 gene (Figure 3A). The sizes of PCR products corresponding to the dominant genes Pi-1 and Pi-2 are 133 bp and 325 bp, for recessive genes are 120 bp 313 bp, respectively. The following 13 hybrids were found to contain Pi-1 gene in homozygous state in their genotypes: 7698/Aisaule var. suberythroseros (well 5), Aru/7702 var. erythroceros (well 6), Aisaule/7689 var. zeravschanica (well 7), Aru/0327 var. vulgaris (well 8), Aru/0327 var. subpyrocarpa (well 9), Bakanasski/7667 var. subjanthoseros (well 10), Bakanasski/7667 var. vulgaris (well 11), Bakanasski/7701 var. vulgaris (well 12), Aisaule/7664 var. italica (well 13), Aisaule/04470 var. subvulgaris (well 15), Aisaule/04470 var. italica (well 16), Aisaule/Don 7712 var. sundensis (well 17), Aisaule/Don 7712 var. italica (well 18) (Figure 3B).
The DNA profiles of the above hybrid lines were like those of differentiator cultivars C101-Lac and C101-A51, indicating successful transfer of blast resistance genes.
The electropherogram of Figure 3C demonstrates the heterozygous state of the hybrid Bakanasski/7683 var. subjanthoseros (well 6) for the Pi-1. The hybrid line Aru/04636 var. vulgaris was found to be dominant homozygote for this gene (well 14).
3.4. Screening of Rice Hybrids for the Presence of Pi-33 and Pi-ta Genes
Figure 4A shows that the hybrid Bakanasski/7684 var. subvulgaris (well 9) has the Pi-33 target gene in its genotype in a homozygous state, and the line Bakanasski/7668 var. subuzbekistanica (well 5) contains the dominant Pi-ta resistance gene. The sizes of PCR products corresponding to the dominant genes Pi-33 and Pi-ta are 198 bp and 400 bp, and for recessive genes are 300 bp and 600 bp, respectively.
The electropherogram in Figure 4B showed that the hybrid line Aisaule/7689 var. zeravschanica (well 7) is heterozygous for the Pi-33 resistance gene. Figure 4B demonstrates the heterozygous state of the Pi-ta allele found in 7 hybrid lines: 7698/Aisaule var. suberythroseros (well 5), Aru/7702 var. erythroceros (well 6), Aisaule/7689 var. zeravschanica (well 7), Aru/0327 var. vulgaris (well 8), Bakanasski/7667 var. vulgaris (well 9), Bakanasski/7701 var. vulgaris (well 10) and Aisaule/7664 var. italica (well 11). Electropherogram Figure 4C demonstrates that the line Aisaule/7663 var. italica (well 18) is heterozygous for the Pi-33 gene. The following lines Aru/7666 var. suberythroseros (well 13), Aru/Don 7712 var. Desvauxii semi-awned (well 15), and Aru/0327 var. subpyrocarpa (well 19) are heterozygous for the Pi-ta blast resistance gene.
The last electropherogram Figure 4D shows that the lines Bakanasski/Don 7712 (well 5), Aru/04468 var. subvulgaris (well 7), Bakanasski/7667 var. subjanthoseros (well 8) and Aru/Don 7712 var. erythroceros (well 13) have a heterozygous state of the Pi-ta allele in their genotypes. Consequently, on electrophoresis of PCR products of the Pi-33 and Pi-ta genes, the presence of both homozygous and heterozygous lines is observed in the hybrid offspring. All other analyzed samples showed DNA profile compliance with the negative control cultivar Flagman, which indicates the presence of a recessive allele of the Pi-33 and Pi-ta genes, respectively.
According to electropherograms of PCR products, 28 hybrid lines were identified whose genotypes contain genes for resistance to blast disease. It was revealed that 7 hybrids of them are pyramidal: 6 hybrids 7698/Aisaule var. suberythroseros, Aru/7702 var. erythroceros, Bakanasski/7667 var. subjanthoseros, Bakanasski/7667 var. vulgaris, Bakanasski/7701 var. vulgaris, Aisaule/7664 var. italica contain 2 blast resistance genes– Pi-1 and Pi-ta, and the hybrid Aisaule/7689 var. zeravschanica 3 genes are pyramided – Pi-1, Pi-33 and Pi-ta. The remaining hybrids have one of the studied genes each.
2.5. Phytopathological Test of Cultivars and Pyramided Lines for Resistance to Blast Disease
Phytopathological testing of local cultivars used as maternal lines showed that the cultivars Bakanasski, Aisaule and Aru are susceptible, and the cultivar Fatima is moderately resistant. Testing of the resistance of hybrid lines allowed us to identify level of resistance of hybrids to the phytopathogen. Figure 5 shows the reaction to disease of resistant control, susceptible control and improved pyramidal rice lines characterized as moderately resistant.
Among the seven pyramided lines selected, one line failed to grow under infectious nursery conditions. In results, of the remaining 6 pyramided lines, 2 lines - Bakanasski/7667 var. vulgaris and Aisaule/7664 var. italica (carrying 2 resistance genes Pi-1 and Pi-ta) and 1 line – Aisaule/7689 var. zeravschanica (carrying 3 genes – Pi-1, Pi-33 and Pi-ta) showed moderate resistance (MR) were developed (Table 1).
2.6. Assessment of the Productivity of Pyramided Rice Lines
The evaluation of the productivity created 6 lines of rice from 7 selected hybrid lines (1 line not grown under the infectious diseases nursery) (Aru/7702 var. erythroceros, Bakanasski/7667 var. subjanthoseros, Bakanasski/7667 var. vulgaris, Bakanasski/7701 var. vulgaris, Aisaule/7664 var. italica, Aisaule/7689 var. zeravschanica) performed by analysis of yield structure elements. The lines were evaluated based on main productivity trait, such as “weight of 1000 seeds, g.” Analysis of variance showed that pyramidal lines were inferior to the standard in yield characterization, since there was significant difference between the groups, p-value < 0.001 (Table 2).
According to the characteristic of yield productivity pyramidal lines are lower in comparison with the control cultivar Bakanasski, which is highly productive for Almaty region.
3. Discussion
There are most of the research works focused on the creation of pyramidal rice lines carrying two or more blast resistance genes using MAS and hybridization methods. MAS analysis facilitates the identification of genes, their introduction and pyramiding into new lines [44,45,46], that significantly speeds up rice breeding. The main goal of rice breeders in combating blast is to develop new cultivars and lines with broad-spectrum and durable resistance [7,8,9,10,11,12,13,14,43]. Rice lines carrying only one major resistance gene cannot withstand a changing population of the fungus [8]. In this case, the development of cultivars carrying not only one gene, but the pyramid of several genes of resistance to blast will help overcome this disease [47,48,49].
However, there is some research on assessing the resistance of created lines to phytopathogens in a natural or artificial infectious background which identify the actual degree of resistance of newly created lines. Jiang, H. et al. [8] developed several lines containing two blast resistance genes (Pi 37/Pid 3), (Pi 5/Pi54), (Pi 54/Pid3) and (Pigm/Pid 37), which showed high resistance to leaf and neck blast under natural infection conditions. Dubina et al. created pyramidal rice lines carrying five blast resistance genes (Pi-1, Pi-2, Pi-33, Pi-ta and Pi-b) in a homozygous state, and obtained cultivars (Pentagen, Magnate, Pirouette, Argamac, Kapitan, Lenaris) [48]. Kostylev, P.I. et al. [49] obtained pyramided rice lines with 2–6 genes of resistance to blast (Pi-1, Pi-2, Pi-33, Pi-ta, Pi-b and Pi-40). Evaluation of the resistance of pyramided lines to fungal infection showed that lines containing 2 or more genes are more resistant compared to monogenic lines. Ji, Z.J. et al. [50] created a pyramidal line with the Pi1 and Pi2 genes, which demonstrated resistance to blast. Positive results obtained by different researchers confirm the effectiveness of using these gene combinations (Pi-1, Pi-2, Pi-33 and Pi-ta) in improving rice protection against blast. The combination of several genes in one genotype has shown the greatest efficiency in the strategy of durable resistance of rice to blast [7,8,9,10,11,12,13,14,43]. However, there is evidence that rice lines carrying both individual resistance genes [44,51] and lines with pyramidal genes can have a broad spectrum of resistance to Magnaporthe oryzae [45,52]. This mainly depends on weather conditions and the degree of pathogenicity of the fungal strains.
It is well known that resistance of rice cultivars is enhanced by pyramiding of several resistance genes [51]. Wang, X. et al. [52] reported the creation of 74 lines by introgression and pyramiding of genes for resistance to biotic factors such as leafhopper (BPH), blast, fire blight (BLB) and aroma gene. These improved rice lines (ILs), containing combinations of 1 to 4 resistance genes to BPH, blast, BLB and aroma gene, were characterized by moderate or high resistance to multiple biotic stresses [52].
It should be noted that the introduction and pyramiding of several genes in lines sometimes does not lead to an increase in the resistance of the lines and can even sometimes have a negative effect. The introduction of several genes into one line, they may be incompatible, suppressing each other's action and leading to sensitivity to the phytopathogen or to a decrease in the yield of the lines [53].
The results of molecular screening of parental samples for the presence of genes showed that locally zoned varieties contain one of the four resistance genes studied – the Pi-2 gene, which is present in only three varieties (Bakanasski, Aisaule and Aru). Introgression of blast resistance genes into local zoned cultivars from differentiation varieties was successful. Seven pyramidal lines of rice are created. Three of them were found to show improved resistance to blast according to the results of the phytopathological test: 2 lines - F2 Bakanasski /7667 var. vulgaris and F2 Aisaule /7664 var. italica, containing 2 resistance genes (Pi-1 and Pi-ta) and 1-line F2 Aisaule /7689 var. zeravschanica, carrying 3 genes (Pi-1, Pi-33 and Pi-ta).
The resistance of pyramidal lines was assessed under infectious nursery conditions, which made it possible to identify the level of resistance of the created lines. The results of the phytopathological testing demonstrated that these three pyramidal lines with pyramidal 2 and 3 resistance genes showed moderately resistance to a mixture of fungal conidia of different isolates of M. oryzae, while 3 local varieties (Bakanasski, Aisaule and Aru) containing 1 resistance gene (Pi-2) – sensitive to phytopathogen. Thus, disease resistance of rice was enhanced by creating pyramidal lines. The test results showed that half of the pyramidal lines created were moderately resistant, and the other half were sensitive. This indicates that the level of resistance of the lines is revealed when encountering a phytopathogen in the conditions of an infectious nursery.
4. Materials and Methods
4.1. Plant Materials
The 35 genotypes of local and foreign selection from the rice collection of the Institute of Plant Biology and Biotechnology (IPBB, Almaty, Kazakhstan): 04636, 19-14, 57-14, 03-27, 04468, 04470, 04469, 95-06, 25-14, 04888, 212-05, 7653, 7662, 7663, 7664, 7666, 7667, 7668, 7679, 7683, 7684, 7686, 7689, 7690, 7695, 7698, 7701, 7702, 7712, 7824, Don 7712, Bakanasski, Fatima, Aisaule, Aru were used in the crossing as parents. 54 hybrid lines of the F2 generation (Bakanasski/7668 var. subuzbekistanica Kond., Bakanasski / 04470 var. vulgaris Koern., Bakanasski/7653 var. subuzbekistanica Kond., Bakanasski/7653 var. subjanthoseros Brsch., Bakanasski/7684 var. subvulgaris Brsch., Bakanasski/7684 var. vulgaris Koern., 7824/Aisaule var. italica Alef., 7824/Aisaule var. subuzbekistanica Kond., Aru/04468 var. subvulgaris Brsch., 7698/Bakanasski var. subvulgaris Brsch., 7698/Bakanasski var. italica Alef., Aru/7701 var. vulgaris Koern., Bakanasski/0327 var. vulgaris Koern., Aisaule/7679 var. subvulgaris Brsch., 7698/Aisaule var. italica Alef., 7698/Aisaule var. suberythroseros Kan., Aru/7702 var. erythroceros Koern., Aisaule/7689 var. zeravschanica Brsch., Aru/0327 var. vulgaris Koern., Aru/0327 var. subpyrocarpa Alef., Bakanasski/7667 var. subjanthoseros Brsch., Bakanasski/7667 var. vulgaris Koern., Bakanasski/7701 var. vulgaris Koern., Aisaule/7664 var. italica Alef., Aisaule/7664 var. subvulgaris Brsch., Aisaule/04470 var. subvulgaris Brsch., Aisaule/04470 var. italica Alef., Aisaule/Don 7712 var. sundensis Koern., Aisaule/Don 7712 var. italica Alef., Fatima/7695 var. italica Alef., Fatima/7695 var. breviaristata Vav., Bakanasski/7683 var. subjanthoseros Brsch., 7698/Aru var. erythroceros Koern., 7698/Aru var. suberythroseros Kan., Bakanasski/Don 7712, Bakanasski/Don 7712 var. Desvauxii Koern., Aru/212-05 var. vulgaris Koern., Aru/7668 var. vulgaris Koern., Aisaule/04468 var. italica Alef., Aru/04636 var. vulgaris Koern., Bakanasski/7690 var. subjanthoseros Brsch., Bakanasski/7690 var. vulgaris Koern., Bakanasski/04468 var. subvulgaris Brsch., Bakanasski/04468 var. vulgaris Koern., Aru/7666 var. suberythroseros Kan., Fatima/7653 var. persica Kan., Fatima/7689 var. italica Alef., Aru/Don 7712 var. erythroceros Koern., Aru/Don 7712 var. Desvauxii Koern., Aru/Don 7712 var. vulgaris Koern., Aru/Don 7712 var. Desvauxii Koern. semi-awned, ♀ Aru/7683 var. janthoceros Koern., Aru/7683 var. vulgaris Koern., Aisaule/7663 var. italica Alef.) obtained by crossing were used for analysis.
4.2. Hybridization Method
Hybridization was carried out using pneumocastration (three-channel pneumocastration) and the "TWEL" plant pollination method [54]. 31 genotypes were used as donors, the genome of which contains genes of resistance to rice blast (Pi-1, Pi-2, Pi-33 and Pi-ta), and recipients were highly productive zoned local cultivars, such as Bakanasski, Fatima, Aisaule and Aru.
4.3. DNA Extraction and Multiplex PCR Analysis
Genomic DNA was extracted from two-week-old seedlings using the cetyltrimethylammonium bromide (CTAB) method [55]. CTAB reagent was obtained from Pan-Reac AppliChem GmbH (Darmstadt, Germany). All PCR analysis were performed using a T100 thermal cycler (BioRad, Germany). Multiplex PCR analysis was performed by adding several primers in PCR mix to determine the presence of two or more genes in a genotype. Each microtube contained 15 µl of PCR reaction mixture consisting of 40–50 ng of template DNA, 1.2 µl of each forward and reverse primer, 1.2 µl of Taq buffer, 0.05 mmol/L dNTPs and 1.0 U of Taq polymerase. The following PCR conditions were used: 94°C for 5 min; 35 cycles of 35 sec at 94°C, 45 sec at 60°C, and 30 sec at 72°C; and an extension of 5 min at 72°C. SSR (simple sequence repeat) markers were used to identify genes. The list of SSR markers (Syntol, Russia) for Pi complex genes is given in Table 3. The annealing temperature is the same for all primers, 60º C.
The following genotypes were used as differentiator cultivars that are carriers of resistance genes: C101-Lac (+) for Pi-1, Pi-33 genes, C101-A51 (+) for Pi-2 gene and IR 36 (+) for Pi-ta gene. The cultivar Flagman (-) was used as a negative control for all studied genes.
4.5. Gel Electrophoresis
The amplification products were separated in 8% polyacrylamide gel (PAGE) in 1.0× TBE buffer by vertical electrophoresis chamber (Helicon, Russia). Electrophoretic gels were examined using the GelDoc XR (Bio-Rad, USA). The GelAnalyzer program was used to analyze gels.
4.6. Phytopathological Screening
Testing of 54 hybrid rice lines for resistance to Magnaporthe oryzae populations was carried out in 2023 in the artificial infectious nursery at the FSBSI “Federal Scientific Rice Centre” (Russia, Krasnodar). The most resistant cv. Avangard served as a positive control, and the highly susceptible cv. Pobeda 65 was used as a negative control.
Rice plants were infected with the fungal culture at the booting stage by spraying with a suspension of a mixture of conidia of 15 isolates of M. oryzae.
The degree of plant damage (in percentage) was considered on the 14th day after inoculation, according to the express method for assessing rice cultivars resistance to blast. The assessment was carried out considering the type of reaction (in points) on a ten-point scale (0-9) of the Standard Evaluation System for Rice [56]. The degree of resistance was indicated using the following designations: resistant lines - R, moderately resistant - MR and susceptible - S.
The intensity of disease development (IDD, %) was calculated using the formula:
IDD = ∑ (a*b)/ n*9,
where - ∑ (a*b) – sum products of the number of infected plants multiplied by the corresponding damage score, n - the number of recorded plants, pieces.
4.7. Analysis of Structural Elements of Yield of Hybrid Rice Lines
Evaluation of the productivity of hybrid lines in comparison with the standard cv. Bakanasski was carried out according to the biometric parameters of the elements of yield structure (“bushiness, pc”, “plant height, cm”, “panicle length, cm”, “number of grains from the main panicle, pc”, “weight of seeds from the main panicle, g”, “weight of 1000 seeds, g” and etc.) [57]
4.8. Statistical Analyses
To characterize the productivity of hybrid lines in comparison with the cv. Bakanasski, which is the standard for the Almaty region, statistical indicators such as the mean value, standard deviation for the main traits characterizing rice productivity, “weight of 1000 seeds, g” were used. Analysis of variance (one-way ANOVA) was performed to compare the mean values of the data groups. Data processing was carried out in Microsoft Excel 2021 using a data analysis package.
5. Conclusions
Breeding programs successfully use both traditional breeding methods and modern MAS analysis tools involving a wide range of genetic diversity. In the context of global climate change, the creation of elite rice lines and varieties resistant to adverse environmental factors is a necessity to meet the ever-increasing demand for food products. It is known that over time, most varieties that have one resistance gene in their genotype cannot withstand changing fungal races and lose resistance, which leads to a decrease in rice yield. In connection with the above-described problem, increasing rice resistance by pyramiding several resistance genes in one genotype is crucial for stable rice production. The created pyramidal lines are of great value in rice breeding for improving the resistance of local varieties.
The use of modern MAS technologies guarantees the acceleration of rice breeding and hence the introduction to the market of competitive rice cultivars with durable immunity to disease and other stress factors. Microsatellite markers are more often used to assess polymorphism and genetic diversity of samples [58]. The created new pyramided lines of rice that are resistant to phytopathogens will be used in rice breeding to create cultivars, which increases the yield and competitiveness of rice in the market and promotes the export of rice products. These pyramidal lines will be used as a source of resistance genes in the breeding process and for germplasm exchange.
Rice is the food source for most people on the planet. The world's population is increasing every year. Improving rice resistance to blast by introgression and pyramiding of genes will help maintain rice yields during epiphytotic periods, which contributes to sustainable agricultural development and achieving zero hunger according to the UN program [https://www.un.org/sustainabledevelopment/sustainable-development-goals/].
Author Contributions
Supervision, project administration and funding acquisition, U.B.; Conceptualization and writing-original draft preparation, M.D., A.A. and D.E.; Investigation and methodology, M.D. and B.Kh.; Software, Z.Z. and M.D.; Writing review and editing, A.A. and Zh.Zh. All authors have read and agreed to the published version of the manuscript.
Funding
This research has funded by the Science Committee of the Ministry of Education and Science of the Republic of Kazakhstan Grant AP14869300.
Data Availability Statement
All data used in this manuscript are presented in the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khush, G.S.; Jena, K.K. Current status and future prospects for research on blast resistance in rice (Oryza sativa L.). In Wang G.L., Valent B. (Eds.). Advances in genetics, genomics and control of rice blast disease. Springer, Dordrecht., 2009, 1-10. [CrossRef]
- Liu, J.; Wang, X.; Mitchell, T.; Hu, Y.; Liu, X.; Dai, L.; Wang, G.L. Recent progress and understanding of the molecular mechanisms of the rice-Magnaporthe oryzae interaction. Molecular Plant Pathology. 2010, 11, 419-427. [CrossRef]
- Li, Y.B.; Wu, C.J.; Jiang, G.H.; Wang, L.Q.; He, Y.Q. Dynamic analyses of rice blast resistance for the assessment of genetic and environmental effects. Plant Breeding. 2007, 126, 541-547. [CrossRef]
- Zhang, T.; Huang, L.; Wang, Y.; Wang, W.; Zhao, X.; Zhang, S.; Zhang, J.; Hu, F.; Fu, B.; Li, Z. Differential transcriptome profiling of chilling stress response between shoots and rhizomes of Oryza longistaminata using RNA sequencing. PLoS One. 2017, 12(11), e0188625. 0188625. [CrossRef] [PubMed]
- Xiao, W.; Yang, Q.; Huang, M.; Guo, T.; Liu, Y.; Wang, J.; Yang, G.; Zhou, J.; Yang, J.; Zhu, X.; Chen, Zh.; Wang, H. Improvement of rice blast resistance by developing monogenic lines, two-gene pyramids and three-gene pyramid through MAS. Rice. 2019, 12(78). [CrossRef]
- Liu, Z.; Zhu, Y.; Shi, H.; Qiu, J.; Ding, X.; Kou, Y. Recent Progress in Rice Broad-Spectrum Disease Resistance. Int. J. Mol. Sci. 2021, 22, 11658. [CrossRef]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Latif, M.A. Marker-assisted introgression of broad-spectrum blast resistance genes into the cultivated MR219 rice variety. J. Sci. Food Agric. 2017, 97, 2810–2818. [CrossRef]
- Jiang, H.; Li, Z.; Liu, J.; Shen, Z.; Gao, G.; Zhang, Q.; He, Y. Development and evaluation of improved lines with broad-spectrum resistance to rice blast using nine resistance genes. Rice. 2019, 12(29). [CrossRef]
- Jain, P.; Dubey, H.; Singh, P.K.; Solanke, A.U.; Singh, A.K.; Sharma, T.R. Deciphering signaling network in broad spectrum near isogenic lines of rice resistant to Magnaporthe oryzae. Sci. Rep. 2019, 9, 16939. [CrossRef]
- Wu, Y.; Xiao, N.; Chen, Y.; Yu, L.; Pan, C.; Li, Y.; Zhang, X.; Huang, N.; Ji, H.; Dai, Z.; Chen, X.; Li, A. Comprehensive evaluation of resistance effects of pyramiding lines with different broad-spectrum resistance genes against Magnaporthe oryzae in rice (Oryza sativa L.). Rice. 2019, 12, 1–13. [CrossRef]
- Yang, D.; Tang, J.; Yang, D.; Chen, Y.; Ali, J.; Mou, T. Improving rice blast resistance of Feng39S through molecular marker-assisted backcrossing. Rice. 2019, 12, 1–16. [CrossRef]
- Wang, L.; Zhao, L.; Zhang, X.; Zhang, Q.; Jia, Y.; Wang, G.; Li, S.; Tian, D.; Li, W.H.; Yang, S. Large-scale identification and functional analysis of NLR genes in blast resistance in the Tetep rice genome sequence. Proc. Natl. Acad. Sci. USA. 2019, 116, 18479–18487. [CrossRef]
- Ramalingam, J.; Palanisamy, S.; Alagarasan, G.; Renganathan, V.G.; Ramanathan, A.; Saraswathi, R. Improvement of stable restorer lines for blast resistance through functional marker in rice (Oryza sativa L.). Genes. 2020, 11, 1266. [CrossRef]
- Peng, M.; Lin, X.; Xiang, X.; Ren, H.; Fan, X.; Chen, K. Characterization and evaluation of transgenic rice pyramided with the Pi Genes Pib, Pi25 and Pi54. Rice. 2021, 14, 1–14. [CrossRef]
- Devanna, N.B.; Vijayan, J.; Sharma, T.R. The blast resistance gene Pi54 of cloned from Oryza officinalis interacts with Avr-Pi54 through its novel non-LRR domains. PLoS One. 2014, 9(8), 104-40. [CrossRef]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G.; Li, Q.; Zhang, J.; Wu, S.; Milazzo, J.; Mao, B.; Wang, E.; Xie, H.; Tharreau, D.; He, Z. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science. 2017, 355, 962–965. [CrossRef]
- Xie, Z.; Yan, B.; Shou, J.; Tang, J.; Wang, X.; Zhai, K.; Liu, J.; Li, Q.; Luo, M.; Deng, Y.; He, Z. A nucleotide-binding site-leucine-rich repeat receptor pair confers broad-spe;ctrum disease resistance through physical association in rice. Philosophical Transactions of the Royal Society B. 2019, 374, 20180308. [CrossRef]
- Zhao, H.; Wang, X.; Jia, Y.; Minkenberg, B.; Wheatley, M.; Fan, J.; Jia, M.H.; Famoso, A.; Edwards, J.D.; Wamishe, Y.; Valent, B.; Wang, G.L.; Yang, Y. The rice blast resistance gene Ptr encodes an atypical protein required for broad-spectrum disease resistance. Nat. Commun. 2018, 9, 2039. [CrossRef]
- Meng, X.; Xiao, G.; Telebanco-Yanoria, M.J.; Siazon, P.M.; Padilla, J.; Opulencia, R.; Bigirimana, J.; Habarugira, G.; Wu, J.; Li, M.; Wang, B.; Lu, G.; Zhou, B. The broad-spectrum rice blast resistance (R) gene Pita2 encodes a novel R protein unique from Pita. Rice. 2020, 13, 1–15. [CrossRef]
- Chen, D.H.; Zeigler, R.S.; Ahn, S.W.; Nelson, R.J. Phenotypic characterization of the rice blast resistance gene Pi-2(t). Plant Disease. 1996, 80, 52–56. https://www.apsnet.org/publications/plantdisease/backissues/Documents/1996Abstracts/PD_80_52.htm.
- Amante-Bordeos, A.; Sitch, L.A.; Nelson, R.; Dalmacio, R.D.; Oliva, N.P.; Aswidinnoor, H.; Leung, H. Transfer of bacterial blight and blast resistance from the tetraploid wild rice Oryza minuta to cultivated rice Oryza sativa. Theor. Appl. Genet. 1992, 84(3/4), 345–354. [CrossRef]
- Deng Y., Zhu X., Shen Y., He Z. Genetic characterization and fine mapping of the blast resistance locus Pigm(t) tightly linked to Pi2 and Pi9 in a broad-spectrum resistant Chinese variety. Theor. Appl. Genet., 2006. 113, 705–713. [CrossRef]
- Deng Y., Zhai K., Xie Z., Yang D., Zhu X., Liu J., Wang X., Qin P., Yang Y., Zhang G., Li Q., Zhang J., Wu S., Milazzo J., Mao B., Wang E., Xie H., Tharreau D., He Z. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science, 2017. 355, 962–965. [CrossRef]
- Liang, Y.; Yang, T.; Tan, L.; Wen, T.; Wu, J.; Jiang, N.; Li, Z.; Dai, L.; Wang, G.; Liu, X. Development of the molecular marker tightly-linked with the broad-spectrum blast resistance Pigm and its breeding practice in rice. Hybrid Rice, 2013, 28, 63–74, (In Chinese with English Abstract). [CrossRef]
- Jia, Y.; Wang, Z.; Singh, P. Development of Dominant Rice Blast Pi-ta Resistance Gene Markers. Crop Sci. 2002, 42, 2145–2149. [CrossRef]
- Mukhina, Zh.M. The use of DNA markers to study the genetic diversity of plant resources, Krasnodar: Prosveshenie-South, 2008, 98 p.
- Xu, X.; Chen, H.; Fujimura, T.; Kawasaki, S. Fine mapping of a strong QTL of field resistance against rice blast, Pikahei-1(t), from upland rice Kahei, utilizing a novel resistance evaluation system in the greenhouse. Theoretical and applied genetics. 2008, 117, 997-1008. [CrossRef]
- Hua, L.; Wu, J.; Chen, C.; Wu, W.; He, X.; Lin, F.; Wang, L.; Ashikawa, I.; Matsumoto, T.; Wang, L.; Pan, Q. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast. Teor. Appl. Genet., 2012. 125(5), 1047–1055. [CrossRef]
- Qu, S.; Liu, G.; Zhou, B.; Bellizzi, M.; Zeng, L.; Dai, L.; Han, B.; Wang, G.L. The broad-spectrum blast resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice. Genetics, 2006. 172, 1901–1914. [CrossRef]
- Chen, J.; Shi, Y.; Liu, W.; Chai, R.; Fu, Y.; Zhuang J., Wu, J. A Pid3 allele from rice cultivar Gumei2 confers resistance to Magnaporthe oryzae. J. Genet. Genomics, 2011. 38, 209–216. [CrossRef]
- He, N.; Huang, F.; Yu, M.; Zhu, Y.; Li, Q.Q.; Yang D. Analysis of a rice blast resistance gene Pita-Fuhui2663 and development of selection marker. Scientific Reports, 2022. 12, 14917. [CrossRef]
- Yang, D.; Li, S.; Lu, L.; Fang, J.; Wang, W.; Cui, H.; Tang, D. Identification and application of the Pigm-1 gene in rice disease resistance breeding. Plant Biol (Stuttg). 2020. 22(6), 1022-1029. [CrossRef]
- Ashkani, S.; Rafii, M.Y.; Rahim, H.A.; Latif, M.A. Genetic dissection of rice blast resistance by QTL mapping approach using an F3 population. Molecular Biology Reports. 2013, 40, 2503-2515. PMid:23203411. [CrossRef]
- Ballini, E.; Morel, J.B.; Droc, G.; Price, A.; Courtois, B.; Notteghem, J.L.; Tharreau, D.A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance. Molecular plant-microbe interactions: MPMI. 2008, 21, 859-868. PMid:18533827. [CrossRef]
- Padmavathi, G.; Ram, T.; Satyanarayana, K.; Mishra, B. Identification of blast (Magnaporthe grisea) resistance genes in rice. Current Science. 2005, 88, 628-630. https://www.jstor.org/stable/24110264.
- Sharma, R.C.; Shrestha, S.M.; Pandey, M.P. Inheritance of blast resistance and associated microsatellite markers in rice cultivar ‘Laxmi’. Journal of Phytopathology. 2007, 155, 749-753. [CrossRef]
- Mohanty, C.R.; Gangopadhyay, S. Testing of blast resistance in F2 rice seedlings in different doses of nitrogen and seasons. Annals of the Phytopathological Society of Japan. 1982, 48(5), 648-658. [CrossRef]
- Flores-Gaxiola, J.A.; Nuque, F.L.; Crill, J.P.; Khush, G.S. Inheritance of blast Pyricularia oryzae resistance in rice. International Rice Research Newsletter. 1983, 8, 5-6. https://zenodo.org/records/7320914.
- Mackill, D.J.; Bonman, J.M. Inheritance of Blast Resistance in Near-Isogenic Lines of Rice. Phytopathology. 1992, 82, 746-749. https://www.apsnet.org/publications/phytopathology/backissues/Documents/1992Articles/Phyto82n07_746.PDF.
- Bonman, J.M.; Khush, G.S.; Nelson, R.J. Breeding rice for resistance to pests. Annual Review of Phytopathology. 1992, 30, 507-528. [CrossRef]
- Lang, N.T.; Luy, T.T.; Ha, P.T.T.; Buu, B.C. Monogenic lines resistance to blast disease in rice (Oryza sativa L.) in Vietnam. Global Science Research Journals. 2019, 1(7), 439–448. http://www.globalscienceeresearchjournals.org/.
- Hasan, N.; Choudhary, S.; Naaz, N.; Sharma, N.; Laskar, R.A. Recent advancements in molecular marker assisted selection and applications in plant breeding programmers. Journal of Genetic Engineering and Biotechnology. 2021, 19, 128. [CrossRef]
- Zelensky, G.L. Rice: biological foundations of breeding and agricultural technology. Monography. Krasnodar. KubSAU, 2016, 177.
- Li, W.; Lei, C.L.; Cheng, Z.J.; Jia, Y.L.; Huang, D.Y.; Wang, J.L.; Wang, J.K.; Zhang, X.; Su N.; Guo, X.P.; Zhai, H.Q.; Wan, J.M. Identification of SSR markers for a broad-spectrum blast resistance gene Pi20(t) for marker-assisted breeding. Mol. Breed. 2008, 22, 141-149. [CrossRef]
- Peng, P.; Jiang, H.; Luo, L.; Ye, C.; Xiao, Y. Pyramiding of Multiple Genes to Improve Rice Blast Resistance of Photo-Thermo Sensitive Male Sterile Line, without Yield Penalty in Hybrid Rice Production. Plants. 2023, 12, 1389. [CrossRef]
- Ospanova, A.; Mynbayeva, D.; Turganova, Ch.; Usenbekov, B.; Amirova, A.; Berkimbay, Kh.; Zhunusbayeva, Zh.; Utepbergenov, E. Molecular screening for the presence of Magnaporthe oryzae resistance Pi-b gene in rice hybrids. International Scientific Forum “Modern Trends in Sustainable Development of Biological Sciences”. BIO Web of Conferences. 2024, 100, 03012. [CrossRef]
- Amirova A.; Usenbekov, B.; Berkimbay, Kh.; Mynbayeva, D.; Atabayeva, S.; Baiseitova, G.; Meldebekova, A.; Zhunusbayeva, Zh.; Kenzhebayeva, S.; Mukhambetzhanov, S. Selection of rice breeding lines for resistance to biotic and abiotic stresses. Brazilian Journal of Biology. 2024, 84, e282495. [CrossRef]
- Dubina, E.; Kostylev, P.; Ruban, M.; Lesnyak, S; Krasnova, E.; Azarin, K. Rice Breeding in Russia Using Genetic Markers. Plants. 2020, 9(11), 1580. [CrossRef]
- Kostylev PI, Krasnova EV, Red'kin AA, et al. Combination of rice blast resistance genes in the genotypes of Russian rice varieties with the use of marker assisted selection. Ecological genetics. 2017, 15(3), 54-63. [CrossRef]
- Ji, Z.J.; Yang, S.D.; Zeng,Y.X.; Liang,Y., YANG, C.D.; Qian, Q. Pyramiding blast, bacterial blight and brown planthopper resistance genes in rice restorer lines. Journal of Integrative Agriculture. 2016, 15(7), 1432-1440. [CrossRef]
- Hittalmani, S.; Parco, A.; Mew, T.; Zeigler, R.; Huang, N. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor. Appl. Genet. 2000, 100, 1121–1128. [CrossRef]
- Wang, X.; Guo, X.; Ma, X.; Luo, L.; Fang, Y.; Zhao, N.; Han, Y.; Wei, Z.; Liu, F.; Qin, B..; Li, R. Development of New Rice (Oryza. sativa L.) Breeding Lines through Marker-Assisted Introgression and Pyramiding of Brown Planthopper, Blast, Bacterial Leaf Blight Resistance, and Aroma Genes. Agronomy. 2021, 11(12), 2525. [CrossRef]
- Xiao, N.; Wu, Y.; Li, A. Strategy for Use of Rice Blast Resistance Genes in Rice Molecular Breeding. Rice Science. 2020, 27(4): 263í277. [CrossRef]
- Los, G.D. Rice hybridization technique. Rice growing. 2007, 10, 42-51 [Los' G. D. Metodika gibridizacii risa. G. D. Los'. Risovodstvo. 2007, 10, 42-51].
- Williams, C.E.; Ronald, P.C. PCR template-DNA isolated quickly from monocot and dicot leaves without tissue homogenization. Nucleic Acids Research. 1994, 22(10), 1917-1918. PMid:8208619. [CrossRef]
- International Rice Research Institute. Standard evaluation system (SES) for Rice, 5th edn. International Rice Research Institute, Manila, 2013, 14-15. http://www.knowledgebank.irri.org/images/docs/rice-standard-evaluation-system.pdf.
- Guidelines for the Conduct of Test for Distinctiveness, Uniformity and Stability On (Oryza sativa L.). Plant Variety Journal of India. 2007, 1(1), 25. https://www.researchgate.net/publication/332736023.
- Zargar, M.; Dyussibayeva, E.; Orazov, A.; Zeinullina, A.; Zhirnova, I.; Yessenbekova, G.; Rysbekova, A. Microsatellite-Based Genetic Diversity Analysis and Population Structure of Proso Millet (Panicum miliaceum L.) in Kazakhstan. Agronomy 2023, 13, 2514. [CrossRef]
Figure 1.
Electropherogram of PCR products of samples for the presence of the Pi-1 and Pi-2 genes. (a): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51 (control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – 04636, 6 – 03-27, 7 – 04468, 8 – 04470, 9 – 04469, 10 – 25-14, 11 – 04888, 12 – 212-05, 13 – 7653, 14 – 7662, 15 – 7663, 16 – 7664, 17 – 7666, 18 – 7667, 19 – 7668; (b): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51 (control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – 7679, 6 – 7683, 7 – 7684, 8 – 7689, 9 – 7690, 10 – 7695, 11 – 7701, 12 – 7702, 13 – 7712, 14 – 7824, 15 – Don 7712, 16 – Bakanasski, 17 – Fatima, 18 – Aisaule, 19 – Aru.
Figure 1.
Electropherogram of PCR products of samples for the presence of the Pi-1 and Pi-2 genes. (a): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51 (control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – 04636, 6 – 03-27, 7 – 04468, 8 – 04470, 9 – 04469, 10 – 25-14, 11 – 04888, 12 – 212-05, 13 – 7653, 14 – 7662, 15 – 7663, 16 – 7664, 17 – 7666, 18 – 7667, 19 – 7668; (b): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51 (control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – 7679, 6 – 7683, 7 – 7684, 8 – 7689, 9 – 7690, 10 – 7695, 11 – 7701, 12 – 7702, 13 – 7712, 14 – 7824, 15 – Don 7712, 16 – Bakanasski, 17 – Fatima, 18 – Aisaule, 19 – Aru.

Figure 2.
Electropherogram of PCR products of samples for the presence of Pi-33 and Pi-ta genes. (a): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci); 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci); 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci); 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci); 5 – 04636; 6 – 19-14; 7 – 57-14; 8 – 03-27; 9 – 04468; 10 – 04470; 11 – 04469; 12 – 25-14; 13 – 04888; 14 – 212-05; 15 – 7653; 16 – 7662; 17 – 7663; 18 – 7664; 19 – 7666; (b): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR 36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – 7667, 6 – 7668, 7 – 7679, 8 – 7683, 9 – 7684, 10 – 7686, 11 – 7689, 12 – 7690, 13 – 7695, 14 – 7698, 15 – 7701, 16 – 7702, 17 – 7712, 18 – 7824; (c): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Don 7712, 6 – Bakanasski, 7 – Fatima, 8 - Aisaule, 9 – Aru.
Figure 2.
Electropherogram of PCR products of samples for the presence of Pi-33 and Pi-ta genes. (a): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci); 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci); 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci); 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci); 5 – 04636; 6 – 19-14; 7 – 57-14; 8 – 03-27; 9 – 04468; 10 – 04470; 11 – 04469; 12 – 25-14; 13 – 04888; 14 – 212-05; 15 – 7653; 16 – 7662; 17 – 7663; 18 – 7664; 19 – 7666; (b): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR 36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – 7667, 6 – 7668, 7 – 7679, 8 – 7683, 9 – 7684, 10 – 7686, 11 – 7689, 12 – 7690, 13 – 7695, 14 – 7698, 15 – 7701, 16 – 7702, 17 – 7712, 18 – 7824; (c): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Don 7712, 6 – Bakanasski, 7 – Fatima, 8 - Aisaule, 9 – Aru.

Figure 3.
Electropherogram of PCR products of hybrids for the presence of Pi-1 and Pi-2 genes. (a): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51(control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – Bakanasski/7668 var. subuzbekistanica, 6 – Bakanasski/04470 var. vulgaris, 7 – Bakanasski/7653 var. subuzbekistanica, 8 – Bakanasski/7653 var. subjanthoseros, 9 – Bakanasski/7684 var. subvulgaris, 10 – Bakanasski/7684 var. vulgaris, 11 – 7824/Aisaule var. italica, 12 – 7824/Aisaule var. subuzbekistanica, 13 – Aru/04468 var. subvulgaris, 14 – 7698/Bakanasski var. subvulgaris, 15 – 7698/Bakanasski var. italica, 16 – Aru/7701 var. vulgaris, 17 – Bakanasski/0327 var. vulgaris, 18 – Aisaule/7679 var. subvulgaris, 19 –7698/Aisaule var. italica (b): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51(control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – 7698/Aisaule var. suberythroseros, 6 – Aru/7702 var. erythroceros, 7 – Aisaule/7689 var. zeravschanica, 8 – Aru/0327 var. vulgaris, 9 – Aru/0327 var. subpyrocarpa, 10 – Bakanasski/7667 var. subjanthoseros, 11 – Bakanasski/7667 var. vulgaris, 12 – Bakanasski/7701 var. vulgaris, 13 – Aisaule/7664 var. italica, 14 – Aisaule/7664 var. subvulgaris, 15 – Aisaule/04470 var. subvulgaris, 16 – Aisaule/04470 var. italica, 17 – Aisaule/Don 7712 var. sundensis, 18 – Aisaule/Don 7712 var. italica, 19 – Fatima/7695 var. italica (c): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51 (control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – Fatima/7695 var. breviaristata, 6 – Bakanasski/7683 var. subjanthoseros, 7 – 7698/Aru var. erythroceros, 8 – 7698/Aru var. suberythroseros, 9 – Bakanasski/Don 7712, 10 – Bakanasski/Don 7712 var. Desvauxii, 11 – Aru/212-05 var. vulgaris, 12 – Aru/7668 var. vulgaris, 13 – Aisaule/04468 var. italica, 14 – Aru/04636 var. vulgaris, 15 – Bakanasski/7690 var. subjanthoseros, 16 – Bakanasski/7690 var. vulgaris, 17 – Bakanasski/04468 var. subvulgaris, 18 – Bakanasski/04468 var. vulgaris.
Figure 3.
Electropherogram of PCR products of hybrids for the presence of Pi-1 and Pi-2 genes. (a): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51(control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – Bakanasski/7668 var. subuzbekistanica, 6 – Bakanasski/04470 var. vulgaris, 7 – Bakanasski/7653 var. subuzbekistanica, 8 – Bakanasski/7653 var. subjanthoseros, 9 – Bakanasski/7684 var. subvulgaris, 10 – Bakanasski/7684 var. vulgaris, 11 – 7824/Aisaule var. italica, 12 – 7824/Aisaule var. subuzbekistanica, 13 – Aru/04468 var. subvulgaris, 14 – 7698/Bakanasski var. subvulgaris, 15 – 7698/Bakanasski var. italica, 16 – Aru/7701 var. vulgaris, 17 – Bakanasski/0327 var. vulgaris, 18 – Aisaule/7679 var. subvulgaris, 19 –7698/Aisaule var. italica (b): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51(control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – 7698/Aisaule var. suberythroseros, 6 – Aru/7702 var. erythroceros, 7 – Aisaule/7689 var. zeravschanica, 8 – Aru/0327 var. vulgaris, 9 – Aru/0327 var. subpyrocarpa, 10 – Bakanasski/7667 var. subjanthoseros, 11 – Bakanasski/7667 var. vulgaris, 12 – Bakanasski/7701 var. vulgaris, 13 – Aisaule/7664 var. italica, 14 – Aisaule/7664 var. subvulgaris, 15 – Aisaule/04470 var. subvulgaris, 16 – Aisaule/04470 var. italica, 17 – Aisaule/Don 7712 var. sundensis, 18 – Aisaule/Don 7712 var. italica, 19 – Fatima/7695 var. italica (c): M – Molecular marker 100 bp; 1 – C101-Lac (control "+" for the Pi-1 gene at the Rm144 and RM224 loci), 2 – Flagman (control "-" for the Pi-1 gene at the Rm144 and RM224 loci), 3 – C101-A51 (control "+" for the Pi-2 gene at loci RM-527 and SSR-140), 4 – Flagman (control "-" for the Pi-2 gene at loci RM-527 and SSR-140), 5 – Fatima/7695 var. breviaristata, 6 – Bakanasski/7683 var. subjanthoseros, 7 – 7698/Aru var. erythroceros, 8 – 7698/Aru var. suberythroseros, 9 – Bakanasski/Don 7712, 10 – Bakanasski/Don 7712 var. Desvauxii, 11 – Aru/212-05 var. vulgaris, 12 – Aru/7668 var. vulgaris, 13 – Aisaule/04468 var. italica, 14 – Aru/04636 var. vulgaris, 15 – Bakanasski/7690 var. subjanthoseros, 16 – Bakanasski/7690 var. vulgaris, 17 – Bakanasski/04468 var. subvulgaris, 18 – Bakanasski/04468 var. vulgaris.

Figure 4.
Electropherogram of PCR products of hybrids for the presence of Pi-33 and Pi-ta genes. (а): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Bakanasski/7668 var. subuzbekistanica, 6 – Bakanasski/04470 var. vulgaris, 7 – Bakanasski/7653 var. subuzbekistanica, 8 – Bakanasski/7653 var. subjanthoseros, 9 – Bakanasski/7684 var. subvulgaris, 10 – Bakanasski/7684 var. vulgaris, 11 – 7824/Aisaule var. italica, 12 – 7824/Aisaule var. subuzbekistanica, 13 – 7698/Bakanasski var. subvulgaris, 14 – 7698/Bakanasski var. italica, 15 – Aru/7701 var. vulgaris, 16 – Bakanasski/0327 var. vulgaris, 17 – Aisaule/7679 var. subvulgaris, 18 – 7698/Aisaule var. italica (в): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – 7698/Aisaule var. suberythroseros, 6 – Aru/7702 var. erythroceros, 7 – Aisaule/7689 var. zeravschanica, 8 – Aru/0327 var. vulgaris, 9 – Bakanasski/7667 var. vulgaris, 10 – Bakanasski/7701 var. vulgaris, 11 – Aisaule/7664 var. italica, 12 – Aisaule/7664 var. subvulgaris, 13 – Aisaule/04470 var. subvulgaris, 14 – Aisaule/04470 var. italica, 15 – Aisaule/Don 7712 var. sundensis, 16 – Aisaule/Don 7712 var. italica, 17 – Fatima/7695 var. breviaristata, 18 – Bakanasski/7683 var. subjanthoseros (с): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Aru/212-05 var. vulgaris, 6 – Aru/7668 var. vulgaris, 7 – Aisaule/04468 var. italica, 8 – Aru/04636 var. vulgaris, 9 – Bakanasski/7690 var. subjanthoseros, 10 – Bakanasski/7690 var. vulgaris, 11 – Bakanasski/04468 var. subvulgaris, 12 – Bakanasski/04468 var. vulgaris, 13 – Aru/7666 var. suberythroseros, 14 – Fatima/7653 var. persica, 15 – Aru/Don 7712 var. Desvauxii semi-awned, 16 – Aru/7683 var. janthoceros, 17 – Aru/7683 var. vulgaris, 18 – Aisaule/7663 var. italica, 19 – Aru/0327 var. subpyrocarpa (d): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Bakanasski/Don 7712, 6 – Fatima/7689 var. italica, 7 – Aru/04468 var. subvulgaris, 8 – Bakanasski/7667 var. subjanthoseros, 9 – Bakanasski/Don 7712 var. Desvauxii, 10 – Aru/Don 7712 var. vulgaris, 11 – 7698/Aru var. erythroceros, 12 – 7698/Aru var. suberythroseros, 13 – Aru/Don 7712 var. erythroceros, 14 – Aru/Don 7712 var. Desvauxii.
Figure 4.
Electropherogram of PCR products of hybrids for the presence of Pi-33 and Pi-ta genes. (а): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Bakanasski/7668 var. subuzbekistanica, 6 – Bakanasski/04470 var. vulgaris, 7 – Bakanasski/7653 var. subuzbekistanica, 8 – Bakanasski/7653 var. subjanthoseros, 9 – Bakanasski/7684 var. subvulgaris, 10 – Bakanasski/7684 var. vulgaris, 11 – 7824/Aisaule var. italica, 12 – 7824/Aisaule var. subuzbekistanica, 13 – 7698/Bakanasski var. subvulgaris, 14 – 7698/Bakanasski var. italica, 15 – Aru/7701 var. vulgaris, 16 – Bakanasski/0327 var. vulgaris, 17 – Aisaule/7679 var. subvulgaris, 18 – 7698/Aisaule var. italica (в): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – 7698/Aisaule var. suberythroseros, 6 – Aru/7702 var. erythroceros, 7 – Aisaule/7689 var. zeravschanica, 8 – Aru/0327 var. vulgaris, 9 – Bakanasski/7667 var. vulgaris, 10 – Bakanasski/7701 var. vulgaris, 11 – Aisaule/7664 var. italica, 12 – Aisaule/7664 var. subvulgaris, 13 – Aisaule/04470 var. subvulgaris, 14 – Aisaule/04470 var. italica, 15 – Aisaule/Don 7712 var. sundensis, 16 – Aisaule/Don 7712 var. italica, 17 – Fatima/7695 var. breviaristata, 18 – Bakanasski/7683 var. subjanthoseros (с): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Aru/212-05 var. vulgaris, 6 – Aru/7668 var. vulgaris, 7 – Aisaule/04468 var. italica, 8 – Aru/04636 var. vulgaris, 9 – Bakanasski/7690 var. subjanthoseros, 10 – Bakanasski/7690 var. vulgaris, 11 – Bakanasski/04468 var. subvulgaris, 12 – Bakanasski/04468 var. vulgaris, 13 – Aru/7666 var. suberythroseros, 14 – Fatima/7653 var. persica, 15 – Aru/Don 7712 var. Desvauxii semi-awned, 16 – Aru/7683 var. janthoceros, 17 – Aru/7683 var. vulgaris, 18 – Aisaule/7663 var. italica, 19 – Aru/0327 var. subpyrocarpa (d): M – Molecular marker 100 bp; 1 - C101-Lac (control "+" for the Pi-33 gene at the Rm310 and RM72 loci), 2 – Flagman (control "-" for the Pi-33 gene at the Rm310 and RM72 loci), 3 – IR36 (control "+" for the Pi-ta gene at the Pita loci), 4 – Flagman (control "-" for the Pi-ta gene at the Pita loci), 5 – Bakanasski/Don 7712, 6 – Fatima/7689 var. italica, 7 – Aru/04468 var. subvulgaris, 8 – Bakanasski/7667 var. subjanthoseros, 9 – Bakanasski/Don 7712 var. Desvauxii, 10 – Aru/Don 7712 var. vulgaris, 11 – 7698/Aru var. erythroceros, 12 – 7698/Aru var. suberythroseros, 13 – Aru/Don 7712 var. erythroceros, 14 – Aru/Don 7712 var. Desvauxii.
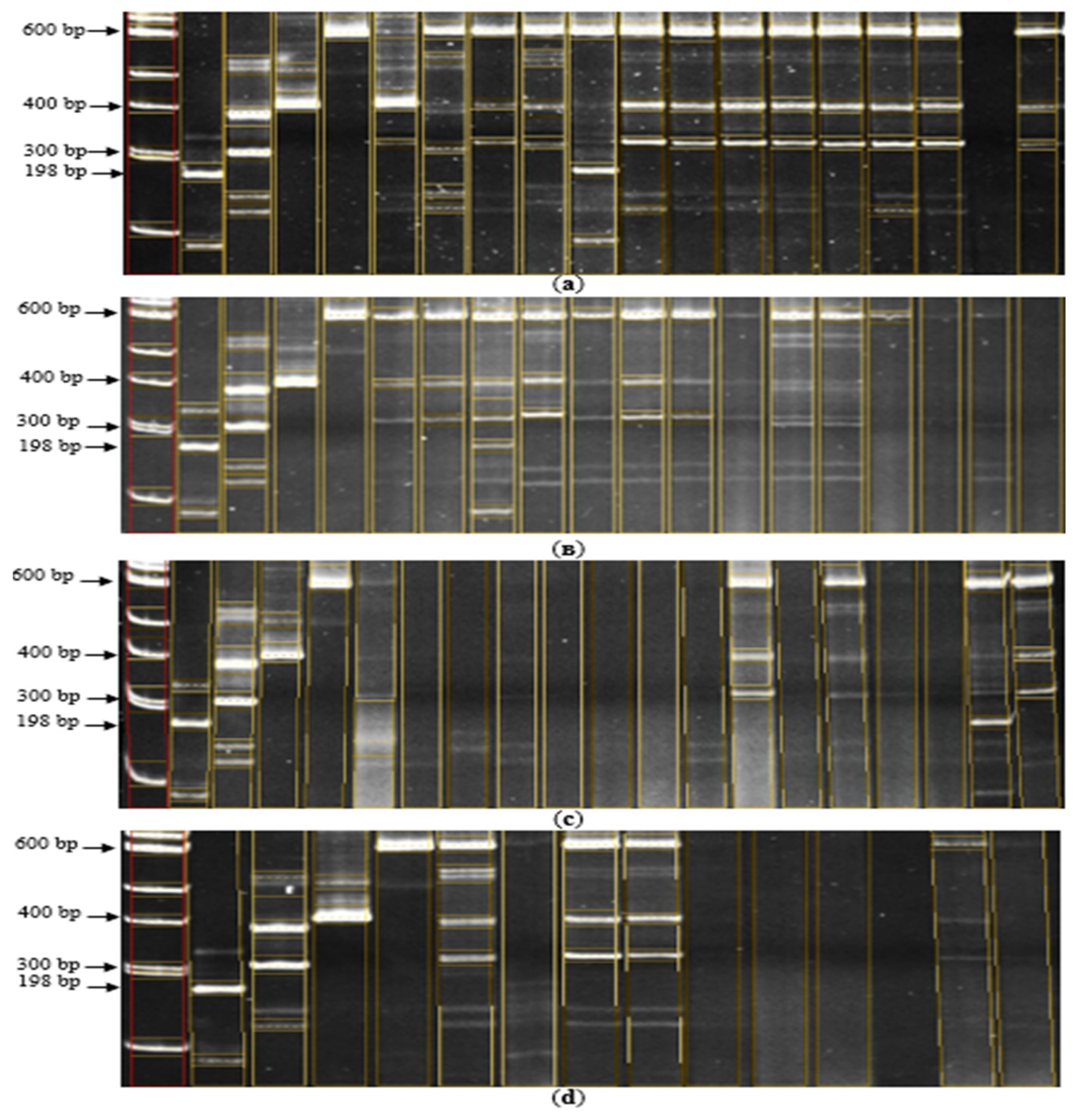
Figure 5.
Degree of leaf resistance to blast: (a) resistant, (b) susceptible, (c) moderately resistant.
Figure 5.
Degree of leaf resistance to blast: (a) resistant, (b) susceptible, (c) moderately resistant.


Table 1.
Phytopathological assessment of blast resistance of obtained pyramidal lines and local rice cultivars.
Table 1.
Phytopathological assessment of blast resistance of obtained pyramidal lines and local rice cultivars.
| Hybrid lines | Blast resistance | |||
|---|---|---|---|---|
| Intensity of disease development, % | Degree of resistance to disease | Resistance genes | Line’s reaction | |
| Bakanasski | 55.6 | susceptible | Pi-2 | S |
| Aisaule | 55.6 | susceptible | Pi-2 | S |
| Aru | 66.7 | susceptible | Pi-2 | S |
| Fatima | 44.4 | moderately resistant | - | MR |
| 7698/Aisaule var. suberythroseros |
- | - | - | This line didn't grow up |
| Aru/7702 var. erythroceros |
61.1 | susceptible | Pi-1, Pi-ta | S |
| Aisaule/7689 var. zeravschanica |
36.7 | moderately resistant | Pi-1, Pi-33, Pi-ta | MR |
| Bakanasski/7667 var. subjanthoseros |
61.1 | susceptible | Pi-1, Pi-ta | S |
| Bakanasski/7667 var. vulgaris |
38.9 | moderately resistant | Pi-1, Pi-ta | MR |
| Bakanasski/7701 var. vulgaris |
72.2 | susceptible | Pi-1, Pi-ta | S |
| Aisaule/7664 var. italica |
44.4 | moderately resistant | Pi-1, Pi-ta | MR |
| MR – moderately resistant, S – susceptible. | ||||
Table 2.
Analyses of the productivity of pyramided rice lines by One-way ANOVA.
| SS | df | MS | F | P-value | F crit | |
| Between | 367.12 | 6 | 61.19* | 27.07 | 0.00 | 3.25 |
| Within | 97.18 | 43 | 2.26 |
Table 3.
List of tested SSR primers for Pi genes of the blast resistance complex.
| Gene | Localization on chromosome | Name of a marker |
Forward primer (5′-3′) Reverse sequence (5′-3′) |
|---|---|---|---|
|
Pi-1 |
11 | Rm 224 | F - аtсgаtсgаtсttсасgаgg R - tgctataaaaggcattcggg |
| Rm144 | F - tgccctggcgcaaatttgatcc R - gctagaggagatcagatggtagtgcatg |
||
| Pi-2 | 6 | Rm 527 | F - ggctcgatctagaaaatccg R - ttgcacaggttgcgatagag |
| SSR 140 | F - aaggtgtgaaacaagctagcaa R - ttctaggggaggggtgtgaa |
||
| Pi-33 | 8 | Rm 310 | F - ссggсgаtаааасааtgаg R - gсаtсggtссtаасtааggg |
| Rm 72 | F - ссggсgаtаааасааtgаg R - gсаtсggtссtаасtааggg |
||
| Pi-ta | 12 | Pita | F1 - gссgtggсttсtаtсtttасatg R1 - аtссааgtgttаgggссаасаttс |
| F2 - ttgасасtсtсаааggасtgggаt R2 - tсааgtсаggttgааgаtgсаtсgа |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



