
Submitted:
06 January 2025
Posted:
06 January 2025
You are already at the latest version
Abstract
Skeletal muscle aging, or sarcopenia, involves progressive muscle mass and function loss, which limits mobility and independence in the aged populations. This decline is driven by chronic inflammation, oxidative stress, and insulin resistance, all of which impair muscle regeneration and accelerate protein breakdown. Mineralocorticoid receptors (MRs), known for roles in electrolyte balance, have emerged as key regulators of these processes in skeletal muscle. MR activation promotes inflammatory signaling, increases oxidative stress, and worsens insulin resistance, accelerating sarcopenia progression. This review examines the impact of MRs on muscle health and highlights the therapeutic potential of targeting these receptors to counteract age-related muscle loss. MR antagonists, such as spironolactone, show promise in reducing inflammation and oxidative damage, potentially slowing sarcopenia. Physical exercise, an established intervention for muscle health, may enhance MR antagonism effects by improving insulin sensitivity and reducing inflammation. However, more research is needed to determine the efficacy and safety of combined MR antagonists and exercise protocols for preventing sarcopenia in older adults.
Keywords:
elderly
; mineralocorticoid
; sarcopenia
1. Introduction
Maintaining muscle mass and functionality is essential during the aging process to avoid decline in mobility, independence, and in the overall quality of life among older adults [1]. Aging naturally leads to a progressive loss of muscle mass and strength that is highly influenced by lifestyle habits and lead to a condition known as sarcopenia [2], which in its turn is associated with increased frailty, a higher risk of falls and injuries, and slower recovery following trauma or surgical procedures [2,3].
Current literature highlights that, in addition to muscle mass loss, aging is linked to metabolic alterations within muscle tissue [4]. These changes include elevated oxidative stress, inflammation and insulin resistance along with impaired autophagy in skeletal muscle [4,5,6]. Such metabolic disruptions impair the muscle's regenerative capacity and enhance the catabolism of muscle proteins, resulting in reduced strength and functionality [4,5,6].
Consequently, practices aiming to maintain adequate muscle mass tend to enhance functional independency so to alleviate the burden on healthcare systems [7]. Older individuals with preserved musculature experience fewer complications related to mobility and metabolic health [1,7]. Furthermore, the preservation of muscle function directly influences the ability to perform physical activities and maintain an active lifestyle, which is crucial for overall cardiovascular and metabolic well-being [8].
In this context, mineralocorticoid receptors (MRs) play a pivotal role in muscle metabolism and function [9,10,11]. MRs influence critical processes necessary for maintaining electrolyte balance, modulating inflammatory responses, and regulating energy metabolism [12,13,14,15]. Traditionally recognized for their role in maintaining sodium and potassium homeostasis and controlling water retention in the kidneys, recent studies have revealed that MRs are also present in the skeletal muscle tissue and significantly contribute to muscle quality indexes [9,10].
2. Mineralocorticoid Receptors (MRs)
MRs exist as two distinct isoforms, MR-1α and MR-1β, which are differentiated by their unique promoter sequences, P1 and P2, respectively [16,17]. These isoforms share core functional domains that exhibit distinct regulatory patterns and tissue-specific expression [16]. P1 is typically active in classic MR-responsive tissues, such as the kidneys, where the receptor plays a pivotal role in blood pressure regulation by electrolyte homeostasis [18]. In contrast, P2 drives expression in non-classical tissues, including skeletal muscle, where MRs influence processes such as inflammation, oxidative stress, and glucose metabolism [19,20]. Specifically, excessive activation of MRs induces insulin resistance in skeletal muscle by impairing GLUT4 translocation to the cell membrane, which reduces glucose uptake by muscle fibers [21]. This dysfunction results in reduced glucose availability for anabolic processes, contributing to the catabolic state characteristic of sarcopenia [22]. Thus, the effects of MRs on glucose metabolism are an extension of their ability to amplify inflammatory and oxidative stress pathways, creating a deleterious cycle that exacerbates muscle atrophy. This duality in promoter activity allows a fine-tuned regulation of MR expression, aligning receptor function with the physiological demands of specific tissues. The differential expression and regulation of MR isoforms may also underlie tissue-specific responses to mineralocorticoids, presenting a potential therapeutic avenue to modulate age-related phenotypes as sarcopenia [23].
This review aims to elucidate the role of MRs in skeletal muscle aging and to explore the therapeutic potential of targeting these receptors to promote healthy aging. By understanding the molecular mechanisms through which MRs influence muscle physiology, we can identify novel interventions to prevent or mitigate sarcopenia, thereby enhancing the quality of life and functional independence of the aging population.
2.1. Skeletal Muscle Aging: Insights into the Drivers of Sarcopenia
Sarcopenia is marked by a systemic and progressive loss of muscle mass, quality, and strength, which significantly impacts mobility and independence, especially among the aged [2]. This degenerative process is driven by several key factors that collectively contribute to sarcopenia. Chronic low-grade inflammation, characterized by elevated levels of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukins (IL-1β, IL-6), accelerates muscle protein degradation and impairs muscle regeneration [24,25]. Additionally, oxidative stress resulting from increased production of reactive oxygen species (ROS) leads to damage in cellular proteins, lipids, and DNA, further compromising muscle integrity [26]. Insulin resistance exacerbates this scenario by reducing the energy available for anabolic processes, thereby promoting muscle atrophy [27]. Finally, dysfunction of satellite cells, which play a crucial role in muscle regeneration, hampers the tissue's ability to recover and adapt, intensifying the loss of muscle mass and strength [28]. As these factors overlap in terms of signaling pathways throughout the aging process, skeletal muscle fibers (mainly of the type II) undergoes a marked reduction in size and quantity, basically due to greater protein degradation rates [29,30]. These changes collectively lead to the gradual loss of muscle mass and strength that underlies the sarcopenic phenotype.
Emerging evidence highlights the intricate role of MRs in skeletal muscle homeostasis, particularly under conditions of aging [31,32]. The interplay between MRs activation and key catabolic processes creates an environment conducive to muscle degradation. This cascade of events not only drives protein catabolism but also disrupts the regenerative capacity of muscle fibers, compounding the loss of muscle mass and function characteristic of sarcopenia [31,32]. Refining our understanding of these mechanisms can inform targeted interventions to disrupt this cycle, offering a more nuanced approach to maintaining muscle health in the aged population.
2.2. MR Activation: Bridging Ion Transport and Insulin Resistance
Beyond their well-established roles in renal regulation [12,13,33], MRs have emerged as key modulators of skeletal muscle physiology [9]. In muscle tissue, MRs influence ion distribution, particularly sodium and potassium, which are critical for maintaining excitability and contractility [23]. This ion balance is integral to muscle function and resilience, particularly under conditions of metabolic stress or aging. By regulating these processes, MRs contribute to the broader homeostatic mechanisms that sustain muscle performance and adaptability, as under conditions of excessive MR activation, as observed in certain pathological states and with aging, an overload of sodium accumulates within muscle cells, leading to intracellular ion imbalance and water retention [9,13,23]. This ionic disruption can cause edema and increased stress on muscle fibers, creating an unfavorable environment for muscle contraction and regeneration, which may exacerbate the loss of muscle mass and function due to atrophy [34,35].
Additionally, MRs influence glucose metabolism in skeletal muscle by modulating insulin sensitivity and glucose uptake via GLUT4 [21,36]. Excessive MRs activation is associated with increased insulin resistance, impairing glucose absorption by muscle cells and thereby reducing the energy available for anabolic processes [21,36]. This effect disrupts muscle energy metabolism, leading to a catabolic state that drives protein degradation and muscle mass loss—common characteristics in aging individuals. Studies suggest that this insulin resistance, amplified by MRs activation, also elevates the risk of related metabolic disorders, such as metabolic syndrome and type 2 diabetes [37,38,39,40].
These adverse effects of excessive MRs activation, including ionic imbalance and insulin resistance, may further be exacerbated by aging, a period during which skeletal muscle naturally experiences a decline in regenerative and functional capacity [41,42]. Prolonged MRs activation thus contributes not only to muscle deterioration but also to metabolic dysfunction within skeletal muscle, positioning MRs as a promising therapeutic target for interventions aimed at preserving muscle metabolism in the elderly.
2.3. Linking MRs to Oxidative Stress and Inflammation
The activation of MRs leads to chronic inflammation and oxidative stress [32,43], processes that accelerate muscle aging and drive the progression of sarcopenia [24,25,26]. In response to MRs activation, a cascade of cellular events is triggered, leading to elevated production of pro-inflammatory mediators and ROS [44]. These factors establish a persistent inflammatory environment that may impair muscle regeneration and promote protein degradation [45]. In this scenario, TNF-α plays a major role in driving muscle catabolism, primarily through its ability to activate proteolytic pathways such as the ubiquitin-proteasome and autophagy-lysosome systems [46]. Elevated TNF-α levels, often observed in chronic inflammation and aging, induce the expression of E3 ubiquitin ligases like MuRF1 and Atrogin-1, which target myofibrillar proteins for degradation [47]. Additionally, TNF-α impairs myogenesis by inhibiting myogenic regulatory factors such as MyoD and myogenin by promoting mitochondrial dysfunction [48], which exacerbates oxidative stress and further contributes to muscle atrophy.
MRs have been implicated in amplifying TNF-α signaling [49]. Thus, MRs activation enhances the expression of inflammation-associated genes, including those encoding key mediators such as pro-inflammatory cytokines and interleukins [50,51]. These pro-inflammatory mediators are known to recruit immune cells and exacerbate tissue damage [52], particularly in aged muscle, where cellular turnover is already limited. This creates a feedback loop where pro-inflammatory mediators, such as TNF-α, induce a catabolic signaling which is sustained, perpetuating muscle protein degradation and impairing regeneration. This chronic inflammatory state negatively impacts muscle function by promoting muscle fiber degradation and inhibiting protein synthesis, processes essential for maintaining muscle mass and strength [53]. Additionally, muscle-derived myokines play a pivotal role in modulating systemic inflammation and metabolic processes, acting as signaling molecules released during muscle contraction which are known to influence muscle regeneration [54,55].
In addition to driving inflammation, excessive MRs activation increases ROS production [44], which may contribute to oxidative stress in muscle tissue. The buildup of ROS inflicts damage on cellular structures, including lipids, proteins, and DNA, compromising muscle cell integrity and diminishing the tissue’s regenerative capacity [56]. With aging, the natural antioxidant defenses become less effective, amplifying the detrimental effects of MRs-induced ROS. This sustained oxidative stress accelerates muscle degeneration, creating a cycle of incomplete repair that perpetuates sarcopenia [57]. This suggests a complex interaction where excessive MRs activation may also alter the balance of myokine secretion, potentially amplifying muscle catabolism and impairing regenerative processes. Thus, this pathological mechanism highlights MRs as a potential therapeutic target for mitigating age-related muscle deterioration by reducing inflammation and oxidative stress, with the goal of preserving muscle mass and function in the elderly.
2.4. Harnessing MR Antagonists for Therapy
The pathological activation of MRs contributes to molecular disturbances which suppress anabolic pathways and enhances catabolic signaling, driving excessive protein degradation in skeletal muscle (Figure 1). A key mechanism underlying this process is the suppression of myogenesis, coupled with increased apoptosis of myocytes [58]. MRs activation directly impairs myogenic differentiation by downregulating critical regulatory factors while increasing oxidative stress that promotes mitochondrial dysfunction [61,62,63]. This dysfunction leads to the release of pro-apoptotic factors, including cytochrome c, which activates caspase-mediated apoptotic pathways [22,64]. These events result in myonuclear loss and a reduction in muscle cross-sectional area and strength, hallmark features of sarcopenia.
MRs antagonists, such as spironolactone [50,59,65] and finerenone [66], show promise in mitigating these effects by reducing ROS production, modulating inflammatory cytokine expression, and limiting apoptosis. Additionally, these agents have been shown to promote anti-inflammatory macrophage phenotypes, reduce muscle fibrosis, and enhance the regenerative capacity of muscle tissue. These findings highlight the therapeutic utility of MRs antagonists in targeting the molecular drivers of sarcopenia [23]. However, targeted clinical studies are still needed to confirm the efficacy and safety of these antagonists in healthy older adults and in the early stages of muscle wasting. Future research should focus on optimizing dosage and safety profiles to minimize adverse effects while promoting muscle preservation. This approach could position MRs antagonists as a viable and effective non-invasive therapy to counteract age-related muscle decline, supporting muscular health and functionality in the aged population.
2.5. Exercise as a Modulator of MRs
Physical exercise plays a critical role in regulating the activity of MRs [65,66,67], and serves as an effective adjunct in preventing sarcopenia associated with aging [68]. In this sense, exercise also acts on counteracting the dysregulation of the hypothalamic-pituitary-adrenal (HPA) axis [69,70], particularly under conditions of sleep disruption [71], which may be associated with MRs activation [72]. Sleep disturbances impair anabolic signaling pathways, including those mediated by IGF-1, while enhancing catabolic processes such as the ubiquitin-proteasome and autophagy-lysosome systems [73]. Regular physical activity restores this balance, reducing HPA axis-driven catabolism and improving muscle health [74], which consequently may reduce MRs expression.
Physical exercise enhances energy metabolism and insulin sensitivity in skeletal muscle [75], which counteracts the adverse effects of excessive MRs activation, avoiding muscle catabolism. By reducing the production of pro-inflammatory mediators and ROS [56,76], exercise can limit the harmful effects linked to MRs activation, thereby preserving muscle fiber integrity and improving muscle functionality. In addition, studies suggest that regular exercise, particularly resistance training [77], helps modulate skeletal muscle metabolism, fostering an environment less prone to inflammation and oxidative stress, both of which are key factors in preserving muscle health during aging.
Combining MRs antagonists with exercise interventions may provide a synergistic approach to addressing the multifactorial nature of sarcopenia. This integrated strategy offers a comprehensive framework for preserving muscle mass, strength, and functionality, ultimately improving the quality of life in aged populations due to functional independence.
2.6. Future Perspectives
Our understanding of the specific role of MRs in the interplay between oxidative stress and insulin resistance across different age groups remains incomplete. Although studies suggest that MRs activation increases oxidative stress and disrupts insulin signaling [31,32], factors that contribute to insulin resistance in skeletal muscle, these interactions have largely been explored in pathological or animal models. How these mechanisms unfold across various aging stages and whether MRs sensitivity to oxidative stress changes with age are still poorly understood. Addressing these gaps requires future research to investigate the potential of antioxidant interventions and MRs antagonists as modulators of insulin response in aging populations. Studies examining whether antioxidants combined with MRs blockers can effectively reduce oxidative stress and restore insulin signaling in healthy older adults and in early stages of insulin resistance could yield promising preventive and therapeutic strategies, ultimately offering new avenues to slow metabolic decline and improve quality of life in aging individuals. Additionally, future studies exploring the interplay between MRs and myokines could provide valuable insights into therapeutic strategies for mitigating muscle aging and enhancing overall health outcomes in aged populations.
While MRs antagonists, such as spironolactone, have shown benefits in specific conditions like heart failure and muscular dystrophies [78,79], data on their impact in healthy older adults is limited. In particular, there is a pressing need for studies focusing on MRs antagonists in the context of sarcopenia across age, which may be an essential factor in assessing the preventive and therapeutic potential of these agents under the aging process. Given the role of MRs in driving inflammation and oxidative stress [32,43], MRs antagonists may be valuable in mitigating age-related muscle mass and strength loss. However, the absence of clinical trials targeting healthy aged individuals limits our ability to fully understand the efficacy and safety of these antagonists in preventing muscle decline. Furthermore, future clinical studies should also investigate aged populations without significant comorbidities to evaluate whether MRs antagonists can serve as effective preventative strategies for delaying muscle wasting progression and enhancing muscle function throughout aging.
Additionally, the combined effects of physical exercise and MRs antagonism on muscle function in the elderly remain largely unexplored. Although the benefits of physical exercise in preserving muscle mass and strength are well-established [68,77,80], and MRs antagonists have shown potential to reduce catabolic signaling in skeletal muscle, the synergistic potential of these approaches has yet to be fully investigated. Emerging evidence suggests that these interventions may complement each other, with exercise potentially mitigating the adverse effects of MRs activation and fostering a more favorable muscle environment in aging populations. To maximize muscle preservation, clinical trials are needed to evaluate the efficacy of integrated protocols combining exercise and MRs antagonists in aged populations. Such studies should aim to identify optimal dosages and frequencies for both exercise and MRs antagonist use to enhance muscle strength, mass, and functionality, offering a comprehensive approach to preventing sarcopenia and physical decline in older adults.
3. Conclusion
MRs are central to skeletal muscle aging, driving inflammation, oxidative stress, and insulin resistance which are factors that contribute to sarcopenia. This review underscores the importance of MRs as therapeutic targets, given their involvement in critical catabolic pathways. Specifically, MRs activation enhances TNF-α signaling, which activates proteolytic systems such as the ubiquitin-proteasome and autophagy-lysosome pathways, while suppressing myogenesis and promoting myocyte apoptosis. These mechanisms collectively contribute to muscle protein degradation and the progressive loss of muscle mass and function. Therapeutic interventions targeting MRs, such as spironolactone, have demonstrated potential in mitigating these detrimental effects (Figure 2). When combined with physical exercise, which enhances muscle integrity, MRs antagonism may offer an effective approach to managing sarcopenia. However, further research is needed to clarify MRs function across aging stages and to establish the safety and efficacy of combining MRs antagonists with exercise in both healthy and sarcopenic aged populations. These findings highlight the central role of MRs in muscle health and reinforce the need for innovative strategies to preserve muscle mass, strength, and overall quality of life in older adults.
References
- Trombetti, A.; Reid, K.F.; Hars, M.; Herrmann, F.R.; Pasha, E.; Phillips, E.M.; Fielding, R.A. Age-Associated Declines in Muscle Mass, Strength, Power, and Physical Performance: Impact on Fear of Falling and Quality of Life. Osteoporos Int 2016, 27, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Kamel, H.K. Sarcopenia and Aging. Nutrition Reviews 2003, 61, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Landi, F.; Liperoti, R.; Russo, A.; Giovannini, S.; Tosato, M.; Capoluongo, E.; Bernabei, R.; Onder, G. Sarcopenia as a Risk Factor for Falls in Elderly Individuals: Results from the ilSIRENTE Study. Clinical Nutrition 2012, 31, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Dirks, A.J.; Hofer, T.; Marzetti, E.; Pahor, M.; Leeuwenburgh, C. Mitochondrial DNA Mutations, Energy Metabolism and Apoptosis in Aging Muscle. Ageing Research Reviews 2006, 5, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Shou, J.; Chen, P.-J.; Xiao, W.-H. Mechanism of Increased Risk of Insulin Resistance in Aging Skeletal Muscle. Diabetol Metab Syndr 2020, 12, 14. [Google Scholar] [CrossRef]
- Chen, M.; Wang, Y.; Deng, S.; Lian, Z.; Yu, K. Skeletal Muscle Oxidative Stress and Inflammation in Aging: Focus on Antioxidant and Anti-Inflammatory Therapy. Front. Cell Dev. Biol. 2022, 10, 964130. [Google Scholar] [CrossRef]
- Álvarez-Bustos, A.; Rodríguez-Sánchez, B.; Carnicero-Carreño, J.A.; Sepúlveda-Loyola, W.; Garcia-Garcia, F.J.; Rodríguez-Mañas, L. Healthcare Cost Expenditures Associated to Frailty and Sarcopenia. BMC Geriatr 2022, 22, 747. [Google Scholar] [CrossRef]
- Tian, D.; Meng, J. Exercise for Prevention and Relief of Cardiovascular Disease: Prognoses, Mechanisms, and Approaches. Oxidative Medicine and Cellular Longevity 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Chadwick, J.A.; Hauck, J.S.; Lowe, J.; Shaw, J.J.; Guttridge, D.C.; Gomez-Sanchez, C.E.; Gomez-Sanchez, E.P.; Rafael-Fortney, J.A. Mineralocorticoid Receptors Are Present in Skeletal Muscle and Represent a Potential Therapeutic Target. FASEB j. 2015, 29, 4544–4554. [Google Scholar] [CrossRef]
- Howard, Z.M.; Gomatam, C.K.; Rabolli, C.P.; Lowe, J.; Piepho, A.B.; Bansal, S.S.; Accornero, F.; Rafael-Fortney, J.A. Mineralocorticoid Receptor Antagonists and Glucocorticoids Differentially Affect Skeletal Muscle Inflammation and Pathology in Muscular Dystrophy. JCI Insight 2022, 7, e159875. [Google Scholar] [CrossRef]
- Howard, Z.M.; Gomatam, C.K.; Piepho, A.B.; Rafael-Fortney, J.A. Mineralocorticoid Receptor Signaling in the Inflammatory Skeletal Muscle Microenvironments of Muscular Dystrophy and Acute Injury. Front. Pharmacol. 2022, 13, 942660. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, E.T.; Berman, S.; Streicher, J.; Estrada, C.M.; Caldwell, J.L.; Ghisays, V.; Ulrich-Lai, Y.; Solomon, M.B. Effects of Combined Glucocorticoid/Mineralocorticoid Receptor Modulation (CORT118335) on Energy Balance, Adiposity, and Lipid Metabolism in Male Rats. American Journal of Physiology-Endocrinology and Metabolism 2019, 317, E337–E349. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Sanchez, E.; Gomez-Sanchez, C.E. The Multifaceted Mineralocorticoid Receptor. In Comprehensive Physiology; Terjung, R., Ed.; Wiley, 2014; pp. 965–994 ISBN 978-0-470-65071-4.
- Zennaro, M.-C.; Caprio, M.; Fève, B. Mineralocorticoid Receptors in the Metabolic Syndrome. Trends in Endocrinology & Metabolism 2009, 20, 444–451. [Google Scholar] [CrossRef]
- Young, M.J. Mechanisms of Mineralocorticoid Receptor-Mediated Cardiac Fibrosis and Vascular Inflammation. Current Opinion in Nephrology & Hypertension 2008, 17, 174–180. [Google Scholar] [CrossRef]
- Pascual-Le Tallec, L.; Lombès, M. The Mineralocorticoid Receptor: A Journey Exploring Its Diversity and Specificity of Action. Molecular Endocrinology 2005, 19, 2211–2221. [Google Scholar] [CrossRef]
- Gaudenzi, C.; Mifsud, K.R.; Reul, J.M.H.M. Insights into Isoform-Specific Mineralocorticoid Receptor Action in the Hippocampus. Journal of Endocrinology 2023, 258, e220293. [Google Scholar] [CrossRef]
- Le Menuet, D.; Viengchareun, S.; Muffat-Joly, M.; Zennaro, M.-C.; Lombès, M. Expression and Function of the Human Mineralocorticoid Receptor: Lessons from Transgenic Mouse Models. Molecular and Cellular Endocrinology 2004, 217, 127–136. [Google Scholar] [CrossRef]
- Ibarrola, J.; Lu, Q.; Zennaro, M.-C.; Jaffe, I.Z. Mechanism by Which Inflammation and Oxidative Stress Induce Mineralocorticoid Receptor Gene Expression in Aging Vascular Smooth Muscle Cells. Hypertension 2023, 80, 111–124. [Google Scholar] [CrossRef]
- Kuhn, E.; Lombès, M. The Mineralocorticoid Receptor: A New Player Controlling Energy Homeostasis. Hormone Molecular Biology and Clinical Investigation 2013, 15, 59–69. [Google Scholar] [CrossRef]
- Hulse, J.L.; Habibi, J.; Igbekele, A.E.; Zhang, B.; Li, J.; Whaley-Connell, A.; Sowers, J.R.; Jia, G. Mineralocorticoid Receptors Mediate Diet-Induced Lipid Infiltration of Skeletal Muscle and Insulin Resistance. Endocrinology 2022, 163, bqac145. [Google Scholar] [CrossRef]
- Marzetti, E.; Privitera, G.; Simili, V.; Wohlgemuth, S.E.; Aulisa, L.; Pahor, M.; Leeuwenburgh, C. Multiple Pathways to the Same End: Mechanisms of Myonuclear Apoptosis in Sarcopenia of Aging. The Scientific World JOURNAL 2010, 10, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Burton, L.A.; McMurdo, M.E.T.; Struthers, A.D. Mineralocorticoid Antagonism: A Novel Way to Treat Sarcopenia and Physical Impairment in Older People?: Mineralocorticoid Antagonism. Clinical Endocrinology 2011, 75, 725–729. [Google Scholar] [CrossRef]
- Bano, G.; Trevisan, C.; Carraro, S.; Solmi, M.; Luchini, C.; Stubbs, B.; Manzato, E.; Sergi, G.; Veronese, N. Inflammation and Sarcopenia: A Systematic Review and Meta-Analysis. Maturitas 2017, 96, 10–15. [Google Scholar] [CrossRef]
- Dalle, S.; Rossmeislova, L.; Koppo, K. The Role of Inflammation in Age-Related Sarcopenia. Front. Physiol. 2017, 8, 1045. [Google Scholar] [CrossRef]
- Bellanti, F.; Buglio, A.L.; Vendemiale, G. Oxidative Stress and Sarcopenia. In Aging; Elsevier, 2020; pp. 95–103 ISBN 978-0-12-818698-5.
- Cleasby, M.E.; Jamieson, P.M.; Atherton, P.J. Insulin Resistance and Sarcopenia: Mechanistic Links between Common Co-Morbidities. Journal of Endocrinology 2016, 229, R67–R81. [Google Scholar] [CrossRef]
- Alway, S.E.; Myers, M.J.; Mohamed, J.S. Regulation of Satellite Cell Function in Sarcopenia. Front. Aging Neurosci. 2014, 6. [Google Scholar] [CrossRef]
- McPhee, J.S.; Cameron, J.; Maden-Wilkinson, T.; Piasecki, M.; Yap, M.H.; Jones, D.A.; Degens, H. The Contributions of Fiber Atrophy, Fiber Loss, In Situ Specific Force, and Voluntary Activation to Weakness in Sarcopenia. The Journals of Gerontology: Series A 2018, 73, 1287–1294. [Google Scholar] [CrossRef]
- Nishikawa, H.; Fukunishi, S.; Asai, A.; Yokohama, K.; Nishiguchi, S.; Higuchi, K. Pathophysiology and Mechanisms of Primary Sarcopenia (Review). Int J Mol Med 2021, 48, 156. [Google Scholar] [CrossRef]
- Jia, G.; Lockette, W.; Sowers, J.R. Mineralocorticoid Receptors in the Pathogenesis of Insulin Resistance and Related Disorders: From Basic Studies to Clinical Disease. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 2021, 320, R276–R286. [Google Scholar] [CrossRef]
- Kuster, G.M.; Kotlyar, E.; Rude, M.K.; Siwik, D.A.; Liao, R.; Colucci, W.S.; Sam, F. Mineralocorticoid Receptor Inhibition Ameliorates the Transition to Myocardial Failure and Decreases Oxidative Stress and Inflammation in Mice With Chronic Pressure Overload. Circulation 2005, 111, 420–427. [Google Scholar] [CrossRef]
- Krishna, G.G.; Kapoor, S.C. Potassium Supplementation Ameliorates Mineralocorticoid-Induced Sodium Retention. Kidney International 1993, 43, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Jurkat-Rott, K.; Weber, M.-A.; Fauler, M.; Guo, X.-H.; Holzherr, B.D.; Paczulla, A.; Nordsborg, N.; Joechle, W.; Lehmann-Horn, F. K + -Dependent Paradoxical Membrane Depolarization and Na + Overload, Major and Reversible Contributors to Weakness by Ion Channel Leaks. Proc. Natl. Acad. Sci. U.S.A. 2009, 106, 4036–4041. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M.; Wallace, H. Ion Channels in Inflammation. Pflugers Arch - Eur J Physiol 2011, 461, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Faught, E.; Vijayan, M.M. The Mineralocorticoid Receptor Functions as a Key Glucose Regulator in the Skeletal Muscle of Zebrafish. Endocrinology 2022, 163, bqac149. [Google Scholar] [CrossRef]
- Feraco, A.; Marzolla, V.; Scuteri, A.; Armani, A.; Caprio, M. Mineralocorticoid Receptors in Metabolic Syndrome: From Physiology to Disease. Trends in Endocrinology & Metabolism 2020, 31, 205–217. [Google Scholar] [CrossRef]
- Bender, S.B.; McGraw, A.P.; Jaffe, I.Z.; Sowers, J.R. Mineralocorticoid Receptor–Mediated Vascular Insulin Resistance. Diabetes 2013, 62, 313–319. [Google Scholar] [CrossRef]
- Korol, S.; Mottet, F.; Perreault, S.; Baker, W.L.; White, M.; De Denus, S. A Systematic Review and Meta-Analysis of the Impact of Mineralocorticoid Receptor Antagonists on Glucose Homeostasis. Medicine 2017, 96, e8719. [Google Scholar] [CrossRef]
- Barrera-Chimal, J.; Lima-Posada, I.; Bakris, G.L.; Jaisser, F. Mineralocorticoid Receptor Antagonists in Diabetic Kidney Disease — Mechanistic and Therapeutic Effects. Nat Rev Nephrol 2022, 18, 56–70. [Google Scholar] [CrossRef]
- Michelucci, A.; Liang, C.; Protasi, F.; Dirksen, R.T. Altered Ca2+ Handling and Oxidative Stress Underlie Mitochondrial Damage and Skeletal Muscle Dysfunction in Aging and Disease. Metabolites 2021, 11, 424. [Google Scholar] [CrossRef]
- Carmeli, E.; Coleman, R.; Reznick, A.Z. The Biochemistry of Aging Muscle. Experimental Gerontology 2002, 37, 477–489. [Google Scholar] [CrossRef]
- Gorini, S.; Kim, S.K.; Infante, M.; Mammi, C.; La Vignera, S.; Fabbri, A.; Jaffe, I.Z.; Caprio, M. Role of Aldosterone and Mineralocorticoid Receptor in Cardiovascular Aging. Front. Endocrinol. 2019, 10, 584. [Google Scholar] [CrossRef] [PubMed]
- Fuller, P.J.; Young, M.J. Mechanisms of Mineralocorticoid Action. Hypertension 2005, 46, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.-J.; Yu, L.-J. Oxidative Stress, Molecular Inflammation and Sarcopenia. IJMS 2010, 11, 1509–1526. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.B.; Li, Y.-P. Tumor Necrosis Factor-α and Muscle Wasting: A Cellular Perspective. Respir Res 2001, 2, 269. [Google Scholar] [CrossRef] [PubMed]
- Clavel, S.; Coldefy, A.-S.; Kurkdjian, E.; Salles, J.; Margaritis, I.; Derijard, B. Atrophy-Related Ubiquitin Ligases, Atrogin-1 and MuRF1 Are up-Regulated in Aged Rat Tibialis Anterior Muscle. Mechanisms of Ageing and Development 2006, 127, 794–801. [Google Scholar] [CrossRef]
- Langen, R.C.J.; Velden, J.L.J.; Schols, A.M.W.J.; Kelders, M.C.J.M.; Wouters, E.F.M.; Janssen-Heininger, Y.M.W. Tumor Necrosis Factor-alpha Inhibits Myogenic Differentiation through MyoD Protein Destabilization. FASEB j. 2004, 18, 227–237. [Google Scholar] [CrossRef]
- Chantong, B.; Kratschmar, D.V.; Nashev, L.G.; Balazs, Z.; Odermatt, A. Mineralocorticoid and Glucocorticoid Receptors Differentially Regulate NF-kappaB Activity and pro-Inflammatory Cytokine Production in Murine BV-2 Microglial Cells. J Neuroinflammation 2012, 9, 260. [Google Scholar] [CrossRef]
- Thangaraj, S.S.; Oxlund, C.S.; Fonseca, M.P.D.; Svenningsen, P.; Stubbe, J.; Palarasah, Y.; Ketelhuth, D.F.J.; Jacobsen, Ib.A.; Jensen, B.L. The Mineralocorticoid Receptor Blocker Spironolactone Lowers Plasma Interferon-γ and Interleukin-6 in Patients with Type 2 Diabetes and Treatment-Resistant Hypertension. Journal of Hypertension 2022, 40, 153–162. [Google Scholar] [CrossRef]
- Sønder, S.U.S.; Mikkelsen, M.; Rieneck, K.; Hedegaard, C.J.; Bendtzen, K. Effects of Spironolactone on Human Blood Mononuclear Cells: Mineralocorticoid Receptor Independent Effects on Gene Expression and Late Apoptosis Induction. British J Pharmacology 2006, 148, 46–53. [Google Scholar] [CrossRef]
- Tu, H.; Li, Y.-L. Inflammation Balance in Skeletal Muscle Damage and Repair. Front. Immunol. 2023, 14, 1133355. [Google Scholar] [CrossRef]
- Beenakker, K.G.M.; Ling, C.H.; Meskers, C.G.M.; De Craen, A.J.M.; Stijnen, T.; Westendorp, R.G.J.; Maier, A.B. Patterns of Muscle Strength Loss with Age in the General Population and Patients with a Chronic Inflammatory State. Ageing Research Reviews 2010, 9, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Gomarasca, M.; Banfi, G.; Lombardi, G. Myokines: The Endocrine Coupling of Skeletal Muscle and Bone. In Advances in Clinical Chemistry; Elsevier, 2020; Vol. 94, pp. 155–218 ISBN 978-0-12-820801-4.
- Lightfoot, A.P.; Cooper, R.G. The Role of Myokines in Muscle Health and Disease. Current Opinion in Rheumatology 2016, 28, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Ji, L.L.; Kavazis, A.N.; Jackson, M.J. Reactive Oxygen Species: Impact on Skeletal Muscle. In Comprehensive Physiology; Prakash, Y.S., Ed.; Wiley, 2011; pp. 941–969 ISBN 978-0-470-65071-4.
- Fulle, S.; Protasi, F.; Di Tano, G.; Pietrangelo, T.; Beltramin, A.; Boncompagni, S.; Vecchiet, L.; Fanò, G. The Contribution of Reactive Oxygen Species to Sarcopenia and Muscle Ageing. Experimental Gerontology 2004, 39, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Burniston, J.; Saini, A.; Tan, L.; Goldspink, D. Aldosterone Induces Myocyte Apoptosis in the Heart and Skeletal Muscles of Rats in Vivo. Journal of Molecular and Cellular Cardiology 2005, 39, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Lastra, G.; Whaley-Connell, A.; Manrique, C.; Habibi, J.; Gutweiler, A.A.; Appesh, L.; Hayden, M.R.; Wei, Y.; Ferrario, C.; Sowers, J.R. Low-Dose Spironolactone Reduces Reactive Oxygen Species Generation and Improves Insulin-Stimulated Glucose Transport in Skeletal Muscle in the TG(mRen2)27 Rat. American Journal of Physiology-Endocrinology and Metabolism 2008, 295, E110–E116. [Google Scholar] [CrossRef]
- Rains, J.L.; Jain, S.K. Oxidative Stress, Insulin Signaling, and Diabetes. Free Radical Biology and Medicine 2011, 50, 567–575. [Google Scholar] [CrossRef]
- Lefranc, C.; Friederich-Persson, M.; Palacios-Ramirez, R.; Nguyen Dinh Cat, A. Mitochondrial Oxidative Stress in Obesity: Role of the Mineralocorticoid Receptor. Journal of Endocrinology 2018, 238, R143–R159. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, D.A.; Choi, E.; Lee, Y.S.; Park, S.J.; Kim, B.-J. Aldosterone Inhibits In Vitro Myogenesis by Increasing Intracellular Oxidative Stress via Mineralocorticoid Receptor. Endocrinol Metab 2021, 36, 865–874. [Google Scholar] [CrossRef]
- Lefranc, C.; Friederich-Persson, M.; Braud, L.; Palacios-Ramirez, R.; Karlsson, S.; Boujardine, N.; Motterlini, R.; Jaisser, F.; Nguyen Dinh Cat, A. MR (Mineralocorticoid Receptor) Induces Adipose Tissue Senescence and Mitochondrial Dysfunction Leading to Vascular Dysfunction in Obesity. Hypertension 2019, 73, 458–468. [Google Scholar] [CrossRef]
- Marzetti, E.; Wohlgemuth, S.E.; Lees, H.A.; Chung, H.-Y.; Giovannini, S.; Leeuwenburgh, C. Age-Related Activation of Mitochondrial Caspase-Independent Apoptotic Signaling in Rat Gastrocnemius Muscle. Mechanisms of Ageing and Development 2008, 129, 542–549. [Google Scholar] [CrossRef]
- Lowe, J.; Kadakia, F.K.; Zins, J.G.; Haupt, M.; Peczkowski, K.K.; Rastogi, N.; Floyd, K.T.; Gomez-Sanchez, E.P.; Gomez-Sanchez, C.E.; Elnakish, M.T.; et al. Mineralocorticoid Receptor Antagonists in Muscular Dystrophy Mice During Aging and Exercise. JND 2018, 5, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Pieronne-Deperrois, M.; Guéret, A.; Djerada, Z.; Crochemore, C.; Harouki, N.; Henry, J.; Dumesnil, A.; Larchevêque, M.; Do Rego, J.; Do Rego, J.; et al. Mineralocorticoid Receptor Blockade with Finerenone Improves Heart Function and Exercise Capacity in Ovariectomized Mice. ESC Heart Failure 2021, 8, 1933–1943. [Google Scholar] [CrossRef] [PubMed]
- Wellhoener, P.; Born, J.; Fehm, H.L.; Dodt, C. Elevated Resting and Exercise-Induced Cortisol Levels after Mineralocorticoid Receptor Blockade with Canrenoate in Healthy Humans. The Journal of Clinical Endocrinology & Metabolism 2004, 89, 5048–5052. [Google Scholar] [CrossRef]
- Yoo, S.-Z.; No, M.-H.; Heo, J.-W.; Park, D.-H.; Kang, J.-H.; Kim, S.H.; Kwak, H.-B. Role of Exercise in Age-Related Sarcopenia. J Exerc Rehabil 2018, 14, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Vgontzas, A.N.; Chrousos, G.P. Sleep, the Hypothalamic–Pituitary–Adrenal Axis, and Cytokines: Multiple Interactions and Disturbances in Sleep Disorders. Endocrinology and Metabolism Clinics of North America 2002, 31, 15–36. [Google Scholar] [CrossRef]
- Duclos, M.; Tabarin, A. Exercise and the Hypothalamo-Pituitary-Adrenal Axis. In Frontiers of Hormone Research; Lanfranco, F., Strasburger, C.J., Eds.; S. Karger AG, 2016; Vol. 47, pp. 12–26 ISBN 978-3-318-05868-0.
- Piovezan, R.D.; Abucham, J.; Dos Santos, R.V.T.; Mello, M.T.; Tufik, S.; Poyares, D. The Impact of Sleep on Age-Related Sarcopenia: Possible Connections and Clinical Implications. Ageing Research Reviews 2015, 23, 210–220. [Google Scholar] [CrossRef]
- Valaiyapathi, B.; Calhoun, D.A. Role of Mineralocorticoid Receptors in Obstructive Sleep Apnea and Metabolic Syndrome. Curr Hypertens Rep 2018, 20, 23. [Google Scholar] [CrossRef]
- Dattilo, M.; Antunes, H.K.M.; Medeiros, A.; Mônico Neto, M.; Souza, H.S.; Tufik, S.; De Mello, M.T. Sleep and Muscle Recovery: Endocrinological and Molecular Basis for a New and Promising Hypothesis. Medical Hypotheses 2011, 77, 220–222. [Google Scholar] [CrossRef]
- Dolezal, B.A.; Neufeld, E.V.; Boland, D.M.; Martin, J.L.; Cooper, C.B. Interrelationship between Sleep and Exercise: A Systematic Review. Advances in Preventive Medicine 2017, 2017, 1–14. [Google Scholar] [CrossRef]
- Borghouts, L.B.; Keizer, H.A. Exercise and Insulin Sensitivity: A Review. Int J Sports Med 2000, 21, 1–12. [Google Scholar] [CrossRef]
- Peake, J.M.; Della Gatta, P.; Suzuki, K.; Nieman, D.C. Cytokine Expression and Secretion by Skeletal Muscle Cells: Regulatory Mechanisms and Exercise Effects. Exerc Immunol Rev 2015, 21, 8–25. [Google Scholar] [PubMed]
- Hurst, C.; Robinson, S.M.; Witham, M.D.; Dodds, R.M.; Granic, A.; Buckland, C.; De Biase, S.; Finnegan, S.; Rochester, L.; Skelton, D.A.; et al. Resistance Exercise as a Treatment for Sarcopenia: Prescription and Delivery. Age and Ageing 2022, 51, afac003. [Google Scholar] [CrossRef] [PubMed]
- Rafael-Fortney, J.A.; Chimanji, N.S.; Schill, K.E.; Martin, C.D.; Murray, J.D.; Ganguly, R.; Stangland, J.E.; Tran, T.; Xu, Y.; Canan, B.D.; et al. Early Treatment With Lisinopril and Spironolactone Preserves Cardiac and Skeletal Muscle in Duchenne Muscular Dystrophy Mice. Circulation 2011, 124, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Pitt, B.; Zannad, F.; Remme, W.J.; Cody, R.; Castaigne, A.; Perez, A.; Palensky, J.; Wittes, J. The Effect of Spironolactone on Morbidity and Mortality in Patients with Severe Heart Failure. N Engl J Med 1999, 341, 709–717. [Google Scholar] [CrossRef]
- Spirduso, W.W.; Cronin, D.L. Exercise Dose-Response Effects on Quality of Life and Independent Living in Older Adults: Medicine and Science in Sports and Exercise 2001, 33, S598–S608. [CrossRef]
Figure 1.
The Role of MRs in Skeletal Muscle. Upregulation of MRs induces an increase in TNF-α levels, creating a pro-inflammatory environment that damages muscle tissue and exacerbates catabolic pathways, including the activation of MuRF1 and Atrogin-1. This process is further compounded by mitochondrial dysfunction, which elevates reactive oxygen species (ROS). Additionally, MRs overactivation suppresses myogenic regulatory factors such as MyoD and myogenin, impairing muscle regeneration, while also reducing GLUT4 activity, leading to disrupted glucose metabolism. Conversely, MRs downregulation promotes the maintenance or activation of anabolic pathways, enhances the activity of antioxidant enzymes, and supports glucose metabolism by increasing GLUT4 activity. This balanced state fosters a protective and regenerative environment within skeletal muscle, counteracting the detrimental effects of MRs overactivation and preserving muscle integrity.
Figure 1.
The Role of MRs in Skeletal Muscle. Upregulation of MRs induces an increase in TNF-α levels, creating a pro-inflammatory environment that damages muscle tissue and exacerbates catabolic pathways, including the activation of MuRF1 and Atrogin-1. This process is further compounded by mitochondrial dysfunction, which elevates reactive oxygen species (ROS). Additionally, MRs overactivation suppresses myogenic regulatory factors such as MyoD and myogenin, impairing muscle regeneration, while also reducing GLUT4 activity, leading to disrupted glucose metabolism. Conversely, MRs downregulation promotes the maintenance or activation of anabolic pathways, enhances the activity of antioxidant enzymes, and supports glucose metabolism by increasing GLUT4 activity. This balanced state fosters a protective and regenerative environment within skeletal muscle, counteracting the detrimental effects of MRs overactivation and preserving muscle integrity.
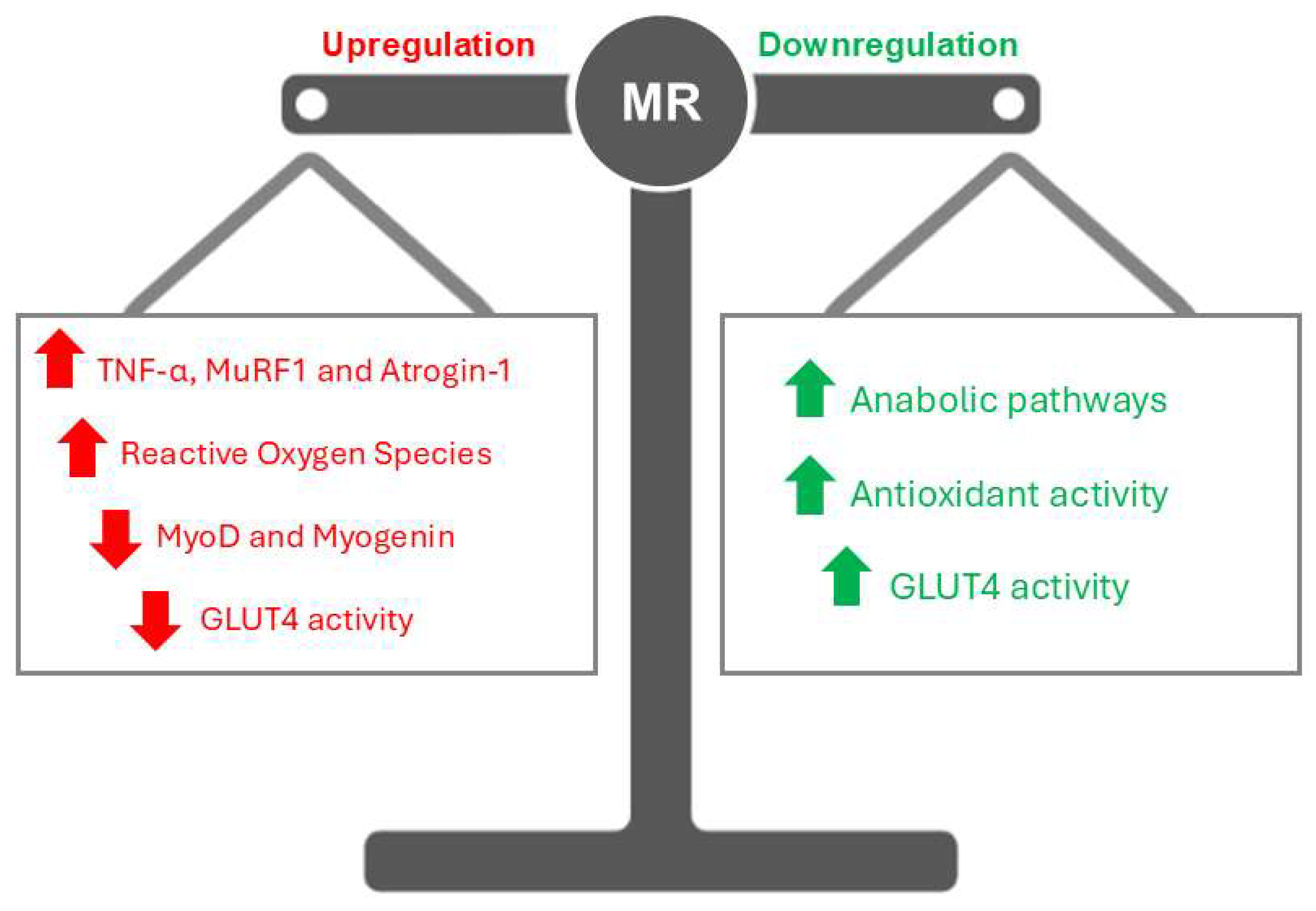
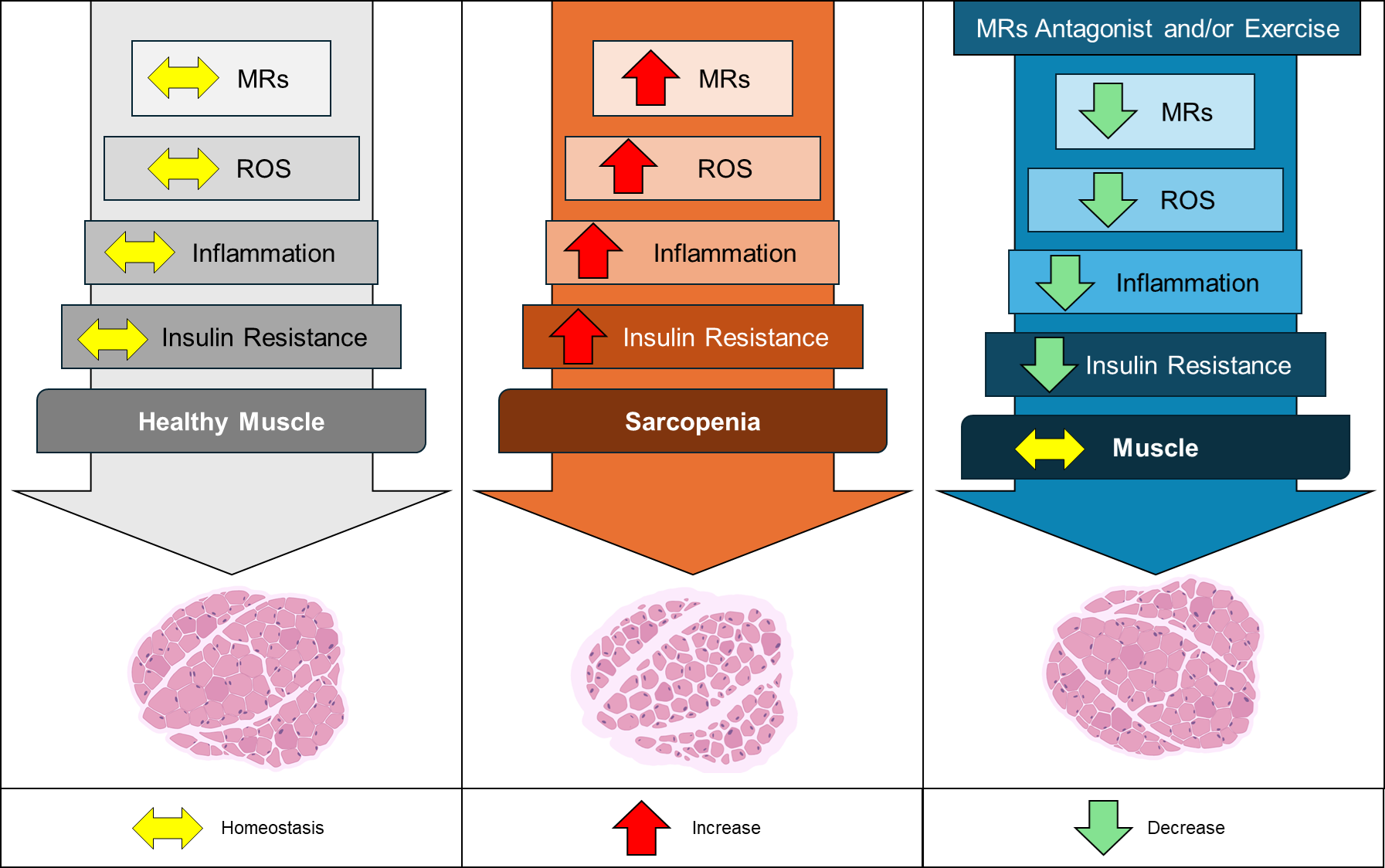
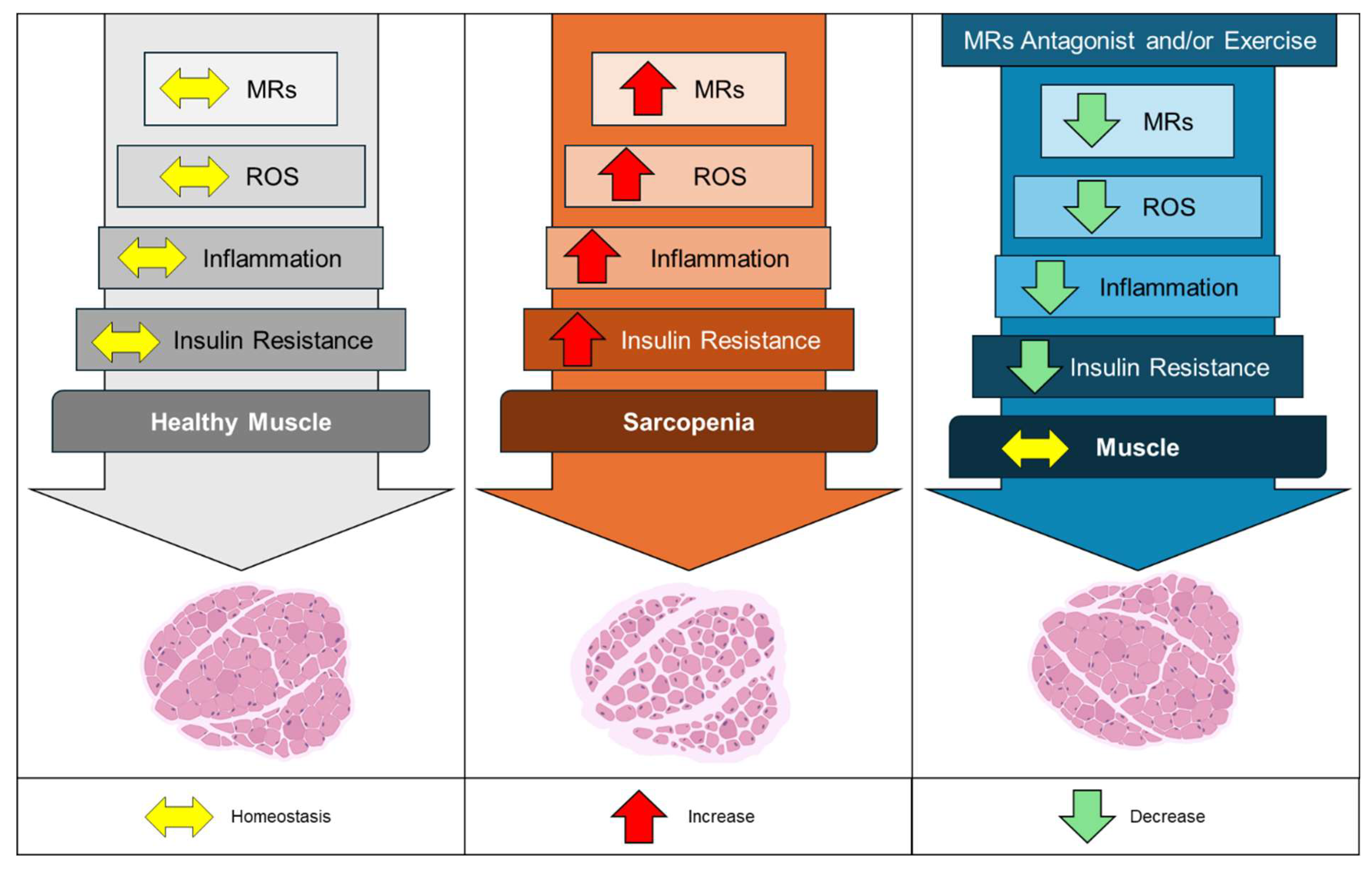
Figure 2.
Effects of MRs in healthy and sarcopenic skeletal muscle. The left panel illustrates a state of homeostasis (yellow arrows) in MRs regulation, mitochondrial reactive oxygen species (ROS) production, inflammatory mediators, and insulin sensitivity, all contributing to healthy muscle function. In the central panel, increased activation and expression of MRs (red arrows) are depicted as driving factors for enhanced mitochondrial ROS generation, elevated muscle inflammation, and insulin resistance. These processes lead to reduced muscle cross-sectional area and diminished muscle strength, culminating in sarcopenia. The right panel demonstrates the therapeutic potential of MRs antagonists, such as spironolactone, as well as regular physical exercise. These interventions reduce (green arrows) MRs activation and expression, decrease ROS levels and inflammatory mediators, and improve insulin sensitivity, thereby restoring muscle homeostasis and promoting healthier muscle function.
Figure 2.
Effects of MRs in healthy and sarcopenic skeletal muscle. The left panel illustrates a state of homeostasis (yellow arrows) in MRs regulation, mitochondrial reactive oxygen species (ROS) production, inflammatory mediators, and insulin sensitivity, all contributing to healthy muscle function. In the central panel, increased activation and expression of MRs (red arrows) are depicted as driving factors for enhanced mitochondrial ROS generation, elevated muscle inflammation, and insulin resistance. These processes lead to reduced muscle cross-sectional area and diminished muscle strength, culminating in sarcopenia. The right panel demonstrates the therapeutic potential of MRs antagonists, such as spironolactone, as well as regular physical exercise. These interventions reduce (green arrows) MRs activation and expression, decrease ROS levels and inflammatory mediators, and improve insulin sensitivity, thereby restoring muscle homeostasis and promoting healthier muscle function.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



