Submitted:
10 February 2025
Posted:
11 February 2025
You are already at the latest version
Abstract
Small mammal species assemble based mainly on plant communities but the structure of their assemblages is shaped by several other factors, modified by larger-scale geography. Despite their ecological significance, they are often neglected elements of biodiversity, therefore, even several Polish national parks have lacked extensive surveys. It also applies to the Wolin National Park (WNP), the only one located on a coastal marine island, known previously for its unique bat fauna. We surveyed small mammals of WNP by live- and pitfall trapping, identifying only nine species, the lowest species richness among five national parks of the region (the remaining hosted 11-13 species, based on trapping data only) and the rarefaction curve showed very low probability of increasing that number by future sampling. Such unexpected paucity of species is probably linked to insular isolation and the location of the park at the edge of regional distributions of the three species. Cluster analysis revealed the most peculiar feature of small mammal assemblages in WNP - a division of the Park into two landscape units, moraine hills and the alluvial delta, where predominant murids were, respectively, Apodemus flavicollis and Apodemus agrarius. That divergence appeared to affect the clustering more than vegetation at the trapping sites.
Keywords:
rodents
; shrews
; Rodentia
; Soricidae
; habitat preferences
; species richness
; Baltic
; Poland
1. Introduction
Small land mammals, especially rodents and shrews, play crucial functions in trophic networks and soil processes in continental terrestrial ecosystems worldwide [1]. They also provide the largest contribution to mammalian diversity globally, along with bats [2,3]. Generally, different habitats, defined e.g. by plant communities, are occupied by different small mammal assemblages, usually consisting of several, co-occurring species [4]. The qualitative and quantitative structure of those assemblages varies with an ensemble of environmental factors - in the temperate zone of Europe primarily composition of herbaceous [5] and tree layer [6], tree cover [Benedek and Lazar 2021], abundance of coarse woody debris [Fauteux et al. 2012], productivity [6], altitude [7], topography, micrometeorology and abundance of epigeic invertebrates [8], large-scale disturbances, including fire [9] and post-fire management [10], activity of keystone species, particularly ecosystem engineers [11,12], intensity of agriculture [13], forest management [14,15] and animal-husbandry-related land use [16]. Effects of those factors may, however, become modified by larger-scale geographical effects, like insular isolation [17]. The combination of multiple factors leads, therefore, to the variable structure of small mammal assemblages even among areas with robustly similar habitat composition, and that variation remains only partially understood, probably due to numerous interactions between those factors.
National parks remain crucial elements of protected area networks globally, contributing significantly to the conservation of biodiversity [18]. In Poland, they are the only protected areas in which all nature is subjected to protection. Therefore, they should be prioritized as subjects of broad-scale inventories of biodiversity, especially those focused on taxonomic groups playing keystone functions in ecological processes. Contrary to more charismatic, large and middle-sized mammals, which were already surveyed in all national parks of Poland, as well as subjected to constant monitoring [19], small rodents and shrews evaded similar attention, so no sufficient inventories of that group were undertaken in several national parks until recently. This has also been the case of Wolin National Park (WNP), located at the northwestern edge of Polish territory and the only national park in the country located on a coastal marine island. This unique position among Polish national parks may result in exceptional biogeographical features that also affect the structure of small mammal assemblages. WNP is known to host unexpectedly depauperate bat fauna, with the structure of their assemblages uniquely uniform among habitats due to the dominance of single, hyperabundant native species [20]. The small mammal fauna of that Park lacked similar insight.
That paper aims to present the structure of small mammal assemblages in a Central European national park that revealed unexpectedly depauperated species composition, compared to regional fauna and local variability in land cover. We also tested if the inter-assemblage similarity of small terrestrial mammals is shaped by plant communities or robust topography in a specific context of coastal landscapes of the Southern Baltic Sea region.
2. Materials and Methods
2.1. Study Area
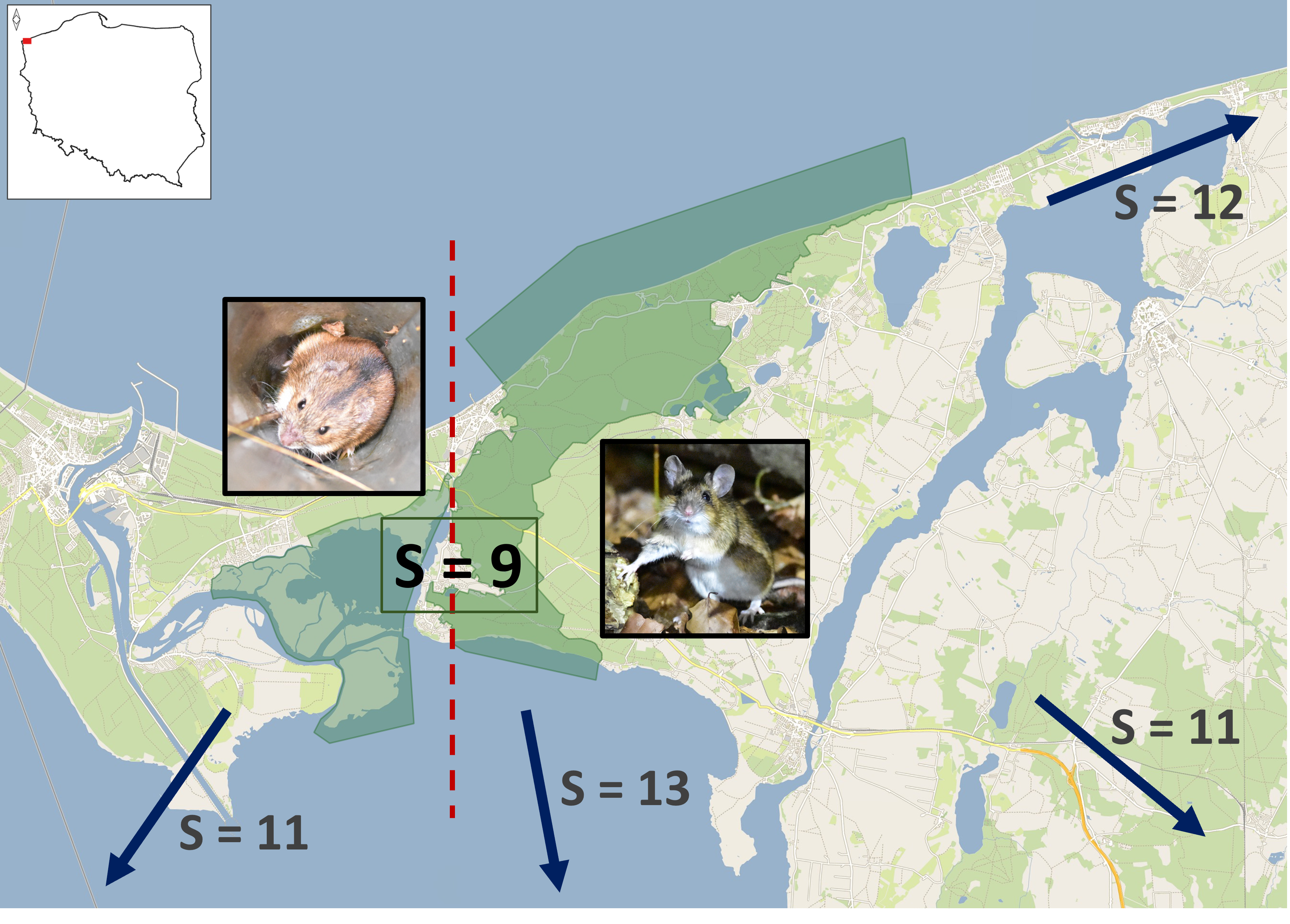
Wolin National Park (WNP) is located along the Baltic Sea coast in northwestern Poland, 53°57′15″N, 14°29′20″E (Figure 1). The park encompasses a significant portion of Wolin Island, the largest Polish island, which covers 265 km². It is separated from the mainland by the Dziwna Channel, which narrows to 90 meters at its least wide point and expands to approximately 4.5 kilometers at its widest.
Established in 1960, WNP covers 10937 hectares, with 4,648 hectares (42.5%) comprising forest ecosystems. Most of the park's woodlands form a continuous block that includes a range of moraine hills, with the highest point reaching 115 m asl. To the north, these woodlands are bordered by active cliffs that rise to 93 meters, composed of clay, sand, and gravel, which drop directly into the Baltic Sea. Lower cliffs, also partially active, define the southern boundary and fall into the Szczecin Lagoon, while dunes can be found to the northeast. The predominantly moraine, woodland part of the Park is adjacent to the flat, alluvial part, located in the regressive delta of the Świna channel, 100-1000 m wide, separating Wolin from the neighboring Uznam Island. Seven lakes, including four postglacial and three post-mining, are located within the hilly part of the park, covering a total surface of 163 ha. Running waters are extremely scarce, restricted to four short streams, discharging the lakes. A significant element of the hydrological system in the delta is a network of brackish channels and open waters of the Szczecin Lagoon, which form the estuary of the Oder River and divide part of the alluvial lands into seven smaller islands. Frequent backflows from the Baltic Sea lead to flooding events in that part of WNP.
According to the Köppen climate classification, the region has a humid continental climate with a warm summer subtype (Dbf). The mean annual temperature is 9.1 °C, with a mean temperature in January of 0.8 °C and a mean temperature in July of 18.2 °C. The mean annual precipitation measures 585 mm, and the area typically experiences an average of 30 days of snow cover each year.
In WNP, the average age of the tree stands is 112 years. They are predominated by Scotch pine Pinus sylvestris, beech Fagus sylvatica, and sessile oak Quercus petraea [https://wopn.gov.pl/statystyka]. The predominant woodland communities are fertile (Galio odorati–Fagetum) and acid-poor (Luzulo pilosae–Fagetum) beech forests, with a significant contribution of acid beech–oak (Fago–Quercetum), birch-oak (Betulo-Quercetum) and suboceanic pine (Leucobryo–Pinetum) forests. Crowberry-pine forests (Empetro nigri-Pinetum) grow on coastal dunes, while sedge-beech (Carici-Fagetum) on the upper edges of marine cliffs. Swamp forests consist almost exclusively of black alder carrs (Ribeso nigri-Alnetum), which occur both around postglacial lakes and on alluvial delta. Compared to the other parts of northern Poland, especially the Pomerania region, the characteristic feature of the Park vegetation is an almost complete lack of moist subatlantic oak-hornbeam (Stellario-Carpinetum) and alder-ash (Fraxino-Alnetum) forests, which are represented only by single patches near postglacial lakes. Among non-forest communities, saltmarsh pastures (Juncetum gerardi), on islands of alluvial delta, are the most noticeable. Significant areas are covered also by reed (Phragmitetum commune) and great fen-sedge (Cladietum marisci) beds. Glycophilous grasslands are represented mostly by false oatgrass (Arrhenaterion elatioris) and purple moor grass (Molinion) meadows. Another characteristic feature of WNP, as one of the two Polish national parks protecting coastal habitats, are diverse and well-developed cliff communities, formed both as swards and shrubs in various stages of succession. On the contrary, non-forest dune communities are poorly represented, with only a single, narrow patch in the central part of the Park coast, where typical zonation from white (Elymo-Ammophiletum) to grey (Helichryso-Jasionetum) dunes occurs [Piotrowska 1966, 1998]. Only a few buildings are present in the WPN, mostly Park offices, foresters’ lodges and - in the delta - cow sheds. However, several settlements, including one city (Międzyzdroje), are located at the Park borders.
2.2. Small Mammals Survey
Data used in the following study were collected during the first-ever broadscale survey of small mammals of the WNP in August and September 2023 on 14 localities (Figure 1), attributed to 17 different terrestrial and wetland habitats of the Park (Table 1).

Table 1.
Habitat characteristics of the study sites at which small mammal trapping was conducted. Landscape units: mh - moraine hills (uplands), ad - alluvial delta (lowlands).
Table 1.
Habitat characteristics of the study sites at which small mammal trapping was conducted. Landscape units: mh - moraine hills (uplands), ad - alluvial delta (lowlands).
| Site | Landscape unit | Habitat characteristics |
| I | mh | Alder-ash riparian forest Fraxino-Alnetum (70 years) along a stream between two lakes, transitioning to the ecotone of black alder swamp forest and reed bed along the shore of eutrophic lake |
| II | mh | Treeless communities on coastal dunes; the site divided into two shorter trap-lines of equal length: IIa – grey dunes Helichryso-Jasionetum, IIb – white dunes Elymo-Ammophiletum |
| III | mh | Mesic meadow Arrhenatherion elatioris on a hill slope, regularly mowed; few scattered shrubs near the trap-line |
| IV | mh | Black alder swamp forest Ribeso nigri-Alnetum (45 years) along the shore of eutrophic lake; hollows filled with water, locally patches of reed Phragmites australis in the herbaceous layer |
| V | mh | Moist subatlantic oak-hornbeam forest Stellario-Carpinetum (40-165 years) on a lakeside terrace and parallel slope; abundant hazelnut Corylus avellana in the undergrowth |
| VI | ad | A mosaic of wet, glycophilous meadows, low reed, and sedge beds, adjacent to the narrow (50-60 m) stripe of trees (pedunculate oaks, black alders) along the ditch; during the trapping meadows were freshly mowed and partially flooded |
| VII | ad | A dike (5-10 m wide) between the two canals, covered by a mosaic of sedge communities and willow shrubs, surrounded by black alder swamp forests Ribeso nigri-Alnetum (96 years) from both sides |
| VIII | mh | Active marine cliff with mosaic of early (Trifolio-Anthyllidetum swards with abundant grasses and field wormwood Artemisia campestris) and later (sea-buckthorn Hippophae rhamnoides shrubland) succession stages |
| IX | mh | Surrounding of forester’s lodge – traps located along buildings’ walls, in the garden, on piles of firewood, under shrubs, and in an orchard |
| X | mh | Fertile, woodruff beach forest Galio-odorati Fagetum (125-165 years) with abundant coarse woody debris of natural origin |
| XI | mh | A complex of acidophilic, broadleaved woodlands (90-165 years); the site divided into two shorter trap-lines of equal length: IIa – acid-poor beech forest Luzulo pilosae-Fagetum with traces of recent active restoration (mostly removal of planted Scotch pine) resulting in abundant woody debris, IIb – acid-poor oak-beech forest Fago-Quercetum |
| XII | mh | A complex of coastal woodlands on dunes; the site divided into two shorter trap-lines of equal length: XIIa – pine forest Empetro nigri-Pinetum (52 years), XIIb – acidic birch-oak forest Betulo-Quercetum (65-135 years) |
| XIII | ad | Halophytic mire Glauco-Puccinietalia, consisting of low sward dominated by Juncus gerardi, intensively grazed by cattle, and patches of higher vegetation, including Schoenoplectus tabernaemontani along a flooded depression. |
| XIV | ad | Reed bed Phragmitetum commune, consisted mostly of high reeds, forming an ecotone (10-40 m wide) between birch-oak forest and mowed, glycophilous meadows |
The methodology was originally designed to maximize the completeness of the species inventory for the Park, thus we allowed minor methodological differences among localities. In general, at every locality, 30 pitfall traps and 17 Sherman live-traps were set. Pitfall trapping followed the methodology used in [12]. The bottom of each pitfall was filled with a ball of local herbaceous plants or moss to improve thermal conditions and 2-3 larvae of mealworm Tenebrio molitor were put in it as a food supply. Both plant stuffing and mealworms were put there to improve the survival of captured shrews. Each live-trap was supplied with a bait, consisting of oat flakes mixed with peanut butter. Traps were set in lines, consisting of trap-points located every 10 m. Each of the first 17 trap-points included a pitfall and a live-trap, located 1 m from each other, later – only a pitfall. When the locality was intended to include two different, neighbouring habitats (II, XI, XII), two parallel trap-lines were set, each consisting of 15 pitfalls and 8-9 live traps. Trapping at each location was usually conducted during two consecutive nights. Traps were opened an hour before sunset and closed an hour after sunrise. The exceptions were localities II, VIII, IX, and XI, where traps were set only for one night. At site V, where we aimed to capture the hazel dormouse Muscardinus avellanarius, live-traps were set in a parallel trap-line from pitfalls, all mounted to branches of hazelnut Corylus avellana, about 1-1,5 m above the ground, while the bait was supplemented with sliced apples. Only live-traps were set at localities VIII and IX, due to hard soils, making setting pitfalls too time-consuming and destructive for the plant cover. All traps were checked every two hours, while captured mammals were identified based on external features, mostly based on the key by Pucek [21], and released on the spot. Some individuals of Apodemus and Microtus/Alexandromys remained identified only to the genus or genus-pair level, as they managed to escape before measurements were taken or non-metric features were examined. These unidentified Apodemus represented exclusively subgenus Sylvaemus [22], i.e. either A. flavicollis or A. sylvaticus (Linnaeus, 1758) but not A. agrarius. After every capture bait or food supply was replenished.
We used capture data to compute the expected species richness, based on the individual abundance rarefaction (species accumulation curve), using PAST ver. 4.07b. This method allowed the in-detail comparison of small mammal species diversity among habitat classes and five northwestern Poland’s (Wolin, Słowiński, Warta Mouth, Drawieński) and northeastern Germany (Lower Oder Valley) national parks [23,24,25,26,27]. To compare the species composition of samples among sites, habitat classes, and topographies (landscape units), we performed cluster analysis in ClustVis, an open-access web tool for visualizing the clustering of multivariate data, provided by the University of Tartu. Samples characterized by relative abundances of small mammal species were clustered with the average linkage method, using the average distance of all possible pairs, with the Manhattan distance as a measure of similarity and the ‘tightest cluster first’ as the method of tree ordering. As individuals were not marked, the relative abundance was calculated based on the number of captures. We divided the Park and its habitats into two landscape units - one represented moraine hills (upland) in the eastern part (10 sites, 13 habitats), and another represented the alluvial, regressive delta in the west (4 sites and habitats). The sites were attributed to four habitat groups: meso- and acidophilic forests on mineral soils (V, X, XI, XII), dry non-forest habitats (II, III, VIII, IX), both exclusive to the upland part of the Park, upland wetland forests, associated with shores of postglacial lakes and streams linking them (I, IV), and exclusively wetland habitats of alluvial delta (VI, VII, XIII, XIV).
3. Results
In total, we captured nine species of small mammals. Three of them belonged to the Soricidae family of Eulipotyphla: common shrew Sorex araneus Linnaeus, 1758, Eurasian pygmy shrew Sorex minutus Linnaeus, 1766 and Eurasian water shrew Neomys fodiens (Pennant, 1771). The remaining six were Rodentia, including four Cricetidae - bank vole Clethrionomys glareolus (Schreber, 1780), root vole Alexandromys oeconomous (Pallas, 1776), short-tailed field vole Microtus agrestis (Linnaeus, 1761), common vole Microtus arvalis (Pallas, 1778) - and two Muridae: striped field mouse Apodemus agrarius (Pallas, 1771) and yellow-necked mouse Apodemus flavicollis (Melchior, 1834). They were represented by 397 captures, among which C. glareolus, A. flavicollis, and A. agrarius were the most numerous. In total, rodents constituted 89.2% of the whole assemblage, with a predominance of Cricetidae (45.6%), followed by Muridae (43.6%). Shrews constituted only 10.8% of all captured mammals, with S. minutus being the most numerous. N. fodiens was the rarest small mammal in our sample, represented only by a single capture. No unambiguous A. sylvaticus were captured, however. The most widespread species were: A. flavicollis (10 localities, 12 habitats), C. glareolus (7 localities, 7 habitats), and S. minutus (6 localities, 6 habitats) (Table 2).
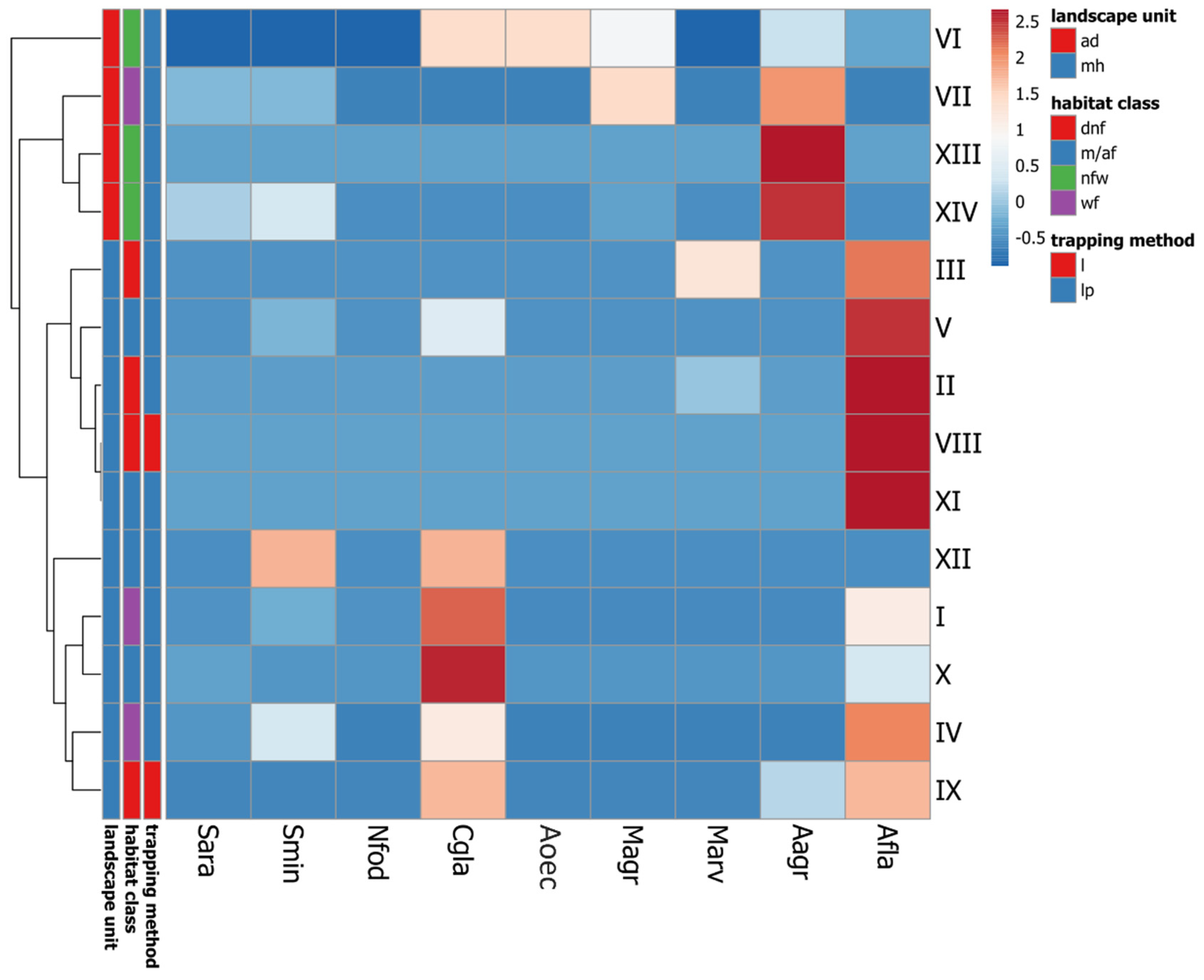
Samples from particular study plots clustered almost exclusively based on location within the two landscape units of the park (i.e. uplands and the alluvial delta) to the point that the alder swamp forest in the delta (VII) appeared much more similar to the nearby open habitats (XIII, XIV) than to swamp and riparian alder forests edging the upland lakes (I, IV). The mosaic of freshwater meadows, sedge and reed beds in the delta (VI) appeared to form a separate branch outside the cluster containing all the other habitats but it still shared much more resemblance to the other delta habitats than to any site located in the upland part of the Park. The two landscape units differed particularly by the relative abundance of the two only small murid rodents recorded (A. flavicollis and A. agrarius).
Figure 2.
Comparison of species composition of small mammals trapped at particular study sites in Wolin National Park, subjected to cluster analysis (clustering method—complete linkage, Manhattan distance, average cluster first). Only individuals identified unambiguously to the species level were taken into account. Number of every site corresponds to the Table 1. Species: Sara - S. araneus, Smin - S. minutus, Nfod - N. fodiens, Cgla - C. glareolus, Aoec - Alexandromys oeconomus, Magr - M. agrestis, Marv - M. arvalis, Aagr - A. agrarius, Afla - A. flavicollis. Landscape unit: mh - moraine hills (upland), ad - alluvial delta (lowland). Habitat class: dnf - deciduous non-forest habitats, m/af - meso- and acidophilic forests on mineral soils, nfw - non-forest wetlands, wf - wetland forests. Trapping method: l - live traps only, lp - live- and pitfall traps. Data from Table S1.
Figure 2.
Comparison of species composition of small mammals trapped at particular study sites in Wolin National Park, subjected to cluster analysis (clustering method—complete linkage, Manhattan distance, average cluster first). Only individuals identified unambiguously to the species level were taken into account. Number of every site corresponds to the Table 1. Species: Sara - S. araneus, Smin - S. minutus, Nfod - N. fodiens, Cgla - C. glareolus, Aoec - Alexandromys oeconomus, Magr - M. agrestis, Marv - M. arvalis, Aagr - A. agrarius, Afla - A. flavicollis. Landscape unit: mh - moraine hills (upland), ad - alluvial delta (lowland). Habitat class: dnf - deciduous non-forest habitats, m/af - meso- and acidophilic forests on mineral soils, nfw - non-forest wetlands, wf - wetland forests. Trapping method: l - live traps only, lp - live- and pitfall traps. Data from Table S1.
Small mammal assemblages in the upland, the eastern part of the park were usually characterized by abundant A. flavicollis, irrespectively of tree cover and humidity. In the mesophilous, deciduous forests with predominant beech and oak in the tree layer, it was outranked, however, by C. glareolus (66.0%) (Figure 3), the effect that can be attributed mostly to the effect of one site with the highest number of small mammal captures during the whole study (X, fertile beech forest, Table 2)). Assemblages inhabiting the upland wetland forests (I, IV) revealed similar dominance of A. flavicollis (34.1%) and C. glareolus (43.9%) but also relatively high dominance of shrews (in total 15.2%, all three species) (Figure 3). The only capture of N. fodiens in the Park was obtained in the alder-ash riparian forest (I). A. agrarius was recorded in the upland part of the Park based on single capture in the garden around the forester’s lodge (IX) (Table 2). The only habitats occupied by M. arvalis were dry non-forest ones (Figure 3), particularly grey dunes (IIa) and mesic meadows (III). Habitats closest to the seashore were occupied only by the two species, A. flavicollis (white dunes, IIb; treeless cliffs, VIII) and S. minutus (coastal dune pine forest XIIa) (Table 2).
On the contrary, wetland habitats of the alluvial delta, i.e. in the western part of the Park, were almost devoid of A. flavicollis (Figure 3), represented there by only a single capture at site VI (Table 2), i.e. located closest to the moraine, hilly uplands. Instead, small mammal assemblages inhabiting that area are characterized by abundant A. agrarius (55.6%) (Figure 3). It was present at all sites, both woodland (VII) and treeless or ecotone ones (VI, XIII, XIV). In the halophytic mires (saltmarsh, XIII) it was the only small mammal recorded (Table 2). A notable feature of small mammal fauna in the delta wetlands was the presence of M. agrestis (three out of four habitats) and the highest relative abundance of shrews among all four habitat groups of the park (20.9%), represented, however, solely by representatives of the genus Sorex (Figure 3). The only habitat occupied by M. oeconomus in the Park was the complex of wet, freshwater meadows and sedge and reed beds (VI) (Table 2).
Figure 4.
Rarefaction curves for the cumulative numbers of species plotted against the numbers of small mammal captures in four different groups of habitats, distinguished by plant communities and topography. Meso- and acidophilic forests on mineral soils: sites V, X, XI, XII. Upland wetland forests: sites I, IV. Dry non-forest habitats: sites II, III, VIII, IX. Wetland habitats on the alluvial delta: sites VI, VII, XIII, XIV.
Figure 4.
Rarefaction curves for the cumulative numbers of species plotted against the numbers of small mammal captures in four different groups of habitats, distinguished by plant communities and topography. Meso- and acidophilic forests on mineral soils: sites V, X, XI, XII. Upland wetland forests: sites I, IV. Dry non-forest habitats: sites II, III, VIII, IX. Wetland habitats on the alluvial delta: sites VI, VII, XIII, XIV.
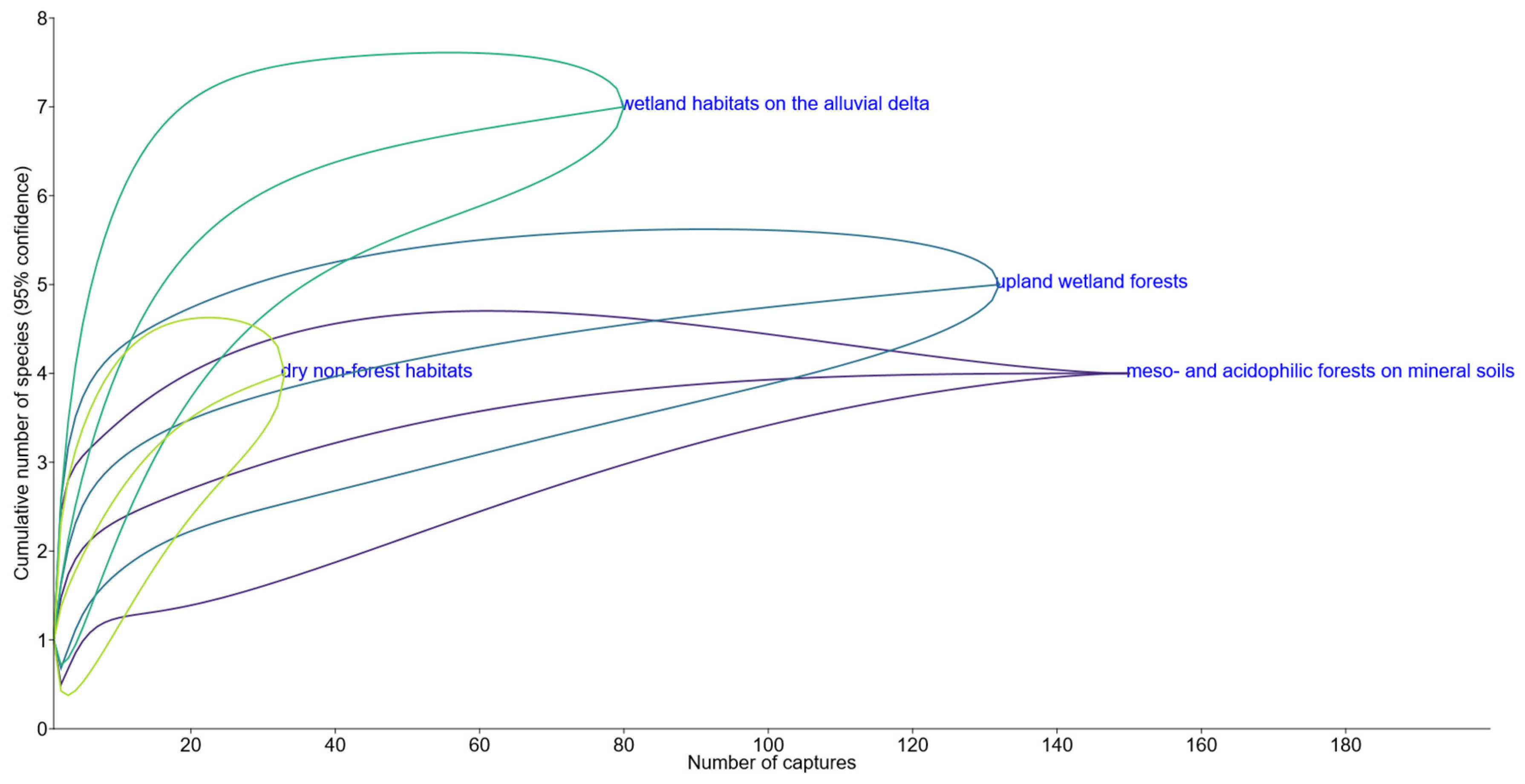
The species accumulation curves revealed the highest diversity in the wetland delta habitats (7 species), followed by upland wetland forests (5); meso- and acidophilic forests on mineral soils appeared to host the poorest assemblages (4). In the majority of habitat groups, curves reached the plateau, suggesting a very low probability of adding any new species if the sample size increases, only the dry non-forest habitats (4 species) remained undersampled (Figure 4).
Wolin NP hosted the poorest small mammal fauna among the four nearby national parks that were compared using species accumulation curves. Lower Oder Valley and Drawieński NP hosted 11 and Słowiński NP hosted 12 species but the difference between them was negligible, with nearly identical trends and strongly overlapping confidence intervals. None of these four parks appeared to be undersampled, as their species accumulation curves reached a plateau already before the sample size reached half of the final number of captures. In the case of WNP, the final number of nine species had been already recorded by 188 captures. Warta Mouth NP hosted the most diverse small mammal fauna, with 13 species, obtained with the lowest sample size (N=80). The species accumulation curve for that park was the only one that did not reach a plateau, therefore the sample is far from saturated (Figure 5).
4. Discussion
4.1. Species Composition
The individual rarefaction curve for the small mammal fauna suggests it is highly unlikely that the number of species would increase significantly if the sampling effort had increased. Thus, nine species recorded during our study probably represent the actual richness of local assemblages. However, it reveals unusually depauperate fauna, compared to the four other national parks of the Lower Oder catchment and Polish Baltic Sea Coast. Seven more species were trapped in those parks, specifically Neomys milleri Mottaz, 1907 (SNP), Crocidura suaveolens (Pallas, 1811) (WMNP, LOVNP), Arvicola amphibius (Linnaeus, 1758) (WMNP), Rattus norvegicus (Berkenhout, 1769) (SNP), Mus musculus Linnaeus, 1758 (DNP), Micromys minutus (Pallas, 1771) and Apodemus sylvaticus (Linnaeus, 1758) (all four other national parks) [23,24,25,26,27]. Five of them were recorded before on the Wolin Island by either trapping or owl pellet analysis, these were A. amphibius, M. musculus, M. minutus, A. sylvaticus [28,29], and R. norvegicus [30]. The most recent, published dates for the occurrence of the first three species on the island are 1962 and 1966, during the extensive survey by the Mammal Research Institute, Polish Academy of Science, and they were reported from 3-7 localities, with M. minutus being the most widely distributed among them [29]. Recently it is usually hard to uncover if those records were obtained from the area recently located within the borders of WNP, as the authors provide only the names of towns and villages, where the material comes from and these settlements are located outside the Park. The exception is A. sylvaticus, whose specimens in the zoological collection of the Mammal Research Institute PAS in Białowieża (no. 61932, 61819, 61723, 61624), are labeled as collected in Wolin National Park in 1966 [31], probably identical with wood mice reported as trapped in the same year in Międzyzdroje [29]. R. norvegicus was recorded by us in August 2023, based on a dead individual found at the lake shore, close to site V [32]. The occurrence of Talpa europaea Linnaeus, 1758 on Wolin Island was mentioned in two sources [33,34] and confirmed, indeed, from the Park itself, by observation of molehills near the sites I and V [32]. Moreover, two arboreal glirid rodents, Glis glis (Linnaeus, 1766) and Muscardinus avellanarius (Linnaeus, 1758) were reported from Wolin Island in the first half of the 20. century [33,35,36], despite not being trapped in any of the national parks of the Lower Oder catchment and Polish Baltic Sea Coast [23,24,25,26,27]. It should be noticed however that G. glis was recorded later in Słowiński National Park through direct observation [37]. Finally, the occurrence of Microtus subterraneus (de Sélys-Longchamps, 1836) was reported from the south-eastern shore of Szczecin Lagoon, about 20 km from the borders of WNP [29], despite not being found in any of the four national parks mentioned above. Based on that data, potential small mammal fauna of WNP may consist of even 19 species but 11 were recorded recently, and only nine - were captured during trapping on 15 sites representing all major habitats. This raises questions about factors behind such paucity of species in recent small mammal assemblages of the Park, comparable to, recently discovered, similarly depauperate bat assemblages (20).
4.2. Potential Methodological Bias Affecting the Structure of Small Mammal Samples
One of the factors behind the differences in the structure of regional small mammal assemblages is the application of traps that are too selective and reduce the detection rate of particular taxa. Usually, small terrestrial mammals are sampled with traps belonging to two types – traps with bait and trigger (live- and snap traps) and pitfalls. Some papers claim that live traps are more efficient in catching rodents, and pitfalls in catching shrews [38; 39]. However, other studies revealed that only snap and live-traps underestimated the abundance and dominance of shrews, while the efficiency of rodent trapping with all three types of traps remained similar [40; 41]. In some extremely diverse biomes, using pitfalls alone may even generate higher estimates of overall small mammal abundance and species richness, as well as rodent abundance and/or species richness than using not only live-traps alone but also both types of traps together [42]. The study in the European boreal zone already revealed that the application of cone traps provides a much more balanced picture of small mammal assemblage, providing abundant material of both shrews and rodents. On the contrary, snap traps captured not only three times less mammals but also material that remained strongly rodent-biased. The difference was even stronger when analyzing particular species, with 70 times higher for the smallest European rodent, M. minutus, in pitfalls than in snap traps [43]. A study on small mammal assemblages inhabiting beaver-created wetlands in Poland provided abundant material of both Central European Neomys species (extremely scarce in WNP) and M. minutus (not detected at all in WNP), despite being based solely on pitfalls as a trapping device [12]. Some species might, indeed, predominantly A. flavicollis, remain undersampled with applications of pitfalls only due to their ability to jump out of the trap [43,44], even if they remain regularly recorded in pitfall-only trapping studies [5,12]. To avoid that potential bias, we applied both live- and pitfall traps in most sites, if topography allowed such an approach. Small mammal surveys in three other national parks, compared to WNP in our study, were based on either solely wooden live traps [SNP - 25, DNP - 23], a combination of wooden and metal live traps [LOVNP - 27] or combination of wooden live traps and pitfalls [WMNP - 24]. May one, therefore, expect that samples from SNP and LOVNP would reveal an underrepresentation of shrews, compared to WNP. Despite that, all that parks were characterized by much higher contribution of shrews to small mammal assemblages (SNP - 34.6% [25], WMNP - 28.8% [24], LOVNP - 21.23% [27], DNP - 18.3% [23]) than in WNP (10.8%) where the application of pitfalls was the most widespread. Thus, the composition of trap types in trapping schemes was probably not a reason for the paucity of species recorded in WNP, including the scarcity of Neomys water shrews and the absence of M. minutus. Despite some claims [45], above-ground live trapping is not necessary for the detection of M. minutus, as the species is regularly captured in ground-level traps [5,12,25,43], thus not applying specific trapping method remains an unlikely explanation of that species’ absence, even if trapping efficiency for the harvest mouse appeared higher in elevated traps than in-ground ones [46].
Neither pitfalls nor live traps are effective trapping devices for the detection of T. europaea and Gliridae, even if they fall, sometimes, into pitfalls [5,47]. Their absence in the WNP sample can be explained by systematic bias in our methodology but these species were neither captured in any other part of the region [23,24,25,26,27]. It is worth noticing, however, that even live trapping dedicated to M. avellanarius (live traps set above the ground, on hazel branches) in its optimal habitat in WNP (site V, oak-hornbeam forest - [48]) failed to detect it. It must be also stressed that R. norvegicus is rarely captured in typical small mammal traps [25,49], probably due to large body size, despite widespread occurrence across the whole Polish territory [29]. However, even if taking into account only the species for which the applied method is considered optimal (i.e. epigeic taxa with body size lower than R. norvegicus), still may one expect to capture 15 species of small mammals, based on their geographical ranges and habitat preferences.
4.3. Factors Leading to Depauperation of Small Mammal Assemblage
The exceptionally low number of species in small mammal fauna of WNP would be explained by the geographical position of the Park if it is located outside of geographical ranges of missing species, recorded in the other national parks of Lower Oder catchment and Polish Baltic Sea Coast. However, it is highly unlikely that biogeography plays a major role here. Most species that were present in the mentioned region but not recorded in WNP cover the entire Poland and Germany by their ranges, this applies to A. amphibius, M. minutus, M. musculus, and A. sylvaticus [29,50]. Nevertheless, three taxa reach their border of distribution near the Oder estuary and their absence in WNP (and - historically - on Wolin and Uznam Islands in general) may be explained by that fact. N. milleri reveals a disjunctive distribution pattern in Europe, with an isolated Pomeranian part of its range [51], ending just at the western Polish border (coinciding roughly with Lower Oder Valley) and not extending to northern Germany at all [50]. The nearest known localities of N. milleri were reported about 30-40 km to NE and SE from the borders of WNP, on the Pomeranian mainland [29]. Among the four compared national parks, only SNP, located furthest to the east, appeared to host a population of N. milleri [25]. Also, M. subterraneus does not occur in northern Germany, thus the western border of its distribution on Central European lowlands almost overlaps with the Polish western national border [50]. C. suaveolens in Poland is widely distributed across the southern and western part of the country but it reaches the northern edge of its range in mainland western Pomerania, with its northernmost record in the region of the Oder estuary located about 30 km from the borders of WNP [52]. M. subterraneus reveals a very patchy distribution in northern Poland, while A. sylvaticus is much rarer and more local in the northern than in the southern part of Poland [29], which can contribute to the lack of these species in recent small mammal samples from WNP. It should be kept in mind, nonetheless, that A. sylvaticus was already recorded in the Park [31]. Even if regional borders of distribution can exclude 3-4 taxa from potential fauna of WNP, still local small mammal assemblages should consist of at least 12 species.
Differences in habitat composition and land cover may account for differences in species composition and diversity of small mammal fauna among compared national parks. It is, however, unlikely in the case of WNP. M. minutus, the most noticeable rodent species, common in the whole of Poland and occurring in all the other national parks of the region, but absent in WNP, clearly would find its preferred habitats here. It selects non-forest habitats with high herbaceous vegetation, including wetland ones, especially reed beds [46,53] and sedge communities [5], but particularly sparse reeds with sedge undergrowth [54], which are abundant in the lowland part of the Park, associated with alluvial delta and represented by the sites XIV and VI. A. amphibius selects simply water bodies and wetland habitats, irrespectively of dominant vegetation [55] and such habitats cover more than a half of the whole park area and are represented by six sites in our sample (I, IV, VI, VII, XIII, XIV). Similarly, the extreme scarcity of N. fodiens, which contributed to generally low species diversity, as revealed by species accumulation curves, requires explanation, as the species is just associated with water [5,56] and various types of wetlands, both forest and non-forest ones. In Poland, it is especially numerous in black alder swamp forests [47,57], reedbeds, and sedge communities [57,58]. Except for site I, where it was captured, it might be expected to occur in abundance in sites IV, VI, VII, and XIV, and at least some of them might also host N. milleri if it occurs in the Park. A. sylvaticus is a species associated with dry, open, or semi-open habitats, including xerothermic swards [47,59], shrubs (e.g. hedgerows), ruderal communities and arable land [47,60]. Several features of that habitats are represented in sites VIII (active marine cliff) and III (mesic meadow) but A. sylvaticus is known also to occupy wetland habitats [61], forests and their ecotones [62], albeit in lower numbers. Therefore we sampled a broad spectrum of potential habitats of that eurytopic species, yet all individuals representing subgenus Sylvaemus that we managed to examine in hand revealed unambiguous morphological features of A. flavicollis. The only group of small mammals that may indeed suffer from the scarcity of their specific habitats are the synanthropic species (M. musculus and C. suaveolens) as rural buildings and their surroundings, e.g. gardens (site IX), are only a few within the borders of the Park and may not be able to sustain their populations. General habitat parameters that are known to be positively correlated with small mammal diversity and density (percentage of actual deciduous tree stands [6] or abundance of coarse woody debris [63,64] do not seem to be factors responsible for paucity of species in the WNP, as the majority of forest cover is composed of broadleaved species, while recent restoration efforts and long term passive protection in the oldest stands led to abundance of deadwood. Finally, many sampled habitats belong to either early successional stages or ecotones that are known to harbor the highest number of species elsewhere in lowland Central Europe [62].
The whole study was conducted during one season, while small terrestrial mammals often reveal long-term population cycles or irregular fluctuations. Differences in population densities between minima and maxima of these cycles can, in the case of rodents, hit one or two orders of magnitude in the temperate broadleaved forest zone of the Palearctic [65,66]. Therefore, surveys conducted during minima of particular species might not only reveal low contribution to the quantitative structure of small mammal assemblage but even not be able to detect them. M. minutus reveals irregular interannual fluctuations, sometimes with maximum increases over five-fold and maximum decreases about fifteen-fold [67]. Although we may have conducted our trapping during a year when the population numbers of M. minutus were low, it's important to note that these minimum numbers do not appear to drop as significantly as the populations of other microtine species or woodland rodents that experience fluctuations associated with mast seeding [65,66]. In Lithuania, the long-term stability of the M. minutus population was revealed by trapping and owl pellet analysis, with only little interannual variation [68]. Both surveys in the compared national parks and other surveys in optimal habitats of M. minutus in northern Poland were conducted in various years but did not fail to detect that species (1997 - 69, 1998 – [23], 2012-2013 – [24], 2008-2014 – [25], 2021 – [26], 2022 – [27], 2020-2021 – [12]). Shrews in Europe reveal no population cycles but some irregular fluctuations, correlated among many distant areas and attributed mostly to effects of the North Atlantic Oscillation [70]. In Lithuania, peaks of their abundance, sometimes ten-fold higher, may occur even 20 years after the minima [71]. Conducting our trapping during one of such minima may partially explain the exceptionally low proportion of shrews, compared to the other national parks of the region. Population outbreak of A. flavicollis and C. glareolus, following the mast year of beech in 2022 [32] led to their dominance in small mammal assemblages in several habitats, as high heavy seed crop usually boosts such outbreaks [72], and consequently to the low proportion of shrews in our samples. However, it does not explain scarcity or (in most cases) the absence of N. fodiens in the Park so rich in aquatic and wetland environments. Water shrews may also reveal strong interannual variation in abundance, reflected in changes of trappability even from 0 to 30 individuals/100 pitfalls but such strong fluctuations were known only from southwestern Siberia [73]. There is not much data about the population dynamics of N. fodiens in Europe but e.g. in Lithuania, the species follows no long-term population trend, while its numbers show only a limited and irregular interannual variation. Nonetheless, there are years in which there are fewer water shrews captured than expected but the difference does not exceed factor 1.3-1.4 [71].
Wolin, from a biogeographic and paleogeographic point of view, is a continental island, while a significant part of WNP is located on smaller, alluvial islands of the regressive delta. The main island itself formed in the Atlantic period (5700-8900 BP), during marine transgression of the Littorina Sea, the previous development stage of the Baltic, that led to the separation of moraine hills from the mainland and from their western counterparts, recently forming Uznam Island [74]. Surrounding brackish channels and lagoons form barriers that obstruct the immigration of small terrestrial mammals into the islands. Although the Dziwna channel is only 90 m wide in the narrowest part, it may act at least as a filter for small mammal movements. A Slovakian river 35 m wide restricted the mobility of small rodents, even if they occasionally crossed it [75] and most terrestrial small mammals do swim effectively [76,77]. The limited immigration and, consequently, lack of replenishment for local populations may lead to extinctions and species turnover, as predicted by the theory of island biogeography [78]. The diversity of insular mammals follows the same patterns as in the case of the other taxa, i.e. it is lower than on the mainland and positively correlated with island size and negatively correlated with island isolation [79,80]. These phenomena are not restricted to geologically old, marine islands distant from the shore. Islands created by the inundation of surrounded land by reservoirs created by hydroelectric dams follow a clear pattern of faunal depauperation, similar to continental, land-bridge islands, with positive species-area relationship and correlation between faunal nestedness and area, resulting from selective extinction [81]. Both diversity and probability of occurrence of particular species of small mammals inhabiting temperate and boreal zone islands located on marine shelf and postglacial lakes are affected either by area only [82,83] or both area and isolation [17,84]. Rodent and shrew populations on these islands suffer from regular extinctions at the scale of 60 [82] or even 5 years [84]. According to our knowledge, the comparable data on small mammals of land-bridge islands of the Baltic Sea were collected in Denmark. The dataset of 31 islands and 12 species revealed the negative effect of isolation on species richness and the presence of 5 species, while the nested species distribution pointed to interspecific differences in both dispersal abilities and extinction risks [85]. Winter ice improves the colonization of temperate zone islands on lakes or coastal marine waters by small, non-volant mammals [83]. However, the number of days with ice cover on the Szczecin Lagoon declined significantly during the last 70 years due to rising temperatures [86], making the dispersal of small mammals to the islands harder and harder, especially when the narrowest points of the Dziwna channel, recently crossed by bridges, are located in the urbanized zones. Scarce, mostly quantitative, historical data do point to the species turnover and either local extinctions or shifts in the structure of small mammal fauna on Wolin Island. Trapping and owl pellet data from 1962 and 1966 from Wolin Island revealed not only the presence of the three species we failed to detect but also a much more widespread occurrence of A. sylvaticus (6/7 localities) than A. flavicollis (2/7 localities) [29], the latter recently being the most widespread small mammal in WNP (10/14 localities). Thus, the low diversity of small mammals of WNP might be attributed to island isolation, probably increasing with worsening ice conditions. Similarly depauperate is the bat fauna of WNP, however, it resulted rather from a combination of natural and anthropogenic disturbances in the Park ecosystems and its neighborhood that led to the expansion of single, hyperabundant species [20], as neither area nor isolation affects the diversity of volant mammals on islands located so close to the shore [83].
4.4. Structure of Small Mammal Assemblages and Habitat Selection by Particular Species
Despite the relative paucity of species and lack of some taxa known elsewhere to be characteristic to specific habitats sampled in the Park, several features of small mammal assemblages and habitat selection of particular species resemble data obtained from the other regions of Central Europe, including Poland. The most numerous species, C. glareolus, and A. flavicollis usually predominate in mesic and acidophilic woodlands, including beech, oak, and hornbeam forests that provide abundant heavy seeds [5,47,87]. Yet, A. flavicollis, considered a woodland species, was abundant - or even dominant - in non-forest habitats as well; it should be noticed, however, that all those patches were located close to the forests. Such ecological plasticity of A. flavicollis was observed in SE Poland, where the species regularly occurred in open habitats, even dominating on arable fields [47] and the species is known to reveal density-dependent spillovers to the habitats outside woodlands during beech mast years [88].
Except for C. glareolus, microtine rodents revealed astonishingly low contribution to small mammal assemblages, compared to the previous studies from Polish lowlands if they covered substantial areas of open habitats and eutrophic wetlands [24,57,61,89]. It may relate to the low abundance of these mammals during the minimum phase of their population cycle [65]. However, their populations, and consequently their percentage in assemblages, might already stabilize at the low level, due to widespread cycle dampening throughout Europe [90]. Nevertheless, their dominance is the highest in the habitats considered typical for these species in Central Europe. M. arvalis in Poland, outside preferred arable land, is often restricted to non-forest dry habitats, like meadows [47,61] or coastal dunes [91], i.e. those occupied in WNP. On the contrary, A. oeconomus is a specialized wetland species, predominating especially in fen communities with sedges and reeds [5,57,61], characteristics much resembling the site where it was captured in the Park (VI). M. agrestis is considered a hygrophilous species that may occur in various habitats, like humid meadows on forest glades [47] and peat bogs [49] but in eastern Poland it becomes a woodland species [5]. It corresponds with its occurrence in wet meadows, reed beds, and black alder swamp forests of the delta (but not the upland ones). Its habitat preferences in WNP align with much more numerous murid, A. agrarius. The latter species, except preferred arable land, is known to occur in various, poorly defined, wetland habitats, reaching high dominance indices in reed beds and meadows [47,57], which may partially correspond with its habitats in alluvial delta on Wolin Island.
The highest diversity of small mammals in the wetland delta, predominated by early succession stages, follows a general trend in the temperate zone [92,93]. Some notable differences between our and earlier studies included a much higher proportion of S. minutus than S. araneus in most habitats. In the majority of earlier studies, the latter species is usually the dominant shrew in Central Europe [5,25,87,94,95] except either the extremely oligotrophic [49] or very wet habitats [12]. Several papers, however, mention the high abundance of S. minutus in black alder swamp forests, which sometimes leads to higher dominance of that species, compared to S. araneus [25,47,57], which corresponds with the situation observed by us in upland wetland forests of WNP.
Data from salt marsh (halophytic meadows) require special attention, as small mammals are rarely sampled in that specific habitat in Europe. Contrary to North America, where a group of unique small mammal taxa is restricted to the coastal salt marshes, no such specialists evolved in Afro-Eurasia [96]. Few species were recorded in the respective habitats of temperate Europe, which seem to be extremely depauperate if compared to freshwater, eutrophic wetlands, like fens. M. arvalis was the most common species in one Dutch coastal salt marsh, accompanied only by scarce M. musculus [97], while on the Danish island of Sprogø C. glareolus predominated, accompanied by single A. sylvaticus [98]. According to our knowledge, A. agrarius has never been reported from such a habitat before, yet in WNP it appears to be the dominant, if not sole, small mammal there. On the contrary, inland salt marshes in Slovakia, harbor 15 species of small mammals (including A. agrarius and five shrews) but the trapping was conducted there during 19 years on 7 sites [99].
4.5. Small Mammal Zonation in the Wolin National Park Forced by Topography?
A unique feature of small mammal fauna in WNP is the division of the Park into two distinct zones - moraine hills (upland), where A. flavicollis is widespread in both forest and non-forest habitats, with A. agrarius almost lacking, and alluvial delta (lowland), where A. agrarius plays a similar role, while A. flavicollis is almost absent, even in woodlands. Additionally, a secondary feature of the lowland assemblages is a significant percentage of M. agrestis, present only there, and scarcity of C. glareolus, otherwise the most numerous species in the study area. We do not know if that division is a short-term phenomenon, associated with e.g. population cycles, desynchronized among habitats, or a more permanent element of the Park biogeography. It might be shaped and maintained by the presence of natural and anthropogenic barriers between the two parts of WNP that restrict the movements of small mammals and the chance for successful immigration. These barriers consist of the city of Międzyzdroje with its densely built-up urban zone, associated road infrastructure, including an intensively used expressway and an interchange during construction [100], and waters of the Lake Wicko Małe, i. e. an extension of Szczecin Lagoon. In consequence, there is no continuity between the forests of the upland part of the island and its lowland counterparts, located on a peninsula with a narrow base, occupied by the city and the roads leading to it, while the only natural habitat connecting both parts of the Park is a narrow stripe of reed beds. Urbanized areas and roads do not represent impermeable barriers for either A. agrarius or A. flavicollis, which both are known to successfully colonize isolated green areas in large cities [101,102]. They may, however, act as a selective filter for mammal species [103], even during population outbreaks, like those following mast years for beech and oak, when dominant woodland rodents may increase their presence in the neighboring suboptimal habitats [88], especially open areas and wetlands.
5. Conclusions
Our study revealed a unique quantitative structure of a small mammal assemblage in a Central European national park, characterized by an exceptionally low species richness and diversity, with some species characteristic for some habitats abundant in the park, notably wetland ones, either extremely scarce or completely absent. Historical data provide evidence for the occurrence of the recently absent species on the coastal marine island, at which the park is located. We suppose that the described species’ paucity, compared to the other national parks of the region, can be explained within the framework of island biogeography, mainly the increased chance of local extinctions and, consequently, species turnover, during minimum phases of their population cycles or fluctuations. The effect appeared to be strong enough even though the water barrier separating the island from the mainland is, at its narrowest point, no wider than some larger rivers in the region. The location of the park at the edge of the regional distribution of three further small mammal species may contribute to the depauperation of local assemblages as well. The hypothesis will, however, remain speculative until the equally extensive sampling in the part of the island outside the park border is conducted. Another distinct feature of small mammal assemblages revealed during our survey, is the effect of broad-scale topography, affecting the clustering more than local vegetation, usually considered a crucial factor shaping the species composition of epigeic rodents and shrews. We link that to the further, intra-insular barriers, acting as dispersal filters, at least partially associated with anthropogenic effects, namely urbanization and development of road infrastructure. Our study confirmed that even the young, land-bridge islands, developed after the last glaciation and located very close to the shore, may develop faunal specificity unknown from similar mainland areas. In the case of Wolin Island, the situation of small terrestrial mammals superficially resembles that of bats, however, we assume a completely different set of factors responsible for that.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Species composition of small mammals trapped at particular study sites, presented as proportion of captures of each species, used for cluster analysis (Figure 2); individuals identified only to genus or genus-group were excluded; Table S2: Species composition of small mammals in the five national parks of the northwestern Poland and northeastern Germany, presented as number of captures of each species, used to plot rarefaction curves (Figure 4).
Author Contributions
Conceptualization: M.C.; methodology, M.C.; small mammal trapping, M.C., Z.W., T.K., M.W., J.B., B.S., K.B., A.R.; preparation of databases, M.C.; statistical analysis, M.C.; writing—original draft preparation, M.C.; writing—review and editing, M.C. and Z.W.; data visualization, M.C.; graphic abstract, map and spatial data, Z.W.; preparation of references, Z.W., T.K.; project administration, M.C. and Z.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Forest Fund of State Forests National Forest Holding (Fundusz Leśny Państwowego Gospodarstwa Leśnego Lasy Państwowe), Grant No. MZ.0290.1.22.2023, through Contract No. 97/2023 between Wolin National Park (the recipient of the grant) and the Polish Society for Nature Protection ‘Salamandra’.
Institutional Review Board Statement
Study was conducted based on the license by Ministry of Climate and Environment and the permission to conduct research issued by the director of the Wolin National Park.
Data Availability Statement
All data are provided in the supplementary material.
Acknowledgments
We would like to thank the staff of Wolin National Park for their permission to conduct the study, organization of funding and accommodation, and help with logistics: Wioletta Nawrocka, Alicja Łepek, Konrad Wrzecionkowski, Marek Szwarc, Tomasz Bajor and Krzysztof Liszka.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| WNP | Wolin National Park |
| DNP | Drawieński National Park |
| LOVNP | Lower Oder Valley National Park |
| SNP | Słowiński National Park |
| WMNP | Warta Mouth National Park |
| NP | National Park |
References
- Hayward, G.F.; Phillipson, J. Community structure and functional role of small mammals in ecosystems. Ecol Small Mammals 1979, 135–211. [Google Scholar]
- Gardezi, T.; da Silva, J. Diversity in relation to body size in mammals: a comparative study. Am. Nat. 1999, 153, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Burgin, C.J.; Colella, J.P.; Kahn, P.L.; Upham, N.S. How many species of mammals are there? J Mammal 2018, 99, 1–14. [Google Scholar] [CrossRef]
- Stephens, R.B.; Anderson, E.M. Habitat associations and assemblages of small mammals in natural plant communities of Wisconsin. J Mammal 2014, 95, 404–420. [Google Scholar] [CrossRef]
- Aulak, W. Small mammal communities of the Białowieża National Park. Acta Theriol. 1970, 15, 465–515. [Google Scholar] [CrossRef]
- Niedziałkowska, M. , Kończak; J., Czarnomska, S.; Jędrzejewska, B. Species diversity and abundance of small mammals in relation to forest productivity in northeast Poland. Ecoscience 2010, 17, 109–119. [Google Scholar] [CrossRef]
- Kamenišťák, J.; Baláž, I.; Tulis, F.; Jakab, I.; Ševčík, M.; Poláčiková, Z.; Klimant, P.; Ambros, M.; Rychlik, L. Changes of small mammal communities with the altitude gradient. Biologia 2020, 75, 713–722. [Google Scholar] [CrossRef]
- Chirichella, R.; Ricci, E.; Armanini, M.; Gobbi, M.; Mustoni, A.; Apollonio, M. Small mammals in a mountain ecosystem: the effect of topographic, micrometeorological, and biological correlates on their community structure. Community Ecol 2022, 23, 289–299. [Google Scholar] [CrossRef]
- Tomassini, O.; Aghemo, A.; Baldeschi, B.; Bedini, G.; Petroni, G.; Giunchi, D.; Massolo, A. Some like it burnt: species differences in small mammal assemblage in a Mediterranean basin nearly 3 years after a major fire. Mamm Res 2024, 69, 283–302. [Google Scholar] [CrossRef]
- Torre, I.; Ribas, A.; Puig-Gironès, R. Effects of post-fire management on a Mediterranean small mammal community. Fire 2023, 6, 34. [Google Scholar] [CrossRef]
- Jasiulionis, M.; Stirkė, V.; Balčiauskas, L. The distribution and activity of the invasive raccoon dog in Lithuania as found with country-wide camera trapping. Forests 2023, 14, 1328. [Google Scholar] [CrossRef]
- Wikar, Z.; Ciechanowski, M.; Zwolicki, A. The positive response of small terrestrial and semi-aquatic mammals to beaver damming. Sci Total Environ 2024, 906, 167568. [Google Scholar] [CrossRef] [PubMed]
- Gentili, S.; Sigura, M.; Bonesi, L. Decreased small mammals species diversity and increased population abundance along a gradient of agricultural intensification. Hystrix 2014, 25, 39–44. [Google Scholar] [CrossRef]
- Wołk, E.; Wołk, K. Responses of small mammals to the forest management in the Białowieża Primeval Forest. Acta Theriol 1982, 27, 45–59. [Google Scholar] [CrossRef]
- Pearce, J.; Venier, L. Small mammals as bioindicators of sustainable boreal forest management. Forest ecology and management 2005, 208, 153–175. [Google Scholar] [CrossRef]
- Ważna, A.; Cichocki, J.; Bojarski, J.; Gabryś, G. Impact of sheep grazing on small mammals diversity in lower mountain coniferous forest glades. Appl Ecol Environ Res 2016, 14, 115–127. [Google Scholar] [CrossRef]
- Adler, G.H.; Wilson, M.L. Small mammals on Massachusetts islands: the use of probability functions in clarifying biogeographic relationships. Oecologia 1985, 66, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Haq SM, A. Multi-benefits of national parks and protected areas: an integrative approach for developing countries. Environ Socio-econ S 2016, 4, 1–11. [Google Scholar] [CrossRef]
- Jamroży, G. Ssaki polskich parków narodowych: drapieżne, kopytne, zajęczaki, duże gryzonie. Instytut Bioróżnorodności Leśnej; Wydział Leśny Uniwersytetu Rolniczego, Uniwersytet Rolniczy im. Hugona Kołłątaja & Vega Studio Adv. Tomasz Müller: Kraków, Poland, 2014. [Google Scholar]
- Ciechanowski, M.; Wikar, Z.; Borzym, K.; Janikowska, E.; Brachman, J.; Jankowska-Jarek, M.; Bidziński, K. Exceptionally Uniform Bat Assemblages across Different Forest Habitats Are Dominated by Single Hyperabundant Generalist Species. Forests 2024, 15, 337. [Google Scholar] [CrossRef]
- Pucek, Z. (1984). Klucz do oznaczania ssaków Polski [Key for identification of Polish mammals]. Państwowe Wydawnictwo Naukowe, Warszawa.
- Michaux, J. R.; Chevret, P.; Filippucci, M. G.; Macholan, M. Phylogeny of the genus Apodemus with a special emphasis on the subgenus Sylvaemus using the nuclear IRBP gene and two mitochondrial markers: cytochrome b and 12S rRNA. Mol Phyl Evol 2002, 23, 123–136. [Google Scholar] [CrossRef]
- Piłacińska, B.; Ziomek, J.; Bajaczyk, R. Drobne ssaki Drawieńskiego Parku Narodowego [Small mammals of the Drawieński National Park]. Bad Fizjogr Pol Zach 1999, 46, 46–95. [Google Scholar]
- Wojtaszyn, G.; Rutkowski, T.; Lesiński, G.; Stephan, W.; Salamandra, P. T. O. P. Soricomorphs and rodents of the Ujście Warty National Park and the surrounding area. Chrońmy Przyr Ojczystą 2015, 71, 179–191. [Google Scholar]
- Rychlik, L. S.; Eichert U., M.; Kowalski, K. Diversity of small mammal assemblages in natural forests and other habitats of the Słowiński National Park, northern Poland - preliminary results. In Nationalpark-Jahrbuch Unteres Odertal; Vössing, A., Ed.; 2020; Volume III, pp. 66–71. [Google Scholar]
- Decher, J.; Bakarr, I.; Hoffmann, A.; Jentke, T.; Klappert, A.; Kowalski, G.; Kuzdrowska, K.; Malinowska, B.; Rychlik, L. S. Aktualisierung unserer Kenntnisse über die Kleinsäugergemeinschaften im Nationalpark Unteres Odertal. In A. Vössing (ed.), Nationalpark-Jahrbuch Unteres Odertal 2021, 18, 145–150. [Google Scholar]
- Hoffmann, A.; Jankowiak, Ł.; Modelska, Z.; Piórkowska, K.; Decher, J.; Jentke, T.; Klappert, A.; Kuzdrowska, K.; Malinowska, B.; Sęk, O. W.; Rychlik, L. S. Diversität von Kleinsäugern im nördlichen Teil des Nationalparks Unteres Odertal. In Vössing, A. (ed.), Nationalpark-Jarbuch Unteres Odertal, 2022, 19, 37–45. [Google Scholar]
- Herold, W. Zur Kleinsäugerfauna der Insel Usedom und Wolin. Dohrniana, Stettin 1934, 13, 176–196. [Google Scholar]
- Pucek, Z.; Raczyński, J. (Eds.) Atlas rozmieszczenia ssaków w Polsce; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1983. [Google Scholar]
- Herold, W. Beiträge zur Säugetierfauna Usedom-Wollins. I Abch Ber Pommerschen Nat-forsch Ges Stettin, 1921, 2, 75–79. [Google Scholar]
- Open Forest Data. Available online: https://dataverse.openforestdata.pl/dataverse/zoo (accessed on 05.05.2024).
- Ciechanowski, M.; Wikar, Z.; Kowalewska, T.; Wojtkiewicz, M.; Brachman, J.; Sarnowski, B.; Borzym, K.; Rydzyńska, A. Department of Vertebrate Ecology and Zoology. University of Gdańsk, Gdańsk, Poland, 2023. Unpublished data.
- Gaffrey, G. Die rezenten wildlebenden Säugetiere Pommerns. Doctoral dissertation, Universität Greifswald, Greifswald, 1944. [Google Scholar]
- Skuratowicz, W. Materiały do fauny pcheł (Aphaniptera) Polski. Acta Parasitol Pol 1954, 2, 65–96. [Google Scholar]
- Herold, W. Zur Verbreitung der Schlagmäuse in Pommern. I Abch Ber Pommerschen Nat-forsch Ges Stettin 1922, 3, 43–50. [Google Scholar]
- Herold, W. Zum Vorkommen von Glis glis (L.). Dohrniana, Stettin 1939, 18. [Google Scholar]
- Goc, M. Stanowisko popielicy szarej Glis glis w Słowińskim Parku Narodowym. Przegląd Przyr 2019, 30. [Google Scholar]
- Rathke, D.; Bröring, U. Colonization of post-mining landscapes by shrews and rodents (Mammalia: Rodentia, Soricomorpha). Ecol Eng 2005, 24, 149–156. [Google Scholar] [CrossRef]
- Nicolas, V.; Colyn, M. Relative efficiency of three types of small mammal traps in an African rainforest. Belg J Zool 2006, 136, 107. [Google Scholar]
- Pucek, Z. Trap response and estimation of numbers of shrews in removal catches. Acta theriol 1969, 14, 403–426. [Google Scholar] [CrossRef]
- O'Brien, C.; McShea, W. J.; Guimondou, S.; Barriere, P.; Carleton, M. D. Terrestrial small mammals (Soricidae and Muridae) from the Gamba Complex in Gabon: species composition and comparison of sampling tecnhiques. Bull Biol Soc Wash 2006, 12, 353–363. [Google Scholar]
- Bovendorp, R.S.; Mccleery, R.A.; Galetti, M. Optimising sampling methods for small mammal communities in Neotropical rainforests. Mamm Rev 2017, 47, 148–158. [Google Scholar] [CrossRef]
- Pankakoski, E. The cone trap—a useful tool for index trapping of small mammals. Ann Zool Fennici 1979, 16, 144–150. [Google Scholar]
- Pelikan, J. , Zejda, J.; Holisova, V. Efficiency of different traps in catching small mammals. Folia Zool 1977, 26, 1–13. [Google Scholar]
- Darinot, F. Dispersal and genetic structure in a harvest mouse (Micromys minutus Pallas, 1771) population, subject to seasonal flooding; TEL - Thèses en ligne: France, 2019. [Google Scholar]
- Occhiuto, F.; Mohallal, E.; Gilfillan, G. D.; Lowe, A.; Reader, T. Seasonal patterns in habitat use by the harvest mouse (Micromys minutus) and other small mammals. Mammalia 2021, 85, 325–335. [Google Scholar] [CrossRef]
- Ziomek, J. Drobne ssaki (Micromammalia) Roztocza. Część I. Micromammalia wybranych biotopów Roztocza Środkowego. Fragm faunist 1998, 41, 93–123. [Google Scholar]
- Juskaitis, R. Peculiarities of habitats of the common dormouse, Muscardinus avellanarius, within its distributional range and in Lithuania: a review. Folia Zool Praha 2007, 56, 337. [Google Scholar]
- Ciechanowski, M.; Cichocki, J.; Ważna, A.; Piłacińska, B. Small-mammal assemblages inhabiting Sphagnum peat bogs in various regions of Poland. Biol Lett 2012, 49, 115–133. [Google Scholar] [CrossRef]
- Mitchell-Jones, A. J.; Mitchell, J.; Amori, G.; Bogdanowicz, W.; Spitzenberger, F.; Krystufek, B.; Reijnders, P J. H.; Spitzenberger, E.; Stubbe, M.; Thissen, J. B. M.; Vohralik, V.; Zima, J. The atlas of European mammals; T & AD Poyser: London; vol. 3. [CrossRef]
- Igea, J.; Aymerich, P.; Bannikova, A. A.; Gosálbez, J.; Castresana, J. Multilocus species trees and species delimitation in a temporal context: application to the water shrews of the genus Neomys. BMC evol biol 2015, 15, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cichocki, J.; Kościelska, A.; Piłacińska, B.; Kowalski, M.; Ważna, A.; Dobosz, R.; Nowakowski, K.; Lesiński, G.; Gabrys, G. Occurrence of lesser white-toothed shrew Crocidura suaveolens (Pallas, 1811) in Poland. Zeszyty Naukowe. Acta Biol. Uniwersytet Szczeciński 2014, 21. [Google Scholar]
- Haberl, W.; Kryštufek, B. Spatial distribution and population density of the harvest mouse Micromys minutus in a habitat mosaic at Lake Neusiedl, Austria. Mammalia 2003, 67, 355–366. [Google Scholar] [CrossRef]
- Surmacki, A.; Gołdyn, B.; Tryjanowski, P. Location and habitat characteristics of the breeding nests of the harvest mouse (Micromys minutus) in the reed-beds of an intensively used farmland. Mammalia 2005, 69, 5–9. [Google Scholar] [CrossRef]
- Brzeziński, M.; Jedlikowski, J.; Komar, E. Space use, habitat selection and daily activity of water voles Arvicola amphibius co-occurring with the invasive American mink Neovison vison. Folia Zool 2019, 68, 21–28. [Google Scholar] [CrossRef]
- van der Putten, T. A.; Verhees, J. J.; Koma, Z.; van Hoof, P. H.; Heijkers, D.; de Boer, W. F.; Esser, H. J.; Hoogerwerf, G.; Lemmers, P. Insights into the fine-scale habitat use of Eurasian Water Shrew (Neomys fodiens) using radio tracking and LiDAR. J Mammal 2025, gyae146. [Google Scholar] [CrossRef]
- Raczyński, J.; Fedyk, S.; Gębczyńska, Z.; Pucek, M. Drobne ssaki środkowego i dolnego basenu Biebrzy. Zeszyty problemowe postępów nauk rolniczych 1983, 255. [Google Scholar]
- Rychlik, L. Habitat preferences of four sympatric species of shrews. Acta Theriol 2000, 45 (Suppl. 1), 173–190. [Google Scholar] [CrossRef]
- Łopucki, R.; Mróz, I.; Klich, D.; Kitowski, I. Small mammals of xerothermic grasslands of south-eastern Poland. Annals of Warsaw University of Life Sciences-SGGW. Anim Sci 2018, 57, 257–267. [Google Scholar] [CrossRef]
- Tattersall, F. H.; Macdonald, D. W.; Hart, B. J.; Manley, W. J.; Feber, R. E. Habitat use by wood mice (Apodemus sylvaticus) in a changeable arable landscape. J Zool 2001, 255, 487–494. [Google Scholar] [CrossRef]
- Gębczyńska, Z.; Raczyński, J. Fauna i ekologia drobnych ssaków Narwiańskiego Parku Narodowego. Parki Narodowe i Rezerwaty Przyrody 1997, 16, 37–61. [Google Scholar]
- Suchomel, J.; Purchart, L.; Čepelka, L. Structure and diversity of small-mammal communities of lowland forests in the rural central European landscape. Eur J For Res 2012, 131, 1933–1941. [Google Scholar] [CrossRef]
- Loeb, S.C. Responses of small mammals to coarse woody debris in a southeastern pine forest. J Mammal 1999, 80, 460–471. [Google Scholar] [CrossRef]
- Sullivan, T. P.; Sullivan, D. S. Maintenance of small mammals using post-harvest woody debris structures on clearcuts: linear configuration of piles is comparable to windrows. Mamm Res 2018, 63, 11–19. [Google Scholar] [CrossRef]
- Jędrzejewski, W.; Jędrzejewska, B. Rodent cycles in relation to biomass and productivity of ground vegetation and predation in the Palearctic. Acta Theriol 1996, 41, 1–34. [Google Scholar] [CrossRef]
- Andreassen, H. P.; Sundell, J.; Ecke, F.; Halle, S.; Haapakoski, M.; Henttonen, H.; Huitu, O.; Jacob, J.; Johnsen, K.; Koskela, E.; Luque-Larena, J.; Lecomte, N.; Leirs, H. , Mariën, J.; Neby, M.; Rätti, O.; Sievert, T.; Singleton, G. R.; van Cann, J.; Vanden Broecke B.; Ylönen, H. Population cycles and outbreaks of small rodents: ten essential questions we still need to solve. Oecologia 2021, 195, 601–622. [Google Scholar] [CrossRef] [PubMed]
- Trout, R.C. A review of studies on populations of wild harvest mice (Micromys minutus (Pallas). Mamm Rev 1978, 8, 143–158. [Google Scholar] [CrossRef]
- Balčiauskas, L. , & Balčiauskienė, L. Long-Term Stability of Harvest Mouse Population. Diversity 2023, 15, 1102. [Google Scholar] [CrossRef]
- Ciechanowski, M.; Fałtynowicz, W.; Zieliński, S. (eds). The nature of the planned reserve “Dolina Mirachowskiej Strugi” in the Kaszubskie Lakeland (northern Poland). Acta Bot Cassubica 2004, 4, 5–137. [Google Scholar]
- Dokulilová, M.; Krojerová-Prokešová, J.; Heroldová, M.; Čepelka, L.; Suchomel, J. Population dynamics of the common shrew (Sorex araneus) in Central European forest clearings. Eur J Wildl Res 2023, 69, 54. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L. Habitat and Body Condition of Small Mammals in a Country at Mid-Latitude. Land 2024, 13, 1214. [Google Scholar] [CrossRef]
- Pucek, Z.; Jędrzejewski, W.; Jędrzejewska, B.; Pucek, M. Rodent population dynamics in a primeval deciduous forest (Białowieża National Park) in relation to weather, seed crop, and predation. Acta Theriol 1993, 38, 199–232. [Google Scholar] [CrossRef]
- Panov, V. V.; Karpenko, S. V. The population dynamics of the water shrew Neomys fodiens (Mammalia, Soricidae) and its helminthes fauna in the northern Baraba. Parazitologiia 2004, 38, 448–456. [Google Scholar]
- Strzelczyk, J.; Łabuz, T. Danielewska, A. & Maciag, M., Ed.; Zmiany linii brzegowej oraz powierzchni wyspy Wolin w holocenie i ich wpływ na osadnictwo od mezolitu do czasów współczesnych (Coastline and the surface of the Wolin Island changes in the Holocene and their impact on settlement from the Mesolithic to modern times). In Najnowsze doniesienia z zakresu ochrony środowiska i nauk pokrewnych; Lublin, Poland, 2020. [Google Scholar]
- Bohdal, T.; Navrátil, J.; Sedláček, F. Small terrestrial mammals living along streams acting as natural landscape barriers. Ekológia (Bratislava) 2016, 35, 191–204. [Google Scholar] [CrossRef]
- Schenk, F. Comparison of spatial learning in woodmice (Apodemus sylvaticus) and hooded rats (Rattus norvegicus). J Comp Psychol 1987, 101, 150. [Google Scholar] [CrossRef]
- Stawski, C.; Koteja, P.; Sadowska, E. T.; Jefimow, M.; Wojciechowski, M. S. Selection for high activity-related aerobic metabolism does not alter the capacity of non-shivering thermogenesis in bank voles. J Comp Physiol A 2015, 180, 51–56. [Google Scholar] [CrossRef]
- Lomolino, M. V.; Brown, J. H.; Sax, D. F. Island biogeography theory. The theory of island biogeography revisited 2010, 13. [Google Scholar] [CrossRef]
- Lomolino, M.V. Mammalian island biogeography: effects of area, isolation and vagility. Oecologia 1984, 61, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Barreto, E.; Rangel, T. F.; Pellissier, L.; Graham, C. H. Area, isolation and climate explain the diversity of mammals on islands worldwide. Proc R Soc B 2021, 288, 20211879. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bao, Y.; Yu, M.; Xu, G.; Ding, P. BIODIVERSITY RESEARCH: Nestedness for different reasons: the distributions of birds, lizards and small mammals on islands of an inundated lake. Div Distrib 2010, 16, 862–873. [Google Scholar] [CrossRef]
- Mallinger, E. C.; Khadka, B.; Farmer, M. J.; Morrison, M.; Van Stappen, J.; Van Deelen, T. R.; Olson, E. R. Longitudinal trends of the small mammal community of the Apostle Islands archipelago. Comm Ecol 2021, 22, 55–67. [Google Scholar] [CrossRef]
- Pichler, T. R.; Mallinger, E. C.; Farmer, M. J.; Morrison, M. J.; Khadka, B.; Matzinger, P. J.; Kirschbaum, A.; Goodwin, K. R.; Route, W. T.; Van Stappen, J.; Timothy R. Van, Deelen; Olson, E. R. Comparative biogeography of volant and nonvolant mammals in a temperate island archipelago. Ecosphere 2022, 13, e3911. [Google Scholar] [CrossRef]
- Peltonen, A.; Hanski, I. Patterns of island occupancy explained by colonization and extinction rates in shrews. Ecology 1991, 72, 1698–1708. [Google Scholar] [CrossRef]
- Christiansen, T.S. Island Biogeography of small mammals in Denmark: Effects of area, isolation and habitat diversity. MSc thesis, University of Aarhus, 2005. [Google Scholar]
- Girjatowicz, J. P.; Świątek, M. Relationship between air temperature change and southern Baltic coastal lagoons ice conditions. Atmosphere 2021, 12, 931. [Google Scholar] [CrossRef]
- Suchomel, J.; Purchart, L.; Čepelka, L.; Heroldová, M. Structure and diversity of small mammal communities of mountain forests in Western Carpathians. Eur J For Res 2014, 133, 481–490. [Google Scholar] [CrossRef]
- Zwolak, R.; Witczuk, J.; Bogdziewicz, M.; Rychlik, L.; Pagacz, S. Simultaneous population fluctuations of rodents in montane forests and alpine meadows suggest indirect effects of tree masting. J Mammal 2018, 99, 586–595. [Google Scholar] [CrossRef]
- Romanowski, J.; Dudek-Godeau, D.; Lesiński, G. The Diversity of Small Mammals along a Large River Valley Revealed from Pellets of Tawny Owl Strix aluco. Biology 2023, 12, 1118. [Google Scholar] [CrossRef]
- Cornulier, T.; Yoccoz, N. G.; Bretagnolle, V.; Brommer, J. E.; Butet, A.; Ecke, F.; Framstad, E.; Henttonen, H.; Hörnfeldt, B.; Huitu, O.; Imholt, C.; Ims, R. A.; Jacob, J.; Jędrzejewska, B.; Millon, A.; Petty, S.; Pietiäinen, H.; Tkadlec, H.; Zub, K.; Lambin, X. Europe-wide dampening of population cycles in keystone herbivores. Science 2013, 340, 63–66. [Google Scholar] [CrossRef]
- Haitlinger, R. Arthropod communities. Acta Theriol 1983, 28 (Suppl. 1), 55–88. [Google Scholar] [CrossRef]
- Grodziński, W. The succession of small mammal communities on an overgrown clearing and landslip in the Western Carpathians. Bull Acad Pol Sc, Cl. II 1958, 6, 10. [Google Scholar]
- Bogdziewicz, M.; Zwolak, R. Responses of small mammals to clear-cutting in temperate and boreal forests of Europe: a meta-analysis and review. Eur J For Res 2014, 133, 1–11. [Google Scholar] [CrossRef]
- Mažeikytė, R. Small mammals in the mosaic landscape of eastern Lithuania: species composition, distribution and abundance. Acta Zool Litu 2002, 12, 381–391. [Google Scholar] [CrossRef]
- Šinkūnas, R.; Balčiauskas, L. Small mammal communities in the fragmented landscape in Lithuania. Acta Zool Litu 2006, 16, 130–136. [Google Scholar] [CrossRef]
- Greenberg, R.; Maldonado, J. E.; Droege, S. A. M.; McDonald, M. V. Tidal marshes: a global perspective on the evolution and conservation of their terrestrial vertebrates. BioScience 2006, 56, 675–685. [Google Scholar] [CrossRef]
- Verkuil, Y. I.; van Guldener, W. E.; Lagendijk, D. G.; Smit, C. Molecular identification of temperate Cricetidae and Muridae rodent species using fecal samples collected in a natural habitat. Mamm Res 2018, 63, 379–385. [Google Scholar] [CrossRef]
- Christensen, J.T.; Jensen, T.S. Småpattedyrfaunaen på Anholt og Sprogø. Flora og Fauna 2024, 129, 3–7. [Google Scholar]
- Ambros, M. Drobné cicavce (Mammalia: Soricomorpha, Rodentia) území európskeho významu: slaniská a slané lúky. Naturae Tutela 2018, 22, 203–214. [Google Scholar]
- Rico, A.; Kindlmann, P.; Sedlacek, F. Barrier effects of roads on movements of small mammals. FOLIA ZOOL-PRAHA 2007, 56, 1. [Google Scholar]
- Andrzejewski, R.; Babińska-Werka, J.; Gliwicz, J.; Goszczyński, J. Synurbization processes in population of Apodemus agrarius. I. Characteristics of populations in an urbanization gradient. Acta Theriol 1978, 23, 341–358. [Google Scholar] [CrossRef]
- Lesiński, G.; Gryz, J.; Krauze-Gryz, D.; Stolarz, P. Population increase and synurbization of the yellow-necked mouse Apodemus flavicollis in some wooded areas of Warsaw agglomeration, Poland, in the years 1983–2018. Urban Ecosyst 2021, 24, 481–489. [Google Scholar] [CrossRef]
- Tangil, B. D.; Rodríguez, A. Environmental filtering drives the assembly of mammal communities in a heterogeneous Mediterranean region. Ecol Appl 2023, 33, e2801. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Location of study sites at which small mammal trapping was conducted within the borders of Wolin National Park. The number of each site corresponds to the Table 1. mh - moraine hills (uplands), ad - alluvial delta (lowlands).
Figure 1.
Location of study sites at which small mammal trapping was conducted within the borders of Wolin National Park. The number of each site corresponds to the Table 1. mh - moraine hills (uplands), ad - alluvial delta (lowlands).
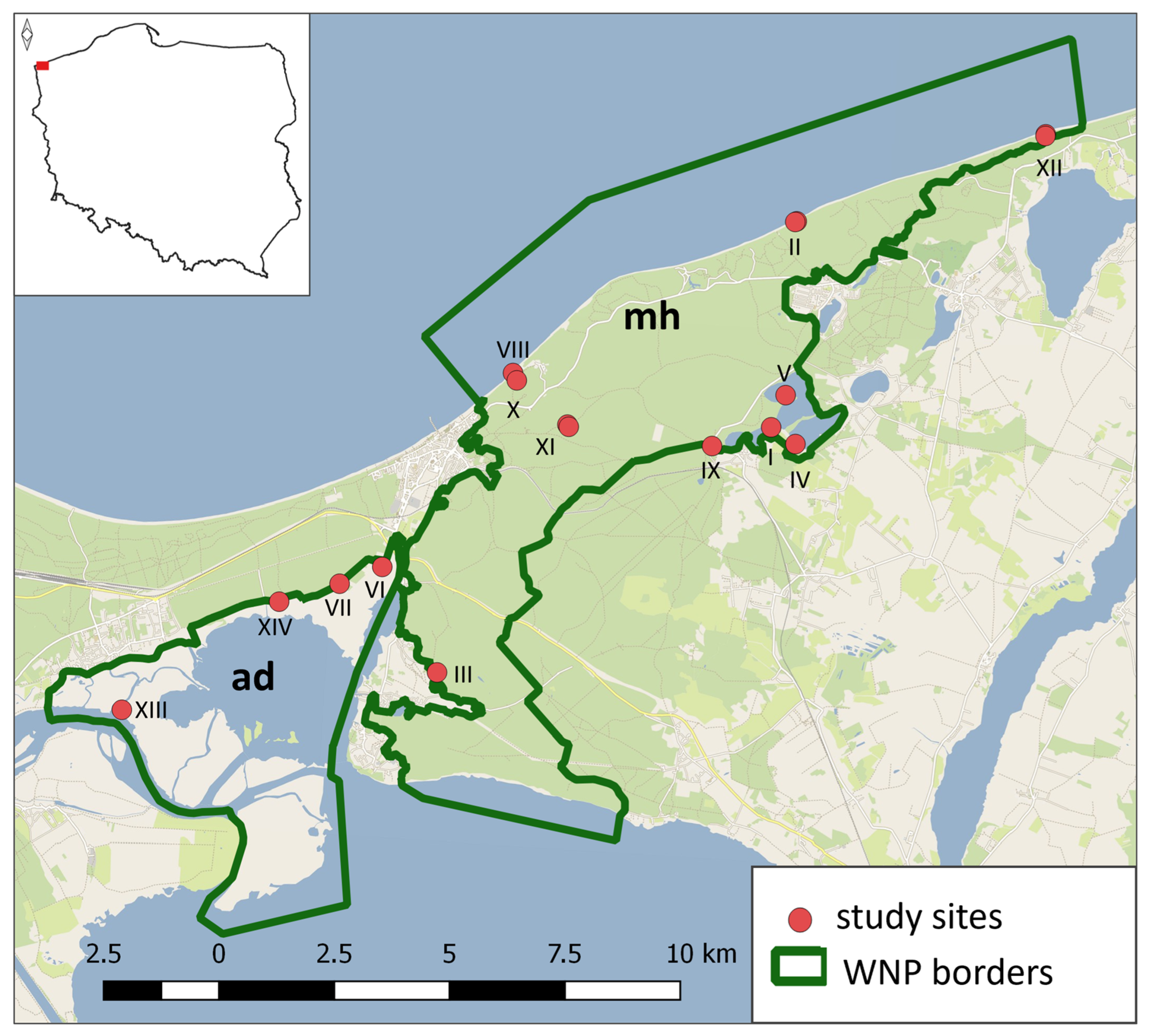
Figure 3.
Structure of small mammal assemblages hosted by four different groups of habitats, distinguished by plant communities and topography. Meso- and acidophilic forests on mineral soils: sites V, X, XI, XII. Upland wetland forests: sites I, IV. Dry non-forest habitats: sites II, III, VIII, IX. Wetland habitats on the alluvial delta: sites VI, VII, XIII, XIV.
Figure 3.
Structure of small mammal assemblages hosted by four different groups of habitats, distinguished by plant communities and topography. Meso- and acidophilic forests on mineral soils: sites V, X, XI, XII. Upland wetland forests: sites I, IV. Dry non-forest habitats: sites II, III, VIII, IX. Wetland habitats on the alluvial delta: sites VI, VII, XIII, XIV.
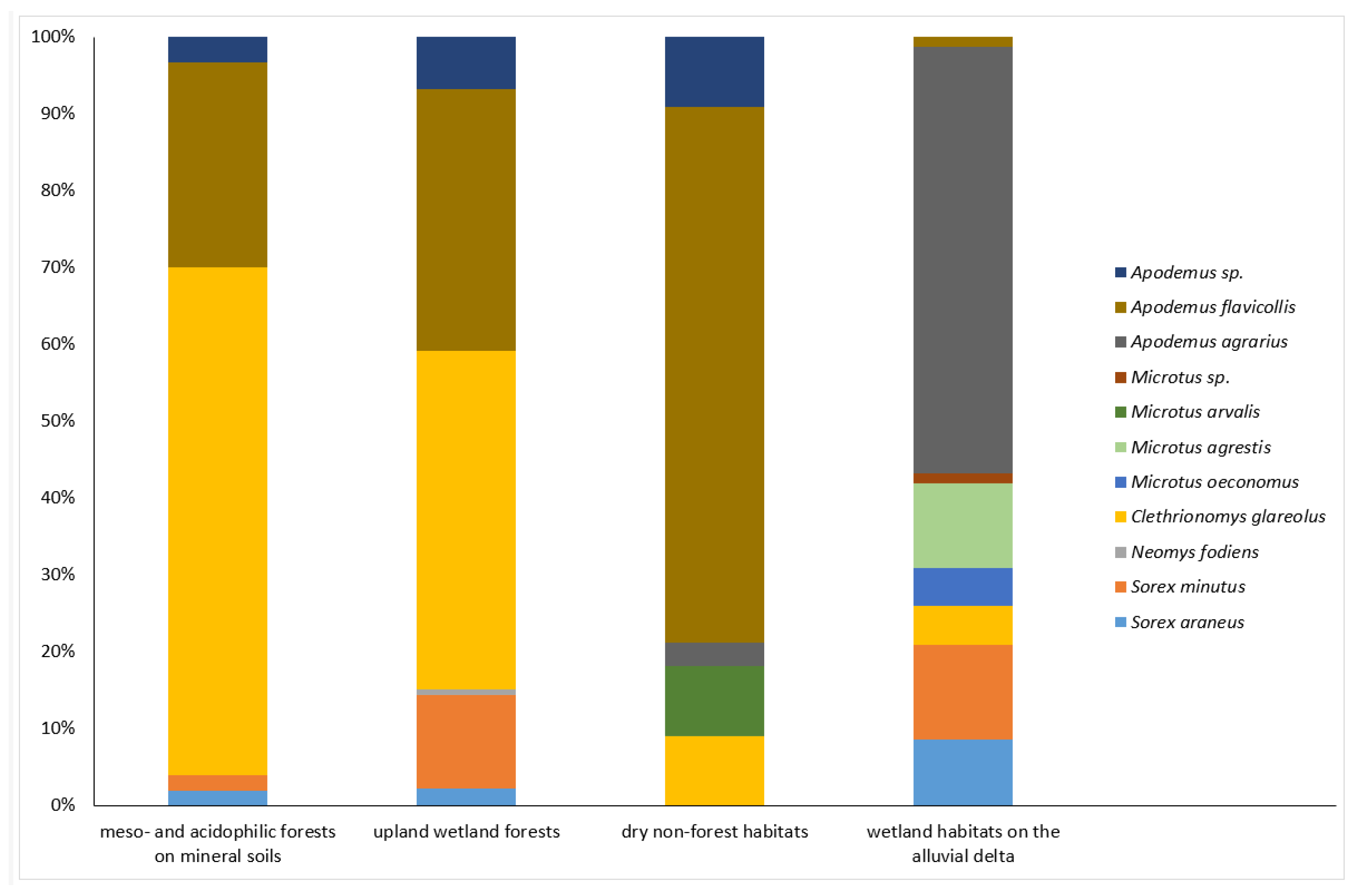
Figure 5.
Rarefaction curves for the cumulative numbers of species plotted against the numbers of small mammal captures in Wolin National Park (WNP) and four other national parks in the Polish Baltic Sea Coast and Lower Oder catchment. DNP - Drawieński National Park, LOVNP - Lower Oder Valley National Park, SNP - Słowiński National Park, WMNP - Warta Mouth National Park (data in Table S2).
Figure 5.
Rarefaction curves for the cumulative numbers of species plotted against the numbers of small mammal captures in Wolin National Park (WNP) and four other national parks in the Polish Baltic Sea Coast and Lower Oder catchment. DNP - Drawieński National Park, LOVNP - Lower Oder Valley National Park, SNP - Słowiński National Park, WMNP - Warta Mouth National Park (data in Table S2).
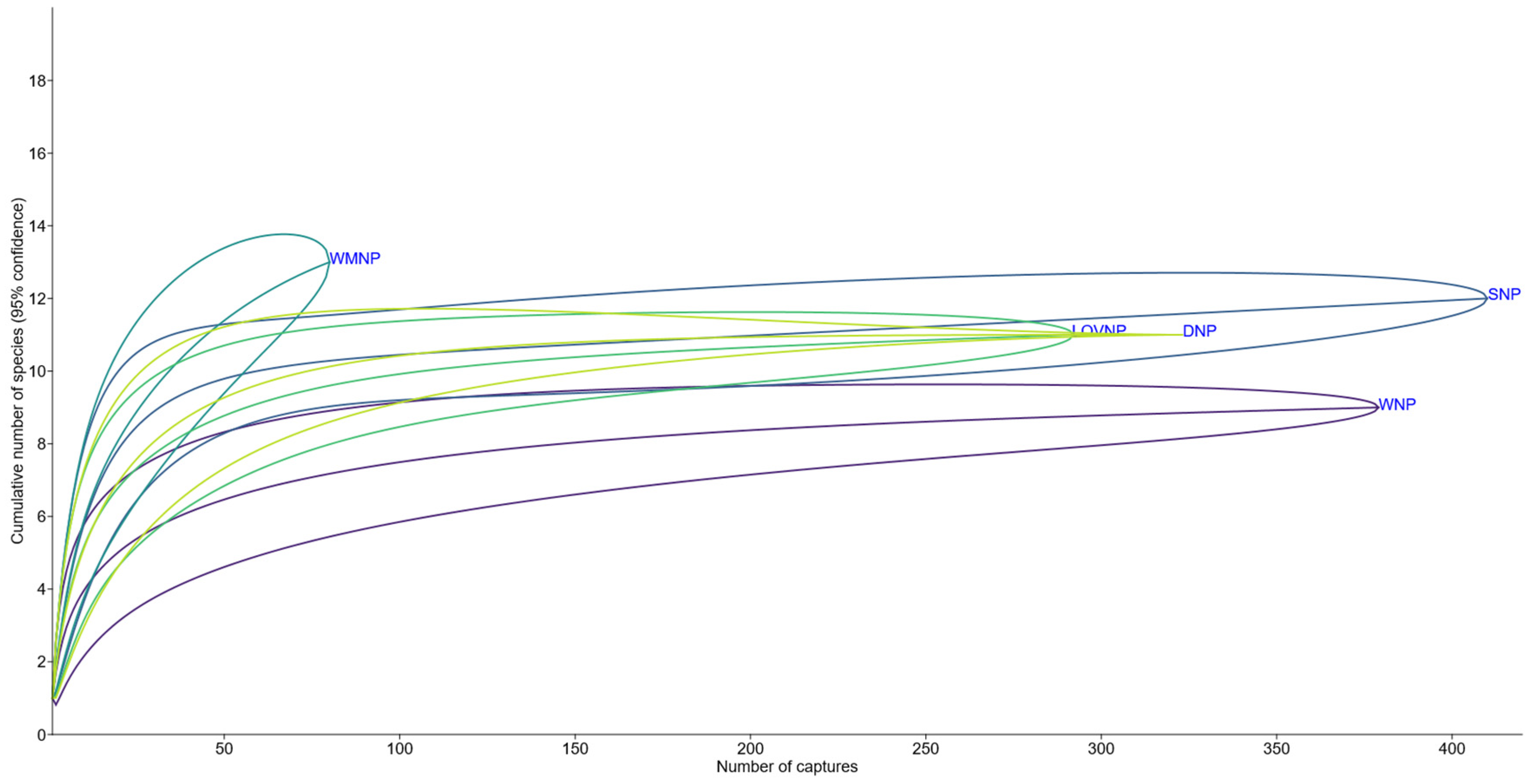
Table 2.
Results of small mammal trapping (number of captures of a particular taxon) in every sampled habitat of the Wolin National Park. Number of every site corresponds to the Table 1. Species: Sara - S. araneus, Smin - S. minutus, Nfod - N. fodiens, Cgla - C. glareolus, Aoec - Alexandromys oeconomus, Magr - M. agrestis, Marv - M. arvalis, A/M - Alexandromys/Microtus, Aagr - A. agrarius, Afla - A. flavicollis, Asp - Apodemus sp. (only representatives of subgenus Sylvaemus).
Table 2.
Results of small mammal trapping (number of captures of a particular taxon) in every sampled habitat of the Wolin National Park. Number of every site corresponds to the Table 1. Species: Sara - S. araneus, Smin - S. minutus, Nfod - N. fodiens, Cgla - C. glareolus, Aoec - Alexandromys oeconomus, Magr - M. agrestis, Marv - M. arvalis, A/M - Alexandromys/Microtus, Aagr - A. agrarius, Afla - A. flavicollis, Asp - Apodemus sp. (only representatives of subgenus Sylvaemus).
| Sara | Smin | Nfod | Cgla | Aoec | Magr | Marv | A/M | Aagr | Afla | Asp | |||
| I | 1 | 4 | 1 | 37 | - | - | - | - | - | 16 | 6 | 65 | 4 |
| Site/habitat | Species | total | S | ||||||||||
| IIa | - | - | - | - | - | - | 1 | - | - | 7 | 1 | 9 | 2 |
| IIb | - | - | - | - | - | - | - | - | - | 1 | - | 1 | 1 |
| III | - | - | - | - | - | - | 2 | - | - | 1 | 2 | 5 | 2 |
| IV | 2 | 12 | - | 21 | - | - | - | - | - | 29 | 3 | 67 | 4 |
| V | - | 2 | - | 6 | - | - | - | - | - | 16 | 2 | 26 | 3 |
| VI | - | - | - | 4 | 4 | 3 | - | 1 | 2 | 1 | - | 15 | 5 |
| VII | 1 | 1 | - | - | - | 4 | - | - | 5 | - | - | 11 | 4 |
| VIII | - | - | - | - | - | - | - | - | - | 12 | - | 12 | 1 |
| IX | - | - | - | 3 | - | - | - | - | 1 | 3 | - | 7 | 3 |
| X | 3 | - | - | 92 | - | - | - | - | - | 21 | 3 | 119 | 3 |
| XIa | - | - | - | - | - | - | - | - | - | 2 | - | 2 | 1 |
| XIb | - | - | - | - | - | - | - | - | - | 1 | - | 1 | 1 |
| XIIa | - | 1 | - | - | - | - | - | - | - | - | - | 1 | 1 |
| XIIb | - | - | - | 1 | - | - | - | - | - | - | - | 1 | 1 |
| XIII | - | - | - | - | - | - | - | - | 8 | - | - | 8 | 1 |
| XIV | 6 | 9 | - | - | - | 2 | - | - | 30 | - | - | 47 | 4 |
| total | 13 | 29 | 1 | 164 | 4 | 9 | 3 | 1 | 46 | 110 | 17 | 397 | 9 |
| % | 3,3 | 7,3 | 0,3 | 41,3 | 1,0 | 2,3 | 0,8 | 0,3 | 11,6 | 27,7 | 4,3 | 100,0 | |
| number of habitats | 5 | 6 | 1 | 7 | 1 | 3 | 2 | - | 5 | 12 | - | 17 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



