
Submitted:
30 December 2024
Posted:
31 December 2024
You are already at the latest version
Abstract
Determining hemicellulose (HM) degradation is crucial for evaluating the nutritional value of ruminant diets. Our previous study showed that oxalic acid (OA) regulates rumen fermentation. Building on this research, the present study examined the effects of OA supplementation on sheep rumen fermentation, microbial diversity, and metabolite production in vitro. Diets with low and high HM levels (10.3% and 17%, respectively) and supplemented with seven concentrations of OA (0, 2.5, 5, 10, 20, 40, 80 mg/kg DM) were evaluated. Tests were conducted under both low (HM10.3%) and high (HM17%) hemicellulose conditions; however, addition of 10 mg/kgDM oxalic acid could have better effects under low hemicellulose (HM10.3%), with higher concentrations of acetic, propionic, and butyric acids, as well as total acids. A 2 × 2 factorial design was used to collect rumen fluid after 12 h of fermentation for analysis of microbial populations and metabolites. OA supplementation at 10 mg/kg DM significantly increased the relative abundances of several bacterial genera, including Prevotella, Butyrivibrio, Ruminococcus, Sharpea, RFN20, Bulleidia, Olsenella and Bifidobacterium (P < 0.05). A positive correlation was observed between Butyrivibrio and Sharpea and the production of isobutyric and isovaleric acids (P < 0.01), indicating these bacteria play a role in volatile fatty acid (VFA) production. Furthermore, rumen metabolites involved in mineral absorption and lipid metabolism, including α-tocopherol, L-glutamic acid, and ginkgolide B, were upregulated. In summary, supplementation with oxalic acid in HM diets alters rumen fermentation, enhances nutrient digestibility, promotes microbial diversity, and influences metabolic pathways. Thus, OA supplementation should be tailored to specific dietary conditions for optimal effects.
Keywords:
Oxalic acid
; Hemicellulose
; Nutrient digestibility
; Microbial diversity
; Metabolites
; In vitro
1. Introduction
Hemicellulose digestibility is a crucial factor for evaluating the nutritional value of ruminant diets. Ruminants rely on microbial communities in the rumen, including anaerobic microorganisms and extracellular enzymes, to break down polysaccharides such as cellulose and hemicellulose, which are otherwise difficult to digest. This fermentation process produces volatile fatty acids (VFAs), essential nutrients that help maintain rumen stability and overall host health[1]. For example, a study by Plata et al. showed that yeast culture (YC) supplementation to an animal diet can promote the degradation of hemicellulose[2]. Similarly, Palmieri et al. found that supplementation of oxalic acid in the diet may enhance rumen fermentation efficiency and improve fiber degradation[3]. Furthermore, our previous study identified oxalic acid as a functional substance of yeast culture, positively influencing animal growth and regulating gastrointestinal function[4].
Interestingly, oxalic acid is recognized as an antinutrient[5]. For example, it can bind with dietary calcium (Ca) or magnesium (Mg) to form insoluble calcium or magnesium oxalate, which may lead to mineral deficiency in the form of low serum calcium or magnesium levels. Additionally, oxalic acid is implicated in renal failure as it is reported to cause sedimentation in the kidneys[6]. The effect of oxalic acid on ruminant growth and development is mainly reflected in insufficient mineral intake and oxalic acid poisoning[7]. For example, addition of 6% oxalate to the diet had no adverse effect on sheep when introduced gradually, following a dose-by-dose approach[8]. However, Frutos et al. found that feeding 0.6 mM/kg live weight/day of oxalic acid increased the abundance of rumen bacteria that degrade active oxalic acid in goat rumen[9]. Moreover, adding free oxalic acid to the feed can cause changes in the decomposition rate of oxalic acid in rumen fluid, especially in goats, where the effect on oxalic acid breakdown is more pronounced[10].
In contrast to its negative effects, adding a certain amount of oxalic acid to diet could improve rumen efficiency and microbial function by breaking down of hemicellulose. This process may help maintain the ability of the rumen to degrade oxalic acid, improving the adaptation of ruminants to its presence. Palmieri et al. reported that roughage supplemented with organic acids improves rumen efficiency by maintaining a higher pH, balancing ammonia-nitrogen (NH3-N), increasing the microbial synthesis of proteins and VFAs, and reducing methane (CH4) production[11].
In this study, we hypothesize that oxalic acid supplementation in diets containing varying levels of hemicellulose will enhance rumen fermentation function, nutrient digestibility, and rumen microbial diversity. This study aimed to provide a valuable reference for the use of oxalic acid in ruminant nutrition, with potential application for improving fiber degradation and overall rumen efficiency.
Materials and Methods
2.1. Animals
Four healthy, quarantine-qualified male Small Tail Han sheep (36 ± 1.5 kg) with permanent rumen fistulae were housed individually, with regular fistula cleaning. They were fed daily at 8 am and 5 pm daily, with free access to clean water. All experiments were conducted at the College of Animal Science and Technology, Jilin Agricultural University, Changchun, China. All experimental procedures were performed in accordance with the Guidelines for the Care and Use of Experimental Animals of Jilin Agricultural University (JLAU-ACUC2024-011).
2.2. Experimental Design
A 2 × 7 full-factor experimental design was adopted, consisting of factors 1 and 2. Factor 1 consisted of two diets with different hemicellulose levels (10.3% and 17%), and Factor 2 was oxalic acid at seven different doses (0, 2.5, 5, 10, 20, 40, and 80 mg/kg DM), with four replicates per group. Samples were collected from three batches at three time points (0, 6, and 12 h). Subsequently, digestion parameters, fermentation parameters, and rumen contents were analyzed to determine the appropriate dose of oxalic acid to positively regulate rumen fermentation, based on previously published research[12]. Oxalic acid was obtained from the Rhawn Reagent Company (Shanghai, China).
2.3. In Vitro Experiment
Before morning feeding, rumen fluid was collected from the fixed rumen fistula using a rigid PVC tube and quickly returned to the laboratory, where it was filtered with a multi-layer gauze. At the same time, an appropriate amount of CO2 was maintained outside the rumen to ensure an anaerobic environment for the rumen flora. According to the method of Menke and Steingass et al.[11], the filtered rumen fluid and the buffer were mixed at a 1:1 ratio. After the mixture was stirred well to create a uniform solution, CO2 was continuously injected into the container containing the substrate and oxalic acid in advance, and the cap was tightly screwed. The container was placed in a gas bath shaking at a controlled temperature (39°C, 80 rpm) for oscillation culturing, and the gas production was monitored and recorded using an ANKOM RF gas production system (ANKOM Technology Corp., Macedon, NY, USA).
2.4. Determination of Rumen Fermentation Parameters
The ANKOM RF gas production system was configured to measure the gas pressure (psi) and record the cumulative gas production of each ANKOM bottle. The GPM software gas production formula is:
Vy = Vx × Ppsi × 0.068004084
where Vy represents the volume of gas generated (in mL). Vx stands for the volume of gas in the upper space of each bottle in mL; Ppsi denotes the cumulative pressure expressed in psi over a given period of time. Throughout this experiment, at each fermentation time point, a Sanxin MP 52304 digital pH meter (Shanghai Sanxin Instrumentation, Inc., Shanghai, China) was used according to the manufacturer's guidelines. The pH of all the bottles in a group were averaged to obtain the pH level based on fermentation time.
Using the method described by Chaney in 1962[13], the ammonia nitrogen (NH3-N) content of each test treatment was determined. A Cary series UV-visible spectrophotometer connected to a computer with Cary series analysis software (Agilent Technologies, California, USA) was used for this purpose.
We used an Agilent 7890 A gas chromatograph (Agilent Technologies, Santa Clara, California, USA) along with a 50-m (inner diameter 0.32 mm) CP-Wax Chrompack silica fused capillary column (Varian, Palo Alto, CA, USA) to measure six major fatty acids (acetic acid, propionic acid, butyric acid, isobutyric acid, valeric acid, and isovaleric acid) and total volatile fatty acid content.
After in vitro fermentation, the substrate was dried at 105°C until absolutely dry; then, in vitro dry matter digestibility (IVDMD) was calculated. The crude protein digestibility (IVCPD), crude fat digestibility (IVEED) and neutral detergent fiber digestibility (IVNDFD) were calculated using the formulas mentioned by Huang et al.[14], DeFeo et al.[15], and Goeser and Combs[16], respectively.
2.5. Determination of Rumen Microbial Diversity
Nucleic acids were extracted using an OMEGA Soil DNA Kit (D5625-01) (Omega Bio-Tek, Norcross, GA, USA). DNA was quantified using an ultraviolet spectrophotometer (Agilent Technologies, California, United States), and the highly variable V3V4 region of the bacterial 16s rDNA gene was used as a template for sequencing. Specific primers, F, ACTCCTACGGGAGGCAGCA, R, and GGACTACHVGGGTWTCTAAT, were selected. NEBQ5 DNA high-fidelity polymerase was used for PCR amplification, and the PCR products were quantified on a FLx800 microplate reader (BioTek, Winooski, VT, US) using a Quant-iT PicoGreen dsDNA Assay kit (Thermo Fisher Scientific, Waltham, MA, USA) and prepared according to the volume of the sample. The TruSeq Nano DNA LT library (Illumina, San Diego, CA, USA) was used to construct the database; then the Agilent Sensitivity DNA kit (Agilent Technologies, Santa Clara, CA, USA) was used to perform quality control of the eligible libraries. Double-terminal sequencing was performed using the NovaSeq 6000 SP kit (500 cycles; Illumina)[17].
2.6. Determination of Rumen Metabolites
After slow thawing at 4°C, an appropriate amount of methanol/acetonitrile/aqueous solution (with acetonitrile and water in a =1:1 volume ratio) was added to the samples. The mixture was next subjected to vortex stirring and ultrasound treatment at low temperature for 30 min. Subsequently, the samples were left to stand at -20°C for 10 min and centrifuged at 4°C for 20 min at a speed of 14,000 g. The supernatant was vacuum dried, and 100 μL of acetonitrile aqueous solution (with acetonitrile and Water in a 1:1 volume ratio) was added for redissolving. After swirling, centrifugation was performed again at 14,000 g for 15 min at 4°C. Finally, the supernatant was collected for further analysis.
The samples were separated using an HILIC column with the 1290 ultra-high performance liquid chromatography system (Agilent). The samples were automatically injected at 4°C. To avoid fluctuations in the instrument detection signal, a random sequence method was employed for continuous analysis of the samples, and quality control samples were inserted into the sample queue to monitor and evaluate the system stability and analyze the reliability of the experimental data. Primary and secondary spectral peaks were extracted using a mass spectrometer (AB Sciex, Path Framingham, MA, USA). The samples were separated using ultra high-performance liquid chromatography (UHPLC) and detected using a TripleTOF-6600 mass spectrometer (AB Sciex) with electrospray ionization (ESI).
2.7. Data Analysis
2.7.1. Rumen Fermentation Parameters
The general linear model of the IBM SPSS Statistics 23 software was used to analyze the relationship between two factors. The statistical analysis model (Yijk= μ + Fi + Vj + Fi × Vj + eijk) was used to analyze the dietary and oxalic acid supplemental doses at different hemicellulose levels and their interaction effects. Here, μ represents the population average. where Fi denotes the dietary effect of different hemicellulose levels (i = 1-2). Vj is the supplemental dose of oxalic acid (j = 1-7). Fi × Vj represents the interaction effect between dietary hemicellulose level and oxalic acid dosage. eijk indicates the residual error. Duncan's multiple comparison method was used to test the significance of factors 1 and 2 and their interactions (factor 1 × factor 2). P < 0.05 was considered as significant and P < 0.01 was considered as an extremely significant difference.
2.7.2. Rumen Microbial Diversity
QIIME2 v2019.4 (https://qiime2.org/) and the R software version 3.2.0 (R Foundation for Statistical Computing, Vienna, Austria) were used to analyze the sequencing results. The demux plug-in was employed for multiplex decomposition of the original sequence. The cutadapt plug-in was used for primer cutting. The DADA2 plug-in was used for quality filtering, denoising, splicing, and elimination of mosaics. The Alpha diversity index was calculated using the ASV table in QIIME2. The Jaccard index, Bray-Curtis index, and UniFrac distance were used to qunatify beta diversity. Principal component analysis was used to visualize the samples. Linear discriminant analysis effect size (LEfSe) was used to measure the differences in microorganisms among the groups. Based on the MetaCyc (https://metacyc.org/) and KEGG (https://www.genome.jp/kegg/pathway.html) databases, functional prediction was performed using PICRUSt2 (https://huttenhower.sph.harvard.edu/picrust/).
2.7.3. Rumen Metabolites
ProteoWizard software was employed to convert the original data into the mzXML format, which was used for alignment and retention of peaks. Peak areas were extracted and corrected for time. The structures of the metabolites were initially identified from the data obtained by XCMS, and the data were preprocessed. Finally, quality evaluation and analysis of the experimental data were performed. Lastly, Mothur software (https://mothur.org/) was used to calculate the metabolome data and Spearman correlation coefficients between microorganisms and bacteria. Based on the Spearman correlation coefficient matrix results, (rho correlation coefficient is between -1 and 1; when -1 < rho < 0, the two are negatively correlated; when 0 < rho <1, the two are positively correlated. When rho = 0, there is no correlation between the two), and R software was used for heat mapping.
3. Results
3.1. Nutritional Digestibility In Vitro
After 6 h of in vitro fermentation, the IVDMD of the treatment groups supplemented with oxalic acid was significantly higher than that of the control group when the diet contained low hemicellulose (HM10.3%) levels (P < 0.05; Table 1). When the fermentation time was extended to 12 h, the IVDMD of the treatment groups supplemented with 2.5, 5, 10, and 20 mg/kg DM oxalic acid at the low hemicellulose (HM10.3%) level also surpassed that of the control group. However, IVDMD was lower in the groups treated with 40 and 80 mg/kg DM oxalic acid than that in the control group. In the treatment groups with the high hemicellulose (HM17%) diet, the IVDMD in the 5 mg/kg DM oxalic acid group was significantly greater than that in the control group (P < 0.05; Table 1).
Different levels of hemicellulose and oxalic acid significantly affected rumen IVCPD, IVEED, and IVNDFD in vitro, and the interaction between hemicellulose levels and oxalic acid dose also significantly affected these three parameters (P < 0.05). After 6 h of fermentation, the IVCPD values in the oxalic acid supplementation groups were greater than that of the control group under the HM10.3% diet. However, under the HM17% diet, the IVCPD values in the 20 and 80 mg/kg DM oxalic acid supplementation groups were significantly lower than that of control group. The IVCPD values of the 2.5, 20, 40, and 80 mg/kg DM oxalic acid-supplementation groups were significantly lower than that of the control group under the HM17% diet (P < 0.05; Table 1). After 12 h of fermentation, the IVCPD of the 10 and 80 mg/kg DM oxalic acid supplementation groups was significantly greater than that of the control group under the HM10.3% diet (P < 0.05; Table 1).
At the HM17% level, the IVEED of the 80 mg/kg DM oxalic acid supplementation group was significantly greater than those of the 2.5, 10, 20, 40 mg/kg DM oxalic acid supplementation and the control groups (P < 0.05) after 6 h of fermentation. With an increase in fermentation time, the IVEED of all oxalic acid supplementation groups was significantly less than that of the control group at 12 h of fermentation with low hemicellulose levels (P < 0.05). The IVEED of the 2.5 and 10 mg/kg DM oxalic acid supplementation groups at the HM17% level was significantly greater than those of the other oxalic acid treatment groups and the control group (P < 0.05; Table 1).
At the HM10.3% level, after 6 h of fermentation, the IVNDFD in the groups supplemented with 2.5, 5, and 80 mg/kg DM oxalic acid was significantly greater than those of the other oxalic acid treatments and the control group (P < 0.05). The IVNDFD values of the groups supplemented with 2.5, 5, 10, and 20 mg/kg DM oxalic acid were significantly higher than those of the groups supplemented with 40 and 80 mg/kg DM oxalic acid, as well as the control group after 12 h of fermentation (P < 0.05). At the HM17% level, the group supplemented with 5 mg/kg DM oxalic acid had a higher IVNDFD value than the groups supplemented with 2.5, 10, 20, and 40 mg/kg DM oxalic acid, and the control (P < 0.05; Table 1).
3.2. Rumen Fermentation Parameters
The effects of dietary hemicellulose levels and oxalic acid supplementation on rumen gas production and ammonia nitrogen concentration in vitro differed significantly between the groups (P < 0.01). Gas production over 6 h of in vitro fermentation in the both hemicellulose groups supplemented with 40 mg/kg DM oxalic acid was higher than that in the other oxalic acid supplementation groups and the control (P < 0.05). After 12 h of fermentation, the gas production of the 40 mg/kg DM oxalic acid group was lower than that of all other oxalic acid dosage groups at both dietary hemicellulose levels (P < 0.05; Table 1). During in vitro fermentation for 6 h, at the HM10.3% level, the ammonia nitrogen concentration of the 5 mg/kg DM oxalic acid group was significantly higher than that of the 20 and 40 mg/kg DM oxalic acid and control groups (P < 0.05); At the HM17% level, the ammonia nitrogen concentrations of the 10 and 20 mg/kg DM oxalic acid supplementation groups were significantly higher than those of 2.5, 5, 40, and 80 mg/kg DM oxalic acid groups (P < 0.05; Table 2).
As the fermentation time increased, acid production gradually increased, and the pH of all groups continued to decrease. The effects of hemicellulose levels on rumen pH and total volatile fatty acid (TVFA) concentrations in vitro were extremely significant. The concentration of TVFA was significantly affected by the interaction between hemicellulose level and oxalic acid dosage (P < 0.01). After 6 h of in vitro fermentation, the pH values of all oxalic acid groups were lower than that of the control group at the HM10.3% level. The pH values of the 40 and 80 mg/kg DM oxalic acid groups were significantly lower than those of the other oxalic acid dosages and the control group (P < 0.05). At the HM17% level, the pH values of the 10 and 80 mg/kg DM oxalic acid groups decreased as time increased. When the fermentation time reached 12 h, there were no significant difference among the groups at the HM10.3% level (P > 0.05). At the HM17% level, the pH values of the 10 and 80 mg/kg DM oxalic acid groups decreased as time passed and were significantly lower than those of the other oxalic acid dosages and control group (P > 0.05;Table 2).
At the HM10.3% level, the TVFA concentration in the 10 mg/kg DM oxalic acid group was significantly higher than that in the other oxalic acid groups and the control (P < 0.05). After 6 h of fermentation (Figure 1A), the TVFA concentration in the oxalic acid groups was significantly higher than that in the control group (P < 0.05). At the HM17% level, the TVFA concentration in the 10 and 20 mg/kg DM oxalic acid groups was significantly higher than that in the other oxalic acid groups and the control (P < 0.05). After 12 h of fermentation (Figure 1B), the TVFA concentration in the 80 mg/kg DM oxalic acid group was significantly higher than that in the other oxalic acid groups and the control (P < 0.05). Additionally, the TVFA concentration in the 2.5 mg/kg DM oxalic acid group also increased with time and was significantly higher than that in the control group (P > 0.05; Table 2).
In the hemicellulose diets supplemented with 10 mg/kg DM oxalic acid, the concentrations of acetic acid, propionic acid, butyric acid, and total acid were significantly higher than those in the other treatment and control groups (P < 0.05; Figure 1). Under the HM10.3% diet, oxalic acid supplementation promoted propionic acid fermentation, enhanced propionic acid production, and decreased ethylene to propylene ratio, which potentially improved rumen fermentation function and potentially enhanced VFA composition potential.
3.3. Microbial Diversity In Vitro
3.3.1. Ruminal Bacterial Alpha Diversity Indexes
Pielou's evenness, Shannon, and Simpson indexes were significantly affected by the interactive effects of different hemicellulose levels in the diet and the addition of oxalic acid (P < 0.01; Figure 2C). Adding 10 mg/kg DM of oxalic acid to a diet with a HM10.3% had no significant effect on the alpha diversity index of rumen microbes (P > 0.05). Adding 10 mg/kg DM of oxalic acid to the diet with HM17% significantly reduced Pielou's evenness, Shannon, and Simpson indexes (P < 0.01). Adding no oxalic acid to the diet at both hemicellulose levels in both groups had no significant effect on these indexes. The HM17% group with 10 mg/kg DM of oxalic acid had significantly higher Pielou's evenness and Shannon index values than the HM10.3% group with added oxalic acid (P < 0.05). Adding 10 mg/kg DM of oxalic acid to both hemicellulose groups significantly reduced the Pielou's evenness, Shannon, and Simpson indexes of the HM17% group compared to that of the HM10.3% group (P < 0.05). In this study, the addition of oxalic acid at the high hemicellulose level reduced the diversity of rumen microbial communities.
3.3.2. Ruminal Bacterial Phylum Composition
The top ten phyla in relative abundance were Bacteroidetes, Firmicutes, Actinobacteria, phylum TM7, Proteobacteria, Spirochaetes, Tenericutes, Verrucomicrobia, Fibrobacteres, and Synergistetes. Bacteroidetes, Firmicutes, and Actinobacteria were the dominant phyla, each with relative abundances greater than 1% (Figure 2A).
Adding 10 mg/kg DM of oxalic acid at the HM10.3% level significantly affected the relative abundances of Bacteroidetes and Proteobacteria during in vitro fermentation of rumen contents (P < 0.05). Addition of 10 mg/kg DM of oxalic acid at the high hemicellulose level significantly affected the relative abundance of Bacteroidetes, Actinobacteria, Phylum TM7, Spirochaetes, and Tenericutes during in vitro rumen content fermentation (P < 0.05). The addition of oxalic acid significantly affected the relative abundances of Bacteroidetes, Firmicutes, Actinobacteria, Phylum TM7, Proteobacteria, Spirochaetes, Tenericutes, Verrucomicrobia, and Fibrobacteres during in vitro rumen fermentation at both hemicellulose levels (P < 0.05). The relative abundances of Bacteroidetes, Actinobacteria, and Phylum TM7 were significantly affected by the interaction between oxalic acid and different hemicellulose levels (P < 0.01). After 12 h of in vitro rumen fermentation, adding 10 mg/kg DM of oxalic acid at the HM10.3% level significantly reduced the relative abundances of Bacteroidetes and Proteobacteria compared with those of the control group (P < 0.05). Adding 10 mg/kg DM of oxalic acid at the HM17% level significantly increased the relative abundances of Bacteroidetes, Spirochaetes, and Tenericutes compared with those of the control group. The relative abundances of Firmicutes, Spirochaetes, and Verrucomicrobia in the HM10.3% groups were significantly higher than those in the HM17.0% groups (P < 0.05), with the relative abundance of Firmicutes differing by up to 8.9%.
3.3.3. Rumen Bacterial Genus Composition
The top ten microbial genera were Prevotella, Butyrivibrio, Ruminococcus, Succiniclasticum, Sharpea, RFN20, Selenomonas, Bulleidia, Olsenella, and Bifidobacterium (Figure 2B). With relative abundances greater than 1%, the dominant bacterial genera also varied depending on the dietary level of hemicellulose and the amount of oxalic acid added. During the 12-h rumen fermentation outside the body, there were five dominant bacterial genera in the HM10.3% group with 10 mg/kg DM of oxalic acid added, namely Prevotella, Butyrivibrio, Ruminococcus, Succiniclasticum and Sharpea. Similarly, there were five dominant bacterial genera in the HM17% group with 10 mg/kg DM of oxalic acid added, namely Prevotella, Butyrivibrio, Succiniclasticum, Sharpea and Bulleidia.
Adding 10 mg/kg DM of oxalic acid to the HM10.3% level significantly affected the relative abundances of Prevotella, Ruminococcus and Selenomonas during in vitro rumen fermentation (P < 0.05). Adding 10 mg/kg DM of oxalic acid to the HM17% diet significantly affected the relative abundances of Prevotella, Olsenella, and Bifidobacterium during in vitro rumen fermentation (P < 0.05). The addition of 10 mg/kg DM of oxalic acid to both hemicellulose levels significantly affected the relative abundances of Prevotella, Butyrivibrio, Ruminococcus, Sharpea, RFN20, Bulleidia, Olsenella and Bifidobacterium during in vitro rumen fermentation (P < 0.05). Prevotella, Ruminococcus, Bulleidia and Olsenella were significantly affected by the interaction between oxalic acid addition and hemicellulose level (P < 0.01). Compared to the control group, adding 10 mg/kg DM of oxalic acid to the HM10.3% significantly affected the relevant parameters.
3.4. The Relationship Between Rumen Fermentation Parameters and Bacterial Genus Composition
By analyzing the correlation between the top ten genera and rumen fermentation parameters under different levels of hemicellulose, we examined whether oxalic acid promoted or inhibited the relationship between them. A 12-h in vitro simulation of rumen fermentation was conducted at the HM10.3% level in the diet (Figure 3A, B). In the control group, Sharpea showed a significant positive correlation with acetic acid and total acid levels (P < 0.05), and a moderate positive correlation with NH3-N (P < 0.05). RFN20 showed a significant positive correlation with NH3-N (P < 0.05). Olsenella showed a significant positive correlation with acetic acid concentration (P < 0.05). In the group treated with 10 mg/kg DM of oxalic acid, Butyrivibrio showed a significant negative correlation with total acid (P < 0.05) and a complex correlation with IVNDFD (P < 0.05). Sharpea also showed a significant positive correlation with the IVNDFD (P < 0.05). Ruminococcus showed a significant negative correlation with propionic acid and total acid, and a complex correlation with the acid-to-propionate ratio (P < 0.05). In the diet with HM17% (Figure 3C, D), the control group showed a significant negative correlation between Butyrivibrio and methane production (P < 0.05), a moderate negative correlation between Sharpea and methane production (P < 0.05). A significant negative correlation was found between Selenomonas and the acid-to-propionate ratio (P < 0.05), and a moderate positive correlation between Olsenella and propionic acid (P < 0.05). Prevotella showed a significant negative correlation with the isobutyric acid content. Bifidobacterium showed a significant positive correlation with the total acid content (P < 0.05) in the control group. The correlation between genera and fermentation parameters was more complex in the groups treated with 10 mg/kg DM oxalic acid.
3.5. Rumen Metabolites In Vitro
Based on the results of the fermentation parameters, the groups supplemented with 10 mg/kg DM in diets with different hemicellulose levels for 12 h were selected for rumen metabolite group analysis. A total of 795 metabolites were detected. Among these, organic acid heterocyclic compounds accounted for the highest proportion, accounting for 21.1% of total metabolites. The other metabolite types accounted for less: 20.9% were attributed to organic acids and their derivatives, 16.5% to phenyl compounds, 6.5% to organic nitrogen compounds, 6.5% to organic oxygen compounds, and 2.6% to nucleosides, nucleotides, and analogs. Lastly, the polyketones of phenylpropane accounted for 2.4%, and hydrocarbons accounted for 1.6% (Figure 4A).
A variety of differential metabolites (standard FC > 1, P < 0.01, VIP > 1.5) were screened after 12 h of in vitro fermentation. Compared to oxalic acid added to the HM10.3% diet, the differential metabolites (+)-.alpha.-tocopherol and Maltotriose were significantly down-regulated. Under the HM17% diet supplemented with oxalic acid, differential metabolites Trp-Cys-Lys, 13-keto-9z,11e-octadecadienoic acid were significantly upregulated, while 2-pyridinecarboxylic, glycocholic acid, and other substances were significantly downregulated. Supplementation with 10 mg/kg DM oxalic acid in both hemicellulose diets was compared with the HM10.3% and 10 mg/kg DM oxalic acid group. The differential metabolites of Maltotriose, L-carnitine, Atrazine desisopropyl, and Ginkgolide B in the HM17% and 10 mg/kg DM oxalic acid group were detected, and other substances were significantly up-regulated (Figure 4B–E).
3.6. Rumen Metabolic Pathway
According to the P and FDR values, several KEGG metabolic pathways were significantly enriched. After 12 h of in vitro fermentation, the differential metabolite enrichment pathways between the HM10.3% and 10 mg/kg DM group and the control group were analyzed. This showed that the differential metabolites were mainly involved in mineral absorption and glycerophospholipid metabolism. The metabolite enrichment pathways of the HM17% and 10 mg/kg DM oxalic acid group compared to the control group were mainly concentrated in riboflavin metabolism, beta-alanine metabolism, histidine metabolism, pyrimidine metabolism, and biosynthesis of cofactors (Figure 4F–I).
3.7. The Correlation Between Rumen Metabolites and Rumen Bacterial
Correlations between the top 50 metabolites and the different bacteria were analyzed. In the HM10.3% diet supplemented with 10 mg/kg DM oxalic acid, ruminal metabolites turkesterone and salicylaldehyde were significantly positively correlated with Prevotella, Ruminococcus and Succinolytica relative abundances (P < 0.05). When 10 mg/kg DM oxalic acid was added to the HM17% diet, rumen metabolites simazine hydroxy and pirimiphos-ethyl were significantly positively correlated with Lactococcus lactis and Aminococcus aminoethyl (P < 0.05). When the two diets were supplemented with 10 mg/kg DM oxalic acid, the vitamin B5 and pelanoic acid contents in the rumen metabolites of the HM17% diet were significantly positively correlated with Ruminococcus and RNF20 bacteria (P < 0.05; Figure 5A, B).
4. Discussion
Maintaining optimal rumen pH is crucial for rumen performance and overall health and ruminant nutrition[18]. Decreased rumen pH and subacute rumen acidosis (SARA) are common challenges[19], especially in intensive sheep production systems. This study found that supplementing oxalic acid at different dietary hemicellulose levels had little effect on rumen pH, which is consistent with previous findings[20]. These results imply that oxalic acid supplementation at different dietary hemicellulose levels does not adversely affect the rumen pH in sheep. This observation indicates that it can be used safely without disrupting rumen stability. The rumen of ruminants contains a complex microecosystem that serves as the primary site for nutrient digestion and absorption[21]. To assess feed degradation rates in the rumen, common indexes such as IVDMD, IVCPD, IVEED, and IVNDFD are often employed to evaluate the nutrient degradation rate in the rumen, providing insights into the efficiency of nutrient background and microbial activity in the rumen[22].
While oxalic acid is not commonly used as a feed additive for ruminants, our study revealed a significant interaction between diets with varying levels of hemicellulose and oxalic acid supplementation. Specifically, oxalic acid supplementation had more pronounced effect in diets with low hemicellulose levels (HM10.3%) compared to in diets with high hemicellulose levels (HM17%). This observation suggests that supplementation of oxalic acid to a low-hemicellulose diet improves the breakdown of carbohydrates, proteins, fats, and neutral detergent fiber. It likely facilitates the colonization of relevant bacteria in the rumen, potentially improving nutrient degradation and microbial activity.
In ruminants, proteins are broken down into peptides and amino acids, which are further degraded into ammonia nitrogen, organic acids, and CO2[23]. Ammonia nitrogen, certain peptides, and free amino acids are crucial precursors for protein formation in microorganisms[24]. Consequently, ammonia nitrogen levels in the rumen serve as indicators of protein utilization in the diet. Ortega et al.[25] observed that under normal conditions, rumen ammonia-nitrogen content remains stable; however, under certain conditions, it can adversely affect protein synthesis. Moreover, at higher nutrient levels, the rate of ruminal protein decomposition surpasses that of protein synthesis, leading to significant nitrogen loss. Belenguer et al.[20] demonstrated that the addition of 4.8 g/d of oxalic acid to rumen fistulas resulted in changes to both ammonia-nitrogen levels and rumen pH within a few hours of feeding. Furthermore, in Belenguer et al.[20], incorporation of oxalic acid into the diet reduced the mean value of ammonia nitrogen in rumen fistulas.
Our study found that addition of oxalic acid at a low hemicellulose level decreased the average rumen ammonia-nitrogen concentration. Conversely, the addition of appropriate oxalic acid at a high hemicellulose level increased the average rumen ammonia-nitrogen concentration. These changes in ammonia-nitrogen levels can be considered a phenotypic response to the dietary treatments, reflecting the adaptive mechanisms to varying oxalic acid and hemicellulose levels. We feel this phenotypic response could be attributed to the relatively small amount of oxalic acid used in this experiment. Furthermore, at high hemicellulose levels, oxalic acid supplementation may enhance the microbial abundance associated with protein degradation, thereby converting feed proteins into oligopeptides and amino acids, which are then degraded into ammonia nitrogen.
It is worth noting that gas production in the rumen is a reliable indicator of microbial activity and carbohydrate fermentation efficiency[26]. In our study, at low hemicellulose levels, the group supplemented with 5 mg/kg DM of oxalic acid showed the highest gas production. Conversely, at high hemicellulose levels, the groups supplemented with 10 and 20 mg/kg DM of oxalic acid exhibited the highest gas production. These results suggest that adding an optimal amount of oxalic acid to diets with varying hemicellulose levels regulates ruminal microbial activity and enhances carbohydrate metabolism.
Notably, 60% of ruminants' energy comes from VFAs, with over 95% being acetic acid, propionic acid, and butyric acid. Among these, propionic acid demonstrates the highest energy conversion rate, followed by acetic acid[20]. Acetic acid contributes to lactose formation, thereby contributing to an improved milk yield in sheep. Additionally, propionic acid serves as a precursor for gluconeogenesis, and higher concentrations of propionic acid are associated with increased fattening effects. In a study by Belenguer, daily injection of 0.6 mmol/kg body weight oxalic acid led to changes in the proportions of certain acids, including an increase in ethylene and propyl and a decrease in butyric acid and trace volatile fatty acids. Our study found that adding 10 mg/kg DM oxalic acid led to higher concentrations of acetic, propionic, butyric, and total acids at both high and low hemicellulosic levels. At low hemicellulose levels, oxalic acid supplementation hindered the transition to propionic acid fermentation, while at high hemicellulose levels it facilitated propionic acid fermentation, resulting in a lower ethylene-to-propylene ratio. These findings suggest that an optimal amount of oxalic acid supplementation simulated rumen microecosystem in vitro enhanced the acid-producing capacity of rumen bacteria positively impact VFA profile.
The rumen ecosystem is home to a diverse array of microorganisms, primarily bacteria[27]. Factors such as diet, additives, host species, and environmental conditions influence microbial diversity[28]. Alpha diversity indices, such as Chao1, Shannon, and Simpson diversity indices, are commonly used to assess species abundance within microbial communities[29]. Previous studies have shown that oxalic acid reduces the microbial diversity in sheep; however, with continued supplementation, the diversity gradually recovers[20]. It has been reported that the addition of oxalic acid to diets with varying hemicellulose levels results in decreased microbial diversity, which is consistent with the findings of the present study. This may be attributed to the slow proliferation of oxalic acid-degrading bacteria, such as oxalicogenes and fecalicococcus, with their diversity gradually rebounding over time. In our experiments, the composition of Bacteroidetes varies across different parts of the rumen; however, Bacteroidetes and Firmicutes were the dominant phyla, as expected[30]. Bacteroidetes (40.95%) and Firmicutes (36.36%) are the prevalent groups in sheep rumen[31]. Prevotella, Butyrivibrio, and Pseudobutyrivibrio are bacterial genera that promote fiber fermentation and are key components of Bacteroides and Firmicutes. Our study found that supplementing with 10 mg/kg DM of oxalic acid at high hemicellulose levels (HM17%) significantly increased the abundance of Bacteroidetes, Actinobacteria, TM7, Proteobacteria, Tenericutes, and Fibrobacteres (P < 0.05). This observation suggests that oxalic acid supplementation at high dietary hemicellulose levels enhances the presence of beneficial bacteria involved in cellulose and hemicellulose fermentation. Additionally, supplementation with 10 mg/kg DM oxalic acid significantly affected the presence of Prevotella, Butyrivibrio, Ruminococcus, Sharpea, RFN20, Bulleidia, Olsenella, and Bifidobacterium (P < 0.05). Some studies have indicated that Prevotella promotes fiber degradation, making it an important genus for enhancing hemicellulose degradation[32].
The presence of oxalic acid can affect the rumen microecological environment, especially in relation to hemicellulose levels. Our correlation analysis revealed that after 12 h of in vitro rumen fermentation, Butyrivibrio in the group supplemented with 10 mg/kg DM of oxalic acid at high hemicellulose levels (HM17%) showed a significant positive correlation with isobutyric acid, butyric acid, isovaleric acid, and total acids (P < 0.05). Similarly, Sharpea exhibited a significant positive correlation with isobutyric acid and isovalent acid (P < 0.05). These results suggest that supplementing oxalic acid at high dietary hemicellulose levels enhances the potential of Butyrivibrio and Sharpea to produce more VFAs.
In the present study, we also identified 34 different bacterial genera between the two groups supplemented with 10 mg/kg DM oxalic acid. This result indicates that oxalic acid supplementation to diets with different hemicellulose levels influences the composition of rumen microflora during in vitro fermentation and may enhance its biological functions. Using non-targeted metabolomics, we found that oxalic acid supplementation had a regulatory effect on rumen metabolites, with organic acid heterocyclic compounds being the primary metabolites. At a low hemicellulose level, the differential metabolite pathway was primarily associated with mineral absorption and lipid metabolism. At high hemicellulose levels the pathway focused on cofactor biosynthesis.
The metabolites of 0 mg/kg DM oxalic acid added to hemicellulose were primarily involved in choline metabolism, anti-folate resistance, and carbohydrate digestion. In the low hemicellulose level, the addition of 10 mg/kg DM oxalic acid notably increased alpha-tocopherol, a form of vitamin E known for its widespread use and high activity[33]. Studies have demonstrated that alpha-tocopherol can disrupt the transport of very low-density lipoprotein (VLDL) and reduce its secretion, thus preventing liver dysfunction[34].
In the low hemicellulose group with no oxalic acid (0 mg/kg DM) supplementation, there was a significant upregulation of L-glutamate, an amino acid crucial for cellular and systemic levels[35]. L-glutamate plays a key role as a source of energy and precursor for various biomolecules in the gut and liver. Interestingly, in the low hemicellulose group supplemented with 10 mg/kg DM oxalic acid, there was a significant upregulation of choline, which has been shown to promote lipid metabolism in dairy cows by affecting transcriptional pathways related to liver lipid metabolism[36]. Conversely, in the high hemicellulose level groups, the addition of 10 mg/kg DM oxalic acid significantly reduced glycinocholic acid, which has been linked to the expression of connective tissue growth factor in hepatocytes and activation of hepatic stellate cells[37]. Additionally, in the high hemicellulose group with no oxalic acid (0 mg/kg DM) oxalic acid group and high hemicellulose group with 10 mg/kg DM oxalic acid groups, there was a significant upregulation of ginkgolide B, a potent platelet-activating factor (PAF) receptor antagonist[38].
5. Conclusions
This study showed that the addition of oxalic acid to hemicellulosic diets increased rumen fermentation and affected microbial diversity and metabolism in vitro. These results were especially pronounced in the diet containing 17% hemicellulose. Supplementation with an appropriate amount of oxalic acid increased the breakdown of dry matter, neutral detergent fiber, and crude protein in the rumen. This supplementation significantly improved ruminal IVCPD, IVEED, ammonia nitrogen concentrations, and acetate, propionate, and TVFA production. These findings suggest that oxalic acid can play a significant role in fostering a favorable rumen environment, enhancing rumen fermentation efficiency, and facilitating hemicellulose digestion. However, to maximize these benefits, it is crucial to determine the optimal dosage of oxalic acid based on specific dietary conditions. Future studies should focus on identifying the ideal concentrations and exploring the long-term effects of oxalic acid supplementation on rumen health and animal performance.
Author Contributions
LZ: Writing - original draft, Data curation, Software, Validation,Visualization. TW,WZ: Writing – review & editing, Resources. ZW: Writing– review & editing, Investigation. YX: Writing – review & editing, Investigation. YP:Writing– Formal analysis, Methodology. MZ: Writing– Formal analysis, Methodology. HJ: Writing – review& editing, Investigation. GQ: Writing – review & editing, Supervision.YZ: Writing – review & editing, Formal analysis, Methodology. ZS: Writing – Funding acquisition, Supervision. XZ,WZ:Writing – review & editing, Formal analysis, Methodology.
Funding
The present work was funded by the Scientific and Technological Developing Scheme of Jilin Province (grant number 20230202075NC), Science and Technology Research Project of the Education Department of Jilin Province (JJKH20240465CY).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, the animal care committee of Jilin Agricultural University (JLAU-ACUC2024-011) approved the comprehensive process used in the management of the experimental animals.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found here: NCBI sequence Read Archive (SRA), accession numbers PRJNA1081127.
Conflicts of Interest
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| HM | Hemicellulose |
| OA | Oxalic acid |
| VFA | Volatile fatty acid |
| YC | Yeast culture |
| IVDMD | Dry matter digestibility |
| IVCPD | Crude protein digestibility |
| IVEED | Crude fat digestibility |
| IVNDFD | Neutral detergent fiber digestibility |
References
- Weimer, P.J. Degradation of Cellulose and Hemicellulose by Ruminal Microorganisms. Microorganisms 2022, 10, 2345. [Google Scholar] [CrossRef] [PubMed]
- Plata, F.P.; Bárcena-Gama, J.R. Effect of a Yeast Culture (Saccharomyces Cerevisiae) on Neutral Detergent Fiber Digestion in Steers Fed Oat Straw Based Diets. Animal Feed Science and Technology 1994, 49, 203–210. [Google Scholar] [CrossRef]
- Palmieri, F.; Estoppey, A.; House, G.L.; Lohberger, A.; Bindschedler, S.; Chain, P.S.; Junier, P. Oxalic Acid, a Molecule at the Crossroads of Bacterial-Fungal Interactions. Advances in applied microbiology 2019, 106, 49–77. [Google Scholar] [PubMed]
- Chen, X.; Xiao, J.; Zhao, W.; Li, Y.; Zhao, W.; Zhang, W.; Xin, L.; Han, Z.; Wang, L.; Aschalew, N.D.; et al. Mechanistic Insights into Rumen Function Promotion through Yeast Culture (Saccharomyces Cerevisiae) Metabolites Using in Vitro and in Vivo Models. Front Microbiol 2024, 15, 1407024. [Google Scholar] [CrossRef] [PubMed]
- Duraiswamy, A.; Sneha A, N.M.; Jebakani K, S.; Selvaraj, S.; Pramitha J, L.; Selvaraj, R.; Petchiammal K, I.; Kather Sheriff, S.; Thinakaran, J.; Rathinamoorthy, S.; et al. Genetic Manipulation of Anti-Nutritional Factors in Major Crops for a Sustainable Diet in Future. Front Plant Sci 2022, 13, 1070398. [Google Scholar] [CrossRef]
- Rahman, M.M.; Abdullah, R.B.; Wan Khadijah, W.E. A Review of Oxalate Poisoning in Domestic Animals: Tolerance and Performance Aspects. Journal of animal physiology and animal nutrition 2013, 97, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.N.; Sun, Y.R.; Tang, Y.M.; Xie, X.L.; Li, Q.Y.; Mou, F.Z.; Sun, H.X. Effect of High Oxalic Acid Intake on Growth Performance and Digestion, Blood Parameters, Rumen Fermentation and Microbial Community in Sheep. Small Ruminant Research 2024, 237, 107324. [Google Scholar] [CrossRef]
- James, L.F.; Butcher, J.E. Halogeton Poisoning of Sheep: Effect of High Level Oxalate Intake. Journal of Animal Science 1972, 35, 1233–1238. [Google Scholar] [CrossRef]
- Frutos, P.; Duncan, A.J.; Kyriazakis, I.; Gordon, I.J. Learned Aversion towards Oxalic Acid-Containing Foods by Goats: Does Rumen Adaptation to Oxalic Acid Influence Diet Choice? Journal of Chemical Ecology 1998, 24, 383–397. [Google Scholar] [CrossRef]
- Duncan, A.J.; Frutos, P.; Young, S.A. Rates of Oxalic Acid Degradation in the Rumen of Sheep and Goats in Response to Different Levels of Oxalic Acid Administration. Animal Science 1997, 65, 451–455. [Google Scholar] [CrossRef]
- Kh, M. Estimation of the Energetic Feed Value Obtained from Chemical Analysis and in Vitro Gas Production Using Rumen Fluid. Anim Res Dev 1988, 28, 7–55. [Google Scholar]
- Aschalew, N.D.; Zhang, L.; Wang, Z.; Xia, Y.; Yin, G.; Dong, J.; Zhen, Y.; Zhang, X.; Wang, T.; Sun, Z.; et al. Effects of Yeast Culture and Oxalic Acid Supplementation on in Vitro Nutrient Disappearance, Rumen Fermentation, and Bacterial Community Composition. Front. Vet. Sci. 2024, 10. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified Reagents for Determination of Urea and Ammonia. Clinical chemistry 1962, 8, 130–132. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Urriola, P.E.; Shurson, G.C. Use of in Vitro Dry Matter Digestibility and Gas Production to Predict Apparent Total Tract Digestibility of Total Dietary Fiber for Growing Pigs. Journal of animal science 2017, 95, 5474–5484. [Google Scholar] [CrossRef]
- DeFeo, M.E.; Shampoe, K.V.; Carvalho, P.H.; Silva, F.A.; Felix, T.L. In Vitro and in Situ Techniques Yield Different Estimates of Ruminal Disappearance of Barley. Translational Animal Science 2020, 4, 141–148. [Google Scholar] [CrossRef]
- Goeser, J.P.; Combs, D.K. An Alternative Method to Assess 24-h Ruminal in Vitro Neutral Detergent Fiber Digestibility. Journal of dairy science 2009, 92, 3833–3841. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.; Stewart, C.J. Microbiota Analysis Using Sequencing by Synthesis: From Library Preparation to Sequencing. Methods Mol Biol 2020, 2121, 165–184. [Google Scholar] [CrossRef]
- Aschalew, N.D.; Wang, T.; Qin, G.X.; Zhen, Y.G.; Zhang, X.F.; Chen, X.; Atiba, E.M.; Seidu, A. Effects of Physically Effective Fiber on Rumen and Milk Parameters in Dairy Cows: A Review. Indian Journal of Animal Research 2020, 54. [Google Scholar]
- Plaizier, J.C.; Krause, D.O.; Gozho, G.N.; McBride, B.W. Subacute Ruminal Acidosis in Dairy Cows: The Physiological Causes, Incidence and Consequences. The Veterinary Journal 2008, 176, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Belenguer, A.; Bati, M.B.; Hervás, G.; Toral, P.G.; Yáñez-Ruiz, D.R.; Frutos, P. Impact of Oxalic Acid on Rumen Function and Bacterial Community in Sheep. animal 2013, 7, 940–947. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Yang, S.; Deng, J.P.; Tang, S.X.; Tan, Z.L. Effects of Three Methane Mitigation Agents on Parameters of Kinetics of Total and Hydrogen Gas Production, Ruminal Fermentation and Hydrogen Balance Using in Vitro Technique. Anim Sci J 2016, 87, 224–232. [Google Scholar] [CrossRef]
- Yin, G.; Sun, Z.; Wang, Z.; Xia, Y.; Cheng, L.; Qin, G.; Aschalew, N.D.; Liu, H.; Zhang, X.; Wu, Q.; et al. Mechanistic Insights into Inositol-Mediated Rumen Function Promotion and Metabolic Alteration Using in Vitro and in Vivo Models. Front Vet Sci 2024, 11, 1359234. [Google Scholar] [CrossRef]
- Tan, P.; Liu, H.; Zhao, J.; Gu, X.; Wei, X.; Zhang, X.; Ma, N.; Johnston, L.J.; Bai, Y.; Zhang, W.; et al. Amino Acids Metabolism by Rumen Microorganisms: Nutrition and Ecology Strategies to Reduce Nitrogen Emissions from the inside to the Outside. Science of The Total Environment 2021, 800, 149596. [Google Scholar] [CrossRef]
- Rodrigues, A.A.; Reis, S.S.; Costa, D.C. da C.C.; Dos Santos, M.A.; Paulino, R. de S.; Rufino, M. de O.A.; Neto, S.G. Yeast-Fermented Cassava as a Protein Source in Cattle Feed: Systematic Review and Meta-Analysis. Trop Anim Health Prod 2023, 55, 67. [Google Scholar] [CrossRef]
- Ortega, M.E.; Stern, M.D.; Satter, L.D. The Effect of Rumen Ammonia Concentration on Dry Matter Disappearance in Situ. J. Dairy Sci 1979, 62, 76. [Google Scholar]
- Keim, J.P.; Alvarado-Gilis, C.; Arias, R.A.; Gandarillas, M.; Cabanilla, J. Evaluation of Sources of Variation on in Vitro Fermentation Kinetics of Feedstuffs in a Gas Production System. Anim Sci J 2017, 88, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Shrivastava, B.; Kumar, N.; Dhewa, T.; Sahay, H. Microbial Feed Additives. Rumen Microbiology: From Evolution to Revolution 2015, 161–175.
- Mahayri, T.M.; Fliegerová, K.O.; Mattiello, S.; Celozzi, S.; Mrázek, J.; Mekadim, C.; Sechovcová, H.; Kvasnová, S.; Atallah, E.; Moniello, G. Host Species Affects Bacterial Evenness, but Not Diversity: Comparison of Fecal Bacteria of Cows and Goats Offered the Same Diet. Animals 2022, 12, 2011. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, J.; Qin, Y.; Wang, Y.; Yu, Z.; Ning, X.; Sun, S. Specific Alterations of Gut Microbiota in Patients with Membranous Nephropathy: A Systematic Review and Meta-Analysis. Frontiers in Physiology 2022, 13, 909491. [Google Scholar] [CrossRef]
- Pinnell, L.J.; Reyes, A.A.; Wolfe, C.A.; Weinroth, M.D.; Metcalf, J.L.; Delmore, R.J.; Belk, K.E.; Morley, P.S.; Engle, T.E. Bacteroidetes and Firmicutes Drive Differing Microbial Diversity and Community Composition Among Micro-Environments in the Bovine Rumen. Front Vet Sci 2022, 9, 897996. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Zhang, X.X.; Li, F.D.; Li, C.; Li, G.Z.; Zhang, D.Y.; Song, Q.Z.; Li, X.L.; Zhao, Y.; Wang, W.M. Characterization of the Rumen Microbiota and Its Relationship with Residual Feed Intake in Sheep. Animal 2021, 15, 100161. [Google Scholar] [CrossRef] [PubMed]
- Strobel, H.J. Pentose Utilization and Transport by the Ruminal Bacterium Prevotella Ruminicola. Arch Microbiol 1993, 159, 465–471. [Google Scholar] [CrossRef]
- Jensen, S.K.; Lauridsen, C. Alpha-Tocopherol Stereoisomers. Vitam Horm 2007, 76, 281–308. [Google Scholar] [CrossRef]
- Clay, R.; Siddiqi, S.; Siddiqi, S.A. α-Tocopherol Reduces VLDL Secretion through Modulation of Intracellular ER-to-Golgi Transport of VLDL. Can J Physiol Pharmacol 2023, 101, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, S.L.; Baldeon, M.E.; Sangronis, E.; Petruschina, A.C.; Reyes, F.G.R. L-Glutamate: A Key Amino Acid for Senory and Metabolic Functions. Arch Latinoam Nutr 2016, 66, 101–112. [Google Scholar] [PubMed]
- Arshad, U.; Zenobi, M.G.; Tribulo, P.; Staples, C.R.; Santos, J.E.P. Dose-Dependent Effects of Rumen-Protected Choline on Hepatic Metabolism during Induction of Fatty Liver in Dry Pregnant Dairy Cows. PLoS One 2023, 18, e0290562. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Wang, J.; Zhang, H.; Chai, Y.; Xu, Y.; Miao, Y.; Yuan, Z.; Zhang, L.; Jiang, Z.; Yu, Q. Glycocholic Acid Aggravates Liver Fibrosis by Promoting the Up-Regulation of Connective Tissue Growth Factor in Hepatocytes. Cell Signal 2023, 101, 110508. [Google Scholar] [CrossRef]
- Shang, Q.; Zhou, X.; Yang, M.R.; Lu, J.-G.; Pan, Y.; Zhu, G.-Y.; Jiang, Z.-H. Amide Derivatives of Ginkgolide B and Their Inhibitory Effects on PAF-Induced Platelet Aggregation. ACS Omega 2021, 6, 22497–22503. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of oxalic acid (OA) supplementation in different level hemicellulose (HM) diets on volatile fatty acid (VFA) production (mmol/L) in vitro :(A) effect of acetic acid, propionic acid, butyric acid, isobutyric acid, isovaleric acid, valeric acid, total acid, and the acetic acid to propionic acid ratio at 6 h. (B) effect of acetic acid, propionic acid, butyric acid, isobutyric acid, isovaleric acid, valeric acid, total acid, and the acetic acid to propionic acid ratio at 12 h.
Figure 1.
Effects of oxalic acid (OA) supplementation in different level hemicellulose (HM) diets on volatile fatty acid (VFA) production (mmol/L) in vitro :(A) effect of acetic acid, propionic acid, butyric acid, isobutyric acid, isovaleric acid, valeric acid, total acid, and the acetic acid to propionic acid ratio at 6 h. (B) effect of acetic acid, propionic acid, butyric acid, isobutyric acid, isovaleric acid, valeric acid, total acid, and the acetic acid to propionic acid ratio at 12 h.
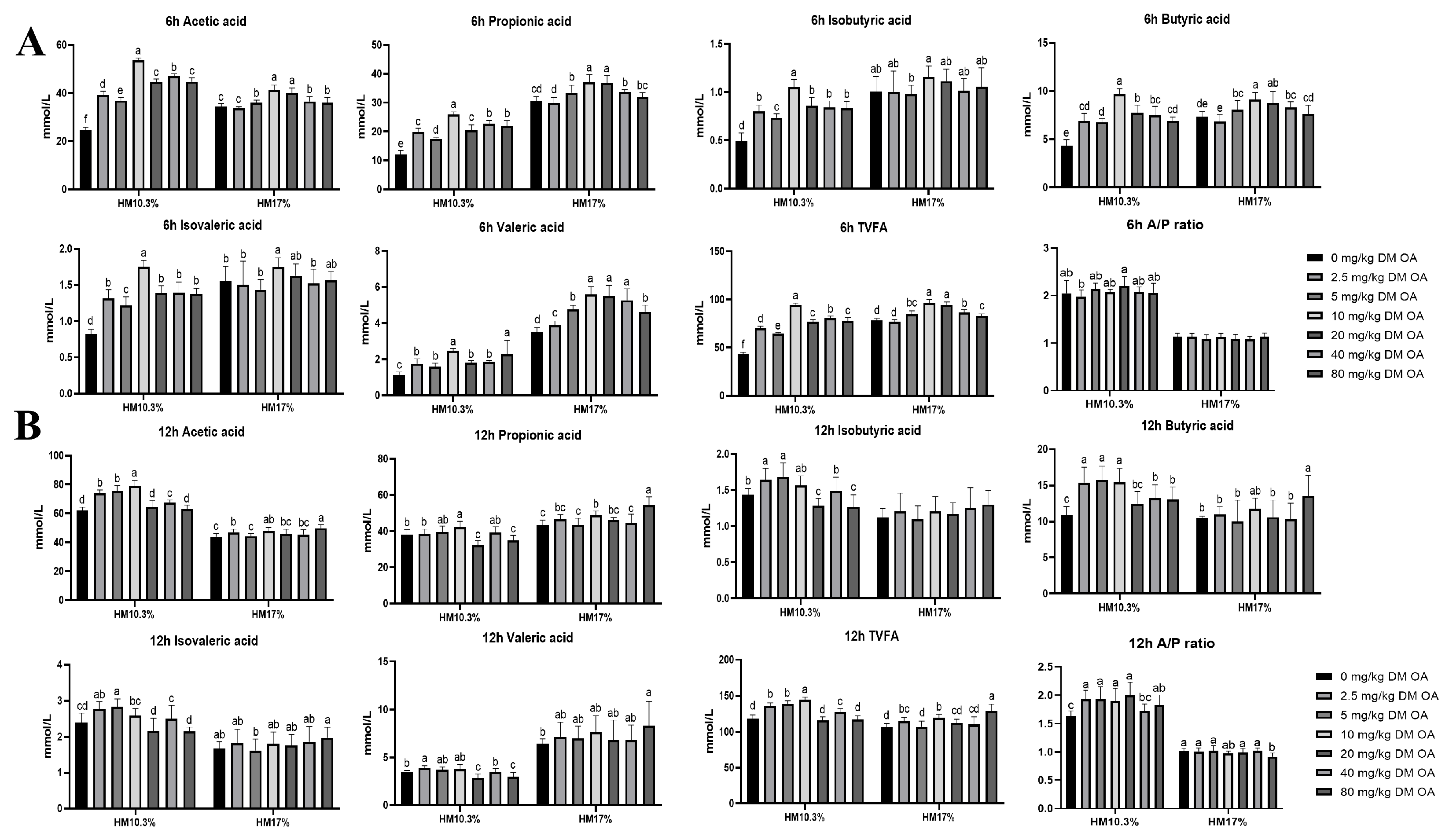
Figure 2.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on rumen microorganisms in vitro : (A) The abundance of bacteria following in vitro fermentation; (B) the abundance of in vitro fermentative bacteria; (C) the 12-h Alpha diversity index of in vitro fermentation; (D) the beta diversity index of in vitro fermentation. D1-0 is the low-hemicellulose 0 mg/kg DM oxalic acid addition group, D1-10 was the low-hemicellulose 10 mg/kg DM oxalic acid addition group, D2-0 was the high-hemicellulose 0 mg/kg DM oxalic acid addition group, and D2-10 was the high-hemicellulose 10 mg/kg DM oxalic acid addition group.
Figure 2.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on rumen microorganisms in vitro : (A) The abundance of bacteria following in vitro fermentation; (B) the abundance of in vitro fermentative bacteria; (C) the 12-h Alpha diversity index of in vitro fermentation; (D) the beta diversity index of in vitro fermentation. D1-0 is the low-hemicellulose 0 mg/kg DM oxalic acid addition group, D1-10 was the low-hemicellulose 10 mg/kg DM oxalic acid addition group, D2-0 was the high-hemicellulose 0 mg/kg DM oxalic acid addition group, and D2-10 was the high-hemicellulose 10 mg/kg DM oxalic acid addition group.
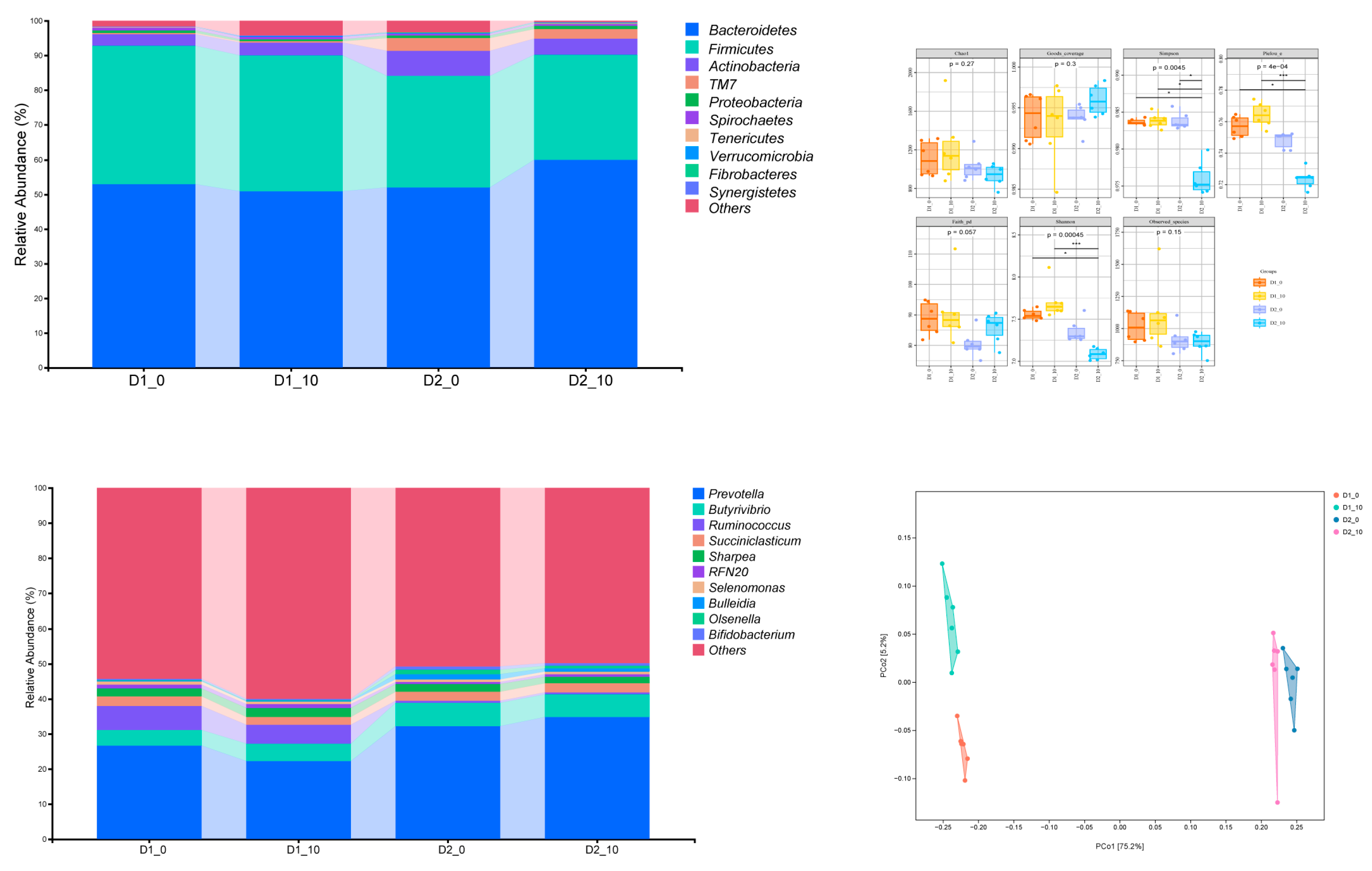
Figure 3.
The correlation between rumen fermentation parameters and bacterial genus composition : (A, B) The correlation analyses of rumen microorganisms and fermentation parameters with oxalic acid (OA) supplementation at the low hemicellulose (HM10.3%) level;( C,D) the correlation analyses of rumen microorganisms and fermentation parameters with OA supplementation at the high hemicellulose (HM17%) level.
Figure 3.
The correlation between rumen fermentation parameters and bacterial genus composition : (A, B) The correlation analyses of rumen microorganisms and fermentation parameters with oxalic acid (OA) supplementation at the low hemicellulose (HM10.3%) level;( C,D) the correlation analyses of rumen microorganisms and fermentation parameters with OA supplementation at the high hemicellulose (HM17%) level.
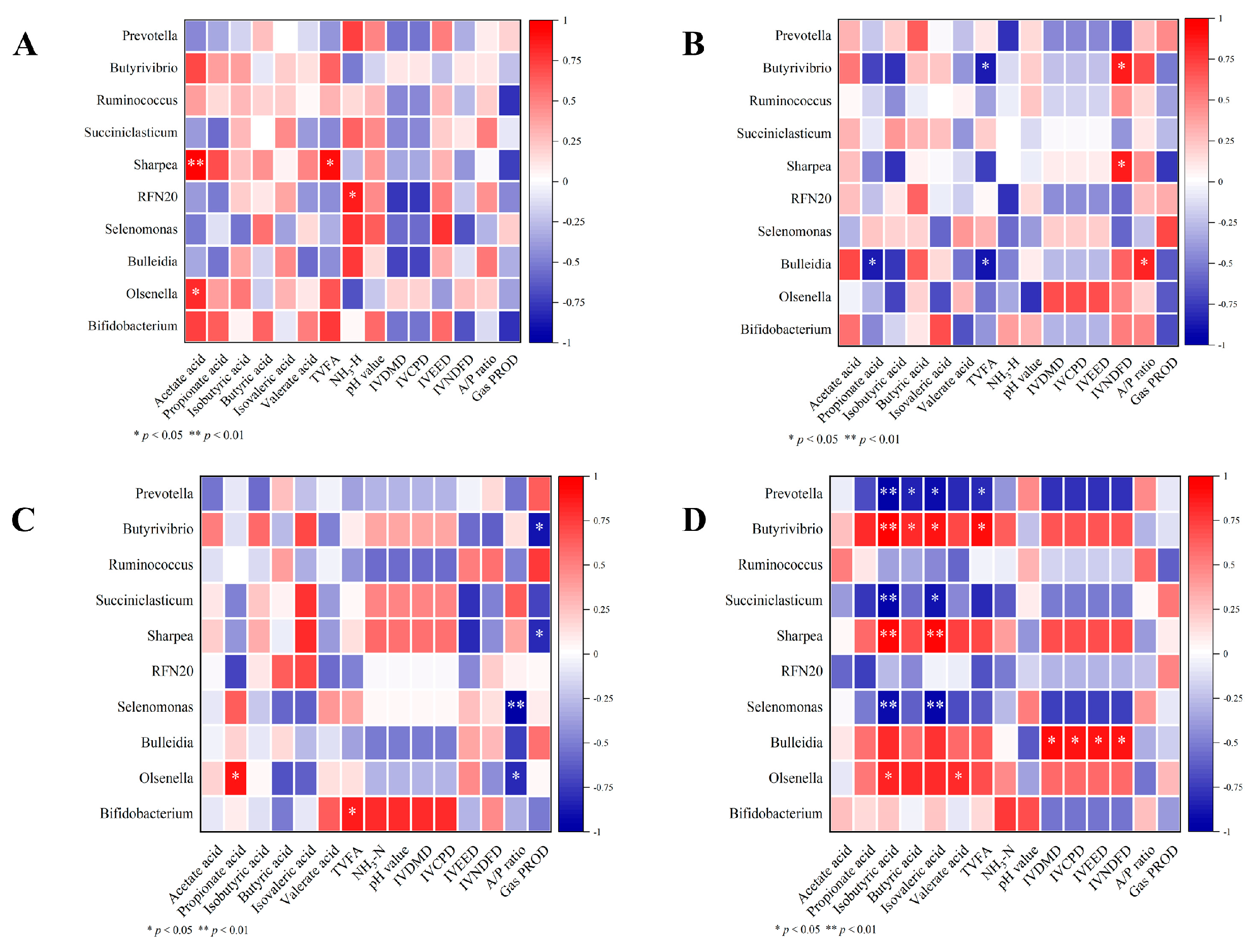
Figure 4.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on rumen metabolites in vitro : (A) metabolite classification statistics; the differential metabolites and metabolic pathway analysis of OA added at the low HM level (B, F); the differential metabolites and metabolic pathway analysis of OA supplementation at high HM level (C, G); different metabolites and metabolic pathway analysis without OA at high and low HM levels (D, H); the differential metabolites and metabolic pathway analysis of OA supplementation at both high and low HM levels (E, I).
Figure 4.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on rumen metabolites in vitro : (A) metabolite classification statistics; the differential metabolites and metabolic pathway analysis of OA added at the low HM level (B, F); the differential metabolites and metabolic pathway analysis of OA supplementation at high HM level (C, G); different metabolites and metabolic pathway analysis without OA at high and low HM levels (D, H); the differential metabolites and metabolic pathway analysis of OA supplementation at both high and low HM levels (E, I).
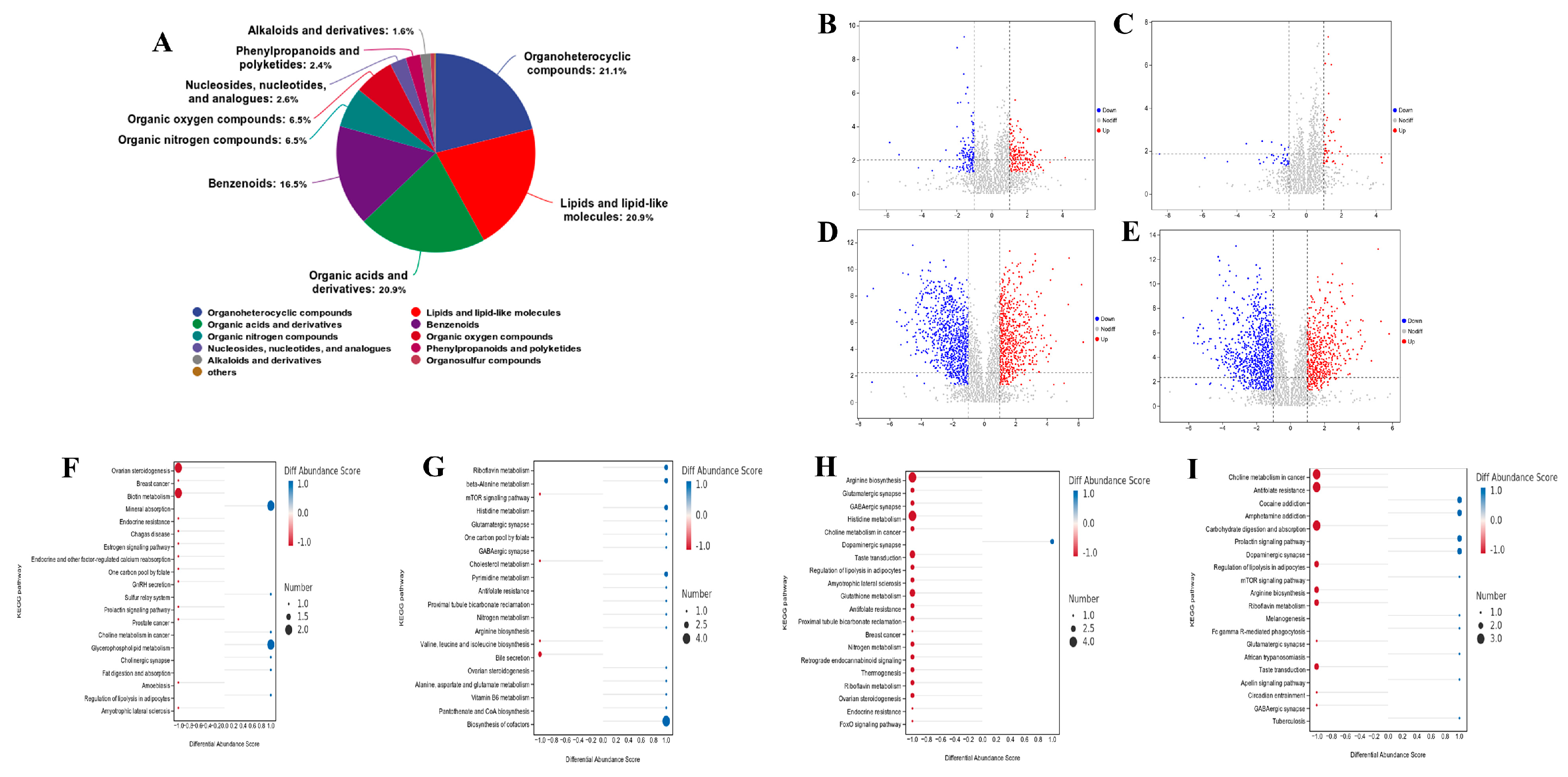
Figure 5.
The correlation between rumen metabolites and bacterial genera composition : (A) The effect of oxalic acid addition on correlations at the low hemicellulose level; (B) the effect of oxalic acid addition on correlations at the high hemicellulose level.
Figure 5.
The correlation between rumen metabolites and bacterial genera composition : (A) The effect of oxalic acid addition on correlations at the low hemicellulose level; (B) the effect of oxalic acid addition on correlations at the high hemicellulose level.
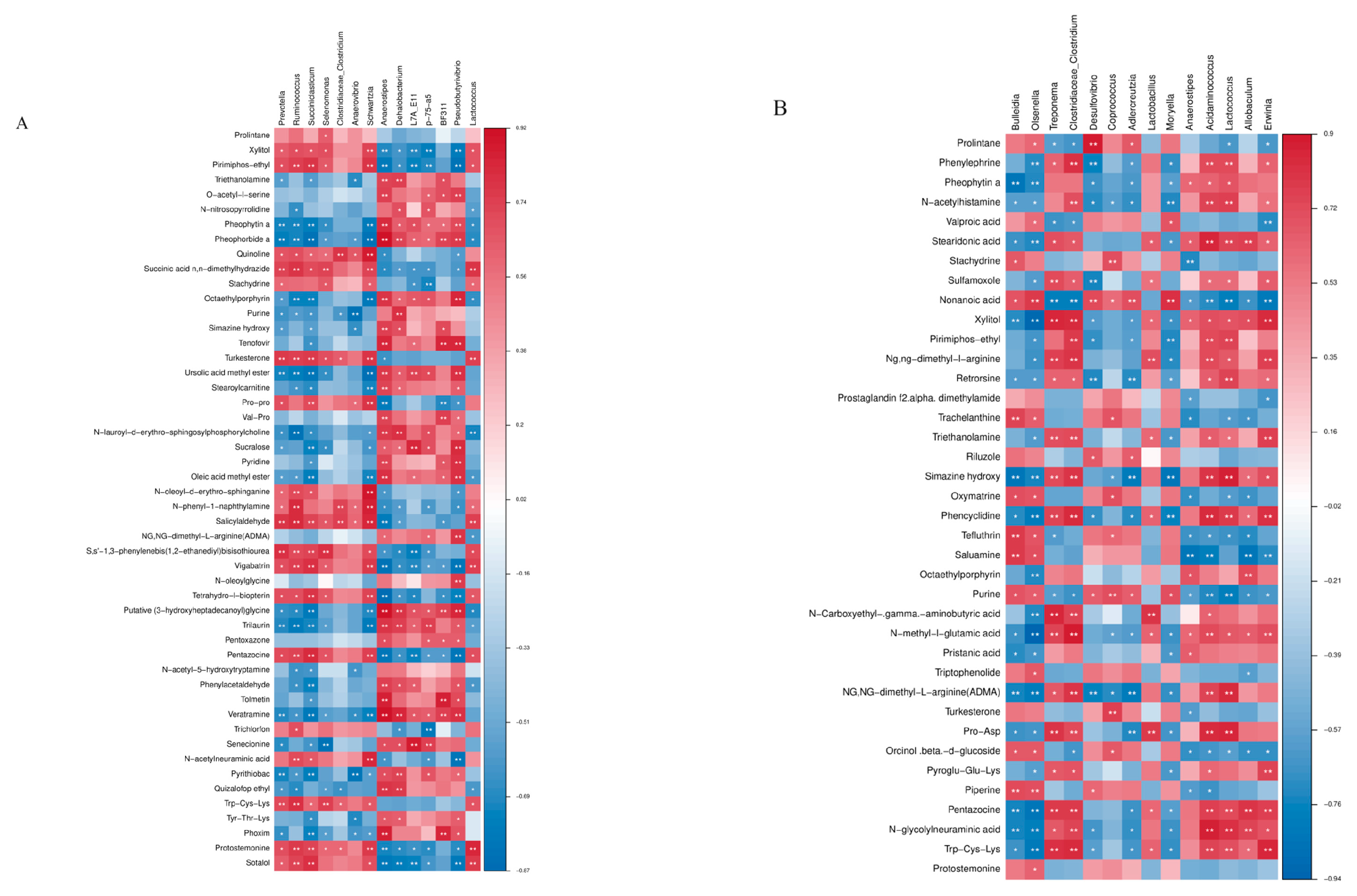

Table 1.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on nutritional digestibility in vitro.
Table 1.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on nutritional digestibility in vitro.
| HM10.3% | HM17% | P-value | |||||||||||||||||
| Item(%) | Time(h) | 0 mg/kg | 2.5 mg/kg | 5 mg/kg | 10 mg/kg | 20 mg/kg | 40 mg/kg | 80 mg/kg | 0 mg/kg | 2.5 mg/kg | 5 mg/kg | 10 mg/kg | 20 mg/kg | 40 mg/kg | 80 mg/kg | SEM | HM | OA | HM×OA |
| IVDMD | 6 | 28.29 | 30.90 | 30.07 | 30.59 | 30.24 | 30.03 | 29.24 | 29.84 | 28.87 | 8.99 | 27.94 | 28.24 | 29.56 | 26.59 | 0.27 | 0.01 | 0.49 | 0.33 |
| 12 | 35.87 | 35.88 | 36.63 | 36.69 | 36.94 | 35.78 | 35.60 | 34.28 | 33.46 | 34.84 | 33.96 | 33.72 | 34.04 | 33.91 | 0.28 | <0.01 | 0.95 | 0.98 | |
| IVCPD | 6 | 23.10c | 30.77a | 29.83ab | 29.41ab | 30.93a | 31.45a | 27.23b | 33.60a | 31.01ab | 31.97a | 31.08ab | 27.22bc | 32.37a | 25.08c | 0.34 | <0.01 | <0.01 | <0.01 |
| 12 | 38.67b | 38.39b | 38.28b | 43.89a | 40.72ab | 40.70ab | 45.08a | 40.47a | 36.75b | 40.44a | 37.72ab | 35.36b | 37.01b | 35.74b | 0.34 | <0.01 | 0.01 | <0.01 | |
| IVEED | 6 | 18.63cd | 27.37a | 19.24c | 18.08d | 20.98b | 28.10a | 16.81e | 27.89c | 30.69b | 31.57ab | 23.36d | 21.59e | 30.87b | 31.96a | 0.47 | <0.01 | <0.01 | <0.01 |
| 12 | 29.05a | 23.25c | 23.65c | 25.63b | 26.50b | 18.50d | 26.43b | 29.10c | 31.37b | 16.84f | 37.82a | 29.33c | 24.86e | 27.48d | 0.44 | <0.01 | <0.01 | <0.01 | |
| IVNDFD | 6 | 12.24c | 15.43a | 15.31a | 13.42b | 12.62bc | 12.67bc | 14.53a | 7.53ab | 8.51a | 6.05ab | 6.47ab | 7.14ab | 7.99ab | 4.79b | 0.36 | <0.01 | <0.01 | <0.01 |
| 12 | 21.53d | 22.31c | 22.92c | 23.59b | 24.62a | 21.07d | 20.30e | 12.10bc | 12.59bc | 16.74a | 13.41bc | 11.25c | 11.09c | 14.71ab | 0.45 | <0.01 | <0.01 | <0.01 | |
Table 2.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on rumen fermentation parameters in vitro .
Table 2.
Effects of oxalic acid (OA) supplementation in different hemicellulose (HM) diets on rumen fermentation parameters in vitro .
| HM10.3% | HM17% | P-value | |||||||||||||||||
| Item | Time (h) | 0 mg/kg | 2.5 mg/kg | 5 mg/kg | 10 mg/kg | 20 mg/kg | 40 mg/kg | 80 mg/kg | 0 mg/kg | 2.5 mg/kg | 5 mg/kg | 10 mg/kg | 20 mg/kg | 40 mg/kg | 80 mg/kg | SEM | HM | OA | HM×OA |
| NH3-N (mg/dL) |
6 | 17.97d | 19.77abc | 21.08a | 20.32ab | 18.50cd | 18.91bcd | 20.20ab | 22.06ab | 20.47c | 19.60c | 23.46a | 22.52a | 19.96c | 20.94bc | 0.18 | <0.01 | <0.01 | <0.01 |
| 12 | 28.70a | 26.05b | 28.23a | 22.87c | 23.12c | 24.30c | 23.80c | 25.04 | 25.85 | 25.33 | 25.38 | 25.09 | 25.63 | 25.63 | 0.19 | 0.672 | <0.01 | <0.01 | |
| Gas PROD (mL) |
6 | 50.94cd | 57.26bc | 61.34b | 59.95b | 46.76d | 135.12a | 51.8cd | 65.2d | 67.45d | 64.45d | 79.36c | 101.25b | 129.86a | 108.92b | 4.48 | <0.01 | <0.01 | <0.01 |
| 12 | 100.05d | 153.35b | 170.94a | 149.92b | 113.46c | 108.31cd | 118.39c | 85.04c | 152.92a | 113.24b | 136.33a | 112.06b | 93.73c | 145.5a | 4.04 | <0.01 | <0.01 | <0.01 | |
| pH value |
6 | 7.10a | 7.07ab | 7.06ab | 7.04ab | 7.04ab | 7.01b | 7.01b | 6.95 | 6.96 | 6.95 | 6.95 | 6.90 | 6.91 | 6.95 | 0.02 | <0.01 | 0.89 | 0.99 |
| 12 | 6.88 | 6.83 | 6.80 | 6.80 | 6.81 | 6.81 | 6.77 | 6.73 | 6.75 | 6.71 | 6.69 | 6.71 | 6.72 | 6.69 | 0.02 | <0.01 | 0.94 | 0.99 | |
| TVFA (mmol/L) |
6 | 43.45f | 69.77d | 64.53e | 94.61a | 76.85c | 80.75b | 78.03c | 78.38d | 76.79d | 84.81bc | 96.19a | 94.00a | 86.36b | 82.78c | 1.14 | <0.01 | <0.01 | <0.01 |
| 12 | 118.36d | 135.83b | 138.92b | 144.61a | 115.59d | 127.38c | 117.32d | 107.10d | 114.62bc | 107.33d | 119.02b | 112.33cd | 110.32cd | 29.06a | 1.09 | <0.01 | <0.01 | <0.01 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



