Submitted:
18 February 2025
Posted:
19 February 2025
You are already at the latest version
Abstract
As an endogenous hormone, auxin plays a crucial role in regulating plants’ growth and development. Auxin synthesis and transport are mediated by gene expression regulation, but the regulatory mechanism is poorly understood. Therefore, we studied this problem using citrus (trifoliate orange, citrus rootstocks) as the experimental material. The experiment involved the treatment of two groups of trifoliate orange with exogenous auxin (indolebutyric acid, IBA) and auxin inhibitor (2-naphthoxyacetic acid, 2-NOA), respectively, in a sand culture system. The aim of this study is to investigate the regulatory mechanisms by which auxin affects seedling growth and mineral nutrition levels in trifoliate orange. The results showed that exogenous auxin significantly enhanced taproot elongation and lateral root density, while also markedly elevating the levels of phosphorus (P), cuprum (Cu), zinc (Zn), and other mineral nutrients in trifoliate orange leaves. In contrast, the auxin inhibitor produced the opposite effects. Exogenous auxin treatment significantly upregulated the expression of genes related to auxin synthesis and transport and led to a significant increase in auxin content. Exogenous auxin inhibitor treatment significantly reduced the expression levels of most auxin synthesis and transport genes and led to a significant decrease in auxin content. According to the results of this study, the synthesis and transport of auxin regulate trifoliate orange root growth, thereby further affecting its absorption of mineral nutrients.
Keywords:
1. Introduction
2. Materials and Methods
2.1. Materials and Growth Conditions
2.2. Experiment Design
2.3. Study of Root Morphology
2.4. Mineral Nutrient Content Analysis
2.5. Root Auxin Content
2.6. Analysis of Gene Expression
2.7. Statistical Analysis
3. Results
3.1. The Morphology of Trifoliate Orange Seedlings
3.2. The Growth of Main and Lateral Roots

| Treatment | Tap root length (cm) | Tap root diameter (cm) | Lateral root length (cm) | Lateral root number (#) |
| CK | 5.24±1.48b | 0.11±0.01b | 0.39±0.08b | 1.80±0.44b |
| 1.0 µmol•L-1 IBA | 6.16±0.39a | 0.13±0.01a | 0.87±0.22a | 3.40±1.63a |
| 50 µmol•L-1 2-NOA | 4.12±0.14c | 0.10±0.01b | 0.35±0.03c | 1.02±0.10c |
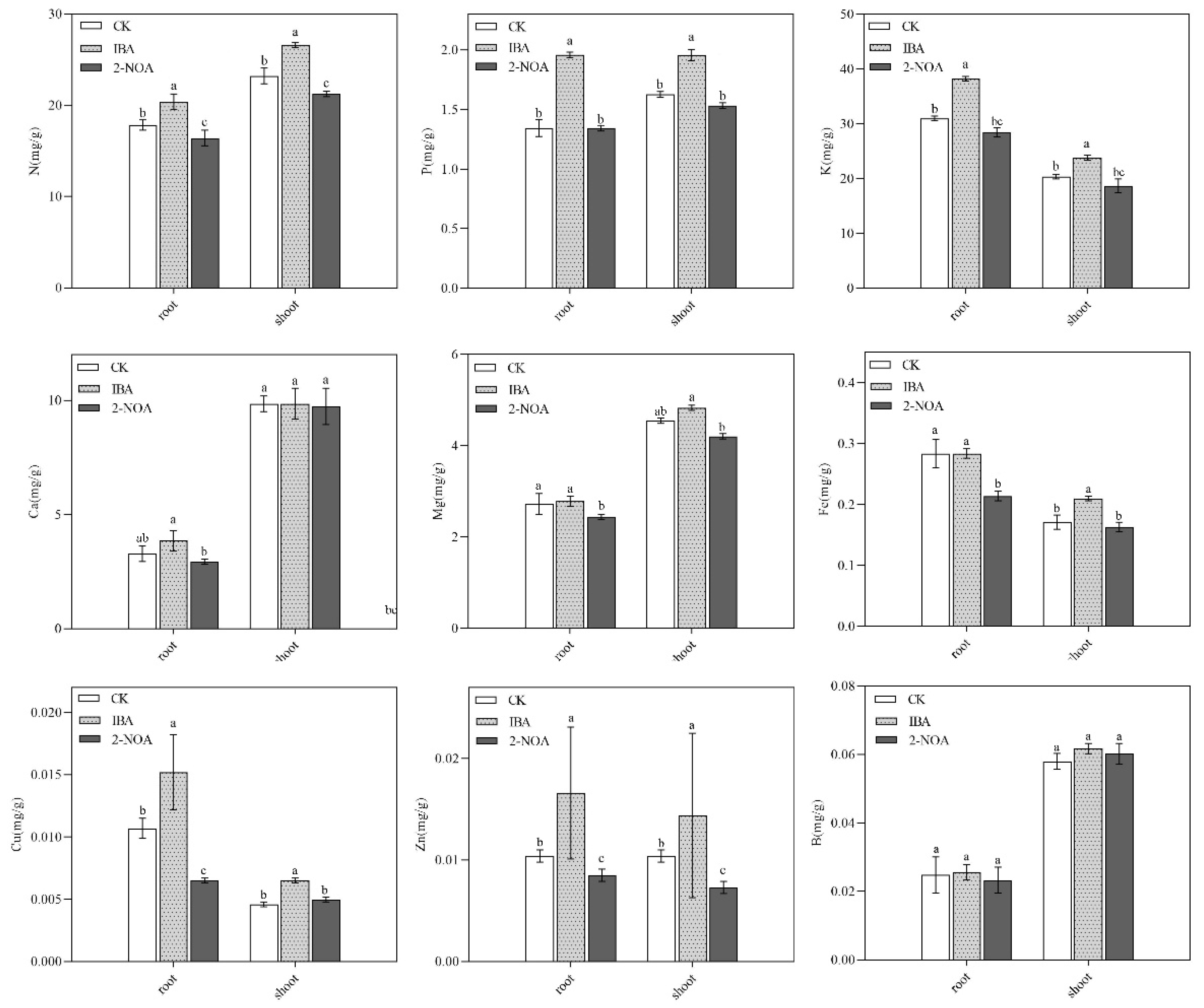
3.3. Variations in Plant Mineral Nutrient Composition
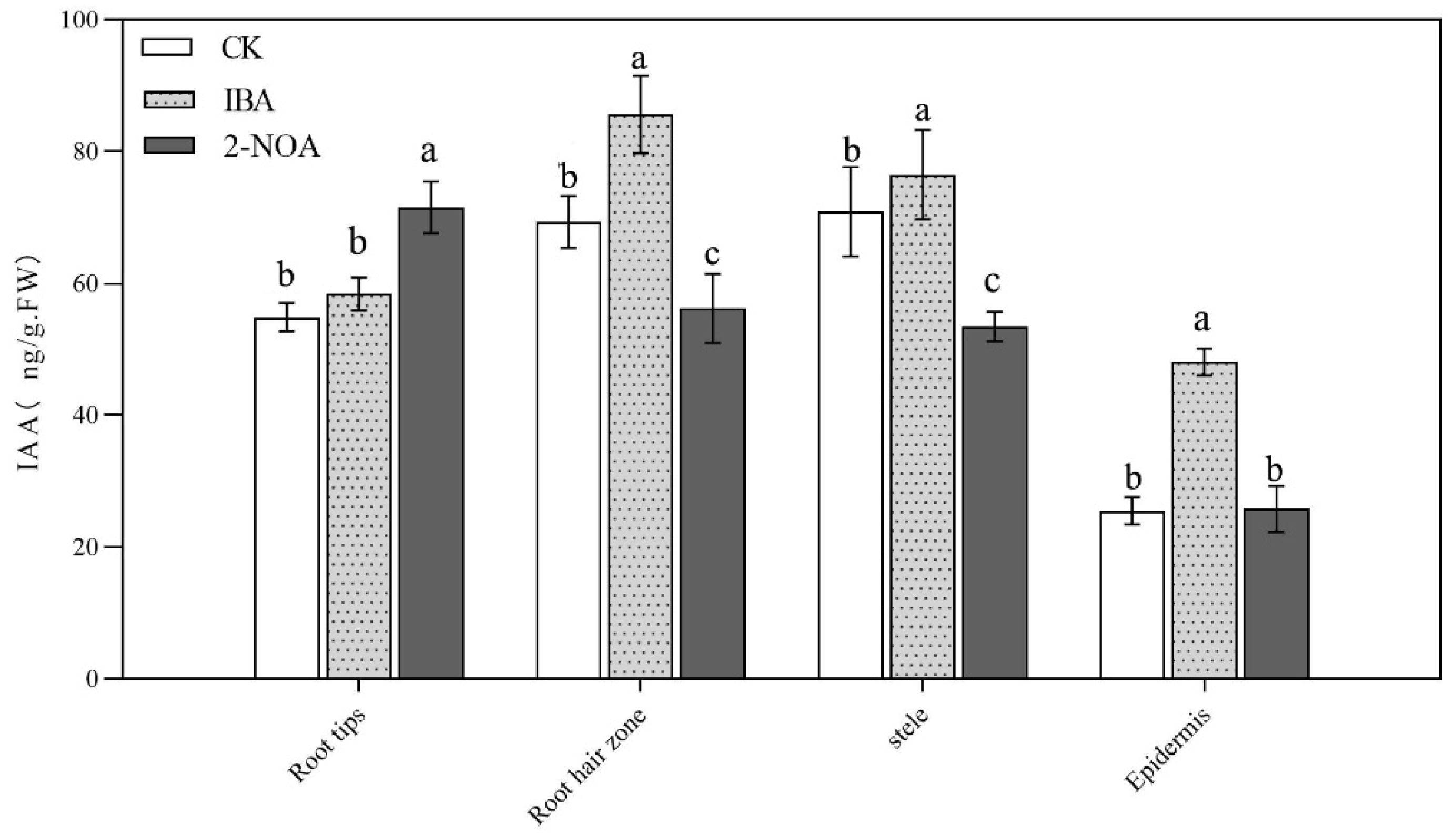
3.4. Measurement of Endogenous Auxin Content in Roots
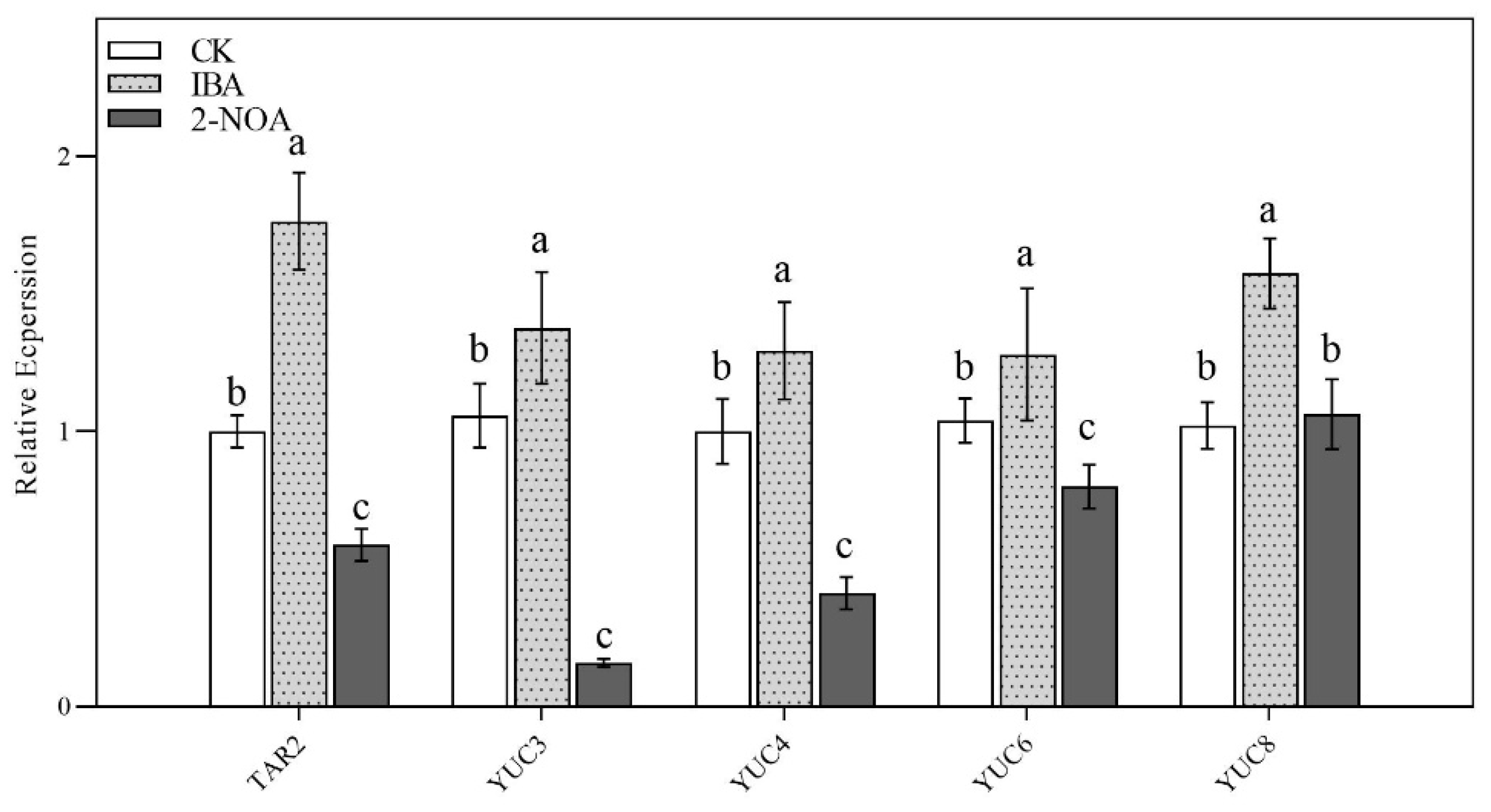
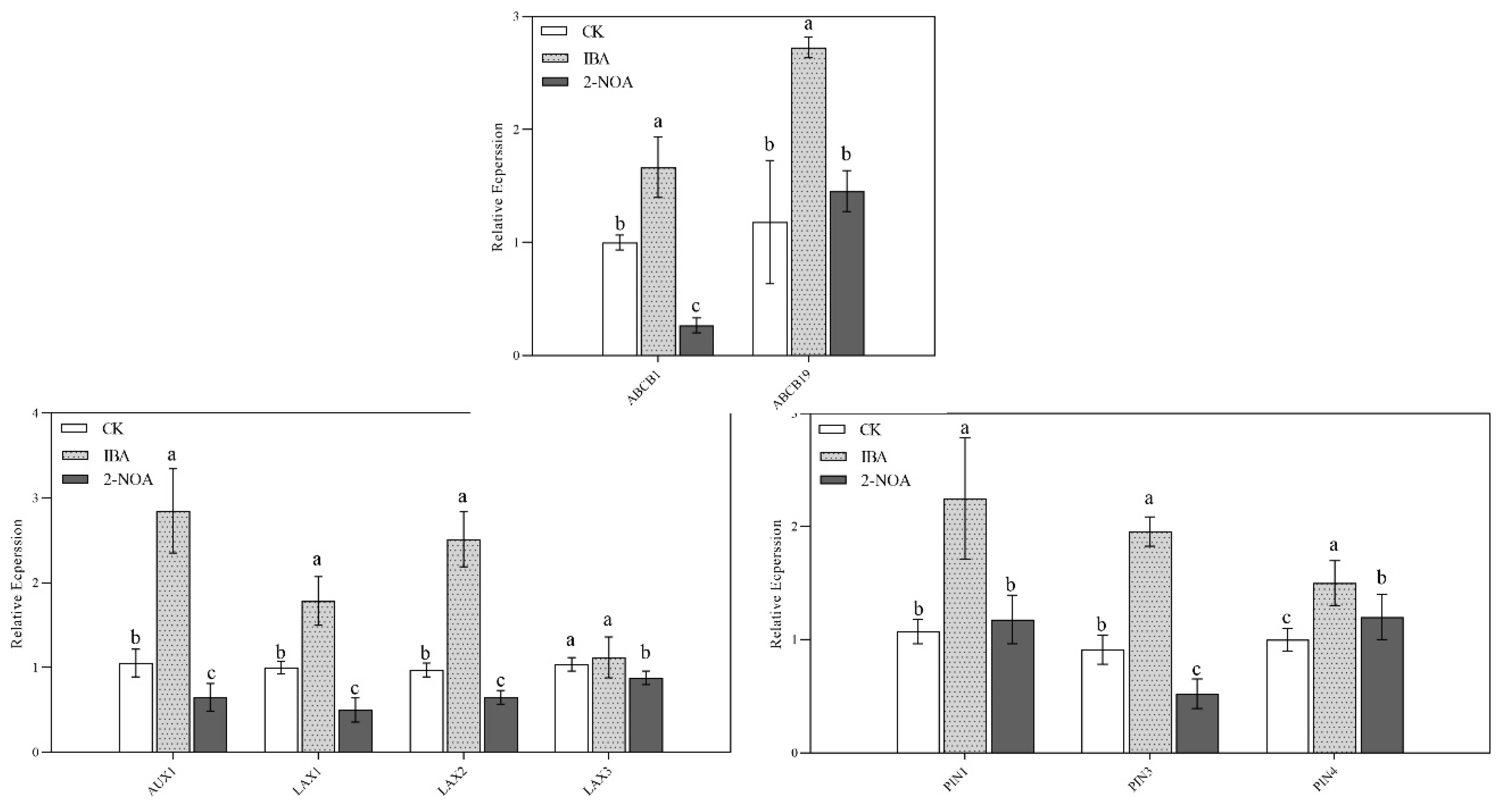
3.5. The Expression Levels of Root Auxin Biosynthesis and Transport Genes
4. Discussion
4.1. Effect of Auxin on the Root Growth of Trifoliate Orange
4.2. Effect of Auxin on Mineral Nutrients of Trifoliate Orange
4.3. Expression of Auxin and Its Related Genes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Quint, M.; Gray, W.M. Auxin signaling. Curr Opin Plant Biol. 2006, 9(5), 448-53. [CrossRef]
- Cavallari, N.; Artner, C.; Benkova, E. Auxin-regulated lateral root organogenesis. Csh Perspect Biol. 2021, 13(7), a039941. https://doi:10.1101/cshperspect.a03994.
- Meirav. Lavy.; Mark, Estelle. Mechanisms of auxin signaling. Development. 2016, 143(18), 3226-3229. [CrossRef]
- Chen, Y.; Barzee, T.J.; Zhang, R.; et al. Citrus: Integrated Processing technologies for food and agricultural by-products. Academic Press. 2019, 217-242. [CrossRef]
- Eekhout, J.P.C.; de, Vente, J. Global impact of climate change on soil erosion and potential for adaptation through soil conservation. Earth-Sci Rev. 2022, 226, 103921. [CrossRef]
- Rivas, M.Á. Friero, I,; Alarcón, M.V.; et al. Auxin-cytokinin balance shapes maize root architecture by controlling primary root elongation and lateral root development. Front Plant Sci. 2022, 13, 836592. [CrossRef]
- Qin, H.; Huang, R. Auxin controlled by ethylene steers root development. Int J. Mol Sci. 2018, 19(11), 3656. [CrossRef]
- Jeon, B.W.; Kim, M.J.; Pandey, S.K.; et al. Recent advances in peptide signaling during Arabidopsis root development. J. Exp Bot. 2021, 72(8), 2889-2902. [CrossRef]
- Thelander, M.; Landberg, K.; Muller, A.; et al. Apical dominance control by TAR-YUC-mediated auxin biosynthesis is a deep homology of land plants. Curr Biol. 2022, 32(17), 3838-3846. e5. [CrossRef]
- Kim, H.; Jang, J.; Seomun, S.; et al. Division of cortical cells is regulated by auxin in Arabidopsis roots. Front Plant Sci. 2022, 13, 953225. [CrossRef]
- Swarup R, Péret B. AUX/LAX family of auxin influx carriers—an overview. Front Plant Sci. 2012, 3, 32237. [CrossRef]
- Singh G, Retzer K, Vosolsobě S, et al. Advances in understanding the mechanism of action of the auxin permease AUX1. Int J. Mol Sci. 2018, 19(11), 3391. [CrossRef]
- Zhou, J.J.; Luo, J. The PIN-FORMED auxin efflux carriers in plants. Int J. Mol Sci. 2018, 19(9), 2759. [CrossRef]
- Vosolsobě, S.; Skokan, R.; Petrášek, J. The evolutionary origins of auxin transport: what we know and what we need to know. J. Exp Bot. 2020, 71(11), 3287-3295. [CrossRef]
- Fageria, N.K.; Moreira, A. The role of mineral nutrition on root growth of crop plants. Adv Agron. 2011, 110, 251-331. [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants. 2021, 10(2), 419. [CrossRef]
- Farhat, N.; Elkhouni, A.; Zorrig, W.; et al. Effects of magnesium deficiency on photosynthesis and carbohydrate partitioning. Acta Physiol Plant. 2016, 38(6), 145. [CrossRef]
- Wang, J.; Yang, S. Dose-dependent responses of Arabidopsis thaliana to zinc are mediated by auxin homeostasis and transport. Environ Exp Bot. 2021, 189, 104554. [CrossRef]
- Borah, S.; Baruah, A.M.; Das, A.K.; et al. Determination of mineral content in commonly consumed leafy vegetables. Food Anal Method. 2009, 2, 226-230. [CrossRef]
- Kramberger, B.; Gselman, A.; Janzekovic, M.; et al. Effects of cover crops on soil mineral nitrogen and on the yield and nitrogen content of maize. Eur J Agron. 2009, 31(2), 103-109. [CrossRef]
- Ali, B.; Sabri, A.N.; Ljung, K.; et al. Auxin production by plant associated bacteria: impact on endogenous IAA content and growth of Triticum aestivum L. Lett Appl Microbiol. 2009, 48(5), 542-547. [CrossRef]
- Xu, Q.; Chen, L.L.; Ruan, X.; et al. The draft genome of sweet orange (Citrus sinensis). Nat Genet. 2013, 45(1), 59-66. [CrossRef]
- Ceng-Hong, H.U.; Shi-Dong, Y.; Cui-Ling, T.; et al. Ethylene modulates root growth and mineral nutrients levels in trifoliate orange through the auxin-signaling pathway. Not Bot Horti Agrobo. 2023, 51(3), 13269-13269. [CrossRef]
- Livak, K.J.; Schmittgen, T.D.; Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔ CT method. Methods. 2001, 25(4), 402-408. [CrossRef]
- Pandey, R. Mineral nutrition of plants. Plant Biology and Biotechnology: Volume I: Plant Diversity, Organization, Function and Improvement. 2015, 499-538. [CrossRef]
- Luo, P.; Di, D.W. Precise regulation of the TAA1/TAR-YUCCA auxin biosynthesis pathway in plants. Int J. Mol Sci. 2023, 24(10), 8514. [CrossRef]
- Li, Y.; Zhu, J.; Wu, L.; et al. Functional divergence of PIN1 paralogous genes in rice. Plant Cell Physiol. 2019, 60(12), 2720-2732. [CrossRef]
- Cavallari, N.; Artner, C.; Benkova, E. Auxin-regulated lateral root organogenesis. Csh Perspect Biol. 2021, 13(7), a039941. [CrossRef]
- Qin, H.; Huang, R. Auxin controlled by ethylene steers root development. Int J. Mol Sci. 2018, 19(11), 3656. [CrossRef]
- Poza-Viejo, L.; Abreu, I.; González-García, M.P.; et al. Boron deficiency inhibits root growth by controlling meristem activity under cytokinin regulation. Plant Sci. 2018, 270, 176-189. [CrossRef]
- Bouain, N.; Krouk, G.; Lacombe, B.; et al. Getting to the root of plant mineral nutrition: combinatorial nutrient stresses reveal emergent properties. Trends Plant Sci. 2019, 24(6), 542-552. [CrossRef]
- Chen, J.; Liu, L.; Wang, Z.; et al. Nitrogen fertilization increases root growth and coordinates the root–shoot relationship in cotton. Front Plant Sci. 2020, 11, 880. [CrossRef]
- Xu, Q.; Fu, H.; Zhu, B.; et al. Potassium improves drought stress tolerance in plants by affecting root morphology, root exudates, and microbial diversity. Metabolites. 2021, 11(3), 131. [CrossRef]
- Omondi, J.O.; Lazarovitch, N.; Rachmilevitch, S.; et al. Phosphorus affects storage root yield of cassava through root numbers. J. Plant Nut. 2019, 42(17), 2070-2079. [CrossRef]
- Qu, S.; Li, H.; Zhang, X.; et al. Effects of Magnesium Imbalance on Root Growth and Nutrient Absorption in Different Genotypes of Vegetable Crops. Plants, 2023, 12(20), 3518. [CrossRef]
- Salas-González, I.; Reyt, G.; Flis, P.; et al. Coordination between microbiota and root endodermis supports plant mineral nutrient homeostasis. Science. 2021, 371(6525), eabd0695. [CrossRef]
- Phuphong, P.; Cakmak, I.; Yazici, A.; et al. Shoot and root growth of rice seedlings as affected by soil and foliar zinc applications. J. Plant Nut. 2020, 43(9), 1259-1267. [CrossRef]
- Sun, J.; Li, Q.; Xu, H.; et al. Analysis of metabolomic changes in xylem and phloem sap of cucumber under phosphorus stresses. Metabolites, 2022, 12(4), 361. [CrossRef]
- Ye, X.; Chen, X.F.; Deng, C.L.; et al. Magnesium-deficiency effects on pigments, photosynthesis and photosynthetic electron transport of leaves, and nutrients of leaf blades and veins in Citrus sinensis seedlings. Plants, 2019, 8(10), 389. [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; et al. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front Plant Sci. 2022, 13, 932861. [CrossRef]
- Li, X.; Kang, H.; Chen, S..; et al. Gln-Lys isopeptide bond and boroxine synergy to develop strong, anti-mildew and low-cost soy protein adhesives. J Clean Prod. 2023, 397, 136505. [CrossRef]
- Yang, X.M.; Ren, G.M.; Luo, K.J. Effects of Sepiolite and Mycorrhizals on Nutrient Element Absorptions of Maize in Heavy Metals Contaminated Soil. Advanced Materials Research. 2013, 800, 153-158. [CrossRef]
- Jiang, H.B.; Lu, X.H.; Deng, B.; et al. Adaptive mechanisms of the model photosynthetic organisms, cyanobacteria, to iron deficiency. Microbial photosynthesis. 2020, 197-244. [CrossRef]
- Rawat, J.; Pandey, N.; Saxena, J.; Role of potassium in plant photosynthesis, transport, growth and yield. Role of potassium in abiotic stress. 2022, 1-14. [CrossRef]
- Sardans, J.; Peñuelas, J.; Potassium control of plant functions: Ecological and agricultural implications. Plants. 2021, 10(2), 419. [CrossRef]
- Du, Y.; Scheres, B. Lateral root formation and the multiple roles of auxin. J. Exp Bot. 2018, 69(2), 155-167. [CrossRef]
- Leftley, N.; Banda, J.; Pandey, B.; et al. Uncovering how auxin optimizes root systems architecture in response to environmental stresses. Csh Perspectt Biol. 2021, 13(11), a040014. [CrossRef]
- Zhang, P.; Sun, L.; Qin, J.; et al. cGMP is involved in Zn tolerance through the modulation of auxin redistribution in root tips. Environ Exp Bot. 2018, 147, 22-30. [CrossRef]
- Xi, D.; Chen, X.; Wang, Y.; et al. Arabidopsis ANAC092 regulates auxin-mediated root development by binding to the ARF8 and PIN4 promoters. J. Integr Plant Biol. 2019, 61(9), 1015-1031. [CrossRef]

| Gene | Accession No. | Forward primer (5’-3’) | Reverse primer (5’-3’) | Amplification size (bp) |
| ABCB1 | Ciclev10010916m | GAGCCATTCACGCCACTTC | TCTTGTAACCGAGCCTTTGAGC | 186 |
| ABCB19 | Ciclev10010931m | GCATGAGTTTGGGTCAGTCTTT | CATCTTCCATTTGTTGGGTCTT | 127 |
| AUX1 | Ciclev10011596m | CTTGACTCTGCCCTATTCATTCTC | TGGACCCAGTAACCCATCAAGC | 205 |
| LAX1 | Ciclev10031413m | TTGGCGGACATGCAGTGAC | CAGCGGCAGCAGAAGGAAT | 123 |
| LAX2 | Ciclev10028271m | TGTGGGAAGATGGGTAGGGAC | TAGTCATGCTCGCCCACCC | 98 |
| LAX3 | Ciclev10001072m | ATCACTTTCGCTCCTGCTGC | CAAACCCAAATCCCACCACTA | 133 |
| PIN1 | Ciclev10007787m | GCTTTGGCAACAGAAGAGGATT | ATTACACTTGTCGGCGGCATA | 94 |
| PIN3 | orange1.1g006199m | CATGCCTCCAGCGAGTGTTAT | TGCCACCTGAAAGCGATTAGA | 126 |
| PIN4 | Ciclev10012938m | ATGGGGTTGAAAACGAAGGG | CCTGATAAGTTTCCTCCACACCA | 167 |
| TAR2 | Ciclev10020085m | CACACACGGCACACCCCTA | GCCTCCCACTCCCCAGATC | 137 |
| YUC3 | Ciclev10006828m | CCTTCAGGTTTAGCCGTTGC | GGAAGTTTGGAAGTTGGCAGA | 157 |
| YUC4 | Ciclev10008466m | GACCATCTGGGTTAGCCGTTT | GTATTTTGGGAAGTTTTCAGGGA | 185 |
| YUC6 | Ciclev10008473m | GTGGTTGCTAAAGTGGCTGC | GTTGAAGGGGACCCAAAAGA | 122 |
| YUC8 | Ciclev10020503m | GTGATAATGGTAGGGGCAGGA | GAATGGCAGGTGAGGGAGC | 183 |
| β-actin | Ciclev10025866m | CCGACCGTATGAGCAAGGAAA | TTCCTGTGGACAATGGATGGA | 190 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).



