
Submitted:
17 March 2025
Posted:
18 March 2025
You are already at the latest version
Abstract
Foodborne illnesses caused by pathogenic Escherichia coli represent a significant challenge for the food industry. In this study, a cocktail bacteriophages with high lytic potential, broad host range, and thermal stability was designed and evaluated for the control of E. coli in lettuce stored at 4°C and 25°C. Phages were isolated from wastewater samples and selected based on their lytic activity against multiple strains. Application of the cocktail achieved significant reductions of E. coli ATCC 25922, reaching a maximum decrease of 2.64 log CFU/mL at 6 hours at 25°C and 2.47 log CFU/mL at 24 hours at 4°C. These results support the use of phage cocktails as a promising strategy for biocontrol of E. coli in fresh produce.
Keywords:
biocontrol
; bacteriophages
; Escherichia coli
1. Introduction
Escherichia coli (E. coli) is a ubiquitous bacterium commonly found in the gastrointestinal tract of humans and animals [1]. Although most E. coli strains are non-pathogenic commensals, some have virulence factors that enable them to cause disease in humans [1,2]. Among these, Shiga toxin-producing E. coli (STEC) is a major cause of foodborne illness, with the World Health Organization estimating that in 2010, it led to over 2.5 million infections, including 3,330 cases of hemolytic uremic syndrome and 269 deaths [3].
E. coli STEC serotype O157:H7 is one of the most important causes of foodborne illness worldwide. In the United States, it accounts for approximately 43% of STEC-related illnesses. However, other serotypes, such as O26, O103, O111, O121 and O145, are gaining more and more relevance [4]. This suggests the need to expand monitoring efforts and implement broader control strategies to mitigate the risks associated with diverse STEC serotypes.
E. coli contamination of food represents a significant public health problem, especially with fresh vegetables such as lettuce, which have been identified as vehicles for this bacterium [5,6]. The food industry has implemented various measures to control E. coli contamination in fresh produce, including good agricultural practices, such as using clean irrigation water and ensuring worker hygiene, as well as post-harvest decontamination strategies, such as washing and disinfection treatments [5,6,7]. However, these methods have limitations in terms of their effectiveness in completely eliminating E. coli. In addition, the use of chemical disinfectants raises concerns about antimicrobial resistance and potential negative effects on food quality [5,8,9,10].
Bacteriophages (phages), viruses that specifically infect and lyse bacteria, have emerged as a promising alternative to traditional methods of pathogen control in food safety [9,11,12]. Phages are ubiquitous in nature, existing wherever their bacterial hosts are found, including food and environmental reservoirs [5,9]. They offer several advantages as biocontrol agents, including their high specificity, which typically targets few strains within a species [8]. Unlike conventional chemical treatments, phages do not disrupt the microbiota present in food or in the human microbiome, making them a selective and environmentally friendly intervention. Additionally, phages have been extensively studied for their safety in food applications and are generally recognized as safe (GRAS) by regulatory agencies [10,13]. Several studies, including human clinical trials, have demonstrated their lack of adverse effects, further supporting their potential as food biopreservatives [14].
Phages have also shown remarkable efficacy in reducing bacterial contamination across various food matrices, significantly enhancing food safety [15,16,17,18,19]. For instance, a study reported that a cocktail of five lytic phages effectively decreased Shigella sonnei counts in lettuce and other foods [20]. Similarly, phage A511 was shown to completely eliminate Listeria monocytogenes in mozzarella cheese brine and chocolate milk, demonstrating its versatility in different food systems [21]. Despite these promising findings, the application of phages to control E. coli in fresh produce, remains underexplored. Addressing this gap is crucial for advancing phage-based biocontrol strategies in fresh food safety.
To enhance phage efficacy, the use of phage cocktails is increasingly being explored to overcome the narrow host range of individual phages and prevent bacterial resistance.
This study aims to isolate, and characterize lytic E. coli phages from environmental samples. Based on these findings, an optimized phage cocktail will be formulated and tested for its ability to control E. coli contamination in lettuce at different temperatures.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
A total of 12 E. coli strains were used including five strains isolated from wastewater samples (WWS) collected at a wastewater treatment plant. In addition, the strain ATCC 14028 of Salmonella enterica serovar Typhimurium was included. (Supplementary material Table S1). Bacterial cultures were grown in Luria-Bertani (LB) broth (5 g/L NaCl, 5 g/L yeast extract, 10 g/L tryptone) or in saline LB broth (5 g/L NaCl, 5 g/L yeast extract, 10 g/L tryptone, CaCl 1% (w/v), MgSO4 10% (w/v)) and incubated at 37°C with continuous shaking. LB agar plates were prepared with 1.5% (w/v) agar, while LB soft agar (used for double-layer plaque assays) contained 0.6% (w/v) agar. Plates were incubated at 37°C for 16 hours.
2.2. Isolation and Purifications of Phages
Bacteriophages were isolated from seven WWS collected in the Santiago Poniente treatment plant over 3-4 week intervals, following the method described by Clokie and Kropinski [22]. A pooled sample was also created by mixing equal parts of the seven individual samples, with all samples stored at 4°C until use. To remove bacterial cells, WWS were centrifuged and filtered through a 0.22 μm membrane filter before enrichment. Briefly, 25 mL of each WWS was mixed with 25 mL of 2X saline LB broth and inoculated with 0.1 mL of a bacterial pool (B1347, B1348, B2494, B2495, B2590, and B2591) or E. coli ATCC 25922 in separate assays. The mixture was incubated at 37°C with shaking for 24 hours. After incubation, the enriched samples were centrifuged and filtered through a 0.22 μm membrane filter, and phages were isolated by serial dilutions in SM buffer (Tris-HCl 50 mM, NaCl 100 mM, MgSO4·7H2O 8 mM pH 7,5 and 0.01% w/v gelatin) followed by double-layer agar plating [23]. Distinct lysis plaques were picked from countable dilution plates. Phage titers were determined as plaque-forming units per milliliter (PFU/mL) using serial dilutions.
2.3. Infection Curves of the Phages
The inhibition effect of the phages in LB broth was determined by monitoring bacterial growth inhibition through the measurement of optical density (OD) at 620 nm in an ELISA plate reader every 30 minutes for 12 hours, following a previously described method with modifications [22]. For each assay, 180 µL of E. coli at a concentration of 1 × 10⁵ CFU/mL was mixed with 20 µL of bacteriophages at concentrations of 1 × 10⁷ PFU/mL, 1 × 10⁶ PFU/mL, 1 × 10⁵ PFU/mL, and 1 × 10⁴ PFU/mL, achieving multiplicities of infection (MOI) of 10, 1, 0.1, and 0.01, respectively. Measurements were performed in triplicate, with shaking for 5 seconds before each measurement.
2.4. Phages Host Range Determination
The host range of the isolated phages was determined using a spot assay. E. coli strains in the exponential growth phase (Figure 1) were used as bacterial hosts. An aliquot of 5 µL of each phage suspension (1 × 10⁷ PFU/mL) was spotted onto a bacterial lawn grown for 16 hours at 37°C. Every test was performed in triplicate.
2.5. Temperature Stability of the Phages
The temperature stability of phages E.1, E.3.1, and E.9 (1 × 10⁷ PFU/mL) was evaluated after incubation in a water bath at 4°C, 25°C, 37°C, 50°C, and 70°C for 1 hour followed by titration using the double-layer agar plate assay. The assay was performed in triplicate.
2.6. One Step Growth Curve
The one-step growth curve of phages E.1, E.3.1, and E.9 was determined following a previously described method with minor modifications [22]. Briefly, E. coli cultures were mixed with phages at an MOI of 0.1 and incubated at 37°C for 5 minutes to allow adsorption. After adsorption, the mixture was diluted 1:100 in LB broth, followed by a 1:10 dilution after 5 minutes. Samples were collected every 5 or 10 minutes for 60 minutes and titered by the double-layer agar plate method. The concentration of phages (PFU/mL), latency period and burst size were calculated as described in a previous method [24]. Every experiment was realized in two different assays, and measurement each was performed in triplicate.
2.7. Cocktail Design
The phage cocktail was prepared by combining equal volumes of phages E.1, E.3.1, and E.9 at the same concentration until the desired final concentration was reached.
2.8. Infection Curves of the Cocktail
The infection curves of the phage cocktail were performed following the methodology described in Section 2.3. Specifically, 20 µL of the phage cocktail at concentrations of 1 × 10⁶ PFU/mL and 1 × 10⁵ PFU/mL was added to 180 µL of E. coli at 1 × 10⁵ CFU/mL, with triplicate measurements taken every 30 minutes.
2.9. Inhibition Effect of Phages Against E. coli in Saline LB Broth (In Vitro)
The inhibitory effect of the phage cocktail against E. coli strains in LB broth was assessed at 4°C and 25°C over 24 hours. A total of 9 mL of E. coli (1 × 10⁴ CFU/mL) was mixed with 1 mL of the phage cocktail (3 × 10⁵ PFU/mL). The bacterial count was determined by serial dilution and growth on LB agar plates incubated for 16 hours at 37°C.
2.10. Inhibition Effect of Phages Against E. coli in Lettuce (Challenge)
The inhibitory effect of the phage cocktail on E. coli in lettuce was assessed using a modified protocol [25]. Approximately 600 g of lettuce was disinfected by immersion in 70% (v/v) ethanol for 5 minutes, followed by centrifugation for 5 minutes and exposure to UV light for 10 minutes on each side. Contamination was performed by immersing lettuce in 100 mL of a 1 × 10⁵ CFU/mL E. coli solution for 10 seconds, followed by drying for 30 minutes in a biosecurity cabinet. Phage treatment was applied by immersing the contaminated lettuce in 100 mL of a 1 × 10⁸ PFU/mL phage solution (MOI 1000). Samples were stored at 4°C for 24 and 48 hours, and at 25°C for 6 and 24 hours. Viable bacteria were enumerated by serial dilution plate counting.
2.11. Statistical Analysis
All data were analyzed using GraphPad Prism 8.0.1. Results are expressed as mean ± standard deviation (SD), and statistical significance was determined by two-way ANOVA, followed by Tukey’s test, with p < 0.05 considered significant.
3. Results
3.1. Isolation and Morphology Plates of the Phages
A total of ten phages (E.1, E.2, E.3, E.3.1, E.4, E.5.1, E.5.2, E.6, E.7.1, and E.9) were isolated after an enrichment process (Table 1) from eight wastewater filtrates collected from the influent of the Santiago Poniente treatment plant in 2023.
The analysis of phage plaque morphology revealed considerable diversity in both shape and size. Phages E.3.1 and E.9 formed clear plaques, characteristic of lytic phages [30]. In contrast, phage E.1 exhibited a “bull’s-eye” morphology, with a clear center and turbid edges. Phage E.6 produced turbid plaques, suggesting it may be temperate. The remaining phages formed plaques with halos, indicating the diffusion of lytic enzymes.
Regarding plaque size, the smallest was E.9 (0.155 cm ± 0.02), while the largest belonged to E.5.2 (0.574 cm ± 0.02), which also exhibited a halo of 1.402 cm ± 0.09.
Individual characterization of each phage was performed, leading to the selection of three candidates for further characterization and cocktail formulation.
- WWS: wastewater sample
- NO: No observed
3.2. Phage Characterization
3.2.1. Host Range
The host range analysis was conducted using 13 bacterial strains (Figure 1). Bacterial sensitivity to each phage was classified into three categories based on plaque clarity: sensitive strains, low-sensitive strains, and non-sensitive strains, as described previously [31].
Results showed that 9 out of 10 phages lysed E. coli ATCC 25922, likely due to the fact that this strain was used for enrichment, making it the most susceptible. On the other hand, strains B1347 was the most resistant, as none of the phages exhibited lytic activity against them. Similarly, the five wild type E. coli isolates obtained in this study from WWS remained unaffected.
Phages E.2, E.3, E.4, E.5.1, E.6, and E.7 had the narrowest host range, lysing only E. coli ATCC 25922. In contrast, phage E.9 displayed the broadest host range, lysing five out of the 13 tested strains. Notably, E.9 was isolated through an enrichment process that did not include E. coli ATCC 25922.
An intriguing finding was that phage E.3.1 exhibited lytic activity against Salmonella enterica serovar Typhimurium (ATCC 14028), suggesting a polyvalent nature, which is uncommon and typically associated with broad-host-range phages.
3.2.2. Infection Curves
Figure 2 presents the infection curves of selected phages at an MOI of 0.1. Phages E.1 and E.3.1 completely inhibited bacterial growth for up to 48 hours, whereas E.6 suppressed growth for only 6 hours before bacterial regrowth occurred. The remaining phages did not significantly affect bacterial growth, displaying similar behavior to the control curve (Supplementary material Figure S1).
3.3. Characterization of Cocktail Phages
Based on the previous analyses, three phages were selected for the cocktail design. Phage E.1 and E.3.1 were chosen due to their clear lysis plaques (Table 1) and their efficient bacterial inhibition capacity under optimal E. coli growth conditions (Figure 2), suggesting their ability to evade bacterial resistance, a key feature in phage biocontrol applications. Phage E.9 was selected for its broad host range (Figure 1), which enhances the control of various E. coli strains when included in the cocktail.
3.3.1. Phage Temperature Stability
The selected phages exhibited high thermal tolerance, remaining viable after exposure to 50°C for one hour. Phages E.3.1 and E.9 demonstrated greater heat resistance, as viable particles were detected even after exposure to 70°C for one hour, although a reduction in infectivity was observed (Figure 3). At 80°C, no viable phages were detected.
3.3.2. Phage Adsorption Rate and One Step Growth Curve
The one-step growth curve experiments revealed distinct kinetic parameters among the three phages. Phage E.1 exhibited a latency period of approximately 15 minutes, followed by a burst size of 5 PFU per infected bacterial cell. In contrast, phages E.3.1 and E.9 had shorter latency periods (10 minutes) and significantly higher burst sizes, estimated at 27 and 43 PFU per infected bacterial cell, respectively (Figure 4).
3.4. Application
3.4.1. Inhibition Effect of the Phages Cocktail Against E. coli in Culture Medium
The infection curves of the phage cocktail showed that bacterial growth was completely inhibited for at least 48 hours in all tested E. coli strains (Supplementary Material Figure S2). Additionally, in vitro assays conducted at low temperatures revealed significant bacterial reductions in most cases. The greatest reduction was observed at 25°C for both E. coli strains, with a reduction of 1.148 Log10 CFU/mL in strain B1348 and 0.723 Log10 CFU/mL in strain ATCC 25922. On the other hand, at 4°C, the cocktail achieved a reduction of 0.466 Log10 CFU/mL in strain ATCC 25922 (Figure 5).
3.4.2. Inhibition Effect of Phages Against E. coli in Lettuce (Challenge)
A cocktail containing 1 × 108 PFU/mL of each selected phage was tested on lettuce under two different temperature conditions to evaluate its effectiveness in reducing E. coli contamination. Each assay was conducted separately using two E. coli strains.
As shown in Figure 6, the phage cocktail significantly reduced E. coli ATCC 25922 counts under all tested conditions. The greatest reduction was 2.64 log CFU/mL at 25°C after 6 hours. A similar reduction (2.47 log CFU/mL) was observed in lettuce stored at 4°C for 24 hours.
For E. coli B1348, reductions were less pronounced but still detectable. At 4°C, bacterial counts decreased by 0.728 log CFU/mL after 24 hours and 0.551 log CFU/mL after 48 hours. At 25°C, a 1.200 log CFU/mL reduction was observed after 6 hours, decreasing to 0.704 log CFU/mL after 24 hours with respect to the control.
4. Discussion
Foodborne illnesses caused by pathogenic E. coli pose a significant challenge to the food industry [5]. In recent years, bacteriophages have emerged as a promising alternative for controlling these pathogens due to their specificity and ability to reduce microbial loads in various environments [5]. In this study, a cocktail of three phages with high lytic potential, broad host range, and thermal stability was designed and evaluated, achieving a significant reduction of E. coli in lettuce stored at 4°C and 25°C. At 4°C, the maximum reduction reached 2.47 log10 CFU/mL after 24 h, while at 25°C, the reduction was 2.64 log10 CFU/mL at 6 h. These results suggest that the phage cocktail maintains its effectiveness across a range of temperatures relevant to fresh food preservation.
For bacteriophage selection and cocktail design, 10 phages against E. coli were isolated from wastewater samples, a source recognized for its high phage diversity [9,26,27]. Previous studies have shown that wastewater treatment systems harbor numerous bacteriophages with biocontrol potential. For instance, Göller et al. (2021) reported the isolation of 155 phages from 46 staphylococcal hosts spanning 17 different species [28]. Following a similar approach, this study focused on selecting phages active against E. coli.
Host range is one of the most relevant characteristics for the selection of phages in biocontrol applications. Our results show that six of the ten isolated phages were only able to lyse the strain in which they were enriched (E. coli ATCC 25922), while the remaining four phages lysed at least two different strains. Among them, phage E.9 exhibited the broadest host range, successfully lysing six of the thirteen strains evaluated. These results suggest that most of the phages obtained have a narrow host range.
Phage E.9 was selected to expand the host range of the cocktail. To achieve this, a pool of 7 wastewater samples was enriched with six E. coli strains. However, it did not exhibit lytic activity against strain ATCC 25922, as this strain had been excluded from the enrichment pool. This finding highlights the influence of the enrichment method on the diversity of phages obtained and their specificity towards certain strains. Given its broad host range, phage E.9 was incorporated into the final cocktail design.
The high specificity and variability in the host ranges of phage isolates have been reported previously by Topka et al. (2019), who demonstrated that bacteriophage vB_EcoS-95, obtained from urban sewage, was capable of lysing strains such as E. coli MG1655 and two clinical isolates but was unable to infect Shiga toxin-producing serotypes, such as O157:H7, or other related bacterial species, such as Shigella flexneri and Salmonella enterica [29]. In contrast, a broader analysis of a collection of bacteriophages showed that most had restricted host ranges, limited to specific strains of E. coli [30]. However, some phages were able to infect both laboratory strains and clinical isolates, highlighting their potential for more versatile cocktail applications [30].
The use of phages with different host ranges in the same cocktail not only broadens their spectrum of action against E. coli, but also helps mitigate the emergence of resistant strains, enhancing the sustainability of biocontrol in food matrices [31].
To form the cocktail, the lytic activity of the selected phages was evaluated at different MOI levels by means of infection curves. Among the characterized phages, E.1 and E.3.1 exhibited complete bacterial lysis across all MOI tested, indicating highly efficient phage-bacteria interactions. The ability to induce complete and sustained lysis is fundamental for applications in food matrices, as it prevents bacterial regrowth and enhances biocontrol efficiency [4,32]. Due to their strong lytic activity, both phages were selected for the formulation of the designed cocktail.
Then, infection curve assays performed using the cocktail designed in this study showed complete lysis of the cultures, demonstrating its effectiveness. This test was performed on three different strains. A noteworthy observation is that strain B1348 was lysed only by phage E9; however, its lysis was not complete (Supplementary material Figure S1). Yet, when the combination of all three phages was used, complete lysis of this strain was achieved, indicating an interaction between the phages, which translates into synergistic activity (Supplementary material Figure S2). This phenomenon has been reported in other studies, such as in the treatment of milk contaminated with E. coli, where a cocktail of three phages reduced the bacterial load to undetectable levels in both raw and UHT-treated milk, while a cocktail of only two phages allowed for bacterial regrowth [33].
The described results align with those reported in the literature [6,34], where the use of phage cocktails with different host ranges has proven to be an effective strategy for the biocontrol of E. coli in diverse environments. The combination of phages with complementary specificities not only expands the range of action against multiple strains, but also helps mitigate the emergence of bacterial resistance by targeting different host cell receptors [35]. In addition, previous studies have shown that these cocktails can maintain sustained lysis in food and environmental matrices, even under different temperature and storage conditions [36]. One example is the use of a three-phage cocktail that significantly reduced E. coli O157:H7 populations in ground beef and lettuce, maintaining this reduction for one week at 4°C [37]. These observations reinforce the potential of phage cocktails as versatile and sustainable tools in biocontrol strategies, contributing to microbiological safety in the food industry and wastewater treatment.
In order to facilitate the application of a phage cocktail at an industrial level, it is essential to consider that the formulation must be both stored and transported. Additionally, to enhance its versatility, it could be applied during industrial processes that involve high temperatures. Therefore, understanding the thermal stability of the cocktail is crucial. The greater tolerance of phages compared to bacteria provides them with a significant advantage in this context, allowing them to withstand elevated temperatures while remaining effective in pathogen control [38]. In this study, the selected phages demonstrated resistance to heat treatments for 1 hour at temperatures between 50°C and 70°C while maintaining their lytic activity. This tolerance aligns with other studies showing that seven E. coli lytic phages remain stable between 25°C and 70°C for up to 6 hours, with temperatures above 70°C reducing phage titers due to thermal effects on phage proteins [35].
In this context, the application of phage cocktails as biocontrol agents in food must consider the distribution and storage conditions they will undergo. In this study, the cocktail was designed for use in vegetables, making it essential that it retain its lytic capacity at typical refrigeration (4°C) and ambient (25°C) temperatures. Phage efficacy is influenced by multiple factors, particularly the interaction between temperature and the metabolic activity of the host bacteria. At low temperatures, such as those found in refrigeration, bacterial replication decreases significantly, which may limit phage infection since phages rely on the bacterial metabolic machinery to replicate and produce new viral particles [9,39]. However, some studies suggest that mechanisms such as lysis from without could explain bacterial reduction even under low-temperature conditions. This mechanism occurs when high phage concentrations induce bacterial cell rupture without requiring intracellular replication, which can be particularly relevant in refrigerated food matrices where bacterial metabolism is minimal [8,35,40]. Nevertheless, this lysis mechanism does not generate new viral particles, meaning that the initial phage concentration must be sufficiently high to effectively eliminate or significantly reduce bacterial populations in the food matrix.
Prior to evaluating the cocktail in the food matrix, we assessed its lytic capacity at 4°C and 25°C in a culture medium. These in vitro results demonstrated that the phage cocktail retained its lytic activity at 4°C, achieving a reduction of 0.466 log10 CFU/mL at 24 hours. At 25°C, the reduction was even greater, reaching 1.148 log10 CFU/mL. This behavior could be explained by the fact that, although metabolic activity of E. coli is reduced at 4°C, the high phage concentration can induce bacterial lysis through this alternative mechanism. While active phage replication does not occur at these temperatures, cell disruption can still proceed, indicating that even with decreased bacterial activity, the cocktail remains effective in eliminating pathogens. Similar studies have reported that phages applied at refrigeration temperatures also exhibit effectiveness. For instance, Kudva et al. (1999) observed reductions of up to 3.4 log10 CFU/mL at 4°C, suggesting that lysis from without may be a key mechanism for bacterial elimination under these conditions [41]. This finding supports our hypothesis that lysis from without plays a crucial role in the ability of the cocktail to reduce bacterial populations during refrigerated storage, even when viral replication is minimal [41].
Our results from application to lettuce show that the cocktail achieved significant reductions under all evaluated conditions for E. coli ATCC 25922, reaching a maximum reduction of 2.64 log10 CFU/mL at 6 hours at 25°C and 2.47 log10 CFU/mL at 24 hours at 4°C. These findings not only confirm the ability of phages to reduce E. coli in food matrices but also highlight their effectiveness over a range of temperatures relevant to fresh produce storage. Similar results have been previously reported in the literature. Zhou et al. (2022) observed reductions of 2.42 log10 CFU/piece at 4°C and 3.83 log10 CFU/piece at 25°C in E. coli contaminated lettuce, while phage VR26 achieved reductions of approximately 2 log10 CFU/piece in lettuce after 2 hours of treatment at 4°C, 8°C, and 22°C, with no survivors detected after 6 hours at 4°C [8]. However, the diversity in infection methods, bacterial load used, and the type of lettuce tested in these studies precludes a direct comparison of the results obtained. Lettuce, being a particularly complex food matrix due to its high roughness, may reduce the possibility of interaction between bacteria and bacteriophages, which could influence the effectiveness of biocontrol. This effectiveness could potentially be improved by using other food matrices, such as cucumber or melon.
The results obtained in the analysis of Safarirad et al. (2021) indicate that factors such as temperature, exposure time, phage application method, phage origin, and vegetable type are determinants in the reduction of E. coli. However, they noted that neither the bacterial strain nor the phage dose significantly influenced the observed reduction, suggesting that, rather than the E. coli strain, environmental factors play a more relevant role in the effectiveness of the treatment [39]. This information is relevant in the context of our study, because although we did not evaluate enterohemorrhagic E. coli, the results imply that factors such as temperature and application method are more influential than the bacterial strain used, which is consistent with our observations with other laboratory strains. Another option is the possibility of incorporating a phage that is capable of lysing this type of strain, either by evaluating phages isolated in these pathogenic variants or by adding a specific phage against enterohemorrhagic E. coli in future formulations. This would broaden the spectrum of action of the cocktail and improve its applicability in the food industry.
However, the marketing of precut vegetables must comply with regulations that require low levels of E. coli in these products, making the use of such phage cocktails a potentially effective strategy to meet these requirements (https://eur-lex.europa.eu/eli/reg/2005/2073/2014-06-01).
In general, trials show that the use of phages in food matrices significantly reduces the amount of E. coli [42,43]. However, results may vary between studies due to methodological differences, such as the initial bacterial load, the temperature range evaluated, and the strain of E. coli used [16]. In addition, discrepancies are observed between in vitro and in vivo assays, as controlled laboratory conditions do not always reflect the actual behavior of phages in food matrices [39]. These differences in experimental approaches, such as the MOI employed, the time of evaluation, the methodology of bacterial quantification, and the manner of application of the phage cocktail, are determinants in the effectiveness of the treatment. Methodological variations make direct comparison between studies difficult and emphasize the need to standardize experimental protocols to more accurately assess the impact of phages on pathogen biocontrol. Despite these variations, the general trend shows that phages have a high potential to reduce pathogens in vegetable matrices and other fresh produce.
On the other hand, the use of polyvalent phages, capable of infecting multiple bacterial species, represents an interesting strategy to broaden the spectrum of action of phage cocktails [6] In our study, one of the phages demonstrated activity against both E. coli and Salmonella, suggesting that their incorporation into biocontrol formulations could be advantageous in food matrices where different pathogens coexist. The ability of these phages to infect multiple species could reduce the need for combining several phages in a single cocktail, simplifying their production and application. However, it is crucial to evaluate whether their efficacy is maintained under different environmental conditions and whether there are risks of horizontal gene transfer between pathogens, an aspect that requires further investigation [4].
In this study, E. coli was observed to grow in the food matrix at 25°C, with an increase of 1.5 log10 CFU/g at 6 hours and 2.4 log10 CFU/g at 24 hours for the E. coli ATCC 25922 strain. This bacterial growth, observed in the absence of phage, suggests that the food matrix provides favorable conditions for E. coli proliferation. It highlights the importance of implementing biocontrol strategies before bacterial populations reach critical levels.
On the other hand, growth of E. coli was also observed at 4°C, indicating that the bacteria remain metabolically active, which could favor the lytic action of the phages. However, if the reduction observed at low temperatures is primarily due to lysis from without, the lack of efficient viral replication may allow the bacterial population to increase over time. This suggests that the cocktail formulation should be optimized to enhance both productive infection and lysis from without, thereby maximizing its effectiveness across a broad temperature range.
An alternative strategy to enhance the efficacy of the phage cocktail would be to combine phages with different technologies or incorporate additives, such as natural antimicrobials or packaging technologies, to improve their stability within the food matrix. These findings highlight the potential of combined strategies to optimize biocontrol effectiveness and prevent bacterial regrowth. Future research should focus on evaluating their impact on sensory quality and investigating ways to improve phage stability, thereby contributing to the development of innovative solutions for food safety and shelf-life extension of fresh produce.
Supplementary Materials
Table S1: Bacterial strains used in this study; Figure S1: Infection curves of phages E.2, E.3, E.4, E.5.1, E.5.2 and E.7.1 at MOI 0,1 against E. coli ATCC 25922 (A), and phage E.9 against E. coli B1348 (B); Figure S2: Infection curves of the cocktail at MOI 1 and 10 against E. coli ATCC 25922 (A), E. coli B1348 (B).
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, V.G. and A.A.; methodology, V.G. and A.A..; validation, V.G. and A.A; formal analysis, V.G..; investigation, V.G; resources, V.G.; data curation, V.G.; writing—original draft preparation, A.A.; writing—review and editing, V.G; visualization, V.G and A.A.; supervision, V.G.; project administration, V.G.; funding acquisition, V.G. All authors have read and agreed to the published version of the manuscripte..
Funding
This research was supported by a research grant 2024 of the Technological Faculty, USACH. and funded by the Laboratorio de Inocuidad Microbiológica de la Universidad de Santiago de Chile, and by Dicyt regular grant number 082571GM_REG.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors wish to thank the University of Santiago de Chile for the scholarship awarded to Alba-Rayen Araya-Acosta.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vila, J.; Sáez-López, E.; Johnson, J.R.; Römling, U.; Dobrindt, U.; Cantón, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martínez-Medina, M.; et al. Escherichia Coli: An Old Friend with New Tidings. FEMS Microbiol Rev 2016, 40, 437–463. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia Coli. Nat Rev Microbiol 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Döpfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis. PLoS Med 2015, 12, e1001921. [Google Scholar]
- Mangieri, N.; Picozzi, C.; Cocuzzi, R.; Foschino, R. Evaluation of a Potential Bacteriophage Cocktail for the Control of Shiga-Toxin Producing Escherichia Coli in Food. Front Microbiol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Tokman, J.I.; Woolston, J.; Sulakvelidze, A. Phage Biocontrol Improves Food Safety by Significantly Reducing the Level and Prevalence of Escherichia Coli O157:H7 in Various Foods. J Food Prot 2020, 83, 668–676. [Google Scholar] [CrossRef]
- Puligundla, P.; Lim, S. Biocontrol Approaches against Escherichia Coli O157:H7 in Foods. Foods 2022, 11. [Google Scholar] [CrossRef]
- Emencheta, S.; Attama, A.; Ezeibe, E.; Onuigbo, E. Isolation of Escherichia Coli Phages from Waste Waters. Hosts and Viruses 2022, 9. [Google Scholar] [CrossRef]
- Zajančkauskaitė, A.; Noreika, A.; Rutkienė, R.; Meškys, R.; Kaliniene, L. Low-Temperature Virus Vb_ecom_vr26 Shows Potential in Biocontrol of Stec O26:H11. Foods 2021, 10. [Google Scholar] [CrossRef]
- Snyder, A.B.; Perry, J.J.; Yousef, A.E. Developing and Optimizing Bacteriophage Treatment to Control Enterohemorrhagic Escherichia Coli on Fresh Produce. Int J Food Microbiol 2016, 236, 90–97. [Google Scholar] [CrossRef]
- Oluwarinde, B.O.; Ajose, D.J.; Abolarinwa, T.O.; Montso, P.K.; Du Preez, I.; Njom, H.A.; Ateba, C.N. Safety Properties of Escherichia Coli O157:H7 Specific Bacteriophages: Recent Advances for Food Safety. Foods 2023, 12. [Google Scholar] [CrossRef]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J. Bacteriophage Therapy. Antimicrob Agents Chemother 2001, 45, 649–659. [Google Scholar] [CrossRef]
- Campbell, A. The Future of Bacteriophage Biology. Nat Rev Genet 2003, 4, 471–477. [Google Scholar] [CrossRef]
- Sada, T.S.; Tessema, T.S. Isolation and Characterization of Lytic Bacteriophages from Various Sources in Addis Ababa against Antimicrobial-Resistant Diarrheagenic Escherichia Coli Strains and Evaluation of Their Therapeutic Potential. BMC Infect Dis 2024, 24. [Google Scholar] [CrossRef] [PubMed]
- Bruttin, A.; Brüssow, H. Human Volunteers Receiving Escherichia Coli Phage T4 Orally: A Safety Test of Phage Therapy. Antimicrob Agents Chemother 2005, 49, 2874–2878. [Google Scholar] [CrossRef]
- Duc, H.M.; Son, H.M.; Yi, H.P.S.; Sato, J.; Ngan, P.H.; Masuda, Y.; Honjoh, K. ichi; Miyamoto, T. Isolation, Characterization and Application of a Polyvalent Phage Capable of Controlling Salmonella and Escherichia Coli O157:H7 in Different Food Matrices. Food Research International 2020, 131. [Google Scholar] [CrossRef] [PubMed]
- Shebs-Maurine, E.L.; Giotto, F.M.; Laidler, S.T.; de Mello, A.S. Effects of Bacteriophages and Peroxyacetic Acid Applications on Beef Contaminated with Salmonella during Different Grinding Stages. Meat Sci 2021, 173. [Google Scholar] [CrossRef] [PubMed]
- Orquera, S.; Gölz, G.; Hertwig, S.; Hammerl, J.; Sparborth, D.; Joldic, A.; Alter, T. Control of Campylobacter Spp. and Yersinia Enterocolitica by Virulent Bacteriophages; 2012; Vol. 6;
- Lu, Y.T.; Ma, Y.; Wong, C.W.Y.; Wang, S. Characterization and Application of Bacteriophages for the Biocontrol of Shiga-Toxin Producing Escherichia Coli in Romaine Lettuce. Food Control 2022, 140, 109109. [Google Scholar] [CrossRef]
- Sharma, M.; Patel, J.R.; Conway, W.S.; Ferguson, S.; Sulakvelidze, A. Effectiveness of Bacteriophages in Reducing Escherichia Coli O157:H7 on Fresh-Cut Cantaloupes and Lettuce †; 2009; Vol. 72;
- Soffer, N.; Woolston, J.; Li, M.; Das, C.; Sulakvelidze, A. Bacteriophage Preparation Lytic for Shigella Significantly Reduces Shigella Sonnei Contamination in Various Foods. PLoS One 2017, 12. [Google Scholar] [CrossRef]
- Guenther, S.; Huwyler, D.; Richard, S.; Loessner, M.J. Virulent Bacteriophage for Efficient Biocontrol of Listeria Monocytogenes in Ready-to-Eat Foods. Appl Environ Microbiol 2009, 75, 93–100. [Google Scholar] [CrossRef]
- Clokie, M.; Kropinski, A. Bacteriophages; Clokie, M.R.J. , Kropinski, A.M., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2009; ISBN 978-1-58829-682-5. [Google Scholar]
- Guo, Y.; Li, J.; Islam, M.S.; Yan, T.; Zhou, Y.; Liang, L.; Connerton, I.F.; Deng, K.; Li, J. Application of a Novel Phage VB_SalS-LPSTLL for the Biological Control of Salmonella in Foods. Food Research International 2021, 147. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Z.; Tian, C.; Chen, X.; Hu, L.; Wei, X.; Li, H.; Lin, W.; Jiang, A.; Feng, R.; et al. Characterizing the Biology of Lytic Bacteriophage VB_EaeM_φEap-3 Infecting Multidrug-Resistant Enterobacter Aerogenes. Front Microbiol 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Tao, Z.; Li, T.; Chen, H.; Zhao, Y.; Sun, X. Isolation and Characterization of Novel Bacteriophage VB_KpP_HS106 for Klebsiella Pneumonia K2 and Applications in Foods. Front Microbiol 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Viazis, S.; Akhtar, M.; Feirtag, J.; Diez-Gonzalez, F. Reduction of Escherichia Coli O157:H7 Viability on Leafy Green Vegetables by Treatment with a Bacteriophage Mixture and Trans-Cinnamaldehyde. Food Microbiol 2011, 28, 149–157. [Google Scholar] [CrossRef]
- Beheshti Maal, K.; Delfan, A.S.; Salmanizadeh, S. Isolation and Identification of Two Novel Escherichia Coli Bacteriophages and Their Application in Wastewater Treatment and Coliform’s Phage Therapy. Jundishapur J Microbiol 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Göller, P.C.; Elsener, T.; Lorgé, D.; Radulovic, N.; Bernardi, V.; Naumann, A.; Amri, N.; Khatchatourova, E.; Coutinho, F.H.; Loessner, M.J.; et al. Multi-Species Host Range of Staphylococcal Phages Isolated from Wastewater. Nat Commun 2021, 12. [Google Scholar] [CrossRef]
- Topka, G.; Bloch, S.; Nejman-Falenczyk, B.; Gasior, T.; Jurczak-Kurek, A.; Necel, A.; Dydecka, A.; Richert, M.; Wegrzyn, G.; Wegrzyn, A. Characterization of Bacteriophage VB-EcoS-95, Isolated from Urban Sewage and Revealing Extremely Rapid Lytic Development. Front Microbiol 2019, 10. [Google Scholar] [CrossRef]
- Jurczak-Kurek, A.; Gasior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of Bacteriophages: Morphological and Biological Properties of a Large Group of Phages Isolated from Urban Sewage. Sci Rep 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Tomat, D.; Casabonne, C.; Aquili, V.; Balagué, C.; Quiberoni, A. Evaluation of a Novel Cocktail of Six Lytic Bacteriophages against Shiga Toxin-Producing Escherichia Coli in Broth, Milk and Meat. Food Microbiol 2018, 76, 434–442. [Google Scholar] [CrossRef]
- Shahdadi, M.; Safarirad, M.; Berizi, E.; Mazloomi, S.M.; Hosseinzadeh, S.; Zare, M.; Derakhshan, Z.; Rajabi, S. A Systematic Review and Modeling of the Effect of Bacteriophages on Salmonella Spp. Reduction in Chicken Meat. Reduction in Chicken Meat. Heliyon 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- McLean, S.; Dunn, L.; Palombo, E. Phage Inhibition of Escherichia Coli in Ultrahigh-Temperature-Treated and Raw Milk. Foodborne Pathog Dis 2014, 10, 956–962. [Google Scholar] [CrossRef]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Tomat, D.; Casabonne, C.; Aquili, V.; Balagué, C.; Quiberoni, A. Evaluation of a Novel Cocktail of Six Lytic Bacteriophages against Shiga Toxin-Producing Escherichia Coli in Broth, Milk and Meat. Food Microbiol 2018, 76, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Ameh, E.M.; Tyrrel, S.; Harris, J.A.; Pawlett, M.; Orlova, E. V.; Ignatiou, A.; Nocker, A. Lysis Performance of Bacteriophages with Different Plaque Sizes and Comparison of Lysis Kinetics After Simultaneous and Sequential Phage Addition. PHAGE: Therapy, Applications, and Research 2020, 1, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.D.; Parks, A.; Abuladze, T.; Li, M.; Woolston, J.; Magnone, J.; Senecal, A.; Kropinski, A.M.; Sulakvelidze, A. Bacteriophage Cocktail Significantly Reduces Escherichia Coli O157. Bacteriophage 2012, 2, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Nime, I.; Pan, F.; Wang, X. Isolation and Characterization of Phage ISTP3 for Bio-Control Application against Drug-Resistant Salmonella. Front Microbiol 2023, 14. [Google Scholar] [CrossRef]
- Safarirad, M.; Shahdadi, M.; Berizi, E.; Mazloomi, S.M.; Hosseinzadeh, S.; Montaseri, M.; Derakhshan, Z. A Systematic Review and Modeling of the Effect of Bacteriophages on E. Coli O157:H7 Reduction in Vegetables. Heliyon 2023, 9. [Google Scholar] [CrossRef]
- Snyder, A.B.; Perry, J.J.; Yousef, A.E. Developing and Optimizing Bacteriophage Treatment to Control Enterohemorrhagic Escherichia Coli on Fresh Produce. Int J Food Microbiol 2016, 236, 90–97. [Google Scholar] [CrossRef]
- Kudva, I.T.; Jelacic, S.; Tarr, P.I.; Youderian, P.; Hovde, C.J. Biocontrol of Escherichia Coli O157 with O157-Specific Bacteriophages; 1999; Vol. 65;
- Abuladze, T.; Li, M.; Menetrez, M.Y.; Dean, T.; Senecal, A.; Sulakvelidze, A. Bacteriophages Reduce Experimental Contamination of Hard Surfaces, Tomato, Spinach, Broccoli, and Ground Beef by Escherichia Coli O157:H7. Appl Environ Microbiol 2008, 74, 6230–6238. [Google Scholar] [CrossRef]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10. [Google Scholar] [CrossRef]
Figure 1.
Heat map showing the host range of the 10 isolated phages. Dark color shows the sensitive strains, medium color the low sensitive strains and light color the no sensitive strains.
Figure 1.
Heat map showing the host range of the 10 isolated phages. Dark color shows the sensitive strains, medium color the low sensitive strains and light color the no sensitive strains.
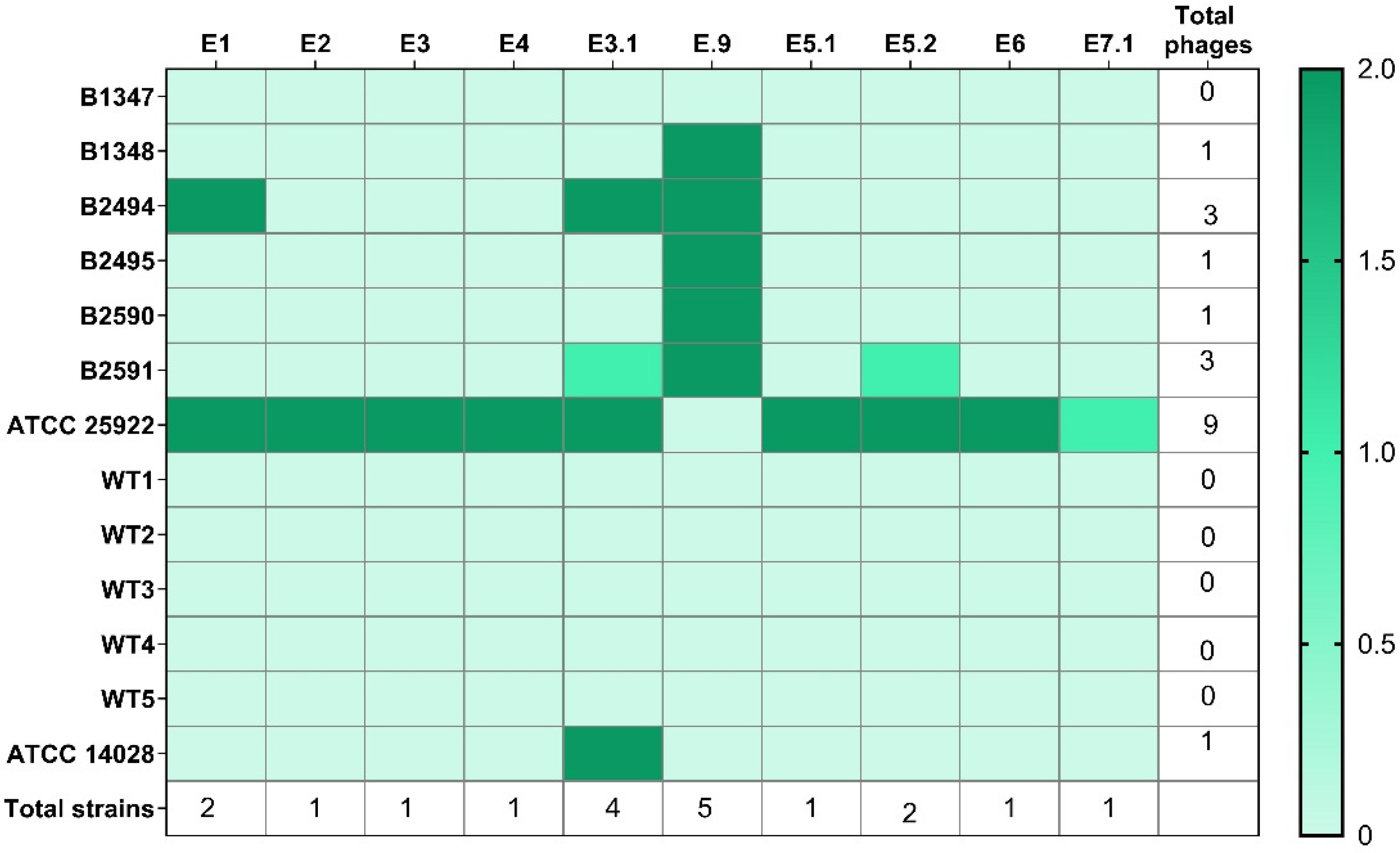
Figure 2.
Infection curves of phages E.1, E.3.1 and E.6 at MOI 0,1 against E. coli strains. The positive control (c+) corresponds to the growth curve of E. coli ATCC 25922.
Figure 2.
Infection curves of phages E.1, E.3.1 and E.6 at MOI 0,1 against E. coli strains. The positive control (c+) corresponds to the growth curve of E. coli ATCC 25922.
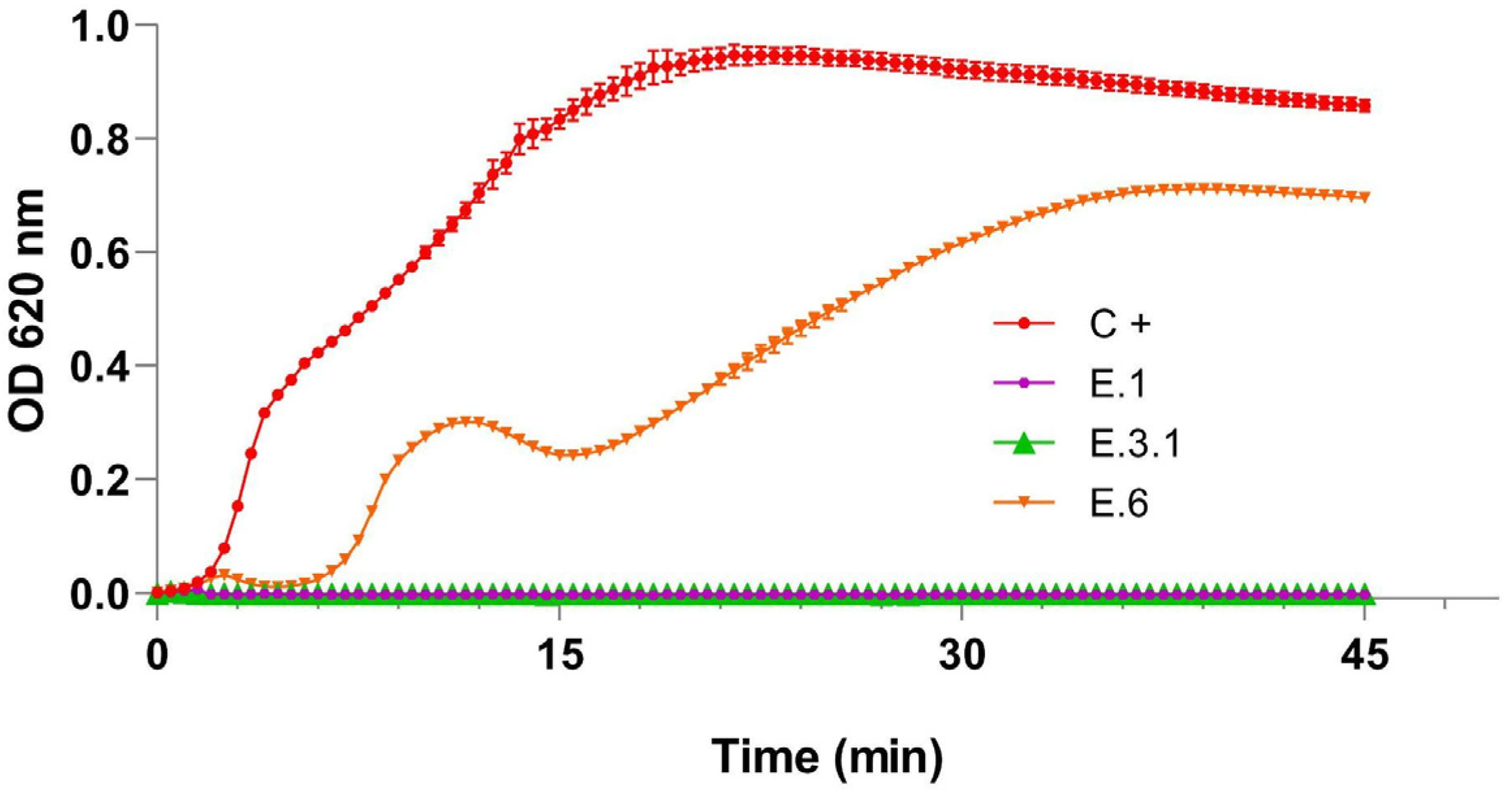
Figure 3.
Thermal stability of phages E.1, E.3.1, and E.9. ****Significant at p < 0.0001.
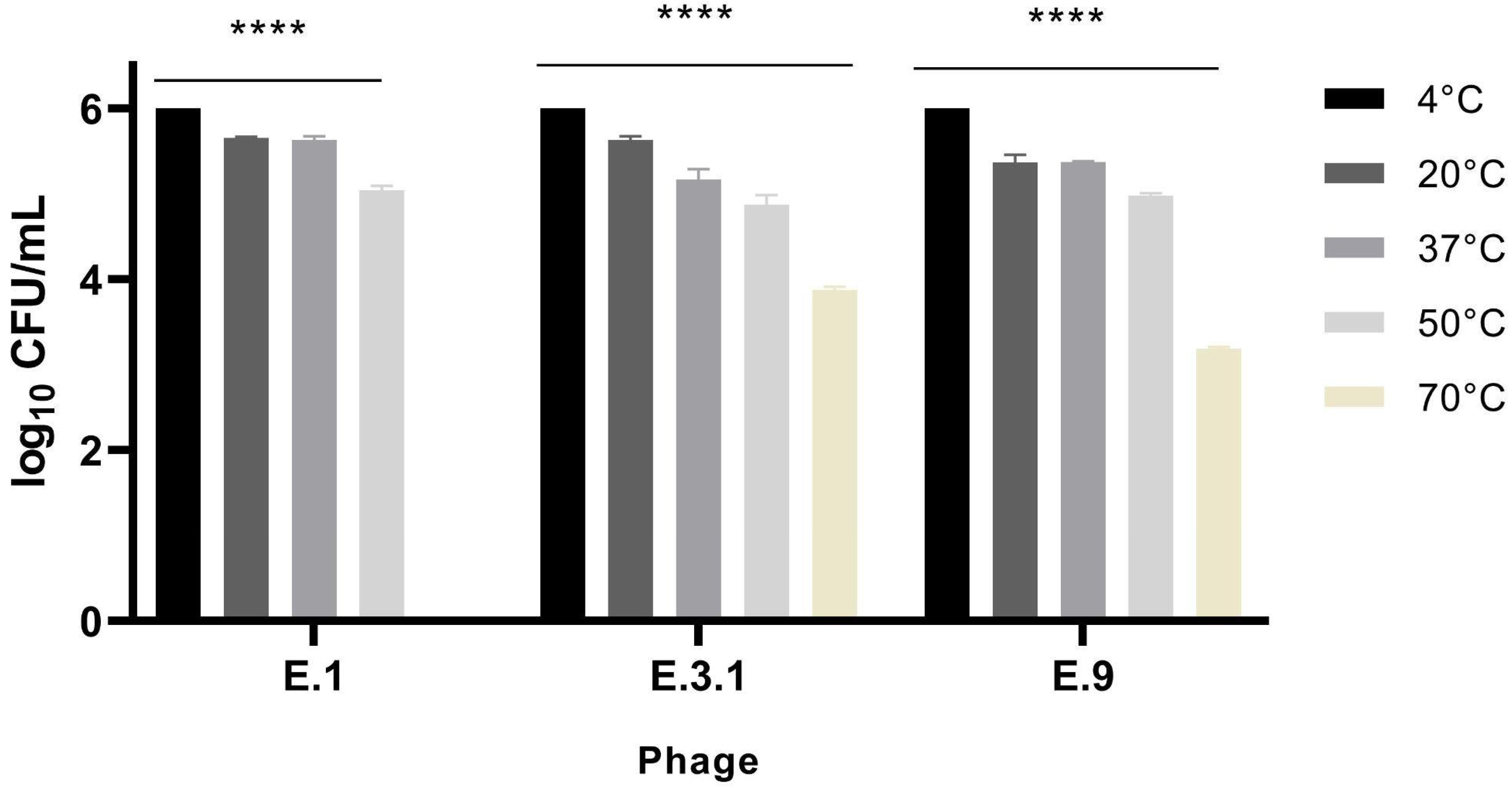
Figure 4.
One Step Growth curve of phage E.1 (A), phage E.3.1 (B) on E. coli ATCC 25922 and phage E.9 on E. coli B1348 (C).
Figure 4.
One Step Growth curve of phage E.1 (A), phage E.3.1 (B) on E. coli ATCC 25922 and phage E.9 on E. coli B1348 (C).
Figure 5.
In vitro assay at low temperature using the E. coli strain ATCC 25922 (A), and the E. coli strain B1348 (B). ****Significant at p < 0.0001; **Significant at p < 0.01; the others comparisons were not significant at p > 0.05.
Figure 5.
In vitro assay at low temperature using the E. coli strain ATCC 25922 (A), and the E. coli strain B1348 (B). ****Significant at p < 0.0001; **Significant at p < 0.01; the others comparisons were not significant at p > 0.05.
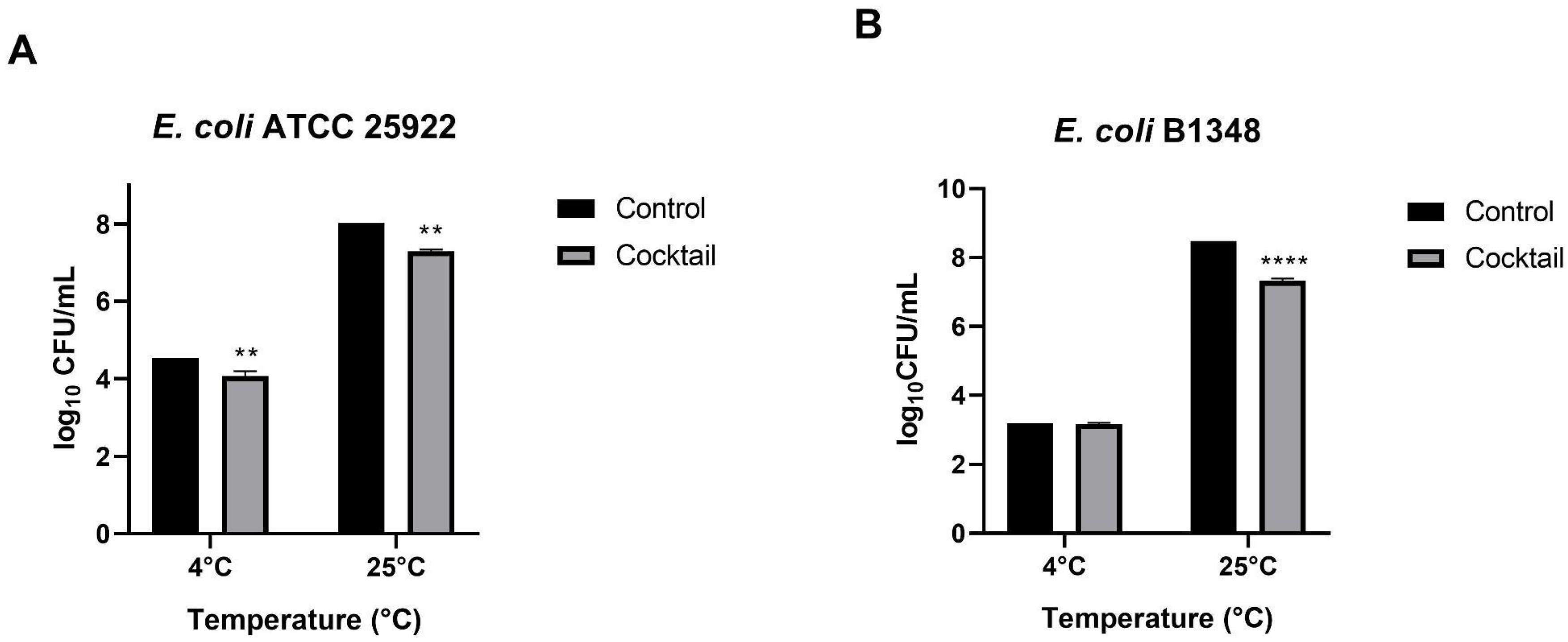
Figure 6.
Effects of the cocktail on the viability of E. coli strains in lettuce. Lettuce at 4°C (A) and 25°C (B) with E. coli ATCC 25922 and lettuce at 4°C (C) and 25°C (D) with E. coli B1348. All the values were tabulated as mean ± SD and a significant difference between variations was denoted by asterisks using two-way ANOVA. ***Significant at p < 0.001; **Significant at p < 0.01; *Significant at p < 0.1; the others comparisons were not significant at p > 0.05.
Figure 6.
Effects of the cocktail on the viability of E. coli strains in lettuce. Lettuce at 4°C (A) and 25°C (B) with E. coli ATCC 25922 and lettuce at 4°C (C) and 25°C (D) with E. coli B1348. All the values were tabulated as mean ± SD and a significant difference between variations was denoted by asterisks using two-way ANOVA. ***Significant at p < 0.001; **Significant at p < 0.01; *Significant at p < 0.1; the others comparisons were not significant at p > 0.05.
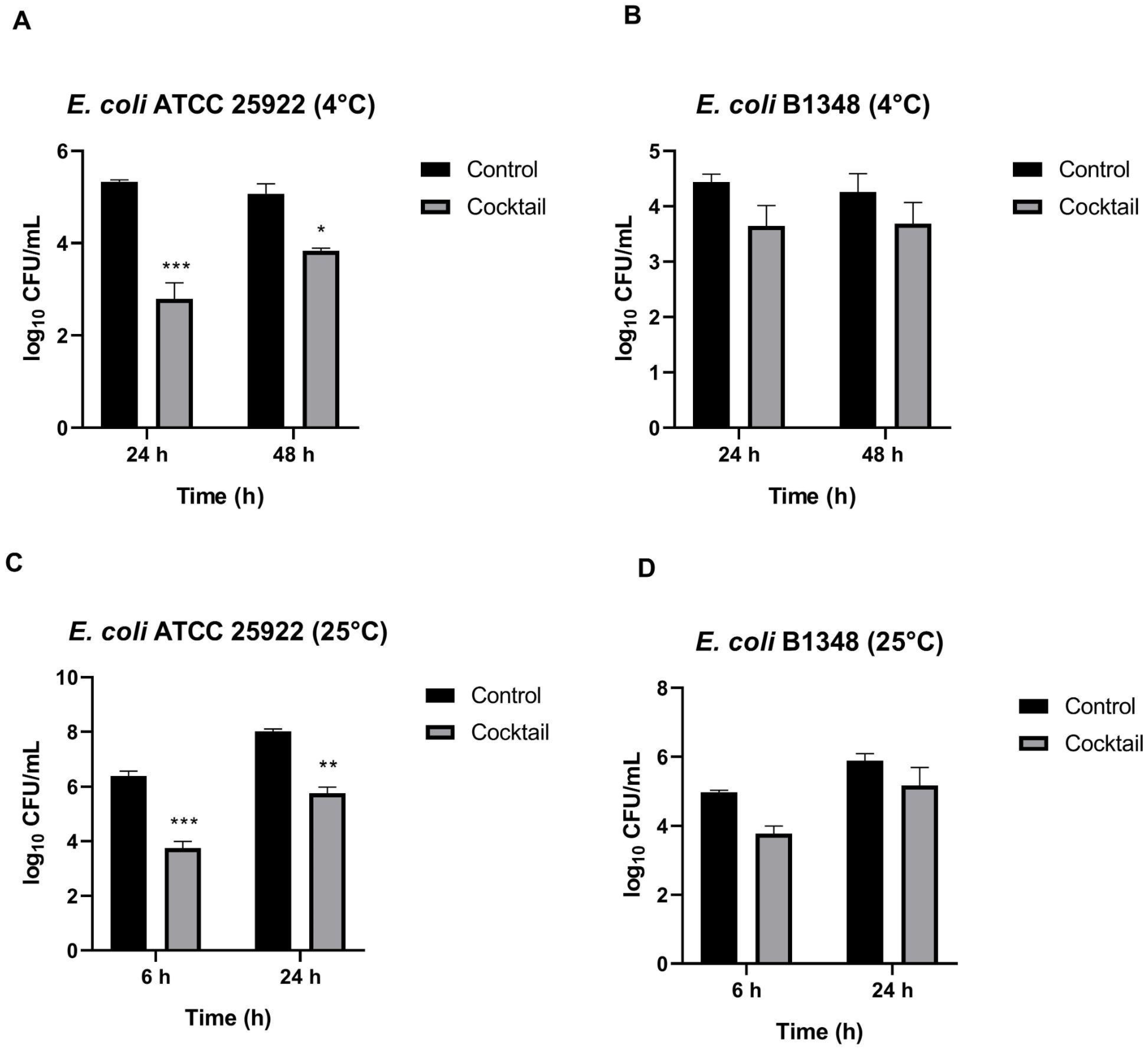

Table 1.
Characterization of lysis plates of isolated phages.
| Phage | WWS (date) | Size (plaque/halo, cm) | Plaque turbidity | Regular border | Halo |
|---|---|---|---|---|---|
| E.3.1 | WWS 3 (29/06/2023) | 0,320 ± 0,02/NO | No | Yes | No |
| E.1 | WWS 4 (19/07/2023) | 0,333 ± 0,05/NO | No | Yes | No |
| E.2 | WWS 4 (19/07/2023) | 0,334 ± 0,07 / 0,892 ± 0,12 | Yes | Yes | Yes |
| E.3 | WWS 4 (19/07/2023) | 0,337 ± 0,07 / 1,122 ± 0,19 | No | Yes | Yes |
| E.4 | WWS 4 (19/07/2023) | 0,353 ± 0,03 / 0,885 ± 0,09 | Yes | No | Yes |
| E.5.1 | WWS 5 (09/08/2023) | 0,165 ± 0,03 / 0,424 ± 0,08 | Yes | No | Yes |
| E.5.2 | WWS 5 (09/08/2023) | 0,574 ± 0,02 / 1,402 ± 0,09 | No | Yes | Yes |
| E.6 | WWS 6 (30/08/2023) | 0,193 ± 0,03/NO | Yes | Yes | No |
| E.7.1 | WWS 7 (22/09/2023) | 0,487 ± 0,06 / 1,223 ± 0,14 | No | Yes | Yes |
| E.9 | WWS 8: Pull de aguas | 0,155 ± 0,02/NO | No | Yes | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



