
Submitted:
05 April 2025
Posted:
07 April 2025
You are already at the latest version
Abstract
Erythropoietin-producing hepatocellular (Eph) receptor A3 (EphA3) is a member of the Eph receptor family, which binds to its respective ligands, ephrins. These interactions are essential for normal development and tissue homeostasis. Dysregulation of EphA3 has been reported to be associated with human hematopoietic malignancies, making it a promising target for therapy and diagnosis. Due to the high similarity of the extracellular domain among Eph receptors (more than 33% amino acid identity), generating highly specific monoclonal antibodies (mAbs) is crucial. We developed anti-human EphA3 mAbs in this study using the Cell-Based Immunization and Screening (CBIS) method. Among them, the clone Ea3Mab-20 (IgG1, kappa) exhibited high affinity and specificity in flow cytometry. The dissociation constant values of Ea3Mab-20 for CHO/EphA3 and Jurkat cells were determined to be 9.0 ± 0.3 × 10−9 M and 1.4 ± 0.1 × 10−9 M, respectively. Ea3Mab-20 showed no cross-reactivity with other Eph receptors in flow cytometry. Furthermore, Ea3Mab-20 demonstrated the suitability for detecting formalin-fixed paraffin-embedded cell samples in immunohistochemistry. Therefore, Ea3Mab-20 is valuable mAb for basic research and is expected to contribute to the clinical application of mAb for cancer therapy and diagnosis.
Keywords:
EphA3
; monoclonal antibody
; Cell-Based Immunization and Screening
; flow cytometry
; immunohistochemistry
1. Introduction
Erythropoietin-producing hepatocellular (Eph) receptors are a family of receptor tyrosine kinases with a single transmembrane domain, classified into A and B categories based on their extracellular domains [1,2,3,4,5,6]. The extracellular domains of Eph receptors share a highly similar architecture, consisting of a ligand binding domain, a cysteine-rich region with Sushi and epidermal growth factor-like domains, and two fibronectin type III domains in tandem [1,2,3,7], with more than 33% amino acid identity [8]. Eph receptors interact with their membrane-bound ephrin ligands, with each receptor having preferred ephrin ligands [2]. The mammalian Eph system comprises 14 receptor tyrosine kinases (nine EphA and five EphB receptors such as EphA1 to EphA8, EphA10, EphB1 to EphB4, and EphB6) and eight cell surface-anchored ephrin ligands (five glycosylphosphatidylinositol-linked ephrin-As such as ephrin A1 to A5 and three transmembrane ephrin-Bs such as ephrin B1 to B3) [4,9]. These interactions are essential for various normal cellular processes during development and serve as key mediators of adult tissue homeostasis [7,10,11,12].
The expression of Eph receptors and ephrin ligands can be either upregulated or downregulated in cancer cells compared to normal tissues [5,7,13,14]. Aberrant EphA3 regulation has been reported in human hematopoietic malignancies and solid cancers [5,14,15]. High expression and oncogenic functions of EphA3 have been reported in acute lymphoblastic leukemia (ALL) [16], glioblastoma [17], gastric cancer [18], head and neck cancer [19], and prostate cancer [20]. Conversely, low expression and a tumor-suppressive role of EphA3 have been observed in small-cell lung cancer [21]. Additionally, the correlation between EphA3 mutational status and tumorigenesis in lung cancer has been reported [22]. These findings highlight EphA3 as an important therapeutic target for cancer treatment.
In the development of therapeutic monoclonal antibodies (mAbs) targeting EphA3, the mouse antibody (clone IIIA4) and its humanized defucosylated mAb ifabotuzumab (KB004) were developed to target EphA3-overexpressing cells [7,23]. Ifabotuzumab selectively binds to EphA3-positive cancer cells [24] and can stimulate antibody-dependent cell-mediated cytotoxicity (ADCC) [25]. This agent also prevents the proliferation of cancer cells and endothelial cells in the tumor vasculature by inhibiting EphA3 signaling [26]. Ifabotuzumab was evaluated in a Phase I clinical trial for the treatment of patients with hematological malignancies, demonstrating some encouraging clinical responses [24]. Moreover, treatment with IIIA4, conjugated to maytansine or lutetium-177, prevented tumor formation in glioblastoma-bearing mice [17]. Additionally, EphA3-targeted chimeric antigen receptor (CAR)-T cells demonstrated robust antigen-specific killing of human glioblastoma and diffuse midline glioma cell lines in animal models [27,28]. The EphA3 CAR, consisted of single-chain variable fragments (scFv) derived from the anti-EphA3 mAbs IIIA4 [27] and 3C3-1 [28], has been reported.
Since the development of therapeutic mAbs requires strict specificity to minimize off-target effects caused by cross-reactivity, we aimed to develop anti-EphA3 mAbs with no cross-reactivity to other Eph receptors. We have previously developed several mAbs against various membrane proteins, including Eph receptors, using the Cell-Based Immunization and Screening (CBIS) method [29,30,31,32,33]. The mAbs obtained using this method are prone to recognize conformational epitopes and are suitable for flow cytometry. Furthermore, some of these mAbs also apply to immunohistochemistry, contributing to therapeutic and diagnostic advancements. Therefore, we employed the CBIS method to generate anti-EphA3 mAbs with strict specificity to develop therapeutic and diagnostic agents targeting EphA3.
2. Materials and Methods
2.1. Plasmid Construction and Establishment of Stable Transfectants
The gene encoding human EPHA3 (NM_005233) was obtained from the RIKEN BioResource Research Center (Ibaraki, Japan). The open reading frames, excluding the signal sequences, were subcloned into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) with an N-terminal PA16 tag [34] or a MAP16 tag [35]. The plasmids were transfected into Chinese hamster ovary (CHO)-K1 and LN229 cells (ATCC, VA, USA), and stable transfectants were established as described previously [29]. Other Eph receptor-expressed CHO-K1 cells (e.g., CHO/EphA2) were established and the cell surface expression was confirmed as reported previously [29]. The human T cell leukemia cell lines Jurkat and MOLT-4 were obtained from the Cell Resource Center for Biomedical Research, Institute of Development, Aging and Cancer at Tohoku University (Miyagi, Japan) and the Japanese Collection of Research Bioresources Cell Bank (Osaka, Japan). These cell lines were cultured as described previously [36].
2.2. Development of Hybridomas
Two six -week-old female BALB/cAJcl mice (CLEA Japan, Tokyo, Japan) were intraperitoneally immunized with LN229/EphA3 cells (1 × 10⁸ cells) with Alhydrogel adjuvant (2%, InvivoGen). After three weekly immunizations (1 × 10⁸ cells), a final booster injection (1 × 10⁸ cells) was administered two days before splenocyte harvesting. The splenocytes were fused to P3X63Ag8.U1 (P3U1, ATCC). Hybridomas were generated as described previously [31].
2.3. Flow Cytometry
Cells were stained with Alexa Fluor 488-conjugated anti-mouse IgG (1:2,000 dilution; Cell Signaling Technology, Inc., Danvers, MA, USA) for 30 minutes at 4°C. Data were acquired using the SA3800 Cell Analyzer (Sony Corporation, Tokyo, Japan) and analyzed using FlowJo software (BD Biosciences, Franklin Lakes, NJ, USA).
2.4. Determination of Dissociation Constant Values Using Flow Cytometry
CHO/EphA3 and Jurkat cells were treated with serially diluted Ea3Mab-20 and IIIA4 (10 to 0.0006 μg/mL). Subsequently, the cells were stained with Alexa Fluor 488-conjugated anti-mouse IgG (1:200 dilution) for 30 minutes at 4°C. The dissociation constant (KD) values were determined as described previously [29].
2.5. Immunohistochemical Analysis
Formalin-fixed paraffin-embedded (FFPE) CHO/EphA3 and CHO-K1 cell blocks were prepared using iPGell (Genostaff Co., Ltd., Tokyo, Japan). Staining was performed using the VENTANA BenchMark ULTRA PLUS (Roche Diagnostics) with the recommended protocol and the ultraView Universal DAB Detection Kit.
3. Results
3.1. Development of Anti-Human EphA3 mAbs
Two BALB/cAJcl mice were immunized with LN229/EphA3 cells (Figure 1A). After harvesting splenocytes from these mice, cell fusion with P3U1 cells was performed (Figure 1B). The resulting hybridomas were seeded into 96-well plates. After colony formation, supernatants were collected and analyzed by flow cytometry-based high-throughput screening to identify those that were negative for CHO-K1 cells and positive for CHO/EphA3 cells (Figure 1C). Subsequently, 20 hybridomas producing anti-EphA3 mAbs were cloned by limiting dilution. Finally, anti-EphA3 mAb clones, including Ea3Mab-20 (IgG1, kappa), were established (Figure 1D).
3.2. Flow Cytometry Using Ea3Mab-20 and IIIA4
We assessed the reactivity of Ea3Mab-20 against CHO/EphA3 and CHO-K1 cells. Ea3Mab-20 recognized CHO/EphA3 cells in a dose-dependent manner at concentrations ranging from 1 to 0.001 μg/mL (Figure 2A). However, Ea3Mab-20 did not bind to CHO-K1 cells at any concentrations (Figure 2B). This result indicates that Ea3Mab-20 recognizes EphA3 on the cell surface. A commercially available anti-human EphA3 mAb (IIIA4) exhibited a similar pattern of reactivity with CHO/EphA3 and CHO-K1 cells (Figure 2). Next, we analyzed the reactivity of Ea3Mab-20 against endogenous EphA3-expressing cells, Jurkat (Figure 3A) and MOLT-4 (Figure 3B) cells [16], using IIIA4 as a positive control (Figure 3). Dose-dependent reactivity was observed, suggesting that Ea3Mab-20 binds to endogenous EphA3.
3.3. Specificity of Ea3Mab-20 Using CHO-K1 Cells Overexpressed Various Eph Receptors
We previously established CHO-K1 that overexpressed each human Eph receptor (EphA1 to A8, A10, B1 to B4, and B6) [29]. Using these cell lines, we analyzed the specificity of anti-EphA3 mAbs. Among the 20 clones producing anti-EphA3 mAbs, 13 highly reactive clones were selected for analysis. Staining data with 10 μg/mL of Ea3Mab-3, 4, 7, 9, 15, and 20 showed no cross-reactivity among the Eph receptors (Figure 4 and Supplementary Table S1). However, IIIA4 exhibited slight but significant reactivity with CHO/EphA6 cells (Supplementary Figure S1), indicating that IIIA4 does not exhibit complete specificity for EphA3. Ea3Mab-20 exhibited the highest binding affinity and specificity among these mAbs.
3.4. Determination of Binding Affinity of Ea3Mab-20 and IIIA4 Using Flow Cytometry
The binding affinity of Ea3Mab-20 and IIIA4 was determined using CHO/EphA3 and Jurkat cells. The average KD values obtained from three independent measurements for Ea3Mab-20 were 9.0 ± 0.3 × 10⁻⁹ M for CHO/EphA3 cells (Figure 5A and Supplementary Figure S2) and 1.4 ± 0.1 × 10⁻⁹ M for Jurkat cells (Figure 5B and Supplementary Figure S2). The average KD values for IIIA4 were 2.4 ± 0.3 × 10⁻⁹ M for CHO/EphA3 cells and 5.7 ± 0.9 × 10⁻11 M for Jurkat cells (Figure 5 and Supplementary Figure S2)
3.5. Immunohistochemistry Using Anti-EphA3 mAbs
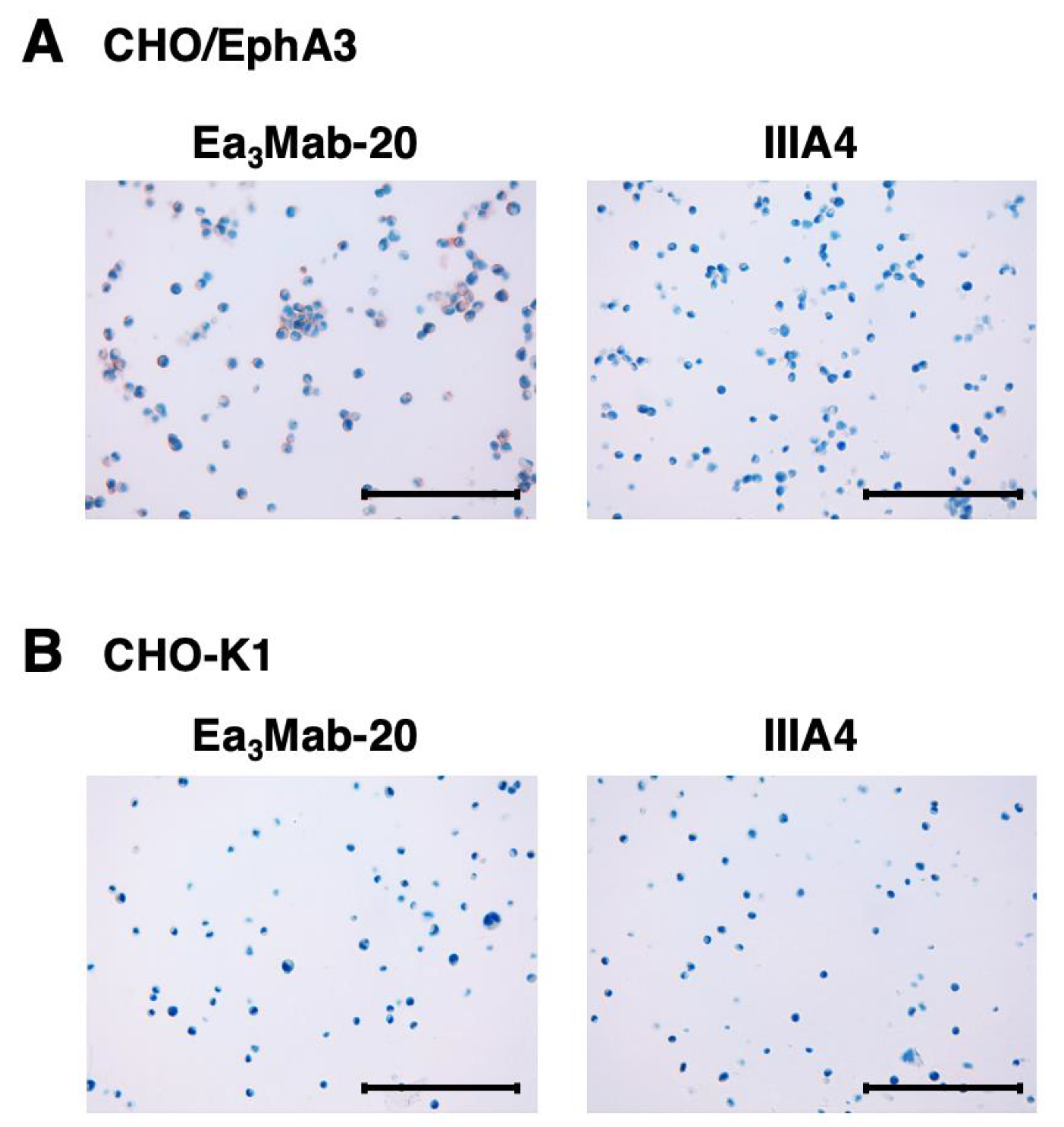
Ea3Mab-20 was assessed for its application in immunohistochemistry using FFPE CHO-K1 and CHO/EphA3 cell sections. We used the VENTANA BenchMark ULTRA PLUS system for the detection. Apparent membranous staining by Ea3Mab-20 was observed in CHO/EphA3 cells (Figure 6A) but not in CHO-K1 cells (Figure 6B). These results indicate that Ea3Mab-20 is suitable for detecting EphA3-positive cells not only in flow cytometry but also in FFPE samples. In contrast, IIIA4 is unavailable for staining FFPE cell sections (Figure 6).
4. Discussion
Using the CBIS method, we generated novel anti-EphA3 mAbs, including Ea3Mab-20, with confirmed specificity for EphA3. Ea3Mab-20 is suitable for flow cytometry (Figure 2 and Figure 3) and immunohistochemistry (Figure 6). Since Ea3Mab-20 does not cross-react with other Eph receptors expressed in CHO-K1 cells (Figure 4), it is versatile for basic research and is expected to contribute to the development of clinical applications of mAbs for cancer therapy and diagnosis.
The cross-reactivity of IIIA4 with EphA6 may be due to the similarity of the extracellular domain among Eph receptors. The amino acid identity and similarity of the extracellular domain among human Eph receptors range from 33 to 65% and 72 to 91%, respectively (Supplementary Figure S3A). The extracellular domain of EphA6 is the most similar in the phylogenetic tree among Eph receptors (Supplementary Figure S3B), exhibiting a high amino acid identity (64%) and similarity (91%) with that of EphA3. This may explain why an anti-EphA3 mAb IIIA4 cross-reacts with EphA6. Additionally, several clones of the obtained anti-EphA3 mAbs showed cross-reactivity with various Eph receptors, including EphA6 (Supplementary Table S1). Since the development of therapeutic mAbs requires strict specificity to avoid off-target effects caused by cross-reactivity, Ea3Mab-20 is a promising candidate for further development as a therapeutic and diagnostic agent.
The binding affinity of IIIA4 is approximately 3.8-fold higher than that of Ea3Mab-20 when analyzed using CHO/EphA3 cells (Figure 5). However, this ratio increases to approximately 24.6-fold when using Jurkat cells endogenously express EphA3 (Figure 5). This discrepancy may be due to the endogenous expression of EphA6 in Jurkat cells [35], which could have influenced the affinity analysis through the cross-reactivity of IIIA4 with EphA6. The mAbs with high specificity for Eph receptors, such as Ea3Mab-20, are crucial for clinical applications, even though the binding affinity of Ea3Mab-20 (KD: 9.0 ± 0.3 × 10⁻⁹ M) is lower than that of IIIA4 (KD: 2.4 ± 0.3 × 10⁻⁹ M) in CHO/EphA3 cells.
Several preclinical studies reported that anti-EphA3 CAR-T cells are effective against glioblastoma [27,28]. Regarding the binding affinity of CARs consisting of scFv, it has been reported that a low-affinity CAR (KD: 1.4 × 10-8 M), which exhibits more than 40-fold lower affinity for CD19 compared to existing scFvs derived from FMC63, enhances CAR-T cell expansion and prolongs persistence in pediatric patients with ALL [37]. Additionally, it has been noted that a faster off-rate, which reflects the rate at which the antibody dissociates from the antigen, is preferred, particularly in CAR-T therapies. The on-rate and off-rate of Ea3Mab-20 should be determined. However, its affinity for CHO/EphA3 and Jurkat cells is on the order of 10⁻⁹ M, which may provide sufficient potential for CAR development.
We have previously developed two methods for epitope mapping: PA insertion for epitope mapping (PAMAP) and RIEDL insertion for epitope mapping (REMAP) [38,39,40,41,42]. These approaches have successfully identified the epitopes of various mAbs, including anti-mouse CD39 mAb (C39Mab-1) [38], anti-CD44 mAbs (C44Mab-5 and C44Mab-46) [39,40], and anti-EGFR mAbs (EMab-51 and EMab-134) [41,42]. Further investigation is necessary to determine the epitope of Ea3Mab-20. If Ea3Mab-20 targets a linear and non-glycosylated epitope, this finding could facilitate the development of broadly applicable and highly specific mAbs against other Eph family members through peptide-based immunization strategies.
To effectively target EphA3-positive cancer cells using Ea3Mab-20 (IgG1), generating a class-switched variant with a mouse IgG2a backbone would be beneficial. Additionally, our previous studies demonstrated that defucosylated IgG2a mAbs enhance ADCC activity and exhibit more potent antitumor effects in mouse xenograft models [43,44]. Developing a class-switched and defucosylated version of Ea3Mab-20 could improve its therapeutic efficacy against EphA3-positive cancers in preclinical research. Furthermore, it is also essential to evaluate whether Ea3Mab-20 demonstrates ADCC activity or drug-induced cytotoxicity when conjugated with cytotoxic agents.
Altogether, Ea3Mab-20 reacts with EphA3 without cross-reactivity with Eph family members and is suitable for flow cytometry and immunohistochemistry. EphA3 is a potential therapeutic target, especially in hematopoietic malignancies and brain cancers. Therefore, Ea3Mab-20 is a highly sensitive and versatile mAb for basic research and is expected to contribute to clinical applications such as antibody therapy and tumor diagnosis.
Credit authorship contribution statement: Hiroyuki Satofuka: Investigation, Funding acquisition, Writing – original draft. Hiroyuki Suzuki: Investigation, Funding acquisition, Writing – review and editing. Miu Hirose: Investigation. Keisuke Shinoda: Investigation. Takuya Nakamura: Investigation. Tomohiro Tanaka: Investigation, Funding acquisition. Mika K. Kaneko: Conceptualization. Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing – review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP24am0521010 (to Y.K.), JP24ama121008 (to Y.K.), JP24ama221339 (to Y.K.), JP24bm1123027 (to Y.K.), and JP24ck0106730 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 24K11652 (to H.Satofuka), 22K06995 (to H.Suzuki), 24K18268 (to T.T), and 22K07224 (to Y.K.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Barquilla, A.; Pasquale, E.B. Eph receptors and ephrins: therapeutic opportunities. Annu Rev Pharmacol Toxicol 2015;55: 465-487. [CrossRef]
- Darling, T.K.; Lamb, T.J. Emerging Roles for Eph Receptors and Ephrin Ligands in Immunity. Front Immunol 2019;10: 1473.
- Liang, L.Y.; Patel, O.; Janes, P.W.; Murphy, J.M.; Lucet, I.S. Eph receptor signalling: from catalytic to non-catalytic functions. Oncogene 2019;38(39): 6567-6584.
- Pasquale, E.B. Eph receptors and ephrins in cancer progression. Nat Rev Cancer 2024;24(1): 5-27. [CrossRef]
- Buckens, O.J.; El Hassouni, B.; Giovannetti, E.; Peters, G.J. The role of Eph receptors in cancer and how to target them: novel approaches in cancer treatment. Expert Opin Investig Drugs 2020;29(6): 567-582.
- Poliakov, A.; Cotrina, M.; Wilkinson, D.G. Diverse roles of eph receptors and ephrins in the regulation of cell migration and tissue assembly. Dev Cell 2004;7(4): 465-480. [CrossRef]
- Pasquale, E.B. Eph receptor signalling casts a wide net on cell behaviour. Nat Rev Mol Cell Biol 2005;6(6): 462-475.
- Beckmann, M.P.; Cerretti, D.P.; Baum, P.; et al. Molecular characterization of a family of ligands for eph-related tyrosine kinase receptors. EMBO J 1994;13(16): 3757-3762.
- Hirai, H.; Maru, Y.; Hagiwara, K.; Nishida, J.; Takaku, F. A novel putative tyrosine kinase receptor encoded by the eph gene. Science 1987;238(4834): 1717-1720.
- Lisabeth, E.M.; Falivelli, G.; Pasquale, E.B. Eph receptor signaling and ephrins. Cold Spring Harb Perspect Biol 2013;5(9).
- Batlle, E.; Wilkinson, D.G. Molecular mechanisms of cell segregation and boundary formation in development and tumorigenesis. Cold Spring Harb Perspect Biol 2012;4(1): a008227. [CrossRef]
- Pasquale, E.B. Eph-ephrin bidirectional signaling in physiology and disease. Cell 2008;133(1): 38-52.
- Hanover, G.; Vizeacoumar, F.S.; Banerjee, S.L.; et al. Integration of cancer-related genetic landscape of Eph receptors and ephrins with proteomics identifies a crosstalk between EPHB6 and EGFR. Cell Rep 2023;42(7): 112670.
- Xi, H.Q.; Wu, X.S.; Wei, B.; Chen, L. Eph receptors and ephrins as targets for cancer therapy. J Cell Mol Med 2012;16(12): 2894-2909. [CrossRef]
- London, M.; Gallo, E. Critical role of EphA3 in cancer and current state of EphA3 drug therapeutics. Mol Biol Rep 2020;47(7): 5523-5533.
- Charmsaz, S.; Al-Ejeh, F.; Yeadon, T.M.; et al. EphA3 as a target for antibody immunotherapy in acute lymphoblastic leukemia. Leukemia 2017;31(8): 1779-1787. [CrossRef]
- Day, B.W.; Stringer, B.W.; Al-Ejeh, F.; et al. EphA3 maintains tumorigenicity and is a therapeutic target in glioblastoma multiforme. Cancer Cell 2013;23(2): 238-248.
- Nasri, B.; Inokuchi, M.; Ishikawa, T.; et al. High expression of EphA3 (erythropoietin-producing hepatocellular A3) in gastric cancer is associated with metastasis and poor survival. BMC Clin Pathol 2017;17: 8.
- Kim, S.H.; Lee, W.H.; Kim, S.W.; et al. EphA3 maintains radioresistance in head and neck cancers through epithelial mesenchymal transition. Cell Signal 2018;47: 122-130.
- Wu, R.; Wang, H.; Wang, J.; et al. EphA3, induced by PC-1/PrLZ, contributes to the malignant progression of prostate cancer. Oncol Rep 2014;32(6): 2657-2665.
- Peng, J.; Wang, Q.; Liu, H.; et al. EPHA3 regulates the multidrug resistance of small cell lung cancer via the PI3K/BMX/STAT3 signaling pathway. Tumour Biol 2016;37(9): 11959-11971.
- Lisabeth, E.M.; Fernandez, C.; Pasquale, E.B. Cancer somatic mutations disrupt functions of the EphA3 receptor tyrosine kinase through multiple mechanisms. Biochemistry 2012;51(7): 1464-1475.
- Vail, M.E.; Murone, C.; Tan, A.; et al. Targeting EphA3 inhibits cancer growth by disrupting the tumor stromal microenvironment. Cancer Res 2014;74(16): 4470-4481.
- Swords, R.T.; Greenberg, P.L.; Wei, A.H.; et al. KB004, a first in class monoclonal antibody targeting the receptor tyrosine kinase EphA3, in patients with advanced hematologic malignancies: Results from a phase 1 study. Leuk Res 2016;50: 123-131.
- Tomasevic, N.; Luehrsen, K.; Baer, M.; et al. A high affinity recombinant antibody to the human EphA3 receptor with enhanced ADCC activity. Growth Factors 2014;32(6): 223-235. [CrossRef]
- Lv, X.Y.; Wang, J.; Huang, F.; et al. EphA3 contributes to tumor growth and angiogenesis in human gastric cancer cells. Oncol Rep 2018;40(4): 2408-2416.
- Lertsumitkul, L.; Iliopoulos, M.; Wang, S.S.; et al. EphA3-targeted chimeric antigen receptor T cells are effective in glioma and generate curative memory T cell responses. J Immunother Cancer 2024;12(8).
- Martins, P.; D'Souza, R.C.J.; Skarne, N.; et al. EphA3 CAR T cells are effective against glioblastoma in preclinical models. J Immunother Cancer 2024;12(8).
- Ubukata, R.; Suzuki, H.; Hirose, M.; et al. Establishment of a highly sensitive and specific anti-EphB2 monoclonal antibody (Eb2Mab-12) for flow cytometry. MI 2025. [CrossRef]
- Tanaka, T.; Suzuki, H.; Li, G.; et al. Ea 1 Mab-30: A Novel Monoclonal Antibody Against Erythropoietin-Producing Hepatocellular Receptor A1 for Versatile Applications Preprint 2025.
- Tanaka, T.; Kaneko, Y.; Yamamoto, H.; et al. Development of a novel anti-erythropoietin-producing hepatocellular receptor B6 monoclonal antibody Eb(6)Mab-3 for flow cytometry. Biochem Biophys Rep 2025;41: 101960. [CrossRef]
- Satofuka, H.; Suzuki, H.; Tanaka, T.; et al. Development of an anti-human EphA2 monoclonal antibody Ea2Mab-7 for multiple applications. Biochemistry and Biophysics Reports 2025;42: 101998.
- Nanamiya, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of an Anti-EphB4 Monoclonal Antibody for Multiple Applications Against Breast Cancers. Monoclon Antib Immunodiagn Immunother 2023;42(5): 166-177.
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; et al. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014;95: 240-247.
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon Antib Immunodiagn Immunother 2016;35(6): 293-299.
- Tanaka, T.; Suzuki, H.; Isoda, Y.; et al. Development of a Sensitive Anti-Human CCR9 Monoclonal Antibody (C(9)Mab-11) by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022;41(6): 303-310.
- Ghorashian, S.; Kramer, A.M.; Onuoha, S.; et al. Enhanced CAR T cell expansion and prolonged persistence in pediatric patients with ALL treated with a low-affinity CD19 CAR. Nat Med 2019;25(9): 1408-1414.
- Okada, Y.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Epitope Mapping of an Anti-Mouse CD39 Monoclonal Antibody Using PA Scanning and RIEDL Scanning. Monoclon Antib Immunodiagn Immunother 2024;43(2): 44-52.
- Asano, T.; Kaneko, M.K.; Takei, J.; Tateyama, N.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C(44)Mab-46) Using the REMAP Method. Monoclon Antib Immunodiagn Immunother 2021;40(4): 156-161. [CrossRef]
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon Antib Immunodiagn Immunother 2021;40(4): 162-167.
- Sano, M.; Kaneko, M.K.; Aasano, T.; Kato, Y. Epitope Mapping of an Antihuman EGFR Monoclonal Antibody (EMab-134) Using the REMAP Method. Monoclon Antib Immunodiagn Immunother 2021;40(4): 191-195.
- Nanamiya, R.; Sano, M.; Asano, T.; et al. Epitope Mapping of an Anti-Human Epidermal Growth Factor Receptor Monoclonal Antibody (EMab-51) Using the RIEDL Insertion for Epitope Mapping Method. Monoclon Antib Immunodiagn Immunother 2021;40(4): 149-155.
- Ishikawa, K.; Suzuki, H.; Ohishi, T.; et al. Antitumor activities of anti-CD44 monoclonal antibodies in mouse xenograft models of esophageal cancer. Oncol Rep 2024;52(5).
- Ishikawa, K.; Suzuki, H.; Ohishi, T.; et al. Anti-CD44 Variant 10 Monoclonal Antibody Exerts Antitumor Activity in Mouse Xenograft Models of Oral Squamous Cell Carcinomas. Int J Mol Sci 2024;25(17). [CrossRef]
Figure 1.
Schematic illustration of anti-EphA3 mAb production using the CBIS method. (A) Two female BALB/cAJcl mice were intraperitoneally injected with LN229/EphA3 cells. (B) Splenocytes were harvested and fused with P3U1 myeloma cells using PEG1500. (C) Hybridoma supernatants were screened by flow cytometry using CHO/EphA3 and parental CHO-K1 cells to identify anti-EphA3-specific mAbs. (D) Hybridoma clones producing antigen-specific mAbs were isolated through the limiting dilution method.
Figure 1.
Schematic illustration of anti-EphA3 mAb production using the CBIS method. (A) Two female BALB/cAJcl mice were intraperitoneally injected with LN229/EphA3 cells. (B) Splenocytes were harvested and fused with P3U1 myeloma cells using PEG1500. (C) Hybridoma supernatants were screened by flow cytometry using CHO/EphA3 and parental CHO-K1 cells to identify anti-EphA3-specific mAbs. (D) Hybridoma clones producing antigen-specific mAbs were isolated through the limiting dilution method.
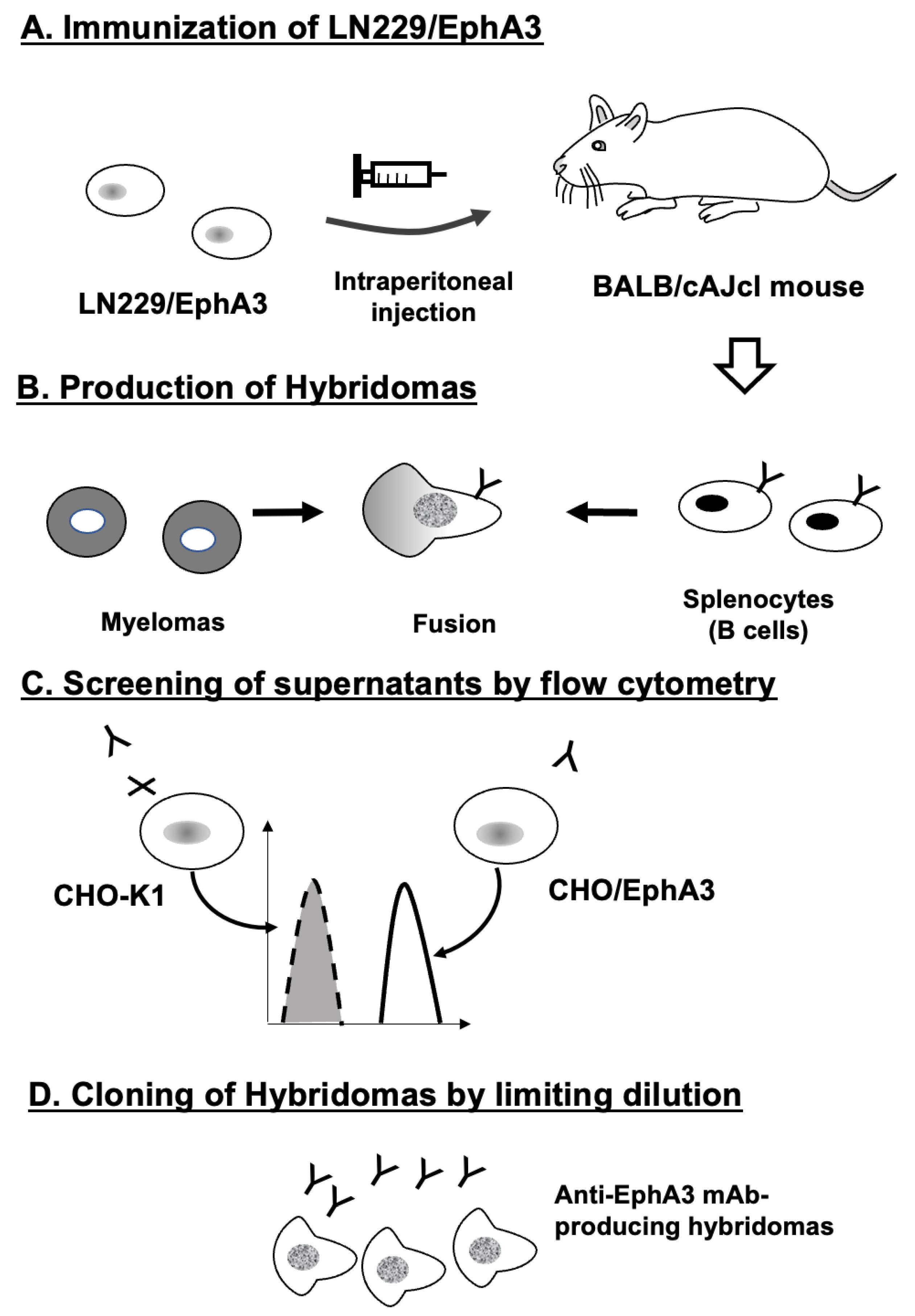
Figure 2.
Flow cytometry analysis of anti-EphA3 mAbs against CHO/EphA3 and CHO-K1 cells. CHO/EphA3 (A) and CHO-K1 (B) cells were treated with Ea3Mab-20 or the commercially available anti-EphA3 mAb IIIA4 at the indicated concentrations. Cells were stained with (red lines) or without (black lines) anti-EphA3 mAbs, followed by staining with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were subsequently acquired using the SA3800 Cell Analyzer.
Figure 2.
Flow cytometry analysis of anti-EphA3 mAbs against CHO/EphA3 and CHO-K1 cells. CHO/EphA3 (A) and CHO-K1 (B) cells were treated with Ea3Mab-20 or the commercially available anti-EphA3 mAb IIIA4 at the indicated concentrations. Cells were stained with (red lines) or without (black lines) anti-EphA3 mAbs, followed by staining with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were subsequently acquired using the SA3800 Cell Analyzer.
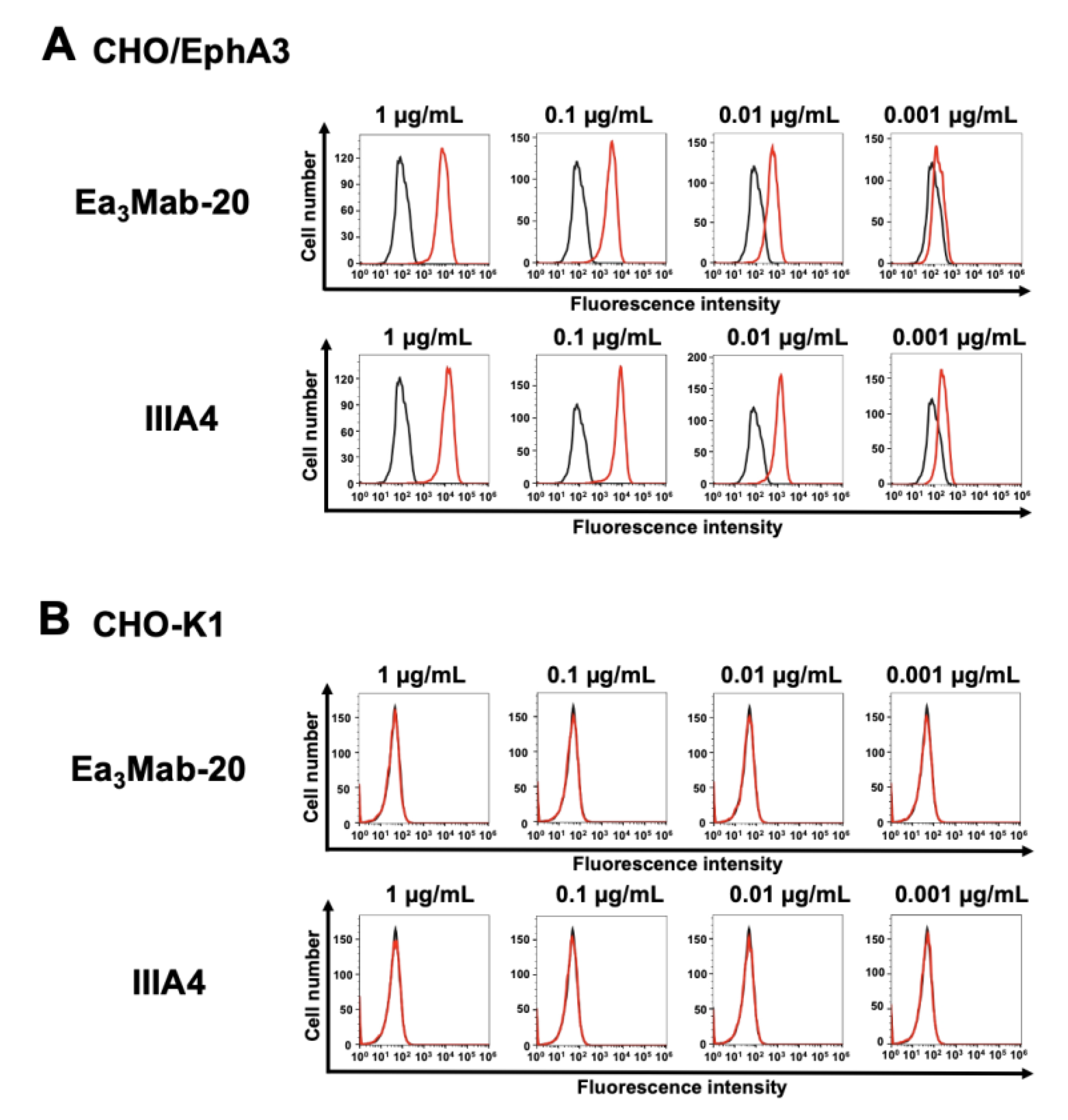
Figure 3.
Flow cytometry analysis of anti-EphA3 mAbs against endogenous EphA3 expressing cancer cells. Jurkat (A) and MOLT-4 (B) cells were treated with Ea3Mab-20 and IIIA4 at the indicated concentrations (red lines). The black line represents the negative control, which was stained without anti-EphA3 mAbs (blocking buffer). The mAb-treated cells were incubated with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were subsequently collected using the SA3800 Cell Analyzer.
Figure 3.
Flow cytometry analysis of anti-EphA3 mAbs against endogenous EphA3 expressing cancer cells. Jurkat (A) and MOLT-4 (B) cells were treated with Ea3Mab-20 and IIIA4 at the indicated concentrations (red lines). The black line represents the negative control, which was stained without anti-EphA3 mAbs (blocking buffer). The mAb-treated cells were incubated with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were subsequently collected using the SA3800 Cell Analyzer.
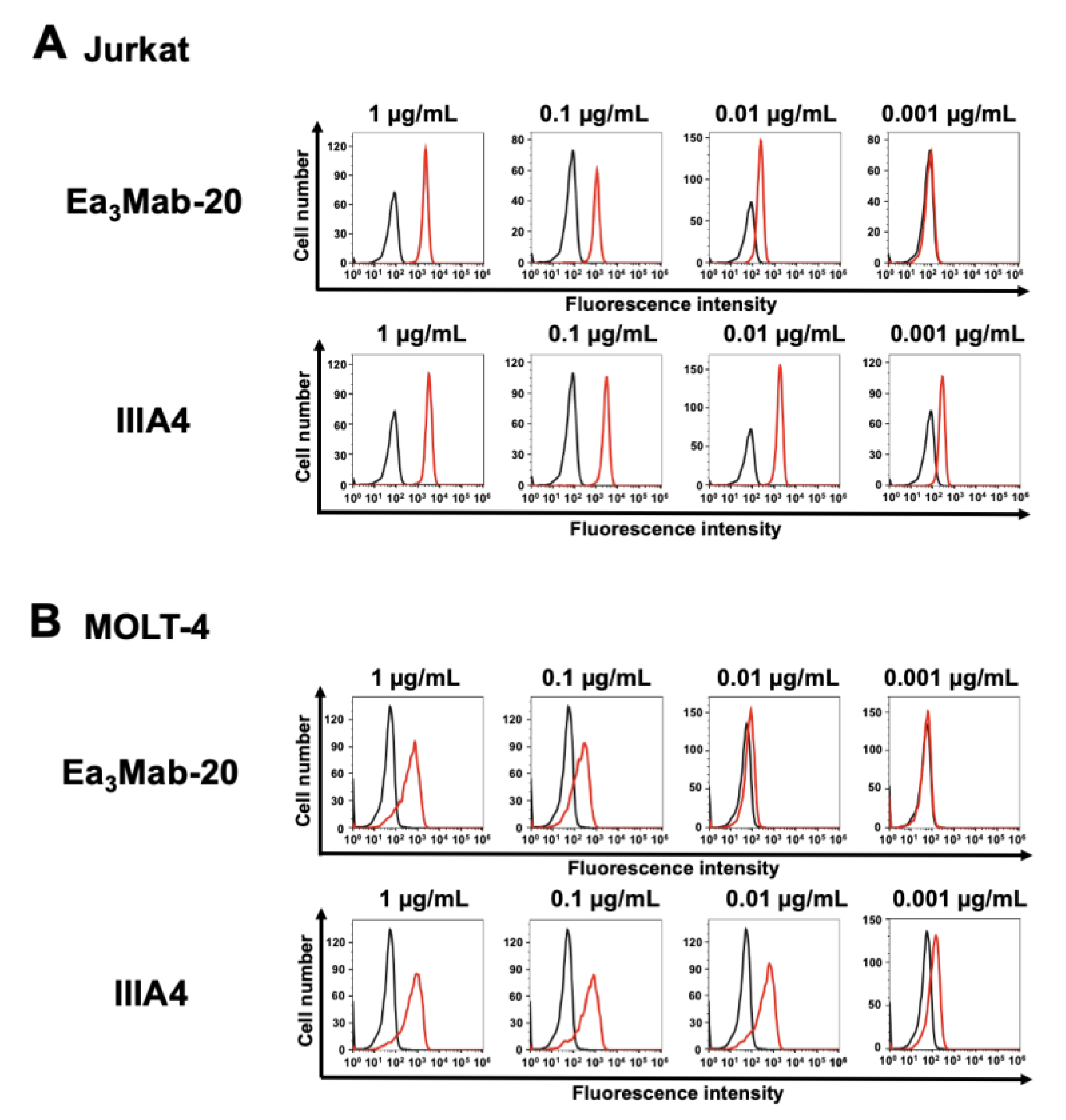
Figure 4.
Cross-reactivity of Ea3Mab-20 in Eph receptor-expressed CHO-K1 cells. The 14 Eph receptors (EphA1 to A8, A10, B1 to B4, and B6)-expressed CHO-K1 and parental CHO-K1 cells were treated with 10 μg/mL of Ea3Mab-20 (red lines). The black line represents the negative control, which was stained without anti-EphA3 mAbs (blocking buffer). The mAb-treated cells were subsequently incubated with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer.
Figure 4.
Cross-reactivity of Ea3Mab-20 in Eph receptor-expressed CHO-K1 cells. The 14 Eph receptors (EphA1 to A8, A10, B1 to B4, and B6)-expressed CHO-K1 and parental CHO-K1 cells were treated with 10 μg/mL of Ea3Mab-20 (red lines). The black line represents the negative control, which was stained without anti-EphA3 mAbs (blocking buffer). The mAb-treated cells were subsequently incubated with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer.
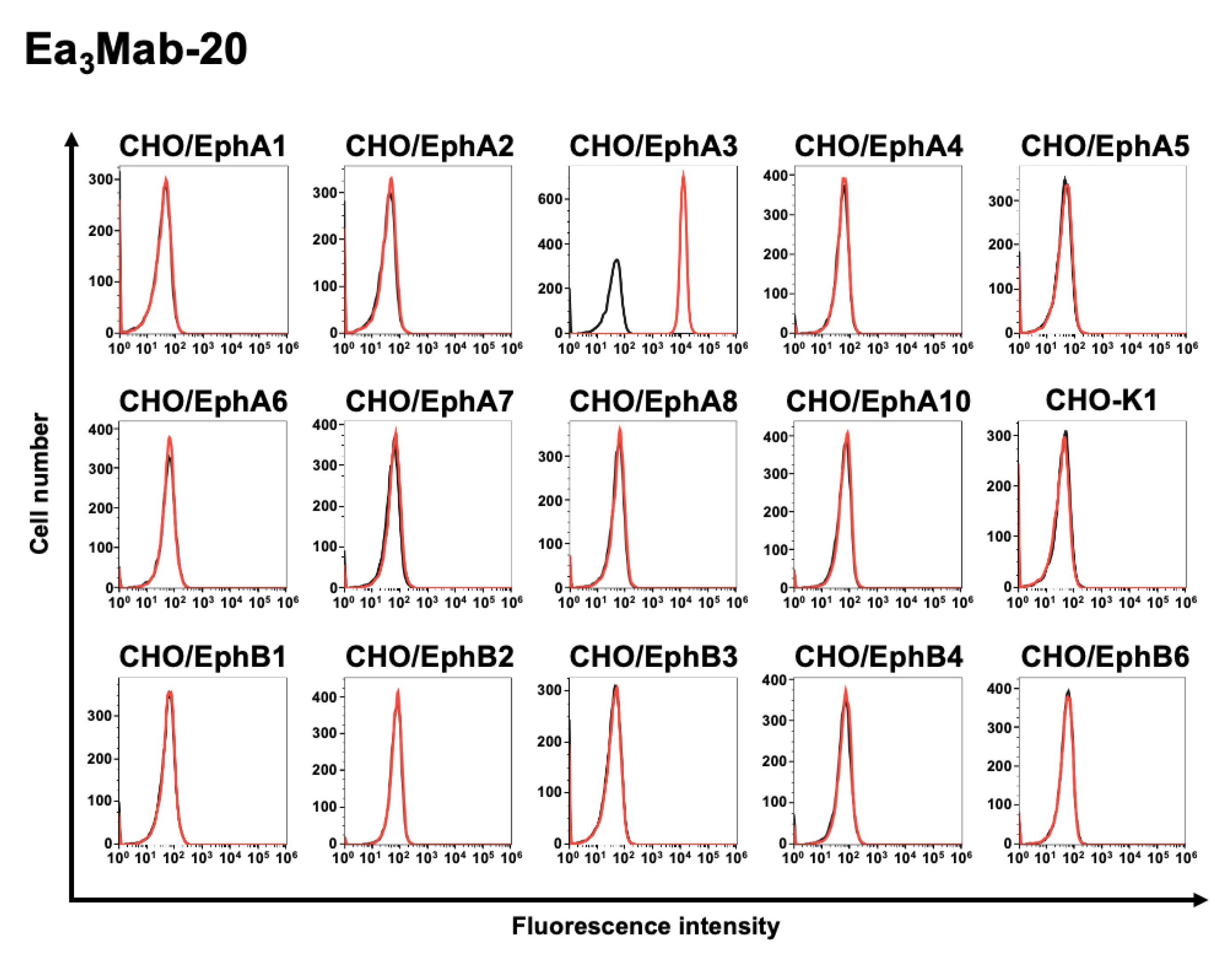
Figure 5.
Measurement of the binding affinity of Ea3Mab-20 and IIIA4. CHO/EphA3 (A) and Jurkat (B) cells were treated with 100 μL of serial dilutions of Ea3Mab-20 and IIIA4 (10 to 0.0006 μg/mL). Subsequently, fluorescence data were collected using the SA3800 Cell Analyzer. The fluorescence data's geometric mean (GeoMean) values were plotted, and the KD values were calculated using GraphPad PRISM 6 software.
Figure 5.
Measurement of the binding affinity of Ea3Mab-20 and IIIA4. CHO/EphA3 (A) and Jurkat (B) cells were treated with 100 μL of serial dilutions of Ea3Mab-20 and IIIA4 (10 to 0.0006 μg/mL). Subsequently, fluorescence data were collected using the SA3800 Cell Analyzer. The fluorescence data's geometric mean (GeoMean) values were plotted, and the KD values were calculated using GraphPad PRISM 6 software.
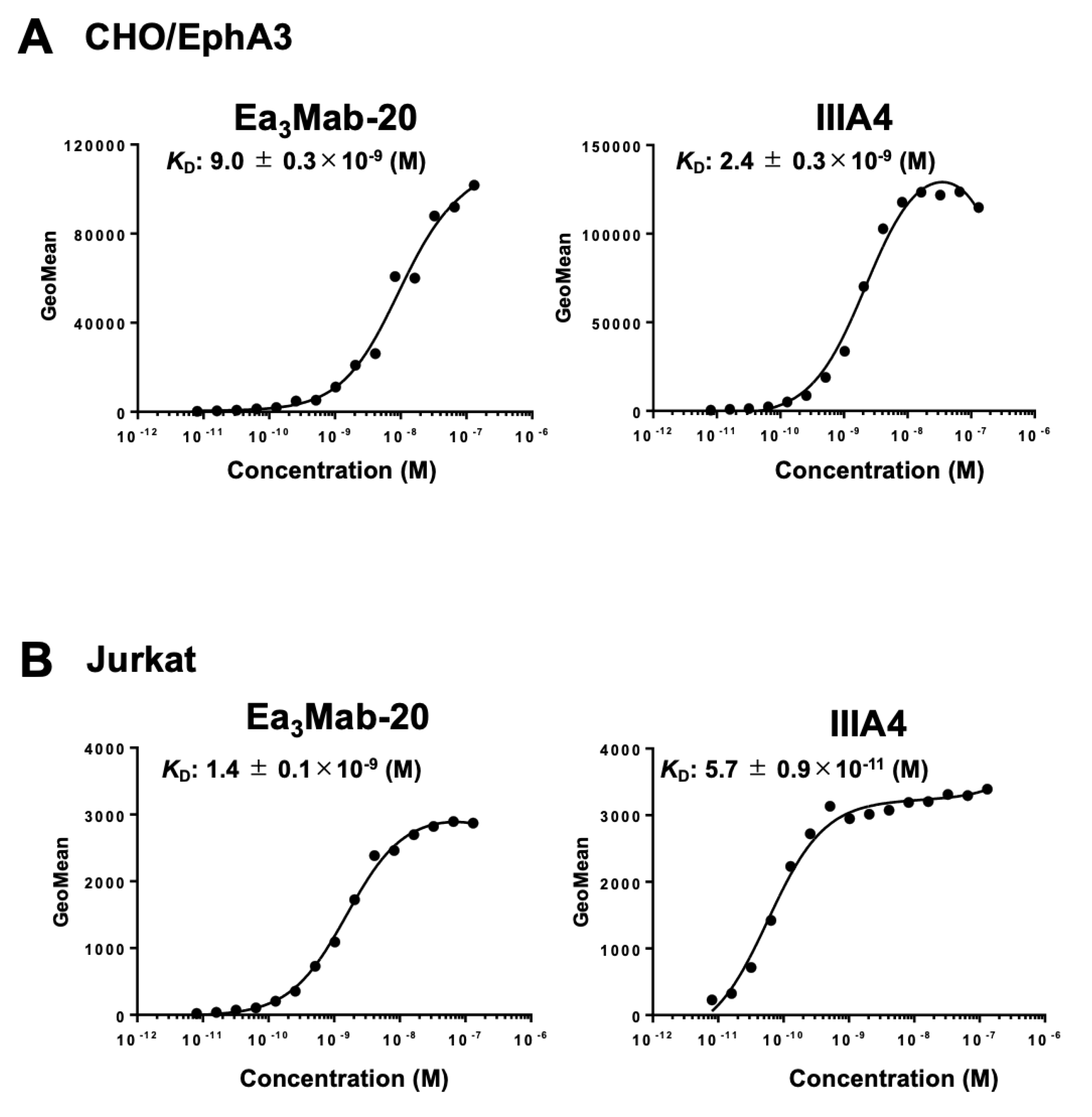
Figure 6.
Immunohistochemical staining of paraffin-embedded cell sections of CHO/EphA3 and CHO-K1 cells. Sections of CHO/EphA3 (A) and CHO-K1 (B) cells were stained with 10 μg/mL of Ea3Mab-20 and IIIA4 using the VENTANA BenchMark ULTRA PLUS. Scale bar = 100 μm.
Figure 6.
Immunohistochemical staining of paraffin-embedded cell sections of CHO/EphA3 and CHO-K1 cells. Sections of CHO/EphA3 (A) and CHO-K1 (B) cells were stained with 10 μg/mL of Ea3Mab-20 and IIIA4 using the VENTANA BenchMark ULTRA PLUS. Scale bar = 100 μm.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



