
Submitted:
12 December 2024
Posted:
12 December 2024
You are already at the latest version
Abstract
Erythropoietin-producing hepatocellular receptor A8 (EphA8) is a type I transmembrane protein that belongs to the largest Eph family among receptor tyrosine kinases. By binding to its membrane-bound ephrin-A or ephrin-B ligands on adjacent cells, Eph receptors form complexes and mediate bidirectional signaling activities, triggering cell-cell adhesion and repulsion. Increased expression of EphA8 correlates with poor prognosis in some types of cancer. Therefore, developing sensitive monoclonal antibodies (mAbs) for EphA8 has been desired for treatment, diagnosis, and further basic research. In particular, there are no anti-EphA8 mAbs that can be used for flow cytometry. In this study, we established a novel specific and sensitive anti-human EphA8 mAb, which applies to flow cytometry, clone Ea8Mab-9 (mouse IgG1, kappa), by the Cell-Based Immunization and Screening (CBIS) method. Ea8Mab-9 reacted with EphA8-overexpressed Chinese hamster ovary-K1 cells (CHO/EphA8) and EphA8-overexpressed LN229 glioblastoma cells (LN229/EphA8) in flow cytometry. Ea8Mab-9 did not recognize other members of Eph receptors. Furthermore, Ea8Mab-9demonstrated a high binding affinity for CHO/EphA8 and LN229/EphA8 with dissociation constants of 1.3×10-9 M and 1.6×10-9 M, respectively. Ea8Mab-9, established by the CBIS method, could be useful for analyzing the EphA8-related biological response using flow cytometry with high affinity and specificity.
Keywords:
EphA8
; CBIS method
; monoclonal antibody
; flow cytometry
1. Introduction
Erythropoietin-producing hepatocellular (Eph) receptors are the most prominent family of receptor tyrosine kinases (RTKs) and regulate tissue homeostasis, including cell proliferation and migration, tissue remodeling, angiogenesis, axon guidance, and synaptic plasticity in the nervous system [1,2,3]. Eph receptors are classified into EphA and EphB subfamilies according to the sequence homology and binding mode to membrane-bound ephrin ligands. The EphA subfamily includes nine molecules, such as EphA1 to EphA8 and EphA10. The EphB subfamily consists of five molecules, such as EphB1 to EphB4 and EphB6. The eight ephrin ligands have been identified, including glycosylphosphatidylinositol (GPI)-anchored ephrin A1 to A5 and transmembrane ephrin B1 to B3. Following the binding to receptors, forward signals are triggered on the receptor side, and reverse signals are generated on the ligand side, controlling various biological homeostasis [3].
EphA8 gene was identified from rat and human DNA encoding as eek (Eph- and elk-related kinase, EEK) in 1991 [4]. Another group has cloned mouse EphA8 molecule in 1997 [5]. EphA8 is one of the members of RTKs; the regulatory mechanism is thought to be based on tyrosine kinase activity. Phosphorylation of Tyr-615 in the EphA8 juxtamemembrane domain mediated a strong association with the SH2 domain of Fyn, a member of Src TKs [6]. Tyr-838 phosphorylation in the EphA8 kinase domain modulates Fyn binding to Tyr-615, resulting in attenuating cell adhesion by cellular cytoskeletal modifications [6]. Interestingly, tyrosine kinase activity-independent functions are also emerging. Ephrin A5-induced EphA8-integrin interaction is promoted by p110γ PI3K in an EphA8 tyrosine kinase-independent manner [7]. Like representative growth factor receptors such as epidermal growth factor receptor (EGFR), Eph receptors play a role in cell proliferation primarily through forward signaling [8,9]. Not only cell-cell attachment and cell motility, EphA8 controls organ development and axon growth [10,11]. EphA8 induces caspase-dependent apoptotic cell death of ephrin A5-expressing neural epithelial cells during early brain development [10]. Loss of EphA8 disrupts axon guidance during the nervous system in mammals [11]. Furthermore, EphA8 facilitates neurite outgrowth by sustaining MAPK activity in neuronal cells [12].
EphA8 expression has also been reported to be associated with cancer. EphA8 upregulation is observed in various cancers, including oral tongue squamous cell carcinoma (OTSCC) [13], ovarian cancer [14], gastric cancer [15], and breast cancer [16]. EphA8 and ephrin A5 contribute to the invasiveness of stem cells isolated from MDA-MB-231, a triple-negative invasive breast cancer cell line [17]. In contrast, tumor suppressor functions of EphA8 have also been proposed. Reducing the expression of miR-10a, a promoter of cancer invasion, increases EphA8 expression and suppresses cancer progression, such as colorectal cancer and glioma [18,19]. Further research is necessary to clarify the role of EphA8 in promoting or suppressing cancer functions.
Previously, we have developed various monoclonal antibodies (mAbs) against human Eph receptors, including EphA2 [20], EphB2 [21], and EphB4 [22], by the Cell-Based Immunization and Screening (CBIS) method. This method can efficiently obtain a wide variety of antibodies that recognize modifications and three-dimensional structures of extracellular domains of membrane protein. Since flow cytometry is used for high-throughput screening in the CBIS method, mAbs that can be used for that application are the first choice. However, anti-EphA8 mAbs suitable for flow cytometry are not yet commercially available.
In this study, we have successfully established an anti-human EphA8 mAb (clone Ea8Mab-9) for flow cytometry using the CBIS method.
2. Materials and Methods
2.1. Cell Lines and Stable Transfectants
LN229, Chinese hamster ovary (CHO)-K1, and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (Manassas, VA). The cDNA encoding human EphA8 (Accession No. NM_020526; Catalog No.: RC220352) was purchased from OriGene Technologies, Inc. (Rockville, MD, USA). The open reading frame of EphA8, excluding the signal sequence, interleukin 2-signal sequence, and PA16 tag [23] or MAP16 tag [24] of the N-terminus of EphA8, were subcloned into the pCAG-Ble vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) using the In-Fusion HD Cloning Kit (Takara Bio, Inc., Shiga, Japan). The plasmid was transfected into the cell lines using a Neon transfection system (Thermo Fisher Scientific, Inc., Waltham, MA, USA). Subsequently, LN229 and CHO-K1, stably overexpressed EphA8, which deleted amino acids 1st to 27th with N-terminal MAP16 tags (from now on described as LN229/EphA8 and CHO/EphA8, respectively) and LN229 which stably overexpressed EphA8 which deleted amino acids 1st to 27th with N-terminal PA16 tag (from now on described as LN229/PA16-EphA8) were established using a cell sorter (SH800; Sony Corp., Tokyo, Japan) by staining tag systems.
Other Eph receptor cDNAs including EphA1 (Catalog No.: RC213689, Accession No.: NM_005232), EphA4 (Catalog No.: RC211230, Accession No.: NM_004438), EphA5 (Catalog No.: RC213206, Accession No.: NM_004439), EphA6 (Catalog No.: RC223510, Accession No.: NM_001080448), EphA7 (Catalog No.: RC226293, Accession No.: NM_004440), EphA10 (Catalog No.: RC218374, Accession No.: NM_001099439) EphB1 (Catalog No.: RC214301, Accession No.: NM_004441), EphB2 (Catalog No.: RC223882, Accession No.: NM_004442), EphB6 (Catalog No.: RC229404, Accession No.: NM_004445), were purchased from OriGene Technologies, Inc. EphA2 (Catalog No.: HGY095959, Accession No.: NM_004431), EphA3 (Catalog No.: HGY053437, Accession No.: NM_005233), and EphB3 (Catalog No.: HGX039581, Accession No.: NM_004443) cDNAs were purchased from RIKEN DNA Bank (Ibaraki, Japan).
EphA2 and EphB3 cDNAs were cloned into a pCAGzeo vector (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). EphB6 cDNA was cloned into a pCMV6 vector. EphA1 cDNA was cloned into a pCAGzeo-ssnPA vector. EphA3, EphA4, EphA5, EphA6, EphA7, EphA8, EphA10, and EphB1 cDNA were cloned into a pCAGzeo_ssnPA16 vector.
The plasmids were also transfected into CHO-K1 cells and stable transfectants were established by staining with an anti-EphA2 mAb (clone SHM16; BioLegend, San Diego, CA, USA), an anti-EphB3 mAb (clone 647354; R&D Systems Inc., Minneapolis, MN, USA), an anti-EphB6 mAb (clone T49-25; BioLegend), and an anti-PA tag [25] mAb (clone NZ-1 for EphA2, EphA3, EphA4, EphA5, EphA6, EphA7, EphA10, and EphB1), and sorted using SH800. After sorting, cultivation in a medium containing 0.5 mg/mL of Zeocin (InvivoGen, San Diego, CA, USA) or 0.5 mg/mL of G418 was progressed. These Eph receptors-overexpressed CHO-K1 (e.g., CHO/EphA1) clones were finally established. CHO/PA16-EphB4 was previously described [22].
CHO-K1, P3U1, and each Eph receptor-overexpressed CHO-K1 cells were also cultured in a Roswell Park Memorial Institute (RPMI)-1640 medium (Nacalai Tesque, Inc.) that was supplemented with 10% heat-inactivated fetal bovine serum (FBS, Thermo Fisher Scientific Inc.), 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). LN229 and LN229/EphA8 were cultured in a Dulbecco’s Modified Eagle Medium (DMEM) (Nacalai Tesque, Inc.) that was supplemented with 10% heat-inactivated FBS (Thermo Fisher Scientific Inc.), 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). Then, cells were cultured in a humidified incubator, supplied with 5% CO2 and 95% air at 37°C.
2.2. Antibodies
The secondary Alexa Fluor 488-conjugated anti-mouse IgG was purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
2.3. Development of Hybridomas
For developing anti-EphA8 mAbs, two 6-week-old female BALB/cAJcl mice purchased from CLEA Japan (Tokyo, Japan) were immunized intraperitoneally with 1 × 108 cells of LN229/PA16-EphA8. The immunogen was harvested after brief exposure to 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). Alhydrogel adjuvant 2% (InvivoGen) was added as an adjuvant in the first immunization. Three additional injections of 1 × 108 cells of LN229/PA16-EphA8 were performed without an adjuvant addition every week. A final booster immunization was performed with 1 × 108 cells of LN229/PA16-EphA8 intraperitoneally two days before harvesting splenocytes from mice. We conducted cell-fusion of the harvested splenocytes from immunized mice with P3U1 cells using polyethylene glycol 1500 (PEG1500; Roche Diagnostics, Indianapolis, IN) under warming conditions.
Hybridomas were cultured in the RPMI-1640 medium, supplemented as shown above. We further added hypoxanthine, aminopterin, and thymidine (HAT; Thermo Fisher Scientific, Inc.), 5% BriClone (NICB, Dublin, Ireland), and 5 μg/mL of Plasmocin (InvivoGen) into the medium. The hybridoma supernatants were screened by flow cytometry using CHO/EphA8 and parental CHO-K1 cells. The cultured supernatant of Ea8Mab-9-producing hybridomas was filtrated and purified using Ab-Capcher Extra (ProteNova, Kagawa, Japan).
2.4. Flow Cytometric Analysis
CHO-K1, CHO/EphA1, CHO/EphA2, CHO/EphA4, CHO/EphA6, CHO/EphA7, CHO/EphA8, CHO/EphB1, CHO/EphB6, LN229, and LN229/EphA8 cells were harvested after brief exposure to 1 mM EDTA. CHO/EphA3, CHO/EphA5, CHO/EphA10, CHO/EphB2, CHO/EphB3, CHO/EphB4 were harvested after brief exposure to 0.25% trypsin and 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). Cells were washed with 0.1% bovine serum albumin in phosphate-buffered saline (PBS) and treated with primary mAb for 30 min at 4°C. Afterward, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG (1:1000) following the collection of fluorescence data using the SA3800 Cell Analyzer (Sony Corp.).
2.5. Determination of Dissociation Constant (KD) by Flow Cytometry
CHO/EphA8 and LN229/EphA8 were suspended in 100 μL serially diluted Ea8Mab-9 (10 µg/mL to 0.0006 µg/mL), after which Alexa Fluor 488-conjugated anti-mouse IgG (1:200) was added. Fluorescence data were subsequently collected using the BD FACSLyric (BD Biosciences, Franklin Lakes, NJ, USA), following the calculation of the dissociation constant (KD) by fitting the binding isotherms into the built-in one-site binding model in GraphPad PRISM 6 (GraphPad Software, Inc., La Jolla, CA, USA).
3. Results
3.1. Development of Anti-EphA8 mAbs Using the CBIS Method
To develop anti-EphA8 mAbs, we employed the CBIS method using EphA8-overexpressed cells. Anti-EphA8 mAbs-producing hybridoma were screened by using flow cytometry (Figure 1). Two female BALB/cAJcl mice were intraperitoneally immunized with LN229/PA16-EphA8 every week for a total of 5 times. Subsequently, hybridomas were seeded into 96-well plates, after which flow cytometric analysis was used to select CHO/EphA8-reactive and parental CHO-K1-nonreactive supernatants of hybridomas. Afterward, we obtained some highly CHO/EphA8-reactive supernatants of hybridomas. We finally established the highest sensitivity clone, Ea8Mab-9 (mouse IgG1, kappa), by limiting dilution and additional analysis.
3.2. Flow Cytometric Analysis
Flow cytometric analysis was conducted using Ea8Mab-9 against CHO-K1, CHO/EphA8, LN229, and LN229/EphA8 cells. Results indicated that Ea8Mab-9 recognized CHO/EphA8 (Figure 2A, upper) and LN229/EphA8 (Figure 2B, upper) dose-dependently. Ea8Mab-9 did not react with parental CHO-K1 (Figure 2A, lower) and LN229 (Figure 2B, lower) cells even at 10 µg/mL of Ea8Mab-9. Ea8Mab-9 could react to EphA8 specifically in flow cytometry.
3.3. Specificity of Ea8Mab-9 to Eph Receptor-Expressed CHO-K1 Cells
We have established the all Eph receptor-overexpressed CHO-K1 cells, EphA1 to A8, A10, B1 to B4, and B6 respectively. Using the fourteen cell lines, the specificity of Ea8Mab-9 was evaluated. As shown in Figure 3, Ea8Mab-9 could recognize only CHO/EphA8, but not other Eph receptors-expressed cell lines. We confirmed the specificity of Ea8Mab-9 among Eph receptors.
3.4. Determination of the Binding Affinity of Ea8Mab-9 by Flow Cytometry
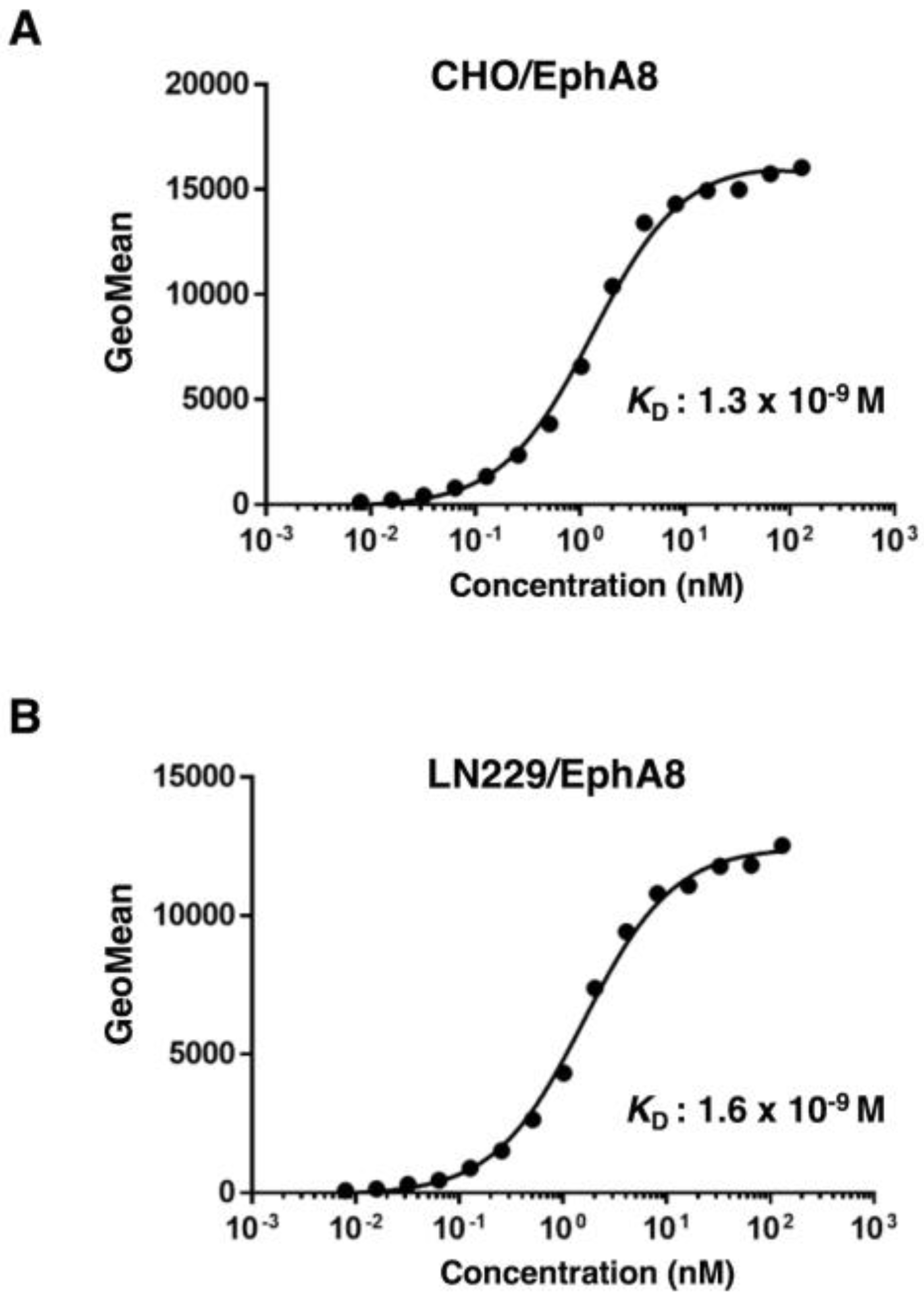
The binding affinity of Ea8Mab-9 was assessed with exogenously EphA8-expressed CHO/EphA8 and LN229/EphA8 using flow cytometry. Results showed that the KD values of Ea8Mab-9 were 1.3×10-9 M (CHO/EphA8) and 1.6×10-9 M (LN229/EphA8) (Figure 4). These results demonstrate that Ea8Mab-9 possesses a high affinity for EphA8.
4. Discussion
In this study, we developed a novel anti-EphA8 mAb, Ea8Mab-9 using the the CBIS method and showed the high affinity and specificity in flow cytometry (Figure 2, Figure 3 and Figure 4). We also established other anti-EphA8 mAb clones for flow cytometry (http://www.med-tohoku-antibody.com/topics/001_paper_antibody_PDIS.htm#EphA8). Ea8Mab-9 and other clones would contribute to the elucidation of EphA8 functions in various researches.
EphA8 regulates brain development and neural outgrowth in normal tissues [10,12], and its overexpression in cancer has been reported [13,14,16]. EphA8 TK-dependent activation appears to be an essential mechanism for its function. Indeed, Tyr-phosphorylated EphA8 regulates cell-cell adhesion by interacting with Fyn [6]. On the other hand, TK-independent roles of EphA8 have also been noted [7]. Recently, it has become evident that the RTKs’ modification mediated by Ser/Thr kinases is closely related to the regulation and function of receptors. EGFR internalization is mediated by Ser/Thr phosphorylation by p38 MAPK [26,27]. EphA2, the most extensively studied Eph receptors, is phosphorylated Ser-897 by p90 ribosomal S6 kinase and correlates tumor malignant progression [28]. Cellular stress triggers these modifications to RTKs [29,30]. The role of EphA8 has also been addressed. In breast cancer, EphA8 suppresses cell apoptosis via Akt activation and correlates with poor prognosis [16]. Simultaneously, EphA8 expression may mediate the resistance to paclitaxel [16]. Also, in normal tissue, EphA8 induces apoptosis of ephrin A5-expressing neural epithelial cells during the brain development of mice [10]. EphA8 has been reported to govern apoptosis in both tumor and normal tissues. Furthermore, EphA8 is involved in gastric cancer malignancy through Akt signaling and interaction with ADAM10 [15]. The expression of EphA8 significantly correlates with only TNM stage, but not other risk factors such as age, gender, drinking, and smoking history in OTSCC patients.[13] Although the TK dependency was not clarified in these reports, the TK-independent regulation of EphA8 by Ser/Thr kinases may function through a non-canonical TK-independent mechanism. Since EphA8 has been reported to cooperate with stem cells [17], the relationship between EphA8-expressing cells and other membrane protein markers such as CD44 and CD133 might be evaluated [31,32]. To analyze the population of EphA8-expressing cancer, it is necessary to detect naïve EphA8 in cancer cells. Because Ea8Mab-9 recognizes EphA8 with high affinity, it will be helpful for diagnosis and experiments using flow cytometry.
Eph receptors have been widely studied about cancer and are attracting attention as therapeutic targets [2,33]. Clinical trials of various modalities, including compounds, antibody drugs, and chimeric antigen receptor (CAR)-T cells, have been conducted targeting Eph receptors and ephrin ligands such as EphA2, EphA3, EphA5, EphB4, ephrin A4, and ephrin B2 [2,34]. However, there are currently no drugs approved specifically for Eph receptors or ephrin ligands. The clinical tests targeting EphA8 have not been confirmed yet. Although the specific antibody is also desired to clarify the character of EphA8 molecule in basic research, commercially available antibodies for EphA8 is not abundant. In particular, no commercially available anti-EphA8 antibodies are suitable for flow cytometry. Ea8Mab-9 has high affinity and is amenable to flow cytometry (Figure 2, Figure 3 and Figure 4). Ea8Mab-9 is expected to lead to a more detailed elucidation of the role of EphA8 in cancer, including the tumor microenvironment and in neuronal research. Furthermore, we have previously enhanced antibody-dependent cellular cytotoxicity (ADCC) and complement-dependent cytotoxicity (CDC) activities by switching isotypes and eliminating fucosylation in mAbs [35,36,37,38]. Since Ea8Mab-9 is mouse IgG1, which lacks ADCC activity, its mouse IgG2a version will be generated to examine the efficacy of antitumor activities in tumor xenograft models in the future study.
Author Contributions
Tomohiro Tanaka: Investigation, Writing – original draft, Haruto Yamamoto: Investigation, Yu Kaneko: Investigation, Keisuke Shinoda: Investigation, Takuya Nakamura: Investigation, Guanjie Li: Investigation, Shiori Fujisawa: Investigation, Hiroyuki Satofuka: Investigation, Mika K. Kaneko: Conceptualization, Funding acquisition, Hiroyuki Suzuki: Funding acquisition, Writing – review and editing, Yukinari Kato: Conceptualization, Funding acquisition, Project administration, Writing – review and editing, All authors have read and agreed to the published version of the manuscript.
Funding Information
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP24am0521010 (to Y.Kato.), JP24ama121008 (to Y.Kato.), JP24ama221339 (to Y.Kato.), JP24bm1123027 (to Y.Kato.), and JP24ck0106730 (to Y.Kato.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 22K06995 (to H.Suzuki), 21K20789 (to T.T.), 24K11652 (to H.Satofuka), 21K07168 (to M.K.K.), and 22K07224 (to Y.Kato.).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care and Use Committee of Tohoku University (Permit number: 2022MdA-001) for studies involving animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
All related data and methods are presented in this paper. Additional inquiries should be addressed to the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest involving this article.
References
- Tuzi, N.L.; Gullick, W.J. eph, the largest known family of putative growth factor receptors. Br J Cancer 1994;69(3): 417-421. [CrossRef]
- Pasquale, E.B. Eph receptors and ephrins in cancer progression. Nat Rev Cancer 2024;24(1): 5-27. [CrossRef]
- Pasquale, E.B. Eph-ephrin bidirectional signaling in physiology and disease. Cell 2008;133(1): 38-52. [CrossRef]
- Chan, J.; Watt, V.M. eek and erk, new members of the eph subclass of receptor protein-tyrosine kinases. Oncogene 1991;6(6): 1057-1061.
- Park, S.; Sánchez, M.P. The Eek receptor, a member of the Eph family of tyrosine protein kinases, can be activated by three different Eph family ligands. Oncogene 1997;14(5): 533-542. [CrossRef]
- Choi, S.; Park, S. Phosphorylation at Tyr-838 in the kinase domain of EphA8 modulates Fyn binding to the Tyr-615 site by enhancing tyrosine kinase activity. Oncogene 1999;18(39): 5413-5422. [CrossRef]
- Gu, C.; Park, S. The EphA8 receptor regulates integrin activity through p110gamma phosphatidylinositol-3 kinase in a tyrosine kinase activity-independent manner. Mol Cell Biol 2001;21(14): 4579-4597.
- Wang, S.D.; Rath, P.; Lal, B.; et al. EphB2 receptor controls proliferation/migration dichotomy of glioblastoma by interacting with focal adhesion kinase. Oncogene 2012;31(50): 5132-5143. [CrossRef]
- Bhatia, S.; Bukkapatnam, S.; Van Court, B.; et al. The effects of ephrinB2 signaling on proliferation and invasion in glioblastoma multiforme. Mol Carcinog 2020;59(9): 1064-1075. [CrossRef]
- Kim, Y.; Park, E.; Noh, H.; Park, S. Expression of EphA8-Fc in transgenic mouse embryos induces apoptosis of neural epithelial cells during brain development. Dev Neurobiol 2013;73(9): 702-712. [CrossRef]
- Park, S.; Frisén, J.; Barbacid, M. Aberrant axonal projections in mice lacking EphA8 (Eek) tyrosine protein kinase receptors. Embo j 1997;16(11): 3106-3114. [CrossRef]
- Gu, C.; Shim, S.; Shin, J.; et al. The EphA8 receptor induces sustained MAP kinase activation to promote neurite outgrowth in neuronal cells. Oncogene 2005;24(26): 4243-4256. [CrossRef]
- Liu, L.; Wang, X.; Ge, W. EphA8 is a Prognostic Factor for Oral Tongue Squamous Cell Carcinoma. Med Sci Monit 2018;24: 7213-7222. [CrossRef]
- Liu, X.; Xu, Y.; Jin, Q.; et al. EphA8 is a prognostic marker for epithelial ovarian cancer. Oncotarget 2016;7(15): 20801-20809. [CrossRef]
- Wang, Y.; Zhou, N.; Li, P.; et al. EphA8 acts as an oncogene and contributes to poor prognosis in gastric cancer via regulation of ADAM10. J Cell Physiol 2019;234(11): 20408-20419. [CrossRef]
- Wang, G.H.; Ni, K.; Gu, C.; et al. EphA8 inhibits cell apoptosis via AKT signaling and is associated with poor prognosis in breast cancer. Oncol Rep 2021;46(2). [CrossRef]
- Lucero, M.; Thind, J.; Sandoval, J.; et al. Stem-like Cells from Invasive Breast Carcinoma Cell Line MDA-MB-231 Express a Distinct Set of Eph Receptors and Ephrin Ligands. Cancer Genomics Proteomics 2020;17(6): 729-738. [CrossRef]
- Ren, W.; Chen, S.; Liu, G.; et al. TUSC7 acts as a tumor suppressor in colorectal cancer. Am J Transl Res 2017;9(9): 4026-4035.
- Yan, Y.; Wang, Q.; Yan, X.L.; et al. miR-10a controls glioma migration and invasion through regulating epithelial-mesenchymal transition via EphA8. FEBS Lett 2015;589(6): 756-765. [CrossRef]
- Satofuka H, S.H., Tanaka T, Li G, Kaneko MK, Kato Y. An Anti-Human EphA2 Monoclonal Antibody Ea2Mab-7 Shows High Sensitivity for Flow Cytometry, Western Blot, and Immunohistochemical Analyses. Preprint 2024.
- Ubukata R, S.H., Hirose M, Satofuka H, Tanaka T, Kaneko MK, Kato Y. Establishment of a Highly-sensitive Anti-EphB2 Monoclonal Antibody Eb2Mab-3 for Flow Cytometry. Preprint 2024.
- Nanamiya, R.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of an Anti-EphB4 Monoclonal Antibody for Multiple Applications Against Breast Cancers. Monoclon Antib Immunodiagn Immunother 2023;42(5): 166-177.
- Yamada, S.; Itai, S.; Nakamura, T.; et al. Detection of high CD44 expression in oral cancers using the novel monoclonal antibody, C(44)Mab-5. Biochem Biophys Rep 2018;14: 64-68.
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon Antib Immunodiagn Immunother 2016;35(6): 293-299. [CrossRef]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; et al. PA tag: a versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr Purif 2014;95: 240-247. [CrossRef]
- Tanaka, T.; Zhou, Y.; Ozawa, T.; et al. Ligand-activated epidermal growth factor receptor (EGFR) signaling governs endocytic trafficking of unliganded receptor monomers by non-canonical phosphorylation. J Biol Chem 2018;293(7): 2288-2301.
- Perez Verdaguer, M.; Zhang, T.; Paulo, J.A.; et al. Mechanism of p38 MAPK-induced EGFR endocytosis and its crosstalk with ligand-induced pathways. J Cell Biol 2021;220(7). [CrossRef]
- Zhou, Y.; Yamada, N.; Tanaka, T.; et al. Crucial roles of RSK in cell motility by catalysing serine phosphorylation of EphA2. Nat Commun 2015;6: 7679. [CrossRef]
- Nishimura, M.; Shin, M.S.; Singhirunnusorn, P.; et al. TAK1-mediated serine/threonine phosphorylation of epidermal growth factor receptor via p38/extracellular signal-regulated kinase: NF-{kappa}B-independent survival pathways in tumor necrosis factor alpha signaling. Mol Cell Biol 2009;29(20): 5529-5539.
- Zhou, Y.; Oki, R.; Tanaka, A.; et al. Cellular stress induces non-canonical activation of the receptor tyrosine kinase EphA2 through the p38-MK2-RSK signaling pathway. J Biol Chem 2023;299(5): 104699. [CrossRef]
- Paya, L.; Rafat, A.; Talebi, M.; et al. The Effect of Tumor Resection and Radiotherapy on the Expression of Stem Cell Markers (CD44 and CD133) in Patients with Squamous Cell Carcinoma. Int J Hematol Oncol Stem Cell Res 2024;18(1): 92-99. [CrossRef]
- da Silva, D.D.; Araldi, R.P.; Belizario, M.R.; et al. DLK1 Is Associated with Stemness Phenotype in Medullary Thyroid Carcinoma Cell Lines. Int J Mol Sci 2024;25(22). [CrossRef]
- Zhou, Y.; Sakurai, H. Emerging and Diverse Functions of the EphA2 Noncanonical Pathway in Cancer Progression. Biol Pharm Bull 2017;40(10): 1616-1624. [CrossRef]
- Janes, P.W.; Vail, M.E.; Gan, H.K.; Scott, A.M. Antibody Targeting of Eph Receptors in Cancer. Pharmaceuticals (Basel) 2020;13(5). [CrossRef]
- Li, G.; Suzuki, H.; Ohishi, T.; et al. Antitumor activities of a defucosylated anti-EpCAM monoclonal antibody in colorectal carcinoma xenograft models. Int J Mol Med 2023;51(2). [CrossRef]
- Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. A Cancer-Specific Monoclonal Antibody against Podocalyxin Exerted Antitumor Activities in Pancreatic Cancer Xenografts. Int J Mol Sci 2023;25(1).
- Arimori, T.; Mihara, E.; Suzuki, H.; et al. Locally misfolded HER2 expressed on cancer cells is a promising target for development of cancer-specific antibodies. Structure 2024;32(5): 536-549.e535. [CrossRef]
- Tanaka, T.; Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. Antitumor activities against breast cancers by an afucosylated anti-HER2 monoclonal antibody H(2) Mab-77-mG(2a) -f. Cancer Sci 2024;115(1): 298-309.
Figure 1.
A schematic diagram of anti-EphA8 mAbs development. The procedure of antibody development using the Cell-Based Immunization and Screening (CBIS) method. (A) LN229/PA16-EphA8 cells were immunized into two mice by intraperitoneal injection. (B) The spleen cells from antigen-immunized mice were fused with myeloma P3U1 cells by PEG1500. (C) The culture supernatants of hybridoma were screened by flow cytometry using CHO-K1 and CHO/EphA8 to select EphA8-specific mAb-producing hybridomas. (D) After limiting the dilution of hybridomas to obtain the single clone and additional screening, Ea8Mab-9 (mouse IgG1, kappa) was finally established.
Figure 1.
A schematic diagram of anti-EphA8 mAbs development. The procedure of antibody development using the Cell-Based Immunization and Screening (CBIS) method. (A) LN229/PA16-EphA8 cells were immunized into two mice by intraperitoneal injection. (B) The spleen cells from antigen-immunized mice were fused with myeloma P3U1 cells by PEG1500. (C) The culture supernatants of hybridoma were screened by flow cytometry using CHO-K1 and CHO/EphA8 to select EphA8-specific mAb-producing hybridomas. (D) After limiting the dilution of hybridomas to obtain the single clone and additional screening, Ea8Mab-9 (mouse IgG1, kappa) was finally established.
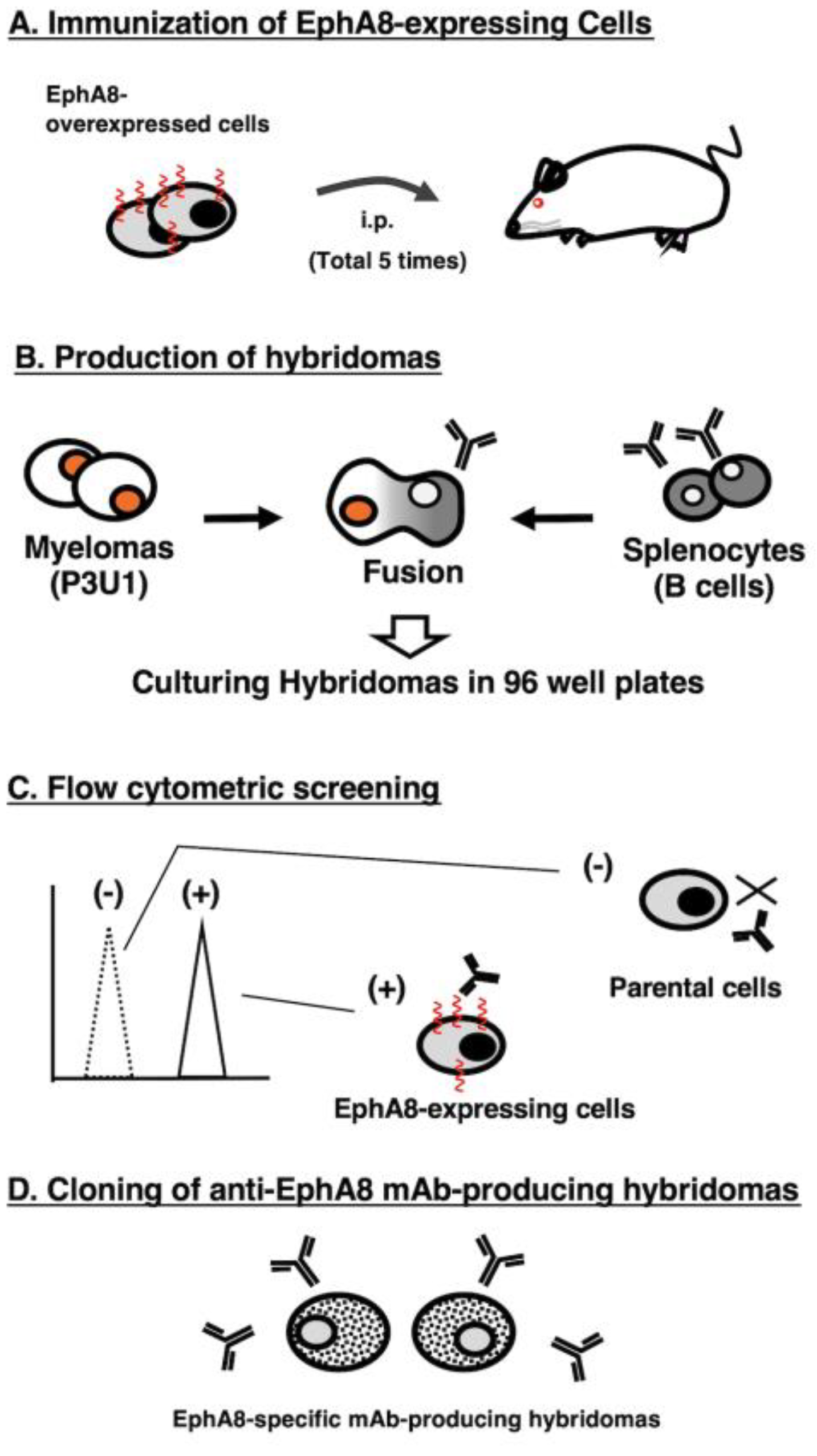
Figure 2.
Flow cytometric analysis of Ea8Mab-9. (A) CHO/EphA8 (upper panels) and CHO-K1 (lower panels) cells were treated with 0.01–10 µg/mL of Ea8Mab-9 (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment). (B) LN229/EphA8 (upper panels) and LN229 (lower panels) cells were treated with 0.01–10 µg/mL of Ea8Mab-9 (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
Figure 2.
Flow cytometric analysis of Ea8Mab-9. (A) CHO/EphA8 (upper panels) and CHO-K1 (lower panels) cells were treated with 0.01–10 µg/mL of Ea8Mab-9 (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment). (B) LN229/EphA8 (upper panels) and LN229 (lower panels) cells were treated with 0.01–10 µg/mL of Ea8Mab-9 (red line), followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. Fluorescence data were collected using the SA3800 Cell Analyzer. Black line, control (no primary antibody treatment).
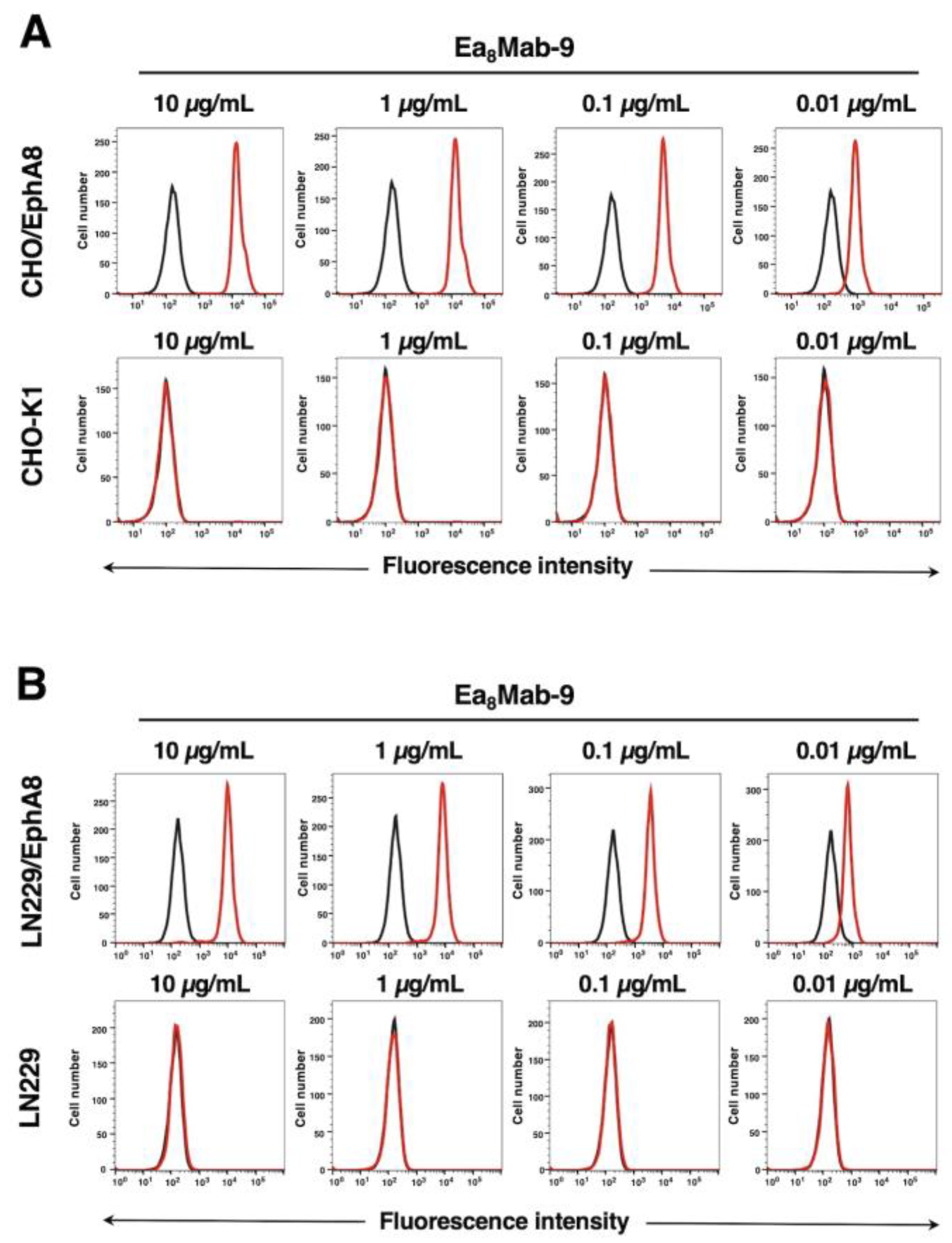
Figure 3.
Flow cytometry of Ea8Mab-9 in Eph receptor-expressed CHO-K1 cells. CHO-K1 cells which expressed each of the fourteen Eph receptors were treated with 10 µg/mL of Ea8Mab-9 (red line) or control blocking buffer (black line) followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488.
Figure 3.
Flow cytometry of Ea8Mab-9 in Eph receptor-expressed CHO-K1 cells. CHO-K1 cells which expressed each of the fourteen Eph receptors were treated with 10 µg/mL of Ea8Mab-9 (red line) or control blocking buffer (black line) followed by the treatment with anti-mouse IgG conjugated with Alexa Fluor 488.
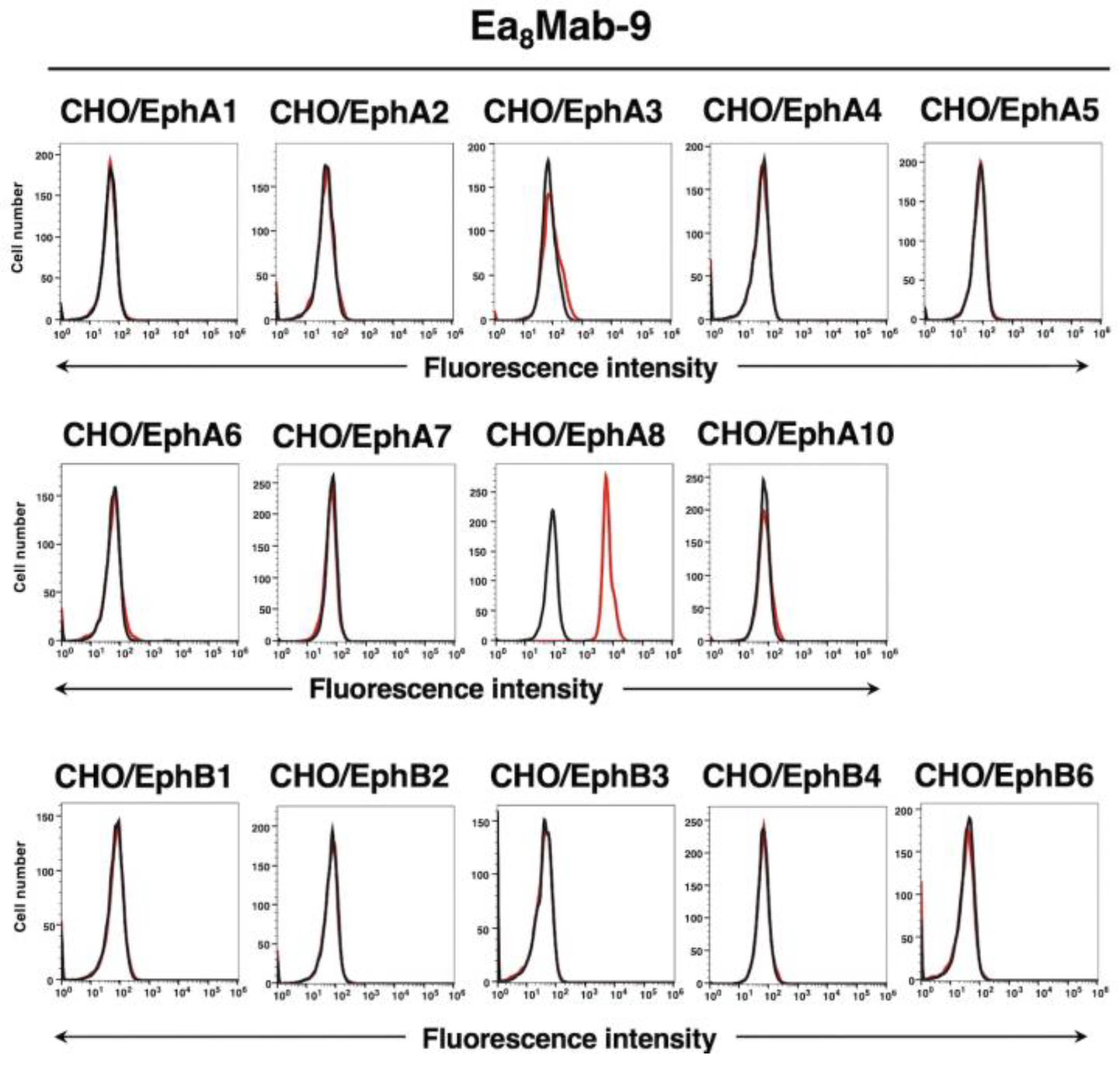
Figure 4.
Determination of the binding affinity of Ea8Mab-9 against EphA8-expressed cells. CHO/EphA8 (A) and LN229/EphA8 (B) cells were suspended in 100 µL of serially diluted Ea8Mab-9 (10 µg/mL to 0.0006 µg/mL). Then, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG. Subsequently, fluorescence data were collected using the BD FACSLyric, following the calculation of the dissociation constant (KD) by GraphPad PRISM 6 software.
Figure 4.
Determination of the binding affinity of Ea8Mab-9 against EphA8-expressed cells. CHO/EphA8 (A) and LN229/EphA8 (B) cells were suspended in 100 µL of serially diluted Ea8Mab-9 (10 µg/mL to 0.0006 µg/mL). Then, cells were treated with Alexa Fluor 488-conjugated anti-mouse IgG. Subsequently, fluorescence data were collected using the BD FACSLyric, following the calculation of the dissociation constant (KD) by GraphPad PRISM 6 software.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



