
Submitted:
18 February 2025
Posted:
19 February 2025
You are already at the latest version
Abstract
Ciliates of the genus Frontonia have been extensively studied to resolve their phylogenetic and evolutionary history, but challenges remain. This study used molecular analyses of SSU rRNA genes, phylogenetic tree reconstruction, molecular dating, and diversification analysis, together with ancestral state reconstruction of morphological traits and habitat preferences. Data included newly sequenced Korean species, GenBank records and published morphological information. Phylogenetic trees revealed paraphyly within Frontonia, identifying four groups that emerged in the Mesozoic era: Group I (~172 mya), Group II (~83 mya), Group III (~115 mya), and Group IV (~190 mya), with a common ancestor dating to ~420 mya in the Palaeozoic era. Diversification analysis revealed higher extinction rates (0.826 and 0.613 species/year) than speciation rates (0.011 and 0.016 species/year). Morphological evolution showed habitat adaptation and plasticity, with habitat transitions unrelated to contractile vacuolar traits. The SSU rRNA gene polymorphism likely contributed to the paraphyletic state of Frontonia. These results highlight the complex evolutionary patterns of the genus, shaped by genetic diversity, morphology, and environmental constrains.
Keywords:
Frontonia
; evolutionary history
; morphological evolution
; diversification analysis
; SSU rRNA gene
; molecular dating
; ancestral state reconstruction
1. Introduction
The research on ciliates, with a particular focus on the genus Frontonia, has indeed advanced significantly due to the integration of molecular and morphological studies. The genus Frontonia, established nearly two centuries ago, represents a diverse group of ciliate species [1,2,3,4,5,6,7]. While various research efforts have contributed to the understanding of their morphological traits, the phylogenetic relationships and evolutionary history of many ciliate lineages, including Frontonia, remain unresolved [8]. Molecular analyses, particularly of the SSU rRNA gene, have provided crucial insights into the phylogenetic relationships within Frontonia. These analyses have revealed the paraphyletic nature of the genus, with several distinct clades identified. Furthermore, certain species consistently cluster with different genera in the SSU rRNA gene phylogenetic trees, suggesting a complex evolutionary history [8,9,10,11,12].
However, the molecular analysis of the SSU rRNA gene has been the primary tool used to construct phylogenetic trees within Frontonia, with limited integration of morphological data beyond species descriptions. To address this gap, a multifaceted approach is proposed in the current study. This approach involves the integration of molecular data, in particular the SSU rRNA gene, for molecular clock analysis. At the same time, morphological data will be used to reconstruct ancestral states, allowing for a more comprehensive understanding of the phylogenetic relationships and evolutionary history of Frontonia. By combining molecular and morphological data, the study aims to elucidate the evolutionary patterns, phylogenetic relationships, and divergence time of the genus Frontonia. These integrated approaches will provide deeper insights into the diversification patterns and evolutionary processes of the lineages in Frontonia.
2. Materials and Methods
Sample Collection and Morphological Study
Newly sequenced species of Frontonia were collected from different locations in South Korea. Detailed information on the collection sites can be found in Table 1. The Frontonia species were cultivated in petri dishes using water collected from their respective habitats. Initial in vivo cell observations were made using both bright field and differential interference contrast microscopy techniques. Subsequent detailed examinations were performed using a compound microscope, with magnifications ranging from 100x to 1000x for both live cells and stained samples. The protargol-impregnated method was employed to visualize the structure of the buccal area, the oral apparatus and the ciliary pattern.
DNA Extraction, Amplification, and Sequencing
The genomic DNA extraction was carried out using the RED Extract-N-Amp Tissue PCR Kit from Sigma (St. Louis, MO, USA) according to the manufacturer's instructions. The polymerase chain reaction (PCR) was performed using the forward primer EUKA (5'- GAC CGT CCT AGT TGG TC-3') [13] or 82F (5'-CTC GGT AAG CGT CAA AG-3') [14] and the reverse primers D1, D2 rev2 (5'-GAC TGC ACG TTT AGC TAG CA-3') or D1, D2 rev4 (5'- GTG CCT GGT TCY TCA GAT TG -3'). PCR amplifications were performed using the TaKaRa ExTaq DNA polymerase kit from TaKaRa Bio-medicals (Otsu, Japan) following a specific protocol: an initial denaturation cycle at 94°C for 2 minutes, followed by 37 cycles of denaturation at 95°C for 30 seconds, annealing at 50°C for 40 seconds, and extension at 72°C for 4 minutes. A final extension step was then carried out at 72°C for 10 minutes [15].
Alignment and Phylogenetic Analysis
In this study, 71 sequences from the species of the genus Frontonia obtained from NCBI and newly sequenced in Korea (Table A1), 20 sequences from related genera (Paranassula, Paramecium, Apofrontonia, Stokesia, Disematosoma, Marituja, Lembadion, Urocentrum, and Tetrahymena) were selected as reference sequences and two species from the genus Tetrahymena were selected as outgroup (T. pyriformis & T. rostrata) (Figure 1). The alignment for this data set was performed by using MAFFT version 7.0 [16]. Subsequent refinement and masking of the alignments were carried out using G-blocks version 0.91b (Castresana, 2000). To determine the best evolutionary models, jModeltest version 2.1.7 [18,19] was employed. Further analysis included maximum likelihood (ML) using PhyML 3.0 software with 1000 non-parametric bootstrap replicates [20]. In addition, Bayesian Inference (BI) analysis was performed using MrBayes version 3.1 [21].
Molecular Dating and Diversification Analysis
The analysis used 33 sequences from representative species of the genus Frontonia, selected on the basis of the availability of both molecular and morphological information (Suppl. Table 1). In addition, 18 sequences from species of related genera were included in the analysis. Divergence times were estimated using a Bayesian framework that implemented in BEAST ver. 2.4.5 [22,23]. The software BEAUti ver.2.4.5 [23] was used to generate the BEAST software input XML file with the following settings; (i) calibrated Yule model, (ii) GTR+I(=0.389)+Γ(=0.412), (iii) four gamma categories for substitution rate heterogeneity, (iv) strict molecular clock, (v) clock rate prior assuming a normal distribution with a mean of 3.88 × 10−4 nucleotide substitutions per site per one million years and a 95% credibility interval ranging from 1.24 × 10−4 to 9.14 × 10−4 (Rataj and Vďačný, 2018), and (vi) Yule birth rate set with shape parameter set to 0.001 and scale parameter set to 1000 (gamma shape parameter).
For the estimation of the divergence time, calibration nodes from the genera Paramecium and Tetrahymena were used. These nodes were based on microfossil evidence from Paramecium triassicum and Tetrahymena rostrata-like species found on an amber Triassic slab during the Upper Triassic period, approximately 220-230 million years ago (Mya) [24]. The mean divergence time for these calibration nodes was estimated to be 225 Mya. Markov Chain Monte Carlo (MCMC) analyses started with a random seed and ran for 600,0000 generations, with trees and all other parameters were saved every 10,000th iteration. The quality of the MCMC analysis was assessed using Tracer version 1.6 [25] to ensure convergence and adequate burn-in. The final maximum credibility tree was generated using TreeAnnotator ver. 1.8.1[22] after discarding the first 10% of sampled trees.
The analysis of speciation rate versus extinction rate in the genus Frontonia was performed using two software programs: Divbayes and SubT [26]. Divbayes used data containing information on the time of ancestral divergence and the corresponding number of species. On the other hand, SubT used 95% height data for each branch obtained from a chronogram generated by the BEAST software, together with species number data used in the analysis, including predicted data for species numbers where actual data were not available (Table A2). These analyses were conducted to explore patterns and dynamics of species diversification through time.
Reconstruction of Ancestral Morphologies
For the prediction of ancestral states, 33 species of the genus Frontonia and Apofrontonia dohrni were selected as representative taxa. The selection was based on the availability and credibility of their molecular and morphological data, as shown in Supplementary Table 2. To perform the ancestral state reconstruction, 12 important and significant characters were selected within the genus Frontonia. These characters included the number of ciliary rows in peroral membranes (PM), the number vestibular kineties (VK), the number of postoral kineties (PK), the number of ciliary rows in peniculi 1-3, the structure of P 3 (shortened and linearity with P1 and P2), the habitat, the number of contractile vacuoles (CV), the number of contractile vacuolar pores (CVP) and the presence of contractile canals (CC). The character matrix and ancestral state reconstruction were performed using the parsimony model in Mesquite software ver. 3.70 [27]. The reconstruction of ancestral states was based on the topology of the best likelihood tree obtained from the RAxML analysis performed on CIPRES [28].
3. Results
3.1. Phylogenetic Analyses
The phylogenetic analyses based on SSU rRNA gene sequences have provided insights into the evolutionary relationships within the genus Frontonia, revealing the existence of four main groups: Group I: This group includes species such as F. canadensis, F. subtropica, F. salmastra, F. sinica, F. magna, F. mengi, F. tchibisovae, and F. lynni. Both Bayesian Inference (BI) and Maximum Likelihood (ML) analyses strongly support this group, with node support values of 1.00/100%. Group II: Comprising species such as F. paramagna, F. leucas, F. vesiculosa, F. vernalis, F. shii, F. paravernalis, F. leucas, F. angusta, and F. cf. leucas, Group II also shows robust node support values of 1.00/100% for both BI and ML. Group III: This group includes species such as F. didieri, F. ocularis, F. elegans, F. pusilla, and F. anatolica, with slightly lower node support values of 0.98/85% for BI and ML compared to Groups I and II. Group IV: Including species such as F. terricola, F. cf. acuminata, F. acuminata, F. atra, F. apoacuminata, F. minuta, and F. cf. atra, Group IV shows moderate node support (BI/ML, 1.00/88%) (Figure 1).
Furthermore, Group III shows a close relationship with the genus Apofrontonia, albeit with lower node support (BI/ML, 0.81/60%). Group IV forms a cluster with the genera Stokesia, Marituja, and Disematostoma, with strong node support (BI/ML, 1.00/100%). These phylogenetic relationships shed light on the evolutionary dynamics and diversification patterns within the genus Frontonia highlighting both strong and moderate support for the identified groups and their relationships with related taxa (Figure 1).
3.2. Estimation of Divergence Times and Diversification Using the SSU rRNA Gene
The divergence time within the genus Frontonia can be estimated using two methods: relaxed clock and strict clock. Relaxed clock is commonly used at higher taxonomic levels such as family or order, while strict clock is favored for intraspecific level analyses where low rates of variation between branches are expected [29,30,31,32]. In the case of the genus Frontonia, the strict clock approach is more suitable for estimating divergence time due to the limited data set within the genus level, and the minimal variation between branches.
The analyses of divergence time in the genus Frontonia performed in this study show that Peniculia is estimated to have originated approximately 750 million years ago (Mya), confirming the previous findings of Rataj and Vďačný (2018) (Figure 2). Using the strict clock approach, the SSU rRNA gene showed a mean clock rate of 2.36 x 10e-4 per year. Node calibrations for the Tetrahymena clade and Paramecium clades indicate their emergence at 223 Mya and 226 Mya, respectively. The common ancestor of the genus Frontonia appeared around 420 Mya and serves as an ancestral point for all members of the Penicullida. Notably, the emergence of three clades within the genus Frontonia occurred relatively recently: Group I around 172 Mya, Group II around 83 Mya, Group III around 115 Mya, and Group IV around 190 Mya (Figure 2).
The results obtained from Divbayes and SubT for speciation and extinction indicate a higher extinction rate compared to the speciation rate with genus Frontonia: 0.826 species/year over speciation rate: 0.011 species/year (DivBayes 1.1) and extinction rate: 0.613 species/year over speciation rate: 0.016 species/year (SubT1.1).
3.3. Reconstruction of Ancestral Character State
The analysis of ancestral states in the genus Frontonia reveals intriguing evolutionary patterns across several morphological characters;
- Vestibular kineties (VK): Group III retains the ancestral state of three rows of VK, whereas Group I and II show increases to four and five rows, respectively. Notably, F. canadensis shows transitions from three to five and back to three rows, consistent with the ancestral state of the genus Frontonia. Group IV undergoes significant changes, expanding to four, five, and even six rows (Figure 3A).
- Postoral kineties (PK): Group I shows various increases in PK number (six to eight rows), Group II retains the ancestral state PK number except for F. magna, Group III decrease PK number except for F, anatolica, whereas Group IV shows both decreasing and increasing tendencies, with some species reaching up to six rows (Figure 3B).
- Peroral membrane (PM): Groups II and III show an increases in PM ciliary rows, although species such as F. sinica, F. salmastra, and F. lynni revert from two rows to one (Figure 3C).
- Peniculi 1 and 2 (P1 and P2): Group I shows a marked increase from four to five rows, while other groups, with a few exceptions, retain the ancestral state (Figure 3D-E).
- Peniculi 3 evolution: Groups I and II show an increases in ciliary rows from three to four or five, whereas F. mengi and F. sinica uniquely decrease to two rows. Group IV members retain the ancestral state with several species increasing to four or five rows, while Group III decreases from the ancestral three ciliary rows to two. Most Frontonia species retain the ancestral linear structure of peniculi 3, with shortening of peniculi 3 considered an ancestral character (Figure 3F-H).
- Habitat adaptation: The genus Frontonia originates from brackish habitats before adapting to freshwater environments. Group III remains in the brackish environment and adapts to marine habitats for several species, while Group I and IV retain a freshwater adaptation, except for F. acuminata, which adapts to a marine environment. Groups II evolves from freshwater to marine and to brackish habitats (Figure 4).
- Contractile vacuole (CV): The ancestral analysis suggests that the genus originally possessed a single CV, a character often retained. Group III, excluding some species, increases from one to two CVs, while unique species in Group IV show up to 10 CVs per individual (Figure 4).
- Contractile vacuolar pores (CVP) and collecting canals (CC): The evolution of CVPs and CCs is not closely correlated with the evolution of CVs (Figure 4).
These results shed light on the evolutionary history and adaptive strategies of Frontonia species, providing valuable insights into morphological evolution and habitat preferences.
4. Discussion
4.1. Evolutionary Patterns of Morphological Characters in the Genus Frontonia
The ancestral state reconstruction analysis provides valuable insights into the evolutionary patterns and morphological adaptations within the genus Frontonia. By examining the ancestral characters of Frontonia species, we can better understand the evolutionary trajectory of their oral apparatus and its adaptation to different habitats.
The results suggest that the common ancestor of Frontonia probably inhabited brackish waters and possessed specific characteristics related to its oral apparatus. This ancestor had a single contractile vacuole (CV), a single contractile vacuolar pore (CVP) and no contractile canals (CC). In addition, the ancestral species had consistent number of vestibular kineties (VK), postoral kineties (PK), and peroral membrane (PM) ciliary rows, as well as distinct structures in peniculi 1, 2, and 3. Frontonia canadensis, while closely resembling the ancestral state, shows evolutionary changes such as variations in the number of ciliary rows in peniculi 3 (P3) and changes in the structure of preoral membrane (PM). These changes highlight the dynamic nature of the morphological evolution of Frontonia [34].
The morphological characteristics of the oral apparatus, including the number of VK, PK, PM ciliary rows, and the structure of peniculi 1-3, serve as crucial markers for distinguishing Frontonia species. Reconstructing ancestral state using these characters reveals distinct evolutionary patterns for each character. While some characters, such as the number of VKs, show predictable evolutionary trends, others, such as the number of PM ciliary row, show reversals, highlighting the complexity of morphological evolution in Frontonia.
Habitat plays an important role in shaping morphological charcteristics of ciliates such as Frontonia [35,36,37]. However, our analysis indicates that there is no direct correlation between habitat type and the number of contractile vacuoles, pores, and canals in Frontonia species. This suggests that other factors, perhaps related to physiological adaptations, also influence the evolution of these traits. The observed stability of certain morphological characters across different habitats is consistent with the neutral morphological theory, suggesting that certain characters may be favored in different ecological contexts [38].
4.2. Evolutionary History of Genus Frontonia
The evolutionary history of the genus Frontonia provides insights into its emergence and divergence over millions of years, and sheds light on the environmental factors and evolutionary processes that have shaped its diversity. The common ancestor of the four groups within genus Frontonia and members of Penicullida emerged and diverged approximately 420 million years ago during the Palaeozoic era (Figure 2) [39]. Each group or clade within Frontonia originated at different times in geological time. Groups I and II shared a common ancestor at the beginning of the Mesozoic era about 230 million years ago, with Group I diverging about 172 million years ago and Group II diverging about 83 million years ago. Group III, morphologically close to the genus Apofrontonia, shared a common ancestor at the beginning of the Mesozoic era, about 185 million years ago, while Group III itself diverged about 115 million years ago. Group IV appeared about 190 million years ago, making it one of the oldest groups within Frontonia. Group I is the youngest, emerging at the end of the Mesozoic era.
The ancestor of the genus Frontonia probably diverged during the Cambrian explosion, a period of significant biological diversification. While species numbers increased during the Cambrian period, environmental changes, possibly driven by glacial cycles, led to mass extinctions in the Paleozoic era following the Cambrian explosion. Although fossil evidence for ciliates is scarce, it is plausible that the harsh environmental conditions limited the survival and development of the ancestral Frontonia populations, resulting in long branches without speciation during the Palaeozoic era [39,40,41].
Diversification tests suggest a higher extinction rate than the speciation rate in Frontonia, indicating the challenges faced by ancestral populations in maintaining their existence over geological time scales. Surviving Frontonia species persisted into the early Mesozoic era, during which time they diverged, coinciding with a warmer Earth and potentially more favorable environmental conditions [42,43].
The current paraphyly within the genus Frontonia may be due to SSU rRNA gene polymorphism in its ancestor, contributing to the high genetic diversity observed in the past. Harsh environmental conditions during the Palaeozoic era probably subjected ancestral Frontonia populations to natural selection, resulting in the decline of many populations. However, surviving populations retained variations in their SSU rRNA gene, potentially leading to divergent evolutionary trajectories. Over time, during the Mesozoic era, these divergent populations underwent speciation, possibly in response to changing environmental conditions, as supported by recent studies [8].
5. Conclusions
This study provides a comprehensive investigation of the phylogenetic, evolutionary and diversification dynamics within the genus Frontonia by integrating molecular data from the SSU rRNA gene with analyses of morphological characters. Reconstructions of ancestral states have provided valuable insights into the unique evolutionary patterns of each morphological character within Frontonia. In particular, habitat transitions do not show a direct correlation with the number of contractile vacuoles, contractile vacuole pores, and contractile canals, suggesting the influence of complex factors on these characters. The historical perspective of the genus Frontonia traces its origins back to the Cambrian explosion, revealing survival challenges during the Paleozoic era and subsequent diversification throughout the Mesozoic era. The observed paraphyly within Frontonia is attributed to SSU rRNA gene polymorphism in its ancestor, reflecting high genetic diversity and adaptation to changing environmental conditions over time.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table A1: Datasheet Genus Frontonia members in this Study; Table A.2. Datasheet that input into speciation rate analysis software.
Author Contributions
Conceptualization, R.K.W. and M.K.S.; methodology, R.K.W. and R.A.; software, R.K.W..; validation, R.K.W. and M.K.S.; formal analysis, R.K.W.; investigation, R.K.W. and R.A.; resources, M.K.S.; data curation, R.K.W.; writing—original draft preparation, R.K.W.; writing—review and editing, R.K.W., M.K.S., and R.A.; visualization, R.K.W.; supervision, M.K.S.; project administration, M.K.S.; funding acquisition, M.K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the NRF Korea funded by the Ministry of Education of the Republic of Korea (RS-2021-NR065806).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank MD Abu Taher (University of Ulsan, University of British Columbia) for providing sample (Frontonia cf. atra)) for analysis in this paper.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Appendix A.1

Table A1.
Datasheet Genus Frontonia members in this Study.
| Species | Accession No. | Origin | Molecular information (Source by publication based on SSU rRNA gene) | Morphological information |
|---|---|---|---|---|
| SSU | ||||
| Frontonia anatolica | MG456578 | Turkey | Kizildang & Yildiz, 2019 | Kizildang & Yildiz, 2019, Yildiz & Senler, 2013 |
| Frontonia anatolica | MG456581 | Turkey | ||
| Frontonia acuminata | MG456579 | Turkey | Kizildang & Yildiz, 2019 | Kizildang & Yildiz, 2019, Foissner, et al., 1994, Kahl, 1931, Pernard, 1922 |
| Frontonia angusta | MG456580 | Turkey | Kizildang & Yildiz, 2019 | Kizildang & Yildiz, 2019, Foissner, et al., 2002, Kahl, 1931, Foissner, et al., 1994 |
| Frontonia canadensis | KJ475313 | Shenzen, China | Zhao et al., 2016 | Pan, X., et al., 2013, Roque. and de Puytorac, 1972. |
| Frontonia canadensis | KJ475311 | Huizhou, China | ||
| Frontonia canadensis | KJ475309 | Huizhou, China | ||
| Frontonia canadensis | KJ475310 | Nansha, China | ||
| Frontonia canadensis | KJ475312 | Hongkang, China | ||
| Frontonia canadensis | FJ868196 | Nansha, China | Fan et al., 2013 | |
| Frontonia cf. acuminata | PV016905 | Gwangju, Korea | present study | present study |
| Frontonia cf. acuminata | PV016906 | Ulsan, Korea | ||
| Frontonia cf. acuminata | PV016907 | Gunsan,Korea | ||
| Frontonia cf. acuminata | PV016908 | Ulsan, Korea | ||
| Frontonia cf. atra | PV016909 | Daejon, Korea | ||
| Frontonia cf. atra | PV016910 | Gunsan, Korea | ||
| Frontonia cf. atra | PV016911 | Masan, Korea | ||
| Frontonia cf. atra | PV016912 | Chungcheongbukdo, Korea | ||
| Frontonia cf. atra | PV016913 | Gyeongsangnamdo, Korea | ||
| Frontonia cf. atra | PV016914 | Ulsan, Korea | ||
| Frontonia cf. atra | PV016917 | Gyeongju, korea | ||
|
Frontonia atra |
MT040844 | Serchio river, Italy | Serra et al., 2021 | Serra et al., 2021, Dragesco & Dragesco-Kerneis, 1986, Foissner, et.al., 1994 |
| Gangneung-si | Omar and Jung, 2021 | |||
| Frontonia didieri | KJ475297 | Qingdao, China | Zhao et al., 2016 | Long et al., 2008 |
| Frontonia didieri | KJ475299 | Qingdao, China | ||
| Frontonia didieri | KJ475298 | Shenzen, China | ||
| Frontonia didieri | DQ885986 | Qingdao, China | Long et al., 2008 | |
|
Frontonia ocularis |
FJ868198 | Guangzhou, China | Pan et al., 2013 | Pan et al., 2013 |
| Gangneung-si | Jung, 2021 | |||
| Frontonia elegans | FJ868200 | Guangzhou, China | Fan et al., 2013 | Fan et al., 2013 |
| Frontonia elegans | KJ475301 | Huizhou, China | Zhao et al., 2016 | |
| Frontonia elegans | KJ475302 | Huizhou, China | ||
| Frontonia elegans | KJ475303 | Huizhou, China | ||
| Frontonia cf. leucas | PV016904 | Gwangju, Korea | present study | present study |
| Frontonia leucas | MG437395 | Turkey | Yildiz & Kizildang , 2017 | Yildiz & Kizildang, 2019 |
| Frontonia leucas | MG437396 | |||
| Frontonia leucas | AM072622 | Italy | Fokin et al., 2006 | Serra et al., 2021, Dragesco & Dragesco-Kerneis, 1986, Foissner, et.al., 1994, Kahl, 1931 |
| Frontonia leucas | KY855558 | Andhra Pradesh, India | Serra et al., 2021 | |
| Frontonia lynni | DQ190463 | Qingdao, China | Long et al., 2008 | Long et al., 2005 |
| Frontonia magna | FJ868199 | Zhuhai, China | Zhao et al., 2016 | Fan et al., 2011 |
| Frontonia magna | FJ876953 | Shenzen, China | Fan et al., 2011 | |
| Frontonia mengi | FJ875141 | Qingdao, China | Fan et al., 2011 | Fan et al., 2011 |
| Frontonia paramagna | KJ475304 | Guangzhou, China | Zhao et al., 2016 | |
| Frontonia paramagna | KJ475306 | Sichuan, China | Zhao et al., 2016 | |
| Frontonia paramagna | JQ868786 | Harbin, China | Chen et al., 2014 | Chen et al., 2014 |
| Frontonia paramagna | KJ475305 | Qingdao, China | Zhao et al., 2016 | |
| Frontonia paramagna | MF279207 | Harbin, China | Cai et al., 2018 | Cai et al., 2018 |
| Frontonia paramagna | KY855554 | Andhra Pradesh, India | Serra et al., 2021 | Serra et al., 2021 |
| Frontonia paramagna | MW031789 | Heilojiang, China | Sun et al.,2021 | Sun et al.,2021 |
| Frontonia shii | MF279208 | Harbin, China | Cai et al., 2018 | Cai et al., 2018 |
| Frontonia sinica | KJ475308 | Qingdao, China | Zhao et al., 2016 | Fan et al., 2013 |
| Frontonia sinica | FJ868197 | Shenzen, China | Fan et al., 2013 | |
| Frontonia subtropica | FJ868202 | Shenzen, China | Fan et al., 2013 | Pan et al., 2013 |
| Frontonia tchibisovae | KJ475318 | Qingdao, China | Zhao et al., 2016 | Long et al., 2008 |
| Frontonia tchibisovae | KJ475316 | Qingdao, China | ||
| Frontonia tchibisovae | KJ475319 | Qingdao, China | ||
| Frontonia tchibisovae | DQ883820 | Yantai, China | Long et al., 2008 | |
| Frontonia terricola | MF926593 | Guangzhou, China | Xu et al., 2018 | Xu et al., 2018, Foissner, 1987, Foissner, et al., 2002 |
| Frontonia pusilla | FJ868201 | Guangzhou, China | Fan et al., 2013 | Fan et al., 2013 |
| Frontonia vernalis | U97110 | UK | Hirt et al., 1997 | |
| Frontonia vernalis | MT040840 | Tusacany, Italy | Serra et al., 2021 | Serra et al., 2021 |
| Frontonia paravernalis | MT040839 | Tusacany, Italy | Serra et al., 2021 | Serra et al., 2021 |
| Frontonia salmastra | MH319376 | Tusacany, Italy | Fokin et al., 2019 | Fokin et al., 2019, Dragesco & Dragesco-Kerneis, 1986 |
| Frontonia vesiculosa | MT040850 | Andhra Pradesh, India | Serra et al., 2021 | Serra et al., 2021, Dragesco & Dragesco-Kerneis, 1986, Kahl, 1932 |
| Frontonia minuta | MT040846 | Serchio river, Italy | Serra et al., 2021 | Serra et al., 2021, Dragesco & Dragesco-Kerneis, 1986 |
| Frontonia fusca | MT040845 | Serchio river, Italy | Serra et al., 2021 | Serra et al., 2021, Fokin, 2008, Kahl, 1932 |
| Frontonia sp. | MT040843 | India | Serra et al., 2021 | Serra et al., 2021 |
| Frontonia sp. | MT040841 | Serchio river, Italy | Serra et al., 2021 | Serra et al., 2021 |
Appendix A.2
Table A2.
Datasheet that input into speciation rate analysis software.
| DivBAyes 1.1 | SubT 1.1 | ||
|---|---|---|---|
| Estimated taxaa | Diversification date | Data of taxab | Subtitution taxac |
| 69 species | 450 Mya | 33 species | 36 species |
References
- Long, H.; Song, W.; Gong, J.; Hu, X.; Ma, H.; Zhu, M.; Wang, M. Frontonia Lynni n. Sp., a New Marine Ciliate (Protozoa, Ciliophora, Hymenostomatida) from Qingdao, China. Zootaxa 2005, 57–64. [Google Scholar] [CrossRef]
- Long, H.; Song, W.; Al-Rasheid, K.A.S.; Wang, Y.; Yi, Z.; Al-Quraishy, S.A.; Lin, X.; Al-Farraj, S.A. Taxonomic Studies on Three Marine Species of Frontonia from Northern China: F. Didieri n. Sp., F. Multinucleata n. Sp. and F. Tchibisovae Burkovsky, 1970 (Ciliophora: Peniculida). Zootaxa 2008, 50, 35–50. [Google Scholar] [CrossRef]
- Fan, X.; Chen, X.; Song, W.; Al-Rasheid, K.A.S.; Warren, A. Two Novel Marine Frontonia Species, Frontonia Mengi Spec. Nov. and Frontonia Magna Spec. Nov. (Protozoa; Ciliophora), with Notes on Their Phylogeny Based on Small-Subunit RRNA Gene Sequence Data. Int. J. Syst. Evol. Microbiol. 2011, 61, 1476–1486. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Lin, X.; Liu, W.; Xu, Y.; Al-Farraj, S.A.; Al-Rasheid, K.A.S.; Warren, A. Morphology of Three New Marine Frontonia Species (Ciliophora; Peniculida) with Note on the Phylogeny of This Genus. Eur. J. Protistol. 2013, 49, 312–323. [Google Scholar] [CrossRef]
- Pan, X.; Liu, W.; Yi, Z.; Fan, X.; Al-Rasheid, K.A.S.; Lin, X. Studies on Three Diverse Frontonia Species (Ciliophora, Peniculida), with Brief Notes on 14 Marine or Brackish Congeners. Acta Protozool. 2013, 52, 35–49. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Y.; Pan, X.; Ding, W.; Al-Rasheid, K.A.S.; Qiu, Z. Morphology and Phylogeny of a New Frontonia Ciliate, F. Paramagna Spec. Nov. (Ciliophora, Peniculida) from Harbin, Ortheast China. Zootaxa 2014, 3827, 375–386. [Google Scholar] [CrossRef]
- Cai, X.; Wang, C.; Pan, X.; El-Serehy, H.A.; Mu, W.; Gao, F.; Qiu, Z. Morphology and Systematics of Two Freshwater Frontonia Species (Ciliophora, Peniculida) from Northeastern China, with Comparisons among the Freshwater Frontonia Spp. Eur. J. Protistol. 2018, 63, 105–116. [Google Scholar] [CrossRef]
- Zhao, Y.; Yi, Z.; Gentekaki, E.; Zhan, A.; Al-Farraj, S.A.; Song, W. Utility of Combining Morphological Characters, Nuclear and Mitochondrial Genes: An Attempt to Resolve the Conflicts of Species Identification for Ciliated Protists Q. Mol. Phylogenet. Evol. 2016, 94, 718–729. [Google Scholar] [CrossRef]
- Kizildag, S.; Yildiz, I. Morphology and Molecular Phylogeny of Four Frontonia Species from Turkey (Protista, Ciliophora). Zootaxa 2019, 4609, 548–564. [Google Scholar] [CrossRef]
- Li, T.; Pan, X.; Lu, B.; Miao, M.; Liu, M. Taxonomy and Molecular Phylogeny of a New Freshwater Ciliate Frontonia Apoacuminata sp. nov. (Protista, Ciliophora, Oligohymenophorea) from Qingdao, PR China. Int. J. Syst. Evol. Microbiol. 2021, 71. [Google Scholar] [CrossRef]
- Serra, V.; D’Alessandro, A.; Nitla, V.; Gammuto, L.; Modeo, L.; Petroni, G.; Fokin, S.I. The Neotypification of Frontonia Vernalis (Ehrenberg, 1833) Ehrenberg, 1838 and the Description of Frontonia Paravernalis sp. nov. Trigger a Critical Revision of Frontoniid Systematics. BMC Zool. 2021, 6, 1–33. [Google Scholar] [CrossRef]
- Sun, M.; Li, Y.; Cai, X.; Liu, Y.; Chen, Y.; Pan, X. Further Insights into the Phylogeny of Peniculid Ciliates (Ciliophora, Oligohymenophorea) Based on Multigene Data. Mol. Phylogenet. Evol. 2021, 154, 107003. [Google Scholar] [CrossRef] [PubMed]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The Characterization of Enzymatically Amplified Eukaryotic 16S-like RRNA-Coding Regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Wang, C.; Pan, X.; El-Serehy, H.A.; Mu, W.; Gao, F.; Qiu, Z. Morphology and Systematics of Two Freshwater Frontonia Species (Ciliophora, Peniculida) from Northeastern China, with Comparisons among the Freshwater Frontonia Spp. Eur. J. Protistol. 2018, 63, 105–116. [Google Scholar] [CrossRef]
- Kim, S.Y.; Choi, J.K.; Dolan, J.R.; Shin, H.C.; Lee, S.; Yang, E.J. Morphological and Ribosomal DNA-Based Characterization of Six Antarctic Ciliate Morphospecies from the Amundsen Sea with Phylogenetic Analyses. J. Eukaryot. Microbiol. 2013, 60, 497–513. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Posada, D. JModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and High-Performance Computing Europe PMC Funders Group. Nat. Methods 2015, 9, 772. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian Evolutionary Analysis by Sampling Trees. BMC Evol. Biol. 2007, 7, 1–8. [Google Scholar] [CrossRef]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian Phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Schönborn, W.; Dörfelt, H.; Foissner, W.; Krienitz, L.; Schäfer, U. A Fossilized Microcenosis in Triassic Amber. J. Eukaryot. Microbiol. 1999, 46, 571–584. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Ryberg, M.; Nilsson, R.H.; Brandon Matheny, P. Divbayes and SubT: Exploring Species Diversification Using Bayesian Statistics. Bioinformatics 2011, 27, 2439–2440. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P. and D.R.M. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61. http://www.mesquiteproject.org 2019.
- Miller, M.A., Pfeiffer, W. and Schwartz, T. No TitleThe CIPRES Science Gateway: A Community Resource for Phylogenetic Analyses. Proc. 2011 TeraGrid Conf. Extrem. Digit. Discov. (pp. 1-8).
- Ho, S.Y.W.; Duchêne, S. Molecular-Clock Methods for Estimating Evolutionary Rates and Timescales. Mol. Ecol. 2014, 23, 5947–5965. [Google Scholar] [CrossRef]
- Rajter, Ľ.; Vďačný, P. Rapid Radiation, Gradual Extinction and Parallel Evolution Challenge Generic Classification of Spathidiid Ciliates (Protista, Ciliophora). Zool. Scr. 2016, 45, 200–223. [Google Scholar] [CrossRef]
- Vďačný, P. Estimation of Divergence Times in Litostomatean Ciliates (Ciliophora: Intramacronucleata), Using Bayesian Relaxed Clock and 18S RRNA Gene. Eur. J. Protistol. 2015, 51, 321–334. [Google Scholar] [CrossRef]
- Vďačný, P.; Breiner, H.-W.; Yashchenko, V.; Dunthorn, M.; Stoeck, T.; Foissner, W. The Chaos Prevails: Molecular Phylogeny of the Haptoria (Ciliophora, Litostomatea). Protist 2014, 165, 93–111. [Google Scholar] [CrossRef]
- Cai, X.; Wang, C.; Pan, X.; El-Serehy, H.A.; Mu, W.; Gao, F.; Qiu, Z. Morphology and Systematics of Two Freshwater Frontonia Species (Ciliophora, Peniculida) from Northeastern China, with Comparisons among the Freshwater Frontonia Spp. Eur. J. Protistol. 2018, 63, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Gao, F.; Liu, W.; Fan, X.; Warren, A.; Song, W. Morphology and SSU RRNA Gene Sequences of Three Frontonia Species, Including a Description of F. Subtropica Spec. Nov. (Ciliophora, Peniculida). Eur. J. Protistol. 2013, 49, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Clamp, J.; Xu, D.; Huang, B.; Shin, M.K. An Integrative Approach to Phylogeny Reveals Patterns of Environmental Distribution and Novel Evolutionary Relationships in a Major Group of Ciliates. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Syberg-Olsen, M.J.; Irwin, N.A.T.; Vannini, C.; Erra, F.; Di Giuseppe, G.; Boscaro, V.; Keeling, P.J. Biogeography and Character Evolution of the Ciliate Genus Euplotes (Spirotrichea, Euplotia), with Description of Euplotes Curdsi Sp. Nov. PLoS One 2016, 11, 1–18. [Google Scholar] [CrossRef]
- Zhao, Y.; Yi, Z.; Warren, A.; Song, W.B. Species Delimitation for the Molecular Taxonomy and Ecology of the Widely Distributed Microbial Eukaryote Genus Euplotes (Alveolata, Ciliophora). Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Lahr, D.J.G.; Laughinghouse, H.D.; Oliverio, A.M.; Gao, F.; Katz, L.A. How Discordant Morphological and Molecular Evolution among Microorganisms Can Revise Our Notions of Biodiversity on Earth. BioEssays 2014, 36, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Babcock, L.E.; Peng, S.C.; Brett, C.E.; Zhu, M.Y.; Ahlberg, P.; Bevis, M.; Robison, R.A. Global Climate, Sea Level Cycles, and Biotic Events in the Cambrian Period. Palaeoworld 2015, 24, 5–15. [Google Scholar] [CrossRef]
- Sepkoski, J.J. Environmental Trends in Extinction During the Paleozoic. American Association for the Advancement of Science Stable 2010, 235, 64–66. [Google Scholar]
- Stanley, E.L.; Bauer, A.M.; Jackman, T.R.; Branch, W.R.; Mouton, P.L.F.N. Between a Rock and a Hard Polytomy: Rapid Radiation in the Rupicolous Girdled Lizards (Squamata: Cordylidae). Mol. Phylogenet. Evol. 2011, 58, 53–70. [Google Scholar] [CrossRef]
- Jenkyns, Hugh, C. Evidence for Rapid Climate Change in the Mesozoic-Palaeogene Greenhouse World. Philosophical Transactions of the Royal Society of London. Series A: Mathematical, Physical and Engineering Sciences 2003, 361, 1885–1916. [CrossRef]
- Price, G.D.; Twitchett, R.J.; Wheeley, J.R.; Buono, G. Isotopic Evidence for Long Term Warmth in the Mesozoic. Sci. Rep. 2013, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic tree generated from Bayesian Inference (BI) using Mr Bayes ver. 3.1 software and Maximum Likelihood (ML) from PHYML ver. 3.0 based on the SSU rRNA (top) and SSU-ITS-LSU rRNA (bottom) gene sequences of the genus Frontonia and related genera. The scale bar indicates the number of base changes per 1000 nucleotide positions in BI analysis. Node support is represented as follows: BI posterior probability/ ML bootstrap. The newly sequenced species of the genus Frontonia are shown in red.
Figure 1.
Phylogenetic tree generated from Bayesian Inference (BI) using Mr Bayes ver. 3.1 software and Maximum Likelihood (ML) from PHYML ver. 3.0 based on the SSU rRNA (top) and SSU-ITS-LSU rRNA (bottom) gene sequences of the genus Frontonia and related genera. The scale bar indicates the number of base changes per 1000 nucleotide positions in BI analysis. Node support is represented as follows: BI posterior probability/ ML bootstrap. The newly sequenced species of the genus Frontonia are shown in red.
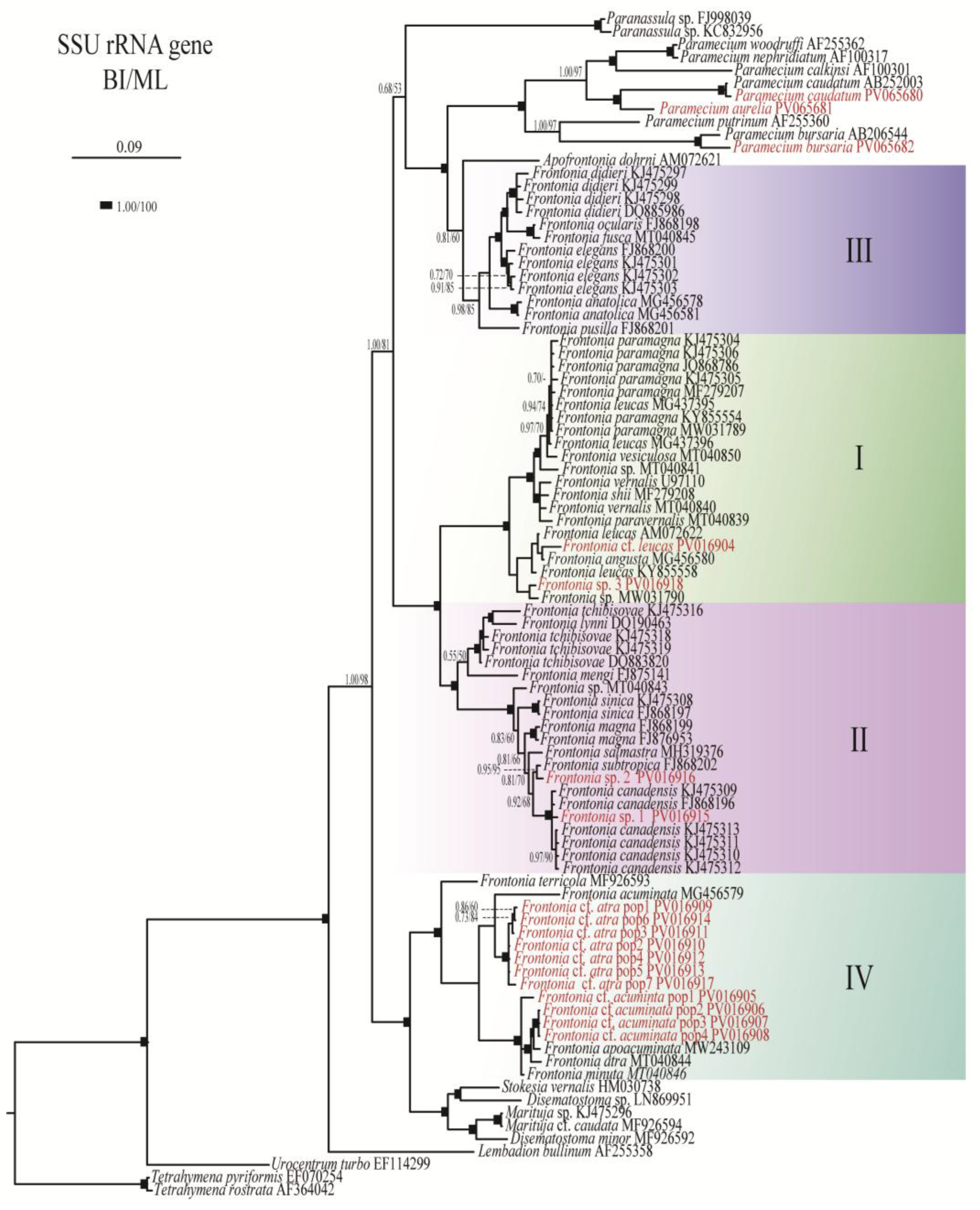
Figure 2.
Maximum credibility tree showing posterior means of divergence times of the genus Frontonia and related genera obtained using Bayesian relaxed molecular dating in BEAST. The 95% credibility intervals are shown as bars for all nodes. The horizontal axis represents the time scale in millions of years.
Figure 2.
Maximum credibility tree showing posterior means of divergence times of the genus Frontonia and related genera obtained using Bayesian relaxed molecular dating in BEAST. The 95% credibility intervals are shown as bars for all nodes. The horizontal axis represents the time scale in millions of years.
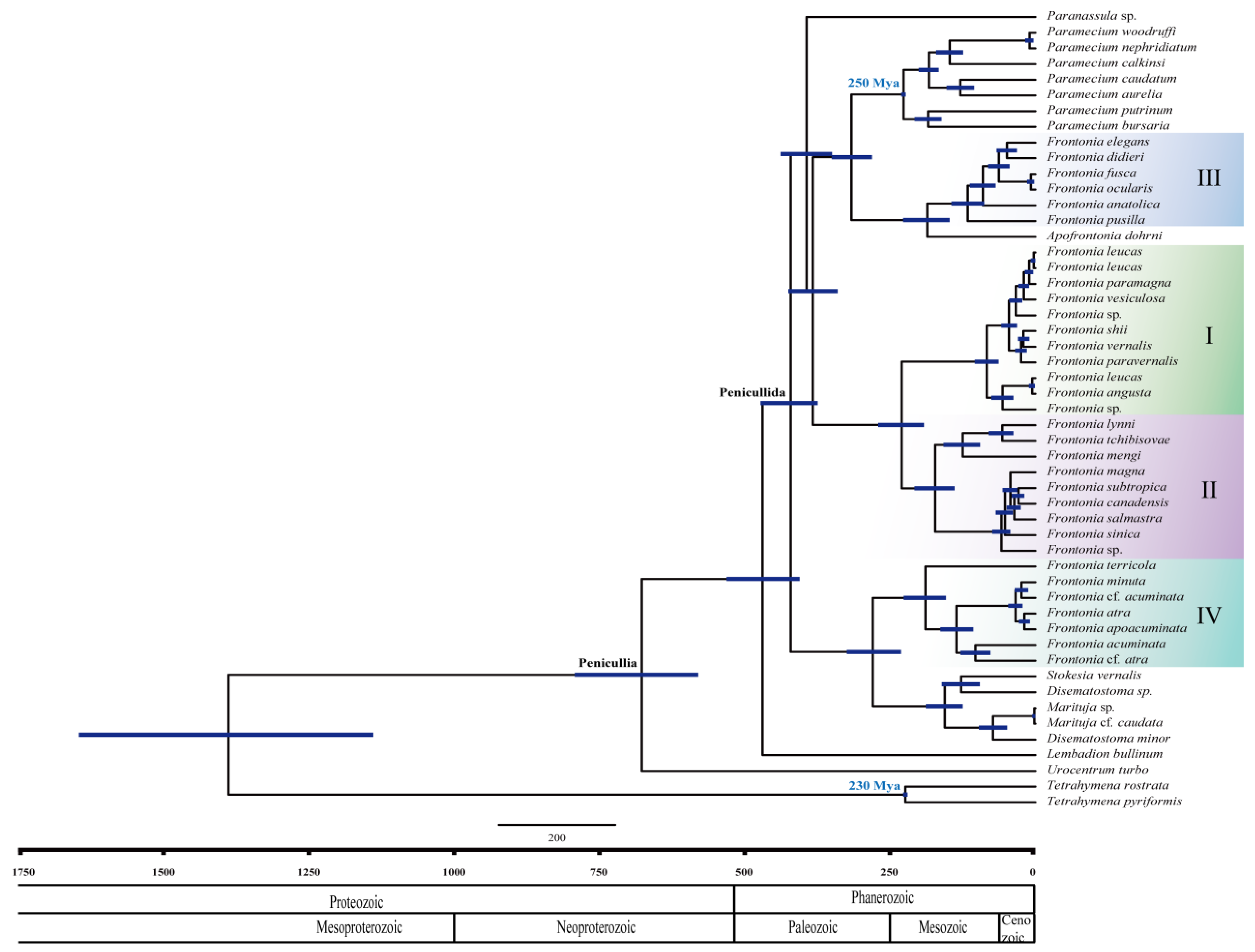
Figure 3.
Ancestral state reconstruction of morphological characteristics; A) vestibular kinety (VK) number, B) postoral kinety (PK) number, C) peroral membrane (PM) row number, D) peniculi 1 (P1) row number, E) peniculi 2 (P2) row number, F) peniculi 3 (P3) row number, G) linearity of peniculi 3 (P3) in relation to peniculi 1 (P1) and peniculi 2 (P2), H) length of peniculi 3 (P3) in relation to peniculi 1 (P1) and peniculi 2 (P2) (shortened), I) F. shii [33] (right) as an example of species in which P3 has the same structure as P1 and P2, F. paramagna [33] (middle) as an example of species in which P3 does not have the same structure as P1 and P2, and F. canadensis [34] (left) as an example of species in which P3 is shortened compared to P1 and P2. *0: no, 1: yes.
Figure 3.
Ancestral state reconstruction of morphological characteristics; A) vestibular kinety (VK) number, B) postoral kinety (PK) number, C) peroral membrane (PM) row number, D) peniculi 1 (P1) row number, E) peniculi 2 (P2) row number, F) peniculi 3 (P3) row number, G) linearity of peniculi 3 (P3) in relation to peniculi 1 (P1) and peniculi 2 (P2), H) length of peniculi 3 (P3) in relation to peniculi 1 (P1) and peniculi 2 (P2) (shortened), I) F. shii [33] (right) as an example of species in which P3 has the same structure as P1 and P2, F. paramagna [33] (middle) as an example of species in which P3 does not have the same structure as P1 and P2, and F. canadensis [34] (left) as an example of species in which P3 is shortened compared to P1 and P2. *0: no, 1: yes.
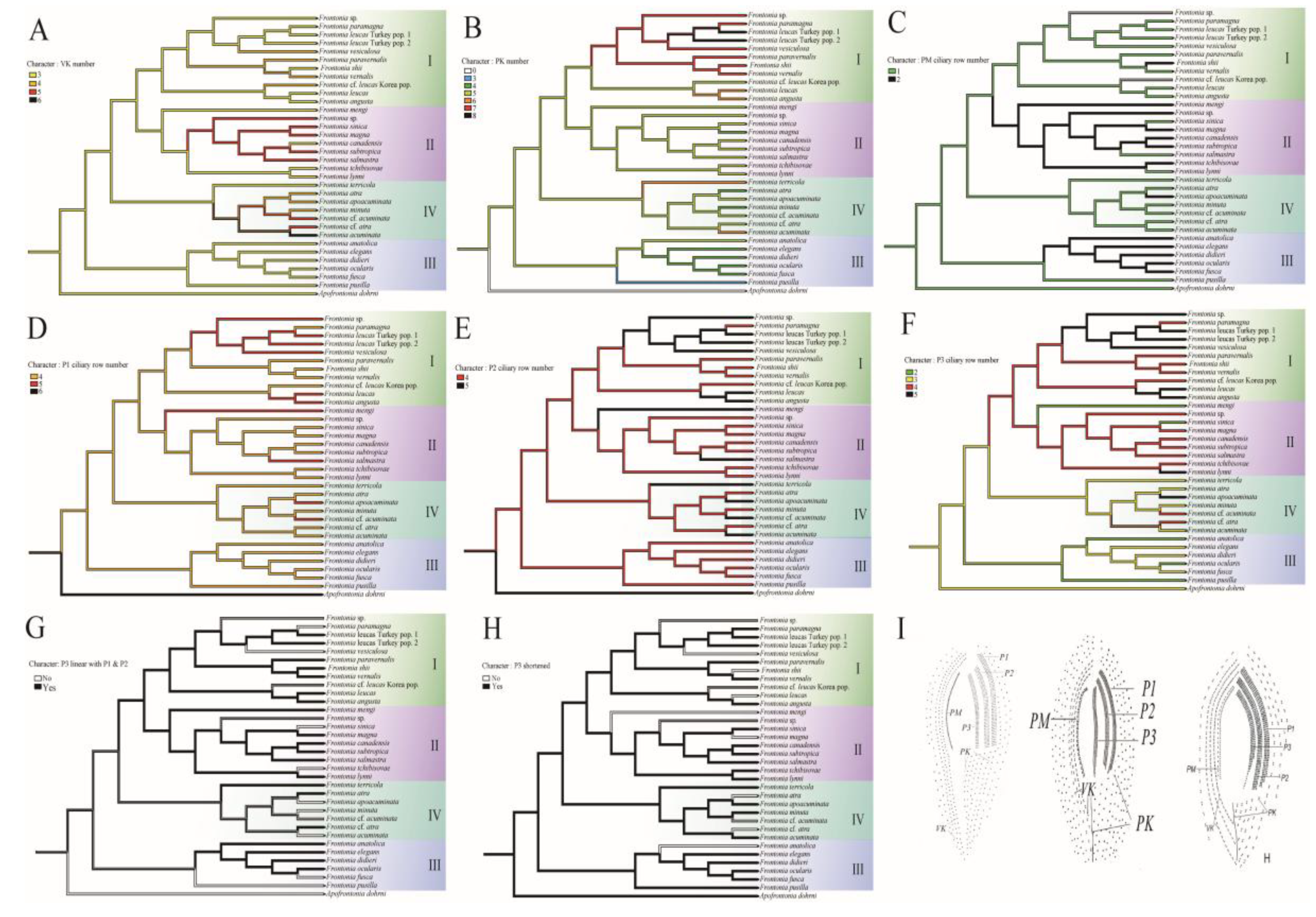
Figure 4.
Reconstruction of ancestral state of morphological characteristics; A) habitat, B) number of contractile vacuole (CV), C) number of contractile vacuolar pore (CVP) and D) presence of contractile canal (CC).
Figure 4.
Reconstruction of ancestral state of morphological characteristics; A) habitat, B) number of contractile vacuole (CV), C) number of contractile vacuolar pore (CVP) and D) presence of contractile canal (CC).
Table 1.
Frontonia species from Korea and their accession number for SSU rRNA gene sequence.
| Taxon | Collection site | accession no. |
|---|---|---|
| Frontonia cf. acuminata pop1 | Gwangju, Korea | PV016905 |
| Frontonia cf. acuminata pop2 | Ulsan, Korea | PV016906 |
| Frontonia cf. acuminata pop3 | Gunsan, Korea | PV016907 |
| Frontonia cf. acuminata pop4 | Ulsan, Korea | PV016908 |
| Frontonia cf. atra pop1 | Daejon, Korea | PV016909 |
| Frontonia cf. atra pop2 | Gunsan, Korea | PV016910 |
| Frontonia cf. atra pop3 | Masan, Korea | PV016911 |
| Frontonia cf. atra pop4 | Chungcheongbukdo, Korea | PV016912 |
| Frontonia cf. atra pop5 | Gyeongsangnamdo, Korea | PV016913 |
| Frontonia cf. atra pop6 | Ulsan, Korea | PV016914 |
| Frontonia cf. atra pop7 | Gyeongju, Korea | PV016917 |
| Frontonia cf. leucas | Gwangju, Korea | PV016904 |
| Frontonia sp. 1 | Ulsan, Korea | PV016915 |
| Frontonia sp. 2 | Ulsan, Korea | PV016916 |
| Frontonia sp. 3 | Gyeongju, Korea | PV016918 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



