
Submitted:
27 May 2025
Posted:
28 May 2025
You are already at the latest version
Abstract
Fragments of urban forests can host a remarkable diversity of insects, even in environments that have been greatly transformed. This study evaluates the diversity, abundance and composition of insects that belong to seven families in four urban forest fragments near Panama City, i.e., Ciudad del Saber (CDS), Parque Natural Metropolitano (PNM), Corozal (COR) and Albrook (ALB). A total of 2,038 individuals were collected via Malaise traps and categorized into 403 morphospecies, 75 genera and 43 subfamilies. The highest richness of morphospecies was observed in CDS (223), whereas PNM exhibited the highest abundance of individuals (862). The alpha diversity indices (Shannon‒Wiener> 4.3; Margalef> 21; Pielou ≈ 1.0; and Simpson's inverse> 0.95) reflected communities that were characterized by high levels of diversity and equity. The level of similarity observed among the fragments was moderate (Diserud-Odegaard index = 0.543), thus indicating differences among the sites evaluated as part of this research in terms of their taxonomic composition. These results provide evidence concerning the variability of entomological communities in tropical urban landscapes and the role of forest fragments as possible reservoirs of biodiversity.
Keywords:
insects
; alpha and beta diversity
; morphospecies
; urban forests
; entomological communities
; Panama
1. Introduction
Panama City is a notable region in the neotropical region of America given that it has conserved considerable natural forest coverage both in its interior and in the surrounding areas [1]. However, these green areas are not exempt from the effects of urban growth, including the division of the landscape into fragments of isolated vegetation [2].
Urbanization is a transformative factor in landscapes that can alter ecological and functional patterns at various scales [3,4,5]. Through mechanisms such as the modification of microclimates, changes in the composition of the soil and the interruption of ecological flows, the urban environment can influence the distribution and composition of biological communities [6].
The reduction in extensive forest areas in isolated patches has been associated with variations in the richness, structure and abundance of species in different taxonomic groups [7,8,9]. However, the magnitude and direction of these changes depend largely on the ecological context and the group of organisms under investigation [7].
In recent decades, the need to include urban environments as valid settings for ecological research has been recognized, given the increasing expansion of such environments and their relevance with respect to efforts to conserve biodiversity [10,11,12]. Studies on urban diversity can facilitate not only the documentation of emerging biological patterns but also the evaluation of the roles of green areas as refuges or ecological corridors in landscapes that have been subjected to high levels of intervention [13,14,15].
This study aimed to estimate the diversity, abundance and taxonomic composition of insects that belong to the families Ichneumonidae, Bethylidae, Asilidae, Dolichopodidae, Curculionidae, Membracidae and Psocidae in four urban forest fragments that are located near Panama City. We sought to answer the following questions: (1) What are the richness, abundance and composition of insects among the fragments under evaluation? (2) What level of similarity is evident among the communities associated with the four fragments under investigation?
Given that the fragments investigated in this research are geographically close to one another, similar in terms of altitude, climate, and type of vegetation and located close to the Panama Canal, for many years, these fragments have maintained some degree of protection; accordingly, we hope to observe a high level of similarity among their entomological communities.
2. Materials and Methods
2.1. Study Area
The study was conducted in four urban forest fragments near Panama City: Metropolitan Natural Park (PNM, Los Momótides Trail), Albrook (ALB), Corozal (COR) and City of Knowledge (Cerro Gun) (Figure 1). These forest fragments are part of a biological corridor that runs along the eastern bank of the Panama Canal [16]. The forests of the Panama Canal Basin are located within protected areas and feature a rainy season that generally extends from May to November and a dry season that extends from December to April. These forests are characterized by different types of vegetation, but on the Pacific slope, which is close to Panama City, dry forest predominates [17]. The four sites under investigation are located within the tropical humid forest life zone [18].
In this study, we used the definition of fragments provided in [19], who defined a fragment as any patch of native vegetation around which most of the original vegetation has been removed. These fragments feature different types of soil and vegetation and vary in terms of size, shape and isolation. Accordingly, conserved places that are separated by a matrix of urban areas are viewed as fragments.
The Metropolitan Natural Park features a surface area of 232 ha and is divided into seven defined management zones: primitive use, priority archaeological interest, intensive use, extensive use, special use, coordination of the Curundú River and buffering influence [20]. The fragment under investigation in the PNM is located at 8° 59′41.55′′N and 79° 32′35.22′′W and features an approximate area of 18.12 ha alongside a perimeter of 1,756 km. The vegetation observed at this site is characterized by a mixture of humid tropical forest and lowland tropical dry forest, and it features few areas of stubble and grasslands alongside a well-defined stratum; this region is used for hiking and is delimited by Juan Pablo Avenue. II, Avenida de la Amistad and the Metropolitan Equestrian Club. The fragment in Corozal (COR) is located at 8° 59′19.34′′N and 79° 34′11.83′′W and features an approximate area of 56.31 ha and a perimeter of 3,028 km. The vegetation observed at this site can be characterized as herbaceous, including late secondary forest and some open areas. This forest fragment is managed by the Institutional Protection Service (SPI) and is used to support practice and training in the jungle. The fragment located in the town of Albrook (ALB) is located at 8° 58′37.49′′N and 79° 33′43.82′′W and features an approximate area of 34.79 ha and a perimeter of 5,003 km. The vegetation observed at this site is heterogeneous and consists of open grasslands, stubble and secondary forests. The site under investigation in this context is managed by the Panama Canal Authority; it is a heavily urbanized area in which many academic, commercial and recreational activities take place. The fragment located in the City of Knowledge (CDS) is known as the Dr. Rodrigo Tarte Biological Reserve. This area is protected by the Ciudad del Saber foundation. It is located at 9° 00 ”33.13′′N and 79° 35′12.96′′W. This site is adjacent to Soberanía National Park, and ex situ conservation activities are conducted at this location by the Ministry of the Environment (Table 1) (Figure 1).
These sites were selected from a qualitative perspective, which focused particularly on the degree of similarity among the biotic and physical characteristics of each site. The four sampling sites selected for this research are characterized by well-preserved secondary forests with similar types of vegetation; these locations have been fragmented by anthropogenic activities and pertain to the protection of the Panama Canal.
2.2. Sampling Method
Townes-type Malaise traps were used in this research as a passive trapping method considering their ability to collect fast-moving flying insects [21,22]. Two traps were installed in each fragment, which were separated from each other by 0.5 km and located at least 0.2 km from the edge of the fragment to minimize the edge effect. The traps were placed in open areas within the forest to intercept flight corridors. The sampling process took place weekly between August 2019 and March 2020, for a period of 33 weeks. The traps remained active continuously, thus generating a total effort of 5,760 hours of sampling and leading to the collection of 264 samples.
The samples thus collected were transported to the building of the Central American Program of Master in Entomology of the University of Panama (PCMENT). In one of the laboratories located in this building, a LEICA S9 stereoscope was used to separate individuals of the families of interest, which were subsequently stored in vials with 95% ethanol. These individuals were subsequently mounted on entomological pins (1, 2 and 3), separated into morphospecies and identified by reference to the relevant literature. The key provided in [23] was used to identify the subfamilies of Ichneumonidae Latreille, 1802. For Dolichopodidae Latreille, 1809; and Asilidae Meigen, 1802 used the genera keys in [24]. For the genera of Bethylidae Latreille, 1802, the key developed in [25,26] was used, and the illustrated key to the subfamilies and genera present in Panama provided in [27]. Was used for Psocoptera Schönherr, 1813 the keys developed in [28]. Were used to identify individuals in the family Curculionidae Linnaeus, 1758, was realized by Alfredo Lanuza, a specialist in Coleoptera. For the genera of Membracidae Latreille, 1802, the work in [29] and Flynn (2012) [30] was used as a reference for identification.
In this study, the morphospecies approach proposed in [31] was employed. This approach facilitates the use of morphological groupings as operational units in the absence of complete taxonomic identification. This method also facilitates the estimation of richness and the analysis of species turnover among sites, provided that each morphospecies is recognized consistently based on visible diagnostic characteristics. With this system, it is possible to construct comparable sets of species and perform biodiversity assessments in habitat fragments, even in the absence of precise information regarding the taxonomic identities of all the organisms involved in this process. Oliver and Beattie demonstrated the efficacy of this approach in the context of studies on ants, beetles and spiders, revealing that it is superior to traditional methods in terms of its ability to detect variations and predict ecological impacts in a more sensitive way.
2.3. Data Analysis
Statistical analyses were performed with the assistance of R version 4.2.2 software and the SpadeR platform. To determine the diversity and composition of the communities of the families under investigation, statistical analyses of alpha (α) diversity were conducted, including by reference to the Shannon‒Wiener index, which can facilitate the integration of both species’ richness and equitability in terms of the distribution of individuals and thus represents a composite measure of community heterogeneity [32]. In turn, Simpson's index can be used to estimate the probability that two randomly selected organisms belong to different species and is interpreted as a weighted average based on the relative abundance of the species present [33]. Similarly, the Pielou equity index was used to evaluate the degree of uniformity in the distribution of individuals among different species. To complement these analyses, accumulation and rarefaction curves were used, which can facilitate estimations of the expected wealth in communities that are homogeneous [34]. With respect to beta diversity, the Bray‒Curtis [35] and Jaccard [36] indices and the Diserud‒Odegaard multiple similarity index [37] were calculated. These metrics can facilitate the quantification of the turnover of species between communities or sites, thus reflecting the degree of biotic change along environmental or spatial gradients [38,39]. In addition, total species richness and true diversity were estimated via the SpadeR platform, and the abundance data pertaining to each site were used to compare the observed values with the expected values. On the other hand, Venn diagrams generated with the assistance of R (specifically, by reference to the matplotlib_venn package) were used to visualize the intersection of morphospecies among the different urban forest fragments, thus allowing the number of exclusive and shared species to be identified and providing a graphic representation of the levels of community overlap among the sites.
3. Results
3.1. Diversity and General Abundance
A total of 2,038 specimens that belonged to the seven families under investigation were collected alongside 43 subfamilies and 75 genera were collected; furthermore, a total of 403 morphospecies were estimated at the four sites under investigation. The Ichneumonidae family exhibited a total of 841 individuals, followed by Curculionidae at 446 individuals, Dolichopodidae at 426 individuals, Bethylidae at 277 individuals, Asilidae at 108 individuals, Psocidae Schönherr, 1817 at 90 individuals, and Membracidae at 64 individuals. The Ichneumonidae family exhibited the greatest number of subfamilies, at a total of 18. The subfamilies that exhibit the highest numbers were Crypthorynchinae Schoenherr, 1825 (Curculionidae), Epyrinae Kieffer, 1914 (Bethylidae) and Smiliinae Stal, 1869 (Membracidae), each of which was associated with six genera. These subfamilies were followed by Amphigerontiinae Pearman, 1936 (Psocidae), Baridinae Schoenherr, 1836, Curculioninae Latreille, 1802 (both from Curculionidae) and Membracinae Rafinesque, 1815 (Membracidae), which were associated with five genera each. On the other hand, the subfamilies Molytinae Schoenherr, 1823 and Conoderinae Schoenherr, 1833 (Curculionidae), alongside Pristocerinae Kieffer, 1914 (Bethylidae), were each associated with four genera.
In terms of abundance, the subfamily Sciapodinae Aldrich, 1905 (Dolichopodidae) was the most represented, at 389 samples, followed by Cryptinae Förster, 1869 (Ichneumonidae), at 243 individuals; Pristocerinae (Bethylidae) at 224; Conoderinae (Curculionidae) at 176; and Molytinae (Curculionidae) at 151 individuals. With respect to the number of morphospecies by genus, Dissomphalus Ashmead, 1893 (Bethylidae), was the most notable at 28 morphospecies, followed by Zygops Schoenherr, 1825 (Curculionidae), at 26, Conotrachelus Dejean, 1835 (Curculionidae), at 25, Lechriops Schoenherr, 1836 (Curculionidae), at 17, and Anthonomus Germar, 1817 (Curculionidae), at 7 morphospecies. In terms of abundance by genus, the most represented example was Condylostylus Bigot, 1859 (Dolichopodidae), at 277 specimens, followed by Zygops at 159, Dissomphalus at 151, Conotrachelus at 105, and Hercostomus Loew, 1857 (Dolichopodidae) at 104 individuals. Finally, the most abundant morphospecies identified during the sampling process were Condylostylus sp1 (Dolichopodidae) at 152 individuals; Cryptinae M1 (Ichneumonidae) at 127; Hercostomus sp1 at 126, Zygops tridentata Gyllenhal, 1836, at 102; and Condylostylus sp3 at 98.
The urban forest fragment that featured the highest abundance of specimens was Metropolitan Natural Park (PNM), at a total of 826 individuals, followed by Ciudad del Saber (CDS) at 676 individuals, Corozal (COR) at 280, and Albrook (ALB) at 256. In terms of the richness of the morphospecies, the fragment that featured the highest number was Ciudad del Saber, at 223 morphospecies, followed by PNM at 211, Corozal at 127, and Albrook, which exhibited the lowest level of richness at 118 morphospecies (Figure 2).
3.2. Alpha and Beta Diversity
The Shannon‒Wiener (H') diversity index values exceeded 3.0 for all fragments, thus indicating highly heterogeneous communities. The highest value was recorded in CDS (4.57), followed by both PNM and COR (4.4) and, at a slightly lower level, ALB (4.3). With respect to Margalef's wealth index, the values ranged between 21.1 and 34.07. Ciudad del Saber exhibited the highest value (34.07), followed by PNM (31.27), Corozal (22.36) and Albrook (21.1) (Table 2) (Figure 3).
The values obtained for the Pielou equity index were close to 1.0 for all the fragments under analysis, thus indicating a relatively uniform distribution of individuals among the present morphospecies. With respect to the Simpson dominance index (D), values close to zero were observed, thus suggesting that no morphospecies predominated significantly in the communities evaluated as part of this research. In addition, the high values of the inverse Simpson index (1-D), which were close to 1.0, reinforce the evidence indicating a high level of diversity and a balanced community structure in the urban forest fragments under investigation (Table 2).
With respect to beta diversity, the Bray‒Curtis dissimilarity index indicated a high level of variation in the composition of morphospecies among the sites (0.7558). The greatest differences were observed between Corozal and Ciudad del Saber, whereas Ciudad del Saber and Parque Natural Metropolitano were the most similar to one another. The Jaccard similarity index confirmed these patterns: CDS and PNM were the most similar fragments (0.3111), followed by Corozal and Albrook (0.289). The lowest level of similarity was observed between CDS and COR (Figure 4).
To complement the alpha and beta diversity indices, an estimate of the true richness and diversity of morphospecies was performed, which revealed marked differences among the urban fragments evaluated in this study. The Metropolitan Natural Park (PNM) and Ciudad del Saber (CDS) exhibited the highest abundances and levels of wealth observed in this context, at 211 and 223 morphospecies, respectively, which accounted for 51.88% and 48.13%, respectively, of the estimated wealth for each site. In contrast, the Corozal (COR) and Albrook (ALB) fragments exhibited a lower proportion of species than our estimations (36.70% and 42.8%, respectively), thus suggesting that a greater number of species were not detected during the sampling period. Despite these differences in richness, the true diversity observed (which was based on the distribution of abundances) was similar in all the fragments, as indicated by percentages that were higher than 95% with respect to the estimated values, thus indicating a relatively balanced community structure in terms of equity (Table 3).
The intersection analysis of species on the basis of Venn diagrams (A – D) allows us to visualize the distribution and coincidence of species among the urban forest fragments evaluated in this research: City of Knowledge (CDS), Metropolitan Natural Park (PNM), Corozal (COR) and Albrook (ALB). Diagram A (CDS – PNM – ALB) indicates a notable intersection between CDS and PNM, two fragments that feature higher levels of forest cover and closer geographic proximity, whereas ALB is characterized by a smaller number of shared species, thus suggesting a certain degree of ecological isolation. In diagram B (CDS – COR – ALB), CDS continues to exhibit the highest number of unique and shared species, and COR exhibits some moderate coincidences, possibly as a result of residual connectivity; ALB once again exhibits a low level of similarity with the other fragments. Diagram C (PNM – COR – ALB) reinforces this trend; namely, PNM appears as a middle point of intersection, whereas COR and ALB are characterized by a reduced coincidence of species. Finally, diagram D (CDS – PNM – COR) reveals the most ecologically relevant combination: CDS and PNM share a significant number of species, thus consolidating their role as nuclei of urban biodiversity, whereas COR is distinguished by a lower level of intersection, thus indicating its relative isolation (Figure 5).
Finally, the Diserud–Odegaard similarity index for all fragments is 0.543 (54%), thus indicating a moderate level of similarity in terms of the composition of the corresponding morphospecies. The multiple dissimilarity index estimated between the fragments is 0.407, thereby reflecting notable differences in the structure of the evaluated communities. We calculated alpha diversity indices (Shannon, Simpson, and Chao1) for each forest fragment, and then compared them using ANOVA and Kruskal-Wallis tests. While all fragments showed high levels of diversity (Shannon > 4.3; Simpson > 0.97), statistical tests did not detect significant differences among sites (Kruskal-Wallis χ² = 3.00, df = 3, p = 0.3916) (Table 4). However, a PERMANOVA based on Bray-Curtis distances revealed significant differences in species composition among fragments (R² = 0.0389, F = 2.91, p = 0.001), suggesting that although overall diversity levels were similar, each fragment harbored distinct insect communities. This pattern was also supported by the normalized Morisita-Horn similarity index (NDMI), which showed higher compositional similarity between Ciudad del Saber and Parque Natural Metropolitano, and lower similarity values between Corozal and the other fragments (Figure 6).
4. Discussion
The results obtained in this research regarding the diversity and composition of seven families of insects in urban forest fragments near Panama City highlights a similarity between communities (Diserud-Odegaard = 0.543), thus contradicting the initial hypothesis that a high level of similarity is to the result of the geographic proximity and shared ecological characteristics among the fragments. This divergence suggests that other factors that were not measured directly in this study could modulate the community structure observed in this context.
Our findings agree with those reported in [40], who demonstrated that the relationship between environmental heterogeneity and diversity is not always linear or predictable, even with respect to similar ecological groups. This finding reinforces the claim that biological communities can respond differently to environmental pressures, even under apparently homogeneous conditions.
In contrast, our results differ from those that have been reported in studies conducted in other contexts, such as in [41], who focused on Sydney, Australia, where wasp communities exhibit high levels of similarity among fragments, thus suggesting some degree of resistance to urban fragmentation. These findings also differ from the results of previous investigations conducted in Panama, such as the studies conducted in [42], who focused on butterflies, in [1], who focused on wasps of the Braconidae family, which have revealed highly similar compositions among fragments (as indicated by dissimilarity rates of 3% and 6%, respectively). This discrepancy could be the result of the fact that a greater number of families were included in our study, thus increasing the level of variability among samples, or to methodological differences pertaining to the calculation of dissimilarity.
At the taxonomic level, the subfamilies Crypthorynchinae (Curculionidae), Pristocerinae (Bethylidae) and Smiliinae (Membracidae) exhibit the greatest number of genera, a finding which is similar to the results of previous studies that have highlighted the high levels of diversity observed in this context in tropical regions [43,44,45,46]). With respect to the richness of the morphospecies, the fragments associated with the City of Knowledge and the Metropolitan Natural Park were associated with the highest values, possibly as a result of their greater connectivity, lower levels of isolation or status as protected areas. In contrast, Corozal and Albrook, which are more firmly embedded in an urban matrix, exhibited less wealth and abundance. This trend was also documented in [47], who noted that the size of the fragment and its connectivity with other forest masses significantly influence the population dynamics of the corresponding organisms.
The families of parasitoid insects, particularly Ichneumonidae and Bethylidae, dominated in terms of diversity and abundance; this finding is in line with the results of studies that have linked the high presence of such insights with the ready availability of hosts, as reported in [1] with respect to braconids in urban areas of Panama.
The alpha diversity indices (i.e., Shannon, Margalef, Simpson and inverse Simpson indices) support the presence of highly diverse entomological communities in all the fragments evaluated as part of this research. Similarly, the high level of equity and low level of dominance observed in this context suggest the absence of dominant species, which could be interpreted as an indicator of ecological stability and functional use of these spaces as possible biological corridors. These results highlight the critical importance of conserving urban forest remnants and promoting effective strategies that can help strengthen their connectivity within the urban landscape. In tropical contexts, in which entomological biodiversity is particularly high, these plant fragments function not only as reservoirs of species but also as key nodes within broader ecological networks. The ability of such fragments to sustain complex insect communities is closely related to factors such as fragment size, habitat quality, and, most notably, the degree of connectivity between a given fragment and other green fragments [48].
Recent studies have reported that well-connected green areas can facilitate gene flow and species dispersal and that they can mitigate the negative effects of isolation, such as the loss of species or decreases in key ecological interactions [49]. Urban vegetation, particularly when it is integrated into ecological corridors, can play a fundamental role in efforts to promote the resilience of entomological communities in response to disturbances associated with urbanization and climate change [15]. In this sense, urban planning must incorporate a landscape approach that prioritizes not only the amount of vegetation cover but also its spatial and functional configuration within the urban fabric, including by recognizing these fragments as key pieces in efforts to conserve biodiversity in tropical cities.
The values of observed richness (as opposed to the estimated richness) indicate that although forest fragments such as PNM and CDS exhibit more diverse entomological communities and are partially represented in the field, an important component of diversity remains unrecorded, which may be attributed to methodological limitations or the seasonal or cryptic behavior of certain species [50,51]). In fragments such as COR and ALB, the low proportion of species observed considering the estimated total suggests a higher level of complexity in the sampling or a degree of fragmentation that affects the detectability of species. However, the high level of agreement observed between the values of true diversity observed and estimated at all sites (> 95%) suggests that the communities in question are composed of species that are characterized by more equitable abundances, thus leading to the absence of excessive dominance. This pattern is characteristic of mature or functionally structured communities, even in urban contexts, and reinforces the need to expand sampling and conservation efforts pertaining to these urban fragments Soanes et al. 2023 [15,49]).
Venn diagrams suggest that the urban forest fragments located in the City of Knowledge (CDS) and the Metropolitan Natural Park (PNM) serve as key nodes of entomological biodiversity within the urban landscape of Panama City. The high levels of richness and significant number of species that are shared among these sites can be attributed to their larger size, better structural connectivity, and relative habitat continuity, given that these factors have been widely recognized as determinants of biological diversity in urban settings [13,49]. In contrast, the fragments located in Albrook and Corozal are characterized by more impoverished and differentiated communities, which could reflect the effects of ecological isolation and anthropogenic pressure that is typical of intensely modified urban matrices [14,48]).
The structure of the intersections observed among the sites highlights the importance of conserving and improving the connectivity among fragments such as CDS and PNM, as this situation can facilitate the movement of species, maintain gene flow and promote more stable and resilient communities [15]. Similarly, the differences observed among the fragments highlight the need to implement ecological restoration measures in locations such as Corozal and Albrook, with the aim of increasing their ability to serve as urban ecological habitats or corridors. This strategy, in addition to efforts to strengthen local biodiversity, can contribute to the ecological sustainability of the urban landscape; accordingly, this approach is in line with contemporary approaches that involve green infrastructure and resilient planning to combat climate change [49,52].
The lack of statistically significant differences in alpha diversity indices among forest fragments suggests a relatively homogeneous level of insect diversity across urban patches. This result aligns with studies indicating that small but structurally complex urban green spaces can support high insect diversity regardless of size or isolation [9,53]. However, the significant differences found in community composition (PERMANOVA) and the contrasting similarity values in the NDMI matrix confirm that each fragment hosts a distinct assemblage of species. Similar patterns have been observed in other tropical urban systems, where beta diversity is driven more by habitat heterogeneity and microclimatic variation than by alpha diversity metrics alone [54,55]. These findings reinforce the importance of preserving multiple forest fragments within urban landscapes, as each contributes uniquely to regional biodiversity.
5. Conclusion
This study provides evidence indicating the high diversity of insects in urban forest fragments located in Panama City, even in environments that have been greatly transformed. The richness of morphospecies, the equity among different communities and the absence of dominance indicates that these fragments host structurally complex entomological communities. Although greater similarity in species composition was expected because of the proximity and shared ecological conditions among the fragments under investigation, the results are characterized by a notable level of heterogeneity, thus suggesting that other unmeasured factors could influence the organization of these communities.
Estimates of the true diversity and richness of these areas revealed that although not all of the expected diversity was captured, the level of representation of the most common species was high, and the communities exhibited a well-balanced internal structure in terms of abundance. In turn, the Venn diagrams confirmed that fragments such as the City of Knowledge and Metropolitan Natural Park share a greater number of species with each other, in contrast to Corozal and Albrook, which are characterized by higher levels of isolation and fewer intersections, possibly because of the degree of fragmentation that these areas exhibit and the surrounding urban matrix. These tools made it possible to identify key nodes of diversity within the urban landscape, as well as fragments that require attention in terms of ecological restoration.
Differences among the investigated fragments in terms of richness and abundance could be related to the degree of isolation, the size of the fragment or the characteristics of the surrounding matrix. However, these hypotheses should be evaluated in future studies based on the integration of landscape metrics and connectivity analysis. The findings presented here highlight the potential role played by urban forest fragments as reservoirs of biodiversity and possible biological corridors. The conservation and management of these areas should be viewed as complementary strategies within sustainable urban development plans in tropical cities such as Panama City.
Author Contributions
Conceptualization, J.A. and E.M.; Methodology, J.A. and E.M.; Investigation, J.A. and E.M.; Formal analysis, J.A. and E.M.; Writing—original draft, J.A. and E.M.; Writing—review and editing, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by multiple grants and funding programs. Financial support was provided under contract No. 001-2019 (Educational Collaboration Agreement No. 49-2018). The author also acknowledges the economic support received from the National Research System (SNI) of SENACYT under contract No. 050-2022. This research was further supported by the Research Mobility Call under contract DDCCT No. 004-2023, corresponding to the project entitled "Urban forests as a resource to face climate change." Additional funding was provided by the University of Panama through the Research Fund Call under code CUFI-2021-EP-CNET-001. The support of all these institutions was essential for the completion of this study.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
I thank the GB Fairchild Invertebrate Museum of the University of Panama, the Central American Master's Program in Entomology, and to the National Secretariat of Science, Technology and Innovation (SENACYT) for the financial support provided under grant contract No. 001-2019 (Educational Collaboration Agreement No. 49-2018). Similarly, I thank the National Research System (SNI) of SENACYT for the economic subsidies received under contract No. 050-2022. This research was supported by the Call for Research Mobility under contract DDCCT No. 004-2023, which corresponds to the project entitled "Urban forests as a resource to face climate change." Similarly, I thank the Call for Research Funds of the University of Panama for the support granted under code CUFI-2021-EP-CNET-001. The support of all these institutions was essential for this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rodríguez, L.A.; Medianero, E. The composition of braconid wasp communities in three forest fragments in a tropical lowland forest of Panama. BMC Ecol Evol. 2022, 22, 98. [Google Scholar] [CrossRef]
- Medianero, E.; Santos Murgas, A. Insectos asociados a los bosques urbanos de la ciudad de Panamá; D'McPherson: Panamá, 2023. [Google Scholar]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of earth's ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Grimm, N.B.; Grove, J.M.; Pickett, S.T.A.; Redman, C.L. Integrated approaches to long-term studies of urban ecological systems. BioScience 2000, 50, 571–584. [Google Scholar] [CrossRef]
- Alberti, M.; Marzluff, J.M.; Shulenberger, E.; Bradley, G.; Ryan, C.; Zumbrunnen, C. Integrating humans into ecology: opportunities and challenges for studying urban ecosystems. BioScience 2003, 53, 1169–1179. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Didham, R.K.; Hammond, P.M.; Lawton, J.H.; Eggleton, P.; Stork, N.E. Beetle species responses to tropical forest fragmentation. Ecol Monogr 1998, 68, 295–323. [Google Scholar] [CrossRef]
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. The ecological impacts of nighttime light pollution: a mechanistic appraisal. Biol. Rev. 2013, 88, 912–927. [Google Scholar] [CrossRef]
- Johnson, M.T.J.; Munshi-South, J. Evolution of life in urban environments. Science 2017, 358, eaam8327. [Google Scholar] [CrossRef]
- Laurance, W.F. Comparative responses of five arboreal marsupials to tropical forest fragmentation. J. Mammal. 1990, 71, 641–653. [Google Scholar] [CrossRef]
- McDonnell, M.J.; Pickett, S.T.A. Ecosystem structure and function along urban–rural gradients: an unexploited opportunity for ecology. Ecology 1990, 71, 1232–1237. [Google Scholar] [CrossRef]
- Bergman, K.-O.; Dániel-Ferreira, J.; Milberg, P.; Öckinger, E.; Westerberg, L. Butterflies in Swedish grasslands benefit from forest and respond to landscape composition at different spatial scales. Landsc. Ecol. 2018, 33, 2189–2204. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Aronson, M.F.J.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the city: key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Liu, M.; Russo, A. Urban green infrastructure: bridging biodiversity conservation and sustainable development through adaptive management approaches. Front. Ecol. Evol. 2023, 11, 1440477. [Google Scholar] [CrossRef]
- Ministerio de Ambiente. Proyecto Corredor Biológico Mesoamericano del Atlántico Panameño, fase II; https://www.gacetaoficial.gob.pa/pdfTemp/28285_A/GacetaNo_28285a_20170524.pdf; Gaceta Oficial Digital No. 28285-A: Panamá, 2017. [Google Scholar]
- Valdéz, L.; Ortiz, O.; Medianero, E. Diversidad y estructura de las comunidades de mariposas diurnas (Lepidoptera: Rhopalocera) en fragmentos de vegetación urbanas adyacentes a la Ciudad de Panamá. In Insectos asociados a los bosques urbanos de la ciudad de Panamá; Medianero, E., Santos Murgas, A., Eds.; D'McPherson: Panamá, 2023; pp. 23–71. [Google Scholar]
- Autoridad Nacional del Ambiente (ANAM). Informe nacional sobre el estado del ambiente 2010; ANAM: Panamá, 2010. [Google Scholar]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: a review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Parque Natural Metropolitano. Plan de manejo del Parque Natural Metropolitano; Parque Natural Metropolitano: Panamá, 1999. [Google Scholar]
- Muirhead-Thomson, R.C. Trap Responses of Flying Insects. The Influence of Trap Design on Capture Efficiency; Academic Press: London, UK, 1991. [Google Scholar]
- Karlsson, D.; Hartop, E.; Forshage, M.; Jaschhof, M.; Ronquist, F. The Swedish Malaise trap project: a 15 year retrospective on a countrywide insect inventory. Biodivers. Data J. 2020, 8, e47255. [Google Scholar] [CrossRef]
- Hanson, P.E.; Gauld, I.D. The Hymenoptera of Costa Rica; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Brown, B.V. Manual of Central American Diptera; NRC Research Press: Ottawa, Canada, 2009. [Google Scholar]
- Evans, H.E. A revision of the genus Apenesia in the Americas (Hymenoptera, Bethylidae). Bull. Mus. Comp. Zool. 1963, 130, 249–359. [Google Scholar]
- Evans, H.E. A synopsis of the American Bethylidae (Hymenoptera, Aculeata). Bull. Mus. Comp. Zool. 1964, 132, 1–222. [Google Scholar]
- Santos Murgas, A.; Gonzales Domínguez, P.E. Biosistemática de la familia Bethylidae (Insecta: Hymenoptera) en Panamá; Tesis de licenciatura, Universidad de Panamá: Panamá, 2001. [Google Scholar]
- Aldrete, A.N.G.; Carrejo, N.S.; Mendivil, J.; Calderón, N.; Saenz, O.; Panche, J.; Román, C.; Obando, R.G. Checklist of ‘Psocoptera’ (Psocodea) of Colombia and identification key to the families. Dugesiana 2018, 25, 77–103. [Google Scholar] [CrossRef]
- Hanson, P.E.; Fernández, F. Los Membracidae (Hemiptera: Auchenorrhyncha) de Costa Rica; Instituto Nacional de Biodiversidad (INBio): Santo Domingo de Heredia, Costa Rica, 2006. [Google Scholar]
- Flynn, D.J. Checklist of treehoppers of Panama (Hemiptera: Membracidae) with a list of checklists and keys to the Nearctic and Neotropical fauna. Zootaxa 2012, 3405, 35–63. [Google Scholar] [CrossRef]
- Oliver, I.; Beattie, A.J. Invertebrate morphospecies as surrogates for species: a case study. Conserv. Biol. 1996, 10, 99–109. [Google Scholar] [CrossRef]
- Moreno, C.E.; Sánchez-Rojas, G.; Pineda, E.; Escobar, F. Evaluación de la diversidad en comunidades biológicas. In Capital natural de México, vol. I: Conocimiento actual de la biodiversidad; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO): México, 2006; pp. 21–30. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, USA, 1988. [Google Scholar]
- Hulbert, S.F.; Morrison, S.J.; Klawitter, J.J. Compatibility of porous ceramics with soft tissue; application to tracheal prostheses. J. Biomed. Mater. Res. 1971, 5, 269–279. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Real, R.; Vargas, J.M. The probabilistic basis of Jaccard's index of similarity. Syst. Biol. 1996, 45, 380–385. [Google Scholar] [CrossRef]
- Diserud, O.H.; Odegaard, F. A multiple-site similarity measure. Biol. Lett. 2007, 3, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Peet, R.K. The measurement of species diversity. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- Moreno, C.E.; Rodríguez, P.; Zuria, I. Diversidad alfa, beta y gamma. In La biodiversidad en un mundo cambiante: fundamentos teóricos y métodos de estudio; Moreno, C.E., Ed.; Universidad Autónoma del Estado de Hidalgo: México, 2018; pp. 119–144. [Google Scholar]
- Heidrich, L.; Barlow, J.; Gómez, C.; Lees, A.C.; Nally, R.M.; Schleuning, M. Disentangling direct and indirect effects of forest fragmentation on plant–frugivore interactions. Ecology 2020, 101, e02980. [Google Scholar] [CrossRef]
- Christie, F.J.; Hochuli, D.F. Responses of wasp communities to urbanization: effects on community resilience and species diversity. J. Insect Conserv. 2009, 13, 213–221. [Google Scholar] [CrossRef]
- Valdés, L. Estructura de las comunidades de mariposas diurnas (Lepidoptera: Rhopalocera) en fragmentos de vegetación en La Ciudad de Panamá; Universidad de Panamá: Panamá, 2018. [Google Scholar]
- Azevedo, C.O. Synopsis of the neotropical Dissomphalus (Hymenoptera, Bethylidae). Zootaxa 2003, 338, 1–74. [Google Scholar] [CrossRef]
- Azevedo, C.O. Characterization of the types of the neotropical Pseudisobrachium (Hymenoptera: Bethylidae), with a key to species. Rev. Bras. Zool. 2008, 25, 737–801. [Google Scholar] [CrossRef]
- Santos Murgas, A.; González, D. Distribución de especies de la subfamilia Epyrinae (Hymenoptera: Bethylidae) en Panamá. Tecnociencia 2006, 8, 37–50. [Google Scholar]
- Grebennikov, V.V.; Newton, A.F. Good-bye Scydmaenidae, or why the ant-like stone beetles should become megadiverse Staphylinidae sensu latissimo (Coleoptera). Eur. J. Entomol. 2009, 106, 275–301. [Google Scholar] [CrossRef]
- Ruiz-Guerra, B.; Renton, K.; Dirzo, R. Consequences of fragmentation of tropical moist forest for birds and their role in predation of herbivorous insects. Biotropica 2012, 44, 228–236. [Google Scholar] [CrossRef]
- Wang, J.; Ren, Z.; Ding, Y. Supporting function of vegetation in urban riparian ecological corridors: a case study in Beijing, China. Sci. Total Environ. 2023, 857, 159377. [Google Scholar] [CrossRef]
- Soanes, K.; Sievers, M.; Chee, Y.E.; Egan, E. Planning for urban biodiversity on shifting social-ecological grounds: a case study of Narrm-Melbourne. Ecol. Solut. Evid. 2023, 4, e70005. [Google Scholar] [CrossRef]
- Magurran, A.E. Species abundance distributions: pattern or process? Funct. Ecol. 2005, 19, 177–181. [Google Scholar] [CrossRef]
- Moreno, C.E.; Barragán, F.; Pineda, E.; Pavón, N.P. Reanálisis de la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas. Rev. Mex. Biodivers. 2011, 82, 1249–1261. [Google Scholar] [CrossRef]
- European Environment Agency. Green spaces and corridors in urban areas. Climate-ADAPT, 2023. Available online: https://climate-adapt.eea.europa.eu/en/metadata/knowledge-items/green-spaces-and-corridors-in-urban-areas (accessed on 18 April 2025).
- Mata, L.; Andersen, A.N.; Morán-Ordóñez, A.; Hahs, A.K.; Backstrom, A.; Ives, C.D.; Bickel, D.; Duncan, D.; Palma, E.; Thomas, F.; et al. Indigenous plants promote insect biodiversity in urban greenspaces. Ecol. Appl. 2021, 31, e02309. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, L.; Zhang, H.; Liu, X.; Li, Q. Seasonal variation and driving factors of beta diversity of stream insects in a subtropical urban landscape. Urban Ecosyst. 2023, 26, 123–135. [Google Scholar] [CrossRef]
- Lagucki, E.; Burdine, J.D.; McCluney, K.E. Urbanization alters communities of flying arthropods in parks and gardens of a medium-sized city. PeerJ 2017, 5, e3620. [Google Scholar] [CrossRef]
Figure 1.
Sampling sites in the urban forests associated with Panama City.
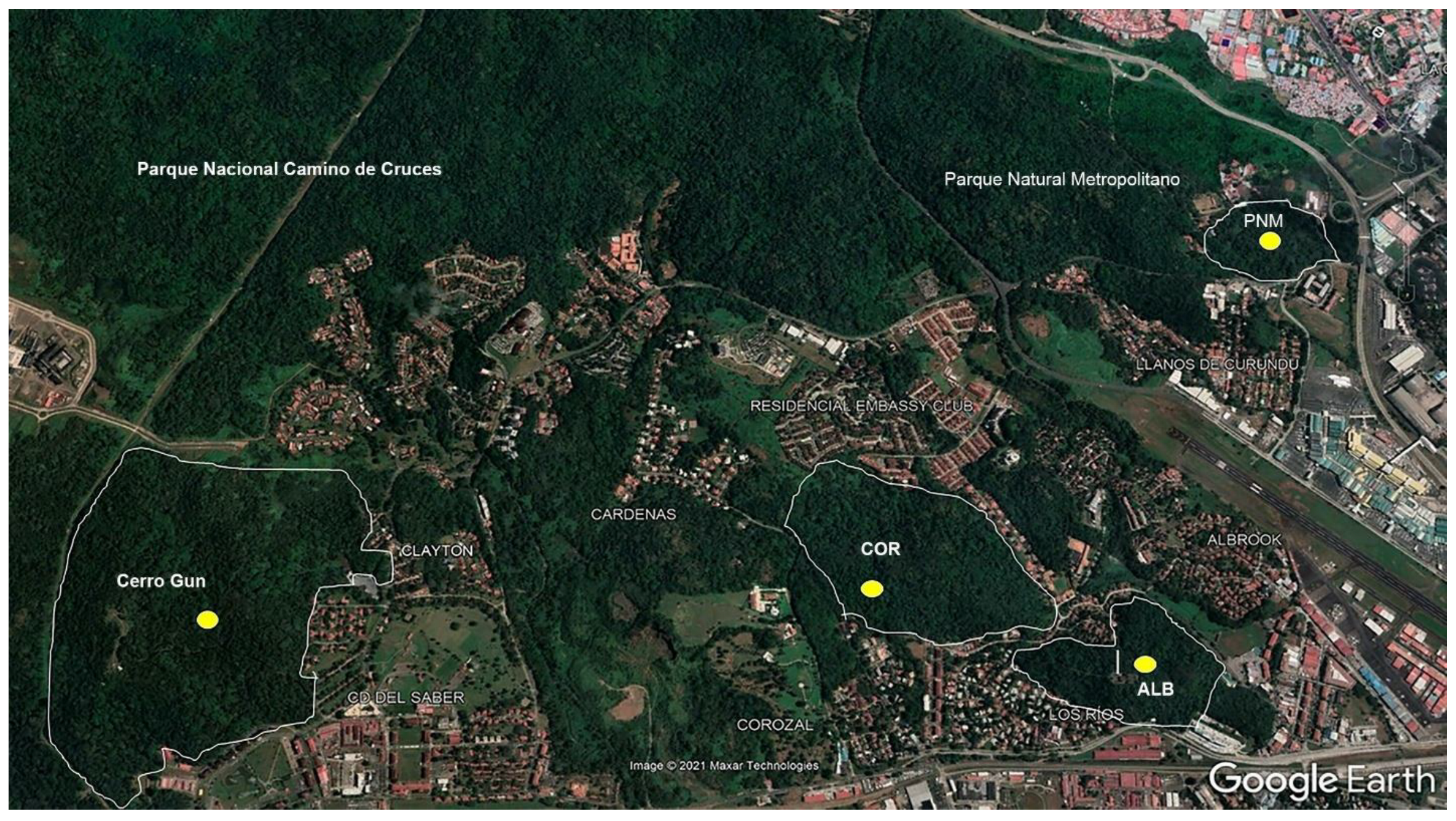
Figure 2.
Number of individuals and morphospecies estimated in each of the four urban forest fragments sampled in Panama City.
Figure 2.
Number of individuals and morphospecies estimated in each of the four urban forest fragments sampled in Panama City.
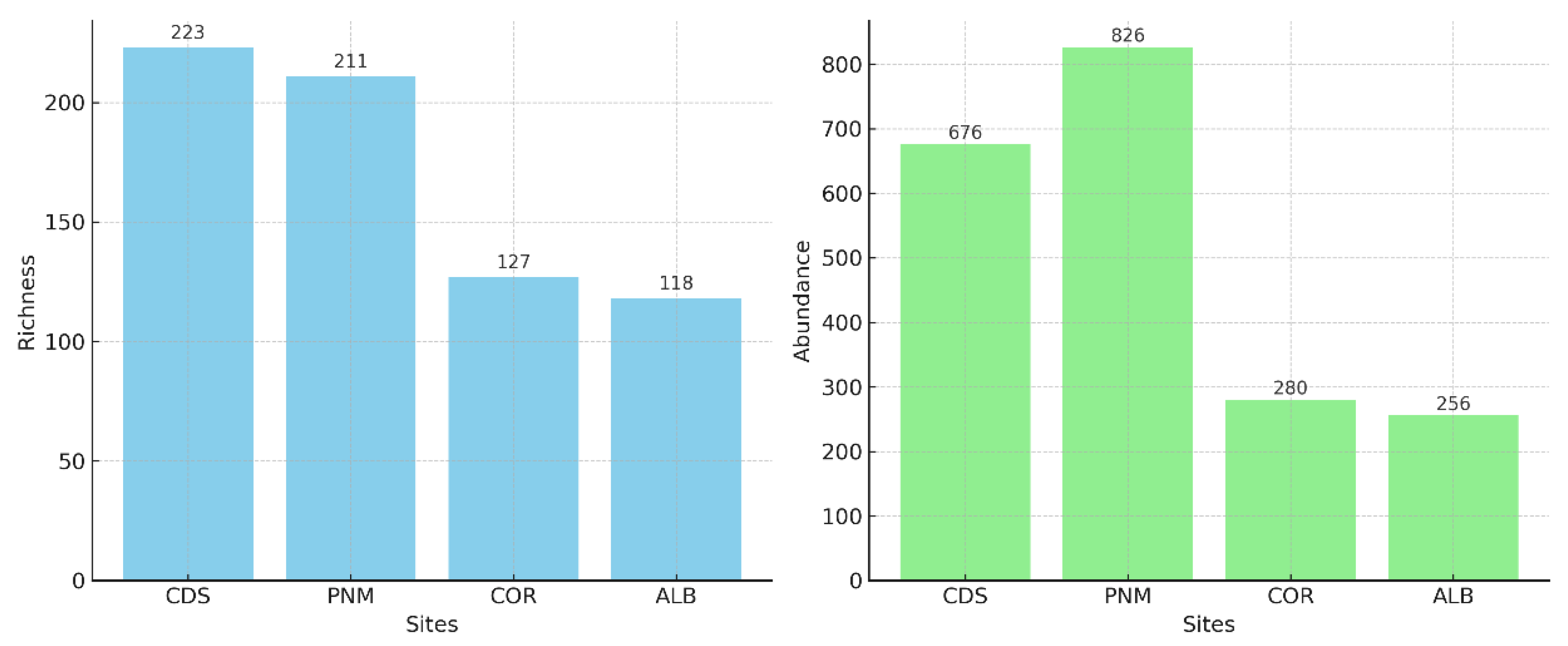
Figure 3.
Shannon index (H'), in which context the diversity of the species in the urban forest fragments studied is observed.
Figure 3.
Shannon index (H'), in which context the diversity of the species in the urban forest fragments studied is observed.
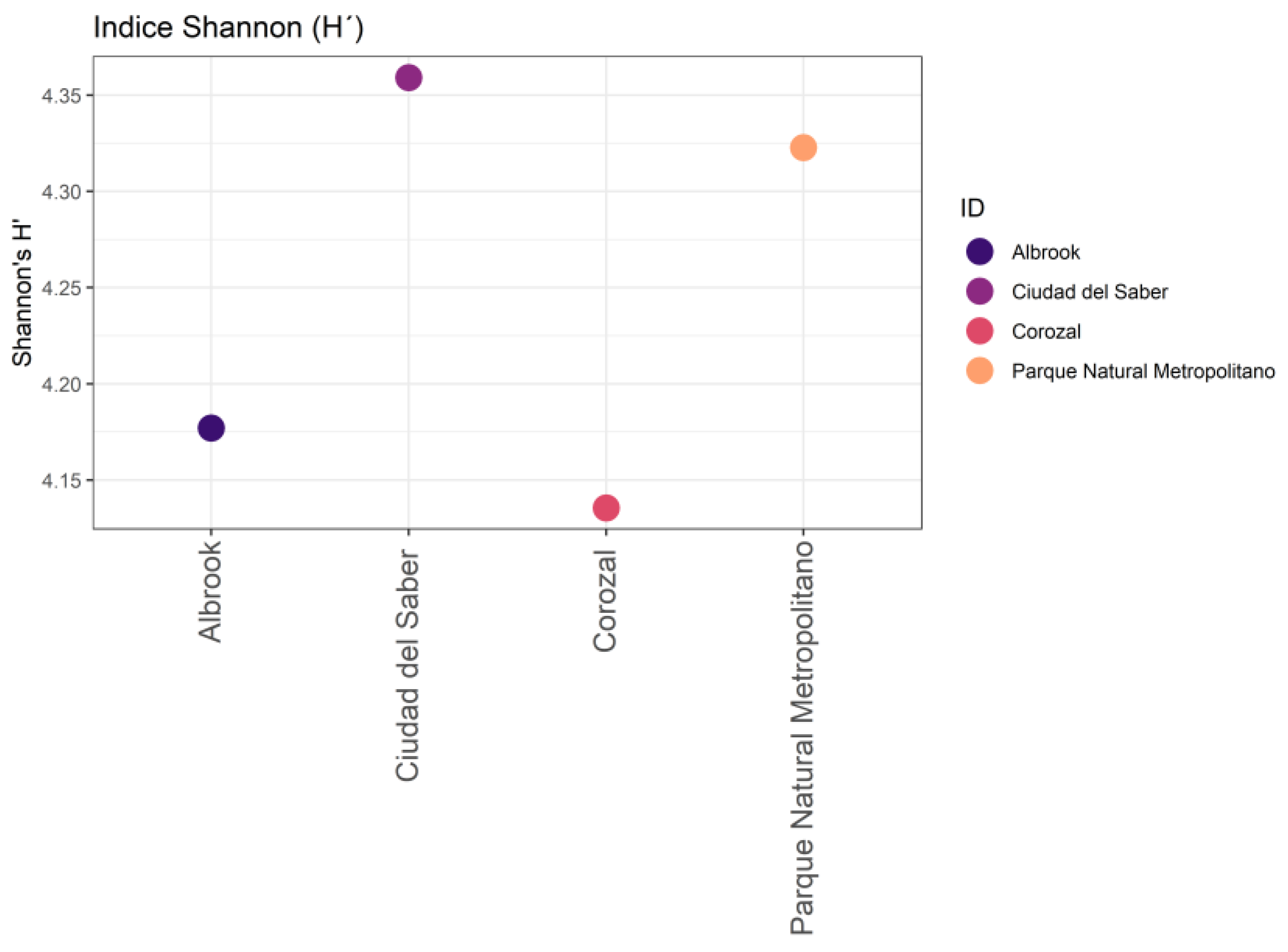
Figure 4.
Bray‒Curtis and Jaccard similarity indices of the morphospecies of the families under investigation in the different study fragments.
Figure 4.
Bray‒Curtis and Jaccard similarity indices of the morphospecies of the families under investigation in the different study fragments.
Figure 5.
Venn diagrams that illustrate the intersections of insect morphospecies among three urban forest fragments in different combinations. The Ciudad del Saber (CDS), Parque Natural Metropolitano (PNM), Corozal (COR) and Albrook (ALB) sites are compared. (A) Intersections among CDS, PNM and ALB; (B) CDS, COR and ALB; (C) PNM, COR and ALB; and (D) CDS, PNM and COR.
Figure 5.
Venn diagrams that illustrate the intersections of insect morphospecies among three urban forest fragments in different combinations. The Ciudad del Saber (CDS), Parque Natural Metropolitano (PNM), Corozal (COR) and Albrook (ALB) sites are compared. (A) Intersections among CDS, PNM and ALB; (B) CDS, COR and ALB; (C) PNM, COR and ALB; and (D) CDS, PNM and COR.
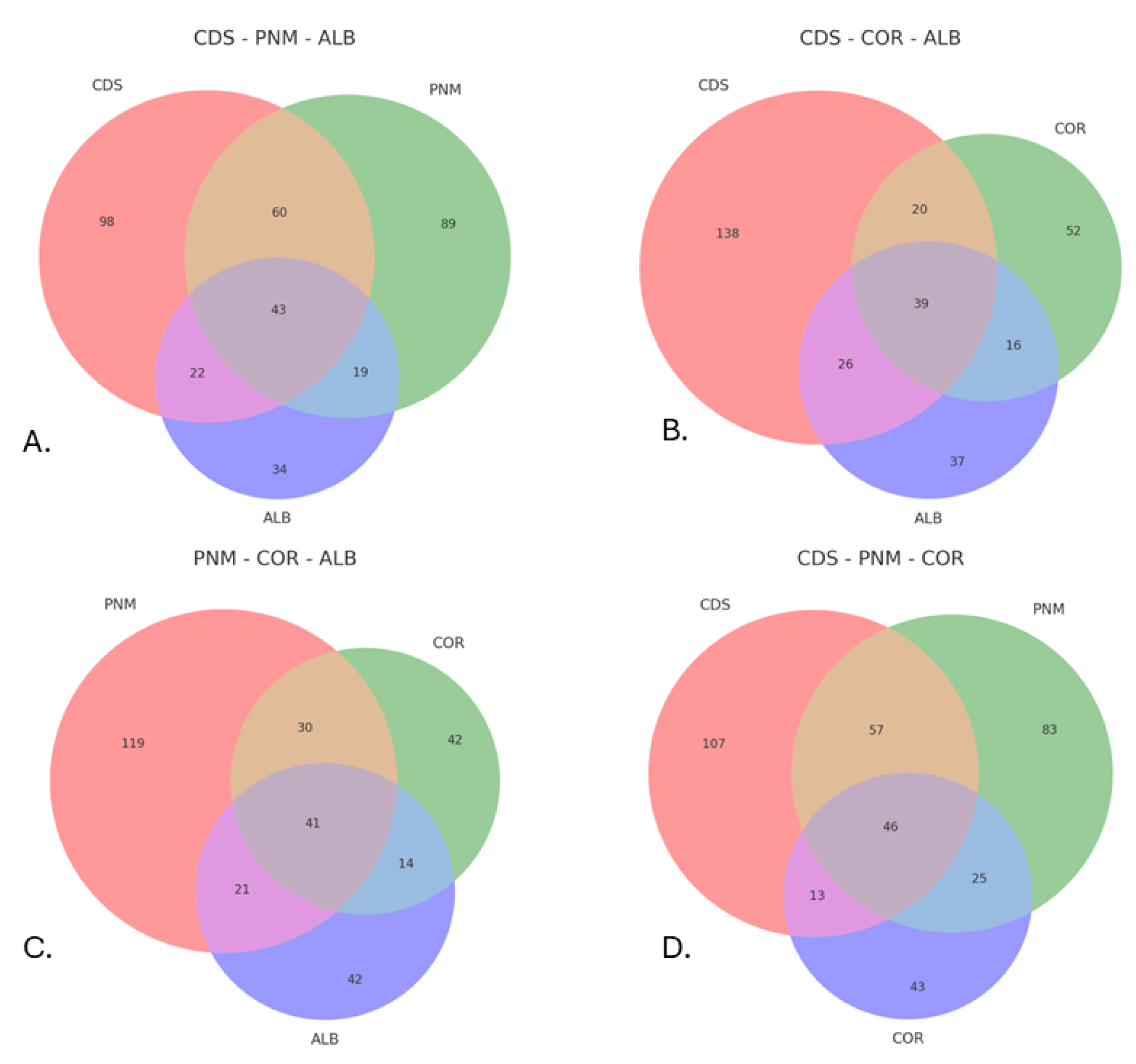
Figure 6.
Heatmap showing the pairwise similarity between insect communities in four urban forest fragments in Panama City, based on the normalized Morisita-Horn index (NDMI). Higher values (close to 1) indicate greater similarity in species composition, while lower values reflect higher dissimilarity. The highest similarity was observed between Ciudad del Saber (CDS) and Parque Natural Metropolitano (PNM), while Corozal (COR) exhibited the greatest dissimilarity relative to the other fragments.
Figure 6.
Heatmap showing the pairwise similarity between insect communities in four urban forest fragments in Panama City, based on the normalized Morisita-Horn index (NDMI). Higher values (close to 1) indicate greater similarity in species composition, while lower values reflect higher dissimilarity. The highest similarity was observed between Ciudad del Saber (CDS) and Parque Natural Metropolitano (PNM), while Corozal (COR) exhibited the greatest dissimilarity relative to the other fragments.
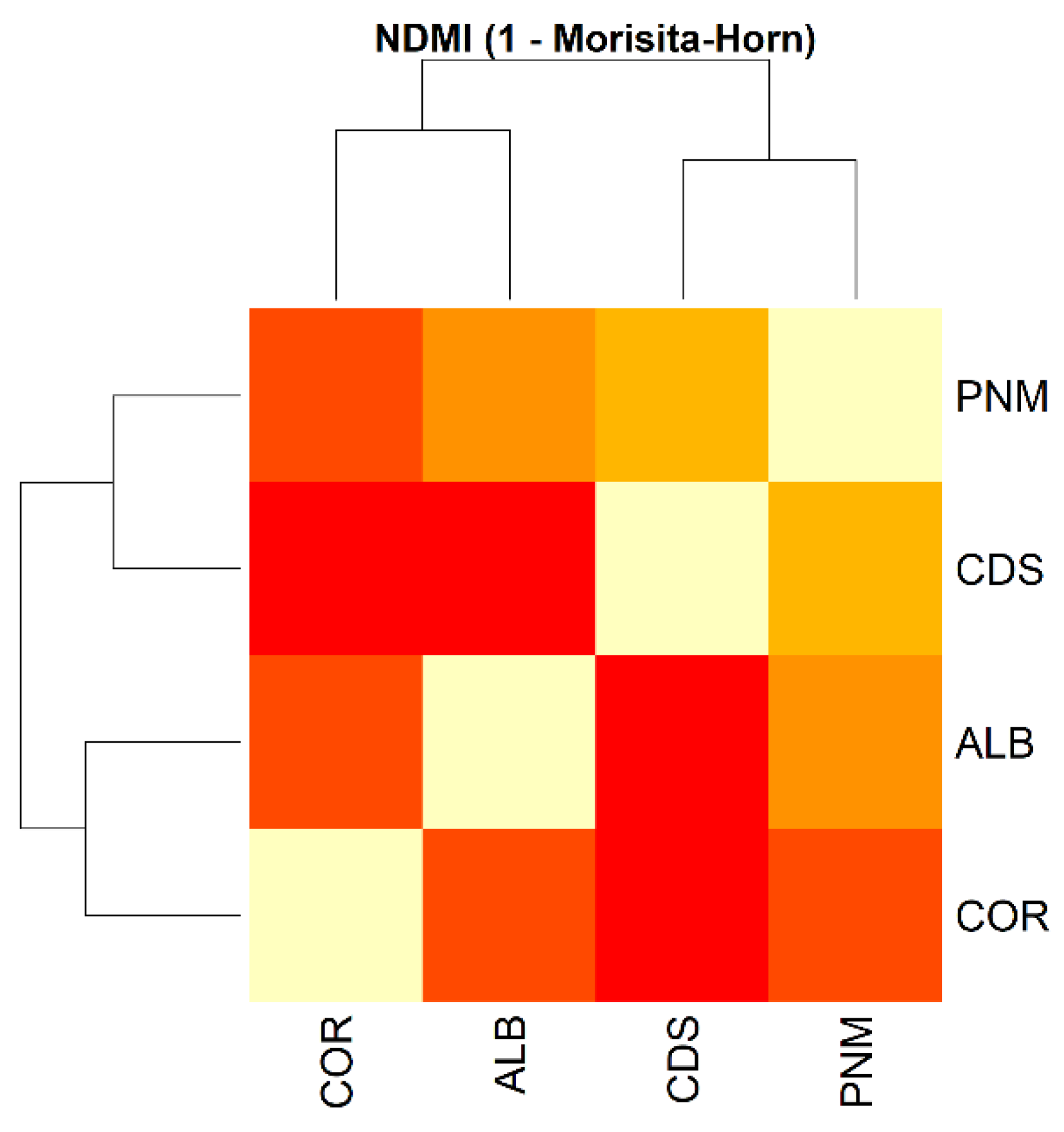

Table 1.
Study sites in Panama City, Panama, Panama.
| Sites | Geographical location | Altitude (meters) | Vegetation | Annual temperature (°C) |
|---|---|---|---|---|
| PNM | 8°59'41.55''N-79°32'35.22''O | 0- 150 | Tropical dry forest | 28 |
| CDS | 9°00'24.3"N-79°35'05.2"W | 0- 100 | Tropical dry forest | 27.5 |
| COR | 08° 59´19.34″N-079°34´11.83″W | 30- 60 | Overlapping deciduous forest. | 26.5 |
| ALB | 08°58'37.49"N-079°33'43.82"W | 20- 80 | Overlapping deciduous forest. | 26.5 |
Table 2.
Alpha (α) diversity indices of the urban forest fragments under investigation.
| Albrook | Ciudad del Saber | Corozal | Parque Natural Metropolitano | |
| Taxas | 118 | 223 | 127 | 211 |
| Individuos | 256 | 676 | 280 | 826 |
| Dominancia (D) | 0.02515 | 0.02836 | 0.01946 | 0.02785 |
| Simpson (1-D) | 0.9749 | 0.9716 | 0.9805 | 0.9721 |
| Shannon (H´) | 4.302 | 4.575 | 4.4 | 4.434 |
| Margalef | 21.1 | 34.07 | 22.36 | 31.27 |
| Equidad (J) | 0.9018 | 0.8461 | 0.9083 | 0.8285 |
Table 3.
Observed, estimated, and exclusive richness, alongside observed and estimated true diversity in Panama (the percentage of richness and diversity obtained in the field with respect to the estimations are shown in parentheses).
Table 3.
Observed, estimated, and exclusive richness, alongside observed and estimated true diversity in Panama (the percentage of richness and diversity obtained in the field with respect to the estimations are shown in parentheses).
| Sites | Abundance | True diversity | |||
| Obs. | Est. | Obs. | Est. | ||
| PNM | 826 | 211 | 406.7 (51.88) |
103.307 | 106.810 (96.72) |
| CDS | 676 | 223 | 463.3 (48.13) | 126.970 | 133.482 (95.12) |
| COR | 280 | 127 | 346.0 (36.70) | 119.806 | 118.624 (1.00) |
| ALB | 256 | 118 | 275.5 (42.8%) | 108.86 | 111.97 (97.22%) |
Table 4.
Alpha diversity indices of insect communities across four urban forest fragments in Panama City. Values are shown for Shannon diversity, Simpson dominance, and Chao1 estimated richness. The final row presents the mean and standard deviation (SD) across all fragments. Although diversity was consistently high across sites, a Kruskal-Wallis test found no statistically significant differences among fragments for the Shannon index (χ² = 3.00, df = 3, p = 0.3916).
Table 4.
Alpha diversity indices of insect communities across four urban forest fragments in Panama City. Values are shown for Shannon diversity, Simpson dominance, and Chao1 estimated richness. The final row presents the mean and standard deviation (SD) across all fragments. Although diversity was consistently high across sites, a Kruskal-Wallis test found no statistically significant differences among fragments for the Shannon index (χ² = 3.00, df = 3, p = 0.3916).
| Sites | Shannon | Simpson | Chao1 |
| Albrook | 4.30 | 0.975 | 256.8 |
| Ciudad del Saber | 4.58 | 0.972 | 421.2 |
| Corozal | 4.40 | 0.981 | 320.7 |
| Parque Natural Metropolitano | 4.43 | 0.972 | 422.5 |
| Mean ± SD | 4.43 ± 0.12 | 0.975 ± 0.004 | ± 78.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



