
Submitted:
14 December 2024
Posted:
16 December 2024
You are already at the latest version
Abstract
Erythropoietin-producing hepatocellular receptor B6 (EphB6) is a member of the largest Eph subfamily of receptor tyrosine kinases. EphB6 is widely expressed in various tissues and regulates cellular homeostasis by interacting with its membrane-bound ephrin ligands and other receptors. EphB6 is involved in cancer pathology despite lacking kinase activity. Therefore, developing sensitive monoclonal antibodies (mAbs) for EphB6 has been desired for treatment, diagnosis, and further analysis of EphB6. This study established a novel specific and sensitive anti-human EphB6 mAb clone Eb6Mab-3 (mouse IgG1, kappa) by the Cell-Based Immunization and Screening (CBIS) method. In flow cytometry, Eb6Mab-3 demonstrated reactivity with EphB6-overexpressed Chinese hamster ovary-K1 cells (CHO/EphB6) and endogenously EphB6-expressing DLD-1 colorectal cancer cells. Cross-reactivity of Eb6Mab-3 was not observed. Eb6Mab-3 demonstrated a moderate binding affinity (dissociation constant; KD) for CHO/EphB6 (KD: 3.8×10-8 M) and a high binding affinity for DLD-1 (KD: 1.9×10-9 M). Eb6Mab-3 can detect EphB6 protein in CHO/EphB6 lysate in western blot. Therefore, Eb6Mab-3, established by the CBIS method, could be valuable for analyzing the EphB6-associated cellular functions and has potential applications in diagnosis and treatment with specificity and high affinity for cancer cells.
Keywords:
Eph receptor
; EphB6
; CBIS method
; monoclonal antibody
; flow cytometry
1. Introduction
Receptor tyrosine kinases (RTKs) play pivotal roles in tissue homeostasis, including cell proliferation, differentiation, migration, tissue remodeling, and angiogenesis [1,2,3,4,5]. Aberrant signaling of abnormal RTKs leads to disordered cellular homeostasis and tumor development [6,7]. Erythropoietin-producing hepatocellular (Eph) receptors belong to the most prominent family of RTKs and exert diverse functions by binding to their ligands, ephrin. The Eph and ephrin families comprise 14 Eph receptors (EphA1 to EphA8, EphA10, EphB1 to EphB4, and EphB6) and eight ephrin ligands [glycosylphosphatidylinositol (GPI)-anchored ephrin A1 to A5 and transmembrane ephrin B1 to B3], respectively [8]. The Eph receptor and ephrin complexes mediate the receptor dimerization and clustering to initiate forward and reverse bidirectional signaling to each expressing cells [9,10]. Eph receptors and ephrin can affect various cell types in healthy tissues and disorders through these signaling [11,12,13].
EphB6 is classified as the tyrosine kinase-inactive pseudokinases as well as EphA10, owing to the containing a pseudokinase domain that lacks catalytic activity in the intracellular domain [14,15,16]. The ligands for EphB6 are still unclear, but ephrinB1 and ephrinB2 are the candidates [17,18]. Despite inactive kinase, EphB6 plays a vital role in controlling the cell functions by binding the ephrins and interacting with other RTKs. EphB6 serves as an oncogene which involved in the development and malignancy of tumors, including colon [19], leukemia [20,21], tongue squamous cell carcinoma [22], breast [23], bladder [24], and non-small cell lung cancer (NSCLC) [25]. EphB6 undergoes tyrosine transphosphorylation upon stimulation with ephrin B1 and ephrin B2 by forming a cluster with EphB1 and EphB4 [26,27]. Also, the interaction of EphB6 and EphA2 has been observed in breast cancer cell lines [28]. These findings suggest that EphB6 possesses valuable functions even in a kinase-dead RTK by cross-talking with partners and might contribute to cancer progression. In clinical thyroid malignant lesions from patients, the higher expression of EphB6 and EphB4 has been observed compared to benign ones and correlated with tumor size [29].
Interestingly, numerous reports have also described the role of EphB6 as a tumor suppressor. Loss of EphB6 brings tumor malignancy and poor prognosis [23,30,31,32,33,34]. Low EphB6 expression is associated with poor TNM stage and tumor grade in ovarian serous carcinoma and neuroblastoma [35,36]. Also, in molecular level analysis, EphB6 suppresses EphA2-promoted anoikis of breast cancer cells by interfering with EphA2-Ephexin 4 interaction [28]. In EphB6 null mice, the T cells had reduced the secretion of interleukin-2 (IL-2), IL-4, and interferon-γ [37]. Conversely, T cell proliferation and lymphokines secretion are enhanced by co-stimulation of EphB6 and T cell receptor [38]. These reports suggest that EphB6 exhibits tumor-suppressive functions within cancer cells and through the immune systems.
Mutations in the EphB6 gene that promote tumor metastasis have been identified in NSCLC patients [25]. The EphB6 mutation mediates paclitaxel resistance by simultaneously upregulating the expression of EphA2 and cadherin 11 [39]. Further analysis is essential to determine whether EphB6 promotes or suppresses tumors, and the development of highly sensitive antibodies against EphB6 is desired for basic research, diagnosis, and treatment.
Previously, we have established monoclonal antibodies (mAbs) against human EphA2 (clone Ea2Mab-7) [40], human EphB2 (clone Eb2Mab-3) [41], and human EphB4 (clone B4Mab-7) [42] by using the Cell-Based Immunization and Screening (CBIS) method. This method can efficiently develop a wide variety of antibodies that recognize linear epitope, structural epitope, and modifications of extracellular domains of membrane protein in a short period. In this study, we have successfully established a novel anti-human EphB6 mAb (clone Eb6Mab-3) using the CBIS method.
2. Materials and Methods
2.1. Cell Lines and Stable Transfectants
Cell lines, including LN229, Chinese hamster ovary (CHO)-K1, and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (Manassas, VA, USA). DLD-1 cells were obtained from the Cell Resource Center for Biomedical Research, Institute of Development, Aging and Cancer, Tohoku University (Miyagi, Japan). The expression plasmid of EphB6 (pCMV6neoEphB6-Myc-DDK, Catalog No.: RC229404, Accession No.: NM_004445, OriGene Technologies, Inc. Rockville, MD, USA) was transfected into cell lines using the Neon transfection system (Thermo Fisher Scientific, Inc., Waltham, MA, USA). Subsequently, LN229 and CHO-K1, which stably overexpressed EphB6 with C-terminal Myc-DDK tags (hereafter described as LN229/EphB6 and CHO/EphB6, respectively) were stained with an anti-EphB6 mAb (clone T49-25; BioLegend, San Diego, CA, USA) and sorted using the SH800 cell sorter (Sony corp., Tokyo, Japan), followed by cultivation in a medium containing 0.5 mg/mL of G418 (Nacalai Tesque, Inc., Kyoto, Japan). The other Eph receptors-overexpressed CHO-K1 (e.g., CHO/EphA1) cell lines and cell culture conditions were previously described [42,43].
2.2. Antibodies
An anti-human EPHB6 mAb (clone T49-25, mouse IgG1, kappa) was purchased from BioLegend. An anti-DYKDDDDK (clone 1E6) mAb was purchased from Wako. An anti-isocitrate dehydrogenase 1 (IDH1) mAb (clone RcMab-1) was developed previously in our lab [44]. A secondary Alexa Fluor 488-conjugated anti-mouse IgG was purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). Secondary horseradish peroxidase-conjugated anti-mouse IgG and anti-rat IgG were obtained from Agilent Technologies Inc. (Santa Clara, CA, USA) and Merck KGaA (Darmstadt, Germany), respectively.
2.3. Development of Hybridomas
For developing anti-EphB6 mAbs, two 6-week-old female BALB/cAJcl mice, purchased from CLEA Japan (Tokyo, Japan), were immunized intraperitoneally with 1 × 108 cells/mouse of LN229/EphB6. The LN229/EphB6 cells as immunogen were harvested after brief exposure to 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). Alhydrogel adjuvant 2% (InvivoGen, San Diego, CA, USA) was added as an adjuvant in the first immunization. Three additional injections of 1 × 108 cells/mouse of LN229/EphB6 were administered intraperitoneally without an adjuvant addition every week. A final booster injection was performed with 1 × 108 cells/mouse of LN229/EphB6 intraperitoneally two days before harvesting splenocytes from mice. We conducted cell-fusion of the harvested splenocytes from immunized mice with P3U1 cells using polyethylene glycol 1500 (PEG1500; Roche Diagnostics, Indianapolis, IN, USA) under heated conditions.
Hybridomas were cultured in the RPMI-1640 medium supplemented as shown above, with additional supplements included hypoxanthine, aminopterin, and thymidine (HAT; Thermo Fisher Scientific, Inc.), 5% BriClone (NICB, Dublin, Ireland), and 5 μg/mL of Plasmocin (InvivoGen) into the medium. The hybridoma supernatants were screened by flow cytometry using CHO/EphB6 and parental CHO-K1 cells. The hybridoma supernatant, containing Eb6Mab-3 in serum free-medium, was filtrated and purified using Ab-Capcher Extra (ProteNova, Kagawa, Japan).
2.4. Flow Cytometric Analysis
Cells were harvested using 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.) or 1 mM EDTA. Subsequently, cells were washed with 0.1% bovine serum albumin in phosphate-buffered saline (PBS) and treated with primary mAbs for 30 min at 4°C. Afterward, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG (1:1000) following the collection of fluorescence data using the SA3800 Cell Analyzer (Sony Corp.).
2.5. Determination of Dissociation Constant (KD) by Flow Cytometry
CHO/EphB6 and DLD-1 cells were suspended in 100 μL serially diluted Eb6Mab-3 (50 µg/mL to 0.003 µg/mL for CHO/EphB6, 100 µg/mL to 0.006 µg/mL for DLD-1) and T49-25 (50 µg/mL to 0.003 µg/mL) after which Alexa Fluor 488-conjugated anti-mouse IgG (1:200) was added. Fluorescence data were subsequently collected using the SA3800 Cell Analyzer, following the calculation of the dissociation constant (KD) by fitting the binding isotherms into the built-in one-site binding model in GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA, USA).
2.6. Western Blot Analysis
Cell lysates (10 µg) were boiled in sodium dodecyl sulfate (SDS) sample buffer (Nacalai Tesque, Inc.). Proteins were electrophoresed on 5%–20% polyacrylamide gels (Wako) and transferred onto polyvinylidene difluoride (PVDF) membranes (Merck KGaA). After blocking with 4% non-fat milk (Nacalai Tesque, Inc.), PVDF membranes were incubated with 5 μg/mL of Eb6Mab-3, 2.5 μg/mL of T49-25, 1 μg/mL of an anti-IDH1 mAb (clone RcMab-1), or 0.5 μg/mL anti-DYKDDDDK (clone 1E6, Wako) mAb, followed by incubation with horseradish peroxidase-conjugated anti-mouse IgG (1:2000; Agilent Technologies Inc.) or anti-rat IgG (1:10000; Merck KGaA). Chemiluminescence signals were developed using ImmunoStar LD (Wako) and imaged with a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
3. Results
3.1. Development of Anti-EphB6 mAbs Using the CBIS Method
To establish anti-EphB6 mAbs, we employed the CBIS method using EphB6-overexpressed cells. Anti-EphB6 mAbs-producing hybridomas were screened by using flow cytometry (Figure 1). Two female BALB/cAJcl mice were intraperitoneally immunized with LN229/EphB6 (1 × 108 cells/time/mouse) every week, a total of 5 times. Subsequently, mouse splenocytes and P3U1 cells were fused by PEG1500. Hybridomas were seeded into 96-well plates, after which the flow cytometric screening was conducted to select CHO/EphB6-reactive and parental CHO-K1-nonreactive supernatants of hybridomas. We obtained some highly CHO/EphB6-reactive supernatants of hybridomas. We finally established the highly sensitive clone Eb6Mab-3 (mouse IgG1, kappa) by limiting dilution and additional analysis.
3.2. Evaluation of Antibody Reactivity Using Flow Cytometry
Flow cytometric analysis was conducted using Eb6Mab-3 and commercially available anti-EphB6 mAb (T49-25) against CHO-K1, CHO/EphB6, and DLD-1 cells. EphB6 is frequently expressed in colorectal cancer [19]. Results indicated that Eb6Mab-3 and T49-25 recognized CHO/EphB6 (Figure 2A) and DLD-1 (Figure 2C) dose-dependently. Reactivity is almost the same between Eb6Mab-3 and T49-25 to CHO/EphB6 (Figure 2A). Eb6Mab-3 showed slightly higher reactivity than T49-25 at 1 µg/mL of mAbs to DLD-1 (Figure 2C). Neither Eb6Mab-3 nor T49-25 reacted with parental CHO-K1 cells (Figure 2B) even at a concentration of 10 µg/mL. Thus, Eb6Mab-3 can detect exogenously and endogenously expressing EphB6 specifically in flow cytometry.
3.3. Specificity of Eb6Mab-3 to Eph Receptor-Overexpressed CHO-K1 Cells
We have established the cell lines of all Eph receptor-overexpressed CHO-K1 cells, EphA1 to A8, A10, B1 to B4, and B6, respectively. Using the 14 cell lines, the specificity of Eb6Mab-3 was analyzed. As shown in Figure 3, 10 µg/mL of Eb6Mab-3 potently recognized CHO/EphB6. Weak recognition to CHO/EphB2 by Eb6Mab-3 was observed.
3.4. Calculation of the Binding Affinity of Anti-EphB6 mAbs Using Flow Cytometry
The binding affinity of Eb6Mab-3 and T49-25 were assessed with exogenously EphB6-expressed CHO/EphB6 and endogenously EphB6-expressing DLD-1 using flow cytometry. The results indicated that the KD values of Eb6Mab-3 for CHO/EphB6 and DLD-1 were 3.8×10-8 M and 1.9×10-9 M, respectively (Figure 4). The KD values of T49-25 for CHO/EphB6 and DLD-1 were 1.5×10-8 M and 1.3×10-8 M, respectively (Figure 5). Although there was no noticeable difference in binding affinity for CHO/EphB6 between Eb6Mab-3 and T49-25, Eb6Mab-3 showed higher binding affinity for DLD-1 than T49-25. These results demonstrate that Eb6Mab-3 can recognize EphB6 with moderate to high affinity to EphB6 on cells.
3.5. Western Blot Analyses Using Anti-EphB6 mAbs
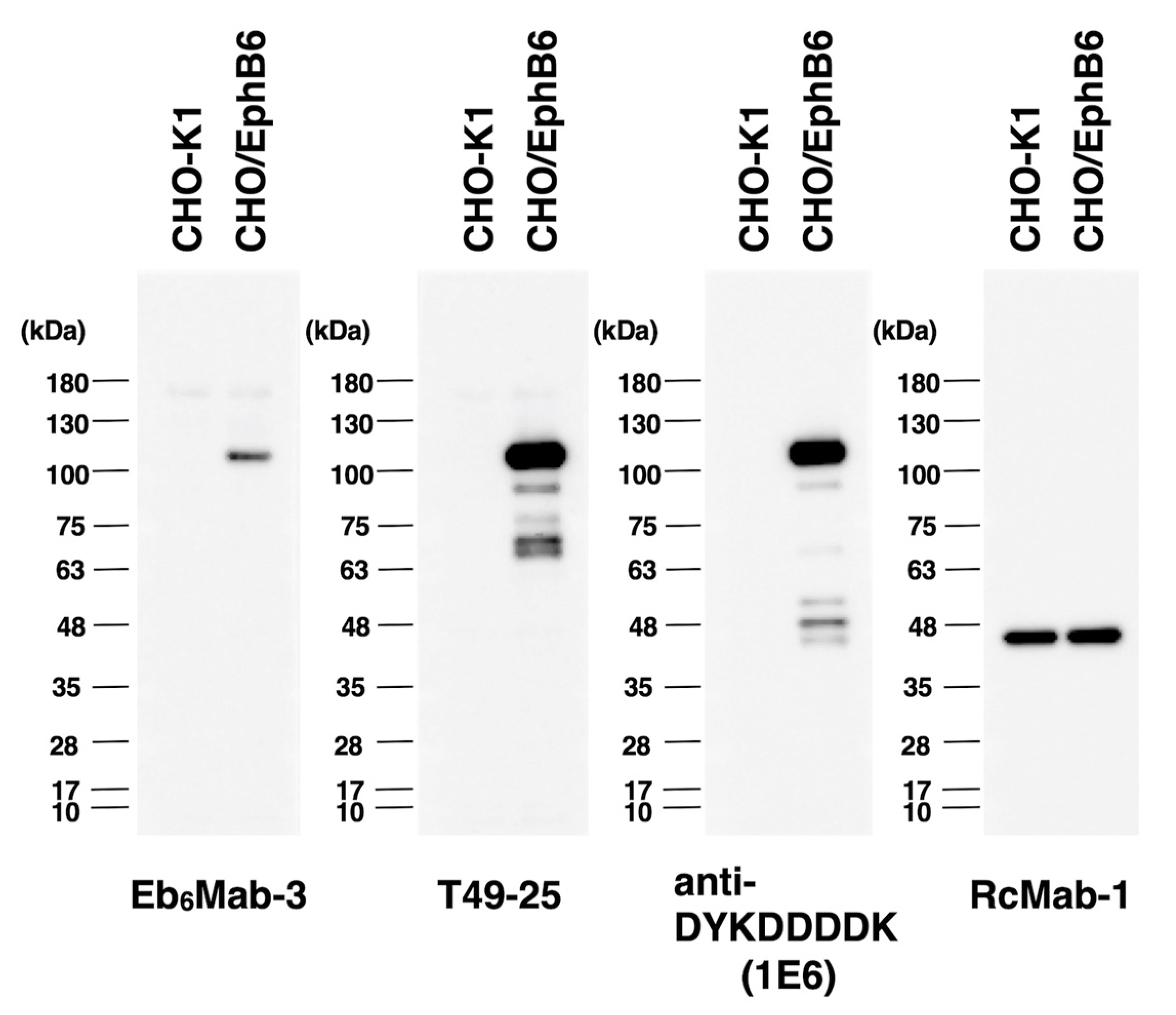
We investigated whether Eb6Mab-3 can be used for western blot analysis by analyzing CHO-K1 and EphB6-overexpressed CHO/EphB6 cell lysates. The estimated molecular weight of EphB6 protein is 110-kDa. As shown in Figure 6, Eb6Mab-3 could detect EphB6 as the band around 100 to 130-kDa (EphB6 + Myc-DDK tags) in CHO/EphB6 cell lysates, while no band was detected in parental CHO-K1 cells. Another anti-EphB6 mAb (clone T49-25) could clearly detect EphB6 as the band around 100 to 130-kDa in CHO/EphB6 cell lysates. In western blot, T49-25 appears to detect EphB6 more sensitively than Eb6Mab-3. An anti-DYKDDDDK mAb was used as a positive control and could detect a band of the same position in CHO/EphB6 cell lysates. An anti-IDH1 mAb (clone RcMab-1) was used for internal control. These results indicate that Eb6Mab-3 can detect EphB6 in EphB6-overexpressing cells in western blot analyses.
Further investigation was conducted to explore other applications, such as immunohistochemistry. Unfortunately, Eb6Mab-3 could not detect EphB6 by immunohistochemistry using cell blocks of CHO/EphB6 (Figure S1).
4. Discussion
Approximately 10% of the protein kinases lack conserved amino acids in the kinase domain, which are required for its activation [45,46]. EphB6 has been identified as one of the RTKs without kinase activity [47]. However, EphB6 expresses numerous tissues and cells to maintain physiological homeostasis, including kidney [48], vascular smooth muscle [49], and T cells [38,50]. The high expression of EphB6 has been observed in the thymus, pancreas, and brain [51,52]. Many reports have demonstrated that EphB6 is fully functional, but its kinase activity is undetected. Human epidermal growth factor receptor 3 (HER3) is one of the most well-known pseudokinases, which belongs to the human epidermal growth factor receptor (EGFR) family [53,54]. HER3 lacks kinase activity but plays pivotal roles such as cell proliferation, physiological homeostasis, and tumor development by forming a heterodimer with human epidermal growth factor receptor 2 (HER2) or EGFR [55,56]. The clinical trial of patritumab deruxtecan (HER3-DXd), HER3 targeting antibody-drug conjugates, is ongoing in locally advanced or metastatic NSCLC with EGFR mutations [57]. Therefore, pseudokinases may become therapeutic targets for cancer. EphB6 has been noted the contribution of oncogenic role in various cancer types, including lung [58], colon [19,59], and breast cancers [60]. Interestingly, crosstalk between EphB6 and EGFR cooperates in cancer progression [61]. Another group indirectly suggests a possible interaction between EphB6 and HER2 by imaging-based analysis [62]. These findings provide further functional expansion for EphB6 and enhance the potential of EphB6 as a therapeutic target for cancer. Furthermore, TK-independent regulation of EphA2, a non-pseudokinase, mediates tumor aggressiveness, including metastasis, invasion, and poor prognosis [63,64]. Kinase activity-independent regulation of Eph receptors might be an attractive target for cancer therapy [65]. In contrast, tumor suppressive functions of EphB6 have been proposed in various cancers. Diminishing expression of EphB6 leads to tumor malignancy [23,35,66]. Further investigation is necessary to clarify these opposing functions of EphB6 in cancer. The management by endocytosis may contribute to modulating the amount of Eph receptors and ephrin expression at the cell surface [67]. In that case, Eb6Mab-3 will contribute to the elucidation of the EphB6-related biological responses by detecting EphB6 with high affinity in flow cytometry (Figure 2, Figure 3, Figure 4 and Figure 5).
Drugs specifically targeting Eph receptors or ephrin ligands have yet to be approved. However, Eph receptors have been widely studied in relation to cancer and considered therapeutic targets [8]. Regarding antibody drugs, the Phase I trial of DS-8895a, a humanized anti-EphA2 defucosylated mAb, has been performed against advanced EphA2-expressing cancer [68,69]. Ifabotuzumab (KB004), an anti-EphA3 mAb, has been tested in advanced hematologic malignancies [70]. The clinical trials targeting EphB6 have not been confirmed to date. However, the development of specific antibodies such as Eb6Mab-3 is also desired to clarify the function of EphB6 in the basic research, diagnosis, and treatment. The expression of EphB6 is inversely related to the expression of the molecules necessary for antitumor immunity, such as chemokine receptors and MHC genes [24]. Eb6Mab-3 might be helpful for the analysis of the immunological cold tumor microenvironment.
Eb6Mab-3 is unsuitable for immunocytochemistry against paraffin-embedded sections of cells (Figure S1). This result suggests that the Eb6Mab-3 recognizes a structural epitope in EphB6. We will determine the epitope of Eb6Mab-3 in a cell-based analysis such as PA scanning and REMAP method [71,72]. Furthermore, we have previously elevated antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) activities by switching isotypes and performing defucosylation in mAbs [73,74,75,76]. Since Eb6Mab-3 is mouse IgG1, which lacks ADCC activity, it will be converted to a mouse IgG2a version to examine the efficacy of antitumor effects in tumor xenograft models in future studies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Tomohiro Tanaka: Investigation, Writing – original draft; Yu Kaneko: Investigation; Haruto Yamamoto: Investigation; Guanjie Li: Investigation; Shiori Fujisawa: Investigation; Hiroyuki Satofuka: Investigation; Keisuke Shinoda: Investigation; Takuya Nakamura: Investigation; Mika K. Kaneko: Conceptualization, Funding acquisition; Hiroyuki Suzuki: Funding acquisition, Writing – review and editing; Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing – review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP24am0521010 (to Y.Kato.), JP24ama121008 (to Y.Kato.), JP24ama221339 (to Y.Kato.), JP24bm1123027 (to Y.Kato.), and JP24ck0106730 (to Y.Kato.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 22K06995 (to H.Suzuki), 21K20789 (to T.T.), 24K11652 (to H.Satofuka), 21K07168 (to M.K.K.), and 22K07224 (to Y.Kato.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Pastore, S.; Mascia, F.; Mariani, V.; Girolomoni, G. The epidermal growth factor receptor system in skin repair and inflammation. J Invest Dermatol 2008;128(6): 1365-1374. [CrossRef]
- Schneider, M.R.; Werner, S.; Paus, R.; Wolf, E. Beyond wavy hairs: the epidermal growth factor receptor and its ligands in skin biology and pathology. Am J Pathol 2008;173(1): 14-24.
- Schneider, M.R.; Wolf, E. The epidermal growth factor receptor ligands at a glance. J Cell Physiol 2009;218(3): 460-466.
- Plum, L.; Schubert, M.; Brüning, J.C. The role of insulin receptor signaling in the brain. Trends Endocrinol Metab 2005;16(2): 59-65. [CrossRef]
- Schlessinger, J. Receptor tyrosine kinases: legacy of the first two decades. Cold Spring Harb Perspect Biol 2014;6(3).
- Bennasroune, A.; Gardin, A.; Aunis, D.; Crémel, G.; Hubert, P. Tyrosine kinase receptors as attractive targets of cancer therapy. Crit Rev Oncol Hematol 2004;50(1): 23-38.
- Templeton, A.J.; Diez-Gonzalez, L.; Ace, O.; et al. Prognostic relevance of receptor tyrosine kinase expression in breast cancer: a meta-analysis. Cancer Treat Rev 2014;40(9): 1048-1055.
- Pasquale, E.B. Eph receptors and ephrins in cancer progression. Nat Rev Cancer 2024;24(1): 5-27. [CrossRef]
- Trinidad, E.M.; Zapata, A.G.; Alonso-Colmenar, L.M. Eph-ephrin bidirectional signaling comes into the context of lymphocyte transendothelial migration. Cell Adh Migr 2010;4(3): 363-367.
- Pasquale, E.B. Eph-ephrin bidirectional signaling in physiology and disease. Cell 2008;133(1): 38-52.
- Edwards, C.M.; Mundy, G.R. Eph receptors and ephrin signaling pathways: a role in bone homeostasis. Int J Med Sci 2008;5(5): 263-272.
- Miao, H.; Wang, B. Eph/ephrin signaling in epithelial development and homeostasis. Int J Biochem Cell Biol 2009;41(4): 762-770.
- McCarron, J.K.; Stringer, B.W.; Day, B.W.; Boyd, A.W. Ephrin expression and function in cancer. Future Oncol 2010;6(1): 165-176.
- Gurniak, C.B.; Berg, L.J. A new member of the Eph family of receptors that lacks protein tyrosine kinase activity. Oncogene 1996;13(4): 777-786.
- Mendrola, J.M.; Shi, F.; Park, J.H.; Lemmon, M.A. Receptor tyrosine kinases with intracellular pseudokinase domains. Biochem Soc Trans 2013;41(4): 1029-1036. [CrossRef]
- Liang, L.Y.; Patel, O.; Janes, P.W.; Murphy, J.M.; Lucet, I.S. Eph receptor signalling: from catalytic to non-catalytic functions. Oncogene 2019;38(39): 6567-6584.
- Munthe, E.; Rian, E.; Holien, T.; et al. Ephrin-B2 is a candidate ligand for the Eph receptor, EphB6. FEBS Lett 2000;466(1): 169-174.
- Liang, L.Y.; Geoghegan, N.D.; Mlodzianoski, M.; et al. Co-clustering of EphB6 and ephrinB1 in trans restrains cancer cell invasion. Commun Biol 2024;7(1): 461.
- Xu, D.; Yuan, L.; Liu, X.; et al. EphB6 overexpression and Apc mutation together promote colorectal cancer. Oncotarget 2016;7(21): 31111-31121.
- El Zawily, A.; McEwen, E.; Toosi, B.; et al. The EphB6 receptor is overexpressed in pediatric T cell acute lymphoblastic leukemia and increases its sensitivity to doxorubicin treatment. Sci Rep 2017;7(1): 14767. [CrossRef]
- Colucci, M.; Trivieri, N.; Mencarelli, G.; et al. A functional role of Ephrin type-B receptor 6 (EPHB6) in T-cell acute lymphoblastic leukemia. Biomark Res 2023;11(1): 92.
- Dong, Y.; Pan, J.; Ni, Y.; et al. High expression of EphB6 protein in tongue squamous cell carcinoma is associated with a poor outcome. Int J Clin Exp Pathol 2015;8(9): 11428-11433.
- Fox, B.P.; Kandpal, R.P. EphB6 receptor significantly alters invasiveness and other phenotypic characteristics of human breast carcinoma cells. Oncogene 2009;28(14): 1706-1713.
- Jia, X.; Zhang, D.; Zhou, C.; et al. Eph receptor B6 shapes a cold immune microenvironment, inhibiting anti-cancer immunity and immunotherapy response in bladder cancer. Front Oncol 2023;13: 1175183.
- Bulk, E.; Yu, J.; Hascher, A.; et al. Mutations of the EPHB6 receptor tyrosine kinase induce a pro-metastatic phenotype in non-small cell lung cancer. PLoS One 2012;7(12): e44591.
- Freywald, A.; Sharfe, N.; Roifman, C.M. The kinase-null EphB6 receptor undergoes transphosphorylation in a complex with EphB1. J Biol Chem 2002;277(6): 3823-3828.
- Truitt, L.; Freywald, T.; DeCoteau, J.; Sharfe, N.; Freywald, A. The EphB6 receptor cooperates with c-Cbl to regulate the behavior of breast cancer cells. Cancer Res 2010;70(3): 1141-1153.
- Akada, M.; Harada, K.; Negishi, M.; Katoh, H. EphB6 promotes anoikis by modulating EphA2 signaling. Cell Signal 2014;26(12): 2879-2884.
- Giaginis, C.; Alexandrou, P.; Poulaki, E.; et al. Clinical Significance of EphB4 and EphB6 Expression in Human Malignant and Benign Thyroid Lesions. Pathol Oncol Res 2016;22(2): 269-275. [CrossRef]
- Mateo-Lozano, S.; Bazzocco, S.; Rodrigues, P.; et al. Loss of the EPH receptor B6 contributes to colorectal cancer metastasis. Sci Rep 2017;7: 43702.
- Liu, J.; Xu, B.; Xu, G.; et al. Reduced EphB6 protein in gastric carcinoma and associated lymph nodes suggests EphB6 as a gastric tumor and metastasis inhibitor. Cancer Biomark 2017;19(3): 241-248.
- Yu, H.; Qin, X.K.; Yin, K.W.; et al. EphB6 deficiency in intestinal neurons promotes tumor growth in colorectal cancer by neurotransmitter GABA signaling. Carcinogenesis 2023;44(8-9): 682-694. [CrossRef]
- Yu, J.; Bulk, E.; Ji, P.; et al. The EPHB6 receptor tyrosine kinase is a metastasis suppressor that is frequently silenced by promoter DNA hypermethylation in non-small cell lung cancer. Clin Cancer Res 2010;16(8): 2275-2283.
- Bailey, C.M.; Kulesa, P.M. Dynamic interactions between cancer cells and the embryonic microenvironment regulate cell invasion and reveal EphB6 as a metastasis suppressor. Mol Cancer Res 2014;12(9): 1303-1313.
- Gu, Y.; Li, F.; Qian, N.; et al. Expression of EphB6 in ovarian serous carcinoma is associated with grade, TNM stage and survival. J Clin Pathol 2016;69(5): 448-453.
- Tang, X.X.; Evans, A.E.; Zhao, H.; et al. High-level expression of EPHB6, EFNB2, and EFNB3 is associated with low tumor stage and high TrkA expression in human neuroblastomas. Clin Cancer Res 1999;5(6): 1491-1496.
- Luo, H.; Yu, G.; Tremblay, J.; Wu, J. EphB6-null mutation results in compromised T cell function. J Clin Invest 2004;114(12): 1762-1773.
- Luo, H.; Yu, G.; Wu, Y.; Wu, J. EphB6 crosslinking results in costimulation of T cells. J Clin Invest 2002;110(8): 1141-1150.
- Yoon, S.; Choi, J.H.; Kim, S.J.; et al. EPHB6 mutation induces cell adhesion-mediated paclitaxel resistance via EPHA2 and CDH11 expression. Exp Mol Med 2019;51(6): 1-12.
- Satofuka H, S.H., Tanaka T, Li G, Kaneko MK, Kato Y. An Anti-Human EphA2 Monoclonal Antibody Ea2Mab-7 Shows High Sensitivity for Flow Cytometry, Western Blot, and Immunohistochemical Analyses. Preprint 2024.
- Ubukata R, S.H., Hirose M, Satofuka H, Tanaka T, Kaneko MK, Kato Y. Establishment of a Highly-sensitive Anti-EphB2 Monoclonal Antibody Eb2Mab-3 for Flow Cytometry. Preprint 2024.
- Nanamiya, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of an Anti-EphB4 Monoclonal Antibody for Multiple Applications Against Breast Cancers. Monoclon Antib Immunodiagn Immunother 2023;42(5): 166-177.
- Tanaka, T.; Yamamoto, H.; Kaneko, Y.; et al. Ea8Mab-9: A Novel Monoclonal Antibody Against Erythropoietin-Producing Hepatocellular Receptor A8 for Flow Cytometry. Preprints 2024. [CrossRef]
- Ikota, H.; Nobusawa, S.; Arai, H.; et al. Evaluation of IDH1 status in diffusely infiltrating gliomas by immunohistochemistry using anti-mutant and wild type IDH1 antibodies. Brain Tumor Pathol 2015;32(4): 237-244.
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002;298(5600): 1912-1934.
- Boudeau, J.; Miranda-Saavedra, D.; Barton, G.J.; Alessi, D.R. Emerging roles of pseudokinases. Trends Cell Biol 2006;16(9): 443-452.
- Strozen, T.G.; Sharpe, J.C.; Harris, E.D.; Uppalapati, M.; Toosi, B.M. The EphB6 Receptor: Kinase-Dead but Very Much Alive. Int J Mol Sci 2021;22(15).
- Ogawa, K.; Wada, H.; Okada, N.; et al. EphB2 and ephrin-B1 expressed in the adult kidney regulate the cytoarchitecture of medullary tubule cells through Rho family GTPases. J Cell Sci 2006;119(Pt 3): 559-570.
- Luo, H.; Wu, Z.; Tremblay, J.; et al. Receptor tyrosine kinase Ephb6 regulates vascular smooth muscle contractility and modulates blood pressure in concert with sex hormones. J Biol Chem 2012;287(9): 6819-6829.
- Freywald, A.; Sharfe, N.; Rashotte, C.; Grunberger, T.; Roifman, C.M. The EphB6 receptor inhibits JNK activation in T lymphocytes and modulates T cell receptor-mediated responses. J Biol Chem 2003;278(12): 10150-10156.
- Matsuoka, H.; Obama, H.; Kelly, M.L.; Matsui, T.; Nakamoto, M. Biphasic functions of the kinase-defective Ephb6 receptor in cell adhesion and migration. J Biol Chem 2005;280(32): 29355-29363.
- Hafner, C.; Schmitz, G.; Meyer, S.; et al. Differential gene expression of Eph receptors and ephrins in benign human tissues and cancers. Clin Chem 2004;50(3): 490-499.
- Mujoo, K.; Choi, B.K.; Huang, Z.; Zhang, N.; An, Z. Regulation of ERBB3/HER3 signaling in cancer. Oncotarget 2014;5(21): 10222-10236.
- Colomba, A.; Claus, J.; Gao, F.; et al. Targeting the HER3 pseudokinase domain with small molecule inhibitors. Methods Enzymol 2022;667: 455-505.
- Pawar, A.B.; Sengupta, D. Resolving the conformational dynamics of ErbB growth factor receptor dimers. J Struct Biol 2019;207(2): 225-233.
- Xie, T.; Lim, S.M.; Westover, K.D.; et al. Pharmacological targeting of the pseudokinase Her3. Nat Chem Biol 2014;10(12): 1006-1012.
- Arter, Z.L.; Nagasaka, M. Spotlight on Patritumab Deruxtecan (HER3-DXd) from HERTHENA Lung01. Is a Median PFS of 5.5 Months Enough in Light of FLAURA-2 and MARIPOSA? Lung Cancer (Auckl) 2024;15: 115-121.
- Yu, J.; Bulk, E.; Ji, P.; et al. The kinase defective EPHB6 receptor tyrosine kinase activates MAP kinase signaling in lung adenocarcinoma. Int J Oncol 2009;35(1): 175-179.
- Wang, J.; Zhang, Y.; Ma, J.; et al. Determining the effects of Ephrin Type B Receptor 6 and Type A Receptor 3 on facilitating colorectal epithelial cell malignant transformation. Neoplasma 2021;68(5): 955-964.
- Toosi, B.M.; El Zawily, A.; Truitt, L.; et al. EPHB6 augments both development and drug sensitivity of triple-negative breast cancer tumours. Oncogene 2018;37(30): 4073-4093.
- Hanover, G.; Vizeacoumar, F.S.; Banerjee, S.L.; et al. Integration of cancer-related genetic landscape of Eph receptors and ephrins with proteomics identifies a crosstalk between EPHB6 and EGFR. Cell Rep 2023;42(7): 112670.
- Kennedy, S.P.; Han, J.Z.R.; Portman, N.; et al. Targeting promiscuous heterodimerization overcomes innate resistance to ERBB2 dimerization inhibitors in breast cancer. Breast Cancer Res 2019;21(1): 43.
- Zhou, Y.; Yamada, N.; Tanaka, T.; et al. Crucial roles of RSK in cell motility by catalysing serine phosphorylation of EphA2. Nat Commun 2015;6: 7679.
- Shi, X.; Lingerak, R.; Herting, C.J.; et al. Time-resolved live-cell spectroscopy reveals EphA2 multimeric assembly. Science 2023;382(6674): 1042-1050.
- Zhou, Y.; Sakurai, H. Emerging and Diverse Functions of the EphA2 Noncanonical Pathway in Cancer Progression. Biol Pharm Bull 2017;40(10): 1616-1624.
- Hafner, C.; Bataille, F.; Meyer, S.; et al. Loss of EphB6 expression in metastatic melanoma. Int J Oncol 2003;23(6): 1553-1559.
- Pitulescu, M.E.; Adams, R.H. Eph/ephrin molecules--a hub for signaling and endocytosis. Genes Dev 2010;24(22): 2480-2492.
- Shitara, K.; Satoh, T.; Iwasa, S.; et al. Safety, tolerability, pharmacokinetics, and pharmacodynamics of the afucosylated, humanized anti-EPHA2 antibody DS-8895a: a first-in-human phase I dose escalation and dose expansion study in patients with advanced solid tumors. J Immunother Cancer 2019;7(1): 219.
- Gan, H.K.; Parakh, S.; Lee, F.T.; et al. A phase 1 safety and bioimaging trial of antibody DS-8895a against EphA2 in patients with advanced or metastatic EphA2 positive cancers. Invest New Drugs 2022;40(4): 747-755.
- Swords, R.T.; Greenberg, P.L.; Wei, A.H.; et al. KB004, a first in class monoclonal antibody targeting the receptor tyrosine kinase EphA3, in patients with advanced hematologic malignancies: Results from a phase 1 study. Leuk Res 2016;50: 123-131.
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon Antib Immunodiagn Immunother 2021;40(4): 162-167.
- Okada, Y.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Epitope Mapping of an Anti-Mouse CD39 Monoclonal Antibody Using PA Scanning and RIEDL Scanning. Monoclon Antib Immunodiagn Immunother 2024;43(2): 44-52.
- Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. A Cancer-Specific Monoclonal Antibody against Podocalyxin Exerted Antitumor Activities in Pancreatic Cancer Xenografts. Int J Mol Sci 2023;25(1).
- Tanaka, T.; Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. A Cancer-Specific Anti-Podoplanin Monoclonal Antibody, PMab-117-mG(2a) Exerts Antitumor Activities in Human Tumor Xenograft Models. Cells 2024;13(22).
- Tanaka, T.; Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. Antitumor activities against breast cancers by an afucosylated anti-HER2 monoclonal antibody H(2) Mab-77-mG(2a) -f. Cancer Sci 2024;115(1): 298-309.
- Arimori, T.; Mihara, E.; Suzuki, H.; et al. Locally misfolded HER2 expressed on cancer cells is a promising target for development of cancer-specific antibodies. Structure 2024;32(5): 536-549.e535.
Figure 1.
A schematic depiction of anti-EphB6 mAbs development by CBIS method. The simplified procedure of antibody development using the CBIS method. (A) LN229/hEphB6 cells were intraperitoneally immunized into two female mice. (B) The spleen cells from antigen-immunized mice were fused with myeloma cells, P3U1, by PEG1500. (C) The culture supernatants of hybridoma were screened by flow cytometry using CHO-K1 and CHO/EphB6 to select EphB6-specific mAb-producing hybridomas. (D) Single hybridoma clones were obtained by limiting dilution, followed by additional screening. Finally, Eb6Mab-3 (mouse IgG1, kappa) was established.
Figure 1.
A schematic depiction of anti-EphB6 mAbs development by CBIS method. The simplified procedure of antibody development using the CBIS method. (A) LN229/hEphB6 cells were intraperitoneally immunized into two female mice. (B) The spleen cells from antigen-immunized mice were fused with myeloma cells, P3U1, by PEG1500. (C) The culture supernatants of hybridoma were screened by flow cytometry using CHO-K1 and CHO/EphB6 to select EphB6-specific mAb-producing hybridomas. (D) Single hybridoma clones were obtained by limiting dilution, followed by additional screening. Finally, Eb6Mab-3 (mouse IgG1, kappa) was established.
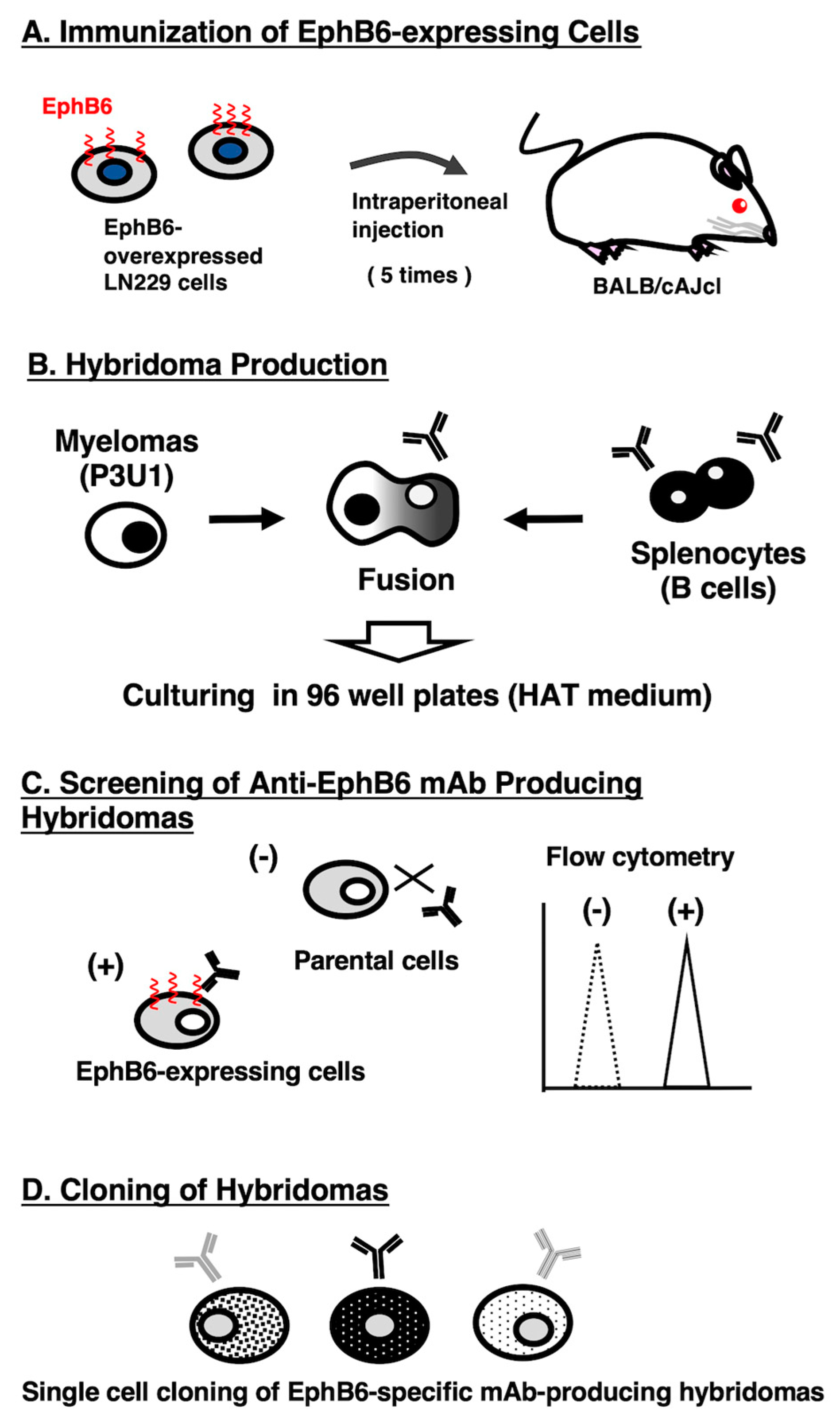
Figure 2.
Flow cytometric analysis of anti-EphB6 mAbs. CHO/EphB6 (A), CHO-K1 (B), and DLD-1 (C) cells were treated with 0.01–10 µg/mL of Eb6Mab-3 or T49-25 (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
Figure 2.
Flow cytometric analysis of anti-EphB6 mAbs. CHO/EphB6 (A), CHO-K1 (B), and DLD-1 (C) cells were treated with 0.01–10 µg/mL of Eb6Mab-3 or T49-25 (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
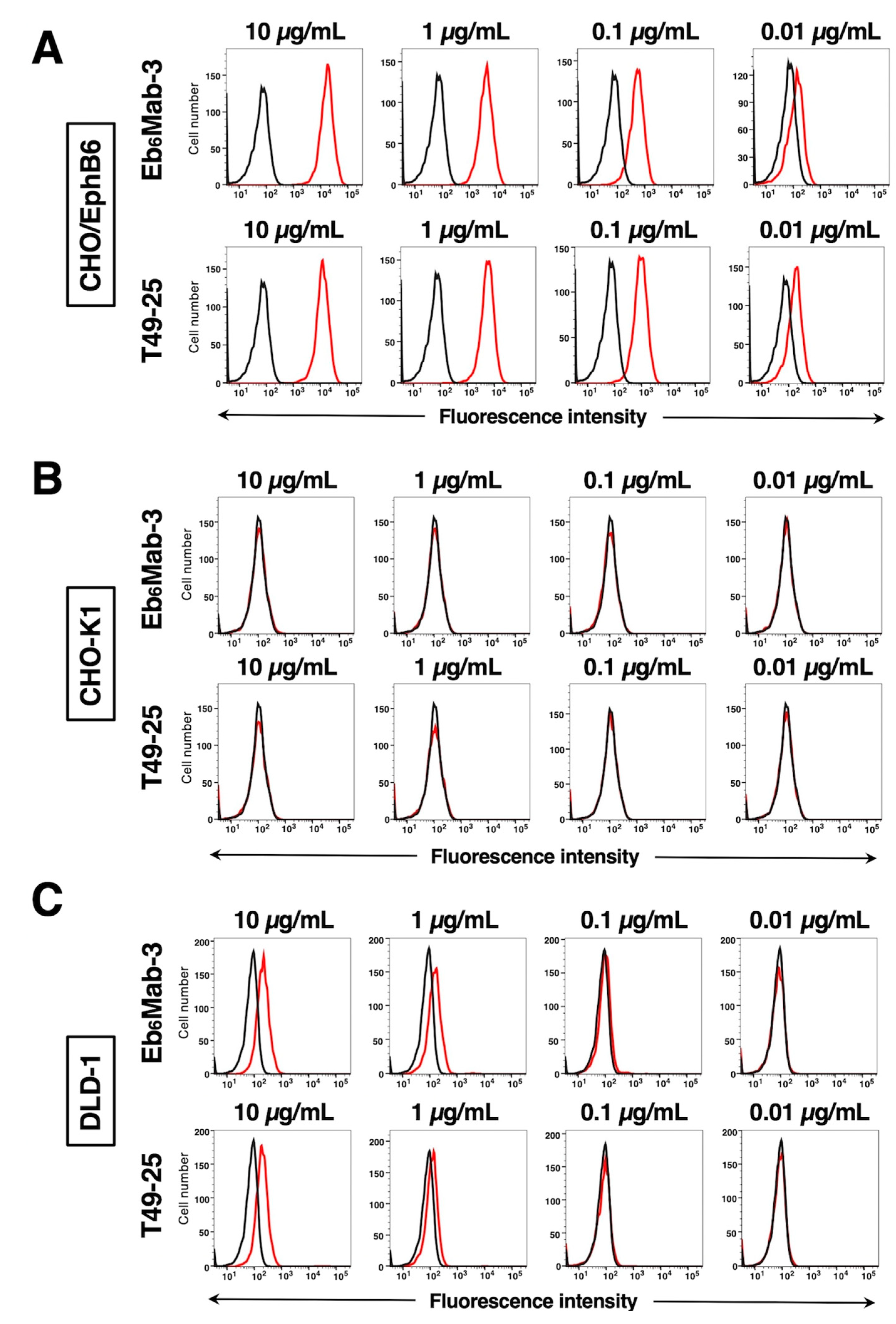
Figure 3.
Flow cytometry of Eb6Mab-3 in Eph receptor-expressed CHO-K1 cells. CHO-K1 cells which overexpressed each of the fourteen Eph receptors were treated with 10 µg/mL of Eb6Mab-3 (red line) or control blocking buffer (black line) followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488.
Figure 3.
Flow cytometry of Eb6Mab-3 in Eph receptor-expressed CHO-K1 cells. CHO-K1 cells which overexpressed each of the fourteen Eph receptors were treated with 10 µg/mL of Eb6Mab-3 (red line) or control blocking buffer (black line) followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488.
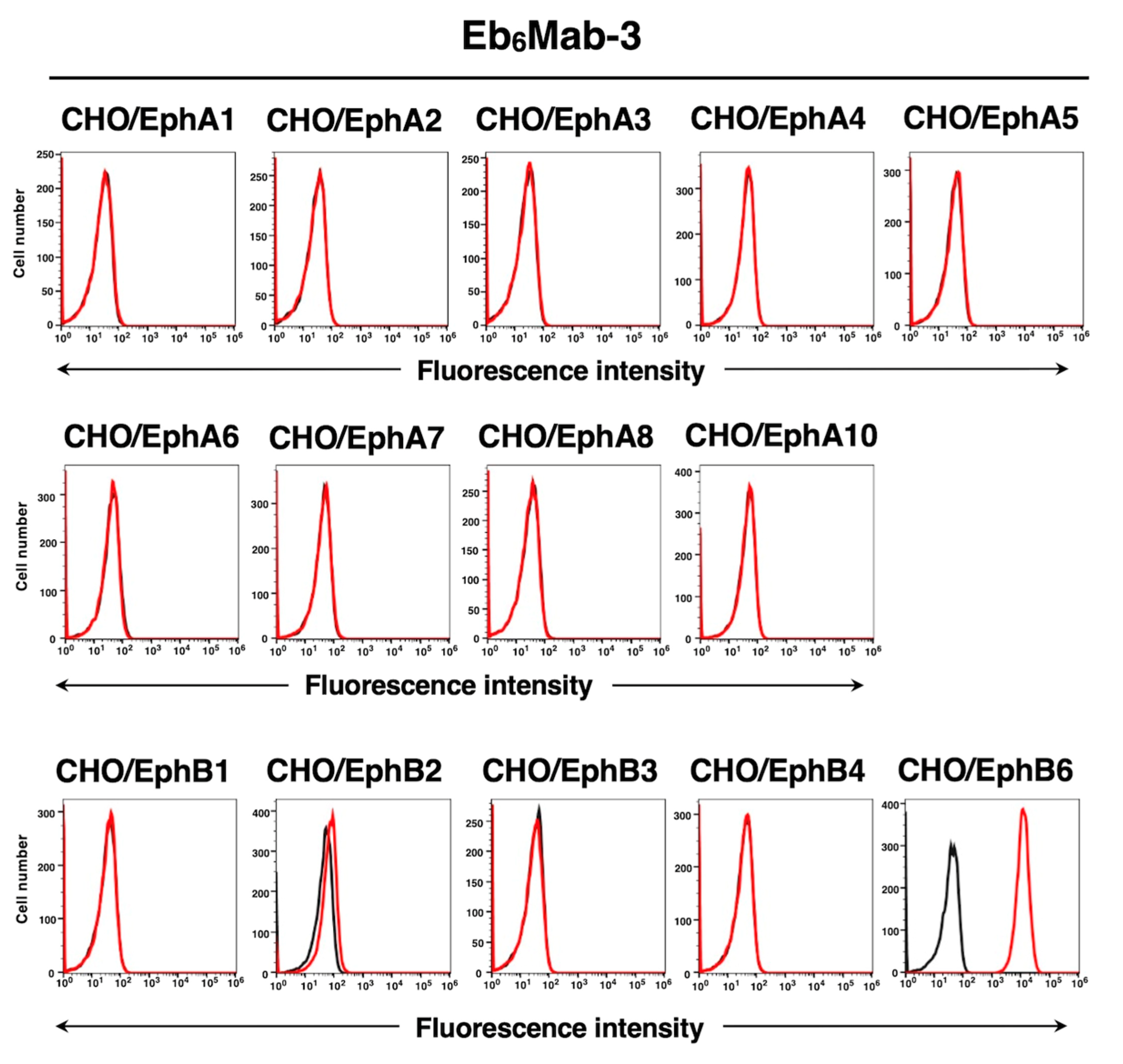
Figure 4.
Determination of the binding affinity of Eb6Mab-3. CHO/EphB6 (A) and DLD-1 (B) cells were suspended in 100 µL of serially diluted 50 µg/mL to 0.003 µg/mL of Eb6Mab-3 for CHO/EphB6, 100 µg/mL to 0.006 µg/mL of Eb6Mab-3 for DLD-1. Then, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG. Subsequently, the geometric mean values from fluorescence data were collected using the SA3800 Cell Analyzer, following the calculation of the KD by GraphPad PRISM 6 software.
Figure 4.
Determination of the binding affinity of Eb6Mab-3. CHO/EphB6 (A) and DLD-1 (B) cells were suspended in 100 µL of serially diluted 50 µg/mL to 0.003 µg/mL of Eb6Mab-3 for CHO/EphB6, 100 µg/mL to 0.006 µg/mL of Eb6Mab-3 for DLD-1. Then, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG. Subsequently, the geometric mean values from fluorescence data were collected using the SA3800 Cell Analyzer, following the calculation of the KD by GraphPad PRISM 6 software.
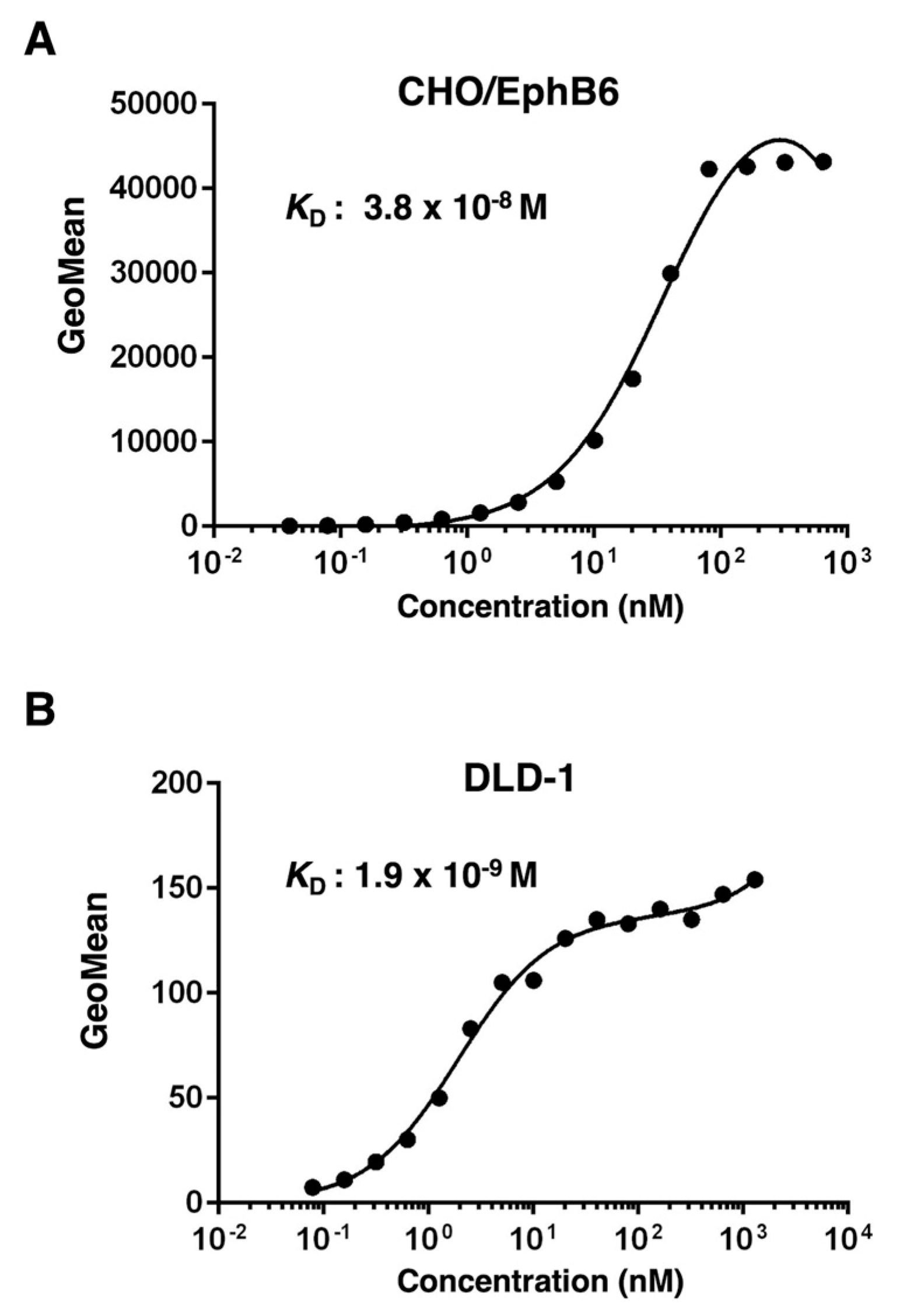
Figure 5.
Determination of the binding affinity of T49-25. CHO/EphB6 (A) and DLD-1 (B) cells were suspended in 100 µL of serially diluted 50 µg/mL to 0.003 µg/mL of T49-25. Then, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG. Subsequently, the geometric mean values from fluorescence data were collected using the SA3800 Cell Analyzer, following the calculation of the KD by GraphPad PRISM 6 software.
Figure 5.
Determination of the binding affinity of T49-25. CHO/EphB6 (A) and DLD-1 (B) cells were suspended in 100 µL of serially diluted 50 µg/mL to 0.003 µg/mL of T49-25. Then, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG. Subsequently, the geometric mean values from fluorescence data were collected using the SA3800 Cell Analyzer, following the calculation of the KD by GraphPad PRISM 6 software.
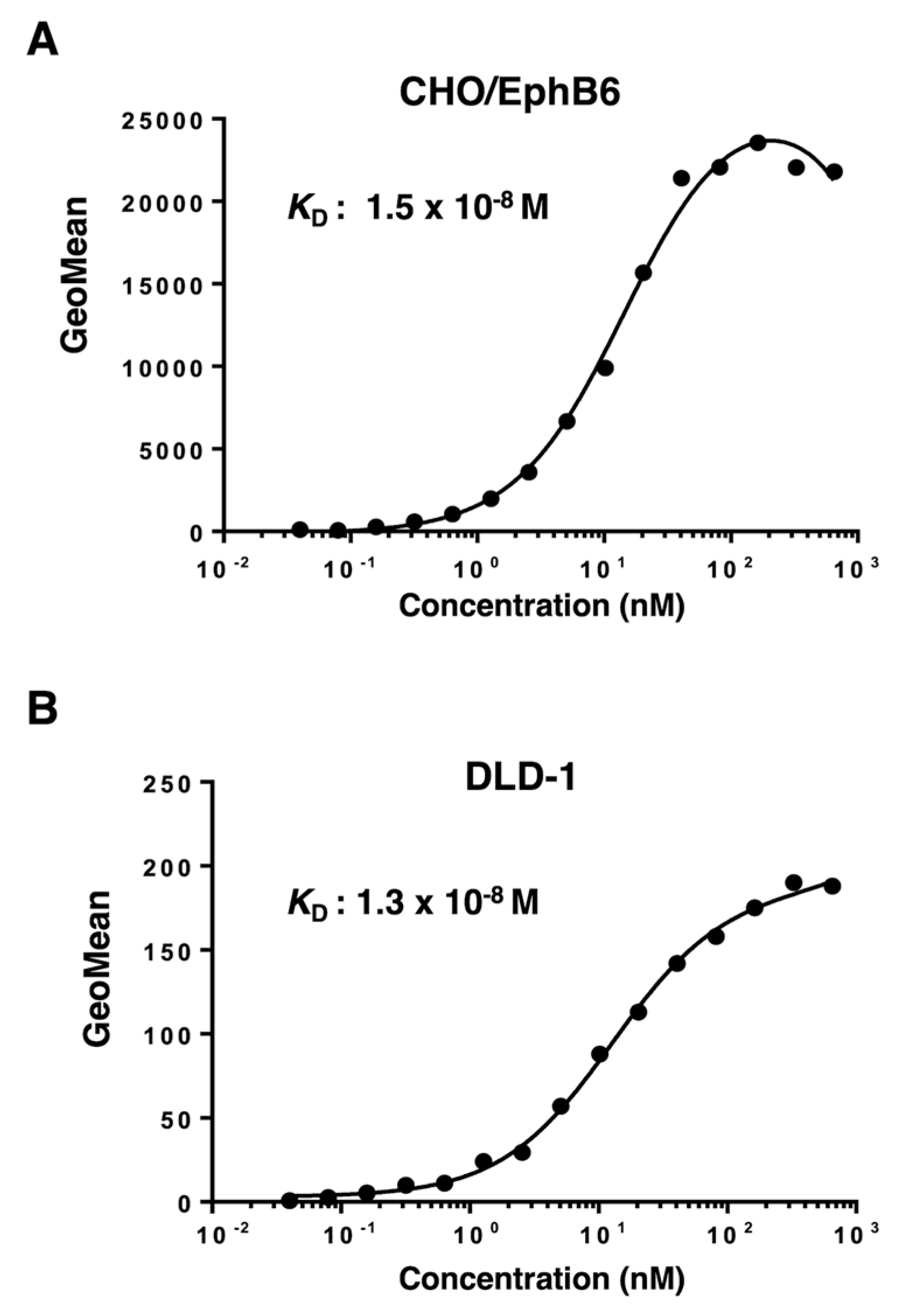
Figure 6.
The detection of EphB6 by western blot analysis. Cell lysates of CHO-K1 and CHO/EphB6 (10 μg/lane) were electrophoresed and transferred onto PVDF membranes. The membranes were incubated with 5 μg/mL of Eb6Mab-3, 2.5 μg/mL of T49-25, 0.5 μg/mL of an anti-DYKDDDDK (clone 1E6) mAb, and 1 μg/mL of RcMab-1 and subsequently with horseradish peroxidase-conjugated anti-mouse or anti-rat immunoglobulins.
Figure 6.
The detection of EphB6 by western blot analysis. Cell lysates of CHO-K1 and CHO/EphB6 (10 μg/lane) were electrophoresed and transferred onto PVDF membranes. The membranes were incubated with 5 μg/mL of Eb6Mab-3, 2.5 μg/mL of T49-25, 0.5 μg/mL of an anti-DYKDDDDK (clone 1E6) mAb, and 1 μg/mL of RcMab-1 and subsequently with horseradish peroxidase-conjugated anti-mouse or anti-rat immunoglobulins.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



