
Submitted:
26 December 2024
Posted:
26 December 2024
You are already at the latest version
Abstract
Data from the literature and our recent studies have shown that dysfunction/deficiency of melatonin accelerates aging or delays lifespan in an age-dependent manner. In the present study, we aimed to determine the effects of pinealectomy on sphingolipid turnover in young adult (3-month-old), middle-aged (14-month-old), and old (18-month-old) rats. Ceramide levels, neutral and acid sphingomyelinase, acid ceramidase and sphingosine-1-phosphate levels in hippocampus and/or plasma, were evaluated by enzyme-linked immunosorbent assay Accumulation of Ceramide and its metabolite second messenger sphingo-sine-1-phosphate in the hippocampus and plasma was associated with increased levels and activity of neutral sphingomyelinase in the hippocampus and plasma, respectively, but not of acid sphingomyelinase, which levels and activity in the hippocampus were reduced in middle-aged and old rats compared to young adult rats. Similar to the aging process, pinealectomy-induced melatonin deficiency increased Ceramide content in the hippocampus of young adult rats compared to the sham group. Pinealectomy also exacerbated an age-related increase in sphingosine-1-phosphate in the hippocampus of 18-month-old rats. However, compared to young adult rats, pinealectomy exerted the opposite effect on age-related changes in Ceramide, neutral, and acid sphingomyelinases in the hippocampus in middle-aged rats. Our results suggest that melatonin deficiency may modulate aging by altering sphingolipid turnover in an age-specific manner.
Keywords:
pinealectomy
; aging
; sphingolipid turnover
; ceramide
; rat
1. Introduction
The world's population is rapidly aging, and it is predicted that by 2050, individuals aged 60 and over will comprise 20-25% of the total population. Age-related neurological complications are common and affect an individual's quality of life. Therefore, understanding the key mechanisms associated with age-related changes, as well as the factors that trigger these inevitable destructive events in brain structure and function, are essential for identifying new therapeutic approaches to meet the needs of aging population. This elucidation will contribute to the development of multimodal health strategies to increase quality of life and lifespan.
The pineal gland, often considered as a key regulator of circadian rhythms, is thought to play a central role in modulating growth, fertility, aging and death [1]. Some studies suggest that the gland follows a biologically "programmed" timeline, particularly influenced by the secretion of melatonin, a hormone known to regulate sleep-wake cycles and possess antioxidant properties. As we age, the pineal gland becomes calcified and melatonin production decreases significantly [2,3,4]. This decline is associated with several aging-related processes, including decreased antioxidant defense system, weakened immune function, and disruptions in circadian rhythms. Decreased melatonin is thought to accelerate aging at the cellular level, leading to the gradual onset of age-related diseases such as neurodegenerative disorders. Accordingly, this gland is seen as a "clock" that "tracks and regulates" the ontogenetic phases of our genetically inherited "program" of life.
On the other hand, the alterations in the sphingolipid (SL) metabolism leading to oxidative dysfunction are critical to age-related processes and diseases [5,6]. There are few data in the scientific literature indicating a link between the melatonin system and SL signaling [7,8]. Elucidating the cellular and molecular mechanisms underlying the most fundamental aspect of our life program will help to develop a rational strategy to improve the health status of the elderly. SLs are a very diverse group of amphipathic lipids that are abundant in cell membranes. They can be classified into two main classes according to the type of polar group in their structure: phosphosphingolipids (PSLs), including sphingomyelin (SM), and glycosphingolipids (GSLs) – cerebrosides and gangliosides [9]. SLs play an important structural role in biological membranes, including neuronal membranes, and are precursors of many bioactive metabolites that regulate various cellular functions including senescence regulation which is critical in aging [6]. In this respect the finely tuned balance between the synthesis and degradation of SLs is normally critical for scores of biological processes [10], so changes in their metabolism can affect homeostasis and brain function.
Sphingomyelin is a major source of ceramides (Cer), that are lipid second messengers formed when SM is cleaved by neutral or acid sphingomyelinases (NSMase and ASMase). The latter enzymes are activated by inflammatory cytokines and oxidative stress (OS) [11] or inhibition of Cer metabolism enzymes such as SM synthase, which converts Cer to SM, and acid ceramidase (ASAH1), which hydrolyzes Cer to sphingosine 1-phosphate (S1P) [12]. Ceramide is an important bioactive molecule involved in the regulation of several physiological processes, including cell proliferation, differentiation, and programmed cell death (apoptosis) [13]. The induced conversion of SM to Cer in many cell types, including neurons, results in toxicity expressed as pro-apoptotic effects. The consequence of this process is tissue damage and organ dysfunction. In parallel, the decreased level of cellular S1P has been shown to promote cellular senescence [6]. On the other hand, low levels of Cer and high level of S1P have trophic effects and promote survival after cell division. The differential regulation of opposing pathways between Cer and S1P is known as the “SL Rheostat” demonstrating Ce’s ability to induce cell growth arrest and apoptosis, while S1P is responsible for optimal cell proliferation and growth as well as the suppression of ceramide mediated growth arrest and apoptosis [14]. One of the major activators of Cer generation is OS caused by reactive oxygen and nitrogen species. A link between aging tissue and NSMase activity has also been reported [15] along with a correlation between NSMase activity and cellular senescence [16]. Although ASMase is more active than NMase, higher activity of both enzymes was observed in the liver of aged rats than in young ones [17]. These enzymes are considered biomarkers of aging because their levels increase with age in mammals and are thought to play a role in age-related diseases, including type 2 diabetes, cardiovascular and immune dysfunction, cancer, and neurodegeneration. However, Cer may not be the only contributing factor to the induction of senescence and aging. ASAH1 which catalyzes Cer into fatty acid and sphingosine has also been shown to be highly upregulated in senescent cells, and silencing ASAH1 in presence of human fibroblasts decreased the expression levels of senescent associated proteins P53, P21, and P16 [18]. The close association between Cer accumulation, increased OS and insulin resistance is intriguing, as these changes are thought to accelerate aging and age-related diseases. In adult rats, accumulation of specific Cer species in mitochondria has been found, a process that correlates with impaired function of complex IV in the electron transport chain [19] and corresponds to increased reactive oxygen species (ROS) during aging. There is a negative correlation between glutathione levels and mitochondrial NSMase activity, suggesting that the therapeutic effect of antioxidant molecules works by preventing NSMase activation and Cer accumulation by maintaining normal glutathione levels. These data support the idea that Cer accumulation is a contributing factor in age-related diseases and that strategies to prevent such accumulation may potentially reduce the incidence and severity of these diseases. Melatonin has been shown to suppress ASMase in the hippocampus of mice and cell lines in a manner similar to antidepressants, while the diurnal regulation of S1P and sphinganine 1-phosphate is dependent on intact melatonin signalling [7].
Previous publications provide a basis for formulating a hypothesis that there is a functional link between the melatonin system and SL pathways that is critical for aging processes, including impaired homeostasis of oxidative status in the central nervous system (CNS). The work of Pierpaoli and Bulian [20] has shown that the role of melatonin in aging is strictly age-specific. This prompted us to plan a comparative analysis of SL metabolism in sham-operated and pinealectomized rats at 3, 14, and 18 months of age, respectively. At these ages, melatonin deficiency was found to accelerate, slow, or have no effect on the aging process in mice and rats [20,21,22]. The relationship between the pineal hormone and the SL pathway in the mechanisms mediating the aging process was investigated by examining the levels of key metabolites of SM such as Cer, S1P, along with enzymes involved in their processing, namely NSMase, ASMase, and ASAH1 at specific age periods. The changes in the oxidative status in the CNS are directly dependent on the perturbation in the SM metabolism during aging, but the role of melatonin deficiency in modulating these processes in an ontogenetic aspect, which we believe is a critical factor in aging, will be clarified in this study.
2. Results
2.1. Melatonin Deficiency Associated with Hormonal Dysfunction (Pinealectomy) Can Modify Age-Related Cer Increase in the Hippocampus but Not in Plasma
We found that aging was associated with increased levels of SM (p < 0.001, 18-month-old vs. 3-month-old sham rats) and Cer levels (p = 0.01, 18-month-old vs. 3-month-old sham rats) in the hippocampus (Figure 1, Figure 2A). Ceramide levels were also increased in the plasma of 18-month-old rats compared to 3-month-old animals (p = 0.042, 18-month-old vs. 3-month-old sham rats) (Figure 2B). The present results demonstrate that pinealectomy is able to modify Cer levels at different age stages in a complex manner. Thus, while pinealectomy in young adult rats resulted in increased Cer in the hippocampus (p = 0.036, 3pinealectomized (pin) rats vs. matched sham rats), melatonin deficiency in the middle-aged rats was associated with decreased Cer content (p = 0.006, 14-month-old pin rats vs. sham rats) (Figure 2A).
2.2. Melatonin Deficiency Reversed Age-Related Decreases in ASAH1 Levels in the Hippocampus but not in the Plasma of 14-Month-old Rats
ASAH1 is also involved in the control of Cer levels by hydrolyzing Cer to sphingosine. There was no significant difference between the sham and pin groups for ASAH1 levels in both the hippocampus and plasma (Figure 3A,B).
Considering that SM catabolism might be the main source of cellular Cer, we further investigated NSMase levels (hippocampus) and activity (plasma) and ASMase levels and activity (hippocampus) of young adult, middle-aged and old sham rats and rats with pinealectomy. Old sham rats showed increased level of NSMase in the hippocampus (p = 0.01, 18-month-old vs 3-month-old sham rats) whereas the activity of the enzyme in plasma showed only slight tendency of increase with the age- (Figure 4A,B). Removal of the pineal gland reduced NSMase levels to those of the control in the middle-aged rats (p < 0.001, 14-month-old pin rats vs sham rats) (Figure 4A).
In contrast to NSMase, which content in the hippocampus increased with age, ASMase exhibited the opposite age-related changes. ASMase levels were decreased in middle-aged and old sham rats compared to young adult rats (p = 0.002 and p = 0.005, respectively) (Figure 5А). Accordingly, the 3- and 14-month-old sham rats showed higher ASMase activity than the 18-month-old matched control (p <0.001, 3- and 14-month-old vs 18-month-old sham rats) (Figure 5B). 5Pinealectomy restored the enzyme levels in the hippocampus to the control levels in the middle-aged rats (p = 0.002, 14-month-old pin rats vs matched sham rats) (Figure 5A).
S1P is a product of Cer degradation by the action of ceramidases. Like its precursor, this signaling molecule was elevated in middle-aged and old rats compared to young adult rats (p < 0.001) (Figure 6). Pinealectomy affected only 18-month-old rats, which showed higher S1P levels than their sham-matched counterparts (p = 0.007).
3. Discussion
The present findings revealed that melatonin deficiency associated with hormonal dysfunction induces an age-specific alteration in SL metabolism in the hippocampus and plasma in rats. It is well-known that neuronal function is closely related to the homeostasis of SLs in both animals and humans while aging is vulnerable to dysregulation of SL metabolism and thus the abnormalities affect hippocampus-based memory. In this study, we showed that old rats had increased levels of SM, Cer and Cer-derivative S1P in both the hippocampus and plasma compared to young adult rats, which was associated with increased activity and content of NSMase, while ASMase activity and levels in the hippocampus were decreased. Excessive accumulation of Cer and its derivative S1P, particularly in the hippocampus, is associated with neurodegeneration, inflammation and apoptosis, which are hallmarks of age-related neurological disorders. Dysregulation in the metabolism of Cer and SM, which are key components of SL signaling, can accelerate the aging process and trigger neurodegeneration, inflammation, and apoptosis [11]. Our results are partly in agreement with the report of Babenko et al.[23], who reported that 24-month-old rats had increased Cer content and Cer/SM ratio in the hippocampus and neocortex compared to 3-month-old rats. Furthermore, these changes were associated with increased NSMase, but not ASMase activity. The cellular Cer can be generated by the hydrolysis of SM catalyzed by SMases. If ASMase activity decreases, it could reduce the production of Cer under acidic conditions. This reduction could counterbalance the amount of Cer generated by NSMase and prevent its overproduction in the aging brain. Therefore, reduced activity of ASMase could be suggested to have an indirect protective effect on the hippocampus by preventing the overproduction of Cer that is under NSMase control in aging. Conversely, activated NSMase leading to elevated Cer levels can induce apoptosis and inflammation, disrupt membrane integrity and signaling pathways critical for cellular functions [24], and can disrupt the balance between endogenous antioxidant molecules such as glutathione and hence the production of free radicals during metabolic activity [25].
Aging is associated with a decrease in glutathione (GSH) levels, leading to OS and activation of NSMase, which hydrolyzes SM to Cer [26]. Chronic inflammation and oxidative damage in the aging brain further enhance NSMase activity. Aging-associated metabolic and inflammatory stress may enhance de novo Cer synthesis from serine and palmitoyl-CoA. This pathway may become overactive in response to insulin resistance or mitochondrial dysfunction, both of which are common in aging. Therefore, our results indicate that NSMase plays a critical role in the elevation of Cer and increase in SM content in the hippocampus of aged rats. In aged brains, OS potentiates NSMase activity and enhances this effect. Other potential mechanisms that could stimulate the accumulation of Cer and SM in the hippocampus of aged rats could be related to a disrupted (upregulated) de novo pathway for Cer synthesis due to cellular stress and chronic inflammation, which are common in the aging brain. The hippocampus is particularly sensitive to Cer-induced neurotoxicity, affecting memory and learning, and our preliminary data confirmed that old rats showed decreased cognitive capacity [27]. While Cer accumulation is more commonly associated with aging, increased SM levels have also been reported as a potential compensatory response to elevated Cer by converting Cer back into SM [28]. Dysregulated NSMase activity in the plasma in our case could lead to inefficient degradation of SM, contributing to its accumulation. Elevated levels of SM can stiffen cellular membranes, interfering with vesicular trafficking and receptor signaling. Excess SM may compete with other phospholipids, disrupting lipid homeostasis and membrane integrity. And finally, the aberrantly high level of total brain SM is a key pathological event leading to neurodegeneration [29].
We found that age-related deterioration of SL turnover could be exacerbated by pinealectomy-induced melatonin deficiency in young adult rats with overproduction of Cer in the hippocampus. However, this effect was not associated with a proportional change in NSMase (increased level) or ASMase (decreased activity and/or level). This discrepancy could be explained by the fact that NSMase level but not its activity was measured in the hippocampus, which is a limitation factor of this study. In addition, endogenous melatonin is known to be a potent scavenger of OS-related molecules[30], and in this regard we previously reported that pinealectomy increased lipid peroxidation in the hippocampus of 3-month-old rats and decreased superoxide dismutase (SOD) activity during the dark phase compared to the corresponding sham rats[31]. The increased OS resulting from pinealectomy at this age period may be a result of the increased Cer production found in the present study.
Curiously, we found that the level of Cer in the hippocampus of middle-aged rats with pinealectomy was decreased compared to the corresponding sham group. In addition, while the level of NSMase was also reduced, the level of ASMase was restored in the hippocampus of 14-month-old pinealectomized rats compared to sham rats of the same age. This result was unexpected considering our previous report showing that this age period is the most susceptible to OS in the hippocampus as a consequence of pinealectomy [32]. As in young adult rats, pinealectomy in middle-aged rats had a devastating effect on liver markers (ALAT and ASAT), heat shock protein 70 in the frontal cortex [22], spatial working memory, and expression of BDNF and its receptor TrkB [27]. Moreover, we recently reported that only at this stage, SM levels in the hippocampus were decreased as a result of pinealectomy compared to the matched control [32]. Importantly, we also found that melatonin deficiency in 14-month-old rats is associated with disturbed antioxidant/oxidant balance in rat hippocampus with decreased GSH and increased lipid peroxidation. Notably, young adult and old rats with pinealectomy were not affected. It appears that endogenous melatonin in mature rats plays a critical role in mitigating free radical production by promoting GSH production. Furthermore, OS-induced changes in the hippocampus resulted in reduced SM levels in the same structure. A previous study demonstrated that the mechanism underlying the antidepressant activity of melatonin is related to suppression of Cer accumulation, possibly via inhibition of SM metabolism [7].
4. Materials and Methods
The experimental protocol was conducted in agreement with the European Communities Council Directive 2010/63/E.U. and approved by the Bulgarian Food Safety Agency (research project: #300/N◦5888–0183).
4.1. Animals
Male Wistar rats of three different ages were purchased from the vivarium of the Institute of Neurobiology, Bulgarian Academy of Sciences. Animals were housed for at least one week prior to surgery in standard Plexiglas cages (n = 3-4) under a 12 h/12 h light/dark regime, light on at 07:00 AM, room temperature at 22-23 °C, with ad libitum access to food and water.
4.2. Experimental Groups and Surgery
Six experimental groups were assigned according to age and surgical procedure as follows: 3-month-old sham group; 3-month-old pin group; 14-month-old sham, 14-month-old pin, 18-month-old sham, and 18-month-old pin groups, respectively. The pinealectomy or sham procedure (the same procedure except for the last step of the pinealectomy) was performed according to the protocol described in our previous studies [32]. Briefly, anesthetized rats (mixture of ketamine, 40 mg/kg, i.p., and xylazine, 20 mg/kg, i.m. for the 3-month-old rats; ketamine, 60 mg/kg, and xylazine, 30 mg/kg, i.m. for the 14-month-old rats) were fixed on a stereotaxic apparatus (Stoelting, Wood Dale, IL, US). After craniotomy with a small dental drill, the pineal gland located under the venous sinus was quickly removed with fine forceps. The rats were treated with antibiotic injection for three days after surgery. At least 2 weeks after the surgical manipulation, the animals were decapitated with a guillotine after light anesthesia with CO2, the two hippocampi were briefly removed, snap frozen in liquid nitrogen and stored at -20 °C until further analysis (6-8 samples/group), and their trunk blood was collected.
4.3. ELISA
4.3.1. Ceramide Levels in Hippocampus
Hippocampal tissue was weighted and homogenized (BANDELIN electronic GmbH&Co. KG, Berlin, Germany) in ice-cold PBS to a final concentration of 60 mg/mL. Then the homogenates were centrifuged for 15 min, 1500 x g, 4°C and the supernatant was collected and assayed immediately or aliquot and stored at -80°C. The samples were processed according to a Rat Ceramide ELISA kit (Cat. No. E02C2522, BlueGene Biotech, Shanghai, China) without further dilution. They were measured at 450 nm in a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland) and the samples’ concentrations were calculated based on a standard curve at ng/mL.
4.3.2. Ceramide Levels in Blood Plasma
The serum was clotted for 10–20 min (room temperature), while the plasma was collected in EDTA tubes and mixed for 20 min. Then the samples were centrifuged for 20 min at 2000–3000 rpm, and the supernatants were collected and stored at −20 °C until assayed. Blood ceramide levels (mmol/L) were determined via a commercial ELISA kit (Cat. No. MBS3809105, MyBioSource, Inc., San Diego, CA, USA). The samples’ OD was measured at 450 nm using a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland).
4.3.3. Sphingomyelin Levels in Hippocampus
Rat hippocampal samples were homogenized to a final concentration of 100 mg/mL in Cell/Tissue extraction buffer (100 mM Tris, pH 7.4; 150 mM NaCl, 1 mM EGTA, 1 mM EDTA, 1% Triton X-100, 0.5% Na deoxycholate, Protease inhibitor cocktail). After centrifugation at 4°C and 10 000 x g for 20 min, the supernatants were transferred to separate tubes and were heated for 5 min at 70°C until they became cloudy. To remove any debris, the samples were centrifuged again at 10 000 x g, 2 min. Then, 100 μL of each supernatant was transferred in duplicates to a 96-well plate. The SM concentration was estimated using a SM kit according to the manufacturer’s instructions (Cat No. MBS265875, MyBioSource, Inc., San Diego, CA, USA). SM concentration was measured by detecting the absorbance at 450 nm using a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland), calculated based on a standard curve and expressed as ng/mL.
4.3.4. Neutral Sphingomyelinase Levels in Hippocampus
Rat hippocampal samples were weighted and homogenized (1:10) with SONOPULS ultrasonic homogenizer (BANDELIN electronic GmbH&Co. KG, Berlin, Germany) in ice-cold PBS. The homogenates were centrifuged at 3000 x g for 10 min at 4°C to obtain the supernatants. The samples were then assayed via Rat Neutral Sphingomyelinase (NSMase) ELISA kit (Cat.No. MBS452865, MyBioSource, Inc., San Diego, CA, USA) according to the protocol. The samples were measured at 450 nm on a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland) and expressed in ng/mL.
4.3.5. Neutral Sphingomyelinase Enzymatic Activity in Blood Plasma
The plasma was collected in EDTA tubes and mixed for 20 min. Then the samples were centrifuged for 20 min at 2000–3000 rpm, and the supernatants were collected and stored at −20 °C until assayed. To measure the NSMase enzymatic activity the samples were processed via an ELISA kit (Cat. No. ab138876, Abcam Limited, Cambridge, UK). The kit uses an indicator to indirectly measure the amount of phosphocholine produced during the hydrolysis of sphingomyelin by NSMase. The samples’ OD was monitored at 655 nm on a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland) and expressed in mU/mL.
4.3.6. Acid Sphingomyelinase Enzymatic Activity in Hippocampus
ASMase enzymatic activity was determined via a commercial kit (ab252889, Abcam Limited, Cambridge, UK). Briefly, 10 mg of rat hippocampal tissue was homogenized in 100 µL ASMase Assay Buffer I supplemented with protease inhibitor cocktail (Cat. No. 11697498001, Roche, Basel, Switzerland) on ice. Then the samples were centrifuged at 10 000xg for 5 min and the supernatant was collected and further processed. The OD of the samples was measured at 570 nm on a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland). The final ASMase activity was calculated based on a standard curve in mU/mL.
4.3.7. Acid Sphingomyelinase Levels in Hippocampus
Rat hippocampal samples were weighted and homogenized (1:10) with SONOPULS ultrasonic homogenizer (BANDELIN electronic GmbH&Co. KG, Berlin, Germany) in ice-cold PBS. The homogenates were centrifuged at 3000 x g for 10 min at 4°C to obtain the supernatants. The samples were then assayed via Rat Acid Sphingomyelinase (ASMase) ELISA kit MyBiosource, Cat. No. MBS017899, Inc., San Diego, CA, USA) according to the protocol. The samples were measured at 450 nm on a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland) and expressed in ng/mL.
4.3.8. Acid Ceramidase Levels in Hippocampus
000
4.3.9. Acid Ceramidase Levels in Blood Plasma
The samples were centrifuged for 20 min at 2000–3000 rpm, and the supernatants were collected and stored at −20 °C until assayed. To determine plasma acid ceramidase (ASAH1) levels a commercial Rat Acid Ceramidase ELISA kit (Cat. No. MBS283260, MyBioSource, Inc., San Diego, CA, USA) was used. The samples’ OD was read at 450 nm using a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland), from which the concentration of ASAH1 was calculated in ng/mL.
4.3.10. Sphingosine-1-Phosphate Levels in Hippocampus
Each of the hippocampal samples were weighted and homogenized (1:10) with SONOPULS ultrasonic homogenizer (BANDELIN electronic GmbH&Co. KG, Berlin, Germany) in ice-cold 0.9% saline (6-8 mice/group). The homogenates were centrifuged at 3000 x g for 10 min at 4°C to obtain the supernatants. Sphingosine-1-Phosphate (S1P) levels were measured by a competitive ELISA assay according to manufacturer’s instructions (Cat. No. abx251939, Abbexa LTD, Cambridge, UK). The OD was measured spectrophotometrically at 450 nm using a microplate reader (Tecan Infinite F200 PRO (Tecan Trading GmbH, Männedorf, Switzerland), from which the concentration of S1P was calculated in ng/mL.
4.4. Statistical Analysis
Two-way analysis of variance (ANOVA) with the factors age (3 levels) and surgery (2 levels) was performed. A post hoc Bonferroni test was applied in case of significance of some of the factors. Data are presented as mean ±S.E.M. Statistical analysis was performed using SigmaStat® 11.0 (Systat Software, San Jose, CA, USA) and GraphPad Prism®6 (GraphPad Software, San Diego, CA, USA). Significant difference was accepted at p ≤ 0.05.
5. Conclusions
Taken together, the results of the present study suggest that melatonin deficiency associated with hormonal dysfunction affects SL signaling in the hippocampus and plasma in an age-specific manner. While the young adult rats with pinealectomy had impaired SL homeostasis, the changes in SL metabolism in middle age seem to be more complex and more affected by melatonin deficiency and deserve further investigation in the future. Considering our previous studies showing that pinealectomy had a deleterious effect on metabolism, hippocampal OS, anxiety responses, cognitive functions, and behavioral signaling in the hippocampus of young adult and middle-aged rats, the results of this study suggest that pinealectomy may modulate aging by altering sphingolipid turnover in an age-specific manner.
Author Contributions
Conceptualization, J.T. and R.T.; methodology, I.G. and R.T.; software, J.T. and I.G.; validation, R.T., J.T. and I.G.; formal analysis, J.T., I.G.; T.V. and S.A.; investigation, R.T., J.T., I.G.; T.V. and S.A.; resources, J.T. and R.T.; data curation, J.T., R.T. and I.G.; writing—J.T.; writing—review and editing, J.T. and R.T.; project administration, J.T.; funding acquisition, J.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Scientific Studies Fund of Bulgaria, grant number KP-06-N41/4 from 30.XI. 2020.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki. The animal study protocol was approved by Bulgarian Food Safety Agency permit No. 300/N◦5888–0183/10.05.2021, in accordance with EC Directive 2010/63/EU.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in thisarticle. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Wurtman, R.J. Age-Related Decreases in Melatonin Secretion—Clinical Consequences. The Journal of Clinical Endocrinology & Metabolism 2000, 85, 2135–2136. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Amaral, F.G.D. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocrine Reviews 2018, 39, 990–1028. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Xu, B.; Zhou, X.; Reiter, R. Pineal Calcification, Melatonin Production, Aging, Associated Health Consequences and Rejuvenation of the Pineal Gland. Molecules 2018, 23, 301. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, R.; Monse, M.; Pohl, E.; Wranik, S.; Wilker, B.; Keitsch, S.; Soddemann, M.; Kornhuber, J.; Kohnen, M.; Edwards, M.J.; et al. Melatonin Acts as an Antidepressant by Inhibition of the Acid Sphingomyelinase/Ceramide System. Neurosignals 2016, 48–58. [Google Scholar] [CrossRef]
- Trayssac, M.; Hannun, Y.A.; Obeid, L.M. Role of Sphingolipids in Senescence: Implication in Aging and Age-Related Diseases. Journal of Clinical Investigation 2018, 128, 2702–2712. [Google Scholar] [CrossRef]
- Li, S.; Kim, H.-E. Implications of Sphingolipids on Aging and Age-Related Diseases. Front. Aging 2022, 2, 797320. [Google Scholar] [CrossRef]
- Hoehn, R.; Monse, M.; Pohl, E.; Wranik, S.; Wilker, B.; Keitsch, S.; Soddemann, M.; Kornhuber, J.; Kohnen, M.; Edwards, M.J.; et al. Melatonin Acts as an Antidepressant by Inhibition of the Acid Sphingomyelinase/Ceramide System. Neurosignals 2016, 48–58. [Google Scholar] [CrossRef]
- Ordoñez, R.; Fernández, A.; Prieto-Domínguez, N.; Martínez, L.; García-Ruiz, C.; Fernández-Checa, J.C.; Mauriz, J.L.; González-Gallego, J. Ceramide Metabolism Regulates Autophagy and Apoptotic Cell Death Induced by Melatonin in Liver Cancer Cells. Journal of Pineal Research 2015, 59, 178–189. [Google Scholar] [CrossRef]
- Mencarelli, C.; Martinez–Martinez, P. Ceramide Function in the Brain: When a Slight Tilt Is Enough. Cell. Mol. Life Sci. 2013, 70, 181–203. [Google Scholar] [CrossRef]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An Overview of Sphingolipid Metabolism: From Synthesis to Breakdown. In Sphingolipids as Signaling and Regulatory Molecules; Chalfant, C., Poeta, M.D., Eds.; Advances in Experimental Medicine and Biology; Springer New York: New York, NY, 2010; Volume 688, pp. 1–23. ISBN 978-1-4419-6740-4. [Google Scholar]
- Cutler, R.G.; Kelly, J.; Storie, K.; Pedersen, W.A.; Tammara, A.; Hatanpaa, K.; Troncoso, J.C.; Mattson, M.P. Involvement of Oxidative Stress-Induced Abnormalities in Ceramide and Cholesterol Metabolism in Brain Aging and Alzheimer’s Disease. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 2070–2075. [Google Scholar] [CrossRef]
- Kitatani, K.; Idkowiak-Baldys, J.; Hannun, Y.A. The Sphingolipid Salvage Pathway in Ceramide Metabolism and Signaling. Cellular Signalling 2008, 20, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Brodowicz, J.; Przegaliński, E.; Müller, C.P.; Filip, M. Ceramide and Its Related Neurochemical Networks as Targets for Some Brain Disorder Therapies. Neurotox Res 2018, 33, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Newton, J.; Lima, S.; Maceyka, M.; Spiegel, S. Revisiting the Sphingolipid Rheostat: Evolving Concepts in Cancer Therapy. Experimental Cell Research 2015, 333, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.; Förl, M.; Winoto-Morbach, S.; Seite, S.; Schunck, M.; Proksch, E.; Schütze, S. Acid and Neutral Sphingomyelinase, Ceramide Synthase, and Acid Ceramidase Activities in Cutaneous Aging. Experimental Dermatology 2005, 14, 609–618. [Google Scholar] [CrossRef]
- Venable, M.E.; Lee, J.Y.; Smyth, M.J.; Bielawska, A.; Obeid, L.M. Role of Ceramide in Cellular Senescence. Journal of Biological Chemistry 1995, 270, 30701–30708. [Google Scholar] [CrossRef]
- Lightle, S.A.; Oakley, J.I.; Nikolova-Karakashian, M.N. Activation of Sphingolipid Turnover and Chronic Generation of Ceramide and Sphingosine in Liver during Aging. Mechanisms of Ageing and Development 2000, 120, 111–125. [Google Scholar] [CrossRef]
- Munk, R.; Anerillas, C.; Rossi, M.; Tsitsipatis, D.; Martindale, J.L.; Herman, A.B.; Yang, J.-H.; Roberts, J.A.; Varma, V.R.; Pandey, P.R.; et al. Acid Ceramidase Promotes Senescent Cell Survival. Aging 2021, 13, 15750–15769. [Google Scholar] [CrossRef]
- Monette, J.S.; Gómez, L.A.; Moreau, R.F.; Dunn, K.C.; Butler, J.A.; Finlay, L.A.; Michels, A.J.; Shay, K.P.; Smith, E.J.; Hagen, T.M. (R)-α-Lipoic Acid Treatment Restores Ceramide Balance in Aging Rat Cardiac Mitochondria. Pharmacological Research 2011, 63, 23–29. [Google Scholar] [CrossRef]
- Pierpaoli, W.; Bulian, D. The Pineal Aging and Death Program: Life Prolongation in Pre-aging Pinealectomized Mice. Annals of the New York Academy of Sciences 2005, 1057, 133–144. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Hrischev, P.; Ivanova, P.; Boyadjiev, N.; Georgieva, K. Metabolic Footprint in Young, Middle-Aged and Elderly Rats with Melatonin Deficit. Physiology & Behavior 2022, 250, 113786. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Krushovlieva, D.; Ivanova, P.; Nenchovska, Z.; Toteva, G.; Atanasova, M. The Role of Melatonin Deficiency Induced by Pinealectomy on Motor Activity and Anxiety Responses in Young Adult, Middle-Aged and Old Rats. Behav Brain Funct 2024, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Babenko, N.A.; Shakhova, E.G. Long-Term Food Restriction Prevents Aging-Associated Sphingolipid Turnover Dysregulation in the Brain. Archives of Gerontology and Geriatrics 2014, 58, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Nikolova-Karakashian, M.; Karakashian, A.; Rutkute, K. Role of Neutral Sphingomyelinases in Aging and Inflammation. In Lipids in Health and Disease; Quinn, P.J., Wang, X., Eds.; Subcellular Biochemistry; Springer Netherlands: Dordrecht, 2008; Volume 49, pp. 469–486. ISBN 978-1-4020-8830-8. [Google Scholar]
- Brunkhorst, R.; Pfeilschifter, W.; Rajkovic, N.; Pfeffer, M.; Fischer, C.; Korf, H.-W.; Christoffersen, C.; Trautmann, S.; Thomas, D.; Pfeilschifter, J.; et al. Diurnal Regulation of Sphingolipids in Blood. Biochimica et Biophysica Acta (BBA) - Molecular and Cell Biology of Lipids 2019, 1864, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Rutkute, K.; Asmis, R.H.; Nikolova-Karakashian, M.N. Regulation of Neutral Sphingomyelinase-2 by GSH: A New Insight to the Role of Oxidative Stress in Aging-Associated Inflammation. Journal of Lipid Research 2007, 48, 2443–2452. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Atanasova, D.; Krushovlieva, D.; Barbutska, D.; Atanasova, M.; Rashev, P.; Nenchovska, Z.; Mourdjeva, M.; Koeva, Y. Age-Related Memory Decline Is Accelerated by Pinealectomy in Young Adult and Middle-Aged Rats via BDNF / ERK / CREB Signalling in the Hippocampus 2024.
- Pujol-Lereis, L.M. Alteration of Sphingolipids in Biofluids: Implications for Neurodegenerative Diseases. IJMS 2019, 20, 3564. [Google Scholar] [CrossRef]
- Gaudioso, Á.; Jiang, X.; Casas, J.; Schuchman, E.H.; Ledesma, M.D. Sphingomyelin 16:0 Is a Therapeutic Target for Neuronal Death in Acid Sphingomyelinase Deficiency. Cell Death Dis 2023, 14, 248. [Google Scholar] [CrossRef]
- Reiter, R.; Carneiro, R.; Oh, C.-S. Melatonin in Relation to Cellular Antioxidative Defense Mechanisms. Horm Metab Res 1997, 29, 363–372. [Google Scholar] [CrossRef]
- Jana, T.; Tzveta, S.; Zlatina, N.; Natasha, I.; Dimitrinka, A.; Milena, A.; Katerina, G. Effect of Endurance Training on Diurnal Rhythms of Superoxide Dismutase Activity, Glutathione and Lipid Peroxidation in Plasma of Pinealectomized Rats. Neuroscience Letters 2020, 716, 134637. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Nenchovska, Z.; Kortenska, L.; Uzunova, V.; Georgieva, I.; Tzoneva, R. Impact of Melatonin Deficit on Emotional Status and Oxidative Stress-Induced Changes in Sphingomyelin and Cholesterol Level in Young Adult, Mature, and Aged Rats. IJMS 2022, 23, 2809. [Google Scholar] [CrossRef]
Figure 1.
The effect of pinealectomy on sphigomielin (SM) content in the hippocampus in 3- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a major Age effect [F1,23 = 60.919, p < 0.001]; *p < 0.001, 18- vs 3-month-old sham rats. 3 m results are published in Int. J. Mol. Sci. 2022, 23, 2809.
Figure 1.
The effect of pinealectomy on sphigomielin (SM) content in the hippocampus in 3- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a major Age effect [F1,23 = 60.919, p < 0.001]; *p < 0.001, 18- vs 3-month-old sham rats. 3 m results are published in Int. J. Mol. Sci. 2022, 23, 2809.
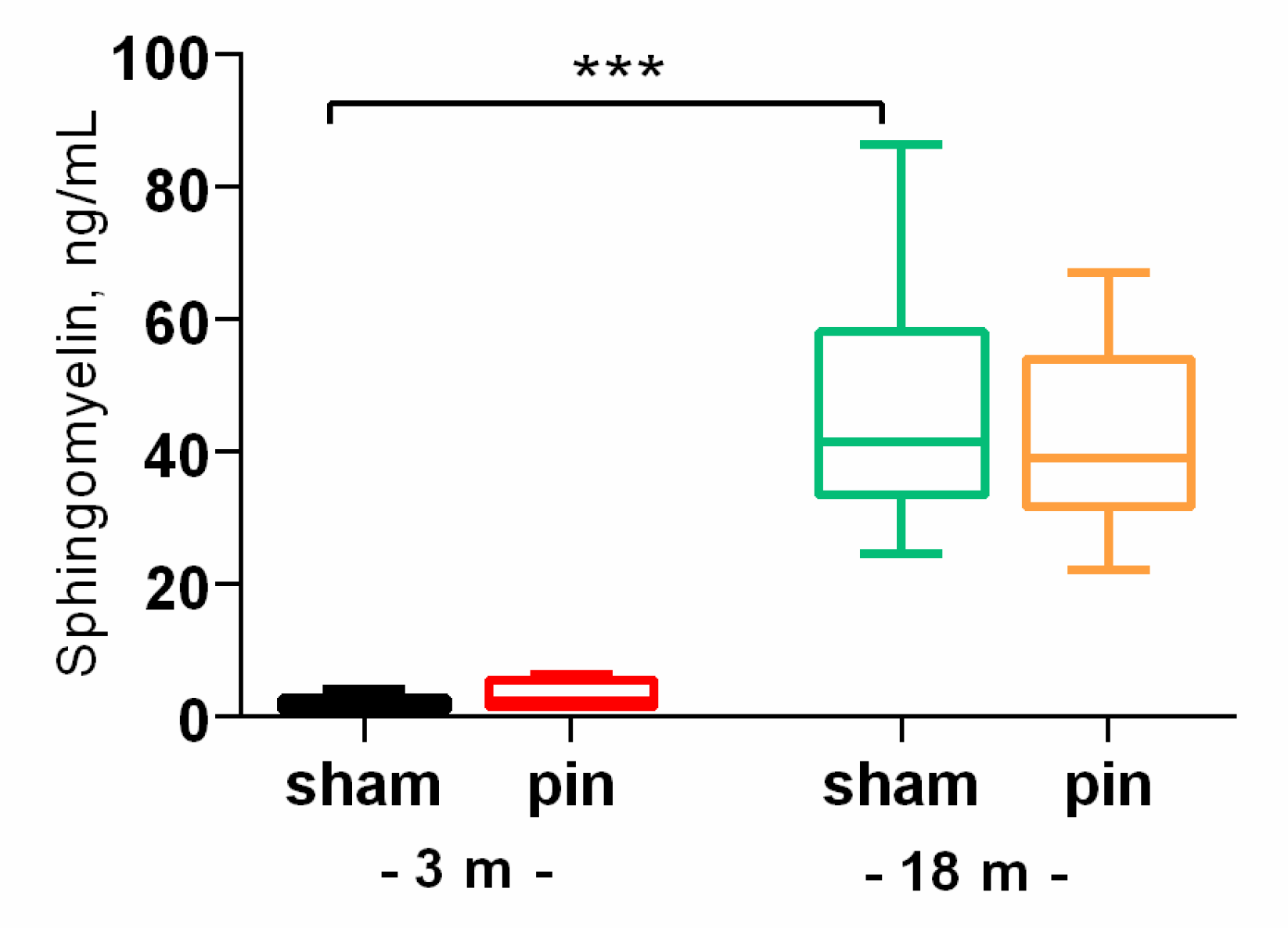
Figure 2.
The effect of pinealectomy on ceramide (Cer) content in the hippocampus and plasma in 3-, 14- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated Age x Surgery interaction [F2,45 = 6.915, p = 0.003] for the hippocampus; *p = 0.01, 18- vs 3-month-old sham rats; *p = 0.036, 3-month-old pin vs sham rats; **p = 0.006, 14-month old pin vs sham rats (A). For plasma a major Age effect [F2,52 = 3.217, p = 0.049] have been detected; *p = 0.042, 18- vs 3-month-old sham rats; ***p < 001, 18- vs 14-month-old sham rats.
Figure 2.
The effect of pinealectomy on ceramide (Cer) content in the hippocampus and plasma in 3-, 14- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated Age x Surgery interaction [F2,45 = 6.915, p = 0.003] for the hippocampus; *p = 0.01, 18- vs 3-month-old sham rats; *p = 0.036, 3-month-old pin vs sham rats; **p = 0.006, 14-month old pin vs sham rats (A). For plasma a major Age effect [F2,52 = 3.217, p = 0.049] have been detected; *p = 0.042, 18- vs 3-month-old sham rats; ***p < 001, 18- vs 14-month-old sham rats.
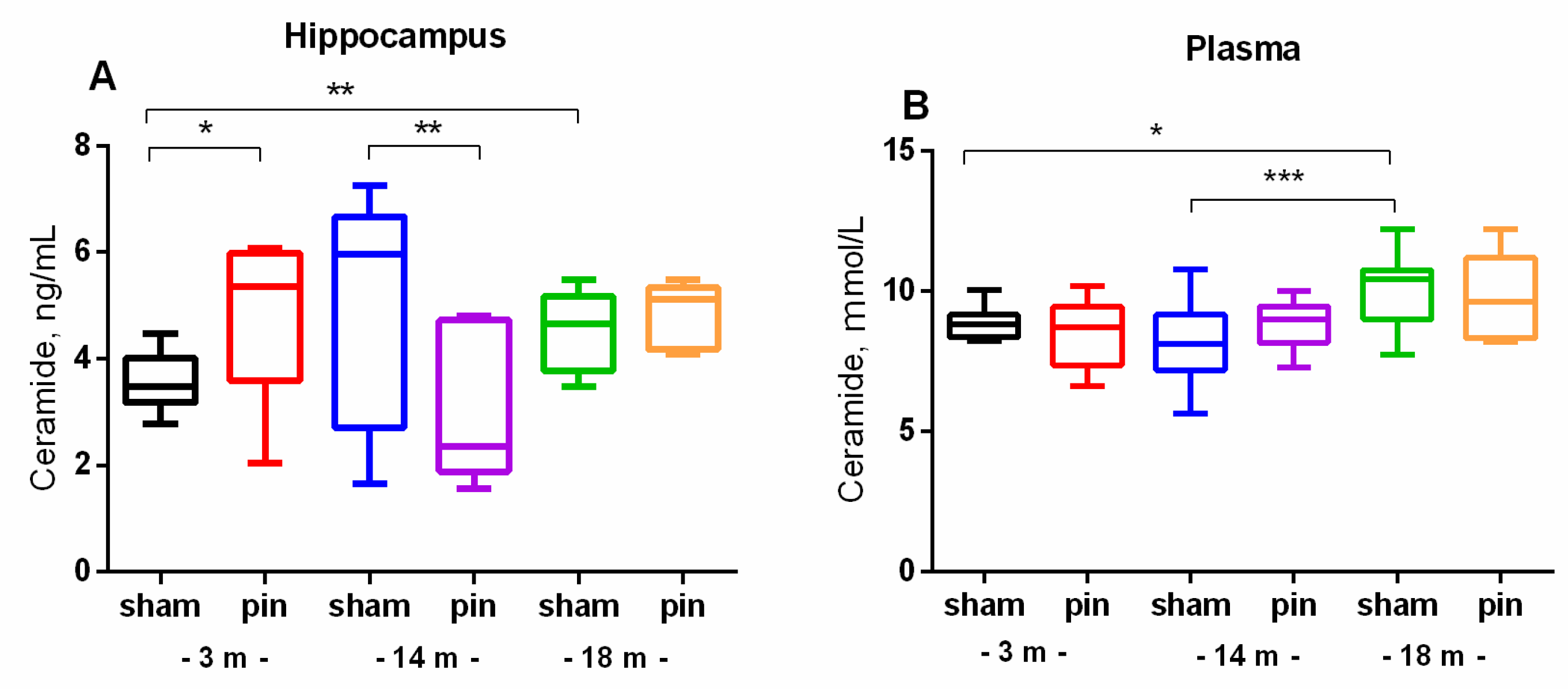
Figure 3.
The effect of pinealectomy on acid ceramidase (ASAH1) content in the hippocampus and plasma in 3-, 14- and 18-month-old rats. Data are presented as mean ± SEM2.3. Melatonin deficiency had beneficial effect on age-associated changes of NSMase and ASMase in the hippocampus only in the middle-aged rats.
Figure 3.
The effect of pinealectomy on acid ceramidase (ASAH1) content in the hippocampus and plasma in 3-, 14- and 18-month-old rats. Data are presented as mean ± SEM2.3. Melatonin deficiency had beneficial effect on age-associated changes of NSMase and ASMase in the hippocampus only in the middle-aged rats.
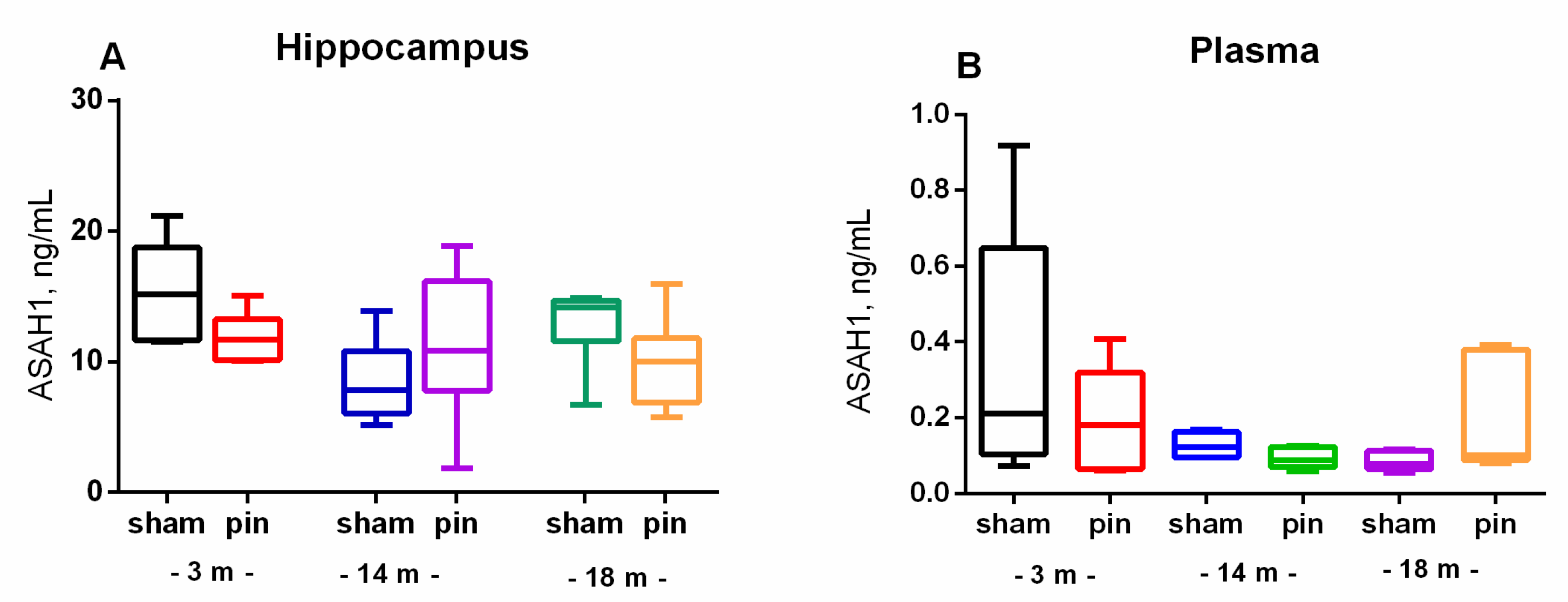
Figure 4.
The effect of pinealectomy on NSMase levels in the hippocampus and NSMase activity in plasma in 3-, 14- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a main Age effect: [F2,35 = 5.496, p = 0.012] as well as Age x Surgery interaction: [F2,35 = 6.267, p = 0.007] for the NSMase level in the hippocampus; **p = 0.01, 18-month-old sham vs 3-month-old sham; ***p < 0.001, 14-month-old pin vs sham rats.
Figure 4.
The effect of pinealectomy on NSMase levels in the hippocampus and NSMase activity in plasma in 3-, 14- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a main Age effect: [F2,35 = 5.496, p = 0.012] as well as Age x Surgery interaction: [F2,35 = 6.267, p = 0.007] for the NSMase level in the hippocampus; **p = 0.01, 18-month-old sham vs 3-month-old sham; ***p < 0.001, 14-month-old pin vs sham rats.
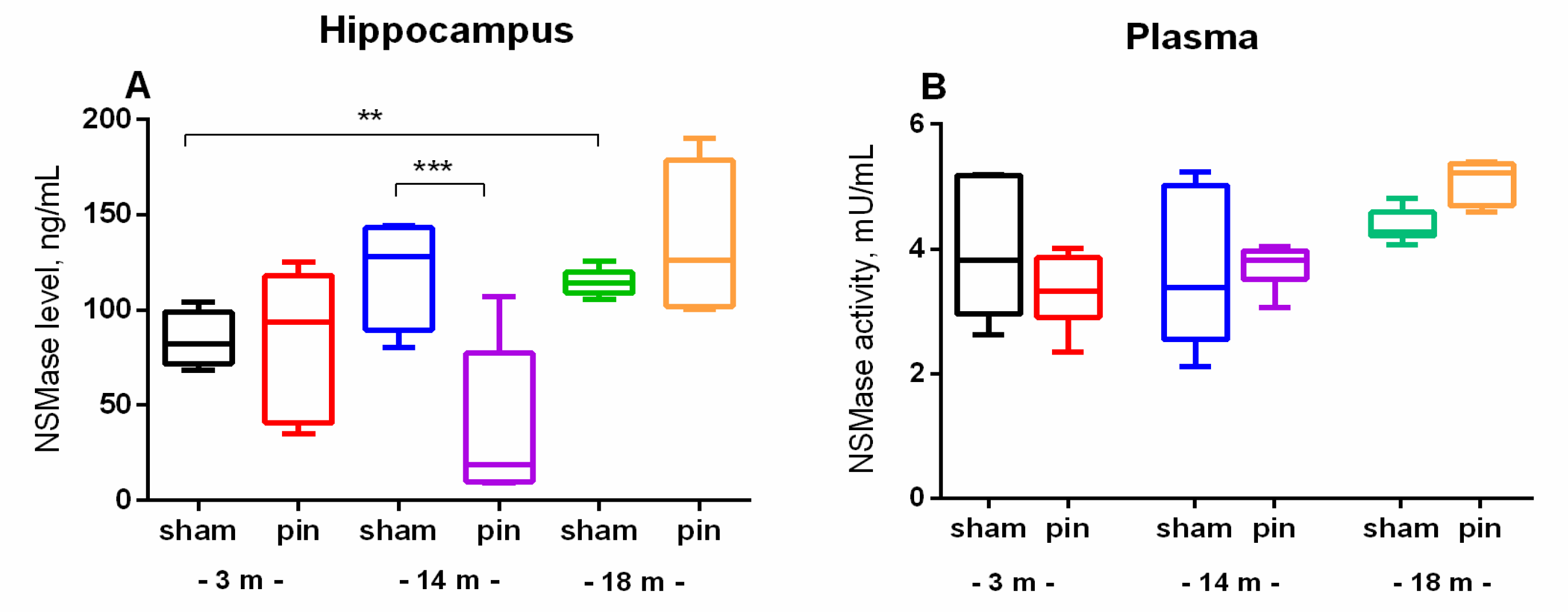
Figure 5.
The effect of pinealectomy on ASMase level (A) and activity (B) in the hippocampus in 3, 14- and 18-month-old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a main age effect [F2,35 = 6,584, p = 0,003], surgery effect [F1,46 = 7,223, p = 0,010] for ASMase level; **p = 0.002, 14-month-old sham rats vs 3-month-old sham rats; **p = 0.005, 18-month-old sham rats vs 3-month-old sham rats (A). Two-way ANOVA showed a major age effect for the ASMase activity: F2,45 = 647.244, p < 0.001], ***p < 0.001, 18-month-old sham rats vs. 3- and 14-month-old sham rats (B). 2.4. Melatonin Deficiency Exacerbates Age-Related Increase in S1P.
Figure 5.
The effect of pinealectomy on ASMase level (A) and activity (B) in the hippocampus in 3, 14- and 18-month-old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a main age effect [F2,35 = 6,584, p = 0,003], surgery effect [F1,46 = 7,223, p = 0,010] for ASMase level; **p = 0.002, 14-month-old sham rats vs 3-month-old sham rats; **p = 0.005, 18-month-old sham rats vs 3-month-old sham rats (A). Two-way ANOVA showed a major age effect for the ASMase activity: F2,45 = 647.244, p < 0.001], ***p < 0.001, 18-month-old sham rats vs. 3- and 14-month-old sham rats (B). 2.4. Melatonin Deficiency Exacerbates Age-Related Increase in S1P.
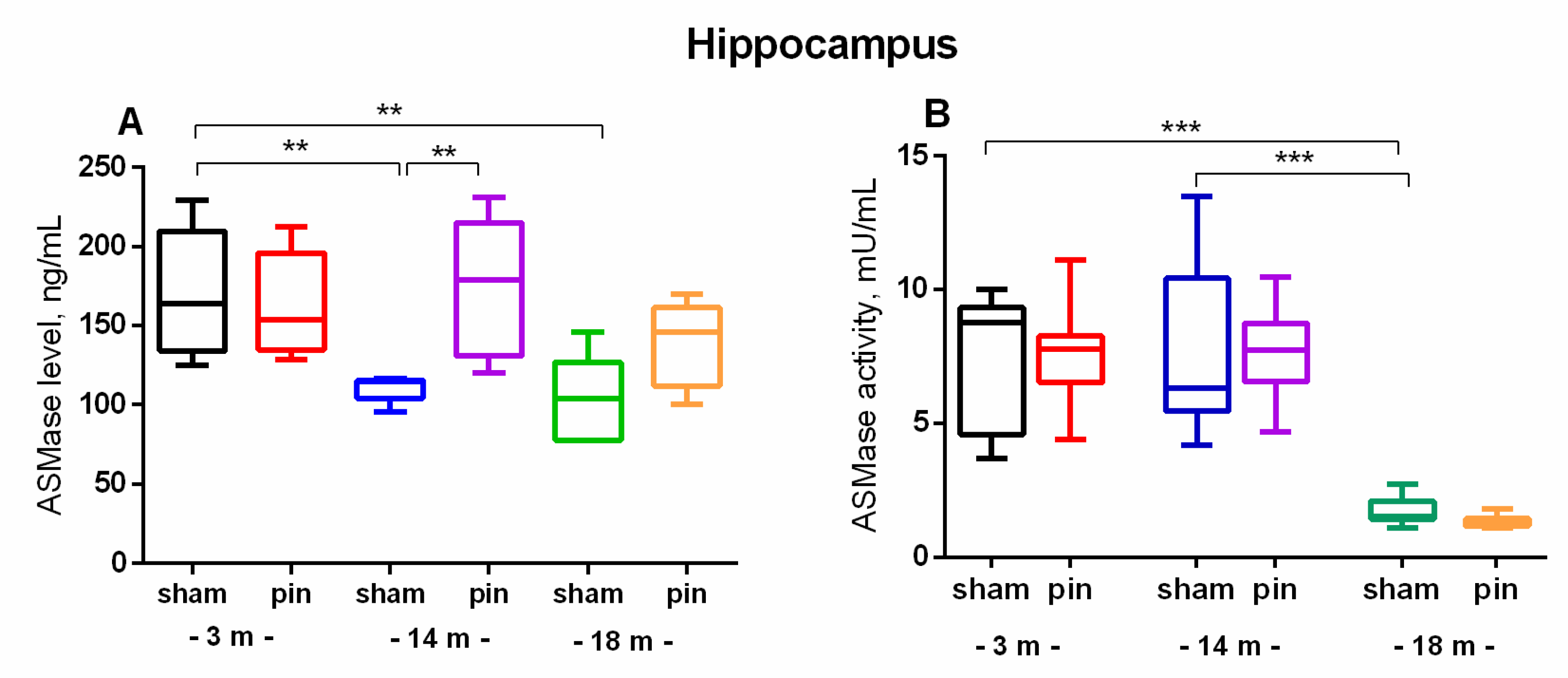
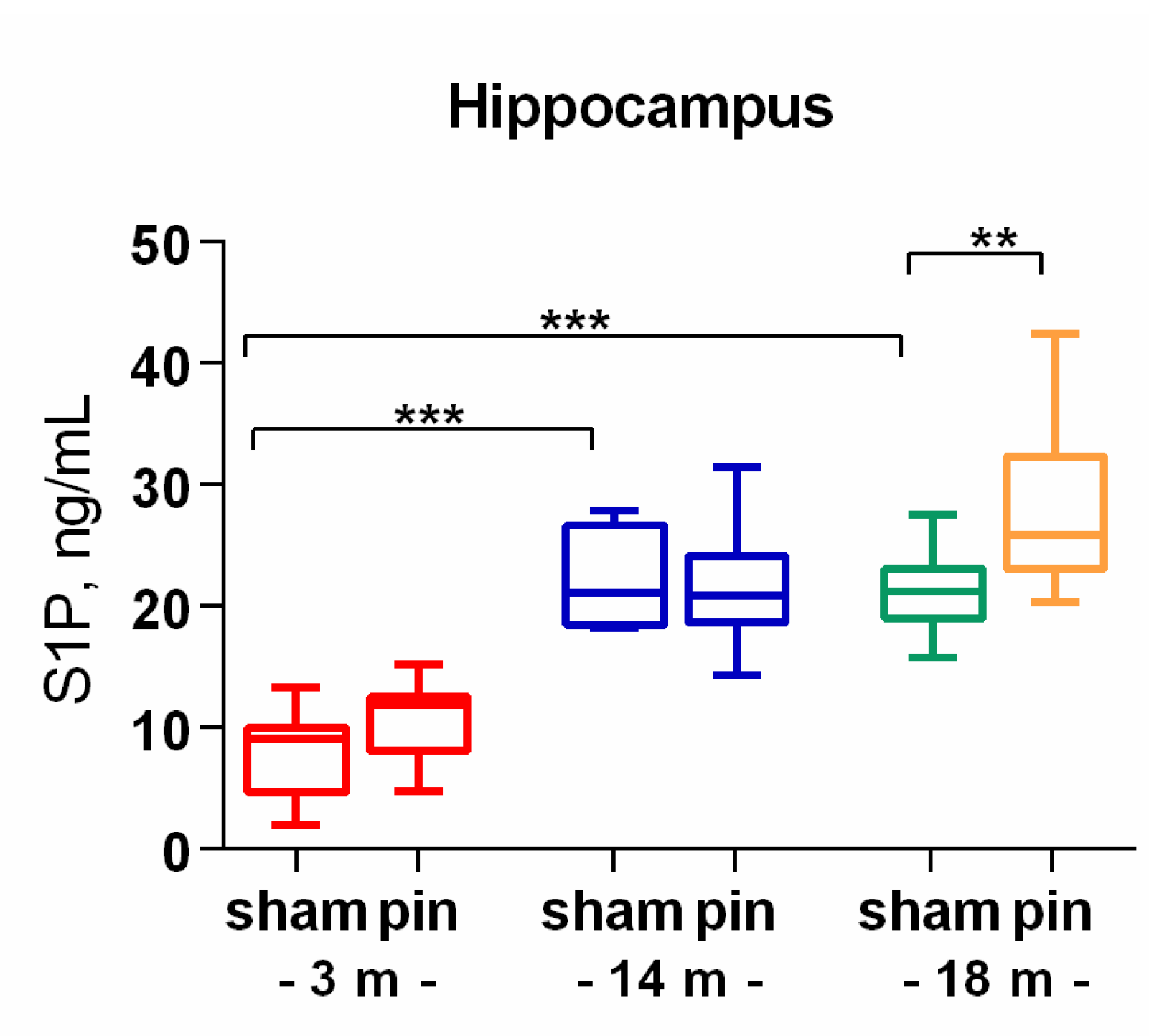
Figure 6.
The effect of pinealectomy on Sphingosine-1-phosphate (S1P) level in the hippocampus in 3-, 14- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a main Age effect [F2,35 = 47,73, p < 0,001], Surgery effect [F1,35 = 4,99, p = 0,05], ***p < 0.001, 14- and 18-month-old sham rats vs. 3-month-old sham rats; **p = 0.007, 18-month-old pin rats vs. 18-month sham rats.
Figure 6.
The effect of pinealectomy on Sphingosine-1-phosphate (S1P) level in the hippocampus in 3-, 14- and 18-month old rats. Data are presented as mean ± SEM. Two-way ANOVA indicated a main Age effect [F2,35 = 47,73, p < 0,001], Surgery effect [F1,35 = 4,99, p = 0,05], ***p < 0.001, 14- and 18-month-old sham rats vs. 3-month-old sham rats; **p = 0.007, 18-month-old pin rats vs. 18-month sham rats.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



