
Submitted:
31 December 2024
Posted:
03 January 2025
You are already at the latest version
Abstract
Objective: Pelleted feed offers several advantages in animal production, yet its hardness can limit intake for young lambs with underdeveloped teeth and digestive systems, particularly when alfalfa inclusion and high compression ratios for small-diameter pellets are involved. This study aimed to evaluate the effects of removing alfalfa from starter feed or post-pelleting crumbling on feed intake, growth, nutrient digestibility, rumen fermentation, and microbial composition in lambs. We hypothesized that crumbling pelleted feed could reduce hardness, thereby increasing intake and enhancing performance.Methods: A total of 117 healthy, 7-day-old Hu lambs (4.02 ± 0.94 kg) were allocated to three groups: (1) a pelleted starter with alfalfa (CON), (2) a non-alfalfa pelleted starter (NA), and (3) a crumbled starter with alfalfa (CA).Results: The CA group exhibited significantly lower pellet hardness compared to CON and NA (P < 0.05), while NA had higher starch gelatinization (P < 0.05). CA notably increased feed intake, particularly after day 21 (P < 0.05), and achieved the highest overall intake and average daily gain (ADG) from days 7–49. Feed type and feed type-sex interactions significantly affected NDF and ADF digestibility, with NA and CA outperforming CON (P < 0.05). Rumen microbiota diversity was influenced by feed composition more than pellet form, with alfalfa inclusion affecting a greater number of microbial genera. Crumbling increased the abundance of Methanobrevibacter (P < 0.05).Conclusion: These findings suggest that post-pelleting crumbling using a roller mill enhances feed intake and growth performance in young lambs, while feed composition plays a predominant role in shaping rumen microbial diversity.
Keywords:
feed technology
; rumen microorganism
; pelleted feeds
; digestibility
; rumen fermentation
; starter
Introduction
Pelleted feed is extensively utilized in the animal feed industry for its ability to reduce ingredient segregation, minimize feed wastage, and enhance palatability and digestibility. In starter diets for lambs, its use has become increasingly common [1,2]. During pelleting, feed ingredients are conditioned under high temperatures for 15 to 20 seconds, which increases starch gelatinization and alters its molecular structure[3] .This process influences fermentation and digestion in the rumen [4]. The feed mixture is then pressed through a die to form cylindrical pellets, which affects their hardness and density [5]. Feed form significantly impacts starch digestion rates in ruminants, with implications for juvenile and adult ruminant health and performance. However, studies on how pelleting affects lamb performance remain limited.
Young lambs’ underdeveloped teeth and digestive systems make pellet hardness a crucial factor in feed intake and digestion. Pellets for young lambs typically have a smaller diameter and are subjected to a higher compression ratio, increasing hardness, which can reduce intake. Starter feeds for lambs often include alfalfa, which improves growth, health, and rumen development, reduces epithelial keratin thickness, and enhances dietary adaptation during weaning [6].However, alfalfa’s high fiber content increases friction during pelleting, producing harder and denser pellets that may be less suitable for young lambs [7]. To address these challenges, producing large pellets using low compression ratio ring dies followed by a post-pelleting crumbling process may improve feed intake in young lambs. This method involves pressing cooled pellets through a roller crumbler and grading sieves to create lamb-appropriate small pellets, reducing fine particles. While commonly used in chick feed production[8], its application to lamb starter feed has not been studied. Recent research highlights the benefits of pelleted feeds on lamb feed intake, growth performance, and rumen fermentation [9], but variations in pelleting processes remain unexplored.
This study aims to evaluate the effects of removing alfalfa from pelleted starter feeds or using post-pelleting crumbling on feed intake, growth performance, nutrient digestibility, rumen fermentation, and microbiota composition in lambs. We hypothesize that post-pelleting crumbling can reduce pellet hardness and density, thereby increasing feed intake, enhancing growth performance, and improving rumen fermentation in young lambs.
Materials and Methods
Ethical Approval
The animal procedures used in this study were reviewed and approved by the Gansu Agricultural University’s Academic Committee, according to guidelines established by the Biological Studies Animal Care and Use Committee of Gansu Province (Approval No. GSAU-Eth-AST-2024-010).
Experimental Treatments, Feeding, and Measurements of Growth Performance
A total of 118 Hu lambs, aged 7 days with similar body conditions and weights (4.02 ± 0.94 kg), were randomly assigned to one of three dietary treatment groups: Control Group (CON, n = 38; 18 males and 20 females): Fed a standard pelleted starter feed containing alfalfa, with a pellet diameter of 4.5 mm. Non-Alfalfa Group (NA, n = 43; 20 males and 23 females): Fed a standard pelleted starter feed without alfalfa, also with a pellet diameter of 4.5 mm. Crumbled Alfalfa Group (CA, n = 37; 18 males and 19 females): Fed a starter feed with the same formulation as the CON group, but processed with a pellet mill using a 6 mm diameter die, followed by crumbling with a roller mill. The particle size of the crumbled starter feed was 4.04 ± 1.45 mm. The composition and nutritional content of the starter feeds are presented in Table 1. All formulations were consistent across the treatments and produced from the same batch of raw materials to minimize variability.
All lambs were allowed free nursing with their mothers after birth. At 7 days of age, they were transferred with their mothers into designated pens (6 m × 4 m) based on their assigned experimental group. Each treatment group was randomly divided into three pens, for a total of nine pens. Pens were equipped with a supplementary feeding area for lambs (1 m × 2 m) where starter feed was provided. This area was designed to restrict access by ewes, ensuring that only lambs could consume the starter feed. Starter feed was supplied daily at 8:00 a.m., and lambs had ad libitum access to nursing, water, and starter feed throughout the experimental period. Ewes were fed the same total mixed ration (TMR) diet, administered twice daily, to ensure uniform maternal nutrition across all groups.
From 7 days of age, lamb body weight, average daily gain (ADG), and starter feed intake were measured every 14 days. The evening before each body weight measurement, all starter feed was removed from the troughs to ensure fasting, and lambs were weighed the following morning in a fasted state. Starter feed intake was calculated as the difference between the amount of feed offered and the feed remaining. Feed offered and refusals were recorded daily to ensure accurate measurements.
Determination of Physical and Chemical Properties of Pellet Feed
For each feed group, three samples of approximately 500 g were collected to evaluate hardness, density, and starch gelatinization. Twenty pellets of uniform length and intact appearance were selected using a quartering method. These pellets were compressed along their diameters until fracture using a hardness tester (YHKC-2A, Taizhou Galaxy Instruments Co.). The mean pressure required for breakage was recorded as the hardness value for each group. A 100 g feed sample was placed into a graduated syringe, and the scale was recorded. A vacuum was applied to measure the inter-pellet void volume. The total pellet volume was then calculated, and density was derived using the formula: ρ=m/V, where ρ is the density (g/cm³), m is the mass of the sample (g), and V is the volume (cm³). Starch gelatinization was measured following the method described by Xiong [10], ensuring consistency with previously established procedures.
Sample Collection and Measurement of Nutrient Digestion
Eighteen lambs (6 from each feed group, with 3 males and 3 females per group) were selected for fecal sample collection. Following weaning at 49 days of age and a 5-day transition period, fecal samples were collected twice daily over four consecutive days using rectal collection. The samples were pooled for each lamb. A portion of the pooled fecal samples was treated with 10% sulfuric acid for nitrogen fixation, followed by crude protein (CP) analysis. The remaining samples were sealed in plastic bags, dried at 65°C, and later used to determine dry matter (DM), neutral detergent fiber (NDF), acid detergent fiber (ADF), and acid-insoluble ash content. Acid-insoluble ash was employed as an internal marker to calculate the apparent digestibility of CP, DM, NDF, ADF, and organic matter (OM). The starter feed and fecal samples were analyzed for: DM: Determined by drying samples at 105°C. CP: Analyzed according to AOAC International (2000) standards [11]. Acid-insoluble ash: Determined using a previously described method [12]. NDF and ADF: Analyzed using a previously described method incorporating heat-stable alpha-amylase and sodium sulfate in the NDF procedure [13]. The apparent digestibility of DM, OM, CP, NDF, and ADF was calculated using the following equation:
Nutrient apparent digestibility (%) = 100%−(acid-insoluble ash content in feces / acid-insoluble ash content in feed × nutrient content in feed / nutrient content in feces) × 100% [12].
Collection of Rumen Fluid and Measurement of Volatile Fatty Acids (VFAs)
At 49 days of age, 50 mL of rumen fluid was collected from each lamb using an oral rumen tube in the morning while the animals were fasting. The collected rumen fluid was immediately transferred into cryovials and frozen at -20°C for subsequent analysis of fermentation parameters and microbial sequencing.
The concentrations of VFAs in the rumen fluid were determined using gas chromatography [14]. The rumen fluid samples were pretreated by filtration through a 0.45 μm disposable filter to remove particulates. The clear supernatant was then transferred to vials for gas chromatography (GC) analysis. VFA concentrations were quantified using a Shimadzu gas chromatograph (GC-2010Plus, Japan) with 2-ethylbutyric acid (2-EB) as the internal standard.
Extraction of Microbial DNA from Rumen Fluid and High-Throughput Sequencing
Total DNA was extracted from rumen fluid using the Omega E.Z.N.A. Stool DNA Kit (Omega Bio-Tek, Winooski, VT, United States). The quality and concentration of the extracted DNA were assessed using a NanoDrop 2000 spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, United States). DNA was diluted to a concentration of 50 ng/μL for downstream applications.
For microbial community analysis, the V3–V4 regions of the 16S rRNA gene were amplified using universal primers 341-F (5’-CCTAYGGGRBGCASCAG-3’) and 806-R (5’-GGACTACNNGGGTATCTAAT-3’). The PCR conditions were as follows: an initial denaturation at 98°C for 1 minute, followed by 30 cycles of denaturation at 98°C for 10 seconds, annealing at 50°C for 30 seconds, and elongation at 72°C for 30 seconds, with a final extension at 72°C for 5 minutes. Barcoded amplicons were pooled in equimolar amounts for Illumina paired-end library preparation. Sequencing was performed on an Illumina NovaSeq 6000 platform (San Diego, CA, United States), producing 250 bp paired-end reads.
Paired-end reads were merged using FLASH (v1.2.11) to create raw tags. Tags were quality-filtered using the QIIME toolkit, resulting in high-quality sequences termed “Effective Tags.” These sequences were denoised with the DADA2 or deblur modules in QIIME2 (version 202202), generating Amplicon Sequence Variants (ASVs). Taxonomic assignment was performed using the SILVA database within mothur, with a confidence threshold of 0.80.
Metrics including Chao1, Shannon, Simpson, ACE, and Observed Species indices were calculated to evaluate within-sample species diversity using QIIME2. Differences in microbial communities across samples were analyzed using Principal Coordinates Analysis (PCoA) and cluster analysis, performed in QIIME (version 1.8.0).
Statistical Analysis
The data were analyzed using SPSS (version 26.0, IBM Corp., Armonk, NY, USA). A two-way analysis of variance (ANOVA) within the general linear model (GLM) framework was applied to assess the main effects of starter feed type and sex, as well as their interactions, on growth performance, nutrient digestion, and rumen fermentation indices. Post hoc multiple comparisons were conducted using the least significant difference (LSD) method. Statistical significance was defined as P < 0.05, while values within the range of 0.05 < P < 0.1 were interpreted as indicating a trend toward significance.
Results
Physical Properties and Starch Gelatinization of Starter Feeds with Different Forms
As shown in Table 2, the hardness of the pelleted feed in the CA group was significantly lower compared to the CON and NA groups (P < 0.05). The degree of starch gelatinization in the NA group was significantly higher than that in the CON and CA groups (P < 0.05). However, no significant differences were observed in the density of the feeds among the three groups (P > 0.05).
Effects of Different Starter Feeds on Lamb Growth Performance
As presented in Table 3, the interaction between feed type and sex did not significantly affect growth performance (P > 0.05). However, feed type significantly influenced the average daily gain (ADG) of lambs during the periods of 7–21 days, 35–49 days, and 7–49 days (P < 0.05). Specifically: From 7–21 days, the ADG of the CA group was significantly lower than that of the CON and NA groups. From 35–49 days, the ADG of the CA group was significantly higher than that of the other two groups. Overall, the ADG of the CA group from 7–49 days was significantly higher than that of the NA group (P < 0.05). At 49 days, the body weight (BW) of the CA group tended to be higher than that of the CON and NA groups (P < 0.1). Sex significantly affected BW at all stages and ADG before 21 days, with males showing higher values than females (P < 0.05).
Effects of Different Starter Feeds on Lamb Starter Feed Intake
As presented in Table 4, feed intake was significantly greater in the CA group compared to the CON and NA groups across all stages (P < 0.05), with a pronounced increase after 21 days, where the intake of the CA group substantially exceeded that of the other two groups (P < 0.05).
Effects of Different Starter Feeds on Nutrient Digestibility in Lambs
As shown in Table 5, feed type and the interaction between feed type and sex significantly influenced the digestibility of neutral detergent fiber (NDF) and acid detergent fiber (ADF). The NDF and ADF digestibility in the NA and CA groups were significantly higher than those in the CON group (P < 0.05). The digestibility of crude protein (CP) in the CA group showed a trend toward being higher compared to the CON and NA groups (P < 0.1). Sex significantly affected CP digestibility, with male lambs exhibiting higher values than female lambs (P < 0.05). However, sex did not significantly affect the digestibility of other nutrients (P > 0.05). Feed type, sex, and their interaction did not significantly influence the digestibility of dry matter (DM) or organic matter (OM) (P > 0.05).
Effects of Different Starter Feeds on Rumen Fermentation in Lambs
As shown in Table 6, the interaction between feed type and sex did not significantly affect the concentration of any volatile fatty acids (VFAs) in lambs (P > 0.05). Feed type also did not significantly influence total acid concentration or the concentrations of most VFAs (P > 0.05), although it significantly affected the concentration of propionic acid (P < 0.05) and showed a trend toward influencing valeric acid concentration (P < 0.1). Specifically, the NA group exhibited lower concentrations of propionic and valeric acids compared to the CON group (P < 0.05). Sex significantly influenced the concentrations of propionic acid and isobutyric acid, with male lambs displaying lower levels than female lambs (P < 0.05).
Effects of Different Starter Feeds on Rumen Microbial Communities in Lambs 16S rDNA Sequencing Results of Rumen Microbes
An average of 105,235 rRNA gene reads were obtained from the rumen fluid samples across all groups. Analysis using QIIME2 software identified 31 phyla, and at the genus level, sequences were classified into 346 different genera. As shown in Figure 1A, a total of 4,057 amplicon sequence variants (ASVs) were identified collectively in the NAF, CF, and CAF groups. Among these, 971 ASVs were shared between the NAF and CF groups, 951 ASVs were shared between the NAF and CAF groups, and 832 ASVs were shared between the CAF and CF groups. Additionally, 596 ASVs were common to all three groups. Figure 1B illustrates the ASV distribution in the NAM, CM, and CAM groups, where a total of 5,201 ASVs were identified. The NAM and CM groups shared 1,148 ASVs, the NAM and CAM groups shared 1,487 ASVs, and the CAM and CM groups shared 1,149 ASVs. A total of 834 ASVs were common to all three groups.
Effects of Different Starter Feeds on the Microbial Diversity Indices in the Rumen of Lambs
As shown in Figure 2, different starter feeds significantly influenced the alpha diversity indices of rumen microbes in lambs. In Figure 2A, the NAM group exhibited a significantly higher Chao1 index compared to the CF and CAF groups (P < 0.01).
In Figure 2B, the Observed Features index for the NAM group was also significantly higher than that of the CF and CAF groups (P < 0.01). In Figure 2C, the Shannon index of the NAM group was significantly greater than that of the CF and CAF groups (P < 0.01). Additionally, the NAF group showed significantly higher values than the CF and CAF groups (P < 0.05). In Figure 2D, the Simpson index for the NAM group was significantly higher than that of the CF and CAF groups (P < 0.05), while the NAF group also showed significantly higher values compared to the CF and CAF groups (P < 0.05).
As shown in Figure 3, PCoA analysis based on weighted UniFrac distances revealed that the first two principal coordinates explained 36.9% and 15% of the variance, respectively. In contrast, PCoA analysis using unweighted UniFrac distances indicated that the first two principal coordinates accounted for 13.89% and 6.61% of the variance. The lack of distinct clustering among different groups suggests that the various starter feeds did not significantly impact the beta diversity of rumen microbes in lambs
Effects of Different Starter Feeds on the Taxonomic Abundance of Rumen Microbiota in Lambs
As shown in Figure 4, the overall structure of the rumen microbial community was similar across all groups. At the phylum level, Firmicutes and Bacteroidota were the dominant phyla in all groups, with relative abundances exceeding 30%. At the genus level, the relative abundances of Christensenellaceae R-7 group and Anaerovorax were significantly higher in the NAM group compared to the CM group (P<0.05). The CAF group had a significantly higher abundance of Methanobrevibacter than the CF group (P<0.05). Additionally, the relative abundance of Saccharofermentans in the NAF group was significantly higher than in the CA group (P<0.05). Rikenellaceae RC9 gut group was more abundant in the NAF group compared to the CF group (P<0.05).
Discussion
The high-temperature conditioning process during feed pelleting is well-recognized for enhancing palatability and digestibility, which is why pelleted feed is a common choice for commercial lamb starter diets. However, the suitability of pelleted feed’s physical characteristics for lambs remains insufficiently explored. Previous research indicates that lambs fed pelleted starters exhibit lower feed intake and average daily gain (ADG) compared to those fed textured starter feeds [15]. Similarly, studies in post-weaning lambs and calves show reduced ADG and dry matter intake [DMI] with pelleted feeds compared to powdered or coarser feed forms [16,17,18]. These findings suggest that pelleted feed does not universally enhance growth performance in young ruminants.
The reduced intake of pelleted feed by lambs may be associated with its hardness. During the processing of pelleted feed, achieving a specific hardness level is crucial, as insufficient hardness can compromise pellet formation rates and durability [19]. This is particularly significant for starter feeds containing alfalfa, as the fiber content in alfalfa increases friction during the pelleting process. While no current studies directly examine the effects of pellet hardness on lamb growth performance, higher hardness may negatively impact feed intake. In this study, removing alfalfa from the feed formulation resulted in a reduction in pellet hardness, though the difference was not statistically significant. Additionally, crumbling the pellets after pelleting substantially decreased their hardness and reduced their density, providing further support for the hypothesis.
This study notably found that feeding lambs crumbled pellets significantly increased their feed intake. Consistent with previous research, the feed intake of all lamb groups increased linearly with age, with particularly rapid growth observed after 21 days of age. In this experiment, the increase in feed intake for the crumbled pellet group after 21 days was significantly higher than that of the control group, with intake during the 21–35 day and 35–49 day periods being 2.30 times and 1.74 times greater, respectively. While limited research exists on the effects of starter feed type on lamb intake and growth, numerous studies on calves indicate that starter feed type significantly influences feed intake and growth [20]. For instance, calves tend to consume more when fed multiple small pellets or powdered feeds compared to standard pellets [21]. Though the use of crumbled pellets in ruminant diets has not been extensively studied, research in poultry has shown that chicks fed crumbled pellets achieved higher weights between 15 and 42 days of age compared to those fed whole pellets [22]. Similarly, studies in broilers during the growing phase reported that weight gain and feed-to-meat ratios were significantly improved with pellets and crumbled pellets compared to meal [23]. In this study, the substantial increase in feed intake for the crumbled pellet group (CA group) led to significantly higher average daily gain (ADG) during the 35—49 day period compared to both the control and NA groups. Crumbled pellets maintain the advantages of pelleted feeds—such as high-temperature conditioning, reduced ingredient separation, and minimized feed waste—while also reducing hardness and particle size. These characteristics likely contributed to the significant increase in feed intake observed in lambs.
In this study, although the hardness of alfalfa-free pellets was lower than that of the control group (with no significant difference), the starch gelatinization degree was significantly higher in the alfalfa-free group. However, the lambs fed alfalfa-free pellets exhibited significantly lower feed intake compared to the control group. This suggests that including alfalfa meal in starter feeds for young lambs benefits feed intake and development, consistent with findings from numerous studies[24]. For example, research on Tibetan sheep demonstrated that adding alfalfa to the diet significantly increased final body weight (P < 0.05) [25]. Previous studies have shown that alfalfa stimulates the proliferation of cellulose-decomposing bacteria in the rumen, promotes the presence of glycolytic bacteria, and increases the production of short-chain fatty acids. These changes alter the rumen fermentation pattern, enhancing the animal’s utilization of fermentation end products and ultimately improving growth performance [26]. The effectiveness of roughage supplementation in promoting the healthy development of young ruminants’ rumen depends on factors such as roughage type, fiber length, and supplementation method [27]. Alfalfa, with its high-quality fiber content, effectively stimulates rumen development. Moreover, alfalfa inclusion increases the concentration of β-hydroxybutyrate (BHBA) in the blood, which may reflect accelerated rumen development, as BHBA levels are closely linked to the metabolic development of the rumen wall and fiber digestion rates [28]. Other research has shown that starter feeds containing straw improve feed intake in calves compared to feeds without straw [29]. These findings underscore that simply removing alfalfa hay from feed formulations to address challenges such as increased friction during pelleting or reduced starch gelatinization after steam absorption is not a viable solution. Instead, post-pelleting crumbling of the pellets offers a more effective approach to improving the feeding outcomes of pelleted feeds.
This study further evaluated the impact of different starter feed types on the nutrient digestibility of lambs. Digestibility is influenced by various factors, including the physical form and composition of the feed. Research indicates that most starter feeds in lamb feeding practices are typically pelleted [30]. However, crumbled feeds or feeds with smaller particle sizes have demonstrated better nutrient digestibility compared to larger pellets [31]. Moreover, incorporating alfalfa into starter feeds has been shown to enhance the digestibility of ADF and NDF [32]. The NDF and ADF digestibility in the NA group (standard pelleted feed without alfalfa) was significantly higher than in the CON group (standard pelleted feed with alfalfa). This difference may be attributed to the higher degree of starch gelatinization observed in the NA group. Previous research has shown that increasing the conditioning time during pelleting enhances starch gelatinization, subsequently improving the digestibility of dry matter and starch in calves [19]. Furthermore, the lower feed intake observed in the NA group may have contributed to the increased digestibility, as studies suggest that higher feed intake reduces retention time in the rumen, which can decrease nutrient digestibility. Interestingly, despite significantly higher feed intake in the CA group (crumbled pellets) compared to the CON group, the digestibility of NDF and ADF remained elevated in the CA group. This finding suggests that crumbled pellet feeds are easier for young lambs to digest and absorb. The physical structure of crumbled pellets appears to be more suitable for the underdeveloped digestive systems of young lambs. Additionally, increased feed intake may promote the physical growth and metabolic functions of the rumen, thereby enhancing fiber digestibility. Supporting this notion, related studies have shown that increased intake of solid feeds contributes to the growth in weight and volume of the rumen, enhancing its metabolic capacity[33].
This experiment further evaluated the effects of different types of starter feeds on the rumen fermentation function of lambs by analyzing volatile fatty acid (VFA) concentrations in rumen fluid. Among the VFAs, the type of starter feed significantly influenced only the propionate concentration, with no significant effects observed on total acid levels, other VFA concentrations, or the acetate-to-propionate ratio. While some studies suggest that concentrates generate higher propionate levels during fermentation [34], the NA group in this study exhibited lower propionate levels, likely due to reduced feed intake. The concentration of VFAs in rumen fluid reflects the balance between microbial fermentation and VFA absorption in the rumen. Other studies have similarly reported that young lambs supplemented with alfalfa exhibited lower propionate concentrations during the transitional period post-weaning [24]. The findings of this study indicate that variations in starter feed types and feed intake have minimal impact on the overall rumen fermentation pattern. Consistent with these results, other research emphasizes that the composition of the diet is the primary determinant of rumen fermentation type, whereas the physical form of the feed exerts minimal influence on fermentation parameters in sheep [35].
Numerous studies have demonstrated that the physical form of feed influences the rumen microbiota in lambs. For example, Liu et al. reported that pelleted feed, compared to mash, affects phenotypic traits such as meat quality by modulating the rumen microbiota [36]. In light of this, we further investigated the effects of post-pelleting crumbling on the rumen microbiota of lambs. Our findings indicate that lamb sex influenced alpha diversity indices of the rumen microbiota. However, within the same sex, only diet composition—not post-pelleting crumbling—affected microbial diversity. Additionally, no significant differences in beta diversity were observed among the experimental groups. Similarly, Liu et al. found no differences in the alpha diversity index of rumen microbiota between lambs fed pelleted versus non-pelleted feeds, aligning with the results of this study. These findings suggest that diet composition plays a predominant role in modulating the rumen microbiota, while the physical form of feed, such as crumbling, has a minimal impact [37].
In terms of microbial composition, the dominant phyla across all groups were Firmicutes and Bacteroidota, with Prevotella being the dominant genus, consistent with previous studies [38]. This study found that pelleting or diet composition influenced the relative abundance of specific phyla and genera. Notably, the microbial groups that differed in male lambs were distinct from those in female lambs. This finding underscores the critical role of animal sex in shaping rumen microbiota and suggests that microbial responses to diet may vary between males and females. Consequently, the inclusion of both sexes in dietary and microbiome studies is vital to obtain a comprehensive understanding of microbial dynamics and dietary impacts, ensuring the broad applicability of findings across populations. Compared to physical feed form, alfalfa inclusion had a more significant influence on a broader range of microbial genera, including the Christensenellaceae R-7 group, Anaerovorax, Saccharofermentans, and Rikenellaceae RC9 gut group, all of which exhibited higher relative abundances in the non-alfalfa group. These results align with other studies, which have similarly reported increased abundances of the Christensenellaceae R-7 group, Saccharofermentans, and Rikenellaceae RC9 gut group in high-concentrate diets. This suggests that these genera are sensitive to the forage-to-concentrate ratio, with the chemical composition of high-concentrate diets creating specific niches that favor their proliferation.
Our study also found that post-pelleting crumbling increased the abundance of Methanobrevibacter. Numerous studies examining the relationship between feed efficiency and rumen microbiota in ruminants have reported an association between Methanobrevibacter abundance and feed conversion efficiency, with higher Methanobrevibacter abundance often observed in low feed-efficiency groups [39] A consistent observation across these studies is that increased feed intake correlates with greater Methanobrevibacter abundance. In this study, the post-pelleting crumbling group exhibited a notable increase in feed intake accompanied by a significant rise in Methanobrevibacter abundance, aligning with these findings. Similarly, research on rabbits demonstrated that smaller pellet sizes enhanced both feed intake and cecum Methanobrevibacter abundance [40], further supporting our results. These findings underscore the role of feed intake as a key factor influencing Methanobrevibacter abundance in the rumen. While the physical form of feed, particularly post-pelleting crumbling, did not significantly affect microbial diversity indices, it did influence the abundance of specific microbial groups. This study suggests that the physical form of feed can indirectly affect microbial composition by altering intake behavior.
Conclusion
This study demonstrates that crumbling pelleted feed significantly improves feed intake and growth performance in young lambs by reducing pellet hardness and enhancing fiber digestibility. Although feed composition exerted a greater influence on rumen microbial diversity than feed form, crumbling specifically increased the abundance of Methanobrevibacter, highlighting its potential to affect microbial composition through altered intake behavior. These findings suggest that crumbled starter feed is a practical and effective strategy for promoting feed intake, nutrient digestibility, and growth performance in lambs.
Funding
This work was supported by the Central Government’s Guiding Fund for Local Science and Technology Development under grant no. 23ZYQC0304, and the Science and Technology Project of Gansu Province - Special Project for Cooperation between the Eastern and Western Regions under grant no. 22CX8NA044, and Discipline Team Project of Gansu Agricultural University under grant no. GAU-XKTD-2022-20.
Acknowledgements
We would like to thank the native English-speaking scientists of Elixigen Company (Huntington Beach, California) for editing our manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Khan MA, Bach A, Weary DM, von Keyserlingk MAG. Invited review: Transitioning from milk to solid feed in dairy heifers. J Dairy Sci 2016,99, 885-902. [CrossRef]
- Adiwinarti R, Rianto E, Purbowati E, Restitrisnani V, Purnomoadi A. Comparative feed management system in sheep fed different physical forms of ration containing Ipomoea aquatica on the performance, rumen characteristics, and chewing activity. J Adv Vet Anim Res 2023,10:677-84. [CrossRef]
- Fahrenholz, AC. Evaluating factors affecting pellet durability and energy consumption in a pilot feed mill and comparing methods for evaluating pellet durability: Kansas State University; 2012.
- Abdollahi MR, Ravindran V, Wester TJ, Ravindran G, Thomas DV. Effect of improved pellet quality from the addition of a pellet binder and/or moisture to a wheat-based diet conditioned at two different temperatures on performance, apparent metabolisable energy and ileal digestibility of starch and nitrogen in broilers. Animal Feed Science and Technology 2012,175:150-7. [CrossRef]
- Svihus B, Kløvstad KH, Perez V, et al. Physical and nutritional effects of pelleting of broiler chicken diets made from wheat ground to different coarsenesses by the use of roller mill and hammer mill. Animal Feed Science and Technology 2004,117:281-93. [CrossRef]
- Li K, Du H, Guo W, Na M, Na R. Initial timing of alfalfa hay supplementation manipulates blood parameters, rumen gene expression, and epithelial microbiota in pre-weaning lambs. J Anim Sci 2024. [CrossRef]
- Buchanan NP, Moritz JS. Main effects and interactions of varying formulation protein, fiber, and moisture on feed manufacture and pellet quality. Journal of Applied Poultry Research 2009,18:274-83. [CrossRef]
- Lv M, Yan L, Wang Z, et al. Effects of feed form and feed particle size on growth performance, carcass characteristics and digestive tract development of broilers. Anim Nutr 2015,1:252-6. [CrossRef]
- Liu T, Li F, Wang W, et al. Early feeding strategies in lambs affect rumen development and growth performance, with advantages persisting for two weeks after the transition to fattening diets. Front Vet Sci 2022,9:925649. [CrossRef]
- Xiong Y, Bartle SJ, Preston RL. Improved enzymatic method to measure processing effects and starch availability in sorghum grain. J Anim Sci 1990,68:3861-70. [CrossRef]
- van Heugten E, Funderburke DW, Dorton KL. Growth performance, nutrient digestibility, and fecal microflora in weanling pigs fed live yeast. J Anim Sci 2003,81:1004-12. [CrossRef]
- Van Keulen J, Young B. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. Journal of animal science 1977, 44:282-7.
- Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 1991, 74:3583-97. [CrossRef]
- Zhang YK, Zhang XX, Li FD, et al. Characterization of the rumen microbiota and its relationship with residual feed intake in sheep. Animal 2021, 15:100161. [CrossRef]
- Li Y, Guo YL, Zhang CX, et al. Effects of physical forms of starter feed on growth, nutrient digestibility, gastrointestinal enzyme activity, and morphology of pre- and post-weaning lambs. Animal 2021, 15:100044. [CrossRef]
- Li B, Sun X, Huo Q, et al. Pelleting of a Total Mixed Ration Affects Growth Performance of Fattening Lambs. Front Vet Sci 2021, 8:629016. [CrossRef]
- Franklin ST, Amaral-Phillips DM, Jackson JA, Campbell AA. Health and performance of Holstein calves that suckled or were hand-fed colostrum and were fed one of three physical forms of starter. J Dairy Sci 2003, 86:2145-53. [CrossRef]
- Porter J, Warner R, Kertz A. Effect of fiber level and physical form of starter on growth and development of dairy calves fed no forage. The Professional Animal Scientist 2007, 23:395-400.
- Soltani E, Naserian AA, Khan MA, Ghaffari MH, Malekkhahi M. Effects of conditioner retention time during pelleting of starter feed on nutrient digestibility, ruminal fermentation, blood metabolites, and performance of Holstein female dairy calves. J Dairy Sci 2020, 103:8910-21. [CrossRef]
- Coverdale JA, Tyler HD, Quigley JD, 3rd, Brumm JA. Effect of various levels of forage and form of diet on rumen development and growth in calves. J Dairy Sci 2004, 87:2554-62. [CrossRef]
- Bach A, Giménez A, Juaristi JL, Ahedo J. Effects of physical form of a starter for dairy replacement calves on feed intake and performance. J Dairy Sci 2007, 90:3028-33. [CrossRef]
- Lang W, Hong P, Li R, et al. Growth performance and intestinal morphology of Hyline chickens fed diets with different diet particle sizes. J Anim Physiol Anim Nutr (Berl) 2019, 103:518-24. [CrossRef]
- Amoozmehr A, Dastar B, Ashayerizadeh O, Mirshekar R, Abdollahi MR. Effect of feed form and nutrient density on growth performance, blood parameters, and intestinal traits in broiler breeder pullets. Poult Sci 2023, 102:102700. [CrossRef]
- Yang B, He B, Wang SS, Liu JX, Wang JK. Early supplementation of starter pellets with alfalfa improves the performance of pre- and postweaning Hu lambs. J Anim Sci 2015, 93:4984-94. [CrossRef]
- Su Y, Sun X, Zhao S, et al. Dietary alfalfa powder supplementation improves growth and development, body health, and meat quality of Tibetan sheep. Food Chem 2022, 396:133709. [CrossRef]
- Yang B, Le J, Wu P, et al. Alfalfa Intervention Alters Rumen Microbial Community Development in Hu Lambs During Early Life. Front Microbiol 2018, 9:574. [CrossRef]
- Castells L, Bach A, Aris A, Terré M. Effects of forage provision to young calves on rumen fermentation and development of the gastrointestinal tract. J Dairy Sci 2013, 96:5226-36. [CrossRef]
- Khan MA, Weary DM, von Keyserlingk MA. Hay intake improves performance and rumen development of calves fed higher quantities of milk. J Dairy Sci 2011, 94:3547-53. [CrossRef]
- Terré M, Castells L, Khan MA, Bach A. Interaction between the physical form of the starter feed and straw provision on growth performance of Holstein calves. J Dairy Sci 2015, 98:1101-9. [CrossRef]
- Karimizadeh E, Chaji M, Mohammadabadi T. Effects of physical form of diet on nutrient digestibility, rumen fermentation, rumination, growth performance and protozoa population of finishing lambs. Anim Nutr 2017, 3:139-44. [CrossRef]
- Mirmohammadi D, Rouzbehan Y, Fazaeli H. The effect of the inclusion of recycled poultry bedding and the physical form of diet on the performance, ruminal fermentation, and plasma metabolites of fattening lambs. J Anim Sci 2015, 93:3843-53. [CrossRef]
- Abouheif MA, Al-Saiady MY, Al-Mufarrej SI, et al. Effect of physical form of diet and frequency of feeding on digesta retention time and digestion in Najdi lambs. 2012.
- Norouzian MA, Valizadeh R, Vahmani P. Rumen development and growth of Balouchi lambs offered alfalfa hay pre- and post-weaning. Trop Anim Health Prod 2011, 43:1169-74. [CrossRef]
- van Ackeren C, Steingaß H, Hartung K, Funk R, Drochner W. Effect of roughage level in a total mixed ration on feed intake, ruminal fermentation patterns and chewing activity of early-weaned calves with ad libitum access to grass hay. Animal Feed Science and Technology 2009, 153:48-59. [CrossRef]
- Trabi EB, Seddik H-e, Xie F, Lin L, Mao S. Comparison of the rumen bacterial community, rumen fermentation and growth performance of fattening lambs fed low-grain, pelleted or non-pelleted high grain total mixed ration. Animal Feed Science and Technology 2019, 253:1-12.
- Liu T, Bu Z, Xiang K, Jia Y, Du S. Effects of non-pelleted or pelleted low-native grass and pelleted high-native grass diets on meat quality by regulating the rumen microbiota in lambs. Microbiol Spectr 2024, 12:e0375823. [CrossRef]
- Palmonari A, Federiconi A, Formigoni A. Animal board invited review: The effect of diet on rumen microbial composition in dairy cows. Animal 2024, 18:101319. [CrossRef]
- Stevenson DM, Weimer PJ. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl Microbiol Biotechnol 2007, 75:165-74. [CrossRef]
- Lopes DRG, de Souza Duarte M, La Reau AJ, et al. Assessing the relationship between the rumen microbiota and feed efficiency in Nellore steers. J Anim Sci Biotechnol 2021, 12:79. [CrossRef]
- Liu S, Yuan M, Jin D, et al. Effects of the particle of ground alfalfa hay on the growth performance, methane production and archaeal populations of rabbits. PLoS One 2018, 13:e0203393. [CrossRef]
Figure 1.
Relationship between different starter feeds and rumen microbial diversity in lambs. Differences in amplicon sequence variants (ASVs) were assessed among the groups. CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs. CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female; M: male.
Figure 1.
Relationship between different starter feeds and rumen microbial diversity in lambs. Differences in amplicon sequence variants (ASVs) were assessed among the groups. CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs. CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female; M: male.
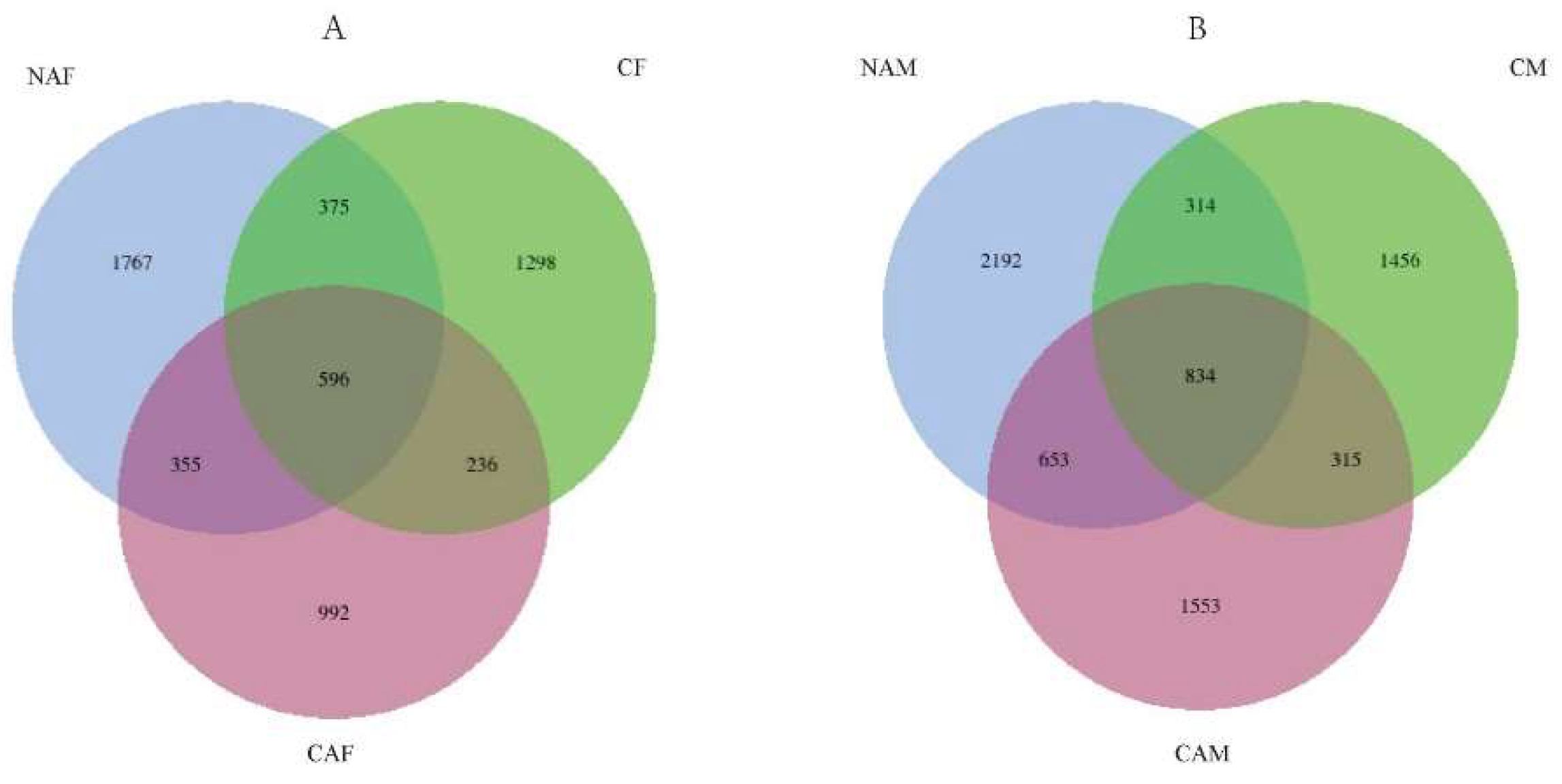
Figure 2.
Rumen microbial diversity indices for lambs fed different starter feeds. Diversity indices include the Chao1 index (A), Observed_features index (B), Shannon index (C), and Simpson index (D). CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs. CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female; M: male. Statistical significance is indicated as follows: P < 0.01 (**), P < 0.05 (*).
Figure 2.
Rumen microbial diversity indices for lambs fed different starter feeds. Diversity indices include the Chao1 index (A), Observed_features index (B), Shannon index (C), and Simpson index (D). CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs. CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female; M: male. Statistical significance is indicated as follows: P < 0.01 (**), P < 0.05 (*).
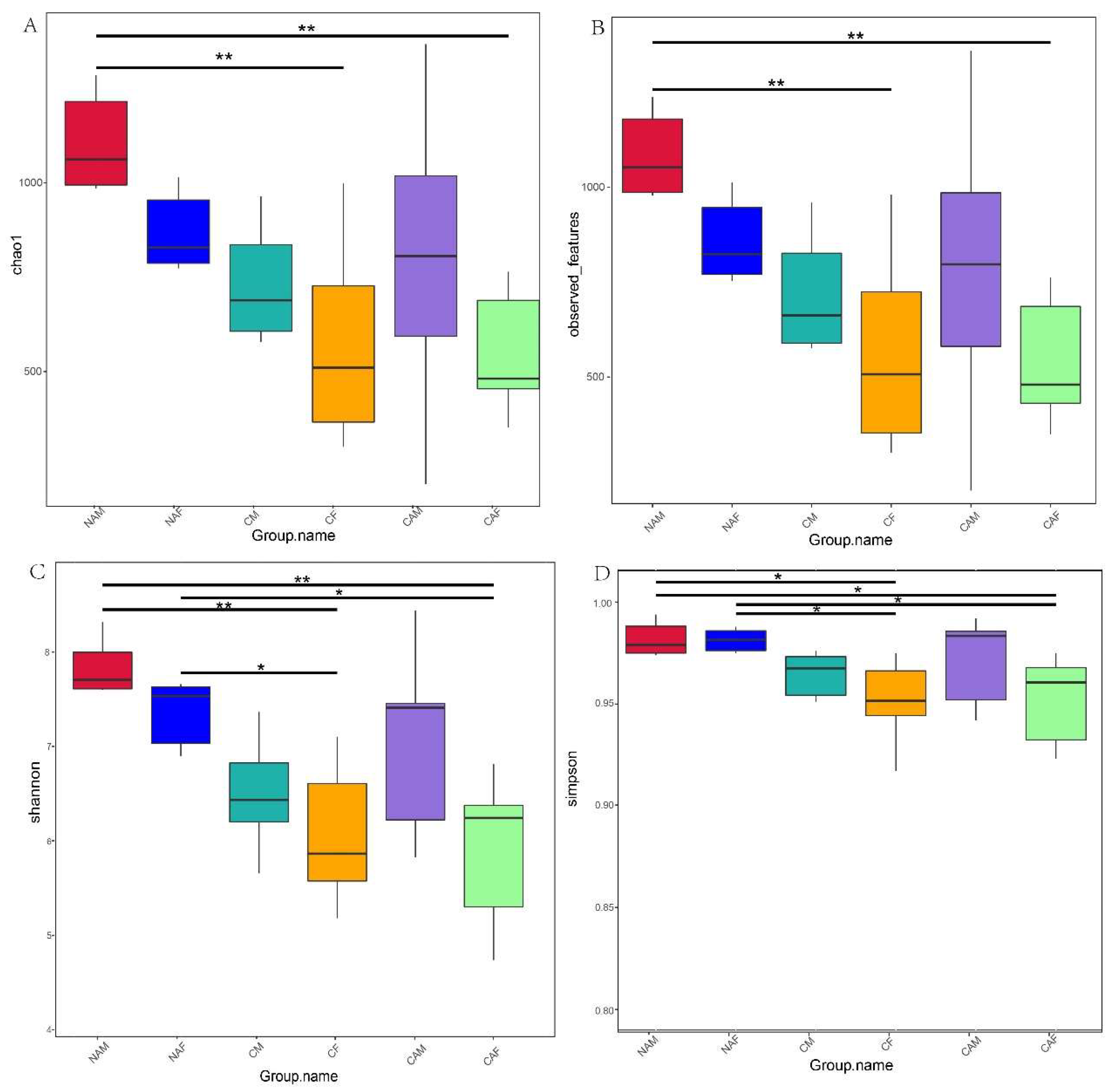
Figure 3.
Beta diversity analysis of rumen microorganisms across different starter feed groups, visualized using principal coordinate analysis (PCoA). CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs. CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female; M: male.
Figure 3.
Beta diversity analysis of rumen microorganisms across different starter feed groups, visualized using principal coordinate analysis (PCoA). CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs. CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female; M: male.
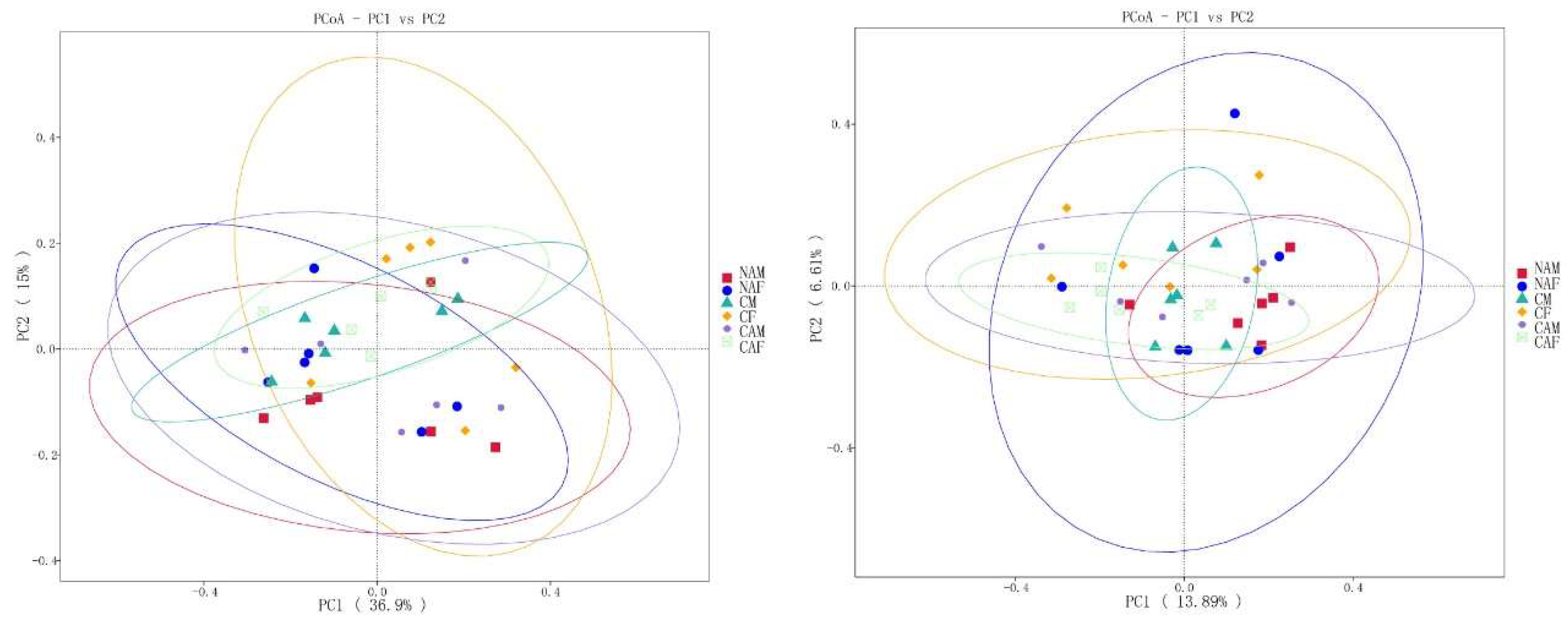
Figure 4.
Relationship between rumen microbial composition and type of starter feed in lambs. A: Differences in microbial composition among the groups at the phylum level. B: Differences in microbial composition among the groups at the genus level. CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs.CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female. M: male. Statistical significance is indicated as follows: P < 0.01 (**), P < 0.05 (*).
Figure 4.
Relationship between rumen microbial composition and type of starter feed in lambs. A: Differences in microbial composition among the groups at the phylum level. B: Differences in microbial composition among the groups at the genus level. CF: Standard pelleted starter feed containing alfalfa, female lambs. CM: Standard pelleted starter feed containing alfalfa, male lambs. NAF: Standard pelleted starter feed without alfalfa, female lambs. NAM: Standard pelleted starter feed without alfalfa, male lambs.CAF: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, female lambs. CAM: Starter feed with the same formulation as the CON group, but crumbled using a roller mill after pelleting, male lambs. F: female. M: male. Statistical significance is indicated as follows: P < 0.01 (**), P < 0.05 (*).
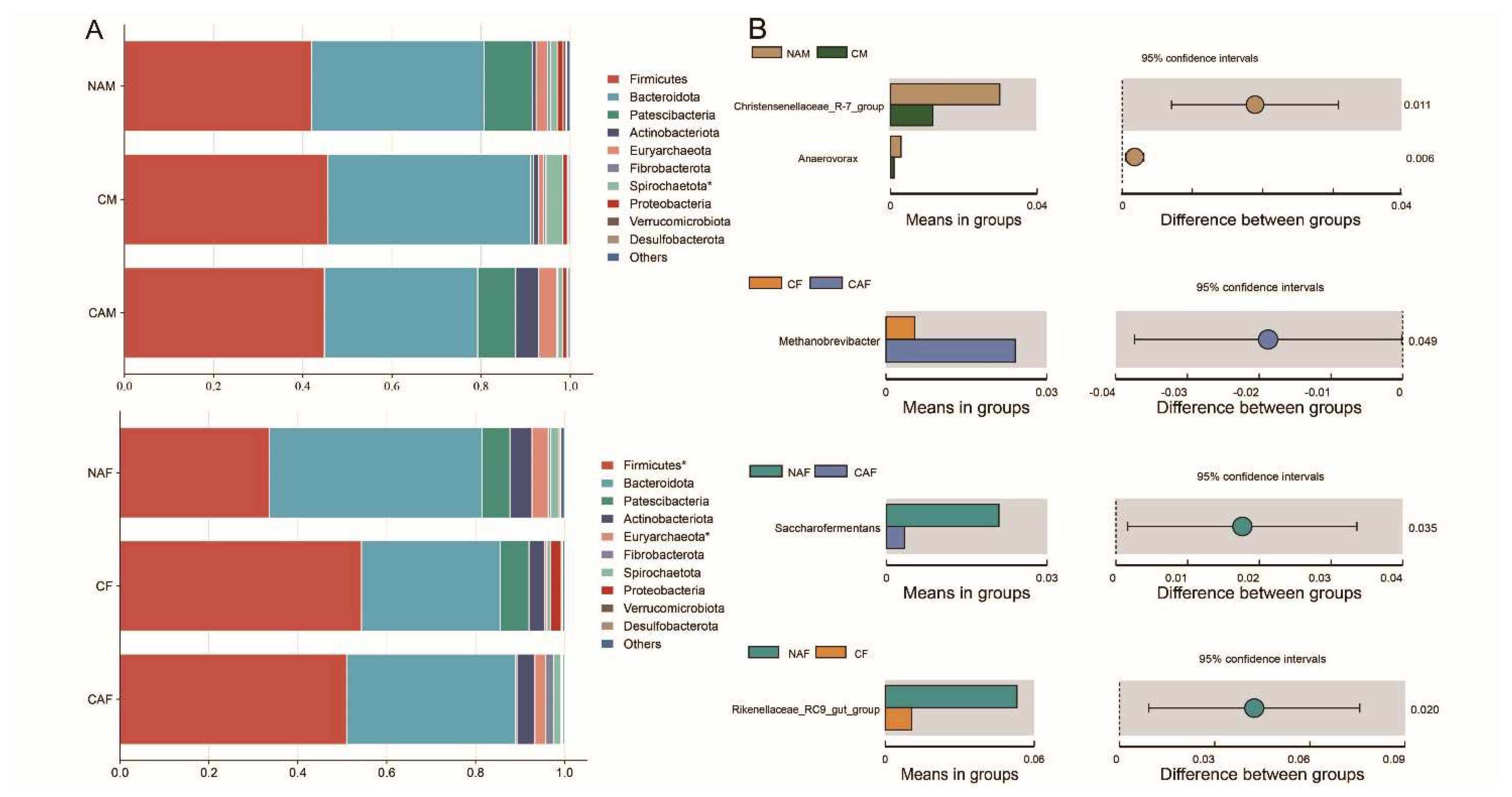

Table 1.
Dietary formulation and nutrient levels (air-dry basis).
| Ingredients | Percentage (%) and Group | |
| CON and CA Group | NA Group | |
| Alfalfa | 7.90 | —— |
| Corn | 37.30 | 40.30 |
| Extruded soybeans | 2.50 | 2.50 |
| Whey powder | 1.50 | 1.50 |
| Corn bran | 6.35 | 9.25 |
| Soybean meal | 18.25 | 19.25 |
| Cottonseed meal | 5.00 | 5.00 |
| DDGS | 12.00 | 13.00 |
| Bio-fermented feed | 2.50 | 2.50 |
| Limestone | 1.70 | 1.70 |
| Premix | 5.00 | 5.00 |
| Total | 100.00 | 100.00 |
| Chemical composition | Content | |
| DM (%) | 93.55 | 93.01 |
| DE (MJ·kg-1) | 12.68 | 13.00 |
| CP (%) | 22.38 | 22.58 |
| NDF (%) | 16.27 | 14.00 |
| ADF (%) | 7.37 | 6.35 |
| Ca (%) | 1.38 | 1.20 |
| P (%) | 0.38 | 0.50 |
Notes: The premix composition per kilogram of diet is as follows: 25 mg Fe as FeSO4·H2O; 40mg Zn as ZnSO4·H2O; 8mg Cu as CuSO4·5H2O; 40mg Mn as MnSO4·H2O; 0.3 mg I as KI; 0.2 mg Se as Na2SeO3; 0.1 mg Co as CoCl2; 940 IU vitamin A; 111 IU vitamin D; 20 IU vitamin E, and; 0.02 mg vitamin B12. DM, CP, NDF, ADF, Ca, and P were measured values, while DE was calculated in reference to Composition and Nutrient Values of Feed in China (The 26th edition).
Table 2.
Physical properties and starch gelatinization of starter feeds with different forms.
| Treatment | Hardness (ѱ N) | Density ( ρ g/cm3) | Starch Gelatinization (%) |
| CON | 230.94a | 0.90 | 22.36b |
| NA | 205.11a | 0.92 | 33.76a |
| CA | 27.34b | 0.82 | 23.65b |
| SEM | 10.473 | 0.073 | 0.020 |
| P-value | <0.001 | 0.397 | 0.002 |
Note: In the same column, different small letter superscripts indicate significant differences (P < 0.05). CON: Fed a standard pelleted starter feed containing alfalfa; NA: Fed a standard pelleted starter feed without alfalfa; CA: Fed a starter feed with the same formulation as the CON group but crumbled using a roller mill after pelleting. a,b Values within a row with different superscripts differ significantly at P < 0.05.
Table 3.
Effects of starter feed form on lamb growth performance and starter intake.
| Items | Groups | Main Effects | SEM | P-value | |||||||||||
| F | M | CON | NA | CA | F | M | Feed | Sex | F×S | ||||||
| CON | NA | CA | CON | NA | CA | ||||||||||
| BW (kg) | |||||||||||||||
| BW7 | 3.83 | 3.81 | 3.79 | 4.20 | 4.10 | 4.52 | 4.02 | 3.95 | 4.15 | 3.81 | 4.27 | 0.090 | 0.636 | 0.008 | 0.564 |
| BW21 | 5.49 | 5.45 | 5.31 | 6.25 | 6.15 | 6.00 | 5.87 | 5.80 | 5.65 | 5.42 | 6.13 | 0.120 | 0.770 | 0.005 | 0.992 |
| BW35 | 7.02 | 6.76 | 6.88 | 8.03 | 7.65 | 8.02 | 7.52 | 7.20 | 7.45 | 6.89 | 7.90 | 0.170 | 0.733 | 0.004 | 0.959 |
| BW49 | 8.48 | 7.93 | 9.43 | 9.43 | 8.96 | 10.06 | 8.96 | 8.44 | 9.74 | 8.61 | 9.48 | 0.230 | 0.080 | 0.061 | 0.934 |
| ADG (kg) | |||||||||||||||
| ADG7-21 | 0.12 | 0.12 | 0.11 | 0.15 | 0.15 | 0.11 | 0.13a | 0.13a | 0.11b | 0.11 | 0.13 | 0.000 | 0.030 | 0.039 | 0.258 |
| ADG21-35 | 0.11 | 0.09 | 0.11 | 0.13 | 0.11 | 0.14 | 0.12 | 0.10 | 0.13 | 0.10 | 0.13 | 0.010 | 0.142 | 0.063 | 0.798 |
| ADG35-49 | 0.10 | 0.08 | 0.18 | 0.10 | 0.09 | 0.15 | 0.10b | 0.09b | 0.16a | 0.12 | 0.11 | 0.010 | 0.000 | 0.463 | 0.398 |
| ADG7-49 | 0.11 | 0.10 | 0.13 | 0.12 | 0.12 | 0.13 | 0.12ab | 0.11b | 0.13a | 0.11 | 0.12 | 0.000 | 0.049 | 0.254 | 0.609 |
Note: In the same row, different small letter superscripts indicate significant differences (P < 0.05). CON: Fed a standard pelleted starter feed containing alfalfa; NA: Fed a standard pelleted starter feed without alfalfa; CA: Fed a starter feed with the same formulation as the CON group but crumbled using a roller mill after pelleting. F: Female; M: Male. a,b Values within a row with different superscripts differ significantly at P < 0.05.
Table 4.
Effects of starter feed form on starter intake.
| Items | Main Effects | SEM | P-value | ||
| CON | NA | CA | |||
| FI 7-21 | 6.66b | 7.32b | 9.44a | 0.191 | <0.001 |
| FI 21-35 | 25.28b | 17.25c | 57.99a | 1.823 | <0.001 |
| FI 35-49 | 92.15b | 50.92c | 159.85a | 5.066 | <0.001 |
| FI 7-49 | 41.36b | 25.16c | 75.76a | 2.281 | <0.001 |
Note: In the same row, different small letter superscripts indicate significant differences (P < 0.05). CON: Fed a standard pelleted starter feed containing alfalfa; NA: Fed a standard pelleted starter feed without alfalfa; CA: Fed a starter feed with the same formulation as the CON group but crumbled using a roller mill after pelleting. a,b Values within a row with different superscripts differ significantly at P < 0.05.
Table 5.
Effects of different food feeds and sex on nutrient digestibility.
| Items | Groups | Main Effects | SEM | P-value | |||||||||||
| F | M | CON | NA | CA | F | M | Feed | Sex | F×S | ||||||
| CON | NA | CA | CON | NA | CA | ||||||||||
| CP | 0.62 | 0.62 | 0.85 | 0.82 | 0.78 | 0.84 | 0.72b | 0.70b | 0.85a | 0.70 | 0.81 | 0.024 | 0.060 | 0.027 | 0.216 |
| NDF | 0.43 | 0.67 | 0.67 | 0.60 | 0.67 | 0.64 | 0.51b | 0.67a | 0.66a | 0.59 | 0.64 | 0.014 | 0.001 | 0.130 | 0.028 |
| ADF | 0.36 | 0.54 | 0.59 | 0.52 | 0.63 | 0.45 | 0.44b | 0.59a | 0.52a | 0.50 | 0.53 | 0.014 | 0.003 | 0.215 | 0.001 |
| DM | 0.80 | 0.79 | 0.76 | 0.73 | 0.76 | 0.80 | 0.77 | 0.78 | 0.78 | 0.78 | 0.76 | 0.018 | 0.956 | 0.681 | 0.382 |
| OM | 0.81 | 0.80 | 0.78 | 0.74 | 0.78 | 0.82 | 0.78 | 0.79 | 0.80 | 0.80 | 0.78 | 0.017 | 0.900 | 0.534 | 0.423 |
Note: In the same row, different small letter superscripts indicate significant differences (P < 0.05). CON: Fed a standard pelleted starter feed containing alfalfa; NA: Fed a standard pelleted starter feed without alfalfa; CA: Fed a starter feed with the same formulation as the CON group but crumbled using a roller mill after pelleting. F: Female; M: Male. a,b Values within a row with different superscripts differ significantly at P < 0.05.
Table 6.
Effects of different starter feed form and sex on rumen fermentation of lambs.
| Items | Groups | Main Effects | SEM | P-value | |||||||||||
| F | M | CON | NA | CA | F | M | Feed | Sex | F×S | ||||||
| CON | NA | CA | CON | NA | CA | ||||||||||
| Acetic acid (mmol/L) |
45.73 | 44.17 | 44.15 | 48.43 | 38.40 | 36.83 | 47.08 | 41.28 | 40.49 | 44.68 | 41.22 | 2.368 | 0.473 | 0.471 | 0.653 |
| Propionic acid (mmol/L) |
14.64 | 12.38 | 15.43 | 15.48 | 10.23 | 9.72 | 15.06a | 11.31b | 12.57ab | 14.15 | 11.81 | 0.518 | 0.019 | 0.031 | 0.050 |
| Isobutyric acid (mmol/L) |
1.78 | 1.48 | 1.90 | 1.42 | 1.15 | 1.07 | 1.60 | 1.31 | 1.48 | 1.72 | 1.21 | 0.098 | 0.497 | 0.014 | 0.520 |
| Butyric acid (mmol/L) |
5.06 | 4.42 | 6.11 | 6.44 | 3.48 | 3.99 | 5.75 | 3.95 | 5.05 | 5.20 | 4.64 | 0.365 | 0.146 | 0.448 | 0.155 |
| Isovaleric acid (mmol/L) |
0.84 | 0.86 | 1.22 | 0.93 | 0.52 | 0.72 | 0.88 | 0.69 | 0.97 | 0.97 | 0.72 | 0.075 | 0.312 | 0.110 | 0.273 |
| Valeric acid (mmol/L) |
0.74 | 0.64 | 0.81 | 0.76 | 0.57 | 0.60 | 0.75a | 0.61b | 0.71a | 0.73 | 0.65 | 0.025 | 0.080 | 0.107 | 0.187 |
| Total acid (mmol/L) |
71.67 | 66.83 | 72.50 | 76.34 | 57.24 | 55.81 | 74.00 | 62.04 | 64.16 | 70.34 | 63.13 | 3.168 | 0.273 | 0.264 | 0.386 |
| Acetate/propionic acid | 3.13 | 3.48 | 2.88 | 3.09 | 3.75 | 3.68 | 3.11 | 3.61 | 3.28 | 3.16 | 3.51 | 0.107 | 0.171 | 0.120 | 0.284 |
Note: In the same row, different small letter superscripts indicate significant differences (P < 0.05). CON: Fed a standard pelleted starter feed containing alfalfa; NA: Fed a standard pelleted starter feed without alfalfa; CA: Fed a starter feed with the same formulation as the CON group but crumbled using a roller mill after pelleting. F: Female; M: Male. a,b Values within a row with different superscripts differ significantly at P < 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



