
Submitted:
13 March 2025
Posted:
14 March 2025
You are already at the latest version
Abstract
To develop a strategy for sustainable bioremediation of heavy metal-contaminated environments it is needed to understand the mechanisms of remediation using microorganisms. A huge bioremediation potential is possessed by fungi. Fusarium culmorum with their wide range of plant hosts can be the basis for creating sustainable phytoremediation technologies and for creating sustainable agriculture methods. Exopolymers (EPS) produced by F. culmorum can be an excellent metal sorbents and basic factors in the biosorption mechanism. The sorption capacities of Zn, Pb, and Cd by the EPS of a pathogenic DEMFc37 strain and two non-pathogenic strains (PGPF-DEMFc2 and DRMO-DEMFc5) were compared, as well as the effects of these metals on EPS synthesis by the three strains was determined. EPS samples were chemically characterized in regards to sugars, protein and phenolic compounds contents and used to study of metal binding. The concentration of metals bound/adsorbed to EPS was determined by Atomic Absorption Spectroscopy. The EPS of all the strains bound more than 80% of Zn as well as 64-84% of Cd and 74-79% of Pb, respectively. Thus, it has been clearly shown, that the use of F. culmorum EPS can be the basis for creating sustainable bioremediation, including phytoremediation.
Keywords:
bioremediation
; extracellular polymers (EPS)
; Fusarium culmorum
; cadmium
; lead
; zinc
; PCA
1. Introduction
Thanks to their many functional groups, microbial exopolymers can be excellent adsorbents of heavy metals (HMs) and play an important environmental role in the acquisition of resistance in metal-contaminated environments.
Extracellular polymeric substances (EPS) produced by microorganisms (bacteria and fungi) are characterised by diverse properties and great plasticity allowing their versatile use. First and foremost, they can protect microorganisms from various abiotic and biotic stresses [1,2] and can shape the soil environment as well as the interactions occurring in the environment between microorganisms and between microorganisms and plants [3]. The type of fungal endophytic strains effect on plants can be strongly modified by heavy metals present in the plant growth environment, whereby contact with metals can increase the resistance of fungi to other factors regulating their growth and development [4]. Toxicity of metals to filamentous fungi is manifested at the morphological level by inhibition of growth of the filaments, their decolourisation, disruption of sporulation and spore germination. At the cellular level, fungi respond by induction of antioxidant processes and the hormesis phenomenon plays an important role in enhancing adaptation to diverse plant host microenvironments [5].
Heavy metals such as copper and zinc are counted among the biological plant protection agents against diseases and are contained in numerous fungicides and the toxic effect of such zinc-based fungicides as zinc pyrithione is due to an increase in the cellular binding of this metal [6]. It is important to consider that most heavy metals (Cd, Cu, Hg, Pb, etc.) are antifungal agents. Some heavy metals, such as copper or zinc, are essential for growth and are physiologically important but most HMs are toxic, with the toxicity of physiological elements being concentration-dependent [7].
Filamentous fungi, including Ascomycota of the genus Fusarium show a high tolerance to the presence of heavy metals in the environment [8]. The problem of HMs accumulation in the frruiting bodies of saprophytic and ectomycorrhizal Basidiomycota fungi, which is species-specific and dependent on the concentration of metals in the substrate, is well known [9,10]. These fungi have been shown to differ in their ability to accumulate metals: Agaricus campestris, Macrolepiota procera and Boletus edulis strongly bound Cd, Pb and Hg, respectively [11].
Heavy metals affect not only the physiology but also the morphology of fungi by affecting both vegetative filaments and sporulation and asexual and sexual reproduction, but there are relatively few studies on this topic [5,11].
EPS primarily alter the surface area of both soil particles and the plants that colonise. They also provide a source of carbon and other biogenic elements (oxygen, hydrogen and nitrogen) and, thanks to numerous ligands, also cations such as sodium, potassium, magnesium, calcium, iron. Still quite poorly understood are the mechanisms of immobilisation (biosorption) of metals by EPS [4,13]. These properties of EPS may result from such physicochemical interaction processes occurring simultaneously and complementing each other, such as mechanical, physical and chemical sorption, complexation, precipitation or ion exchange. Physicochemical properties such as the ability to form a gel and to undergo denaturation and renaturation processes, during which the number of helices making up the EPS changes, e.g., from triple to single, are important to this process.
The metal complexation ability of EPS is mainly influenced by the presence of at least 10 functional groups such as amine -NH2, cyclic nitrogen =N-, carboxyl -COOH, amide -CONH2, cabonyl =CO, hydroxyl -OH, sulfonic -SO3H, sulfhydryl (thiol) -SH, phosphate-PO(OH)2, =POOH, phosphodiester [14,15,16]. It can be assumed that the relatively high tolerance of fungi to heavy metals is precisely related to the great diversity of functional groups in the wall polymers and especially in the exopolymers, which by binding heavy metal ions contribute to the resistance of fungi to these metals. However, most studies so far focus on the use of endophytic bacteria in the bioremediation of metal-contaminated soils [17,18]. Fungi, on the other hand, tend to be used in the bioremediation of sites contaminated by organic pollutants of the petroleum type and mycoremediation mainly uses their enzymes [19].
Among numerous metabolites, phytopathogenic fungal strains e.g., Plenodomus (Leptosphaeria): P. biglobosus and P. lingam infecting winter oilseed rape (Brassica napus L.) have been already shown to commonly produce EPS, and the level of those polymers is species-dependent [20].
The ability to produce EPS with antioxidant properties was demonstrated for three endophytic Fusarium culmorum fungal strains with different effects on cereal plant growth, i.e., two non-pathogenic strains: Plant Growth Promoting (PGPF)- DEMFc2 and Deleterious Rhizosphere Microorganism (DRMO) DEMFc5 and one pathogenic strain DEMFc37 colonising the rhizosphere, the border cell zone and, with various intensity, cereal plant tissues [21,22,23]. These strains have a strong environmental impact as they are potent of producing numerous CWDEs capable of degrading the plant and fungal cell wall [24,25] and a number of phytohormones [26] as well as affecting plants indirectly through the induction of resistance. The EPS of these strains is not only important in supporting the colonisation of soil and plant roots but also in increasing their tolerance to various stress conditions, including the presence of heavy metals. These strains, and the EPS they produce, could play a significant role in the rhizosphere and endosphere of cereal plants in heavy metal-contaminated areas by assisting in the phytoremediation of soils.
The aim of this study was to compare the metal (Zn, Pb, and Cd) absorption capacity of EPS of these three endophytic F. culmorum strains. Another objective was to demonstrate the influence of tested heavy metals on the EPS production capacity.
2. Materials and Methods
2.1. Fungal Strains
The experiments were carried out on three F. culmorum (W.G. Smith) strains exerting distinct effects on the growth of rye, wheat, and barley [21]: one non-pathogenic growth promoting PGPF (plant growth promoting fungi) strain DEMFc2 (CBS 120098, NCBI DQ453700), one non-pathogenic strain detrimental to growth DRMO (deleterious rhizosphere microorganism) DEMFc5 (CBS 120101, DQ450880), and one pathogenic isolate DEMFc37. The DEMFc2 and DEMFc5 strains were isolated from the rhizosphere of healthy rye (Secale cereale L.), and the DEMFc37 (CBS 120103, DQ450878) strain was isolated from winter wheat (Triticum aestivum L.) [27]. Location: Poland, Lublin region. Latitude, longitude coordinates where collected as followes: 51.15 N; 22.34 E. Additional geographic data: Lublin Plateau, continental temperate climate (-20°C to +30°C). The strains were deposited in the Fungal Collection of the Department of Environmental Microbiology (DEM) and in the Centraalbureau voor Schimmelcultures Collections (CBS), P.O. Box 85167, NL-3508 AD Utrecht, The Netherlands. The nucleotide sequences (18S rRNA, internal transcribed spacer 1; 5.8S rRNA, internal transcribed spacer 2; and the 28S rRNA gene [a partial sequence]) of the isolates were deposited in the NCBI GenBank.
2.2. Growth of Fungal Strains on a Medium with the Addition of Heavy Metals
Medium used to cultivation of fungal strains was Czapek-Dox [28,29] contained sucrose (30g), peptone (7.5g), dipotassium phosphate (1g), magnesium sulphate heptahydrate (0.5g), potassium chloride (0.5g), ferrous sulfate (0.01g). Initial pH was 7.0. Cultures were grown until three day of growth at 20°C. The medium was sterilized by heating to 121°C for 30 min. The optimal medium composition and cultivation time were determined in the previously published studies [23]. After sterilization, lead, zinc and cadmium ions in the form of nitrates (NO3-) were added to the medium at final concentrations of 2,4,6,8,10 ug / mL.
2.3. EPS Isolation and Chemical Characterisation
EPS was isolated from the tested strains from liquid cultures on Chapek-Dox medium as was described previously [23]. EPS preparation obtained directly after alcoholic precipitation and lyophilisation was referred to as crude EPS (C), and the preparation subjected to the further dialysis, freeze-drying and proteolysis, was mentioned as proteolysed EPS (P).
The total sugar in EPS preparations (C and P) was determined with the Dubois method [30], protein content was determined using Bradford method [31], and phenolic compounds were determined with the Folin-Ciocalteau method [32], as was described previously in Jaroszuk-Sciseł et al. [23].
For sugar analysis, the EPS samples (C and P) were hydrolysed with 2M trifluoroacetic acid (TFA) (100°C, 4 h). The sugars were converted into (amino)alditol acetates by N-acetylation, reduction with sodium borodeuteride (room temp., 18 h) and acetylation with a mixture of acetic anhydride/pyridine (1:1, v/v; 85°C, 30 min.) [33].
Methylation of proteolyzed samples (P) was performed according to the method of Hakomori [34], and the methylated polysaccharides were recovered by extraction with chloroform. Then, the resulting partly methylated polymers hydrolysed in 2M TFA (100°C, 4 h), reduced with sodium borodeuteride and converted into acetate derivatives.
Obtained (amino)alditol acetates, and partly methylated sugar derivatives were analysed by GLC-MS on a 7890A gas chromatograph (Agilent Technologies, USA) connected to a 5975C MSD detector (inert XL EI/CI, Agilent Technologies, USA). The chromatograph was equipped with an HP-5MS capillary column (Agilent Technologies, 30 m × 0.25 mm). Helium was a carrier gas, with a flow rate of 1 ml min-1. The temperature program was as follows: 150°C for 5 min, raised to 310°C at a rate of 5°C min-1, and the final temperature was kept for 10 min.
2.4. Estimation of the Sugar Polymer Masses by a Gel Permeation Chromatography (GPC)
The average molecular mass of the EPS (P) samples was determined by a gel permeation chromatography with a Sepharose CL-6B column (0.7 cm × 80 cm). Samples (5 mg) were dissolved in 1 ml of 0.5 M NaOH solution, and eluted with 0.5 M NaOH at a flow rate of 0.2 ml min−1. Fractions containing 1 ml of eluate (40 drops) were subjected to carbohydrate estimation by the phenol–sulphuric acid method [30]. Dextrans of known molecular masses (2 mln Da and 10 kDa) were used as standards to calibrate the column.
2.5. Binding of Metal Ions
The tested EPS lyophilisates were weighed in an amount of 1 mg/mL into an aqueous solution of lead, zinc and cadmium ions at a final concentration of 30 mg/mL. The resulting suspension (5 mL) was placed in cellulose dialysis tubes with a 12 kDa pore diameter (Sigma) and then shaken in flasks containing 30 mL of deionized water for a period of 24 and 48 hours. After this incubation time, the concentration of non-binding to EPS ions was determined in the solution outside the dialysis tube by Atomic Absorption Spectroscopy (ASA) using an Agilent AAS - 240 AA spectrometer. The percent ion binding was determined by the formula [10]:
% ion removal from solution
R = 100% × [(Ci − Cf)/Ci]
R - percentage of ion binding
Ci - initial concentration
Cf - final concentration
2.6. Statistical Analysis
All experiments were conducted in 3 biological replicates. Data are presented as mean values with standard deviation (SD). PCA were performed using the open-source software RStudio for Windows version 2024.12.0+467 (Posit, PBC, GNU Affero General Public Licence v3).
3. Results
3.1. Isolation and Chemical Characterization of F. culmorum EPS Preparations
EPS preparations obtained in the metal-free medium were used to study metal binding after 24- and 48-h incubation of lyophilised preparations at 20°C: (1) crude [C] and (2) proteolysed [P], for which the general composition and detailed characterization of sugar components were determined (Table 1, Table 2).
The crude EPS preparation of F. culmorum DEMFc2 strain differed significantly from the C preparation produced by the other two strains by having a 2 to 4-fold higher protein content and a slightly higher content of phenolic compounds. In contrast, the C preparation of EPS of strain DRMO DEMFc5 was characterised by a lower sugar content (Table 1). On the other hand, the proteolysed EPS of strain DEMFc2 was 4-fold lower than the P EPS of strain DEMFc5 and 5-fold lower than that of the pathogenic strain DEMFc37 and also had a lower phenolic compound content than the P preparations of the other two F. culmorum strains.
Sugar analysis of crude, lyophilised EPS preparations (Table 2) revealed that glucose (Glc) was the main component only in EPS from DEMFc2 strain, whereas crude EPS derived from DEMFc5 and DEMFc37 strains contained mainly mannose (Man) (Table 2). Galactose (Gal) comprised the minor structural component of all crude preparations. Samples after proteolysis showed slightly different composition, with an equivalent amounts of Man and Glc in case of DEMFc2 and DEMFc5 EPS preparations, whereas EPS from DEMFc37 contained mainly Man. Small amounts of glucosamine (GlcN) was present only in proteolysed EPS from DEMFc2 strain. These results are in general agreement with our previous findings [23].
Analysis of molecular mass distribution of EPSs after proteolysis, showed the presence of three fractions in preparation from DEMFc37 strain: high molecular weight (HMW) – around 1 MDa, medium molecular weight (MMW) – around 2 KDa and low molecular weight (LMW) – around 5 Da fractions. EPS from DEMFc2 contained only HMW and MMW fractions, whereas DEMFc5 contained only MMW fraction (Table 2).
Linkages analysis of EPS polymers (Table 2) showed, the presence of terminal hexose (Man), →3)-linked, →4)-linked, and →3,4)- as well as →3,6)-branched sugars in proteolysed preparation from DEMFc2 strain. EPS from DEMFc5 strain contained additionally →2)-linked sugar residues and lack of →3,4)-branched sugar units. Moreover, methylation analysis of the EPS from DEMFc37 strain showed the presence of terminal Man as the main component, small amounts of terminal Gal, →2), →4)- and →6)-linked sugar units, branched: →3,4)- and →3,6)-residues, as well as →2,3,4,6)-branched hexose units. That suggests, that analysed preparation from DEMFc37 strain was highly branched.
3.2. F. culmorum Strains Growth and EPS Synthesis on a Media Free of Heavy Metals and with the Addition of Heavy Metals
3.2.1. Growth of F. culmorum Strains
A strong inhibition of the growth of F. culmorum strains was observed in cultures with 2, 4, 6, 8 and 10 µg/mL of Cd and Pb, with a significant decrease already observed at the lowest applied concentration of 2 µg/mL (Figure 1 and Figure 2).
In contrast, the presence of Zn ions introduced at the same concentrations as Cd and Pb even caused an increase in the biomass of strains DEMFc5 (by about 20%) and DEMFc37 (usually by 70%). In contrast, in the PGPF strain DEMFc2 at a concentration of 2 µg Zn/mL, the biomass was 82% of the control (in metal-free culture) and dropped to 41% at 10 µg/mL (Figure 2).
3.2.2. EPS Synthesis by F. culmorum Strains
EPS was obtained in cultures of F. culmorum strains on Czapek-Dox medium without metals and in the presence of individual metals at concentrations of 2, 4, 6, 8, and 10 µg/mL.
The strongest inhibition of EPS production was found in cultures supplemented with lead ions (Figure 1 and Figure 2). It has been shown that the presence of lead had a strong effect on limiting EPS synthesis by the three investigated F. culmorum strains, while the effects of cadmium and zinc on EPS production by the individual F. culmorum strains were different (Figure 1 and Figure 2). Already at 2 µg/mL, a 90% decrease in exopolymer concentration was observed, but even at a concentration of 10 µg/mL Pb EPS was obtained (at ~0.05 g/L).
A positive correlation of EPS concentration of PGPF strain (DEMFc2) with cadmium concentration (2-10 μg/L) in the medium was observed (Figure 1). In cultures of strain DEMFc2 with Cd and Zn ions, EPS concentrations were up to 60% higher than in cultures without metals (Figure 2).
In contrast, the EPS concentration of the DRMO strain (DEMFc5) was even 2-fold lower in the presence of Cd than in the control, although no correlation was observed with the Cd concentration. The DEMFc5 strain in cultures with Cd and Zn produced EPS at 60-80% of the level in control cultures.
The concentration of EPS of the pathogenic DEMFc37 strain in the presence of Cd was strongly negatively correlated with the metal concentration. In the presence of Cd ions in the DEMFc37 strain cultures, the decrease in EPS concentration was correlated with an increase in Cd concentration and at a dose of 10 µg Cd/mL there was a 10-fold decrease in EPS concentration compared to metal-free cultures. A similar relationship was observed when introducing Zn ions into the culture (Figure 1 and Figure 2).
In cultures of PGPR DEMFc2 with Cd ions, a 2-fold increase in the efficiency (mg EPS/g biomass) of EPS production was recorded and in cultures of this strain with zinc ions this efficiency was even higher, reaching a value of 455% at the highest dose of 10 µg Zn/mL (Figure 1 and Figure 2). In the case of DRMO strain DEMFc5, a huge increase (up to 490%) in EPS generation efficiency was recorded in cultures with Cd ions (Figure 2).
3.3. Binding of Metal Ions – Chelating Properties of F. culmorum EPS
F. culmorum EPS obtained in the metal-free medium was used to study metal binding after 24- and 48-h incubation of lyophilised preparations at 20°C: (1) proteolysed and (2) crude, for which the general composition and detailed characterization of sugar components were determined. The concentration of metals bound/adsorbed to EPS placed in cellulose dialysis tubes was determined by Atomic Absorption Spectroscopy (ASA). The EPS of all the strains bound more than 80% of Zn as well as 64, 78, and 84% of Cd and 74, 77, and 79% of Pb, respectively, after 24 hours of shaking EPS in the metal salt solution.
Crude EPS of F. culmorum strains strongly sorbed metal ions after their 24-hour incubation in metal solution. The degree of immobilisation of the metals by the tested EPS after the longer incubation time (48 h) was several percent lower than after 24 h. After 48 hours of shaking, the degree of immobilization of the metals by these tested EPS was reduced compared to the immobilization noted after 24 hours, and as after 24 hours, differences were noted for EPS obtained from cultures of individual strains of F. culmorum. EPS of DEMFc2, DEMFc5 and DEMFc37 strains bound 30, 60 and 76% of zinc, 60, 70 and 70% of cadmium, and 45, 47 and 37% of lead, respectively.
Crude EPS preparations of strains DEMFc5 and DEMFc37 sorbed 80 85% of cadmium, lead and zinc ions from solution. Crude EPS of strain DEMFc2 showed greater variation in the level of biosorption of heavy metal ions, with the weakest sorption of Cd ions, (64%) and stronger sorption of Pb and Zn ions, at 74% and 81% respectively (Figure 3).
The deproteinised EPS sorbed metals weaker than crude EPS after 24 h incubation with metal ion solutions, while after 48 h incubation crude and deproteinised preparations were able to sorb at similar levels as for Pb ions, but in the case of the deproteinised EPS preparations of strain DEMFc2 they sorbed more strongly than crude EPS and even the sorption level increased relative to 24 h incubation.
It was reported that the sorption of Cd by the deproteinised EPS of all three strains increased with time and by crude slightly decreased. In contrast, Pb and Zn sorption decreased with incubation time with the exception of Pb sorption by the deproteinised EPS preparation produced by DEMFc37.
The strongest(by 50%) decrease in sorption with incubation time was observed for the crude EPS of strain DEMFc2.
3.4. Relationship Between Metal Biosorption and Type and Composition of EPS
Through PCA, it has been clearly demonstrated that, the process of biosorption of metals (Cd, Zn and Pb) by EPS DEMFc2 occurs differently than by two remaining strains (DEMFc5 and DEMFc37). Biosorption by EPS DEMFc2 depends on glucose (Figure 4A), as well as phenolic compounds and proteins (Figure 4C), while EPS DEMFc5 and DEMFc37 depend on the total concentration of sugars (Figure 4C), and among monomers on mannose, galactose and glucosamine (Figure 4A). No clear correlations can be observed between the biosorption of individual metals and sugar monomers (Figure 4B), nor with the main components of the EPS (proteins, phenolic compounds) or with the total sugar content of the EPS (Figure 4D).
Crude EPS carried out a stronger biosorption process than proteolysis EPS. The biosorption process by proteolysed EPS preparations was more closely related to the total sugar content (Figure 5B), and among the sugar components it was most strongly associated with galactose and glucose (Figure 5A). In the case of crude EPS, this process depends mainly on phenolic compounds and proteins, and in proteolysis on the content of sugars (Figure 5B), with the strongest influence of galactose and mannose (Figure 5A).
The relationship between EPS concentration, mycelial biomass and medium pH was completely different for the PGPF strain DEMFc2 than for the DRMO and pathogenic strains (Figure 6A). In the case of the DEMFc2 strain, there was a correlation between Cd concentration and EPS mycelial biomass, and the effects of Pb and Zn were strongly related to pH value and less strongly influenced the EPS concentration produced by this strain (Figure 6B.) Clearly, the effects of the individual heavy metals in the case of DRMO DEMFc5 were different: Zn had a strong positive effect on EPS concentrations and mycelial biomass and lead influenced pH value, while Cd had no clear effect on these parameters (Figure 6C). It was noticeable that, in the pathogen, the individual metals influenced the same parameters as in the DRMO strain, but the direction of the relationship between Zn concentration and biomass and EPS concentration was opposite (Figure 6D).
Zn had the strongest effect on the biomass and EPS concentration produced by the F. culmorum strains and Pb affected pH values (Figure 7A.) Cd had a different effect on the pathogen strain than on the biomass and EPS concentration of the DEMFc2 and DEMFc5 strains (Figure 7B). The dependence of the PGPF strain DEMFc2 on lead concentration was quite different from that of the DRMO and pathogen strain, in which Pb strongly affected pH as well as biomass and EPS concentration (Figure 7C). Zn had a clear effect on the biomass and pH values in the DRMO strain culture and on the EPS concentration in the pathogen culture, while Zn had a completely different effect on the PGPF strain (Figure 7D).
4. Discussion
A huge bioremediation potential is possessed by fungi inhabiting different environments: soil bulk, rhizosphere and plant endosphere, which are very poorly studied so far, compared to bacteria. The Fusarium culmorum fungi with their wide range of plant hosts, on which they can act phytopathogenically, but also non-pathogenically by promoting (PGPF) or weakening growth as deleterious microorganisms (DRMO), can be the basis for creating sustainable phytoremediation technologies and at the same time, by making plants resistant not only to abiotic but also biotic stresses, for creating sustainable agriculture methods. Particularly valuable in these methods is the possibility of using the exopolymers (EPS) produced by these fungi, which, thanks to their numerous functional groups, can be excellent metal sorbents and basic factors in the biosorption mechanism. In order to gain a detailed understanding of these mechanisms, the sorption capacities of Zn, Pb, and Cd by the EPS of a pathogenic DEMFc37 strain and two non-pathogenic strains: Plant Growth Promoting (PGPF)- DEMFc2 and Deleterious Rhizosphere Microorganism (DRMO) -DEMFc5, were compared, as well as the effects of these metals on EPS synthesis by the three strains was determined.
EPS was obtained in cultures of these strains on Czapek-Dox medium without metals and in the presence of individual metals at concentrations of 2, 4, 6, 8, and 10 µg/mL. EPS obtained in the metal-free medium was used to study metal binding after 24- and 48-h incubation of lyophilised preparations at 20°C: (1) proteolysed and (2) crude, for which the general composition and detailed characterization of sugar components, were determined. The concentration of metals bound/adsorbed to EPS placed in cellulose dialysis tubes was determined by Atomic Absorption Spectroscopy (ASA). It has been shown that the presence of lead had a strong effect on limiting EPS synthesis by the three investigated F. culmorum strains, while the effects of cadmium and zinc on EPS production by the individual strains were different. A statistically significant positive correlation of the concentration of EPS of the PGPF strain (DEMFc2) with the cadmium and zinc concentration in the medium was observed. In contrast, the concentration of EPS of the DRMO strain (DEMFc5) and the pathogenic strain DEMFc37 in the culture with these metals decreased two-fold and five-fold, respectively.
The EPS of all the F. culmorum strains tested sorbed the metals in solution rapidly (after 24-hour incubation) and to a very high (70-80%) degree. The crude EPS of all the strains bound more than 80% of Zn as well as 64, 78, and 84% of Cd and 74, 77, and 79% by EPS of DEMFc2, DEMFc5, DEMFc37, respectively, after 24 hours of shaking EPS in the metal salt solution. Anyway, the degree of immobilisation of the metals by the tested EPS after the longer incubation time (48 h) was several percent lower than after 24 h.
It should be emphasised that the crude EPS preparation of F. culmorum DEMFc2 strain did not bind more metals than the crude EPS of the other strains despite significantly (2- to 4-fold) higher protein content and also phenolic compounds. It is most likely that the biosorption of the three metals tested in this study by crude EPS of DEMFc2 was mainly mediated by the polysaccharide part, in which glucose predominates, while in crude EPS of the other strains the predominant sugar is mannose.
A similar relationship was described by da Silva et al. [35] for EPS obtained from cultures of Colletotrichum sp. The polymers of this strain were able to bind 79% of cadmium ions and 98% of lead ions. Similarly, a high affinity for metal ions was described by Vimalnat and Subramanian [36] for EPS obtained from a culture of Pseudomonas aeruginosa, which at high polymer concentrations in solution bound lead ions at 90-95%.
In the case of the benthic fungus Ascomycota Aspergillus penicillioides (F12), its ability to biosorb lead very rapidly (5.74 h) was demonstrated and optimisation of ecological parameters [pH, time (h) and temperature (°C) showed that the EPS of A. penicillioides under optimal conditions of pH (8.85) and temperature (32°C) a could sorb 73.14 % lead [37]. Paria et al. [37] also noted that at an EPS concentration of 0.5 mg/L of the fungus A. penicillioides, the highest (88.4%) flocculation capacity and as much as 50 % emulsifying activity of this fungal exopolymer was recorded.
The metals tested (Zn, Pb and Cd) were bound by hydroxyl, carboxyl and amino groups in the exopolymers of the fungus Ascomycota Pestolatiopsis sp. KCTC8637 [38] and Zygomycota Mucor rouxii [39].
In the case of pullulan, an EPS produced by another Ascomycota representative Aureobasidium pullulans on a metal-containing substrate, sorption of metals (Fe, Cu, Zn, Mn, Pb, Cd, Ni, Cr) from the substrate by pullulan was found, but at very low concentrations of hundredths of a ppm, while most of the metals from the substrate were sorbed by the biomass of this fungus [40]. In contrast, purified pullulan devoid of uronic acids and their carboxyl groups did not bind any of these metals.
Examination of the EPS biosorption capacity of F. culmorum strains showed that the biosorption capacity of the three metals tested (Cd, Zn, Pb) was present in both crude preparations and EPS after deproteinisation. In most cases, a 20-30% decrease in the amount of bound metal ions was observed. Only in the case of the EPS of the PGPF strain DEMFc2 was an increase in the affinity for cadmium ions observed in the deproteinised EPS.
The binding of Cd, Co and Cu by Saccharomyces cerevisiae was mainly mediated by mannan hydroxyl and protein thiol groups [41]. PCA indicated a relationship between heavy metal biosorption and mannan content in proteolysed EPS preparations of F. culmorum analysed in this study.
An alkali-soluble polysaccharide obtained from the cell wall of Boletus edulis capable of biosorbing Ni, Zn and, to the greatest extent, Cd and Pb was described as a (1→3)-linked α-d-glucan decorated by α-(1→3)- d-mannose chains [10]. The amide, phosphate and carboxyl groups of the EPS of the bacteria Bacillus subtilis and Pseudomonas putida were responsible for Zn binding [42] and thiol groups complexed especially strongly Cd, but also Zn and Pb as well as other metals (e.g., Co and Ni) in the envelope polymers of Shewanella oneidensis [47]. During biosorption of heavy metal cations (e.g., Cd, Zn, Pb) occurring by ion exchange sorption, there is a non-specific exchange between heavy metal cations present in solution and cation (H, Ca, K) functional groups by living and dead fungal cells of Saccharomyces cerevisiae and Rhizopus arrhizus [14].
Therefore, the ability of biosorption of metals by the F. culmorum EPS was found and although the concentration of immobilized metals decreased with time, the degree of immobilization was significant, especially in the case of cadmium and EPS of the pathogenic DEMFc37 strain.
PCA indicates a very clear and strong biosorption capacity for metals by crude EPS and indicates that this process depends most strongly on the protein components and phenolic compounds of the exopolymer. In contrast, the biosorption process of the tested heavy metals by proteolysed EPS preparations depends mainly on the total sugar content. The biosorption by crude EPS and proteolysed EPS was influenced by other sugar monomers. In the case of proteolysed EPS, these were galactose and mannose, while in the case of crude EPS, the biosorption process depended most strongly on glucose and arabinose.
The process of biosorption of metals (Cd, Zn and Pb) by EPS DEMFc2 occurs differently than by two remaining strains (DEMFc5 and DEMFc37). Biosorption by EPS DEMFc2 depends on phenolic compounds and proteins as well as glucose, while EPS DEMFc5 and DEMFc37 depend on the total concentration of sugars, and among monomers on mannose galactose and glucosamine.
Therefore, the use of plant inoculation with the PGPF strain as well as the use of EPS of this strain and other F. culmorum strains can be the basis for creating sustainable bioremediation, including phytoremediation and, at the same time, stimulating plant protection against phytopathogens through the induction of resistance.
5. Conclusions
F. culmorum strains with different plant effects (beneficial, harmful and pathogenic) were able to grow and produce EPS in the presence of heavy metals (Cd, Zn and Pb). The EPS produced by these F. culmorum strains sorbed Cd, Zn and Pb, irrespective of the type of their interaction with the plants. Differences could be observed between the PGPF strain DEMFc2 and the DRMO strains DEMFc5 and pathogenic DEMFc37, both in terms of the effects of heavy metals on fungal biomass and EPS production, as well as the biosorption of heavy metals by the EPS of the PGPF strain versus the biosorption of HMs by the EPS of the DRMO strain and the pathogen. The results of our study on the influence of metals on EPS production and biosorption capacity of EPS obtained from Fusarium culmorum strains culture contribute a great deal to the knowledge of plant interaction with rhizosphere and endophytic fungi and also open up new possibilities for the development of alternative methods for the bioremediation of soil contaminated by heavy metals, not only by the bioaugmentation technique but also by combining this technique with phytoremediation, which gives hope for the restoration of contaminated soils to agricultural use.
Author Contributions
Conceptualization, J.J-Ś., and A.N.; methodology J.J-Ś.,I.K., M.P-S., D.K. and A.N.; software, M.P-S.; validation, A.N., I.K. and J.J-Ś..; formal analysis, M.P.-S.; investigation, A.N., I.K., M.P.-S. and J.J-Ś..; resources, D.K.; data curation, A.N., I.K, and J.J-Ś..; writing—original draft preparation, J.J-Ś, A.N. and I.K..; writing—review and editing, J.J-Ś, A.N. and I.K..; visualization, J.J-Ś, M.P.-S. and A.N..; supervision, J.J-Ś. and I.K.; project administration, J.J-Ś. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The raw data of this article will be made available by the authors upon request.
Acknowledgments
The authors want to acknowledge M.Sc. Marzena Gęca form the Department of Inorganic Chemistry, Faculty of Chemistry at M. C.-S. University, Poland for technical support in ASA experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Awasthi, S.; Srivastava, P.; Mishra, P.K. Application of EPS in Agriculture: An Important Natural Resource for Crop Improvement. Agric. Res. Technol. Open Access J. 2017, 8, 8–10. [Google Scholar] [CrossRef]
- Barcelos, M.C.S.; Vespermann, K.A.C.; Pelissari, F.M.; Molina, G. Current Status of Biotechnological Production and Applications of Microbial Exopolysaccharides. Crit. Rev. Food Sci. Nutr. 2020, 60, 1475–1495. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.Y.A.; Raaijmakers, J.M.; Kuramae, E.E. Microbial Extracellular Polymeric Substances: Ecological Function and Impact on Soil Aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef] [PubMed]
- Gajewska, J.; Floryszak-Wieczorek, J.; Sobieszczuk-Nowicka, E.; Mattoo, A.; Arasimowicz-Jelonek, M. Fungal and oomycete pathogens and heavy metals: An inglorious couple in the environment. IMA Fungus 2022, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Vashistha, A.; Chaudhary, A. Effects of Biocontrol Agents And Heavy Metals in Controlling Soil-Borne Phytopathogens. Int. J. Herb. Med. 2019, 7(3), 41–45. [Google Scholar]
- Robinson, J.R.; Isikhuemhen, O.S.; Anike, F.N. Fungal–Metal Interactions: A Review of Toxicity and Homeostasis. J. Fungi 2021, 7, 225. [Google Scholar] [CrossRef]
- Balsalobre, L.; de Siloniz, M.I.; Valderrema, M.J.; Benito, T.; Larre, M.T.; Peinado, J.M. Occurence of Yeast in Municipal Wastes and Their Behavior in Presence Of Cadmium, Copper And Zinc. J. Basic Microbiol. 2003, 43, 185–193. [Google Scholar] [CrossRef]
- Ezzouhri, L.; Castro, E.; Moya, M.; Espinola, F.; Lairini, K. Heavy Metal Tolerance of Filamentous Fungi Isolated from Polluted Sites in Tangier, Morocco. Afr. J. Microbiol. Res. 2009, 3, 35–48. [Google Scholar]
- Falandysz, J.; Zhang, J.; Wang, Y.Z.; Saba, M.; Krasinska, G.; Wiejak, A.; Li, T. Evaluation of Mercury Contamination in Fungi Boletus Species from Latosols, Lateritic Red Earths, and Red and Yellow Earths in the Circum-Pacific Mercuriferous Belt of Southwestern China. PLoS ONE 2015, 10, e0143608. [Google Scholar] [CrossRef]
- Choma, A.; Nowak, K.; Komaniecka, I.; Waśko, A.; Pleszczyńska, M.; Siwulski, M.; Wiater, A. Chemical Characterization of Alkali-Soluble Polysaccharides Isolated from a Boletus edulis (Bull.) Fruiting Body and Their Potential for Heavy Metal Biosorption. Food Chem. 2018, 266, 329–334. [Google Scholar] [CrossRef]
- Širić, I.; Humar, M.; Kasap, A.; Kos, I.; Mioč, B.; Pohleven, F. Heavy Metal Bioaccumulation by Wild Edible Saprophytic and Ectomycorrhizal Mushrooms. Environ. Sci. Pollut. Res. 2016, 23, 18239–18252. [Google Scholar] [CrossRef]
- Lundy, S.D.; Payne, R.J.; Giles, K.R.; Garrill, A. Heavy Metals Have Different Effects on Mycelial Morphology of Achlya bisexualis as Determined by Fractal Geometry. FEMS Microbiol. Lett. 2001, 201, 259–263. [Google Scholar] [CrossRef]
- Osińska-Jaroszuk, M.; Jarosz-Wilkołazka, A.; Jaroszuk-Ściseł, J.; Szałapata, K.; Nowak, A.; Jaszek, M.; Ozimek, E.; Majewska, M. Extracellular Polysaccharides from Ascomycota and Basidiomycota: Production Conditions, Biochemical Characteristics, and Biological Properties. World J. Microbiol. Biotechnol. 2015, 31, 1823–1844. [Google Scholar] [CrossRef] [PubMed]
- Fomina, M.; Gadd, G.M. Biosorption: Current Perspectives on Concept, Definition and Application. Bioresour. Technol. 2014, 160, 3–14. [Google Scholar] [CrossRef]
- Volesky, B. Biosorption and me. Water Res. 2007, 41, 4017–4029. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.H.; Ismail, I.M.; Mostafa, T.M.; Sulaymon, A.H. Biosorption of Heavy Metals: A Review. J. Chem. Sci. Technol. 2014, 3, 74–102. [Google Scholar]
- Liu, L.; Quan, S.; Li, L.; Lei, G.; Li, S.; Gong, T.; Zhang, Z.; Hu, Y.; Yang, W. Endophytic Bacteria Improve Bio- and Phytoremediation of Heavy Metals. Microorganisms 2024, 12, 2137. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Singh, V.; Pandit, S. Advanced Omics Approach and Sustainable Strategies for Heavy Metal Microbial Remediation in Contaminated Environments. Bioresour. Technol. Rep. 2025, 29, 102040. [Google Scholar] [CrossRef]
- Dinakarkumar, Y.; Gnanasekaran, R.; Reddy, G.K.; Vasu, V.; Balamurugan, P.; Murali, G. Fungal Bioremediation: An Overview of the Mechanisms, Applications and Future Perspectives. Environ. Chem. Ecotoxicol. 2024, 6, 293–302. [Google Scholar] [CrossRef]
- Nowak, A.; Kutyła, M.; Kaczmarek, J.; Jaroszuk-Ściseł, J.; Jędryczka, M. Differences in the Production of Extracellular Polymeric Substances (EPS) and Other Metabolites of Plenodomus (Leptosphaeria) Infecting Winter Oilseed Rape (Brassica napus L.). Metabolites 2023, 13, 759. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E.; Winiarczyk, K.; Baturo, A.; Łukanowski, A. Colonization of Root Tissues and Protection Against Fusarium Wilt of Rye (Secale cereale) by Nonpathogenic Rhizosphere Strains of Fusarium culmorum. Biol. Control. 2008, 45, 297–307. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E.; Rodzik, B.; Winiarczyk, K. Interactions Between Rye (Secale cereale) Root Border Cells (RBCs) and Pathogenic and Nonpathogenic Rhizosphere Strains of Fusarium culmorum. Mycol. Res. 2009, 113, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Jaroszuk-Ściseł, J.; Nowak, A.; Komaniecka, I.; Choma, A.; Jarosz-Wilkołazka, A.; Osińska-Jaroszuk, M.; Tyśkiewicz, R.; Wiater, A.; Rogalski, J. Differences in Production, Composition, and Antioxidant Activities of Exopolymeric Substances (EPS) Obtained from Cultures of Endophytic Fusarium culmorum Strains with Different Effects on Cereals. Molecules 2020, 25, 616. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E.; Słomka, A.; Janczarek, M.; Rodzik, B. Activities of Cell Wall Degrading Enzymes in Autolyzing Cultures of Three Fusarium culmorum Isolates: Growth-promoting, Deleterious and Pathogenic to Rye (Secale cereale). Mycologia 2011, 103, 929–945. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E. Hydrolysis of Fungal and Plant Cell Walls by Enzymatic Complexes from Cultures of Fusarium Isolates with Different Aggressiveness to Rye (Secale cereale). Arch. Microbiol. 2012, 194, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Jaroszuk-Ściseł, J.; Kurek, E.; Trytek, M. Efficiency of indoleacetic acid, gibberellic acid and ethylene synthesized in vitro by Fusarium culmorum strains with different effects on cereal growth. Biologia 2014, 69, 281–292. [Google Scholar] [CrossRef]
- Kurek, E.; Machowicz, Z.; Kulpa, D.; Slomka, A. The Microorganisms of Rye [Secale cereal L.] Rhizosphere. Acta Microbiol. Pol. 1994, 2, 1053–1061. [Google Scholar]
- Czapek, F. Untersuchungen über die Stickstoffgewinnung. Hofmeisters Beiträge Zur Chem. Phys. U. Pathol. Bd. Ii 1902, 1, 559. [Google Scholar]
- Dox, A.W. The Intracellular Enzyms of Penicillium and Aspergillus: With Special Reference to Those of Penicillium camemberti; US Department of Agriculture, Bureau of Animal Industry: Washington, DC, USA, 1910; Volume 111.
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.t.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On tyrosine and tryptophane determinations in proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative Determination of Monosaccharides as Their Alditol Acetates by Gas Liquid Chromatography. Anal. Chem. 1965, 37, 1602–1604. [Google Scholar]
- Hakomori, S. A Rapid Permethylation of Glycolipid, and Polysaccharide Catalyzed by Methylsulfinyl Carbanion in Dimethyl Sulfoxide. J. Biochem. 1964, 55, 205–208. [Google Scholar]
- Da Silva, L.J; de Rezende Pinto, F.; do Amaral, L.A.; Gracia-Cruz, C.H. Biosorption of Cadmium (II) and Lead (II) from Aqueous Solution Using Exopolysaccharide And Biomass Produced by Colletotrichum sp. Desalin. Water Treat. 2014, 52(40-42), 7878-7886. [CrossRef]
- Vimalnath, S.; Subramanian, S. Studies on the Biosorption of Pb(II) Ions From Aqueous Solution Using Extracellular Polymeric Substances (EPS) of Pseudomonas aeruginosa. Colloids Surf. B Biointerfaces 2018, 172, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Paria, K.; Pyne, S.; Chakraborty, S.K. Optimization of Heavy Metal (Lead) Remedial Activities of Fungi Aspergillus penicillioides (F12) Through Extra Cellular Polymeric Substances. Chemosphere 2022, 286, 131874. [Google Scholar] [CrossRef]
- Moon, S.H.; Park, C.S.; Kim, Y.J.; Park, Y. Il Biosorption Isotherms of Pb (II) and Zn (II) on Pestan, an Extracellular Polysaccharide, of Pestalotiopsis sp. KCTC 8637P. Process Biochem. 2006, 41, 312–316. [Google Scholar] [CrossRef]
- Abdel-Aziz, M.S.; Hamed, H.A.; Mouafi, F.E.; Gad, A.S. Acidic pH-shock Induces the Production of an Exopolysaccharide by the Fungus Mucor rouxii: Utilization of Beet-Molasses. NY Sci. J. 2012, 5, 52–61. [Google Scholar]
- Radulović, M.D.; Cvetković, O.G.; Nikolić, S.D.; Dordević, D.S.; Jakovljević, D.M.; Vrvić, M.M. Simultaneous Production of Pullulan and Biosorption of Metals by Aureobasidium pullulans strain CH-1 on Peat Hydrolysate. Bioresour. Technol. 2008, 99, 6673–6677. [Google Scholar] [CrossRef]
- Brady, D.; Stoll, A.; Starke, L.; Duncan, J. Chemical and Enzymatic Extraction of Heavy Metal Binding Polymers from Isolated Cell Walls of Saccharomyces cerevisiae. Biotechnol. Bioeng. 1994, 44, 297–302. [Google Scholar] [CrossRef]
- Li, C.C.; Wang, Y.J.; Du, H.; Cai, P.; Peijnenburg, W.J.G.M.; Zhou, D.M. Influence of Bacterial Extracellular Polymeric Substances on the Sorption of Zn on γ-Alumina: A Combination of FTIR and EXAFS Studies. Environ. Pollut. 2017, 220, 997–1004. [Google Scholar] [CrossRef]
- Nell, R.M.; Fein, J.B. Influence of Sulfhydryl Sites on Metal Binding by Bacteria. Geoch. Cosm. Act. 2017, 199, 210–221. [Google Scholar] [CrossRef]
Figure 1.
Fungal dry biomass [g/L], EPS yield: EPS concentration [g/L], EPS concentration converted into fungal biomass dry weight [mg/g], and final pH of the post-culture liquid on Czapek-Dox medium with different concentration of A. cadmium, B. lead and C. zinc (2,4,6,8,10 µg/mL). Standard deviations are shown as deviation bars (n = 9).
Figure 1.
Fungal dry biomass [g/L], EPS yield: EPS concentration [g/L], EPS concentration converted into fungal biomass dry weight [mg/g], and final pH of the post-culture liquid on Czapek-Dox medium with different concentration of A. cadmium, B. lead and C. zinc (2,4,6,8,10 µg/mL). Standard deviations are shown as deviation bars (n = 9).
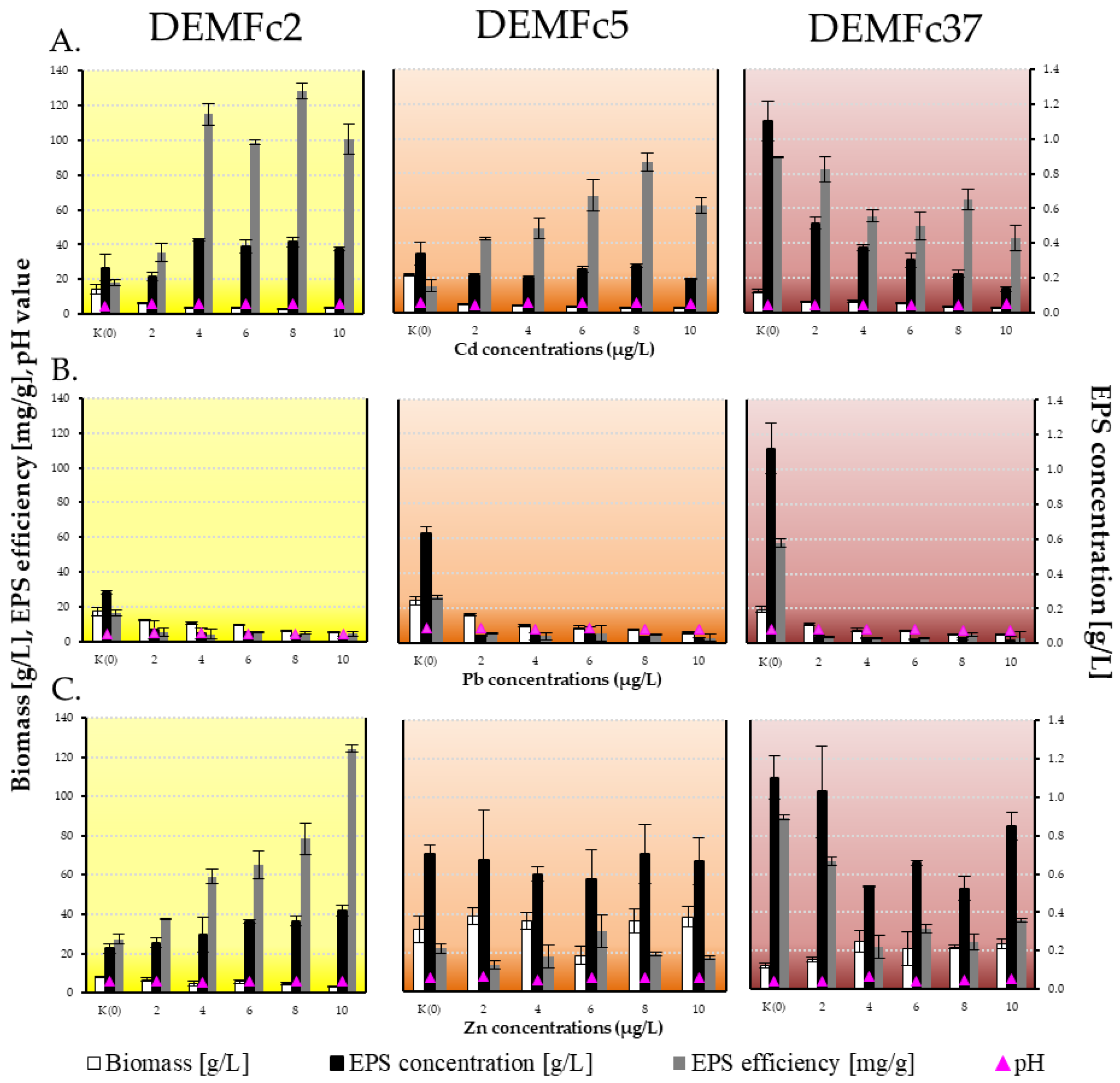
Figure 2.
Percentage against control of fungal dry biomass [g/L], EPS yield: EPS concentration [g/L], EPS concentration converted into fungal biomass dry weight [mg/g] on Czapek-Dox medium with different concentration of A. cadmium, B. lead and C. zinc (2,4,6,8,10 µg/mL). Standard deviations are shown as deviation bars (n = 9).
Figure 2.
Percentage against control of fungal dry biomass [g/L], EPS yield: EPS concentration [g/L], EPS concentration converted into fungal biomass dry weight [mg/g] on Czapek-Dox medium with different concentration of A. cadmium, B. lead and C. zinc (2,4,6,8,10 µg/mL). Standard deviations are shown as deviation bars (n = 9).
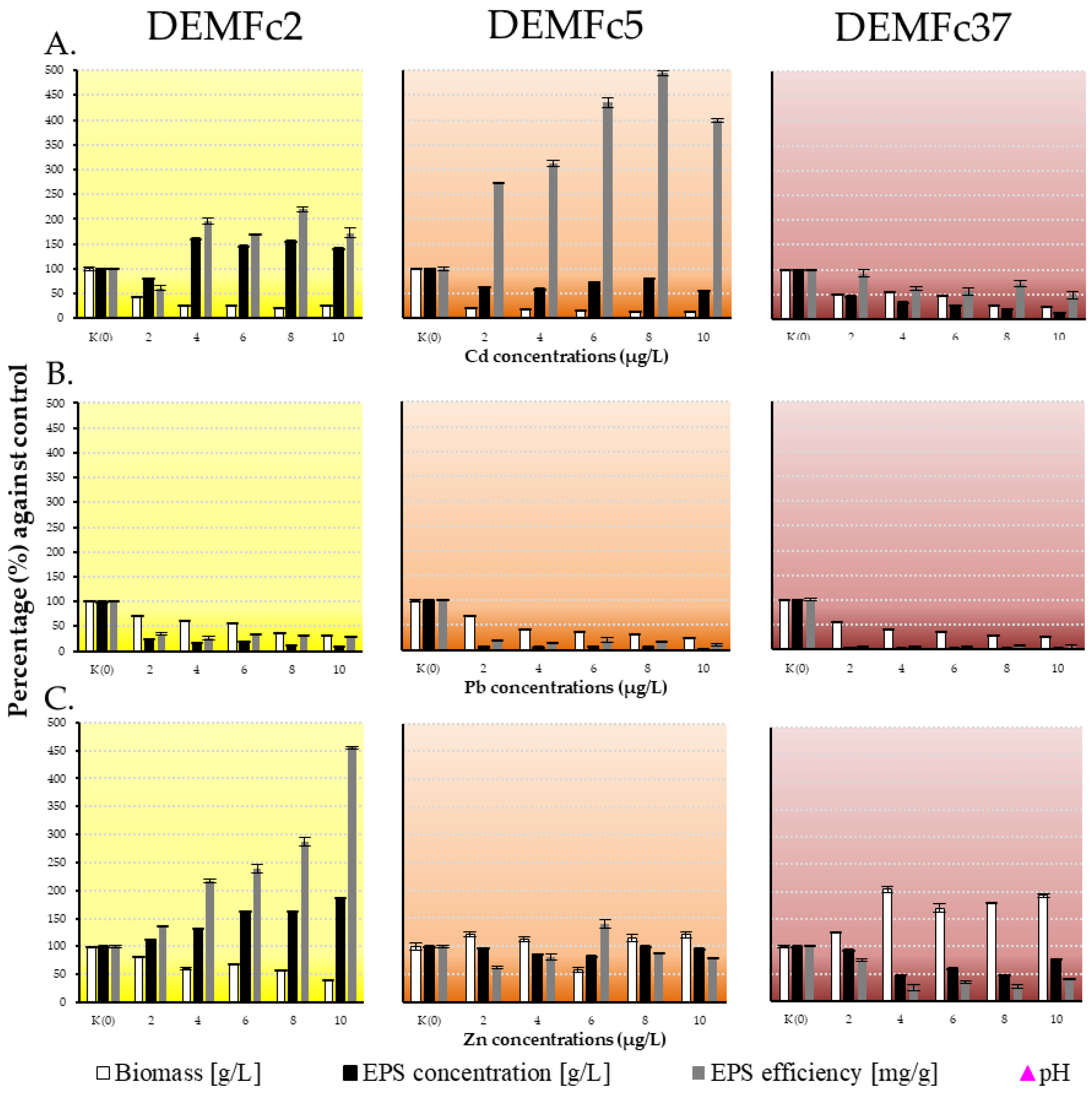
Figure 3.
Percentage of A. cadmium, B. lead and C. zink ion binding after 24 and 48 hour of incubation by Crude (C), and Proteolysed (P) EPS obtained from cultures of three F. culmorum strains: DEMFc2, DEMFc5, and DEMFc37.
Figure 3.
Percentage of A. cadmium, B. lead and C. zink ion binding after 24 and 48 hour of incubation by Crude (C), and Proteolysed (P) EPS obtained from cultures of three F. culmorum strains: DEMFc2, DEMFc5, and DEMFc37.
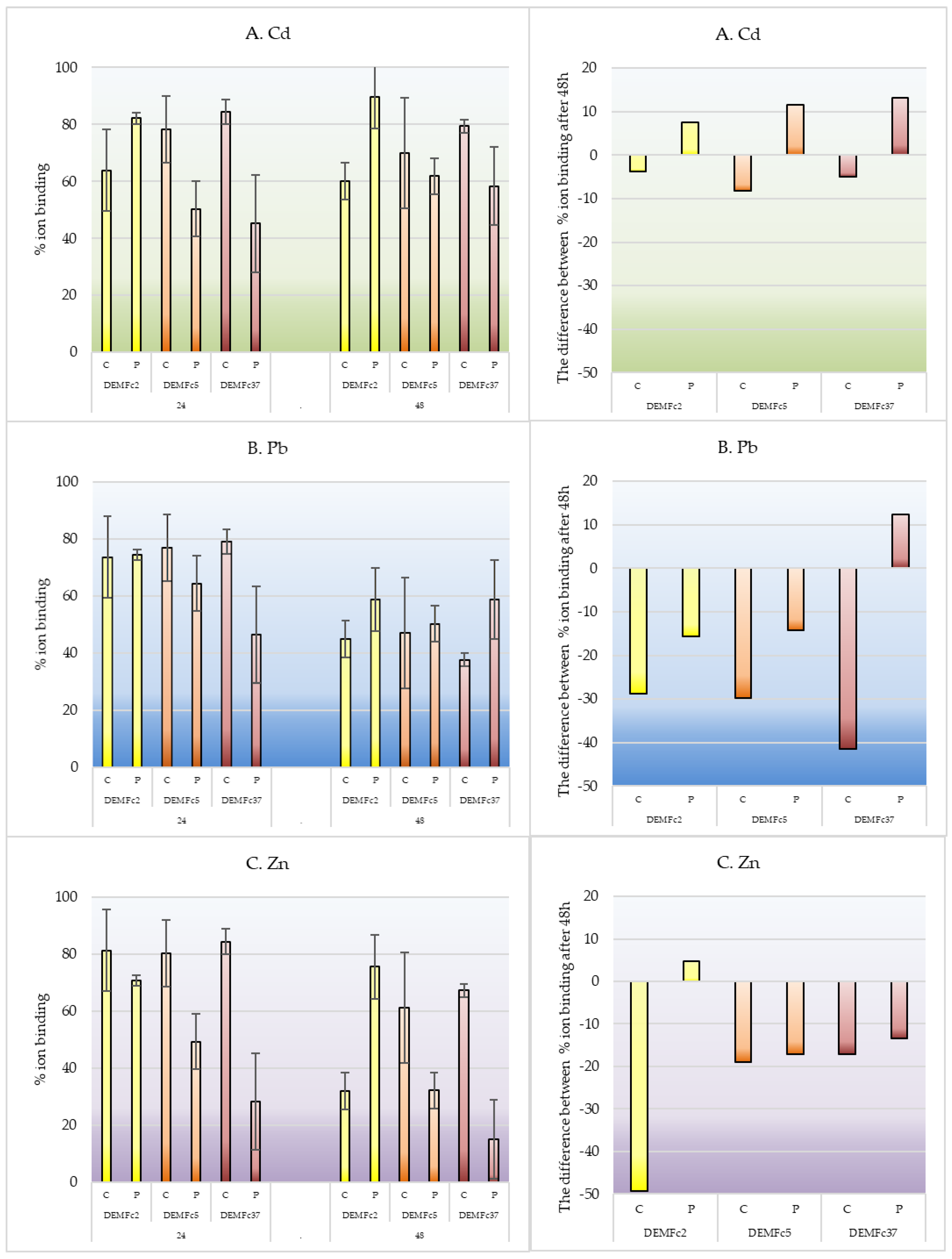
Figure 4.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. metal binding capacity (biosorption) and EPS-boosting monomers (Glc, Man, Gal, GlcN) within the tested strains (DEMFc2, DEMFc5, DEMFc37); B. metal binding capacity (biosorption) and EPS-bubbling monomers (Glc, Man, Gal, GlcN) within the tested metals (Cd, Pb, Zn); C. metal binding capacity (biosorption), EPS-building components (protein, total sugar, phenolic compounds) within the tested strains (DEMFc2, DEMFc5, DEMFc37); D. metal binding capacity (biosorption), growth components, obtained EPS (protein, total sugar, phenolic compounds) in the range of the tested metals (Cd, Pb, Zn).
Figure 4.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. metal binding capacity (biosorption) and EPS-boosting monomers (Glc, Man, Gal, GlcN) within the tested strains (DEMFc2, DEMFc5, DEMFc37); B. metal binding capacity (biosorption) and EPS-bubbling monomers (Glc, Man, Gal, GlcN) within the tested metals (Cd, Pb, Zn); C. metal binding capacity (biosorption), EPS-building components (protein, total sugar, phenolic compounds) within the tested strains (DEMFc2, DEMFc5, DEMFc37); D. metal binding capacity (biosorption), growth components, obtained EPS (protein, total sugar, phenolic compounds) in the range of the tested metals (Cd, Pb, Zn).
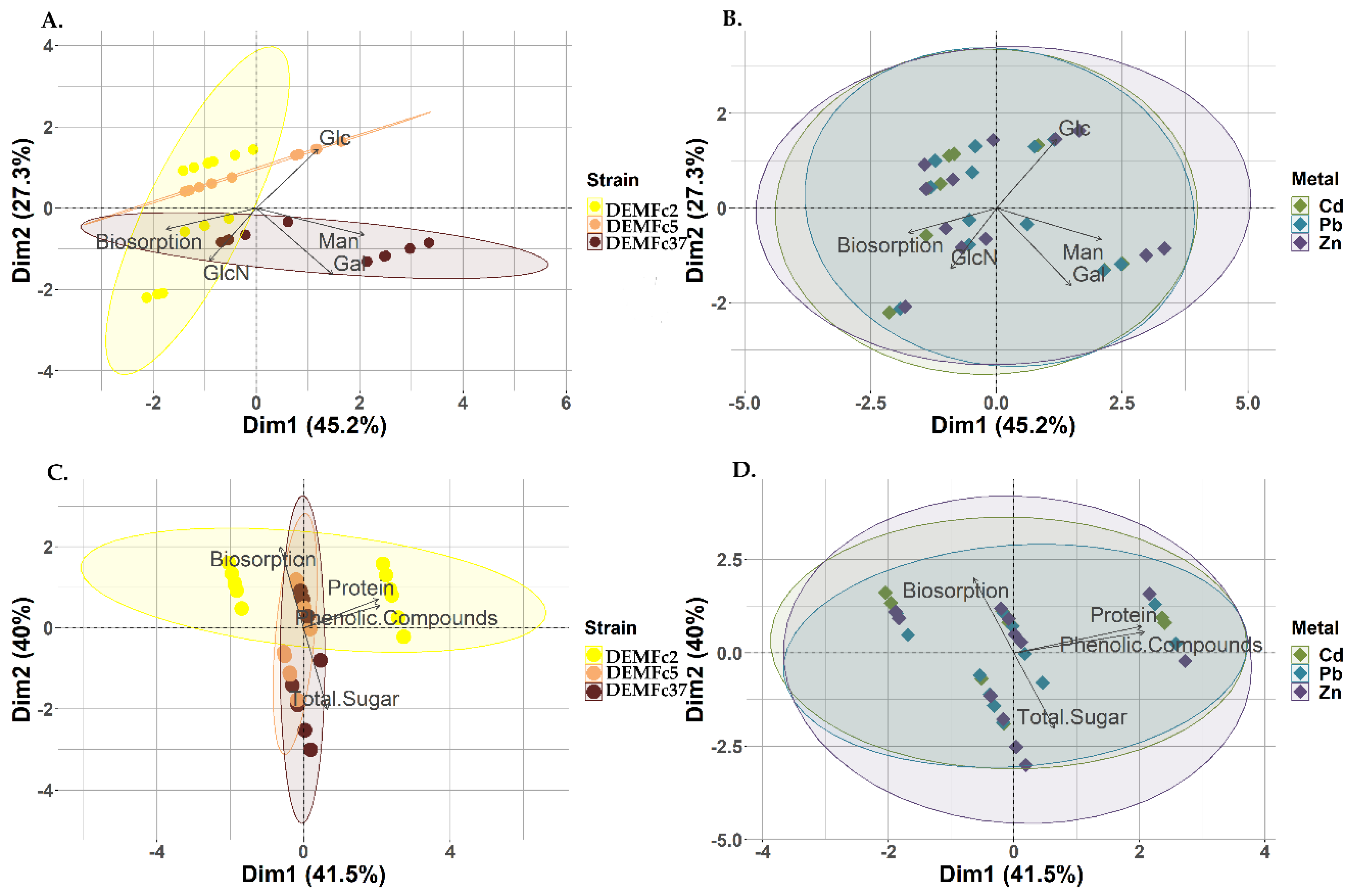
Figure 5.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. metal binding capacity (biosorption) and EPS-bubbling monomers (Glc, Man, Gal, GlcN) within the types of EPS tested (C, P); B. metal binding capacity (biosorption), bulking components of the resulting EPS (protein, total sugar, phenolic compounds) within the types of EPS (C, P) tested.
Figure 5.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. metal binding capacity (biosorption) and EPS-bubbling monomers (Glc, Man, Gal, GlcN) within the types of EPS tested (C, P); B. metal binding capacity (biosorption), bulking components of the resulting EPS (protein, total sugar, phenolic compounds) within the types of EPS (C, P) tested.
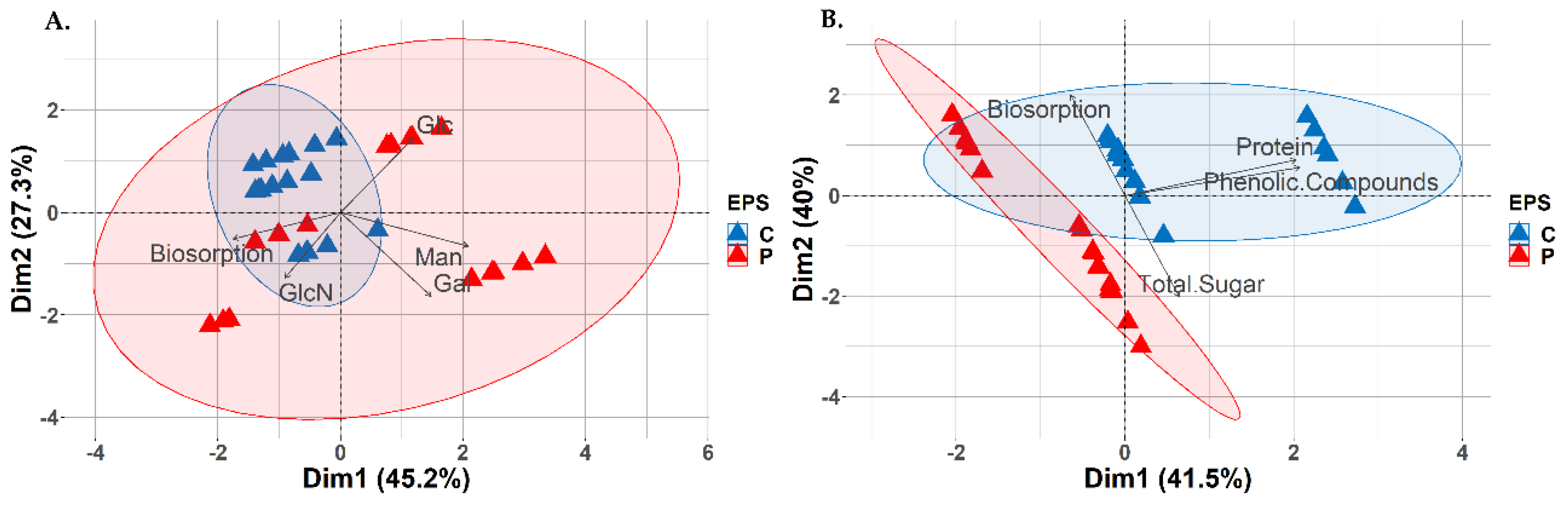
Figure 6.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH) within the tested strains (DEMFc2, DEMFc5, DEMFc37); B. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH)for the DEMFc2 strain within the tested metals (Cd, Pb, Zn); C. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH) for the DEMFc5 strain within the tested metals (Cd, Pb, Zn); D. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH) for the DEMFc37 strain within the tested metals (Cd, Pb, Zn).
Figure 6.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH) within the tested strains (DEMFc2, DEMFc5, DEMFc37); B. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH)for the DEMFc2 strain within the tested metals (Cd, Pb, Zn); C. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH) for the DEMFc5 strain within the tested metals (Cd, Pb, Zn); D. The amount of synthesized (EPS), the biomass obtained from the mycelium (Biomass), the final pH value (pH) for the DEMFc37 strain within the tested metals (Cd, Pb, Zn).
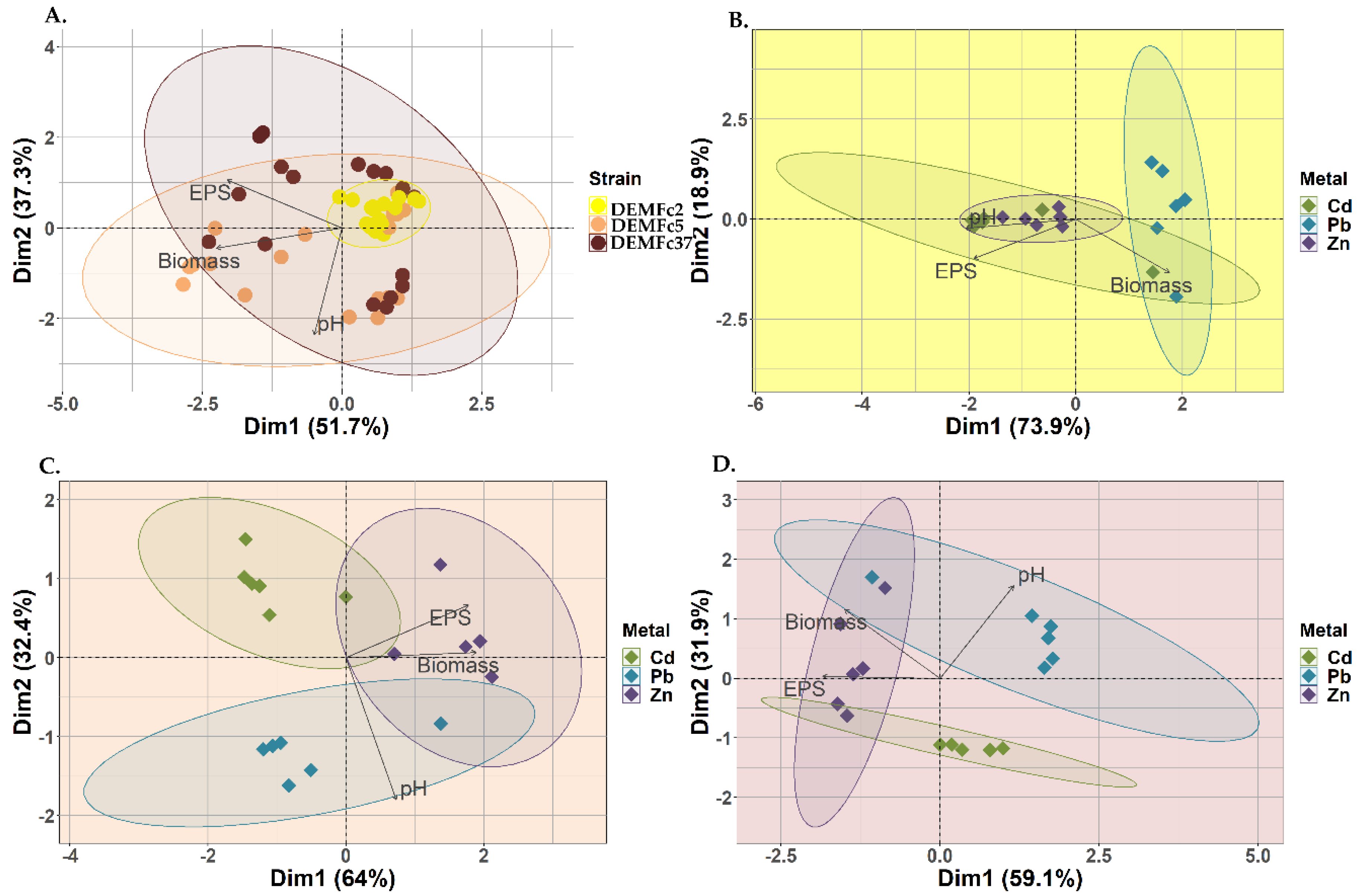
Figure 7.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) within the tested strains (DEMFc2, DEMFc5, DEMFc37); B. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) or the metal (Cd) within the tested strains (DEMFc2, DEMFc5, DEMFc37); C. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) or the metal (Pb) within the tested strains (DEMFc2, DEMFc5, DEMFc37); D. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) or the metal (Zn) within the tested strains (DEMFc2, DEMFc5, DEMFc37).
Figure 7.
Biplot of the Principal Component Analysis (PCA) describing corelations between: A. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) within the tested strains (DEMFc2, DEMFc5, DEMFc37); B. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) or the metal (Cd) within the tested strains (DEMFc2, DEMFc5, DEMFc37); C. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) or the metal (Pb) within the tested strains (DEMFc2, DEMFc5, DEMFc37); D. The amount of synthesized EPS, the biomass obtained (Biomass), the final pH value (pH) or the metal (Zn) within the tested strains (DEMFc2, DEMFc5, DEMFc37).
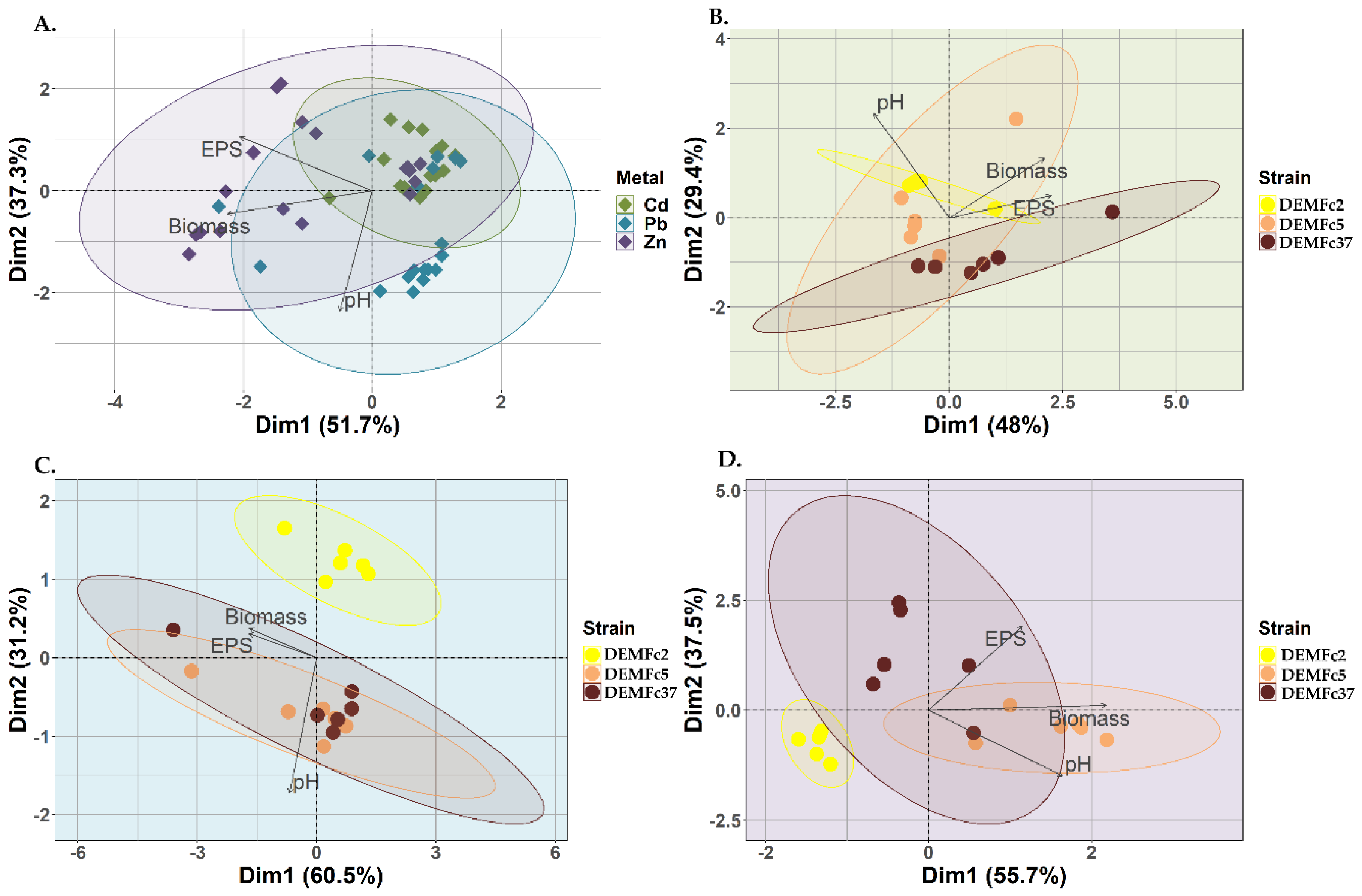

Table 1.
Composition, of water fraction of crude (C) and proteolysed (P) EPS preparations from F. culmorum strains: DEMFc2, DEMFc5 and DEMFc37.
Table 1.
Composition, of water fraction of crude (C) and proteolysed (P) EPS preparations from F. culmorum strains: DEMFc2, DEMFc5 and DEMFc37.
| Strain | DEMFc2 | DEMFc5 | DEMFc37 | |||
| Component | Composition of water fraction of EPS (µg/mL) | |||||
| C | P | C | P | C | P | |
| Protein | 2.3 | 0 | 1.2 | 0 | 0.44 | 0 |
| Phenolic compounds | 31.6 | 14.4 | 26.9 | 20.7 | 27.7 | 20.5 |
| Total sugar | 143.7 | 52.4 | 103.9 | 200.8 | 147.1 | 260.2 |
Table 2.
Sugar composition, molecular weights and linkage analysis of crude (C) and proteolysed (P) EPS preparations from F. culmorum strains: DEMFc2, DEMFc5 and DEMFc37.
Table 2.
Sugar composition, molecular weights and linkage analysis of crude (C) and proteolysed (P) EPS preparations from F. culmorum strains: DEMFc2, DEMFc5 and DEMFc37.
| Strain | DEMFc2 | DEMFc5 | DEMFc37 | |||
| Component | Sugar composition [µg/mg] ±SD* | |||||
| C | P | C | P | C | P | |
| Mannose (Man) | 6.97 (SD=1.670) | 12.40 (SD=0.552) | 29.22 (SD=3.466) | 58.39 (SD=3.828) | 37.39 (SD=4.957) | 128.99 (SD=19.94) |
| Glucose (Glc) | 36.14 (SD=4.230) | 12.69 (SD=1.419) | 18.18 (SD=3.200) | 75.48 (SD=9.038) | 15.18 (SD=3.760) | 34.04 (SD=5.726) |
| Galactose (Gal) | 3.12 (SD=1.290) | 8.01 (SD=0.329) | 2.80 (SD=0.129) | 6.09 (SD=0,531) | 9.41 (SD=1.299) | 13.59 (SD=1.574) |
| Glucosamine (GlcN) | - | 1.09 (SD=0.155) | - | - | - | - |
| Fraction** | Molecular weights [kDa] | |||||
| P | P | P | ||||
| HMW | 1000-735 | - | 1000-735 | |||
| MMW | 15.8 | 73.5 - 34.1 | 18.5 | |||
| LMW | - | 5.4 | ||||
| Type of linkage | Glycosidic bonds [%] | |||||
| P | P | P | ||||
| tHex I (1→(Man)) | 19.5 | 21.13 | 43.56 | |||
| tHex III (1→(Gal)) | - | - | 2.46 | |||
| →2)Hex(1→ | - | 19.81 | 21.32 | |||
| →3)Hex (1→ | 26.81 | 12.67 | - | |||
| →4)Hex(1→ | 22.83 | 35.36 | 4.83 | |||
| →6)Hex (1→ | - | - | 6.38 | |||
| →3,4)Hex(1→ | 14.97 | - | 1.79 | |||
| →3,6)Hex(1→ | 15.89 | 11.03 | 11.46 | |||
| →2,3,4,6)Hex(1→ | - | - | 8.2 | |||
* SD values were
calculated based on the results from three independent repetitions of sugar
analysis. **HMW – high molecular weight; MMW – medium molecular weight; LMW –
low molecular weight.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



