
Submitted:
14 July 2024
Posted:
16 July 2024
You are already at the latest version
Abstract
A mass mortality event of Asian seabass Lates calcarifer juveniles occurred in Zhuhai, the main Asian seabass cultured area in mainland China. The fish samples were pooled for pathogen identification and both high viral loads of ISKNV and NNV were detected by real-time microfluidic quantitative PCR. Immunohistochemistry and immunofluorescence showed that strong ISKNV signals were examined in spleen and liver, while strong NNV signals were detected in brain and eye. The multi-tissue homogenates were inoculated into MFF-1 cell and SSN-1 cell, respectively. After several viral passages, both ISKNV and NNV were purely isolated from each other, and designated as ASB-ISKNV-23 and ASB-NNV-23, respectively. The whole genome sequences of ASB-ISKNV-23 and ASB-NNV-23 were determined and annotated. The result showed that ASB-ISKNV-23 and ASB-NNV-23 are composed of 112,236 bp and 4,538 bp, respectively. Phylogeny analysis showed that ASB-ISKNV-23 belongs to ISKNV-II sub-genotype and ASB-NNV-23 belongs to RGNNV genotype. Collectedly, coinfection of ISKNV-II and RGNNV were firstly documented in mass mortality of Asian seabass in mainland China. Importantly, both ISKNV-II and RGNNV were purely isolated using two different permissive cell lines. Our study provides useful information for better understanding the complex pathogenesis regarding the coinfection with ISKNV and NNV in farmed fish.
Keywords:
Infectious spleen and kidney necrosis virus genotype II (ISKNV-II)
; Red grouper type nervous necrosis virus (RGNNV)
; Asian sea bass Lates calcarifer
; Co-infection
; Mass mortality
1. Introduction
Asian seabass (Lates calcarifer), also known as barramundi, is one of the most important aquaculture species in several Sout-east (SE) Asian countries. Because of its fast growth and highly economic value, the fish is widely cultured, however, as the scale of farming grows, disease problems are increasing and causing huge economic losses to these farming countries [1]. Lymphocystis virus (LCDV), infectious spleen and kidney necrosis virus (ISKNV), red sea bream iridovirus (RSIV) and scale drop disease virus (SDDV) of the Iridoviridae family and nervous necrosis virus (NNV) of the Nordaviridae family were the most commonly concerned viral pathogens, causing high mortalities in Asian seabass production [2,3,4,5,6]. ISKNV and NNV have been prevalent in Chinese fish farms for decades, and both of them are important pathogens associated the most widespread and serious viral infectious diseases in China’s juvenile shellfish farming sector [7,8]. ISKNV, the type species in genus Megalocytivirus, is a large dsDNA viruses, and the viral particle is icosahedral symmetry with a diameter about 150 nm [9,10]. Nowadays, ISKNV has been classified into three genotypes (ISKNV, RSIV and TRBIV) and further six sub-genotypes (ISKNV-I, -II, RSIV-I, -II and TRBIV-I, -II) based on the conserved major capsid protein (mcp) gene [4,11,12]. In ISKNV genotype, ISKNV-I was prevalent in Thailand and ISKNV-II was identified in Vietnam [2,13,14]. In mainland China, we recently reported that the infection with ISKNV-II causing severe mass mortalities of Asian seabass juveniles [15], and an effective inactivated ISKNV-SDDV bivalent vaccine against ISKNV-I, ISKNV-II, RSIV-II and SDDV was developed in Asian seabass infection model [16]. NNV is a small, spherical, and non-enveloped RNA virus with a dual segments of single stranded (+) RNA genome. NNV mainly contained four main genotypes, namely RGNNV, BFNNV, TPNNV, and SJNNV [17]. Outbreaks of NNV-associated diseases have been reported to occur mainly in larvae and the early juvenile fry stages [18]. Diseased fish often exhibit abnormal swimming behavior, loss of appetite, and dark body. Histopathological analysis revealed extensive necrosis of the central nervous system (CNS) with extensive vacuolation in brain and neurodegeneration, accompanied by retinal vacuolation [19,20]. RGNNV has been reported to be the causative agent of mass mortality of Asian seabass larvae or fry [6,21].
Co-infection means that the host is infected with two or more different pathogens at the same case. It has been reported that in aquatic animals, the mortality rate of co-infection was generally higher than that of a single infection [22]. The coinfections of different virus have been documented in some cultured fish, such as co-infection of grouper iridovirus in genus Ranavirus (GIV-R) and RGNNV in juvenile orange-spotted grouper [23], co-infection of largemouth bass virus and rhabdovirus from moribund largemouth bass (Micropterus salmoides) [24]. The co-infection of ISKNV and NNV was reported to cause ~50% mortality in cultured Asian seabass in Thailand [25]. The co-infection of two different viruses involves very complex infection mechanisms. Specifically, it is always not easy to completely isolate the individual virus at cell culture level due to the lack of differentiated in vitro proliferation systems [23,24]. In this work, a very severe mass mortality of Asian seabass juveniles was documented in Zhuhai, South China, and about 100 thousand Asian seabass died out within one week. Pathogen isolation and identification showed that both high viral loads of ISKNV and NNV were detected in infected fish tissues. Importantly, ISKNV-II and RGNNV were purely cultured from the same sample fish using two different permissive cell lines. As far as we know, it is the first time to isolate and characterize both ISKNV and NNV in co-infected Asian seabass in mainland China.
2. Materials and Methods
2.1. Fish Sampling and Pathogen Detection
The mass mortality event occurred in April, 2023. About 100, 000 Asian seabass juveniles with body length about 2-4 cm were transported from a fish farm in Yangjiang to a fish farm in Zhuhai (about 220 km between two fish farms) for a commercial breeding. Three days after the transfer of Asian seabass juveniles to the fish farm in Zhuhai, disease occurred. The clinical symptoms of diseased fish included anorexia and drowsiness, accompanied by body blackening, lying down, or spinning in pond (Figure 1A1 and A2). In the next 2-5 days, a large-scale death occurred, with almost all fish dying within one week. The farmer had to give up his breeding plan and removed the small amounts of residual fish. The diseased fish samples were sent to our laboratory for pathogen identification. The visceral organs including liver, spleen, kidney together with eyes and brain from 3 fish were sampled to be homogenized in sterile PBS with a 1: 9 ratio (w/v). After centrifugation at 5,000 x g for 5 minutes, 100 µL of supernatant was added to a reaction well of a microfluidic quantitative PCR testing equipment (Helixgene Biotechnology Co., LTD, China) for rapid viral pathogen detection. For gene sequence cloning, DNA/RNA extraction was performed by using VAMNE MagUltra FFPE DNA Extraction Kit (Vazyme, China) and Simply RNA Tissue Kit (Promega, USA), and cDNA was prepared by using AccurSTART Reverse Transcriptase (Vazyme, China). PCR was performed with a universal primer set of ISKNV/RSIV major capsid protein gene (mcp) , mcp-F: 5′ -TCA TTG TCA TCA TCA TGT CTG C-3′ and mcp-R: 5′ -AGA CAC ACG GGG CAA TC-3′ [26], and a primer set of VNNV coat protein gene (cp), NNVcp-F: 5'-ATG GTA CGC AAA GGT GAG AA-3', NNVcp-R: 5'-TTA GTT TTC CGA GTC AAC CC-3' [7]. And then the PCR products were detected by agarose gel electrophoresis.
2.2. Immunohistochemistry (IHC) and Immunofluorescence Assay (IFA)
The partial liver, spleen, eye and brain tissues were sampled and fixed in 4% paraformaldehyde (PFA) for immunohistochemistry (IHC) and immunofluorescence assays. Immunohistochemistry assay was performed by using Super Sensitive IHC Detection System Kit (Mouse/Rabbit). Mouse monoclonal antibody (mAb) against ISKNV-VP23 [15] and rabbit anti-NNV-CP polyclonal antibody (pAb) [27] were used as primary antibodies and HRP-labelled goat anti mouse/rabbit IgG was used as the secondary antibodies. Then DAB staining was performed according to instructions.
For IFA, the ISKNV-VP23 mAb and NNV-CP pAb were used as primary antibodies, AlexaFluor555-conjugated (red fluorescence) goat anti-rabbit IgG, AlexaFluor488-conjugated (green fluorescence) goat anti-rabbit IgG and AlexaFluor488-conjugated (green fluorescence) goat anti-mouse IgG were used as secondary antibodies. The nucleus was stained by 4′ ,6-diamidino-2-phenylindole (DAPI) (Abcam, China). Sections were visualized under a confocal laser scanning immunofluorescence microscopy (Leica TCS SP8 STEP 3×, German).
2.3. Virus Isolation, Concentration, Whole Genome Determination and Annotation
Mandarinfish fry (MFF-1) cell line which is highly sensitive to ISKNV/RSIV/SDDV was cultured in ambient air with 5% CO2 at 26 ℃ within Dulbecco Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) (Gibco Invitrogen) [28]. SSN-1 cells [29] were cultured at 26 °C within L15 medium supplemented with 10% FBS. When MFF-1 cells and SSN-1 cells were confluent in tissue dishes with a diameter of 10-cm, 200 μL of multi-tissue (including liver, spleen, brain and eye) homogenate, or liver and spleen homogenate being filtered through 0.22 μm syringe-driven filter unit (Millipore, Merk) was added to MFF-1 cells, and 200 μL of filtered eye and brain homogenate was added to SSN-1 cells. The inoculated cells were observed daily under an inverted microscope. When cells exhibited apparent cytopathic effect (CPE), the infected cells were harvested into -80 ℃ for about 12 hours and thawed at room temperature. The yielded virus after the initial inoculation was labeled as viral passage 1. After three cycles of frozen/thawed treatment, the suspension was inoculated into fresh confluent MFF-1 or SSN-1 for another round of viral passage and the yielded virus was labeled as viral passage 2. In this way, both viruses were passaged at least for three rounds of passage (viral passage 4). IFA was implemented to monitor the infection of ISKNV in MFF-1 cell and NNV SSN-1 cell, respectively.
For viral concentration, the infected cells were frozen/thawed for three times before concentration. Then the suspensions were centrifuged at 8,000× g for 40 min at 4 ℃. The supernatant was then centrifuged at 150,000× g for 1.5 h at 4 ℃. The roughly purified virus was resuspended in sterile PBS for genomic DNA/RNA preparation and library construction, de novo assembly and annotation as our recent description [8]. The genomic RNA was reverse-transcribed into cDNA for genomic sequencing. The whole genome sequencing was conducted by Beijing Novogene Bioinformatics Technology Co., Ltd and Geneplus Clinical Laboratory, Shenzhen, China. The annotation results of viral genome was converted using GB2sequin [30] for submission to the NCBI network service. Genomic map was done by SnapGene 6.0 and the online tool CGView (https://paulstothard.github.io/cgview/) [31].
2.4. Construction of Phylogenetic Tree
The complete sequences of the major capsid protein (mcp) gene of ISKNV and the coat protein (cp) gene of NNV were analyzed by the BLAST program of the NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi) for homology. Two phylogenetic trees of ASB-ISKNV-23 and ASB-NNV-23 isolates in this study were constructed with other 30 isolates from genus Megalocytivirus and 18 isolates from the genus β-Nordavirus, respectively. Sequence alignment was performed by MEGA 6.0 software and phylogenetic tree was constructed by the Maximum Likelihood method.
3. Results
3.1. Pathogen Detection
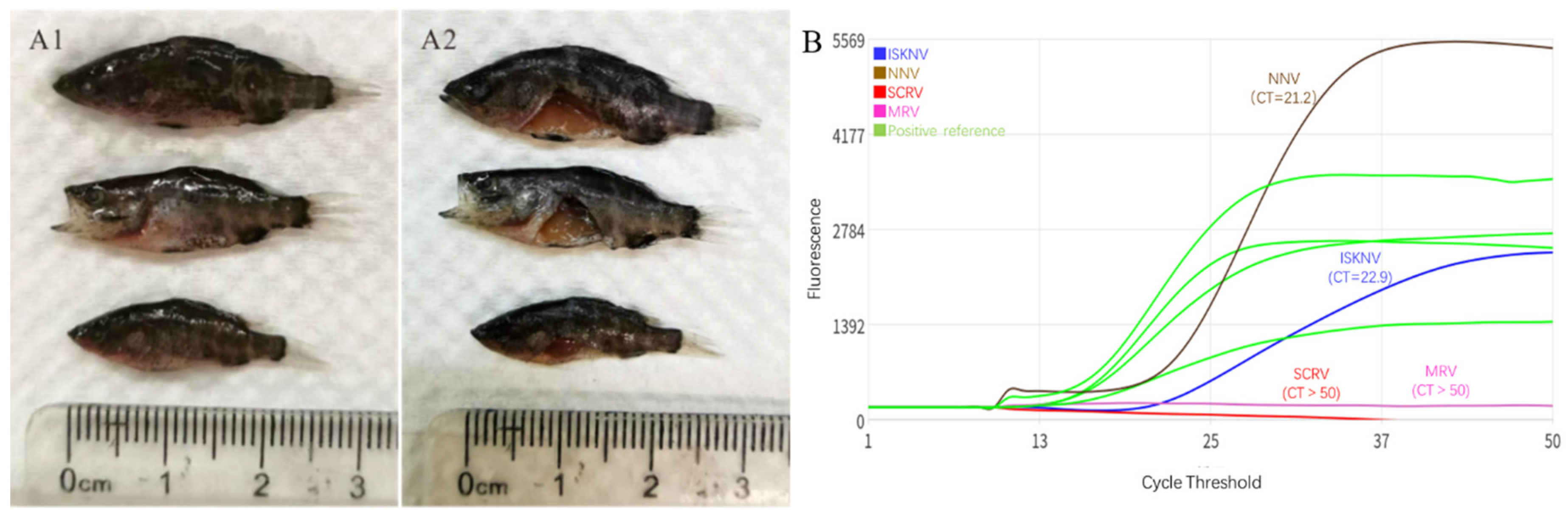
The mixed multi-tissues of liver, spleen, kidney, brain and eye from three diseased fish samples were homogenized for pathogen detection. Four common aquatic viruses i.e. ISKNV, NNV, Siniperca chuatsi rhabdovirus (SCRV) and mandarinfish ranavirus (MRV) were listed as the potential target viruses by using a rapid detection kit of microfluidic real-time quantitative PCR. As a result, the obtained CT values of ISKNV, NNV, SCRV and MRV were 22.9, 21.2,>50 and>50 respectively (Figure 1B), indicating that both high viral loads of ISKNV and NNV were detected from the multi-tissue homogenate. Then, conventional PCR or RT-PCR were performed to detect the mcp gene of ISKNV and the cp gene of NNV. As a result, both positive amplification bands were obtained (Figure 2).
Figure 1.
Clinical samples and pathogen detection of diseased L. calcarifer. A1, three sample fish and their autopsies (A2) with the featured clinical sign of darken body. B, rapid detection of four common viruses by using microfluidic real time quantitative PCR. The CT values of ISKNV, NNV, SCRV and MRV were 22.9 and 21.2, >50 and>50respectively, suggesting high viral loads of ISKNV and NNV in these mixed multi-tissue homogenates.
Figure 1.
Clinical samples and pathogen detection of diseased L. calcarifer. A1, three sample fish and their autopsies (A2) with the featured clinical sign of darken body. B, rapid detection of four common viruses by using microfluidic real time quantitative PCR. The CT values of ISKNV, NNV, SCRV and MRV were 22.9 and 21.2, >50 and>50respectively, suggesting high viral loads of ISKNV and NNV in these mixed multi-tissue homogenates.
3.2. IHC and IFA Assays of the Naturally Diseased L. calcarifer
By IHC, numerous strong ISKNV signals were detected in liver and spleen tissues of diseased L. calcarifer by anti-ISKNV VP23 mAb, indicating the existence of severe ISKNV infection in these tissues (Figure 3E and F). Meanwhile, strong NNV signals were detected in brain and eye tissues by using anti-NNV CP pAb (Figure 3G and H). Besides, cellular vacuolation with nuclear pyknosis and nuclear fragmentation was observed in the brain gray matter, indicating a featured pathological symptom of NNV infection (Figure 3H). By IFA, numerous green fluorescent signals (associated with anti-ISKNV VP23 mAb), were observed in spleen tissue and some in liver tissue. Meanwhile, strong NNV-CP associated green fluorescent signals were observed in brain and eye tissues. Specially, the green fluorescence signals in eye tissue were mainly concentrated in the retina area (Figure 4).
3.3. Isolation of ISKNV and NNV Using Different Permissive Cell Lines
For viral isolation and culture, the filtered multi-tissue homogenate was inoculated into MFF-1 cells. As a result, slight cytopathic effect (CPE) was observed within 3 days after the initial infection, and advanced CPE was observed at 7 dpi (data not shown). To identify whether co-infection of ISKNV and NNV occurred in MFF-1 cell, double immunofluorescence assay assay stained with mouse anti-ISKNV-VP23 mAb and rabbit anti-NNV-CP pAb were performed. As a result, both green fluorescence (associated with ISKNV-VP23) and red fluorescence (associated with NNV-CP) were observed in infected MFF-1 cells (Figure 6A), indicating the presence of active ISKNV and NNV in infected MFF-1 cell after the initial inoclution. To isolate the individual ISKNV and NNV, the filtered homogenate from liver and spleen tissues, and the filtered homogenate from brain and eye tissues were inoculated into MFF-1 cell and SSN-1 cell, respectively. As a result, both CPEs appeared (Figure 5).
3.4. Whole Genome Determination and Phylogenetic Tree Construction
After 2 rounds of viral passages, the propagations of ISKNV in MFF-1 and NNV in SSN-1 showed stably. Double immunofluorescence assay showed that ISKNV-associated strong green fluorescence signals were only observed in infected MFF-1 cells (Figure 6B) and NNV associated strong red fluorescence signals were only observed in infected SSN-1 cells (Figure 6C), indicating that the individual ISKNV and NNV were isolated from each other in MFF-1 cell and in SSN-1 cell, and designated as ASB-ISKNV-23 and ASB-NNV-23, respectively. The whole genome determinations showed that the full length of ASB-ISKNV-23 genome (accession No. PP151097.1) is 112,236 bp, with GC content of 55% and encodes 120 open reading frames (ORFs) (Figure 7), and the full length of ASB-NNV-23 genome was 4,441 bp, with GC content of 53%. The ASB-NNV-23 genome contained two RNA strands, RNA1 (accession no. PP214961.1) with a length of 3,054 bp encoding the RNA polymerase, the RNA2 (accession no. PP214962.1) with a length of 1,387 bp encoding the capsid protein (Figure 8)
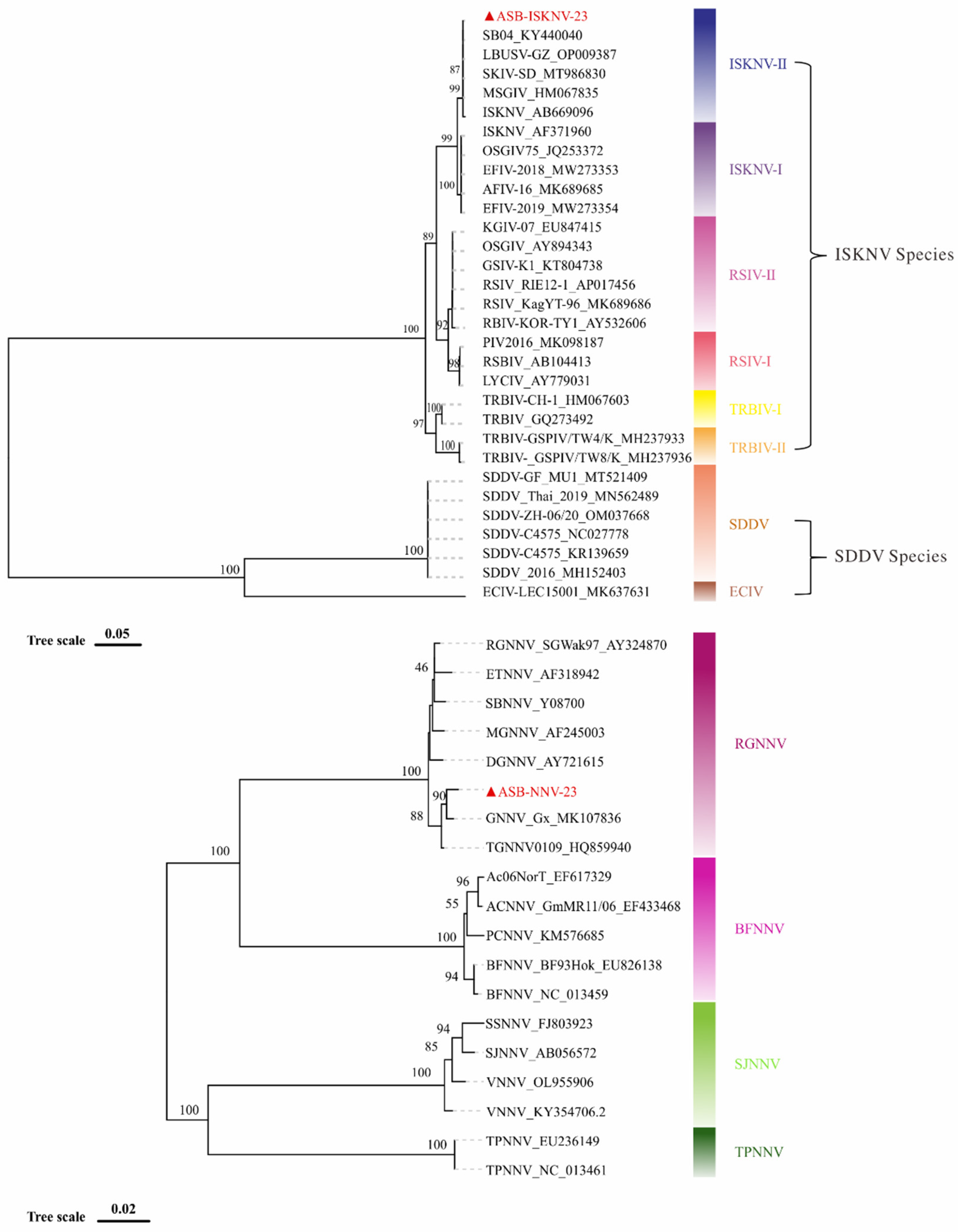
Based on the mcp gene, phylogenetic analysis showed that ASB-ISKNV-23 belongs to ISKNV-II sub-genotype of ISKNV species in genus Megalocytivirus (Figure 9 upper). The complete mcp gene sequence of ASB-ISKNV-23 is 1362 bp, which shares 99.93% similarity with that of a largemouth bass original ISKNV isolate of LBUSV-GZ (Accession no. PP009387.1). The sequence alignment showed the only difference between these two sequences was in 1203rd base, where A mutates into G (data not shown). The phylogenetic analysis based on the capsid protein gene (cp) showed that ASB-NNV-23 was clustered into the clade of RGNNV genotype of Nordavirus (Figure 9 down). The full length of cp gene of ASB-NNV-23 is 1,017 bp, sharing 99.31% similarity with that of a hybrid grouper isolate of GNNV-Gx (Accession no.MK107836.1).
4. Discussion
Several mass mortality events of juvenile Asian seabass have been documented in Zhuhai, the most popular breeding area of commercial Asian seabass in mainland China. In these events, ISKNV sub-genotype II (ISKNV-II) was evidenced and characterized as the causative agent [15]. In this study, the diseased fish samples from a severe mass mortality of Asian seabass juvenile were taken out for pathogen iditenfication and both high viral loads of ISKNV and NNV were confirmed. IHC and IFA showed that strong ISKNV signals were observed in spleen and liver tissues, whereas strong NNV signals were observed in eye and brain tissues. Furthermore, double stained IFA showed that both ISKNV and NNV signals were observed in mixed multi-tissue homogenate (liver, spleen, brain and eyes)-infected MFF-1 cells. Interestingly, in most cases, the green fluorescence associated with ISKNV and the red fluorescence associated with NNV were not observed on the same cell (Figure 6A), indicating that ISKNV and NNV may interfere with each other at cell culture level. MFF-1 cell line is highly susceptible for megalocytiviruses i.e. ISKNV [28], RSIV [23,26] as well as SDDV [11]. After the initial infection of mixed multi-tissue homogenate, although strong NNV signals were also observed, however the NNV signal gradually weakens and eventually disappears with viral passages (data not shown). After two rounds of viral passage, no NNV signal was observed in infected MFF-1 cell, suggesting that MFF-1 cell is not permissive for NNV replication, which was consistent with a previous report [32]. On the other hand, NNV was evidenced to be able to attach but not to penetrate even non-permissive human cell lines [33]. Thus, it is not unexpected that NNV signal was observed in non-permissive MFF-1 cell after the initial inoculation and then disappeared after viral passage. Both IHC and IFA showed that ISKNV was distributed highly in spleen and liver (Figure 3 and Figure 4), whereas NNV was highly concentrated in brain and eye. To effectively isolate the individual ISKNV and NNV, the liver and spleen homogenate, and the brain and eye homogenate were inoculated into MFF-1 cell and SSN-1 cell, respectively. As two rounds of viral passages, ISKNV and NNV were isolated, respectively. SSN-1 is a highly susceptible cell line for NNV [29] and no data showed that SSN-1 supported effective replication of ISKNV/RSIV. Our study showed that using brain and eye homogenate to inoculate SSN-1 cells, advanced CPE was observed at 2 dpi (Figure 4). Double stained IFA showed that only NNV signals were observed in infected SSN-1 cells. To conclude, using two different permissive cell line, ISKNV and NNV were individually isolated from the same case of diseased Asian sea bass sample. Co-infection of ISKNV and NNV has ever been documented in diseased Asian sea bass in Thailand [25]. In Jitrakorn’s report, yellow-striped grunt (HaemuLon flavoliireatum) fin original GF cell, the first marine fish cell line in the world [34], was used to culture both viruses. As a result, both ISKNV and NNV grew well in the same GF cell system, however the individual virus was not purely isolated from each other. The detailed mechanism of coinfection of ISKNV and NNV in GF cell remained unclear [25].
As reported, in Atlantic salmon (Salmo salar), infectious salmon anemia (ISA) may be avirulent in the co-infections of togavirus-like viruses and ISA [35]. On the contrary, co-infection with snakehead retrovirus (SnRV) and grouper nerve necrosis virus (GNNV) could increase the infection of GNNV [36]. Co-infections were common in different type pathogens, which were lot of evidences of virus-bacterial or virus-parasitic co-infections [37,38,39,40]. Compared with single-infection, co-infection may lead to worsened clinical symptoms and higher mortality rate. Previous study also showed that the concurrence of Sparus aurata papillomavirus 1 (SaPV1) and/or S. aurata polyomavirus 1 (SaPV1) could be frequently detected in LCDV-affected gilthead sea bream (S. aurata), might reflecting an opportunistic increased capacity of these viruses to replicate in the presence of an LCDV infection [41]. However, the possible role of SaPyV1 and SaPV1 in lymphocystis disease development still remain largely unknown.
Largely different from other coinfection, in this study, the featured histopathology triggered by ISKNV and NNV could be observed clearly in individual target tissues, although the interaction and infection dynamics of these two pathogens remain unclear. The high viral loads of both ISKNV and NNV in infected tissues together with the featured individual histopathology indicated that both ISKNV and NNV were co-causative agents for this mass mortality event. It is noting that the resultant conclusion of this study is not the norm in production practice. In practice, NNV disease and iridovirus disease usually occur in staggered peaks. For an instance of grouper breeding, the replication of NNV always follows the embryo development of grouper once it appears, due to its vertical transmission characteristics, whereas outbreak of iridovirus occurs just in a certain period of rearing [42]. A molecular epidemiology of ISKNV/RSIV and NNV in diseased spotted sea bass Lateolabrax maculatus also showed that both ISKNV/RSIV and NNV could be detected easily by real-time quantitative PCR in the same diseased fish sample, however high viral loads of both viruses is just an occasional event, and only the dominant virus can be cultured at cell culture level, indicating that the single virus rather than dual viruses leads to the lethal infection (unpublish data).
Genomic determination showed that the full length of ASB-ISKNV-23 genome is 112,236 bp, with 55% GC content and 120 ORFs (Figure 6). The full length of ASB-NNV-23 genome is 4,441 bp with the GC content of 53%, containing two RNA strands, RNA1 with a length of 3,054 bp encoding the RNA polymerase and the RNA2 with a length of 1,387 bp encoding the capsid protein (Figure 7). Phylogenetic analysis showed ASB-ISKNV-23 belongs to ISKNV-Ⅱ,and ASB-NNV-23 belongs to RGNNV. In mainland China, the earliest ISKNV isolate from freshwater or brackish cultured fish was confirmed as ISKNV-I [11,28]. In Zhuhai, Asian sea bass L. calcarifer, spotted sea bass L. maculatus as well as yellowfin sea bream Acanthopagrus latus are cultured in nearly pure freshwater pond with no or very low salinity (<3‰). However, ISKNV-I becomes rare, while ISKNV-II, RSIV-I as well as RSIV-II are becoming prevalent until recently [8,15]. Fortunately, the inactivated vaccine based on the ISKNV-I isolate confers the same highly efficient immune protection against various sub-genotype isolates including ISKNV-II, RSIV-I, RSIV-II as well as ISKNV-I itself in different vaccination fish models [8,16]. As for NNV, in China, RGNNV has been the prevalent isolate for a long time, and rare other genotype NNV was detected of confirmed [7]. In our team, a total of 98 NNV isolates have been isolated and identified through SSN-1-based cell culture in a recent molecular epidemiology investigation of diseased spotted sea bass L. maculatus in Zhuhai. Although significant sequence mutations were observed among these isolates, all these isolates belong to RGNNV genotype, indicating that RGNNV is still the prevalent domestic isolate in mainland China (data not shown).
5. Conclusion
We are the first to document the co-infection of ISKNV and NNV in a mass mortality of Asian sea bass juvenile in mainland China. The high viral loads of both viruses together with their featured immunohistochemistry and immunofluorescence supported that the coinfection of ISKNV-II and RGNNV inflicted severe damage to target tissues and then resulted in outcome of the mass mortality. Moreover, the individual ISKNV and NNV were purely isolated from each other using two different permissive cell lines and whole genomes of both viruses were also determined and characterized. Taken togather, this finding contributes important data for better understanding the complex pathogenesis regarding the coinfection with ISKNV and NNV in farmed fish.
Author Contributions
Conceptualization, Y.L. and C.D.; methodology, Q.S., S.D., X.L., W.Z., F.Y., Y.F., ; software, D.S.,W.Z., and Q.S. ; validation, C.D. , Y.L. and Q.S..; formal analysis, C.D., D.S., and Q.S.; investigation, Y.F., Y.L, X.L. and Q.S.; resources, Y.L., X.L., C.D., and Q.S.; data curation, C.D., Y.L. and Q.S.; writing—original draft preparation, Q.S., D.S., and C.D.; writing—review and editing, Y.L. and C.D.; visualization, Q.S., and W.Z.; supervision, C.D. and Y.L.; project administration, Q.S., C.D. and Y.L.; funding acquisition, C.D. and Y.L. ; All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai) (SML2023SP236), the key areas R&D Program of Guangdong Province under No. 2021B0202040002; Guangdong Provincial Special Fund for innovative team for research and development of common key technologies for monitoring and comprehensive prevention and control of aquatic diseases [Grant Number 2019KJ141].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The whole genome sequences of ASB-ISKNV-23, ASB-NNV-23-RNA1 and ASB-NNV-23-RNA2 have been deposited into NCBI nucleotide GenBank with accession Nos. PP151097.1, PP214961.1 and PP214962.1, respectively.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Islam MA, Bosu A, Hasan MM, Yasmin F, Khan ABS, Akhter M, et al. Culture technique of seabass, Lates calcarifer in Asia: A review. International Journal of Science and Technology Research Archive. 2023;4(1):006-17. [CrossRef]
- Dong HT, Jitrakorn S, Kayansamruaj P, Pirarat N, Rodkhum C, Rattanarojpong T, et al. Infectious spleen and kidney necrosis disease (ISKND) outbreaks in farmed barramundi (Lates calcarifer) in Vietnam. Fish & Shellfish Immunology. 2017;68:65-73. PubMed PMID: WOS:000411299500008. [CrossRef]
- Pearce M, Humphrey JD, Hyatt AD, Williams LM. Lymphocystis disease in captive barramundi Lates calcarifer. Aust Vet J. 1990;67(4):144-5. Epub 1990/04/01. [CrossRef] [PubMed]
- de Groof A, Guelen L, Deijs M, van der Wal Y, Miyata M, Ng KS, et al. A Novel Virus Causes Scale Drop Disease in Lates calcarifer. PLoS Pathog. 2015;11(8):e1005074. Epub 2015/08/08. [CrossRef] [PubMed] [PubMed Central]
- Girisha SK, Puneeth TG, Nithin MS, Kumar BTN, Ajay SK, Vinay TN, et al. Red sea bream iridovirus disease (RSIVD) outbreak in Asian seabass (Lates calcarifer) cultured in open estuarine cages along the west coast of India: First report. Aquaculture. 2020;520. doi: ARTN 734712.
- PubMed PMID: WOS:000515498200020. [CrossRef]
- Hick P, Schipp G, Bosmans J, Humphrey J, Whittington R. Recurrent outbreaks of viral nervous necrosis in intensively cultured barramundi (Lates calcarifer) due to horizontal transmission of betanodavirus and recommendations for disease control. Aquaculture. 2011;319(1-2):41-52. PubMed PMID: WOS:000294751500008. [CrossRef]
- Ma H, Wen W, Su Y, Feng J, Xu L, Peng C, et al. Epidemiological characterization of VNNV in hatchery-reared and wild marine fish on Hainan Island, China, and experimental infection of golden pompano (Trachinotus ovatus) juveniles. Arch Virol. 2015;160(12):2979-89. [CrossRef] [PubMed]
- Fu W, Li Y, Fu Y, Zhang W, Luo P, Sun Q, et al. The Inactivated ISKNV-I Vaccine Confers Highly Effective Cross-Protection against Epidemic RSIV-I and RSIV-II from Cultured Spotted Sea Bass Lateolabrax maculatus. Microbiol Spectr. 2023;11(3):e0449522. Epub 2023/05/24. [CrossRef] [PubMed] [PubMed Central]
- He JG, Zeng K, Weng SP, Chan SM. Systemic disease caused by an iridovirus-like agent in cultured mandarinfish, Siniperca chuatsi (Basillewsky), in China. J Fish Dis. 2000;23, 219-22.
- He JG, Deng M, Weng SP, Li Z, Zhou SY, Long QX, et al. Complete genome analysis of the mandarin fish infectious spleen and kidney necrosis iridovirus. Virology. 2001;291(1):126-39. [PubMed]
- Fu Y, Li Y, Fu W, Su H, Zhang L, Huang C, et al. Scale Drop Disease Virus Associated Yellowfin Seabream (Acanthopagrus latus) Ascites Diseases, Zhuhai, Guangdong, Southern China: The First Description. Viruses. 2021;13(8). Epub 2021/08/29. [CrossRef] [PubMed] [PubMed Central]
- Koda SA, Subramaniam K, Pouder DB, Yanong RP, Waltzek TB. Phylogenomic characterization of red seabream iridovirus from Florida pompano Trachinotus carolinus maricultured in the Caribbean Sea. Arch Virol. 2019;164(4):1209-12. [CrossRef] [PubMed]
- Kerddee P, Dinh-Hung N, Dong HT, Hirono I, Soontara C, Areechon N, et al. Molecular evidence for homologous strains of infectious spleen and kidney necrosis virus (ISKNV) genotype I infecting inland freshwater cultured Asian sea bass (Lates calcarifer) in Thailand. Arch Virol. 2021;166(11):3061-74. Epub 2021/09/01. [CrossRef] [PubMed]
- Thanasaksiri K, Takano R, Fukuda K, Chaweepack T, Wongtavatchai J. Identification of infectious spleen and kidney necrosis virus from farmed barramundi Lates calcarifer in Thailand and study of its pathogenicity. Aquaculture. 2019;500:188-91. PubMed PMID: WOS:000452969500022. [CrossRef]
- Zhu ZM, Duan C, Li Y, Huang CL, Weng SP, He JG, et al. Pathogenicity and histopathology of infectious spleen and kidney necrosis virus genotype II (ISKNV-II) recovering from mass mortality of farmed Asian seabass, Lates calcarifer, in Southern China. Aquaculture. 2021;534. doi: ARTN 736326.
- PubMed PMID: WOS:000614762200002. [CrossRef]
- Fu Y, Li Y, Zhang W, Fu W, Li W, Zhu Z, et al. Effectively protecting Asian seabass Lates calcarifer from ISKNV-I, ISKNV-II, RSIV-II and SDDV by an inactivated ISKNV-I and SDDV bivalent vaccine. Aquaculture. 2023;566.
- Nishizawa T, Furuhashi M, Nagai T, Nakai T, Muroga K. Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Appl Environ Microbiol. 1997;63(4):1633-6. [PubMed]
- Munday BL, Kwang J, Moody N. Betanodavirus infections of teleost fish: a review. Journal of Fish Diseases. 2002;25(3):127-42. PubMed PMID: WOS:000174711900001. [CrossRef]
- Le Breton A, Grisez L, Sweetman J, Ollevier F. Viral nervous necrosis (VNN) associated with mass mortalities in cage-reared sea bass, Dicentrarchus labrax (L.) Journal of Fish Diseases. 1997;20(2):145-51. [CrossRef]
- Nguyen HD, Nakai T, Muroga K. Progression of striped jack nervous necrosis virus (SJNNV) infection in naturally and experimentally infected striped jack Pseudocaranx dentex larvae. . Diseases of Aquatic Organisms. 1996;24(2):99-105. [CrossRef]
- Azad IS, Shekhar MS, Thirunavukkarasu AR, Jithendran KP. Viral nerve necrosis in hatchery-produced fry of Asian seabass Lates calcarifer: sequential microscopic analysis of histopathology. Diseases of Aquatic Organisms. 2006;73(2):123-30. PubMed PMID: WOS:000244271900004. [CrossRef]
- Kotob MH, Gorgoglione B, Kumar G, Abdelzaher M, Saleh M, El-Matbouli M. The impact of Tetracapsuloides bryosalmonae and Myxobolus cerebralis co-infections on pathology in rainbow trout. Parasite Vector. 2017;10. doi: ARTN 442.
- PubMed PMID: WOS:000412088400002. [CrossRef]
- Ma H, Xie J, Weng S, Zhou T, He J. Co-infection of megalocytivirus and viral nervous necrosis virus in a very severe mass mortality of juvenile orange-spotted groupers (Epinephelus coioides). Aquaculture. 2012;358-359:170-5.
- Jin YQ, Bergmann SM, Mai QY, Yang Y, Liu WQ, Sun DL, et al. Simultaneous Isolation and Identification of Largemouth Bass Virus and Rhabdovirus from Moribund Largemouth Bass (Micropterus salmoides). Viruses-Basel. 2022;14(8). doi: ARTN 1643.
- PubMed PMID: WOS:000845304100001. [CrossRef]
- Jitrakorn S, Gangnonngiw W, Bunnontae M, Manajit O, Rattanarojpong T, Chaivisuthangkura P, et al. Infectious cell culture system for concurrent propagation and purification of ISKNV and nervous necrosis virus from Asian Sea bass (Lates calcarifer). Aquaculture. 2020;520. doi: ARTN 734931.
- PubMed PMID: WOS:000515498200037. [CrossRef]
- Dong C, Weng S, Luo Y, Huang M, Ai H, Yin Z, et al. A new marine megalocytivirus from spotted knifejaw, Oplegnathus punctatus, and its pathogenicity to freshwater mandarinfish, Siniperca chuatsi. Virus Res. 2010;147(1):98-106. Epub 2009/11/10. [CrossRef] [PubMed]
- Lai YX, Jin BL, Xu Y, Huang LJ, Huang RQ, Zhang Y, et al. Immune responses of orange-spotted grouper,Epinephelus coioides, against virus-like particles of betanodavirus produced in Escherichia coli. Vet Immunol Immunop. 2014;157(1-2):87-96. PubMed PMID: WOS:000329952600010. [CrossRef]
- Dong C, Weng S, Shi X, Xu X, Shi N, He J. Development of a mandarin fish Siniperca chuatsi fry cell line suitable for the study of infectious spleen and kidney necrosis virus (ISKNV). Virus Res. 2008;135(2):273-81. Epub 2008/05/20. [CrossRef] [PubMed]
- Frerichs GN, Rodger HD, Peric Z. Cell Culture Isolation of Piscine Neuropathy Nodavirus from Juvenile Sea Bass, Dicentrarchus labrax. Journal of General Virology. 1996;77(9):2067-71.
- Lehwark P, Greiner S. GB2sequin - A file converter preparing custom GenBank files for database submission. Genomics. 2019;111(4):759-61. [CrossRef]
- Stothard P, Wishart DS. Circular genome visualization and exploration using CGView. Bioinformatics. 2005;21(4):537-9. Epub 2004/10/14. [CrossRef] [PubMed]
- Huang R, Zhu G, Zhang J, Lai Y, Xu Y, He J, et al. Betanodavirus-like particles enter host cells via clathrin-mediated endocytosis in a cholesterol-, pH- and cytoskeleton-dependent manner. Vet Res. 2017;48(1):8. [CrossRef] [PubMed] [PubMed Central]
- Adachi K, Ichinose T, Watanabe K, Kitazato K, Kobayashi N. Potential for the replication of the betanodavirus redspotted grouper nervous necrosis virus in human cell lines. Archives of Virology. 2008;153(1):15-24. PubMed PMID: WOS:000252473800002. [CrossRef]
- Clem LW, Moewus L, Sigel MM. Studies with Cells From Marine Fish in Tissue Culture. Proc Soc Exp Biol Med. 1961;108 762-6. [CrossRef]
- Kibenge FS, Munir K, Kibenge MJ, Joseph T, Moneke E. Infectious salmon anemia virus: causative agent, pathogenesis and immunity. Animal Health Research Reviews. 2004;5(1):65-78. [CrossRef]
- Lee KW, Chi SC, Cheng TM. Interference of the life cycle of fish nodavirus with fish retrovirus. Journal of General Virology. 2002;83:2469-74. PubMed PMID: WOS:000178203200016. [CrossRef]
- Long A, Garver KA, Jones SRM. Synergistic osmoregulatory dysfunction during salmon lice (Lepeophtheirus salmonis) and infectious hematopoietic necrosis virus co-infection in sockeye salmon (Oncorhynchus nerka) smolts. Journal of Fish Diseases. 2019;42(6):869-82. PubMed PMID: WOS:000467439700009. [CrossRef]
- Lee KK, Yang TI, Liu PC, Wu JL, Hsu YL. Dual challenges of infectious pancreatic necrosis virus and Vibrio carchariae in the grouper, Epinephelus sp. Virus Research. 1999;63(1-2):131-4. PubMed PMID: WOS:000082717700016. [CrossRef]
- Parkingking R, Takano R, Nishizawa T, Mori K, Iida Y, Arimoto M, et al. Experimental coinfection with aquabirnavirus and viral hemorrhagic septicemia virus (VHSV), Edwardsiella tarda or Streptococcus iniae in Japanese flounder Paralichthys olivaceus. Fish Pathology. 2003;38(1):15-21. PubMed PMID: WOS:000181894100003.
- Ogut H, Cavus N. A comparison of ectoparasite prevalence and occurrence of viral haemorrhagic septicemia virus (VHSV) in whiting. Rev Biol Mar Oceanog. 2014;49(1):91-6. PubMed PMID: WOS:000335226000010. [CrossRef]
- López-Bueno A, Mavian C, Labella AM, Castro D, Borrego JJ, Alcami A, et al. Concurrence of Iridovirus, Polyomavirus, and a Unique Member of a New Group of Fish Papillomaviruses in Lymphocystis Disease-Affected Gilthead Sea Bream. Journal of Virology. 2016;90(19):8768-79. PubMed PMID: WOS:000383761900033. [CrossRef]
- Kuo HC, Wang TY, Hsu HH, Chen PP, Lee SH, Chen YM, et al. Nervous necrosis virus replicates following the embryo development and dual infection with iridovirus at juvenile stage in grouper. PLoS One. 2012;7(4):e36183. [CrossRef] [PubMed] [PubMed Central]
Figure 2.
Detection of ISKNV and NNV using conventional PCR and RT-PCR. Left, the amplification result of ISKNV-mcp gene using a universal prime set for ISKNV/RSIV; Red arrow indicates the expected amplification bands. Right, the resultant amplification of NNV cp gene. “+” positive control; “-” negative control.
Figure 2.
Detection of ISKNV and NNV using conventional PCR and RT-PCR. Left, the amplification result of ISKNV-mcp gene using a universal prime set for ISKNV/RSIV; Red arrow indicates the expected amplification bands. Right, the resultant amplification of NNV cp gene. “+” positive control; “-” negative control.
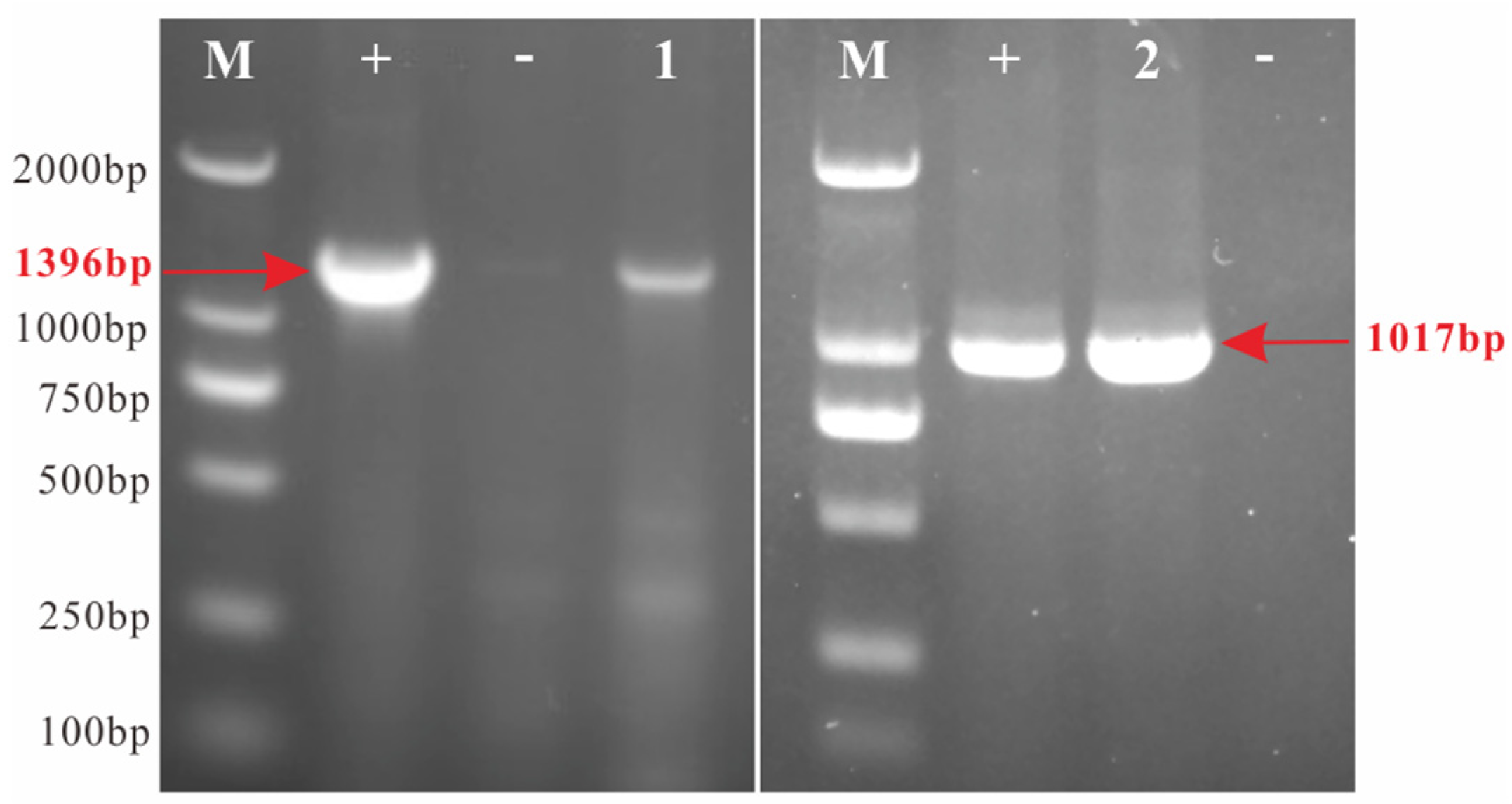
Figure 3.
Detection of ISKNV and NNV of the naturally diseased L. calcarifer by immunohistochemistry. Left (A-D), liver, spleen, eye and brain tissues from healthy fish; Right (E-H), liver, spleen, eye and brain tissues from the naturally diseased fish. Uniform, tightly arranged distribution of cells could be seen in healthy liver tissue (A); Several ISKNV signals (red arrow) and a loose mesh-like structure were observed in diseased liver tissue (E). Numerous ISKNV signals were concentrated in spleen tissue (F). The retina, where a lot of NNV viruses were concentrated and stained as dark brown (red arrow(G)). Compared with healthy fish (D), the brain tissue of diseased fish showed vacuolation (red arrow) (H).
Figure 3.
Detection of ISKNV and NNV of the naturally diseased L. calcarifer by immunohistochemistry. Left (A-D), liver, spleen, eye and brain tissues from healthy fish; Right (E-H), liver, spleen, eye and brain tissues from the naturally diseased fish. Uniform, tightly arranged distribution of cells could be seen in healthy liver tissue (A); Several ISKNV signals (red arrow) and a loose mesh-like structure were observed in diseased liver tissue (E). Numerous ISKNV signals were concentrated in spleen tissue (F). The retina, where a lot of NNV viruses were concentrated and stained as dark brown (red arrow(G)). Compared with healthy fish (D), the brain tissue of diseased fish showed vacuolation (red arrow) (H).
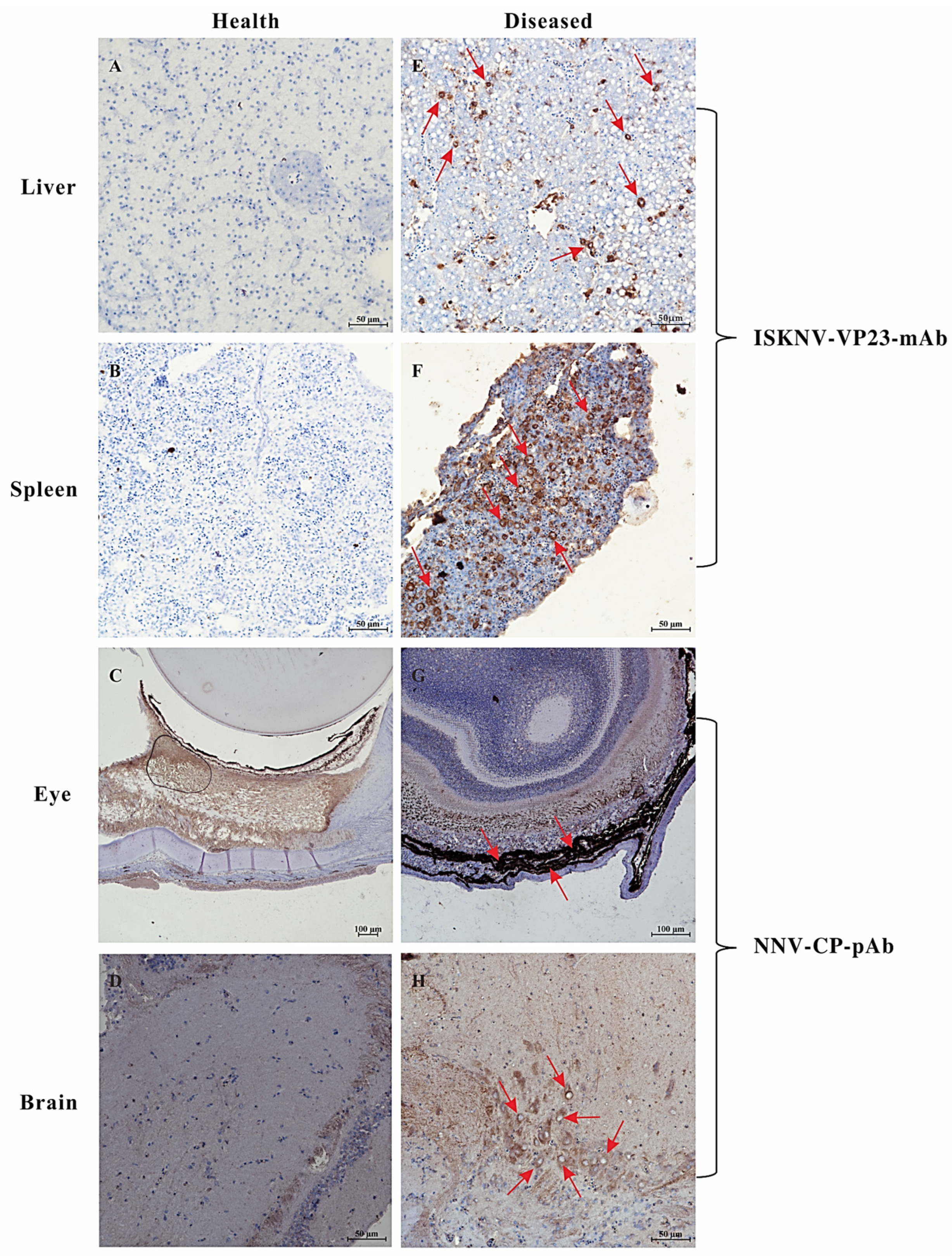
Figure 4.
Detection of ISKNV and NNV of the naturally diseased L. calcarifer by immunofluorescence assay. The liver and spleen tissues were stained by green fluorescence, which is associated with anti-ISKNV-VP23-mAb. The brain and eye tissues were stained by green fluorescence, which is associated with anti-NNV-CP-pAb.
Figure 4.
Detection of ISKNV and NNV of the naturally diseased L. calcarifer by immunofluorescence assay. The liver and spleen tissues were stained by green fluorescence, which is associated with anti-ISKNV-VP23-mAb. The brain and eye tissues were stained by green fluorescence, which is associated with anti-NNV-CP-pAb.
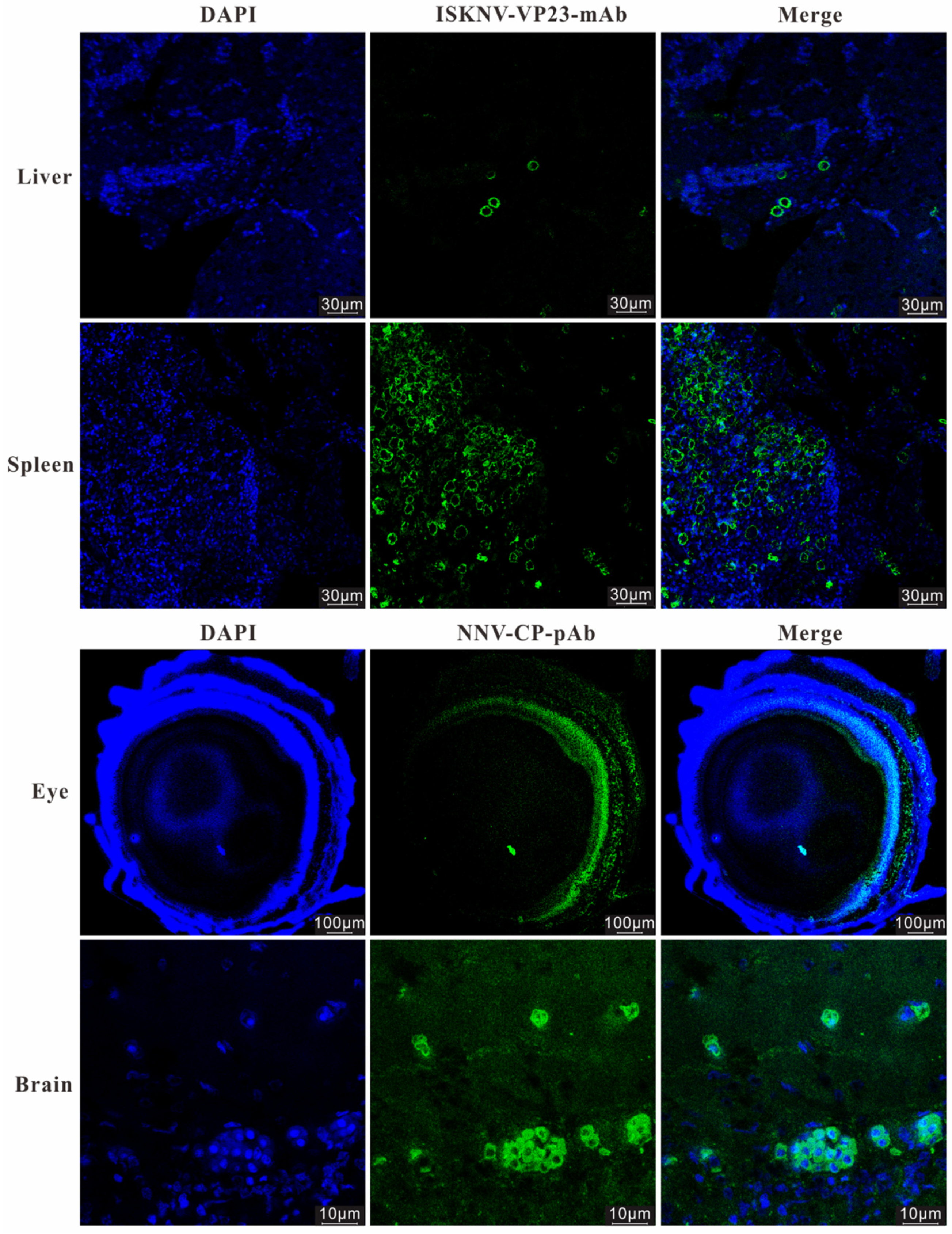
Figure 5.
Cytopathic effects of infected MFF-1 cells and SSN-1 cells incubation with homogenate of liver and spleen, and homogenate of brain and eye, respectively. Obvious CPEs were observed in MFF-1 cell and SSN-1 cell at 4 dpi and 2 dpi, respectively. .
Figure 5.
Cytopathic effects of infected MFF-1 cells and SSN-1 cells incubation with homogenate of liver and spleen, and homogenate of brain and eye, respectively. Obvious CPEs were observed in MFF-1 cell and SSN-1 cell at 4 dpi and 2 dpi, respectively. .
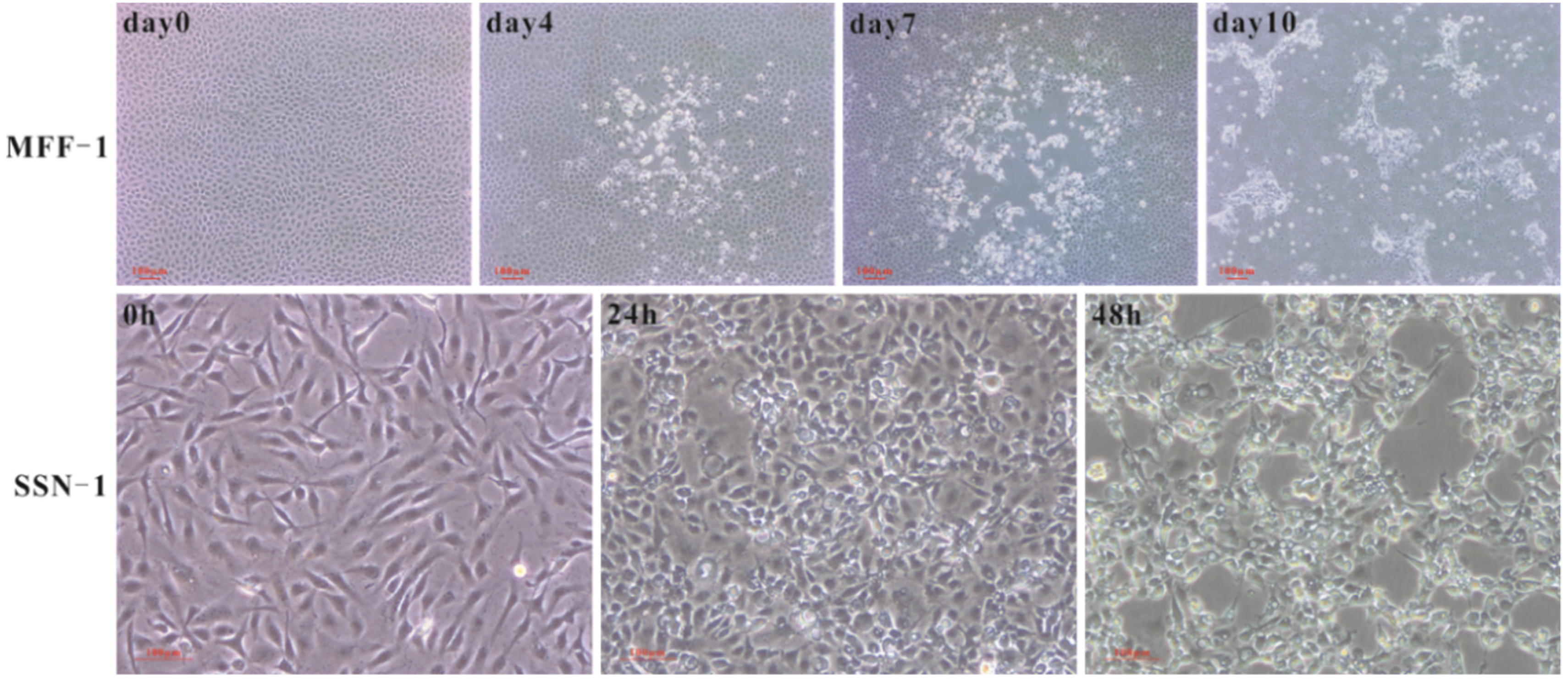
Figure 6.
Double immunofluorescence detection of ISKNV and NNV in infected MFF-1 and SSN-1 cell lines. A, MFF-1 cells inoculation with mixed multi-tissue homogenate at 3 dpi; B, MFF-1 cells inoculation with liver and spleen homogenate at viral passage 3. C, SSN-1 cells inoculation with brain and eye homogenate at viral passage 3.
Figure 6.
Double immunofluorescence detection of ISKNV and NNV in infected MFF-1 and SSN-1 cell lines. A, MFF-1 cells inoculation with mixed multi-tissue homogenate at 3 dpi; B, MFF-1 cells inoculation with liver and spleen homogenate at viral passage 3. C, SSN-1 cells inoculation with brain and eye homogenate at viral passage 3.
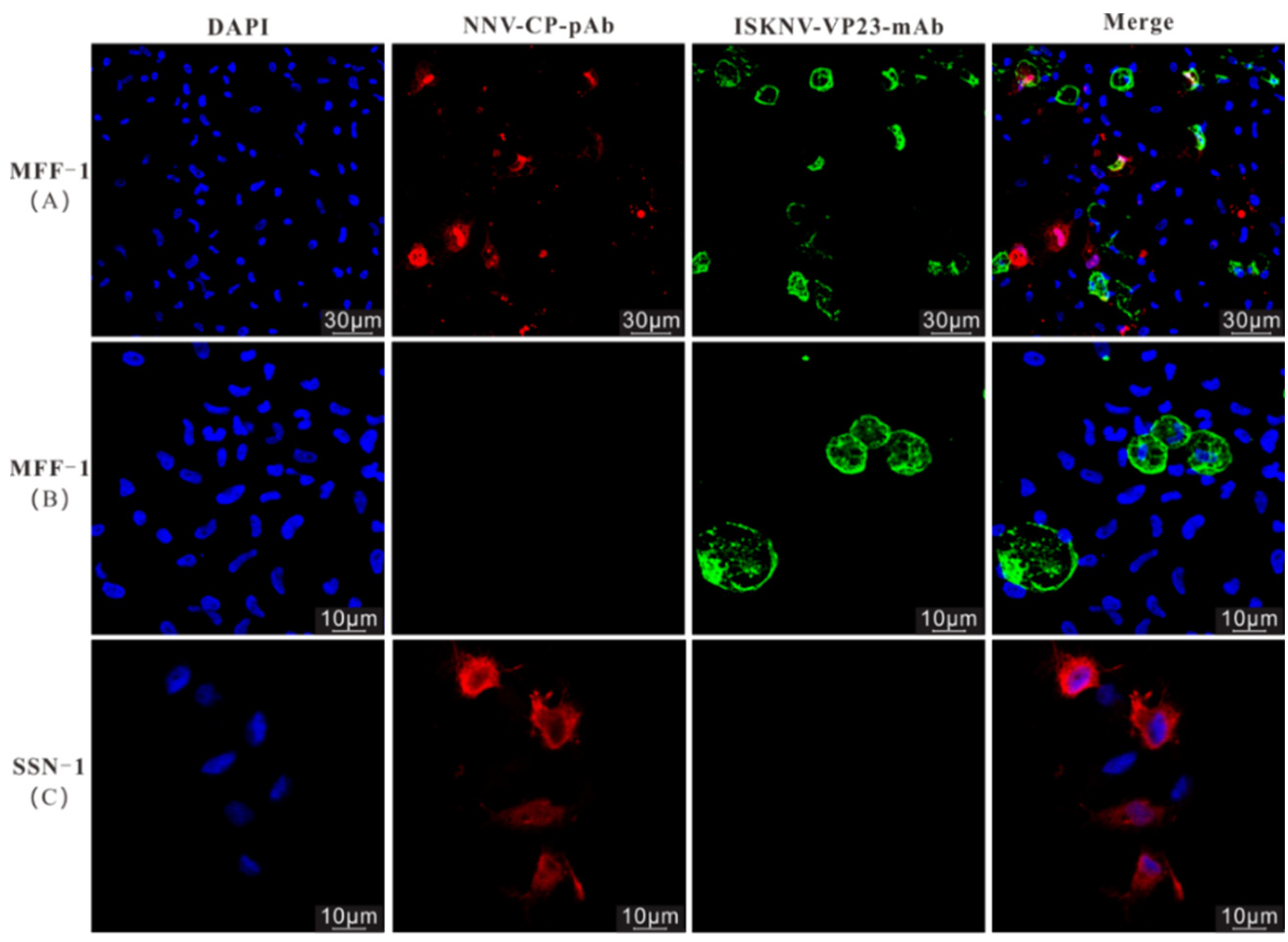
Figure 7.
The genomic map of ASB-ISKNV-23. The red arrow and the blue arrow represent the OFR of ASB-ISKNV-23. Dark blue circle represents GC content. Yellow and green circles represent GC Skew+ and GC Skew-, respectively.
Figure 7.
The genomic map of ASB-ISKNV-23. The red arrow and the blue arrow represent the OFR of ASB-ISKNV-23. Dark blue circle represents GC content. Yellow and green circles represent GC Skew+ and GC Skew-, respectively.
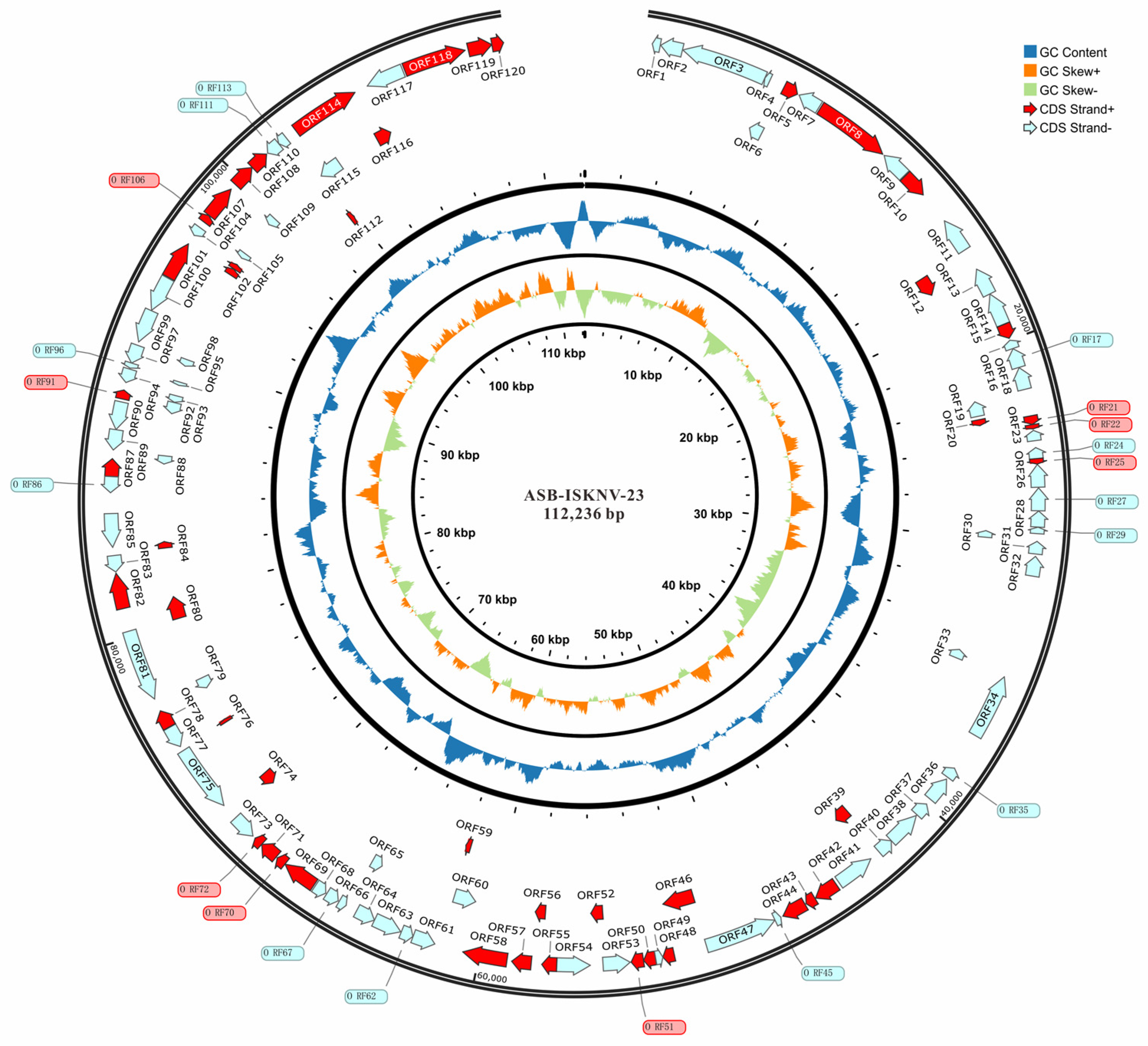
Figure 8.
The genomic map of ASB-NNV-23. Yellow and purple arrows represent the ORF of ASB-NNV-23.
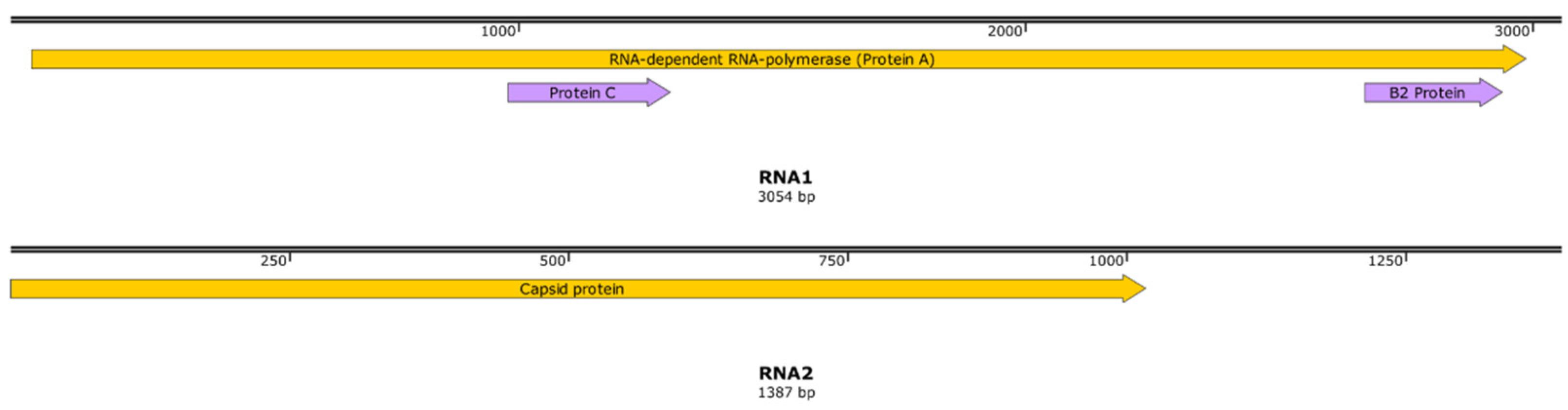
Figure 9.
Phylogenetic trees of ASB-ISKNV-23 and ASB-NNV-23. Phylogenetic analysis showed that ASB-ISKNV-23 (upper) is an ISKNV-II sub-genotype isolate in ISKNV species, and ASB-NNV-23 (down) is an RGNNV genotype isolate.
Figure 9.
Phylogenetic trees of ASB-ISKNV-23 and ASB-NNV-23. Phylogenetic analysis showed that ASB-ISKNV-23 (upper) is an ISKNV-II sub-genotype isolate in ISKNV species, and ASB-NNV-23 (down) is an RGNNV genotype isolate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



