
Submitted:
05 January 2025
Posted:
06 January 2025
You are already at the latest version
Abstract
The covert mortality nodavirus (CMNV), an emerging pathogen that infects several species severely in aquaculture including marine shrimps, freshwater prawns and crabs, has been detected in both wild Artemia and commercial Artemia cysts. Utilization of Artemia from unknown sources can impose considerable biosecurity risks in hatchery operations, therefore effective hatching methods to eliminate the pathogenic potential from CMNV-positive (CMNV+) Artemia cysts are urgently needed. In this work, we assessed the efficacy of three treatment methods of disinfecting CMNV+ Artemia cysts during hatching: (l) Decapsulation of Artemia cysts with Na2CO3 (sodium carbonate) and NaClO (sodium hypochlorite) at various durations before hatching; (2) Application of different concentrations of formalin in hatching water; (3) Combinations of decapsulating Artemia cysts and formalin-treated hatching water. Hatching CMNV+ Artemia with disinfected seawater only served as the control. The results showed that: (l) The virus located on the cyst shells attached/infected Artemia larvae during hatching and remained active for a prolonged time of 7 days. (2) The viral load of empty shells decreased with the decapsulation treatment time. After a 45-minute treatment of decapsulation, CMNV on shells, as well as larvae, were eliminated successfully. Furthermore, decapsulation shortened the hatching time of the cysts from 19 h to 12 h, (3) 10 ppm formalin in the hatching water could block the transmission of CMNV from the shells to the newly hatched larvae, although at this level it was insufficient to eliminate the virus from the shells completely. While use of 30 ppm formalin or higher dosage could eliminate CMNV, however, it also reduced the hatching rates of the Artemia cysts. (4) Combination of decapsulation (treated with Na2CO3-NaClO for 15 min) and 10 ppm formalin in hatching water effectively eliminated the CMNV. This study developed a practical, effective, and reliable treatment method for hatching Artemia to ensure biosecurity in aquaculture hatcheries.
Keywords:
Covert mortality nodavirus (CMNV)
; Artemia cyst
; Decapsulation
; Biosecurity
1. Introduction
In 2008–2009, a disease called Covert Mortality Disease (CMD) broke out in shrimp farming areas in several southern provinces of China. Shrimps infected with CMD often exhibit symptoms such as paler body color, empty intestines, atrophy of the hepatopancreas, slow growth, and eventually die covertly in deep water areas at the bottom of the aquaculture pond [1]. The disease can result in a 60–80% mortality rate and has caused serious economic loss to China's shrimp farming industry [2]. A new nodavirus named Covert mortality nodavirus (CMNV) was later identified as the viral pathogen of CMD [2]. Epidemiological investigations showed that CMNV was not only prevalent in China, but also in other shrimp farming countries, e.g. Vietnam, Ecuador, India, Thailand, Indonesia and Mexico [3,4]. In addition to shrimp, CMNV can also infect other cultured crustaceans, such as the freshwater prawn Macrobrachium rosenbergii [4,5]. In shrimp ponds, several coexisting species including brine shrimp Artemia sinica, barnacle Balanus sp., the rotifer Brachionus urceus, the polychaete Perinereis aibuhitensis, and the amphipod Corophium sinense, could be the vectors and reservoir of CMNV [6,7]. We can also detect CMNV in commercial Artemia cysts, the important aquatic feed, therefore, utilization of Artemia from unknown sources can impose considerable biosecurity risks on aquaculture.
In aquaculture breeding, Artemia cysts are widely used due to their easy storage, convenient hatching, and high nutritional value [8]. Before hatching into nauplii, Artemia cysts were usually decapsulated (i.e., removal of the outer layer of the cyst shell), which can bring many advantages such as disinfection of the cysts, possible direct ingestion and digestion of decapsulated cysts by the larvae [9]. The decapsulation solution usually contains hypochlorite. Although hypochlorite can disinfect the cysts [9], whether CMNV can be eliminate during decapsulation is still unknown. Here we developed an effective and reliable treatment method for decapsulating and hatching Artemia cysts to ensure biosecurity in aquaculture hatcheries.
2. Materials and Methods
2.1. Artemia Cysts and the Detection of CMNV
2.1.1. Artemia Cysts
A can of Artemia cyst (500 g) was provided by Guangdong Haid Group Ltd. (Guangzhou, China).
2.1.2. RNA Extraction
The RNA extraction was conducted using TaKaRa MiniBEST Viral RNA/DNA Extraction Kit Ver.5.0 (TaKaRa, Beijing, China) according to kit instructions. Briefly, 10 mg of Artemia cysts was grinded in liquid nitrogen. After grinding, 200 μL of PBS solution, 200 μL of Buffer VGB, 20 μL Proteinase K (20 mg/mL) and 1.0 μL Carrier RNA were added to the grinded tissue. And then the solution was incubated at 56℃ for 10 min. After centrifuging at 12,000 rpm for 2 min, the precipitate was washed with Buffer RWA and Buffer RWB. Finally, the RNA was eluted in spin column by centrifuging at 12,000 rpm for 2 min, and stored with RNase free dH2O at -20℃.
2.1.3. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
The CMNV detection was performed using CMNV Fluorescent RT-PCR Detection Kit (Qingdao Lijian Bio-Tech Co., Ltd., Qingdao, China). Firstly, to prepare the RT-PCR reaction liquid, the RT-PCR enzyme reaction liquid was mixed with the TagMan fluorescence primer probes, which target on the CMNV RNA1 and RNA2 genome segments [10]. Each PCR reaction tube was added with 20 μL RT-PCR reaction liquid, and then 5 μL RNA sample. The RT-PCR was performed on a StepOne Real-Time PCR System (Thermo Fisher Scientific, USA) and the procedure was as follow: reverse transcription at 50℃ for 15 minutes, initial denaturation at 95℃ for 1 minute, a total of 40 reaction cycles including denature at 95℃ for 10 seconds, extension at 60℃ for 30 seconds. FAM fluorescence signals were collected at 60℃.
2.1.4. Result Determination
When the sample had a typical amplification curve and the cycle threshold (Ct) value ≤37, it could be judged as CMNV-positive (CMNV+). When there was no Ct value or typical amplification curve of the sample, it was determined as CMNV-negative (CMNV-). The Artemia cysts in this study showed a typical amplification curve and had a Ct value of 33.0, we therefore determined that they are CMNV+. These CMNV+ Artemia cysts were then used for the following treatments and assessing the efficacy of disinfecting CMNV.
2.2. Treatments of CMNV+ Artemia Cysts
Three treatments were applied to assess their efficacy of disinfecting CMNV+ Artemia cysts during hatching: (l) Before hatching with disinfected seawater, the Artemia cysts were decapsulated with 1% Na2CO3 (sodium carbonate) and 21% NaClO (sodium hypochlorite) for 5, 15, 30 and 40 min, respectively; (2) Application of different concentrations (10, 30 and 60 ppm) of formalin in the hatching water; (3) Combinations of decapsulating Artemia cysts and formalin-treated hatching water, in which two combinations were applied, ① 5 min–decapsulation followed by adding 5 ppm formalin in the hatching water; ② 15 min–decapsulation followed by adding 10 ppm formalin in the hatching water. Hatching CMNV+ Artemia with disinfected seawater only served as control.
For each treatment, 0.8 gram cysts were used and the cysts were hatched at 25–26 °C. After hatching, 100 larvae/unhatched cysts were randomly selected and observed under stereomicroscope to count the hatching rate. The Artemia larvae were fed with green alga (Chlorella sp.) after hatching.
2.3. CMNV Detection and Viral Load Quantification
CMNV detection and viral load quantification were performed for (1) the newly hatched larvae (L); (2) the empty shells picked out after hatching (S); (3) the larvae cultured for 3 days (the shells stay and the larvae were cultured with the shells, we name this group as 3M). For the control group, we also detected the larvae cultured for 5 days (5M) and 7 days (7M).
CMNV detection was the same as described in section 2.1. To determine the viral load, a standard curve was generated by plotting a graph between different concentrations (in the range of 100–108 copy numbers) of purified CMNV plasmids (provided as the positive control in the kit) [11].
3. Results
3.1. No Treatment (the Control Group)
After hatching, CMNV was detected positive in both the shells (S) and the newly hatched larvae (L) samples, but the viral load of the larvae is only 0.03 copy number/ng RNA, which is 13.7 times lower than that of the shells (Figure 1 and supplementary Table S1). This result indicates that the viruses originally attached on the shells only, and then transmitted to the larvae during hatching. The viral load of 3 day-larvae (3M) is much higher than that of newly hatched larvae, suggesting that the viruses were reproducing. CMNV was also detected on 5 day-larvae (5M) and 7 day-larvae (7M), although the viral load decreased (supplementary Table S1).
3.2. Decapsulation with Na2CO3-NaClO
The result of decapsulating with Na2CO3-NaClO for 5 minutes was similar to that of control group. The CMNV was still detected positive on the shells (S), in newly hatched larvae (L) and 3 day-larvae (3M) samples, but with a reduction in viral load (Figure 1 and supplementary Table S1). When treated for 15 min, the shells were CMNV+, but CMNV was not detected on the newly hatched larvae, indicating that the treatment might block the virus’ transmission from shells to larvae during hatching. But 3 day-larvae (3M) were CMNV+, indicating that the CMNV+ shells are still capable of contaminating the larvae. When treated for 30 min, newly hatched larvae and 3 day-larvae (3M) were both CMNV-, although shells were still CMNV+. When treated for 45 min, the shells, newly hatched larvae and 3 day-larvae were all CMNV- (Figure 1 and supplementary Table S1).
Besides, the hatching rates of Na2CO3-NaClO treating groups and the control group were all above 94% (supplementary Table S1), suggesting that the decapsulating procedure doesn’t affect the hatching rate. Moreover, the decapsulating may shorten the hatching time, when decapsulating for 5 min, the hatching time decreased from 19 h to 17 h; when the treatments ≥ 15 min, the hatching time reduced to 12 h (supplementary Table S1).
3.3. Adding Formalin in the Hatching Water
When 10 ppm formalin was applied, the newly hatched larvae (L) were CMNV-, but the shells (S) and 3 day-larvae (3M) were CMNV+. When formalin concentrations ≥ 30 ppm, the newly hatched larvae and 3 day-larvae (3M) were both CMNV- although the shells were still CMNV+ (Figure 2 and supplementary Table S1), demonstrating that the treatments can disinfect the viruses on the shells and block their transmission to the larvae. However, these treatments decreased the hatching rate (Figure 3 and supplementary Table S1). When 60 ppm formalin was applied, the hatching rate decreased remarkedly from 94% to 40%.
3.4. Combination of Decapsulation and Adding Formalin in the Hatching Water
For the treatment of 5 min–decapsulation followed by adding 5 ppm formalin in the hatching water, the newly hatched larvae and 3 day-larvae were both CMNV- although the shells were CMNV+ (Figure 4 and supplementary Table S1). For the treatment of 15 min + 10 ppm formalin, the shells, the newly hatched larvae and 3 day-larvae are all CMNV-. Besides, this treatment shortened the hatching time to 12 h and did not affect the hatching rate, and the larvae developed faster than the control group (Figure 5).
4. Discussion
Covert mortality nodavirus (CMNV) has a wide range of hosts, multiple vectors and various modes of transmission, leading to long-term widespread of the virus in crustacean culture areas [7]. In shrimp ponds, in addition to cultured shrimp, CMNV was also detected positive in the coexisting species Perinereis aibuhitensis and Artemia sinica, which are the important fresh food for shrimp culture [6]. Yang et al. (2023) recommended disinfection of P. aibuhitensis eggs with peroxymonosulfate (15 ppm for 1 min) to eliminate CMNV [12]. In this work, we also detected the presence of CMNV in the commercial Artemia cysts. Furthermore, we demonstrated that the CMNV probably attached on the cyst shells, and then contaminated the larvae during hatching period as the viral load of shells are 13.7 times higher than that of newly hatched larvae.
To disinfect the CMNV on the shell of Artemia cysts, the easiest way definitely is decapsulation, which also bring the advantages of direct ingestion and digestion o f decapsulated cysts by shrimp larvae [9]. In this study, we also added formalin in the hatching water to assure the elimination of CMNV. We found that the best combination is 15 min–decapsulation process plus 10 ppm formalin treatment in the hatching water, by which not only effectively eliminated the CMNV, but also shortened the hatching time, and the treated larvae developed faster than the control (i.e., larvae hatched from untreated CMNV+ cysts).
Artemia are highly nutritious, their dormant eggs (the cysts) are easy to preserve, and the hatching process is simple, making them widely used in aquaculture breeding, especially for shrimp breeding [13]. However, Artemia and shrimp both belong to crustacean so that naturally there is a risk of Artemia spreading shrimp pathogens [14]. Previous research showed that Artemia can be infected by pathogenic bacteria commonly found in aquaculture, such as Vibrio parahaemolyticus, V. campbellii, and V. harveyi, and pose a risk of spreading acute hepatopancreatic necrosis disease (AHPND) [15]. Artemia may also be contaminated by Enterocytozoon hepatopenaei (EHP) [16], a highly contagious microsporidian parasite in shrimp farming. At present, it has not been confirmed that Artemia are susceptible to common shrimp viruses [14]. In this study, we found that the CMNV+ Artemia cysts of control group can also develop normally (although slower) into larvae, and then grew up to adult with no significant sign of viral infection. However, the risk of Artemia serving as vectors to transmit shrimp pathogens cannot be overlooked. Besides CMNV, Artemia may also be the vectors of multiple shrimp viruses, e.g. white spot syndrome virus (WSSV) [17], Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) [18]. Therefore, utilization of Artemia from unknown sources must be cautious, especially in the culture of specific-pathogen-free (SPF) shrimps.
Currently, the specific-pathogen-free (SPF) technology has been widely used in livestock and aquaculture breeding. It is also because of the establishment of the SPF breeding technology that the white leg shrimp (Penaeus vannamei) has become the world's most widely cultivated and highest-yielding aquaculture species [19]. During the breeding and culture of SPF shrimps, we detected the commercial Artemia cysts for the presence of common specific pathogens, including CMNV, AHPND, EHP, WSSV, MrNV, XSV, DIV1 (decapod iridescent virus 1) and IHHNV (infectious hypodermal and hematopoietic necrosis virus), and we found the Artemia cysts are CMNV-positive in this work. Therefore, we developed this hatching protocol to eliminate CMNV to ensure the biosecurity of producing SPF shrimps. It is yet to know that the disinfection protocol and hatching method for Artemia cysts described in this study could eliminate other common pathogens, and therefore would confer a much broader application prospects in aquaculture industry.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org
References
- Xu, Z.; Ji, F. Comprehensive control of the covert mortality disease of Pacific white shrimp. Fish Guide to be Rich 2009, 1, 60. [Google Scholar]
- Zhang, Q.; Liu, Q.; Liu, S.; Yang, H.; Liu, S.; Zhu, L.; Yang, B.; Jin, J.; Ding, L.; Wang, X. A new nodavirus is associated with covert mortality disease of shrimp. Journal of General Virology 2014, 95, 2700–2709. [Google Scholar] [CrossRef]
- Wang, C.S. Chapter 38 - Nodaviruses of crustaceans. In Aquaculture Virology (Second Edition), Kibenge, F.S.B., Godoy, M.G., Eds.; Academic Press: 2024; pp. 621–641.
- Zhang, Q.; Xu, T.; Wan, X.; Liu, S.; Wang, X.; Li, X.; Dong, X.; Yang, B.; Huang, J. Prevalence and distribution of covert mortality nodavirus (CMNV) in cultured crustacean. Virus Research 2017, 233, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wang, C.; Yao, L.; Wang, W.; Zhao, W.; Jia, T.; Yu, X.; Yang, G.; Zhang, Q. Investigation on natural infection of covert mortality nodavirus in farmed giant freshwater prawn (Macrobrachium rosenbergii). Animals 2022, 12, 1370. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.X.; Wan, X.Y.; Xia, J.T.; Yao, L.; Xu, R.D.; Wang, W.; Yu, X.T.; Zhang, Q.L. Investigation of the prevalence of covert mortality nodavirus (CMNV) in shrimp from 2021 to 2022. Progress in Fishery Sciences 2024, 45, 195–203. [Google Scholar]
- Liu, S.; Wang, X.; Xu, T.; Li, X.; Du, L.; Zhang, Q. Vectors and reservoir hosts of covert mortality nodavirus (CMNV) in shrimp ponds. Journal of Invertebrate Pathology 2018, 154, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Lavens, P.; Sorgeloos, P. The history, present status and prospects of the availability of Artemia cysts for aquaculture. Aquaculture 2000, 181, 397–403. [Google Scholar] [CrossRef]
- Sorgeloos, P.; Bossuyt, E.; Laviña, E.; Baeza-Mesa, M.; Persoone, G. Decapsulation of Artemia cysts: a simple technique for the improvement of the use of brine shrimp in aquaculture. Aquaculture 1977, 12, 311–315. [Google Scholar] [CrossRef]
- Xu, T.; Liu, S.; Li, X.; Zhang, Q. Genomic characterization of covert mortality nodavirus from farming shrimp: Evidence for a new species within the family Nodaviridae. Virus research 2020, 286, 198092. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, S.; Yang, H.; Zhu, L.; Wan, X.; Li, X.; Huang, J. Reverse transcription loop-mediated isothermal amplification for rapid and quantitative assay of covert mortality nodavirus in shrimp. Journal of invertebrate pathology 2017, 150, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zeng, C.; Xia, J.; Liu, Q.; Fang, J.; Zhang, Q. Disinfection of Perinereis aibuhitensis eggs with peroxymonosulfate to eliminate covert mortality nodavirus (CMNV). Aquaculture 2023, 572, 739539. [Google Scholar] [CrossRef]
- Millamena, O.M.; Bombeo, R.F.; Jumalon, N.A.; Simpson, K. Effects of various diets on the nutritional value of Artemia sp. as food for the prawn Penaeus monodon. Marine Biology 1988, 98, 217–221. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, C.; Sui, L.; Ye, S.; Dong, X.; Huang, J. Research progress on the risk of Artemia acting as a kind of live feed to spread shrimp pathogens. China Anim Health Inspect 2020, 37, 61–67. [Google Scholar]
- Defoirdt, T.; Crab, R.; Wood, T.K.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Quorum sensing-disrupting brominated furanones protect the gnotobiotic brine shrimp Artemia franciscana from pathogenic Vibrio harveyi, Vibrio campbellii, and Vibrio parahaemolyticus isolates. Applied and environmental microbiology 2006, 72, 6419–6423. [Google Scholar] [CrossRef]
- Karthikeyan, K.; Sudhakaran, R. Exploring the potentiality of Artemia salina to act as a reservoir for microsporidian Enterocytozoon hepatopenaei of penaeid shrimp. Biocatalysis and agricultural biotechnology 2020, 25, 101607. [Google Scholar] [CrossRef]
- Zhang, J.-S.; Dong, S.-L.; Dong, Y.-W.; Tian, X.-L.; Cao, Y.-C.; Li, Z.-J.; Yan, D.-C. Assessment of the role of brine shrimp Artemia in white spot syndrome virus (WSSV) transmission. Veterinary research communications 2010, 34, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Sudhakaran, R.; Yoganandhan, K.; Ahmed, V.I.; Hameed, A.S. Artemia as a possible vector for Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus transmission (XSV) to Macrobrachium rosenbergii post-larvae. Diseases of aquatic organisms 2006, 70, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Liang, Y.; Huang, J. Global Biosecurity Strategy and Enlightenments from Worldwide Practice. Engineering Science in China 2016, 18, 110–114. [Google Scholar]
Figure 1.
The disinfection efficacy of decapsulation with Na2CO3-NaClO at various durations. S: the shells picked out after hatching; L: the newly hatched larvae; 3M: 3 day-larvae.
Figure 1.
The disinfection efficacy of decapsulation with Na2CO3-NaClO at various durations. S: the shells picked out after hatching; L: the newly hatched larvae; 3M: 3 day-larvae.
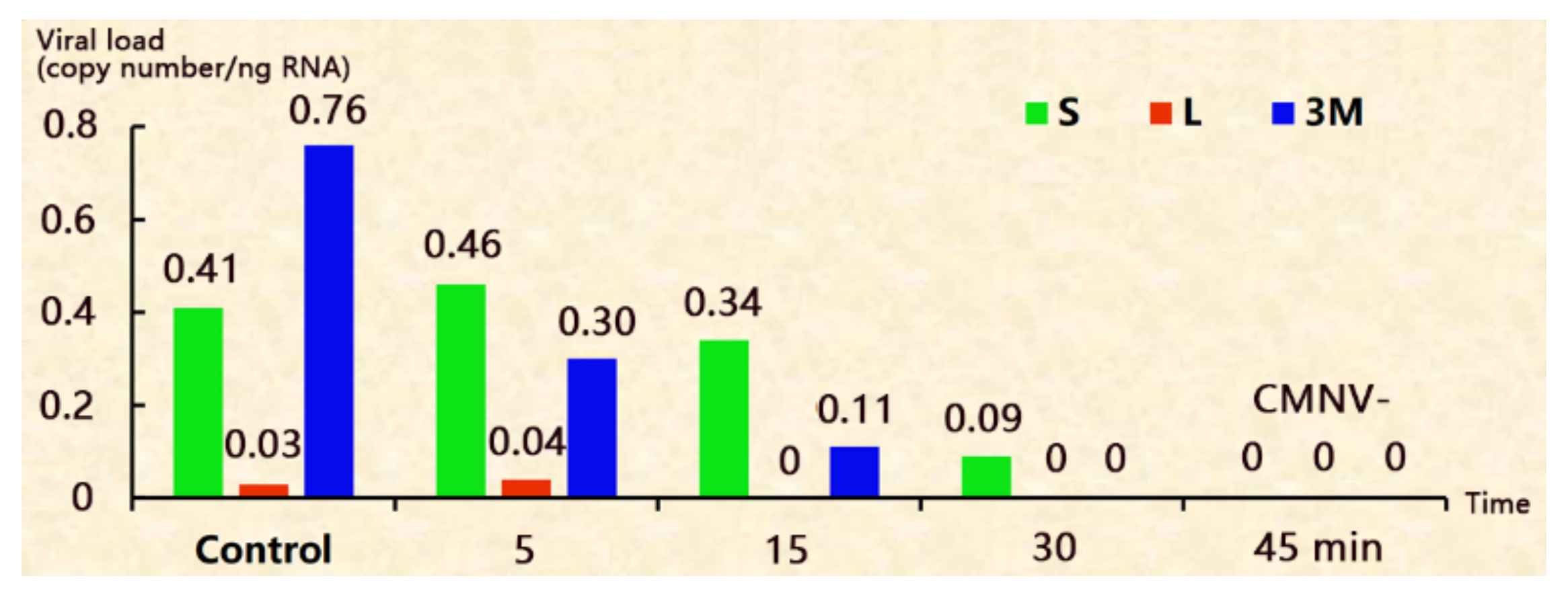
Figure 2.
The disinfection efficacy of adding formalin in the hatching water. S: the shells picked out after hatching; L: the newly hatched larvae; 3M: 3 day-larvae.
Figure 2.
The disinfection efficacy of adding formalin in the hatching water. S: the shells picked out after hatching; L: the newly hatched larvae; 3M: 3 day-larvae.
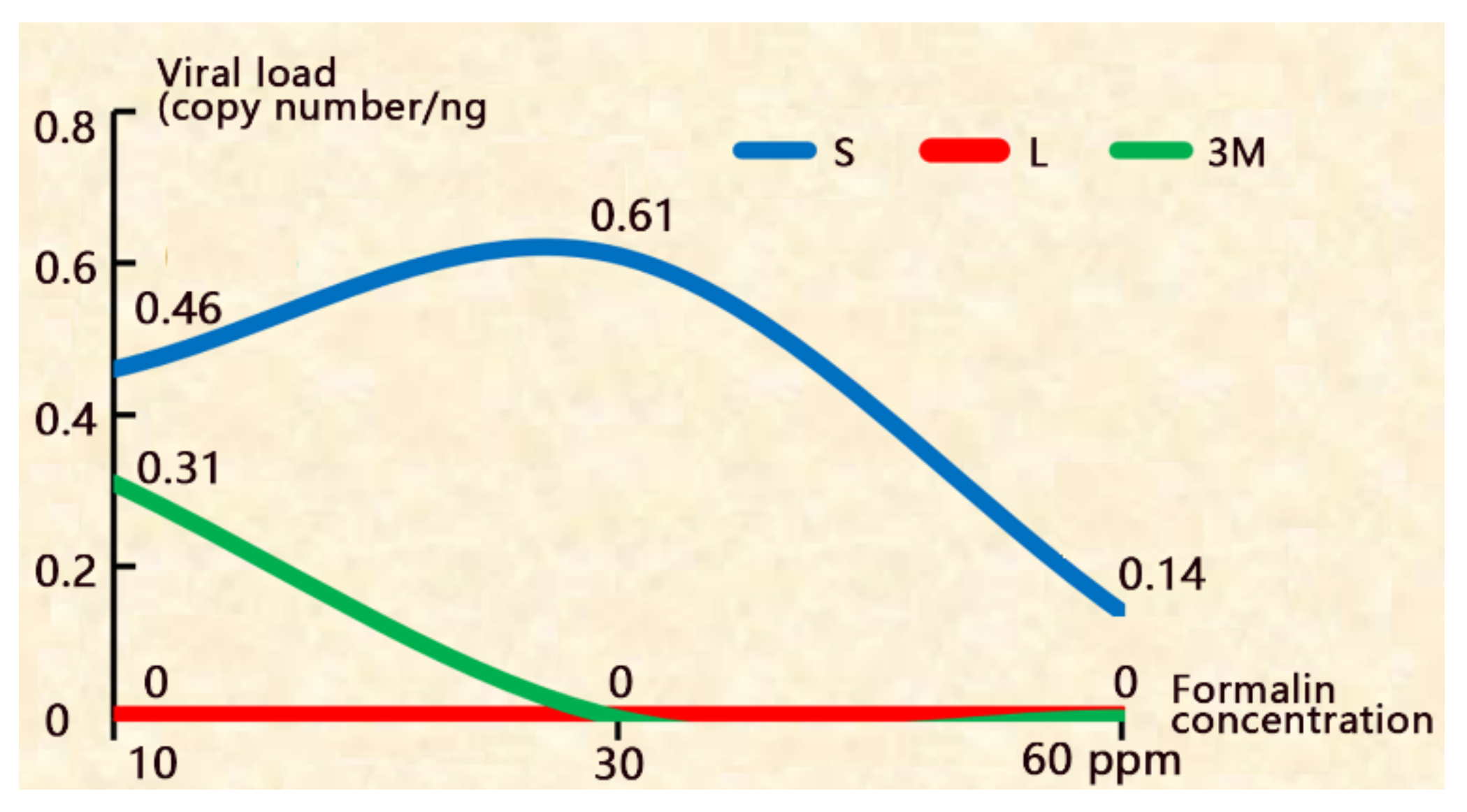
Figure 3.
The hatching rate decreased with the increase of formalin concentration.
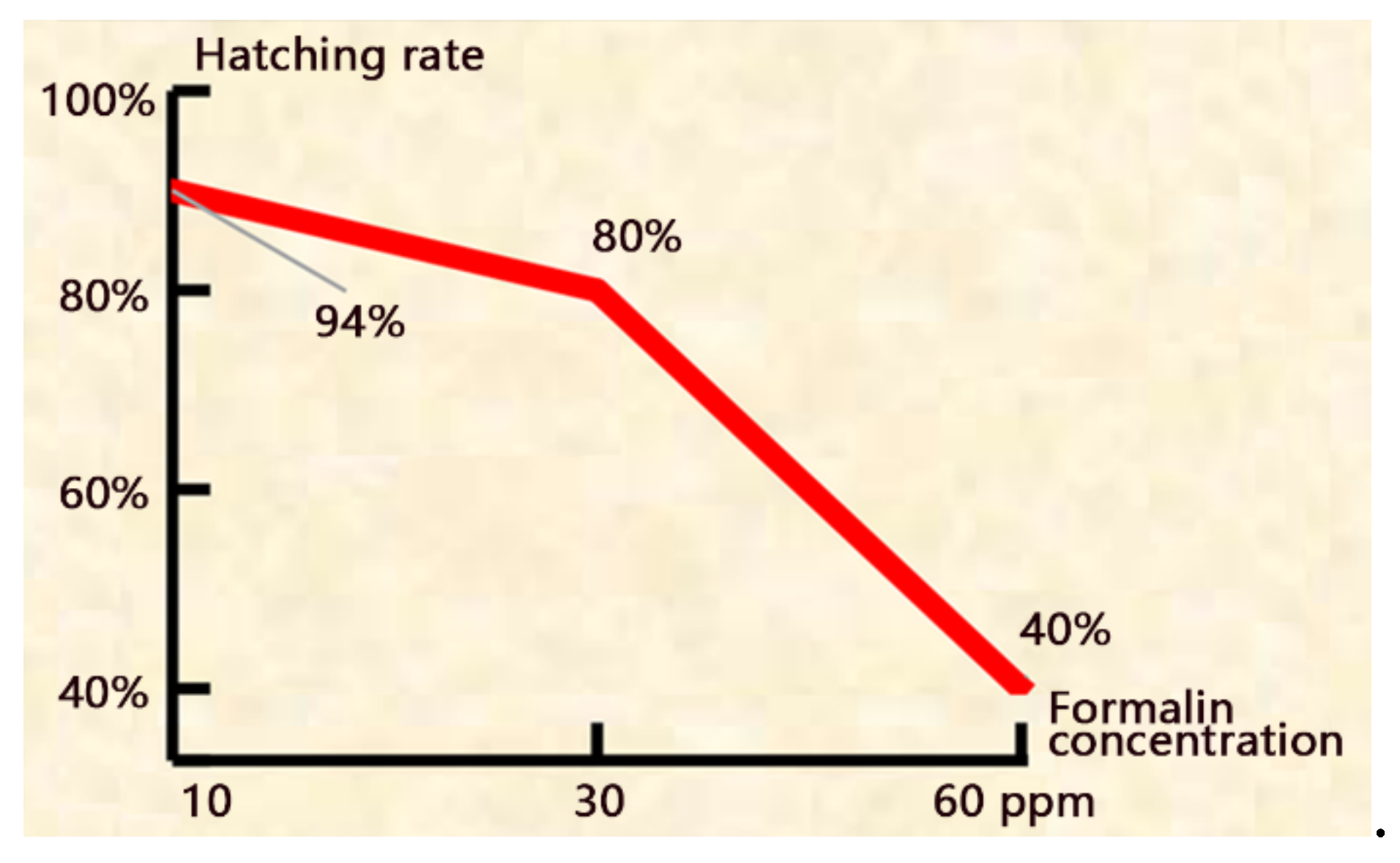
Figure 4.
The disinfection efficacy of combinations of decapsulation and adding formalin in the hatching water. S: the shells picked out after hatching; L: the newly hatched larvae; 3M: 3 day-larvae.
Figure 4.
The disinfection efficacy of combinations of decapsulation and adding formalin in the hatching water. S: the shells picked out after hatching; L: the newly hatched larvae; 3M: 3 day-larvae.
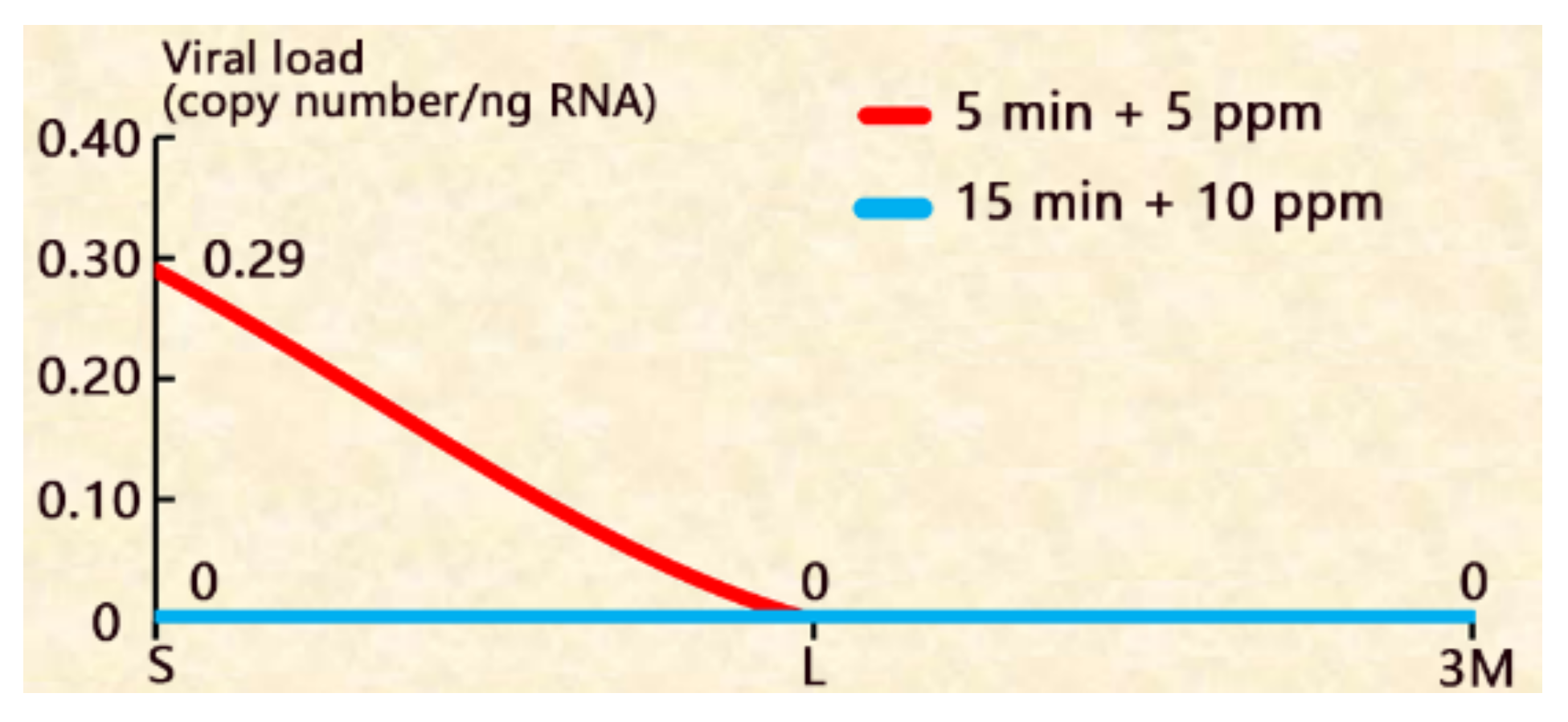
Figure 5.
Three day-larvae of control (A) and 15 min + 10 ppm treatment (B), the latter developed faster.
Figure 5.
Three day-larvae of control (A) and 15 min + 10 ppm treatment (B), the latter developed faster.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



