
Submitted:
07 March 2025
Posted:
07 March 2025
You are already at the latest version
Abstract
Erythropoietin-producing hepatocellular receptor A1 (EphA1) is one of the Eph receptor family members, the largest group of receptor tyrosine kinases. EphA1 is expressed in various tissues and regulates cellular homeostasis by interacting with its membrane-bound ephrin ligands and other receptors. EphA1 critically correlates with the pathogenesis in several disorders, including Alzheimer’s disease and cancers. Therefore, establishing sensitive monoclonal antibodies (mAbs) for EphA1 has been desired for basic research, diagnosis, and treatment. In this study, a novel specific and sensitive anti-human EphA1 mAb, clone Ea1Mab-30 (mouse IgG1, kappa), was established by the Cell-Based Immunization and Screening (CBIS) method. Ea1Mab-30 demonstrated reactivity with an EphA1-overexpressed Chinese hamster ovary-K1 cell line (CHO/EphA1), an endogenously EphA1-expressing bladder carcinoma cell line (5637), and a colorectal adenocarcinoma cell line (Caco-2) in flow cytometry. Crossreactivities of Ea1Mab-30 with other Eph receptors were not observed. Furthermore, Ea1Mab-30 demonstrated high binding affinities (dissociation constant; KD) for CHO/EphA1 (KD: 8.9×10-9 M) and 5637 (KD: 1.7×10-9 M) in flow cytometric analysis. Furthermore, Ea1Mab-30 detected EphA1 protein in CHO/EphA1 and 5637 lysates using western blot analysis. Ea1Mab-30 also clearly stained EphA1 of formalin-fixed paraffin-embedded CHO/EphA1 using immunohistochemistry. Ea1Mab-30, established by CBIS method, could help analyze the EphA1-contributed cellular functions and have potential applications in pathological diagnosis and treatment with specificity and high affinity for EphA1-expressing cells.
Keywords:
Eph receptor
; EphA1
; CBIS method
; monoclonal antibody
; flow cytometry
; western blot
; immunohistochemistry
1. Introduction
Erythropoietin-producing hepatocellular (Eph) receptors are single-pass transmembrane glycoproteins, which belong to the most prominent family of receptor tyrosine kinases (RTKs) [1,2]. Eph receptors play pivotal roles in tissue homeostasis, including cell proliferation, migration, and angiogenesis by binding to their ligands, ephrin [1,2]. Since it was first identified in hepatocellular carcinoma cell line in 1987, the fourteen Eph receptors (EphA1 to EphA8, EphA10, EphB1 to EphB4, and EphB6) and eight ephrin ligands [glycosylphosphatidylinositol (GPI)-anchored ephrin A1 to A5 and transmembrane ephrin B1 to B3] have been identified so far [3]. Eph receptors are topics of potential targets for diseases such as cancers because the aberrant functions of Eph receptors disrupt cellular homeostasis and induce tumor development.
EphA1 was firstly isolated from an erythropoietin-producing human hepato-cellular carcinoma cell line (ETL-1) [3]. EphA1 is expressed in epithelial and nervous tissues, including skin, neuron, and colon crypt base, and controls many cellular functions, such as cell morphology, motility, axonal guidance, and synaptic plasticity [4,5,6,7,8]. EphA1 expression is also confirmed in T-lymphocytes, monocytes [9]. EphA1-knockout (KO) mice exhibited abnormal tail development as a kinky-tail, and an uterovaginal defection caused by a deficiency in apoptosis during reproductive tract canalization [10]. EphA1 plays an essential role in normal tissue development. According to the genome-wide association studies (GWAS) of Alzheimer’s disease (AD) patients, EphA1 has been considered as an associated factor for AD [11,12,13]. AD is a progressive age-associated neurodegenerative disorder leading to loss of autonomy, which pathologically defined by extensive neuronal loss, the accumulation of neurofibrillary tangles, and extracellular amyloid plaques in the brain [14,15,16]. Missense mutations in EphA1 gene have also been reported that may contribute to the development of AD by disrupting receptor signaling [17]. Some single nucleotide polymorphisms (SNPs), rs11767557 and rs11771145, which affect the EphA1 gene, have been reported to decrease the risk of late-onset AD (patients of over 65 years old) [11,13,18]. Furthermore, rs11767557 variant is associated with EphA1 transcription in human whole blood data analysis. Meanwhile, rs11767557 SNP has not shown the significant effect of EphA1 transcription in brain tissue [19]. Moreover, the EphA1 transcription is not altered in brain with AD [20]. The discrepancies in these results may be due to the size and heterogeneity of the samples Since few molecular analyses have been conducted to clarify the regulation of EphA1 in AD, further investigation is required to determine whether EphA1 is involved in the risk of AD.
The EphA1 upregulation is also correlated in various cancers, including esophageal [21], colon [3], gastric [22,23], and prostate cancers [24,25]. EphA1 high expression associates with unfavorable overall survival and higher tumor stage [21,26]. EphA1 has been implicated as a marker for aggressive ovarian cancer stem cells with co-expression with aldehyde dehydrogenase 1A1, CD9, and CD24 [27]. Furthermore, ADAM12 metalloproteinase-cleaved ephrinA1 facilitates tumor cell entry into the lungs by activating EphA1 [28]. While EphA1 is focused as a cancer-promoting factor, tumor suppressor functions of EphA1 have also been proposed. Reduced expression of EphA1 correlates with tumor invasion, lymph node metastasis, and poor outcomes in several cancers, including colon, skin, and ovarian cancers [29,30,31,32,33]. The inhibition of cell spreading and Rac1 activation, which promotes cell motility, is observed by EphA1 activation [5]. It is necessary to clarify the role of EphA1 in promoting or suppressing tumors.
Previously, we have successfully developed several monoclonal antibodies (mAbs) against Eph receptors, including human EphA2 (clone Ea2Mab-7) [34], human EphB2 (clone Eb2Mab-12) [35], human EphB4 (clone B4Mab-7) [36], and human EphB6 (clone Eb6Mab-3) [37] by using the Cell-Based Immunization and Screening (CBIS) method. The CBIS method is an efficient strategy for developing antibodies with a wide variety of epitopes such as structural epitopes and linear epitopes, as well as modifications of cell surface membrane proteins in a short period. In this study, we have established a novel anti-human EphA1 mAb clone Ea1Mab-30 using the CBIS method.
2. Materials and Methods
2.1. Cell Lines
Chinese hamster ovary (CHO)-K1, LN229 glioblastoma, P3X63Ag8U.1 (P3U1) myeloma, and Caco-2 colorectal adenocarcinoma were obtained from the American Type Culture Collection (Manassas, VA, USA). 5637 bladder carcinoma was obtained from the Cell Resource Center for Biomedical Research, Institute of Development, Aging and Cancer, Tohoku University (Miyagi, Japan). The complementary DNA (cDNA) of EphA1 (Catalog No.: RC213689, Accession No.: NM_005232) plus an N-terminal MAP tag (GDGMVPPGIEDK) [38] recognized by an anti-MAP tag mAb (PMab-1) or an N-terminal PA tag (GVAMPGAEDDVV) [39] recognized by an anti-PA tag mAb (NZ-1), were subcloned into a pCAGzeo vector [FUJIFILM Wako Pure Chemical Corporation (Wako), Osaka, Japan]. Afterward, plasmids were transfected into CHO-K1 and LN229 cells using the Neon transfection system (Thermo Fisher Scientific, Inc., Waltham, MA). Stable transfectants [CHO/MAP-EphA1, CHO/PA-EphA1 (CHO/EphA1), LN229/MAP-EphA1, and LN229/PA-EphA1 (LN229/EphA1)] were subsequently selected using a cell sorter (SH800, Sony Corp., Tokyo, Japan) using anti-tag mAbs, PMab-1 and NZ-1, respectively. After sorting, cultivation in a medium containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA, USA) was conducted.
GeneArt™ CRISPR nuclease vectors with OFP plasmid which target EphA1 (TrueGuide™ Synthetic sgRNA, CRISPR466292_SGM) was purchased from Thermo Fisher Scientific Inc. The EphA1 KO 5637 cell line (BINDS-38) was generated by transfecting the vector using the Neon Transfection System (Thermo Fisher Scientific, Inc.). Stable transfectants were established by cell sorting using SH800 (Sony Corp.).
Other Eph receptor-expressing CHO-K1 cells (e.g., CHO/EphA2) were established as previously reported [35].
CHO-K1, P3U1, 5637, BINDS-38, and all Eph receptor-overexpressed CHO-K1 cells were cultured in a Roswell Park Memorial Institute (RPMI)-1640 medium (Nacalai Tesque, Inc.) that was supplemented with 10% fetal bovine serum which inactivated by heating (FBS, Thermo Fisher Scientific Inc.), antibiotics consisting of 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). LN229, LN229/MAP-EphA1, LN229/EphA1, and Caco-2 cells were cultured in a Dulbecco’s Modified Eagle Medium (DMEM) (Nacalai Tesque, Inc.) that was supplemented with 10% FBS which inactivated by heating (Thermo Fisher Scientific Inc.), antibiotics consisting of 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). Then, all cells were cultured in 5% CO2 and 95% air under humidified conditions at 37°C.
2.2. Antibodies
A purified anti-human EphA1 Antibody (clone A20047D, rat IgG2a, kappa) was purchased from BioLegend (San Diego, CA, USA). An anti-isocitrate dehydrogenase 1 (IDH1) mAb (clone RcMab-1) was developed previously [40]. Alexa Fluor 488-conjugated anti-mouse IgG and Alexa Fluor 488-conjugated anti-rat IgG were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). Secondary horseradish peroxidase-conjugated anti-mouse IgG and anti-rat IgG were obtained from Agilent Technologies Inc. (Santa Clara, CA, USA) and Merck KGaA (Darmstadt, Germany), respectively.
2.3. Development of Hybridomas
The Animal experiments were approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2019NiA-001) and were carried out in accordance with the NIH (National Research Council) Guide for the Care and Use of Laboratory Animals. For developing anti-EphA1 mAbs, two 5-week-old female BALB/cAJcl mice purchased from CLEA Japan (Tokyo, Japan) were immunized with CHO/MAP-EphA1 (1 × 108 cells/mouse) via the intraperitoneal route starting at 6-week-old. The CHO/MAP-EphA1 cells as immunogen were harvested after brief treatment with 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.) to avoid antigen digestion. Imject Alum Adjuvant (Thermo Fisher Scientific Inc.) was added to the immunogen cells in the first immunization. Three additional injections of CHO/MAP-EphA1 (1 × 108 cells/mouse) were conducted intraperitoneally without an adjuvant addition every week. A last booster injection was also performed with 1 × 108 cells/mouse of CHO/MAP-EphA1 via intraperitoneal route two days before harvesting spleen cells from mice. We gently executed cell-fusion of P3U1 myeloma cells with the harvested splenocytes from immunized mice using polyethylene glycol 1500 (PEG1500; Roche Diagnostics, Indianapolis, IN, USA) under heated conditions.
Hybridomas were cultured in the RPMI-1640 medium added some supplements as shown above, with additional supplements included hypoxanthine, aminopterin, and thymidine (HAT; Thermo Fisher Scientific, Inc.), 5% BriClone (NICB, Dublin, Ireland), and 5 μg/mL of Plasmocin (InvivoGen) as antibiotics into the medium. On days 6 to 7 after cell-fusion, the hybridoma supernatants were screened by flow cytometry using LN229/MAP-EphA1 and parental LN229 cells. The hybridoma supernatant containing Ea1Mab-30 in serum-free medium was filtered and purified using Ab-Capcher Extra (ProteNova, Kagawa, Japan).
2.4. Flow Cytometry
Cells were harvested using 1 mM EDTA (Nacalai Tesque, Inc.) to avoid enzymatic degradation of cell surface proteins because EphA1 is a trypsin-sensitive protein in our preliminary analysis. Subsequently, cells were gently washed with 0.1% bovine serum albumin in phosphate-buffered saline (PBS) and reacted with primary mAbs for 30 min at 4°C. After that, cells were treated with Alexa Fluor 488-conjugated anti-mouse or rat IgG (dilution rate; 1:1000) following the fluorescence data collection, using the SA3800 Cell Analyzer (Sony Corp.).
2.5. Determination of the Binding Affinity by Flow Cytometry
CHO/EphA1 and 5637 cells were suspended in 100 μL serially diluted of Ea1Mab-30 (20 µg/mL to 0.0012 µg/mL) and A20047D (25 µg/mL to 0.0015 µg/mL), after which Alexa Fluor 488-conjugated anti-mouse or rat IgG (dilution rate; 1:200) was treated. Fluorescence data were subsequently collected, using the BD FACSLyric (BD Biosciences, Franklin Lakes, NJ, USA), following the analyzing of the dissociation constant (KD) by fitting the binding isotherms into the built-in; one-site binding model in GraphPad PRISM 8 (GraphPad Software, Inc., La Jolla, CA, USA).
2.6. Western Blot Analysis
Cells were lysed and boiled in sodium dodecyl sulfate (SDS) sample buffer (Nacalai Tesque, Inc.). Denatured proteins were electrophoresed on 5%–20% polyacrylamide gels (Wako) and transferred onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA). After blocking with 4% non-fat milk (Nacalai Tesque, Inc.), PVDF membranes were reacted with Ea1Mab-30 (1 μg/mL), A20047D (1 μg/mL), an anti-IDH1 mAb (clone RcMab-1, 1 μg/mL), or an anti-PA tag mAb (clone NZ-1, 1 μg/mL), followed by incubation with horseradish peroxidase-conjugated anti-mouse IgG for Ea1Mab-30 (dilution rate; 1:2000; Agilent Technologies Inc.) or anti-rat IgG for A20047D, RcMab-1, and NZ-1 (dilution rate; 1:10000; Merck KGaA). Chemiluminescence signals were visualized using ImmunoStar LD (Wako) and imaged with a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
2.7. Immunohistochemistry
The formalin-fixed paraffin-embedded (FFPE) CHO/EphA1 and CHO-K1 cell blocks were prepared using iPGell (Genostaff Co., Ltd., Tokyo, Japan). The antigen retrieval of paraffin-embedded cell sections was conducted by autoclave in a citrate buffer (pH 6.0; Nichirei Biosciences, Inc., Tokyo, Japan). After blocking using the SuperBlock T20 Blocking Buffer (Thermo Fisher Scientific Inc.), the sections were treated with Ea1Mab-30 (5 μg/mL) and A20047D (5 μg/mL) and then reacted with the Envision+ Kit (Agilent Technologies Inc.) for Ea1Mab-30 or Histofine Simple Stain Mouse MAX PO (Rat) (Nichirei Biosciences, Inc.) for A20047D. Color development was performed using 3,3’-diaminobenzidine tetrahydrochloride (Agilent Technologies Inc.), and counterstaining was carried out using hematoxylin (Merck KGaA).
3. Results
3.1. Development of Anti-EphA1 mAbs Using the CBIS Method
To establish mAbs targeting EphA1, we employed the CBIS method using EphA1-overexpressed cell lines. Anti-EphA1 mAbs-producing hybridoma were screened by flow cytometric analysis (Figure 1). Two female BALB/cAJcl mice were immunized with CHO/MAP-EphA1 (1 × 108 cells/time/mouse) every week via intraperitoneal route for a total of 5 times. Subsequently, splenocytes isolated from immunized mice and P3U1 cells were fused by PEG1500 and fused cells were seeded into 96-well plates. After confirming hybridoma formation, flow cytometric high throughput screening was conducted to select LN229/MAP-EphA1-reactive and parental LN229-nonreactive supernatants of hybridomas. Afterward, we obtained highly LN229/MAP-EphA1-reactive supernatants of the hybridoma. Finally, we established and selected the highly sensitive clone Ea1Mab-30 (mouse IgG1, kappa) by limiting dilution and additional analysis.
3.2. Investigation of Antibody Reactivities Using Flow Cytometry
We checked the reactivity of Ea1Mab-30 against CHO-K1 and CHO/EphA1 cells by flow cytometric analysis. A commercially available anti-EphA1 mAb from BioLegend, A20047D, was used for comparison with Ea1Mab-30. A20047D is currently the only commercially available mAb that can be used for flow cytometry. Results showed that Ea1Mab-30 and A20047D recognized CHO/EphA1 dose-dependently (Figure 2A). Reactivity is almost the same for Ea1Mab-30 and A20047D to CHO/EphA1 (Figure 2A). Neither Ea1Mab-30 nor A20047D reacted with parental CHO-K1 cells (Figure 2B). Thus, Ea1Mab-30 can detect exogenously expressed EphA1 specifically in flow cytometry.
3.3. The Reactivity of Anti-EphA1 mAbs Using Flow Cytometry Against Endogenous EphA1
Next, we confirmed the reactivity of Ea1Mab-30 and A20047D in flow cytometry against 5637 and Caco-2 cells which are endogenously EphA1 expressing cells. The high expression of Eph receptor and ephrin have been reported in bladder cancers [41]. We initially confirmed that EphA1 was endogenously expressed in the bladder cancer cell line, 5637. Furthermore, the expression of EphA1 in colorectal cancer, including Caco-2 cells, has been described in previous reports [30,42]. Results showed that Ea1Mab-30 and A20047D recognized endogenously expressing EphA1 in 5637 (Figure 3A) and Caco-2 (Figure 3B) cancer cells. Ea1Mab-30 showed slightly higher reactivity than A20047D on all concentrations (Figure 3A,B). Neither Ea1Mab-30 nor A20047D reacted with BINDS-38, EphA1-KO 5637 cells, even at a concentration of 10 µg/mL (Figure 3C). Thus, Ea1Mab-30 can detect endogenously expressing EphA1 specifically in flow cytometry.
3.4. Specificity of Ea1Mab-30 to Eph Receptor-Overexpressed CHO-K1 Cells
We have established the cell lines of all Eph receptor-overexpressed CHO-K1 cells, EphA1 to A8, A10, B1 to B4, and B6, respectively. Using the fourteen cell lines, the specificity of Ea1Mab-30 was analyzed. As shown in Figure 4, 10 µg/mL of Ea1Mab-30 potently recognized CHO/EphA1. Ea1Mab-30 never reacted to other Eph receptors. This result complemented the fact that Ea1Mab-30 is an EphA1-specific mAb.
3.5. Calculation of the Binding Affinity of Anti-EphA1 mAbs Using Flow Cytometry
The binding affinity of Ea1Mab-30 and A20047D was assessed with exogenously EphA1-expressed CHO/EphA1 and endogenously EphA1-expressing 5637 using flow cytometry. The results indicated that the KD values of Ea1Mab-30 and A20047D for CHO/EphA1 were 8.9×10-9 M and 8.6×10-9 M, respectively (Figure 5A). The KD values of Ea1Mab-30 and A20047D for 5637 were 1.7×10-9 M and 2.7×10-9 M, respectively (Figure 5B). Although there was no noticeable difference in binding affinity for CHO/EphA1 between Ea1Mab-30 and A20047D, Ea1Mab-30 possessed higher binding affinity for 5637 than A20047D. These results demonstrate that Ea1Mab-30 can recognize EphA1 with high affinity to EphA1 on cells.
3.6. Western Blot Analysis Using Anti-EphA1 mAbs
We investigated whether Ea1Mab-30 can be used for western blot analysis by analyzing LN229, EphA1-overexpressed LN229/EphA1, 5637, and EphA1-KO BINDS-38 cell lysates. The estimated molecular weight of EphA1 protein is 108 kDa. As shown in Figure 6A, Ea1Mab-30 could detect exogenously expressed-EphA1 as the band around 100 to 130 kDa in LN229/EphA1 cell lysates, while no band was detected in parental LN229 cells. Another anti-EphA1 mAb A20047D could not detect any band in LN229/EphA1 cell lysates. Western blot has not been confirmed to apply in the supplier data source from BioLegend. An anti-PA tag mAb NZ-1 was used as a positive control and could also detect a band of the same position around 100 to 130 kDa in LN229/EphA1 cell lysates. An anti-IDH1 mAb (clone RcMab-1) was used for internal control. Furthermore, Ea1Mab-30 could detect endogenously expressing-EphA1 as the band around 100 to 130 kDa in 5637 cell lysates, while no band was detected in BINDS-38, EphA1-KO 5637 cells (Figure 6B). A20047D could not show any band in 5637 cell lysates. These results demonstrate that in western blot analysis, Ea1Mab-30 can detect EphA1 in exogenously and endogenously EphA1-expressing cells.
3.7. Immunohistochemistry Using Anti-EphA1 mAbs
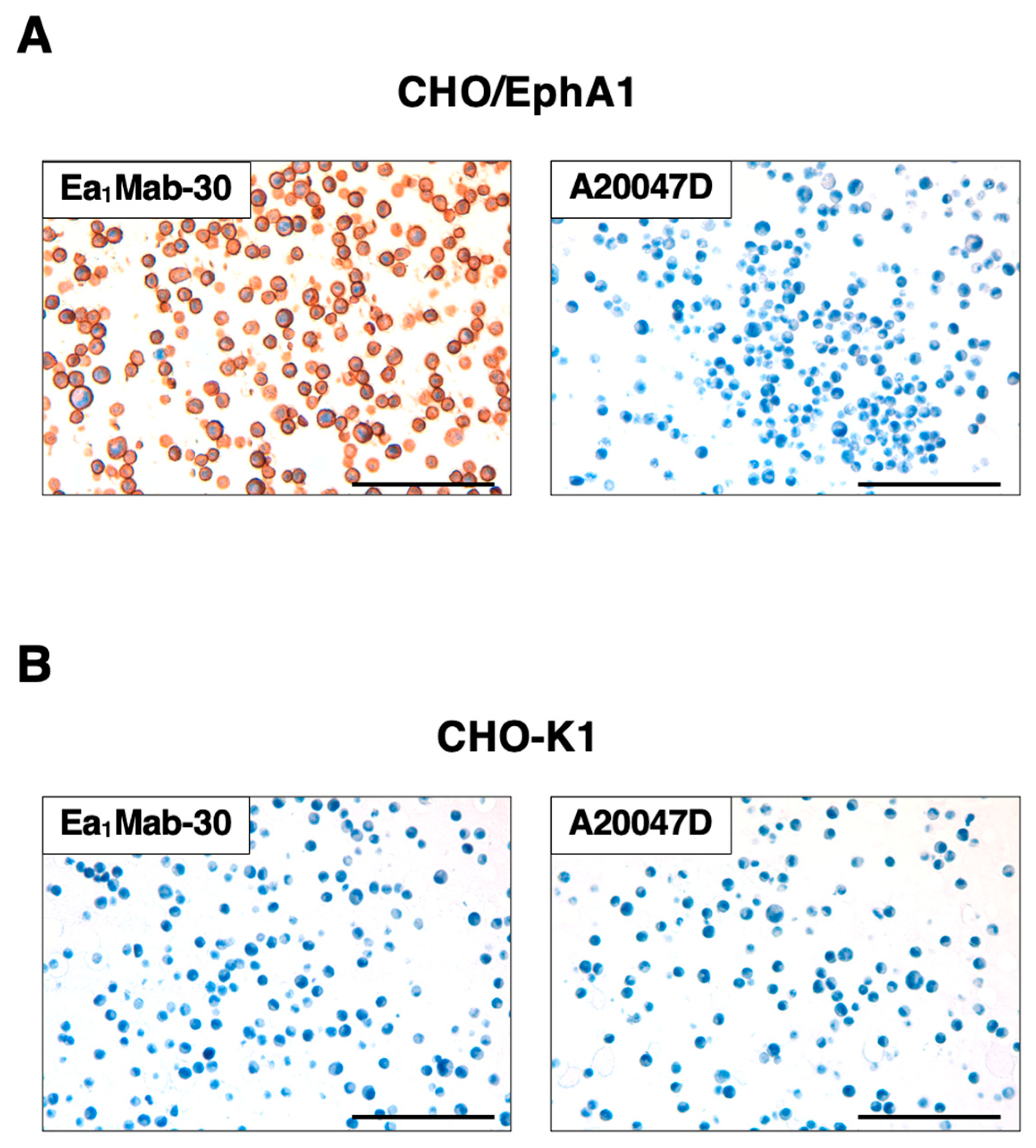
To evaluate whether Ea1Mab-30 can be used for immunohistochemistry (IHC), the FFPE sections of CHO-K1 and CHO/EphA1 were stained with Ea1Mab-30. Apparent membranous staining by Ea1Mab-30 was observed in CHO/EphA1 (Figure 7A). Another anti-EphA1 mAb, A20047D did not stain CHO/EphA1 sections (Figure 7A). Both Ea1Mab-30 and A20047D did not react with the parental CHO-K1 sections (Figure 7B). These results indicate that Ea1Mab-30 applies to IHC for detecting EphA1-positive cells in paraffin-embedded cell samples.
4. Discussion
Currently, no mAbs targeting EphA1 can be used for flow cytometry, western blot, and immunohistochemistry. We successfully established a novel anti-human EphA1 mAb, Ea1Mab-30, which is applicable for various experiments for the first time. Ea1Mab-30 clearly and specifically detected EphA1 in flow cytometry (Figure 2, Figure 3 and Figure 4), western blot (Figure 6), and immunohistochemistry (Figure 7). Furthermore, Ea1Mab-30 recognized EphA1 with higher affinity than A20047D, the only commercially available mAb currently suitable for flow cytometry (Figure 5). Ea1Mab-30 could contribute to elucidating the EphA1-related cellular functions and diagnosis of disorders such as AD and tumors. Overexpression of EphA1 is observed in various cancers [23,33,43], EphA1 can be a target for cancer treatments. To evaluate the antitumor efficacy of Ea1Mab-30, it will be converted to a mouse IgG2a version to elevate antibody-dependent cellular cytotoxicity in the next step [44,45].
Since overexpression of Eph receptors has been observed regardless of cancer type, they have been widely studied in cancer research and emerged as therapeutic targets [46,47,48,49]. However, specific drugs targeting Eph receptors or ephrin ligands have yet to be approved. However, some clinical trials have been conducted such as DS-8895a (a humanized anti-EphA2 afucosylated mAb) against advanced EphA2-expressing cancer and Ifabotuzumab (an anti-EphA3 mAb) against advanced hematologic malignancies [48,50]. EphA1-targeting therapeutic challenge has not been confirmed yet. One reason could be that EphA1 has a dual role in promoting and suppressing cancer.
AD is the most common chronic neurodegenerative disease characterized by cognitive decline and behavioral change. The most well-known neuropathological hallmarks of AD are aggregation of extracellular amyloid-β peptides into amyloid plaques, and cytotoxic intracellular neurofibrillary tau tangles [51,52]. Large number of GWAS for AD have been conducted and suggested numerous loci as the risk of AD including EphA1 [13,14,18,53]. Some SNPs near EphA1 gene, rs11767557 and rs11771145, have been demonstrated in GWAS to decrease the risk of AD [13,18,54]. Several EphA1 missense mutations are considered to increase the risk of AD. P460L mutation in EphA1 is associated with AD by dysregulating EphA1 activity [55]. P460L variant triggers disruptions of EphA1, including reducing tyrosine phosphorylation, cell surface expression, and decreasing T-cell recruitment to the brain by affecting brain endothelial barrier [55]. In Drosophila models, P460L mutation causes some AD-like symptoms such as hyperexcitability, less sleep, a more vigorous circadian rhythm and overactivity of clock neuron in the absence of memory loss [56]. P460 locates in the second fibronectin type III repeat (FNIII-2) of EphA1 adjacent to the cell membrane and highly conserved across species [3,55,57]. Thus, this missense mutation might be necessary for neuropathological research in animal models. EphA-ephrinA axis allows forward and reverse bidirectional signaling in cell-cell contact, followed by playing a role in synaptic formation and plasticity, axonal guidance, and brain development [5,7,58]. EphA1 is an attractive molecule for analyzing AD pathology, but there are few reports of post-mortem brain-tissue analysis. Moreover, EphA1 promotes the pathogenesis of Parkinson’s disease, the second most common neurodegenerative disease, through CXCL12-CXCR4 axis [59]. The developed Ea1Mab-30 will be helpful for the analysis of the diseases.
Although EphA1 is characterized as the negative risk factor of AD, its structural alteration with missense mutations may provoke the AD [60]. As with cancer, to clarify the critical mutation of AD-related proteins is essential to make an early diagnosis and treatment. Furthermore, N414 and N478 in EphA1 are glycosylated [17]. Although these N-glycosylation are impaired by P460L mutation, epigenetic modification might contribute to normal EphA1 functions. We have previously identified cancer specific aberrant glycosylation of podoplanin [61] and misfolding of HER2 [44] through the establishment of mAbs targeting these characteristic epitopes. Research into disease-specific epitopes for EphA1, such as those for cancer and AD, is expected to progress. We plan to obtain mutated-EphA1 specific mAbs to clarify the relation between EphA1 and diseases.
Author Contributions
Tomohiro Tanaka: Investigation, Funding acquisition, Writing: original draft Hiroyuki Suzuki: Investigation, Funding acquisition, Writing – review and editing. Guanjie Li: Investigation. Shiori Fujisawa: Investigation. Mika K. Kaneko: Conceptualization, Funding acquisition. Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing – review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP24am0521010 (to Y.K.), JP24ama121008 (to Y.K.), JP24ama221339 (to Y.K.), JP24bm1123027 (to Y.K.), and JP24ck0106730 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 22K06995 (to H.S), 24K18268 (to T.T), and 22K07224 (to Y.K.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2019NiA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Wang, H.U.; Chen, Z.F.; Anderson, D.J. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell 1998, 93, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Sawamiphak, S.; Seidel, S.; Essmann, C.L.; Wilkinson, G.A.; Pitulescu, M.E.; Acker, T.; Acker-Palmer, A. Ephrin-B2 regulates VEGFR2 function in developmental and tumour angiogenesis. Nature 2010, 465, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H.; Maru, Y.; Hagiwara, K.; Nishida, J.; Takaku, F. A novel putative tyrosine kinase receptor encoded by the eph gene. Science 1987, 238, 1717–1720. [Google Scholar] [CrossRef] [PubMed]
- Coulthard, M.G.; Lickliter, J.D.; Subanesan, N.; Chen, K.; Webb, G.C.; Lowry, A.J.; Koblar, S.; Bottema, C.D.; Boyd, A.W. Characterization of the Epha1 receptor tyrosine kinase: expression in epithelial tissues. Growth Factors 2001, 18, 303–317. [Google Scholar] [CrossRef]
- Yamazaki, T.; Masuda, J.; Omori, T.; Usui, R.; Akiyama, H.; Maru, Y. EphA1 interacts with integrin-linked kinase and regulates cell morphology and motility. J Cell Sci 2009, 122, 243–255. [Google Scholar] [CrossRef]
- Martínez, A.; Otal, R.; Sieber, B.A.; Ibáñez, C.; Soriano, E. Disruption of ephrin-A/EphA binding alters synaptogenesis and neural connectivity in the hippocampus. Neuroscience 2005, 135, 451–461. [Google Scholar] [CrossRef]
- Lai, K.O.; Ip, N.Y. Synapse development and plasticity: roles of ephrin/Eph receptor signaling. Curr Opin Neurobiol 2009, 19, 275–283. [Google Scholar] [CrossRef]
- Kosinski, C.; Li, V.S.; Chan, A.S.; Zhang, J.; Ho, C.; Tsui, W.Y.; Chan, T.L.; Mifflin, R.C.; Powell, D.W.; Yuen, S.T.; et al. Gene expression patterns of human colon tops and basal crypts and BMP antagonists as intestinal stem cell niche factors. Proc Natl Acad Sci U S A 2007, 104, 15418–15423. [Google Scholar] [CrossRef]
- Sakamoto, A.; Sugamoto, Y.; Tokunaga, Y.; Yoshimuta, T.; Hayashi, K.; Konno, T.; Kawashiri, M.A.; Takeda, Y.; Yamagishi, M. Expression profiling of the ephrin (EFN) and Eph receptor (EPH) family of genes in atherosclerosis-related human cells. J Int Med Res 2011, 39, 522–527. [Google Scholar] [CrossRef]
- Duffy, S.L.; Coulthard, M.G.; Spanevello, M.D.; Herath, N.I.; Yeadon, T.M.; McCarron, J.K.; Carter, J.C.; Tonks, I.D.; Kay, G.F.; Phillips, G.E.; et al. Generation and characterization of EphA1 receptor tyrosine kinase reporter knockout mice. Genesis 2008, 46, 553–561. [Google Scholar] [CrossRef]
- Lambert, J.C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; DeStafano, A.L.; Bis, J.C.; Beecham, G.W.; Grenier-Boley, B.; et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer's disease. Nat Genet 2013, 45, 1452–1458. [Google Scholar] [CrossRef]
- Talebi, M.; Delpak, A.; Khalaj-Kondori, M.; Sadigh-Eteghad, S.; Talebi, M.; Mehdizadeh, E.; Majdi, A. ABCA7 and EphA1 Genes Polymorphisms in Late-Onset Alzheimer's Disease. J Mol Neurosci 2020, 70, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Naj, A.C.; Jun, G.; Beecham, G.W.; Wang, L.S.; Vardarajan, B.N.; Buros, J.; Gallins, P.J.; Buxbaum, J.D.; Jarvik, G.P.; Crane, P.K.; et al. Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer's disease. Nat Genet 2011, 43, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Karch, C.M.; Goate, A.M. Alzheimer's disease risk genes and mechanisms of disease pathogenesis. Biol Psychiatry 2015, 77, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Sadigh-Eteghad, S.; Sabermarouf, B.; Majdi, A.; Talebi, M.; Farhoudi, M.; Mahmoudi, J. Amyloid-beta: a crucial factor in Alzheimer's disease. Med Princ Pract 2015, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer's disease: progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Matsumoto, M.; Gomez-Soler, M.; Lombardi, S.; Lechtenberg, B.C.; Pasquale, E.B. Missense mutations of the ephrin receptor EPHA1 associated with Alzheimer's disease disrupt receptor signaling functions. J Biol Chem 2024, 301, 108099. [Google Scholar] [CrossRef]
- Hollingworth, P.; Harold, D.; Sims, R.; Gerrish, A.; Lambert, J.C.; Carrasquillo, M.M.; Abraham, R.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; et al. Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer's disease. Nat Genet 2011, 43, 429–435. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, Y.; Wang, L.; Xu, J.; Chen, X.; Bao, Y.; Hu, Y.; Jin, S.; Tian, R.; Bai, W.; et al. Alzheimer's Disease rs11767557 Variant Regulates EPHA1 Gene Expression Specifically in Human Whole Blood. J Alzheimers Dis 2018, 61, 1077–1088. [Google Scholar] [CrossRef]
- Karch, C.M.; Jeng, A.T.; Nowotny, P.; Cady, J.; Cruchaga, C.; Goate, A.M. Expression of novel Alzheimer's disease risk genes in control and Alzheimer's disease brains. PLoS One 2012, 7, e50976. [Google Scholar] [CrossRef]
- Wang, J.; Ma, J.; Dong, Y.; Shen, Z.; Ma, H.; Wang, X.; Shi, S.; Wu, J.; Lu, G.; Peng, L.; et al. High expression of EphA1 in esophageal squamous cell carcinoma is associated with lymph node metastasis and advanced disease. Apmis 2013, 121, 30–37. [Google Scholar] [CrossRef]
- Wang, J.; Dong, Y.; Wang, X.; Ma, H.; Sheng, Z.; Li, G.; Lu, G.; Sugimura, H.; Zhou, X. Expression of EphA1 in gastric carcinomas is associated with metastasis and survival. Oncol Rep 2010, 24, 1577–1584. [Google Scholar] [CrossRef]
- Wang, Y.C.; Dai, Y.; Xu, G.L.; Yu, W.; Quan, R.L.; Zhao, Y.J. Association Between EphA1 and Tumor Microenvironment in Gastric Carcinoma and its Clinical Significance. Med Sci Monit 2020, 26, e923409. [Google Scholar] [CrossRef]
- Robinson, D.; He, F.; Pretlow, T.; Kung, H.J. A tyrosine kinase profile of prostate carcinoma. Proc Natl Acad Sci U S A 1996, 93, 5958–5962. [Google Scholar] [CrossRef]
- Peng, L.; Wang, H.; Dong, Y.; Ma, J.; Wen, J.; Wu, J.; Wang, X.; Zhou, X.; Wang, J. Increased expression of EphA1 protein in prostate cancers correlates with high Gleason score. Int J Clin Exp Pathol 2013, 6, 1854–1860. [Google Scholar]
- Koh, H.M.; Jang, B.G.; Lee, D.H.; Hyun, C.L.; Kim, D.C. The association between ephrin receptor-A1 expression and survival in patients with cancer: a meta-analysis. Transl Cancer Res 2022, 11, 1587–1594. [Google Scholar] [CrossRef]
- Nagare, R.P.; Sneha, S.; Krishnapriya, S.; Ramachandran, B.; Murhekar, K.; Vasudevan, S.; Shabna, A.; Ganesan, T.S. ALDH1A1+ ovarian cancer stem cells co-expressing surface markers CD24, EPHA1 and CD9 form tumours in vivo. Exp Cell Res 2020, 392, 112009. [Google Scholar] [CrossRef]
- Ieguchi, K.; Tomita, T.; Omori, T.; Komatsu, A.; Deguchi, A.; Masuda, J.; Duffy, S.L.; Coulthard, M.G.; Boyd, A.; Maru, Y. ADAM12-cleaved ephrin-A1 contributes to lung metastasis. Oncogene 2014, 33, 2179–2190. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, J.; Sheng, Z.; Li, G.; Ma, H.; Wang, X.; Zhang, R.; Lu, G.; Hu, Q.; Sugimura, H.; et al. Downregulation of EphA1 in colorectal carcinomas correlates with invasion and metastasis. Mod Pathol 2009, 22, 151–160. [Google Scholar] [CrossRef]
- Herath, N.I.; Doecke, J.; Spanevello, M.D.; Leggett, B.A.; Boyd, A.W. Epigenetic silencing of EphA1 expression in colorectal cancer is correlated with poor survival. Br J Cancer 2009, 100, 1095–1102. [Google Scholar] [CrossRef]
- Hafner, C.; Becker, B.; Landthaler, M.; Vogt, T. Expression profile of Eph receptors and ephrin ligands in human skin and downregulation of EphA1 in nonmelanoma skin cancer. Mod Pathol 2006, 19, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zou, Y.; Wan, L.; Lu, M.; Liu, Y.; Huang, G.; Wang, J.; Xi, Q. Decreased Eph receptor-A1 expression is related to grade in ovarian serous carcinoma. Mol Med Rep 2018, 17, 5409–5415. [Google Scholar] [CrossRef] [PubMed]
- Gajdzis, M.; Theocharis, S.; Gajdzis, P.; Cassoux, N.; Gardrat, S.; Donizy, P.; Klijanienko, J.; Kaczmarek, R. Ephrin Receptors (Eph): EphA1, EphA5, and EphA7 Expression in Uveal Melanoma-Associations with Clinical Parameters and Patient Survival. Life (Basel) 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Satofuka H, S.H., Tanaka T, Li G, Kaneko MK, Kato Y. An Anti-Human EphA2 Monoclonal Antibody Ea2Mab-7 Shows High Sensitivity for Flow Cytometry, Western Blot, and Immunohistochemical Analyses. Preprint 2024. [CrossRef]
- Ubukata, R.; Suzuki, H.; Hirose, M.; Satofuka, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Establishment of a highly sensitive and specific anti-EphB2 monoclonal antibody (Eb2Mab-12) for flow cytometry. MI 2025. [Google Scholar] [CrossRef]
- Nanamiya, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of an Anti-EphB4 Monoclonal Antibody for Multiple Applications Against Breast Cancers. Monoclon Antib Immunodiagn Immunother 2023, 42, 166–177. [Google Scholar] [CrossRef]
- Tanaka, T.; Kaneko, Y.; Yamamoto, H.; Li, G.; Fujisawa, S.; Satofuka, H.; Shinoda, K.; Nakamura, T.; Kaneko, M.K.; Suzuki, H.; et al. Development of a novel anti-erythropoietin-producing hepatocellular receptor B6 monoclonal antibody Eb6Mab-3 for flow cytometry. Biochemistry and Biophysics Reports 2025, 41, 101960. [Google Scholar] [CrossRef]
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon Antib Immunodiagn Immunother 2016, 35, 293–299. [Google Scholar] [CrossRef]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; Nogi, T.; Kato, Y.; Takagi, J. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014, 95, 240–247. [Google Scholar] [CrossRef]
- Ikota, H.; Nobusawa, S.; Arai, H.; Kato, Y.; Ishizawa, K.; Hirose, T.; Yokoo, H. Evaluation of IDH1 status in diffusely infiltrating gliomas by immunohistochemistry using anti-mutant and wild type IDH1 antibodies. Brain Tumor Pathol 2015, 32, 237–244. [Google Scholar] [CrossRef]
- Chu, M.; Zhang, C. Inhibition of angiogenesis by leflunomide via targeting the soluble ephrin-A1/EphA2 system in bladder cancer. Sci Rep 2018, 8, 1539. [Google Scholar] [CrossRef]
- Herath, N.I.; Boyd, A.W. The role of Eph receptors and ephrin ligands in colorectal cancer. Int J Cancer 2010, 126, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Y.; Cao, G.; Zhang, X.; Xu, H.; Xu, H.; Wang, J. Expression of the EphA1 protein is associated with Fuhrman nuclear grade in clear cell renal cell carcinomas. Int J Clin Exp Pathol 2015, 8, 6821–6827. [Google Scholar] [PubMed]
- Arimori, T.; Mihara, E.; Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Takagi, J.; Kato, Y. Locally misfolded HER2 expressed on cancer cells is a promising target for development of cancer-specific antibodies. Structure 2024, 32, 536–549.e535. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. A Cancer-Specific Anti-Podoplanin Monoclonal Antibody, PMab-117-mG(2a) Exerts Antitumor Activities in Human Tumor Xenograft Models. Cells 2024, 13. [Google Scholar] [CrossRef]
- Liang, Z.; Wang, X.; Dong, K.; Li, X.; Qin, C.; Zhou, H. Expression Pattern and Prognostic Value of EPHA/EFNA in Breast Cancer by Bioinformatics Analysis: Revealing Its Importance in Chemotherapy. Biomed Res Int 2021, 2021, 5575704. [Google Scholar] [CrossRef]
- Phan, N.N.; Liu, S.; Wang, C.Y.; Hsu, H.P.; Lai, M.D.; Li, C.Y.; Chen, C.F.; Chiao, C.C.; Yen, M.C.; Sun, Z.; et al. Overexpressed gene signature of EPH receptor A/B family in cancer patients-comprehensive analyses from the public high-throughput database. Int J Clin Exp Pathol 2020, 13, 1220–1242. [Google Scholar]
- Pasquale, E.B. Eph receptors and ephrins in cancer progression. Nat Rev Cancer 2024, 24, 5–27. [Google Scholar] [CrossRef]
- McCarron, J.K.; Stringer, B.W.; Day, B.W.; Boyd, A.W. Ephrin expression and function in cancer. Future Oncol 2010, 6, 165–176. [Google Scholar] [CrossRef]
- Janes, P.W.; Vail, M.E.; Gan, H.K.; Scott, A.M. Antibody Targeting of Eph Receptors in Cancer. Pharmaceuticals (Basel) 2020, 13. [Google Scholar] [CrossRef]
- Masters, C.L.; Bateman, R.; Blennow, K.; Rowe, C.C.; Sperling, R.A.; Cummings, J.L. Alzheimer's disease. Nat Rev Dis Primers 2015, 1, 15056. [Google Scholar] [CrossRef]
- Bellenguez, C.; Grenier-Boley, B.; Lambert, J.C. Genetics of Alzheimer's disease: where we are, and where we are going. Curr Opin Neurobiol 2020, 61, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Laws, S.M.; Miles, L.A.; Wiley, J.S.; Huang, X.; Masters, C.L.; Gu, B.J. Genomics of Alzheimer's disease implicates the innate and adaptive immune systems. Cell Mol Life Sci 2021, 78, 7397–7426. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.C.; Heath, S.; Even, G.; Campion, D.; Sleegers, K.; Hiltunen, M.; Combarros, O.; Zelenika, D.; Bullido, M.J.; Tavernier, B.; et al. Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer's disease. Nat Genet 2009, 41, 1094–1099. [Google Scholar] [CrossRef] [PubMed]
- Owens, H.A.; Thorburn, L.E.; Walsby, E.; Moon, O.R.; Rizkallah, P.; Sherwani, S.; Tinsley, C.L.; Rogers, L.; Cerutti, C.; Ridley, A.J.; et al. Alzheimer's disease-associated P460L variant of EphA1 dysregulates receptor activity and blood-brain barrier function. Alzheimers Dement 2024, 20, 2016–2033. [Google Scholar] [CrossRef]
- Buhl, E.; Kim, Y.A.; Parsons, T.; Zhu, B.; Santa-Maria, I.; Lefort, R.; Hodge, J.J.L. Effects of Eph/ephrin signalling and human Alzheimer's disease-associated EphA1 on Drosophila behaviour and neurophysiology. Neurobiol Dis 2022, 170, 105752. [Google Scholar] [CrossRef]
- Sahoo, A.R.; Buck, M. Structural and Functional Insights into the Transmembrane Domain Association of Eph Receptors. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, A.K.; Ip, N.Y. Bidirectional signaling of ErbB and Eph receptors at synapses. Neuron Glia Biol 2008, 4, 211–221. [Google Scholar] [CrossRef]
- Ma, J.; Wang, Z.; Chen, S.; Sun, W.; Gu, Q.; Li, D.; Zheng, J.; Yang, H.; Li, X. EphA1 Activation Induces Neuropathological Changes in a Mouse Model of Parkinson's Disease Through the CXCL12/CXCR4 Signaling Pathway. Mol Neurobiol 2021, 58, 913–925. [Google Scholar] [CrossRef]
- Yao, M.; Miller, G.W.; Vardarajan, B.N.; Baccarelli, A.A.; Guo, Z.; Liu, Z. Deciphering proteins in Alzheimer's disease: A new Mendelian randomization method integrated with AlphaFold3 for 3D structure prediction. Cell Genom 2024, 4, 100700. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneko, M.K. A cancer-specific monoclonal antibody recognizes the aberrantly glycosylated podoplanin. Sci Rep 2014, 4, 5924. [Google Scholar] [CrossRef]
Figure 1.
A schematic depiction of anti-EphA1 mAbs development by CBIS method. The simplified procedure of antibody development using the CBIS method. (A) CHO/MAP-EphA1 cells were immunized into two female mice via the intraperitoneal route. (B) The spleen cells isolated from antigen-immunized mice were fused with mouse myeloma cells, P3U1, by PEG1500. (C) The culture supernatants of hybridoma were screened by flow cytometric analysis using LN229 and LN229/MAP-EphA1 to select EphA1-specific mAb-producing hybridomas. (D) Single hybridoma clones were obtained by limiting dilution, followed by additional screening. Finally, Ea1Mab-30 (mouse IgG1, kappa) was established.
Figure 1.
A schematic depiction of anti-EphA1 mAbs development by CBIS method. The simplified procedure of antibody development using the CBIS method. (A) CHO/MAP-EphA1 cells were immunized into two female mice via the intraperitoneal route. (B) The spleen cells isolated from antigen-immunized mice were fused with mouse myeloma cells, P3U1, by PEG1500. (C) The culture supernatants of hybridoma were screened by flow cytometric analysis using LN229 and LN229/MAP-EphA1 to select EphA1-specific mAb-producing hybridomas. (D) Single hybridoma clones were obtained by limiting dilution, followed by additional screening. Finally, Ea1Mab-30 (mouse IgG1, kappa) was established.
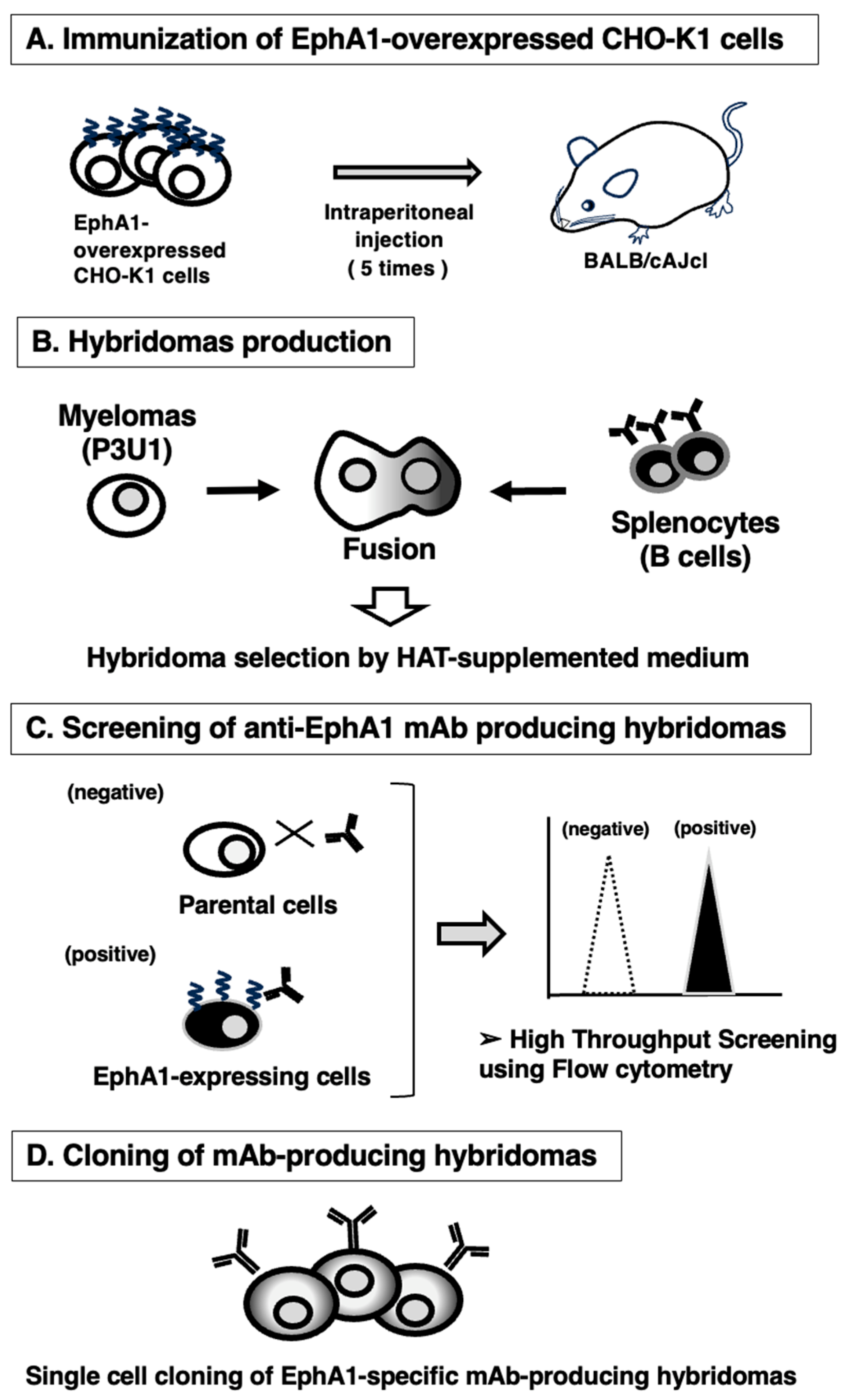
Figure 2.
Flow cytometric analysis of anti-EphA1 mAbs against exogenously expressed EphA1. CHO/EphA1 (A) and CHO-K1 (B) cells were treated with 0.01–10 µg/mL of Ea1Mab-30 or A20047D (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse or rat IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
Figure 2.
Flow cytometric analysis of anti-EphA1 mAbs against exogenously expressed EphA1. CHO/EphA1 (A) and CHO-K1 (B) cells were treated with 0.01–10 µg/mL of Ea1Mab-30 or A20047D (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse or rat IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
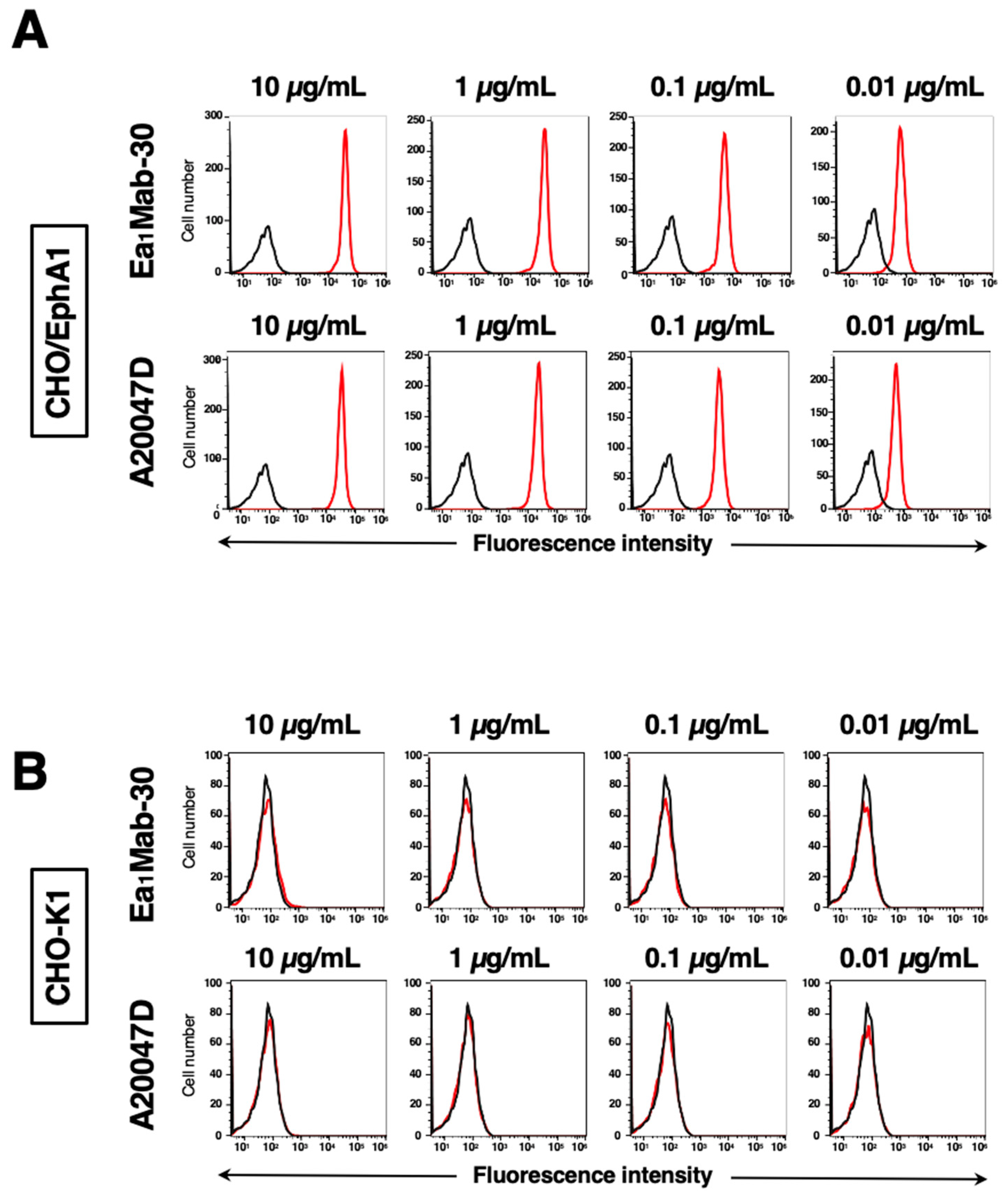
Figure 3.
Flow cytometric analysis of anti-EphA1 mAbs against endogenously expressing EphA1 in cancer cells. 5637 (A), Caco-2 (B), and BINDS-38 (C) cells were treated with 0.01–10 µg/mL of Ea1Mab-30 or A20047D (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse or rat IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
Figure 3.
Flow cytometric analysis of anti-EphA1 mAbs against endogenously expressing EphA1 in cancer cells. 5637 (A), Caco-2 (B), and BINDS-38 (C) cells were treated with 0.01–10 µg/mL of Ea1Mab-30 or A20047D (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse or rat IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
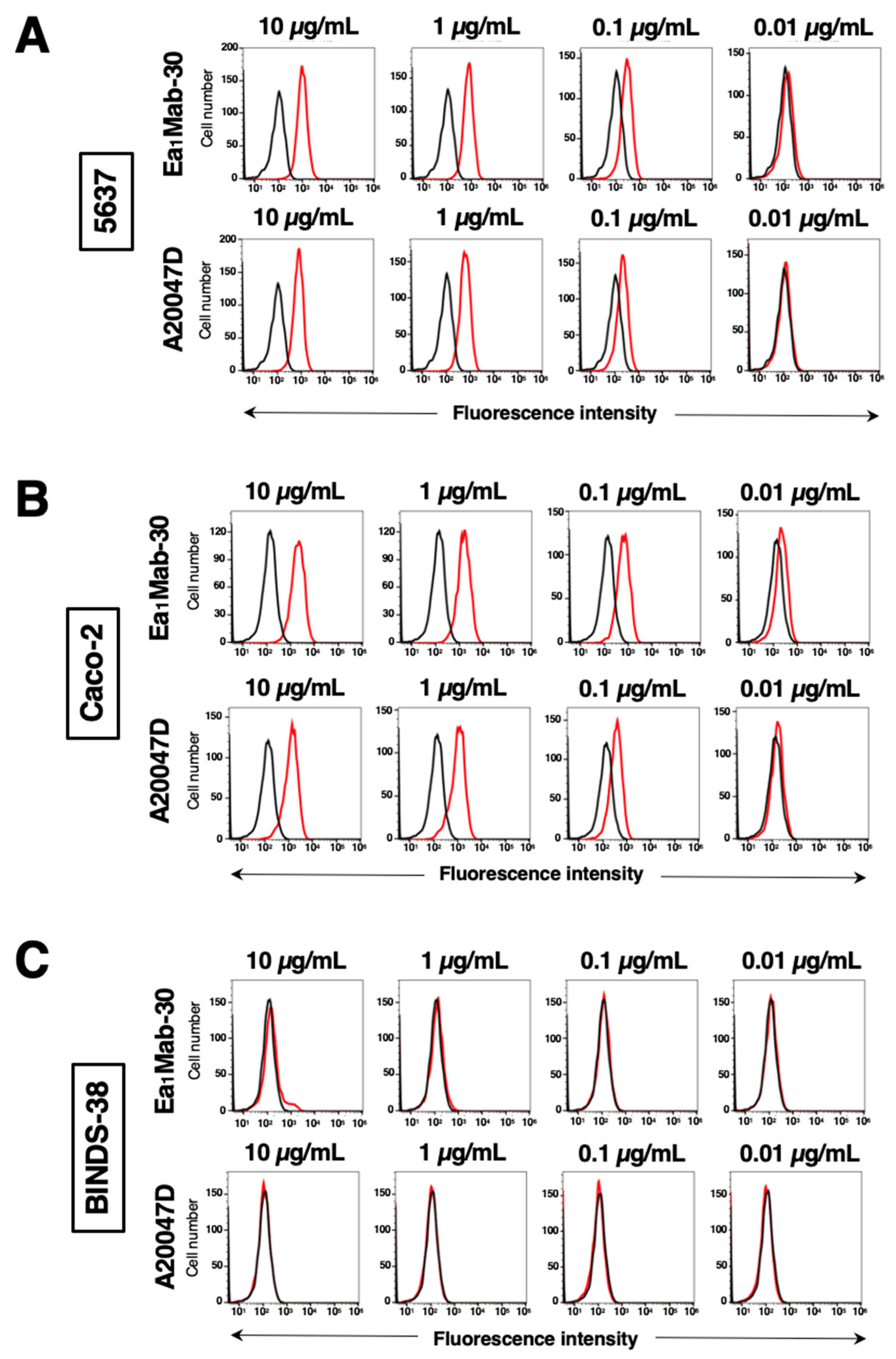
Figure 4.
Cross reactivity of Ea1Mab-30 in Eph receptor-expressed CHO-K1 cells. CHO-K1 cells, which overexpressed each of the fourteen Eph receptors, were treated with 10 µg/mL of Ea1Mab-30 (red line) or control blocking buffer (black line) followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
Figure 4.
Cross reactivity of Ea1Mab-30 in Eph receptor-expressed CHO-K1 cells. CHO-K1 cells, which overexpressed each of the fourteen Eph receptors, were treated with 10 µg/mL of Ea1Mab-30 (red line) or control blocking buffer (black line) followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
Figure 5.
Evaluation of the KD values of anti-EphA1 mAbs. (A) CHO/EphA1 cells were suspended in 100 µL of serially diluted 20 µg/mL to 0.0012 µg/mL of Ea1Mab-30 and 20 µg/mL to 0.0012 µg/mL of A20047D. (B) 5637 cells were suspended in 100 µL of serially diluted 25 µg/mL to 0.0015 µg/mL of Ea1Mab-30 and 25 µg/mL to 0.0015 µg/mL of A20047D. After treatments of primary mAbs, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG (for Ea1Mab-30) or anti-rat IgG (for A20047D). Subsequently, the geometric mean values from fluorescence data were collected using the BD FACSLyric, following the calculation of the KD by GraphPad PRISM 8 software.
Figure 5.
Evaluation of the KD values of anti-EphA1 mAbs. (A) CHO/EphA1 cells were suspended in 100 µL of serially diluted 20 µg/mL to 0.0012 µg/mL of Ea1Mab-30 and 20 µg/mL to 0.0012 µg/mL of A20047D. (B) 5637 cells were suspended in 100 µL of serially diluted 25 µg/mL to 0.0015 µg/mL of Ea1Mab-30 and 25 µg/mL to 0.0015 µg/mL of A20047D. After treatments of primary mAbs, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG (for Ea1Mab-30) or anti-rat IgG (for A20047D). Subsequently, the geometric mean values from fluorescence data were collected using the BD FACSLyric, following the calculation of the KD by GraphPad PRISM 8 software.
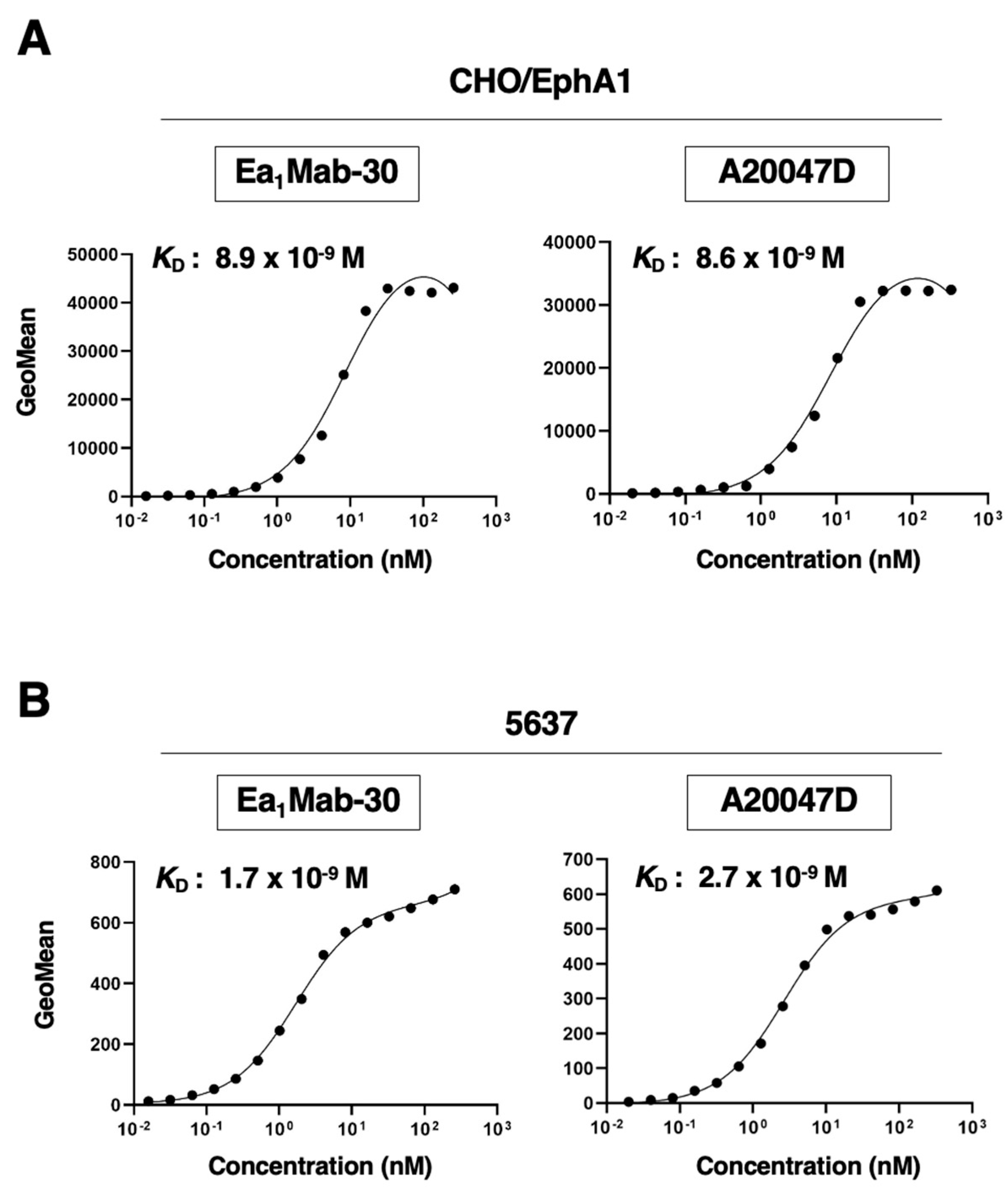
Figure 6.
Detection of EphA1 protein using Ea1Mab-30 by western blot analysis. Cell lysates (10 μg/lane) of LN229, LN229/EphA1 (A), 5637, and BINDS-38 (B) were electrophoresed on polyacrylamide gels and transferred onto PVDF membranes. The membranes were incubated with Ea1Mab-30 (1 μg/mL), A20047D (1 μg/mL), an anti-PA tag mAb (clone NZ-1, 1 μg/mL), and RcMab-1 (1 μg/mL) and subsequently with horseradish peroxidase-conjugated anti-mouse or anti-rat immunoglobulins.
Figure 6.
Detection of EphA1 protein using Ea1Mab-30 by western blot analysis. Cell lysates (10 μg/lane) of LN229, LN229/EphA1 (A), 5637, and BINDS-38 (B) were electrophoresed on polyacrylamide gels and transferred onto PVDF membranes. The membranes were incubated with Ea1Mab-30 (1 μg/mL), A20047D (1 μg/mL), an anti-PA tag mAb (clone NZ-1, 1 μg/mL), and RcMab-1 (1 μg/mL) and subsequently with horseradish peroxidase-conjugated anti-mouse or anti-rat immunoglobulins.
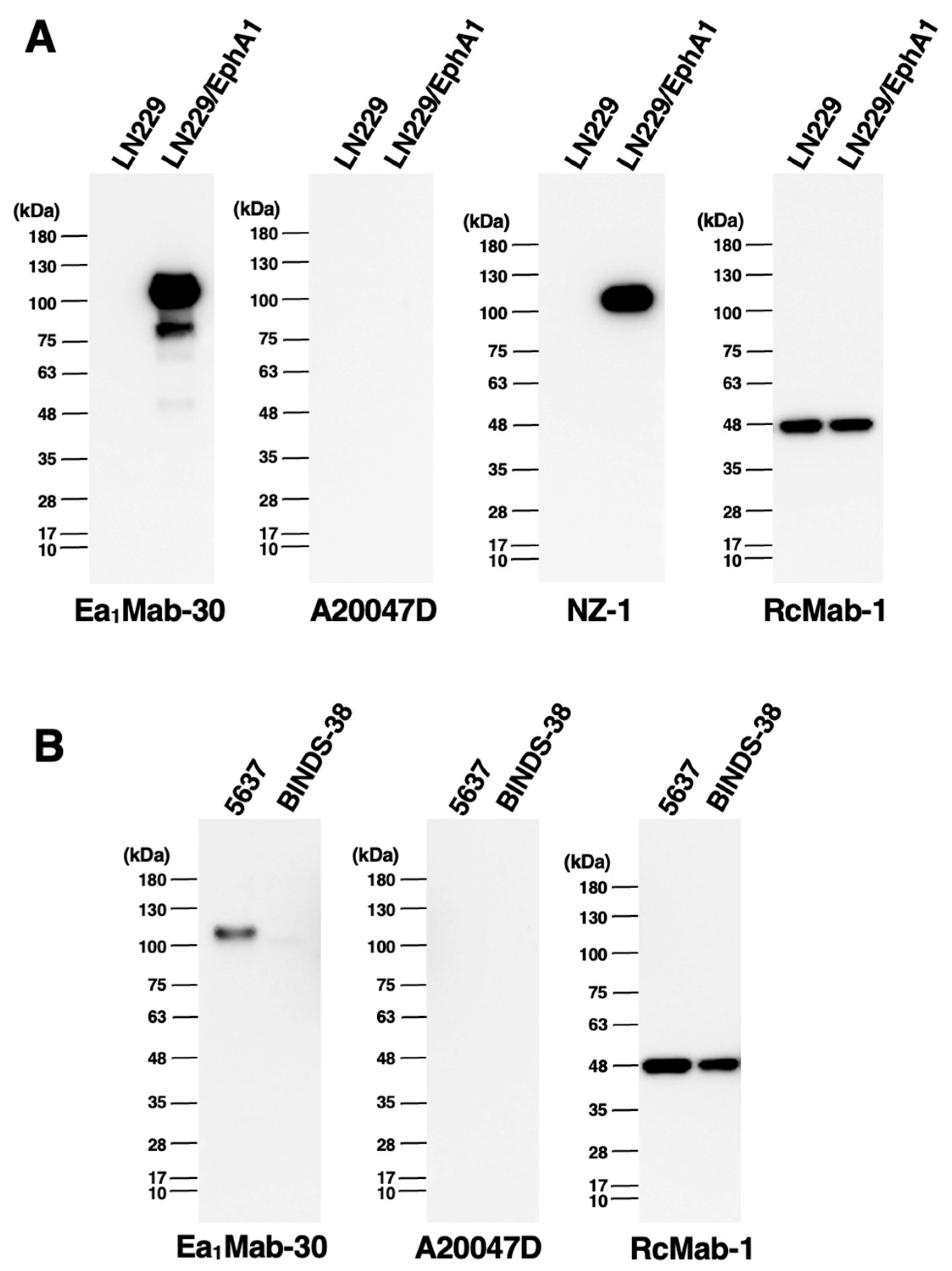
Figure 7.
Immunohistochemical staining of paraffin-embedded sections of CHO/EphA1 and CHO-K1. The sections of CHO/EphA1 (A) and CHO-K1 (B) cells were treated with Ea1Mab-30 (5 μg/mL) or A20047D (5 μg/mL), followed by that with the Envision + Kit (for Ea1Mab-30) or Histofine Simple Stain Mouse MAX PO (Rat) (for A20047D). Color was developed using DAB, and counterstaining was performed using hematoxylin. Scale bar = 100 μm.
Figure 7.
Immunohistochemical staining of paraffin-embedded sections of CHO/EphA1 and CHO-K1. The sections of CHO/EphA1 (A) and CHO-K1 (B) cells were treated with Ea1Mab-30 (5 μg/mL) or A20047D (5 μg/mL), followed by that with the Envision + Kit (for Ea1Mab-30) or Histofine Simple Stain Mouse MAX PO (Rat) (for A20047D). Color was developed using DAB, and counterstaining was performed using hematoxylin. Scale bar = 100 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



