
Submitted:
20 December 2024
Posted:
23 December 2024
You are already at the latest version
Abstract
The impact of the world’s growing population on food systems and the role of dietary patterns in the managing of non-communicable diseases underscore the need to explore sustainable and dietary protein sources. Although microalgae have stood out as alternative sources of proteins and bioactive peptides, some species such as Nannochloropsis gaditana remain unexplored. This study aimed to characterize N. gaditana’s proteome and evaluate its potential as a source of bioactive by using an in silico approach. A total of 1955 proteins were identified and classified into functional groups of cellular component, molecular function, and biological process. In silico gastrointestinal digestion of identified proteins demonstrated that 202 hydrophobic and low molecular size-peptides with potential bioactivity were released. Of them, 27 exhibited theorical antioxidant, antihypertensive, antidiabetic, antiinflammatory, and/or antimicrobial activities. Seven of 27 peptide showed ≥20% intestinal absorption, suggesting potential systemic effects, while the rest could act at local level. Molecular docking demonstrated strong affinities with key enzymes such as MPO, ACE, and DPPIV. Resistance to the digestion, capacity to be absorbed and multifunctionality were demonstrated for peptide FIPGL. This study highlights N. gaditana's potential as a sustainable source of novel potential bioactive peptides with promising local and systemic biological effects.
Keywords:
microalgae
; proteomics
; functional analysis
; in silico digestion
; Nannochloropsis sp.
1. Introduction
In recent years, non-communicable diseases (NCD) have been considered a serious disease burden recognized worldwide. In fact, with more than 50% of all deaths being attributable to them, NCD such as stroke, cancer, ischemic heart disease, chronic kidney disease, and auto-immune and neurodegenerative conditions are now some of the biggest threats and challenges to human health [1]. The main risk factors of NCDs can be classified into the categories of self-management, genetics, medical conditions, and socio-demographic and environmental factors, including diet [2]. Therefore, the role of dietary patterns and evidence-based nutrition interventions in the prevention and management of NCDs should be a global health priority.
Predictive models of global population have raised concerns about the sustainability of food systems. Among the myriads of strategies, the gradual transition from meat to plant-based consumption supported and coordinated by the main health and environmental institutions is considered an effective approach [3]. Proteins derived from animal sources are highly valued for their nutritional and functional qualities; however, livestock production significantly impacts the environment, highlighting the importance of balancing all dietary protein sources to meet human needs [4]. Among the different alternative protein sources currently studied, microalgae are considered one of the most promising due to their optimal nutrition profile, minimal environmental footprint, and significant biological potential [5].
Compared to dry plant seeds, microalgae have been reported to have remarkably higher protein contents, ranging from 30% to 70% of dry weight (with an average value of nitrogen-to-protein conversion factor of 4.78) [6]. In addition, microalgae offer significantly greater protein yield per unit area compared to wheat or beef, owing to their simplified growth processes, reduced energy expenditure, and superior photosynthetic efficiency [7]. Furthermore, microalgae proteins are recognized for their excellent promising functional properties, low antigenicity [8] and high nutritional quality, providing all 20 amino acids (AA) and varying amounts of essential AA (EAA) [9]. The AA composition of many microalgae proteins such as Nannochloropsis gaditana is similar to that of typical plant protein sources, such as soybean [10], and closely aligns with the FAO/WHO/UN reference profile [11]. Thus, microalgal proteins can be regarded as an alternative protein source with nutritional and biological attributes after being broken down in the gastrointestinal tract, liberating EAA and potential bioactive peptides [12].
Bioactive peptides are small fragments, between 2 and 20 AA, which do not exert any effect within their origin protein but can exert multiple biological properties once released. Different bioactivities have been reported for bioactive peptides such as antioxidant, anti-inflammatory, antidiabetic, and immunomodulatory, among others [13]. In the case of microalgae proteins, several studies have mentioned their ability to produce bioactive peptides after enzymatic hydrolysis. Thus, proteins from Chlorella vulgaris, Arthrospira platensis, and A. maxima have been demonstrated to generate bioactive peptides with anti-inflammatory and antioxidant activities after their hydrolysis with pepsin, trypsin or the combination of alcalase and papain [14,15,16]. Similarly, N. oculata has been reported to generate protein hydrolyzates and peptides with angiotensin-converting enzyme (ACE)-1 inhibitory, antithrombotic, and calcium/calmodulin-dependent kinase II (CAMKII) inhibitory activities [17,18]. Saleh et al. reported that a protein hydrolyzate from N. gaditana with alcalase exerted high in vitro antioxidant activity [19].
Proteases, such as pepsin in the stomach and trypsin and chymotrypsin in the small intestine, play a crucial role in breaking down proteins into smaller fragments. In the case of microalgae, several studies have investigated the release of bioactive peptides from microalgae under simulated gastrointestinal conditions. For example, Li et al. [20] described ACE and dipeptidyl peptidase (DPP)-IV inhibitory peptides contained in C. pyrenoidosa digests. The study of Pei et al. [21] identified the antioxidant and anti-inflammatory peptide EMFGTSSET released after simulating gastrointestinal digestion of Isochrysis zhanjiangensis. The susceptibility to the action of digestive enzymes has also been demonstrated for other microalga species such as N. oculata [22]. However, to our knowledge, no studies have demonstrated the potential of N. gaditana as a source of bioactive peptides after its gastrointestinal digestion.
The study of bioactive peptides using in vitro, cell, or animal models is expensive, time-consuming, and might also have ethical considerations. Hence, in silico bioinformatic approaches, which cannot only help to predict the formation, breakdown, and absorption of bioactive peptides from any known protein source but can also help to analyze the structure/function relationship, have gained a new meaning in this scientific field as they can serve as a screening tool to accelerate the process of discovery of potentially encrypted bioactive peptides [23]. Therefore, the goal of this work was to carry out a complete characterization of the microalga N. gaditana proteome, and to demonstrate its potential as a source of bioactive peptides through an in silico gastrointestinal digestion strategy, supporting the value of this microalga as a sustainable alternative protein with both nutritional and health-promoting properties.
2. Materials and Methods
2.1. Samples and Reagents
Commercial N. gaditana microalgae biomass was supplied by AlgaEnergy S.A. (Madrid, Spain). Acetonitrile (ACN), trifluoroacetic acid (TFA), calcium chloride (CaCl2), Tris-HCl, formic acid, dithiothreitol (DTT), and iodoacetamide were purchased from Sigma-Aldrich (St. Louis, MO, USA).
2.2. Proteomic Identification and Characterization
2.2.1. Protein Content and AA Profile
The protein concentration of N. gaditana biomass was measured via the bicinchoninic acid (BCA) method, which was performed using the Pierce BCA kit (Thermo Scientific, Waltham, MA, USA) [24]. The standard used was bovine serum albumin (BSA) at concentrations ranging from 50 to 1000 µg/mL. The AA content was determined in duplicate by using a Biochrom 30 series Amino Acid Analyser (Biochrom, Cambridge, UK), and following the previously described methodology [25].
2.2.2. In-Gel Digestion (Stacking Gel)
N. gaditana biomass was suspended in 50 µL of sample buffer and then, applied onto 1.2-cm wide wells of a conventional SDS-PAGE gel (0.75 mm-thick, 4% stacking, and 10% resolving, Bio-Rad, Hercules, CA, USA) following the methodology previously reported [26]. Then run was stopped when the front entered 3 mm into the resolving gel, so the whole proteome became concentrated in the stacking/resolving gel interface. The unseparated protein bands were visualized by Coomassie staining (Bio-Rad), excised, cut into cubes (2 x 2 mm), and placed in 0.5 mL microcentrifuge tubes [27]. The gel pieces were distained in ACN:H2O (1:1, v:v), reduced and alkylated (disulfide bonds from cysteinyl residues were reduced with 10 mM DTT for 1 h at 56˚C, and then thiol groups were alkylated with 10 mM iodoacetamide for 30 min at room temperature in darkness) and digested in situ with sequencing grade trypsin (Promega, Madison, WI, USA) as described by Shevchenko et al. [28]. The gel pieces were shrunk by removing all liquid using sufficient ACN. ACN was pipetted out, and the gel pieces were dried in a speedvac and re-swollen in 100 mM Tris-HCl pH 8, 10 mM CaCl2 with 60 ng/mL trypsin at 5:1 protein:enzyme (w/w) ratio. The tubes were kept in ice for 2 h and incubated at 37°C for 12 h. Digestion was stopped by the addition of 1% TFA. Whole supernatants were dried down and then desalted using OMIX Pipette tips C18 (Agilent Technologies, Santa Clara, CA, USA) before the analysis. Digestion was done in the presence of 0.2% RapiGest (Waters, Milford, MA, USA).
2.2.3. Reverse Phase-Liquid Chromatography RP-LC-MS/MS Analysis (Dynamic Exclusion Mode)
The desalted protein digest was dried, resuspended in 10 μL of 0.1% formic acid, and analyzed by RP-LC-MS/MS in an Easy-nLC 1200 system coupled to an ion trap LTQ-Orbitrap-Velos-Pro hybrid mass spectrometer (Thermo Scientific, Waltham, MA, USA). The peptides were concentrated (on-line) by reverse phase chromatography using a 0.1mm × 20 mm C18 RP precolumn (Thermo Scientific) and then separated using a 0.075mm x 250 mm bioZen 2.6 µm Peptide XB-C18 RP column (Phenomenex, Torrance, CA, USA) operating at 0.25 μL/min. Peptides were eluted using a 180-min dual gradient. The gradient profile was set as follows: 5−25% solvent B for 135 min, 25−40% solvent B for 45 min, 40−100% solvent B for 2 min, and 100% solvent B for 18 min (Solvent A: 0.1% formic acid in water, solvent B: 0.1% formic acid, 80% ACN in water). Electrospray ionization (ESI) was done using a Nano-bore emitters Stainless Steel ID 30 μm (Proxeon, Odense, Denmark) interface at 2.1 kV spray voltage with S-Lens of 60%. The Orbitrap resolution was set at 30,000 [29]. Peptides were detected in survey scans from 400 to 1600 amu (1 μscan), followed by twenty data-dependent MS/MS scans (Top 20), using an isolation width of 2 u (in mass-to-charge ratio units), normalized collision energy of 35%, and dynamic exclusion applied for 60 seconds periods. Charge-state screening was enabled to reject unassigned and singly charged protonated ions.
2.2.4. Data Processing
Peptide identification from raw data was carried out using PEAKS Studio v11.5 search engine (Bioinformatics Solutions Inc., Waterloo, Ontario, Canada). Database search was performed against UniProt-Nannochloropsis gaditana.fasta (15363 entries); UniProt release 11/2023. The following constraints were used for the searches: tryptic cleavage (semi specific) after arginine (R) and Lysine (K), up to two missed cleavage sites, and tolerances of 20 ppm for precursor ions and 0.6 Da for MS/MS fragment ions. The searches were performed allowing optional methionine (M) oxidation and cysteine (C) carbamidomethylation. False discovery rates (FDR) for peptide spectrum matches (PSM) and protein were limited to 0.01. Only those proteins with at least two unique peptides being discovered from LC/MS/MS analyses were considered reliably identified [27,30,31,32].
2.3. Proteomic Functional Analysis
The proteomic functional analysis of the identified proteins from N. gaditana biomass was carried out using the functional analysis module from OmicsBox 3.3.2 bioinformatic software (Biobam, Valencia, Spain) through the BLAST2GO methodology [33]. The identified proteins were classified into three different functional groups: cellular component, molecular function, and biological process. The database to perform the alignment was Non-redundant protein sequences (nr v5), filtered by taxonomy: N. gaditana (code 72520). The specific workflow followed for the functional analysis is shown in Supplementary Figure 1.
2.4. In silico Gastrointestinal Digestion
The in silico gastrointestinal digestion of proteins was performed usig the Rapid Peptides Generator (RPG) software v22.2.3 [34]. By using the Python (3.12) system, the complete data obtained from the proteomic identification was entered in fasta format and firstly digested with pepsin (code 34) to simulate gastric digestion. Afterward, only peptides released from proteins that generated ≥ 100 peptides/protein after the gastric phase were selected to be concurrently digested with trypsin (code 42) and chymotrypsin high specificity (code 15). In all in silico gastric and gastrointestinal digests, free AA and duplicate peptides were deleted and the resulting peptides were classified based on their size and AA sequence.
2.5. Bioactivity Prediction by In Silico Analysis
For the prediction of the bioactivity, an in silico analysis of the peptides released during the gastric and gastrointestinal phases was conducted using the server based on a novel N-to-1 neural network Peptide Ranker [35]. Peptides with ≥ 0.8 value of the probability of being bioactive were selected to estimate their: i) antioxidant activity by using the Antioxidative peptide predictor (AnOxPP) database [36], ii) antiinflammatory activity by PreAIP database [37], iii) antimicrobial activity by the CAMPR4 database [38], iv) antihypertensive activity by the AHTpin database [39] and v) DPP-IV inhibitory activity by using the StackDPPIV [40].
2.6. Physicochemical and Pharmacokinetic Analysis
The peptides with 4 or more predicted bioactivities were selected and SwissADME molecular sketcher, based on ChemAxon’s Marvin JS 24.3.0 (re389bc4da912), 2024-07-17, was used to obtain the canonical SMILE for each peptide and to predict the physicochemical and pharmacokinetic properties such as molecular formula, weight, lipophilicity, water solubility, bioavailability score, AMES toxicity and the percentage of human intestinal absorption through the SwissADME web tool [41,42].
2.7. Peptide Molecular Docking
The peptide molecular interactions with human angiotensin-converting enzyme (ACE), DPP-IV, and myeloperoxidase (MPO) enzymes were predicted using molecular docking. The crystal structure for ACE (2C6N), DPP-IV (1NU8), and MPO (3F9P) was obtained from a protein data bank (http://www.rcsb.org/) [43]. Peptides were designed using Marvin Sketch software (Chemaxon, version 19.22.0), flexible torsions, charges, and grid size were assigned using Autodock Tools, and docking calculations were performed using AutoDock Vina where the binding pose with the lowest binding energy was selected as representative to visualize in the Discovery Studio 2016 Client (Dassault Systemes Biovia Corp R) as it was previously described [44,45].
To clarify the experimental design, the workflow of the present research is summarized in Figure 1.
3. Results and Discussion
3.1. Proteome of the Microalga Nannochloropsis gaditana
The protein content of N. gaditana biomass was 21.69%. Values reported in the literature ranged from 19-24% for N. oculata [46] to 44.9% for N. gaditana [47]. Levasseur et al. [48] also compiled intermediate percentages for this genus. The protein content of Nannochloropsis sp. microalgae has been demonstrated to be affected by numerous biotic and abiotic factors and genomic variation among species [49]. These factors can also impact the AA profile and, consequently the protein quality [50]. The AA profile of the N. gaditana used in the present research is shown in Table 1. All AA were analyzed except tryptophan, as the methodology conditions degraded it. The measurement showed a high AA content, leucine being the major EAA followed by lysine and phenylalanine. Among non-essential AA (NEAA), glutamic acid was the most predominant followed by aspartic acid and proline. Comparing the results obtained with the recommendation of the FAO/WHO/United Nations University (UNU) report (2007), the requirements established for each AA were covered [51].
After the in-gel digestion and consequent RP-LC-MS/MS analysis of N. gaditana biomass, by using PEAKS and SPIDER search tools [52], a total of 13,097 peptide-spectrum-matches (PSMs), 12,574 scans (which account for MS/MS scans that have a peptide spectrum associated with them), 6,991 features identified with database search only, 6,903 peptide sequences with modifications not including isoleucine/leucine differentiation and 5,820 without modifications and isoleucine/leucine differentiation, were identified.
A total of 1,955 proteins were detected through PEAKS software. Since sequences constituted the key for homology-based protein identification, PEAKS works with protein groups that contain all the proteins that have been identified with the same peptides [53,54]. In our case, 1,439 protein groups were identified. Of 1,955 proteins, 1,258 proteins were identified with two or more unique peptides Their correspondent accession number, -10logP value, number of peptides and unique peptides, the type of modification present in the peptide (PTM), the average mass, and the description of all proteins detected in N. gaditana biomass were collected and available in the Supplementary Table 1.
N. gaditana proteome was first studied by Fernández-Acero et al. [55], identifying 1,950 proteins. The differences found in comparison with our results could be due to factors such as the strain and the growing conditions, which have been demonstrated to affect microalgae macronutrient distribution [56]. Hulatt et al. [57] identified 3,423 proteins in N. gaditana biomass, where the abundance of 1,543 and 113 of them was demonstrated to be influenced by the amount of nitrogen and phosphorous available in the culture conditions, respectively. In general, the limited count of proteins and peptides identified by the database search should be attributed to the incompleteness of N. gaditana proteome sequenced and the differences in the experimental procedures. Thus, mapping N. gaditana proteome may aid to understand variables affecting its proteins and to explore their properties as sources of bioactive peptides.
3.2. Functional Analysis of Nannochloropsis gaditana Proteome
To elucidate the functionality of the identified proteins in N. gaditana, an alignment of the protein sequences detected was carried out using the BLAST2GO methodology, classifying proteins with differentiated biological functions into three groups: biological process, cellular component, and molecular function. The functional distribution of the identified proteins at levels 1 and 2 of each functional group are shown in Figure 2 and the detailed complete level distribution of each function is available in Supplementary Figure 2, Figure 3 and Figure 4.
The distribution of proteins at level 1 was quite homogeneous, since the percentages associated with each function were similar (Figure 2A) with 1,309 sequences for biological process (31.56%), 1,424 sequences for molecular function (34.44%) and 1,415 sequences for cellular component (31.56%). However, within the sequences associated with cellular components at level 2 (Figure 2B), a noticeable difference was observed since the majority were found within cellular entities (74.13%) and not forming complexes (25.87%). Regarding cellular entities, intracellular anatomical structures and organelles were the most abundant groups (Supplementary Figure 1). Concerning the biological process group level 2 classification (Figure 2C), protein sequences related to cellular processes were the most abundant (66.24%) in contrast with proteins related to localization, response to stimulus, and regulation processes. Regarding those cellular processes, metabolic processes in which energy is obtained from the metabolization of organic substances, were the most abundant. Finally, concerning molecular function level 2 classification (Figure 2D), the major percentage of sequences were associated with binding processes (47.30%) and catalytic activities (44.32%) which are related to proteins associated with cyclic organic compounds, necessary for photosynthesis, mobilizing metabolites and building structural components, such as the cell wall, among other previously studied functions [58]. Our results were in tune with the GO classification conducted by Fernández-Acero et al. [55] and Hulatt et al. [50] as the major percentage of proteins that belonged to the biological process group where related to biosynthetic processes, carbon fixation, ATP synthesis, and AA regulation, and the major percentage of proteins who belonged to the molecular process group where related with ion binding processes.
3.3. In Silico Gastrointestinal Digestion
The in silico gastrointestinal digestion of the complete set of 1,955 proteins detected through PEAKS software was conducted. This allowed the inclusion of proteins that could be underrepresented or absent due to technical limitations, such as low abundance, suboptimal extraction methods, or inefficient ionization in mass spectrometry analysis [59]. In this way, we would be ensuring that even proteins with marginal or undetected peptides were considered, potentially uncovering biologically relevant or functional insights [60]. Moreover, the additional 697 proteins with fewer than two unique peptides may still hold critical roles in metabolic pathways or cellular processes but be overlooked in traditional proteomics workflows. Therefore, by considering the entire dataset, researchers can identify proteins with potential bioactivity, nutritional value, or industrial applications that might otherwise be disregarded.
Consequently, an in silico gastric digestion with pepsin was initially done generating a total of 164,503 sequences that, after removing free AA and duplicate peptides resulted in 47,982 sequences. Then, the number of peptides generated from each identified protein was counted and 24 proteins were selected as the most peptide-generating proteins (with ≥ 100 peptides/protein) after the gastric phase (Table 2).
The average mass of the selected proteins ranged from 118 – 552 kDa. The three most peptide-generating proteins where zinc finger ZZ-type (that generated 376 peptides), acetyl CoA carboxylase (207 peptides) and bromodomain containing 1 (169 peptides), which average masses were slightly higher than the rest. These three proteins have been previously described in other microalgae species such as Schizochytrium sp., C. vulgaris, Chlamydomonas reinhardtii, mainly exhibiting a regulatory role in microalgae lipid metabolism and biosynthesis [61,62,63,64]. In addition, Lin et al. [65] specifically highlighted the involvement of acetyl CoA carboxylase in the type II fatty acid synthesis pathway, which occurs in the plastids of the cells of N. gaditana. These proteins would be of great importance since the microalga N. gaditana has aroused great interest due to its high lipid content and its potential as a sustainable source of omega-3 fatty acids.
Proteins susceptibility to pepsin action can be influence by several structural and chemical factors, such as protein molecular weight, AA composition and acidic conditions among others. Pepsin is a protease that specifically cleaves peptide bonds at the amino-terminal side of hydrophobic and aromatic residues sites such as phenylalanine, tyrosine and leucine breaking polypeptide chains into smaller peptides [66]. Generally, larger proteins commonly provide more cleavage sites for pepsin due to their longer polypeptide chains [67]. Therefore, the susceptibly of the three proteins with most peptides generated could be explain by their high average mass (552, 235, 245 kDa respectively) and the amount of hydrophobic AA found in N.gaditana biomass, such as leucine or phenylalanine. As well, in the RPG software pepsin was selected at pH 1.3 (code 33) trying to assure its high cleavage affinity and most optimal conditions. Nevertheless, information regarding N.gaditana protein susceptibility to pepsin is still scarce and should be further explored.
The selected proteins generated a total of 3160 peptides during the gastric phase. Of them, 104 were predicted to be active by the Peptide Ranker software tool with a score ≥ 0.8. These peptides were analyzed by using the AnOxPP, PreAIP, CAMPR4, AHTpin and StackDPPIV databases to predict their antioxidant, antiinflammatory, antimicrobial, antihypertensive and antidiabetic activities, respectively. Three peptides, FHPKF, FIPGL and FRPLPPLA, were predicted to potentially exert the five bioactivities assayed while two sequences, PLPAGRF and HHDCF, were predicted to have four of those bioactivities. In addition, a second phase simulating the intestinal digestion was conducted on 3160 peptides generated during the gastric phase, using trypsin and chymotrypsin as pancreatic enzymes. A total of 6224 peptides were generated after eliminating free AA and duplicates. Of these peptides, 54 were also present in the in silico gastric digest, indicating their capacity to resist the action of trypsin and chymotrypsin. After being analyzed with the Peptide Ranker software tool, 202 of 6224 peptides showed a score ≥ 0.8. The peptide profile is shown in Figure 3.
Figure 3.
Peptide profile of the peptides released after in silico gastrointestinal digestion from Nannochloropsis gaditana biomass and predicted as bioactive using the Peptide Ranker software tool. A) size distribution expressed as number of amino acids; B) amino acid distribution.
Figure 3.
Peptide profile of the peptides released after in silico gastrointestinal digestion from Nannochloropsis gaditana biomass and predicted as bioactive using the Peptide Ranker software tool. A) size distribution expressed as number of amino acids; B) amino acid distribution.
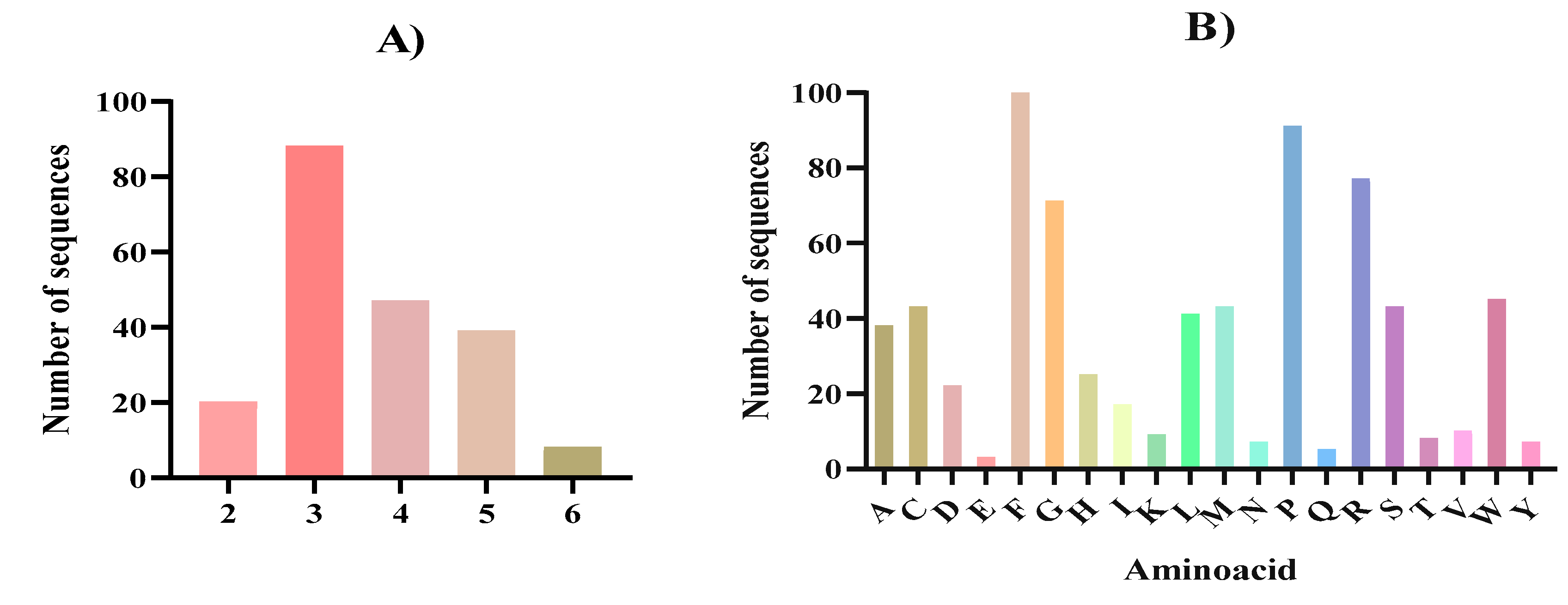
Most of the potentially bioactive peptides released after the complete gastrointestinal digestion were tripeptides (88), followed by tetrapeptides (47) and pentapeptides (39) (Figure 3A). The major AA present were phenylalanine (F) followed by proline (P), arginine (R) and glycine (G) (Figure 3B). The high presence of phenylalanine (F) and proline (P) observed in the in silico gastrointestinal digest agreed with the high content of these AA present in the N. gaditana biomass (Table 1). It has been previously described that numerous bioactive peptides present a structure-activity relation, being a key part of their potential biological effects [68,69]. In tune with our results, in general and specially in the case of the antioxidant and ACE inhibitory activities, it has been described that active fragments are either dipeptides or tripeptides containing hydrophobic AA, such as phenylalanine or proline [70,71]. In the case of microalgae derived bioactive peptides, several microalgae species like Chlorella sp., Navicula sp., and Arthorspira sp. have been described as sources of small bioactive peptides with hydrophobic AA [72,73] which suggests that N. gaditana could also have the potential to become a source of bioactive peptides. After the in silico digestion and bioactivity prediction analysis, from 202 peptides, two sequences (FIPGL and YLPPR) were predicted to exert five activities while 25 sequences showed four of five activities (Table 3).
Regarding the potentially bioactive dipeptides generated, a search of their sequences in BIOPEP-UWM database [79] showed that the dipeptide FK was previously described in sweet potato hydrolyzates and associated with ACE inhibitory and anti-hyperuricemic properties [80,81]. Dipeptide FR has also been described to display ACE and DPPIV inhibitory actions [82,83], as it has been demonstrated for sequence GF [83,84]. ACE inhibitory effects were also observed for RF peptide [85,86]. In the case of the tripeptide ARF, it has been recently described that this specific sequence can not only inhibit ACE but also its homologue ACE2 [87]. Regarding the rest of peptides, to our knowledge, no previous research reported their presence or activity, which reinforces the potential of N. gadinata as a sustainable source of both known and new bioactive peptides. The predicted bioactivity of peptides may be influenced by the position of AA within their sequence. For instance, the placement of hydrophobic or branched-chain AA at the C-terminal, or the presence of aliphatic AA at the N-terminal, can enhance radical scavenging of reactive oxygen species or the ACE inhibitory activity of the peptides [88].
In our case, six peptides contained phenylalanine (FHPR, FSPR, FIPGL, FARPGL, FGPQGG and FLPPAL) and three peptides contained proline (PCMIR, PFGNR and PRPMR) at the N-terminal residue. Moreover, peptides with phenylalanine contained other hydrophobic AA at the C-terminal, such as leucine (L) and glycine (G), suggesting that the position of the AA in the sequence could be a key parameter behind the predicted bioactivities.
In silico proteolysis does not guarantee that the resultant bioactive peptides are safe for use in food and pharmaceutical applications. For this reason, we conducted a physicochemical and pharmacokinetic analysis of the selected 27 peptides (Table 3). The SwissADME evaluation provides information on the likelihood that a molecule can be safely used as an oral drug. Ideally, the molecular mass of a potential drug should be less than 500 Da, its lipophilicity should be ranged from −0.7 to 5.0, and its water solubility should not be exceeding 6 [42]. As can be observed in Table 3, all dipeptides met these criteria and presented a bioavailability score of 0.55, suggesting that these peptides could potentially be absorbed through the intestinal tract. In addition, other three peptides (ARF, VPGF, and FIPGL) showed intestinal absorption values higher than 20%. One them, ARF, showed the highest absorption value (71.49%) although its lipophilicity value was not into the ideal range. FIPGL was the potentially bioactive peptide with the largest sequence and its intestinal absorption value was higher than 20%, meeting SwissADME of drug-likeness criteria. The rest of peptides exhibited poor intestinal absorption values, and some were predicted to be incapable of being absorbed through the intestinal tract (Table 3). However, these peptides should not be discarded since they could display bioactive effects locally over the cells and tissues of the gastrointestinal tract without being absorbed into the bloodstream [89]. None of the 27 peptides analyzed showed any AMES toxicity.
3.4. Peptide Molecular Docking
Since in silico analysis only predicted the potential bioactive peptides released after simulated gastrointestinal digestion of N. gaditana proteins, without analyzing their interactions with biological targets, we decided to evaluate the potential mechanism of action through molecular docking of the ACE, DPPIV and MPO enzymes as a target-specific analysis for the antihypertensive, antidiabetic and antioxidant activities. Seven peptides showing an intestinal absorption value higher than 20% (GF, FK, FR, RF, ARF, VPGF and FIPGL) were selected. The Gibbs free energy (ΔG) quantifies the thermodynamic feasibility of a ligand binding to a target, and it is a central parameter related to binding affinity, where the interaction with the lowest ΔG is typically considered the most stable and biologically relevant position [90]. All the dipeptides and the ARF tripeptide showed high binding affinity with the three enzymes tested. Gibbs free energy values ranged from -10.7 to -7.9 kcal/mol for ACE, from -6.7 to -1.7 kcal/mol for DPPIV, and from -8 to -6.6 kcal/mol for MPO, suggesting that they might display their corresponding potential antioxidant, antidiabetic and antihypertensive activities through the inhibition of these well described enzymes at a systemic level. In the case of VPGF and FIPGL, both peptides showed high binding affinities for MPO and ACE. The molecular interactions of FIPGL with ACE and MPO enzymes are shown in Figure 4.
FIPGL peptide demonstrated to be resistant to the action of trypsin and chymotrypsin and potentially capable to exert multifunctional properties, making it an interesting peptide to be further explored. In the case of MPO (Figure 4A and 4B), FIPGL showed high MPO binding affinity, with a Gibbs free energy value of -9.6 kcal/mol. The peptide-enzyme interaction would be through conventional hydrogen bonds (I160, N162), alkyl and Pi-alkyl interactions (I160, R31, A35), carbon hydrogen bonds (D321) as well as van de Walls forces (Figure 4B). In the human body, the MPO normally interacts with hydrogen peroxide (H₂O₂) released by the phagocytic cells when they encounter foreign particles, forming a highly reactive enzyme-substrate complex [91]. The elevated MPO activity has been linked to various pathological conditions, increasing the risk of oxidative stress, and previous antioxidant peptides have been developed to use MPO as a therapeutic target [92]. Therefore, the demonstrated interaction between FIPGL and the MPO might influence the catalytic turnover or substrate access of the MPO, favoring the decrease of its oxidative capacity. This could be a possible mechanism of action behind FIPGL’s in silico predicted antioxidant activity. Nevertheless, other mechanisms of action such as neutralizing ROS, chelating pro-oxidant metal ions or modulating endogenous antioxidant defense systems could be also behind the predicted antioxidant activity.
Figure 4.
Molecular docking of the potential bioactive peptide FIPGL from Nannochloropsis gaditana digest with the human myeloperoxidase (MPO) and the angiotensin converting enzyme (ACE). A) 3D molecular docking representation of FIPGL with MPO; B) interaction type between FIPGL and MPO; C) 3D molecular docking representation of FIPGL with ACE; D) interaction type between FIPGL and ACE.
Figure 4.
Molecular docking of the potential bioactive peptide FIPGL from Nannochloropsis gaditana digest with the human myeloperoxidase (MPO) and the angiotensin converting enzyme (ACE). A) 3D molecular docking representation of FIPGL with MPO; B) interaction type between FIPGL and MPO; C) 3D molecular docking representation of FIPGL with ACE; D) interaction type between FIPGL and ACE.
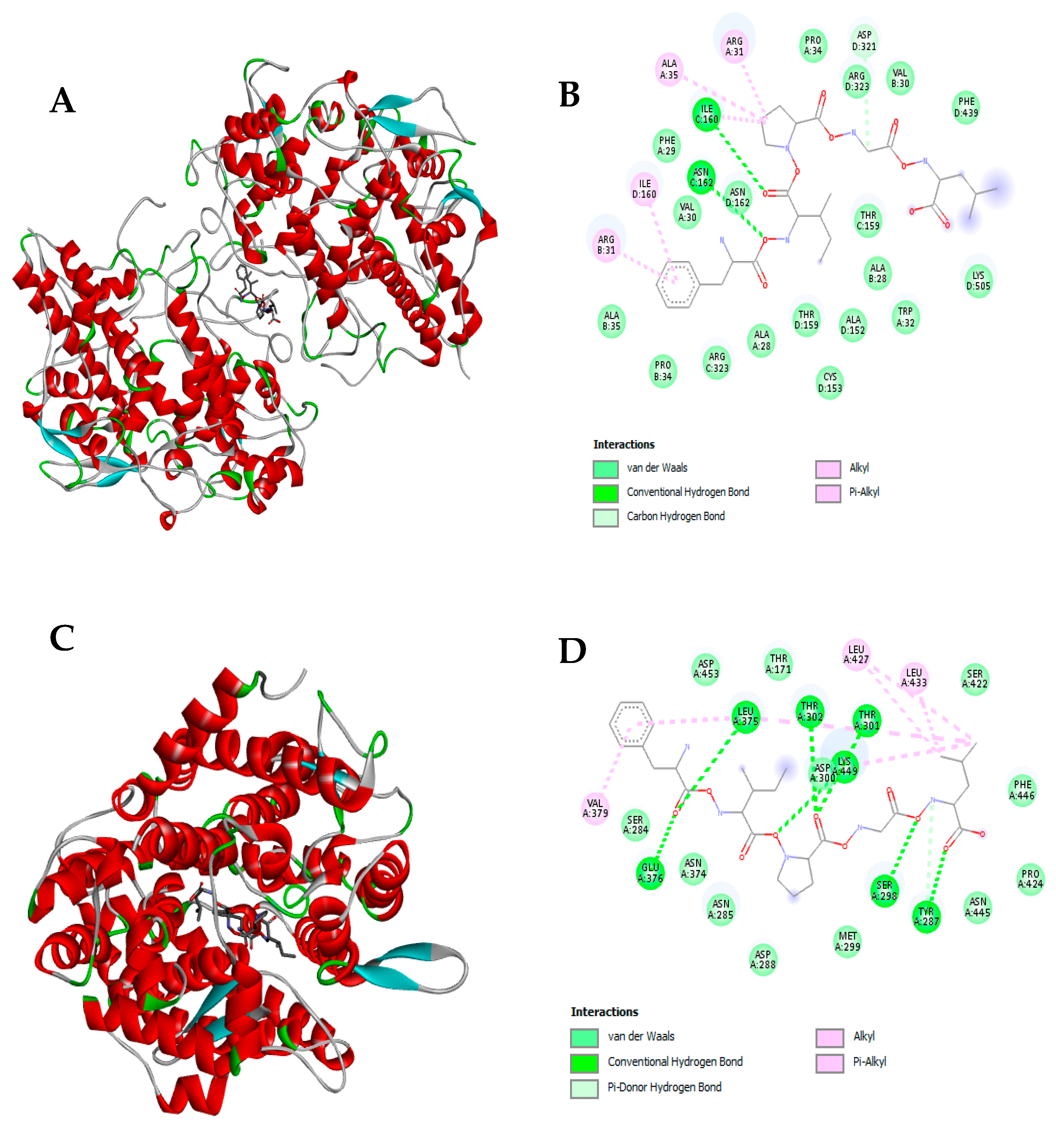
In relation to ACE (Figure 4C and 4D), FIPGL also showed high binding affinity with a Gibbs free energy value of -8.5 kcal/mol. In Figure 4D, FIPGL aminoacidic interactions with ACE were mainly mediated by hydrogen bonds (L375, E376, T301, T302, K449, S298, and Y287), alkyl and Pi-alkyl interactions (V379, L427, L433) and van de Walls forces.
ACE converts angiotensin-I into angiotensin-II, a vasoconstrictor, and its overproduction is one of the main causes of high blood pressure in patients with hypertension. Thus, suitable bioactive food-derived peptides have been studied as inhibitors of this enzyme and natural therapeutic alternatives [93]. The presence and positioning of hydrophobic AA residues such as isoleucine, phenylalanine, or proline would be a crucial structural feature for the ACE inhibitory potential [94]. A closer look regarding the structure of ACE has revealed that the active site consists of a zinc coordination site, several hydrogens bonding sub-sites, a proline binding pocket, and a hydrophobic pocket (with “F” residues) with a preference for aromatic AA like phenylalanine [95]. Consequently, it is imperative for a potent peptide inhibitor to have a hydrophobic residue with an aromatic side chain, such as phenylalanine and proline in its sequence for effective binding. Therefore, FIPGL, could be consider a potentially strong ACE inhibitory peptide. Our results were in tune with previous works carried out with other Nannochloropsis species like N. oculata that have been reported to generate protein hydrolyzates and peptides with ACE1 inhibitory properties [17,96]. Still, quantitative structure–activity relationship models of FIPGL and ACE should be done to further explore the mechanisms of action behind the predicted antihypertensive activity. Finally, regarding the DPPIV, both peptides did not show any binding affinity, obtaining positive Gibbs free energy (data not shown). This result suggested that the predicted antidiabetic activity of these peptides could be related with other pathways, such as enhancement of insulin secretion, mimicking of incretin hormones, and/or inhibition of key enzymes involved in carbohydrate digestion, such as α-amylase and α-glucosidase.
5. Conclusions
Our proteomic analysis was able to deepen the scarce knowledge available on the N. gaditana proteome. A total of 1,955 proteins were identified, of which 1,258 were identified with two or more unique peptides. The functional analysis of identified proteins demonstrated that most of them were related with cellular anatomic structures and cellular and binding processes. Furthermore, in silico gastrointestinal digestion and bioactivity prediction revealed the release of small-sized peptides mainly composed of hydrophobic AA, like phenylalanine and proline, with multiple potential biological activities. The SwissADME analysis showed that these peptides could exert their predicted bioactivities at both local and/or systemic level. Additionally, the molecular docking of potentially absorbable peptides showed high molecular affinity with MPO, ACE and DPPIV enzymes. It was especially remarkable in the case of the FIPGL peptide, which showed resistance to the action of digestive enzymes, capability to be absorbed through the gastrointestinal tract and multifunctionality. This peptide demonstrated strong affinity for MPO and ACE, potential targets of its predicted biological action. Overall, our proteomic and in silico approach suggests, for the first time, the potential of N. gaditana as a sustainable source of bioactive peptides with multiple health benefits.
Supplementary Materials
The following supporting information can be downloaded at website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, P.G.-C. and B.H.-L.; methodology, S.P., L.A.-P., and E.M.-L; software, S.G.F.; formal analysis, S.P. and L.A.-P.; investigation, S.P., L.A.-P., and E.M.-L.; writing—original draft preparation, S.P., and L.A.-P.; writing—review and editing, E.M.-L, P.G.-C., and B.H.-L.; supervision, P.G.-C. and B.H.-L.; funding acquisition, P.G.-C. and B.H.-L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported through Project PID2021-122989OB-I00 financed by MICIU/AEI/10.13039/501100011033 and FEDER, EU. Samuel Paterson gratefully acknowledges the Comunidad Autónoma de Madrid for a predoctoral contract (PIPF-2022/BIO-24996).
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bennett, J.M.; Reeves, G.; Billman, G.E.; Sturmberg, J.P. Inflammation–Nature’s Way to Efficiently Respond to All Types of Challenges: Implications for Understanding and Managing “the Epidemic” of Chronic Diseases. Front Med (Lausanne) 2018, 5, 316. [Google Scholar] [CrossRef] [PubMed]
- Budreviciute, A.; Damiati, S.; Sabir, D.K.; Onder, K.; Schuller-Goetzburg, P.; Plakys, G.; Katileviciute, A.; Khoja, S.; Kodzius, R. Management and Prevention Strategies for Non-Communicable Diseases (NCDs) and Their Risk Factors. Front Public Health 2020, 8, 574111. [Google Scholar] [CrossRef] [PubMed]
- Floret, C.; Monnet, A.F.; Micard, V.; Walrand, S.; Michon, C. Replacement of Animal Proteins in Food: How to Take Advantage of Nutritional and Gelling Properties of Alternative Protein Sources. Crit Rev Food Sci Nutr 2023, 63, 920–946. [Google Scholar] [CrossRef] [PubMed]
- Calvez, J.; Azzout-Marniche, D.; Tomé, D. Protein Quality, Nutrition and Health. Front Nutr 2024, 11, 1406618. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, D.; He, N.; Khoo, K.S.; Ng, E.P.; Chew, K.W.; Ling, T.C. Application Progress of Bioactive Compounds in Microalgae on Pharmaceutical and Cosmetics. Chemosphere 2022, 291, 132932. [Google Scholar] [CrossRef] [PubMed]
- Mosibo, O.K.; Ferrentino, G.; Udenigwe, C.C. Microalgae Proteins as Sustainable Ingredients in Novel Foods: Recent Developments and Challenges. Foods 2024, 13, 733. [Google Scholar] [CrossRef]
- Chronakis, I.S.; Madsen, M. Algal Proteins. Handbook of Food Proteins 2011, 353–394. [Google Scholar] [CrossRef]
- Rzymski, P.; Budzulak, J.; Niedzielski, P.; Klimaszyk, P.; Proch, J.; Kozak, L.; Poniedziałek, B. Essential and Toxic Elements in Commercial Microalgal Food Supplements. J Appl Phycol 2019, 31, 3567–3579. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial Applications of Microalgae. J Biosci Bioeng 2006, 101, 87–96. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Navarro-Juárez, R.; López-Martínez, J.C.; Campra-Madrid, P.; Rebolloso-Fuentes, M.M. Functional Properties of the Biomass of Three Microalgal Species. J Food Eng 2004, 65, 511–517. [Google Scholar] [CrossRef]
- Medina, C.; Rubilar, M.; Shene, C.; Torres, S.; Verdugo, M. Protein Fractions with Techno-Functional and Antioxidant Properties from Nannochloropsis Gaditana Microalgal Biomass. J Biobased Mater Bioenergy 2015, 9, 417–425. [Google Scholar] [CrossRef]
- Zaky, A.A.; Paterson, S.; Gómez-Cortés, P.; Hernández-Ledesma, B. Bioactive Peptides Released from Microalgae during Gastrointestinal Digestion. Protein Digestion-Derived Peptides. [CrossRef]
- Du, C.; Gong, H.; Zhao, H.; Wang, P. Recent Progress in the Preparation of Bioactive Peptides Using Simulated Gastrointestinal Digestion Processes. Food Chem 2024, 453, 139587. [Google Scholar] [CrossRef]
- Sheih, I.C.; Wu, T.K.; Fang, T.J. Antioxidant Properties of a New Antioxidative Peptide from Algae Protein Waste Hydrolysate in Different Oxidation Systems. Bioresour Technol 2009, 100, 3419–3425. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X. Isolation and Identification of Anti-Proliferative Peptides from Spirulina Platensis Using Three-Step Hydrolysis. J Sci Food Agric 2017, 97, 918–922. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X. Separation, Antitumor Activities, and Encapsulation of Polypeptide from Chlorella Pyrenoidosa. Biotechnol Prog 2013, 29, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.; Mora, L.; Lucakova, S. Identification of Bioactive Peptides from Nannochloropsis Oculata Using a Combination of Enzymatic Treatment, in Silico Analysis and Chemical Synthesis. Biomolecules 2022, 12, 1806. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.S.; Ryu, B.M.; Kim, S.K. Purification of Novel Anti-Inflammatory Peptides from Enzymatic Hydrolysate of the Edible Microalgal Spirulina Maxima. J Funct Foods 2013, 5, 1336–1346. [Google Scholar] [CrossRef]
- Md Saleh, N.I. Optimization of Enzymatic Hydrolysis for the Production of Antioxidative Peptide from Nannochloropsis Gaditana Using Response Surface Methodology. Pertanika J Sci & Technol 2019, 27, 41–55. [Google Scholar]
- Li, Y.; Aiello, G.; Fassi, E.M.A.; Boschin, G.; Bartolomei, M.; Bollati, C.; Roda, G.; Arnoldi, A.; Grazioso, G.; Lammi, C. Investigation of Chlorella Pyrenoidosa Protein as a Source of Novel Angiotensin I-Converting Enzyme (Ace) and Dipeptidyl Peptidase-Iv (Dpp-Iv) Inhibitory Peptides. Nutrients 2021, 13, 1624. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Cai, S.; Ryu, B.; Zhou, C.; Hong, P.; Qian, Z.J. An ACE Inhibitory Peptide from Isochrysis Zhanjiangensis Exhibits Antihypertensive Effect via Anti-Inflammation and Anti-Apoptosis in HUVEC and Hypertensive Rats. J Funct Foods 2022, 92, 105061. [Google Scholar] [CrossRef]
- Cavonius, L.R.; Albers, E.; Undeland, I. In Vitro Bioaccessibility of Proteins and Lipids of PH-Shift Processed Nannochloropsis Oculata Microalga. Food Funct 2016, 7, 2016–2024. [Google Scholar] [CrossRef] [PubMed]
- Peredo-Lovillo, A.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Romero-Luna, H.E. Conventional and in Silico Approaches to Select Promising Food-Derived Bioactive Peptides: A Review. Food Chem X 2021, 13, 100183. [Google Scholar] [CrossRef] [PubMed]
- Paterson, S.; Fernández-Tomé, S.; Galvez, A.; Hernández-Ledesma, B. Evaluation of the Multifunctionality of Soybean Proteins and Peptides in Immune Cell Models. Nutrients 2023, 15, 1220. [Google Scholar] [CrossRef] [PubMed]
- Official Methods of Analysis, 22nd Edition (2023) - AOAC INTERNATIONAL Available online:. Available online: https://www.aoac.org/official-methods-of-analysis/ (accessed on December 2024).
- Torres-Sánchez, E.; Morato, E.; Hernández-Ledesma, B.; Gutiérrez, L.F. Proteomic Analysis of the Major Alkali-Soluble Inca Peanut (Plukenetia Volubilis) Proteins. Foods 2024, 13, 3275. [Google Scholar] [CrossRef] [PubMed]
- Sanchiz, Á.; Morato, E.; Rastrojo, A.; Camacho, E.; la Fuente, S.G. De; Marina, A.; Aguado, B.; Requena, J.M. The Experimental Proteome of Leishmania Infantum Promastigote and Its Usefulness for Improving Gene Annotations. Genes (Basel) 2020, 11, 1–20. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins Silver-Stained Polyacrylamide Gels. Anal Chem 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Pisa, D.; Marina, A.I.; Morato, E.; Rábano, A.; Rodal, I.; Carrasco, L. Evidence for Fungal Infection in Cerebrospinal Fluid and Brain Tissue from Patients with Amyotrophic Lateral Sclerosis. Int J Biol Sci 2015, 11, 546. [Google Scholar] [CrossRef]
- Tran, N.H.; Qiao, R.; Xin, L.; Chen, X.; Liu, C.; Zhang, X.; Shan, B.; Ghodsi, A.; Li, M. Deep Learning Enables de Novo Peptide Sequencing from Data-Independent-Acquisition Mass Spectrometry. Nat Methods 2019, 16, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.H.; Zhang, X.; Xin, L.; Shan, B.; Li, M. De Novo Peptide Sequencing by Deep Learning. Proc Natl Acad Sci U S A 2017, 114, 8247–8252. [Google Scholar] [CrossRef]
- Tran, N.H.; Rahman, M.Z.; He, L.; Xin, L.; Shan, B.; Li, M. Complete De Novo Assembly of Monoclonal Antibody Sequences. Sci Rep 2016, 6, 31730. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J Mol Biol 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Maillet, N. Rapid Peptides Generator: Fast and Efficient in Silico Protein Digestion. NAR Genom Bioinform 2020, 2, lqz004. [Google Scholar] [CrossRef] [PubMed]
- Mooney, C.; Haslam, N.J.; Pollastri, G.; Shields, D.C. Towards the Improved Discovery and Design of Functional Peptides: Common Features of Diverse Classes Permit Generalized Prediction of Bioactivity. PLoS One 2012, 7, e45012. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Jiao, L.; Wang, R.; Zhao, Y.; Hao, Y.; Liang, G. Prediction of Antioxidant Peptides Using a Quantitative Structure-Activity Relationship Predictor (AnOxPP) Based on Bidirectional Long Short-Term Memory Neural Network and Interpretable Amino Acid Descriptors. Comput Biol Med 2023, 154, 106591. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.S.; Hasan, M.M.; Kurata, H. PreAIP: Computational Prediction of Anti-Inflammatory Peptides by Integrating Multiple Complementary Features. Front Genet 2019, 10, 129. [Google Scholar] [CrossRef] [PubMed]
- Gawde, U.; Chakraborty, S.; Waghu, F.H.; Barai, R.S.; Khanderkar, A.; Indraguru, R.; Shirsat, T.; Idicula-Thomas, S. CAMPR4: A Database of Natural and Synthetic Antimicrobial Peptides. Nucleic Acids Res 2023, 51, D377–D383. [Google Scholar] [CrossRef]
- Kumar, R.; Chaudhary, K.; Singh Chauhan, J.; Nagpal, G.; Kumar, R.; Sharma, M.; Raghava, G.P.S. An in Silico Platform for Predicting, Screening and Designing of Antihypertensive Peptides. Sci Rep 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Charoenkwan, P.; Nantasenamat, C.; Hasan, M.M.; Moni, M.A.; Lio’, P.; Manavalan, B.; Shoombuatong, W. StackDPPIV: A Novel Computational Approach for Accurate Prediction of Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides. Methods 2022, 204, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I. Using In Silico Approach for Metabolomic and Toxicity Prediction of Alternariol. Toxins (Basel) 2023, 15, 421. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci Rep 2017, 7, 42717. [Google Scholar] [CrossRef]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The Protein Data Bank. Nucleic Acids Res. 2000, 58, 899–907. [Google Scholar] [CrossRef]
- Luna-Vital, D.; Weiss, M.; Gonzalez de Mejia, E. Anthocyanins from Purple Corn Ameliorated Tumor Necrosis Factor-α-Induced Inflammation and Insulin Resistance in 3T3-L1 Adipocytes via Activation of Insulin Signaling and Enhanced GLUT4 Translocation. Mol Nutr Food Res 2017, 61, 1700362. [Google Scholar] [CrossRef]
- Villaseñor, V.M.; Navat Enriquez-Vara, J.; Urías-Silva, J.E.; del Carmen Lugo-Cervantes, E.; Luna-Vital, D.A.; Mojica, L. Mexican Grasshopper (Sphenarium Purpurascens) as Source of High Protein Flour: Techno-Functional Characterization, and in Silico and in Vitro Biological Potential. Food Research International 2022, 162, 112048. [Google Scholar] [CrossRef]
- Cavonius, L.R.; Albers, E.; Undeland, I. In Vitro Bioaccessibility of Proteins and Lipids of PH-Shift Processed Nannochloropsis Oculata Microalga. Food Funct 2016, 7, 2016–2024. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; Sáez, M.I.; Martínez, T.F.; Acién, F.G.; Alarcón, F.J. Differential Hydrolysis of Proteins of Four Microalgae by the Digestive Enzymes of Gilthead Sea Bream and Senegalese Sole. Algal Res 2019, 37, 145–153. [Google Scholar] [CrossRef]
- Levasseur, W.; Perré, P.; Pozzobon, V. A Review of High Value-Added Molecules Production by Microalgae in Light of the Classification. Biotechnol Adv 2020, 41, 107545. [Google Scholar] [CrossRef]
- Paterson, S.; Gómez-Cortés, P.; de la Fuente, M.A.; Hernández-Ledesma, B. Bioactivity and Digestibility of Microalgae Tetraselmis Sp. and Nannochloropsis Sp. as Basis of Their Potential as Novel Functional Foods. Nutrients 2023, 15, 477. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.; Silva, J.; Santos, T.; Gangadhar, K.N.; Raposo, A.; Nunes, C.; Coimbra, M.A.; Gouveia, L.; Barreira, L.; Varela, J. Nutritional Potential and Toxicological Evaluation of Tetraselmis Sp. CtP4 Microalgal Biomass Produced in Industrial Photobioreactors. Molecules 2019, 24, 3192. [Google Scholar] [CrossRef] [PubMed]
- Millward, D.J.; Layman, D.K.; Tomé, D.; Schaafsma, G. Protein Quality Assessment: Impact of Expanding Understanding of Protein and Amino Acid Needs for Optimal Health. Am J Clin Nutr 2008, 87, 1576S–1581S. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Ma, B.; Zhang, K. Spider: software for protein identification from sequence tags with de novo sequencing error. J Bioinform Comput Biol 2011, 3, 697–716. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.U.; Yi, E.K.J.; Amin, N.D.M.; Ismail, M.N. An Empirical Analysis of Sacha Inchi (Plantae: Plukenetia Volubilis L.) Seed Proteins and Their Applications in the Food and Biopharmaceutical Industries. Appl Biochem Biotechnol 2024, 196, 4823–4836. [Google Scholar] [CrossRef]
- Torres-Sánchez, E.; Morato, E.; Hernández-Ledesma, B.; Gutiérrez, L.F. Proteomic Analysis of the Major Alkali-Soluble Inca Peanut (Plukenetia Volubilis) Proteins. Foods 2024, 13, 3275. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Acero, F.J.; Amil-Ruiz, F.; Durán-Peña, M.J.; Carrasco, R.; Fajardo, C.; Guarnizo, P.; Fuentes-Almagro, C.; Vallejo, R.A. Valorisation of the Microalgae Nannochloropsis Gaditana Biomass by Proteomic Approach in the Context of Circular Economy. J Proteomics 2019, 193, 239–242. [Google Scholar] [CrossRef]
- Ferreira, M.; Teixeira, C.; Abreu, H.; Silva, J.; Costas, B.; Kiron, V.; Valente, L.M.P. Nutritional Value, Antimicrobial and Antioxidant Activities of Micro- and Macroalgae, Single or Blended, Unravel Their Potential Use for Aquafeeds. J Appl Phycol 2021, 33, 3507–3518. [Google Scholar] [CrossRef]
- Hulatt, C.J.; Smolina, I.; Dowle, A.; Kopp, M.; Vasanth, G.K.; Hoarau, G.G.; Wijffels, R.H.; Kiron, V. Proteomic and Transcriptomic Patterns during Lipid Remodeling in Nannochloropsis Gaditana. Int J Mol Sci 2020, 21, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Wu, S.; Li, X.; Ge, B.; Zhou, C.; Yan, X.; Ruan, R.; Cheng, P. The Structure, Functions and Potential Medicinal Effects of Chlorophylls Derived from Microalgae. Marine Drugs 2024, 22, 65. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.G. In Silico Proteome Cleavage Reveals Iterative Digestion Strategy for High Sequence Coverage. Int Sch Res Notices 2014, 2014, 960902. [Google Scholar] [CrossRef]
- Kruchinin, A.G.; Bolshakova, E.I.; Barkovskaya, I.A. Bioinformatic Modeling (In Silico) of Obtaining Bioactive Peptides from the Protein Matrix of Various Types of Milk Whey. Fermentation 2023, 9, 380–380. [Google Scholar] [CrossRef]
- Han, X.; Liu, Y.; Chen, Z. Zinc Finger Protein LipR Represses Docosahexaenoic Acid and Lipid Biosynthesis in Schizochytrium Sp. Appl Environ Microbiol 2022, 88, e0206321. [Google Scholar] [CrossRef]
- Wang, R.; Li, J.; Zhang, F.; Miao, X. Non-Tandem CCCH-Type Zinc-Finger Protein CpZF_CCCH1 Improves Fatty Acid Desaturation and Stress Tolerance in Chlamydomonas Reinhardtii. J Agric Food Chem 2023, 71, 17188–17201. [Google Scholar] [CrossRef]
- Ashtiani, F.R.; Jalili, H.; Rahaie, M.; Sedighi, M.; Amrane, A. Effect of Mixed Culture of Yeast and Microalgae on Acetyl-CoA Carboxylase and Glycerol-3-Phosphate Acyltransferase Expression. J Biosci Bioeng 2021, 131, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Bardani, E.; Kallemi, P.; Tselika, M.; Katsarou, K.; Kalantidis, K. Spotlight on Plant Bromodomain Proteins. Biology 2023, 12, 1076. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Li, Y.; Wu, X.; Xu, W.; Zhang, Z.; Zhu, H.; Zhou, H. NgAP2a Targets KCS Gene to Promote Lipid Accumulation in Nannochloropsis Gaditana. Int J Mol Sci 2024, 25, 10305. [Google Scholar] [CrossRef] [PubMed]
- Heda, R.; Toro, F.; Tombazzi, C.R.; Physiology, Pepsin. StatPearls 2023. Available online: https://www.statpearls.com (accessed on December 2024).
- Salelles, L.; Floury, J.; Le Feunteun, S. Pepsin Activity as a Function of PH and Digestion Time on Caseins and Egg White Proteins under Static in Vitro Conditions. Food Funct 2021, 12, 12468–12478. [Google Scholar] [CrossRef] [PubMed]
- Purohit, K.; Reddy, N.; Sunna, A. Exploring the Potential of Bioactive Peptides: From Natural Sources to Therapeutics. Int J of Mol Sci 2024, 25, 1391. [Google Scholar] [CrossRef]
- Yao, S.; Agyei, D.; Udenigwe, C.C. Structural Basis of Bioactivity of Food Peptides in Promoting Metabolic Health. Adv Food Nutr Res 2018, 84, 145–181. [Google Scholar] [CrossRef]
- Bhuyan, B.J.; Mugesh, G. Antioxidant Activity of Peptide-Based Angiotensin Converting Enzyme Inhibitors. Org Biomol Chem 2012, 10, 2237–2247. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Li, B.; He, J.; Qian, P. Quantitative Structure–Activity Relationship Study of Antioxidative Peptide by Using Different Sets of Amino Acids Descriptors. J Mol Struct 2011, 998, 53–61. [Google Scholar] [CrossRef]
- Fernando, R.; Sun, X.; Rupasinghe, H.P.V. Production of Bioactive Peptides from Microalgae and Their Biological Properties Related to Cardiovascular Disease. Macromol 2024, 4, 582–596. [Google Scholar] [CrossRef]
- Pedroni, L.; Perugino, F.; Galaverna, G.; Dall’Asta, C.; Dellafiora, L. An In Silico Framework to Mine Bioactive Peptides from Annotated Proteomes: A Case Study on Pancreatic Alpha Amylase Inhibitory Peptides from Algae and Cyanobacteria. Nutrients 2022, 14, 4680. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and Computational Approaches to Estimate Solubility and Permeability in Drug Discovery and Development Settings. Adv Drug Deliv Rev 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Moriguchi, I.; Hirano, H.; Nakagome, I. Comparison of Reliability of Log P Values for Drugs Calculated by Several Methods. Chem Pharm Bull (Tokyo) 1994, 42, 976–978. [Google Scholar] [CrossRef]
- Moriguchi, I.; Hirono, S.; Liu, Q.; Nakagome, Izum. ; Matsushita, Y. Simple Method of Calculating Octanol/Water Partition Coefficient. Chem Pharm Bull (Tokyo) 1992, 40, 127–130. [Google Scholar] [CrossRef]
- Martin, Y.C. A Bioavailability Score. J Med Chem 2005, 48, 3164–3170. [Google Scholar] [CrossRef]
- Ji, D.; Xu, M.; Udenigwe, C.C.; Agyei, D. Physicochemical Characterisation, Molecular Docking, and Drug-Likeness Evaluation of Hypotensive Peptides Encrypted in Flaxseed Proteome. Curr Res Food Sci 2020, 3, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int J Mol Sci 2019, 20, 5978. [Google Scholar] [CrossRef]
- He, W.; Su, G.; Sun-Waterhouse, D.; Waterhouse, G.I.N.; Zhao, M.; Liu, Y. In Vivo Anti-Hyperuricemic and Xanthine Oxidase Inhibitory Properties of Tuna Protein Hydrolysates and Its Isolated Fractions. Food Chem 2019, 272, 453–461. [Google Scholar] [CrossRef]
- Chao, W.-W.; Lin, B.-F.; Juan, H.-F.; Huang, S.-C.; Tsai, W.-H.; Chen, Y.-C.; Wang, C.-C. Sweet Potato Storage Root Defensin and Its Tryptic Hydrolysates Exhibited Angiotensin Converting Enzyme Inhibitory Activity in Vitro. Bot Stu 2012, 53, 215–222. [Google Scholar]
- Peng, H.; Carretero, O.A.; Vuljaj, N.; Liao, T.D.; Motivala, A.; Peterson, E.L.; Rhaleb, N.E. Angiotensin-Converting Enzyme Inhibitors: A New Mechanism of Action. Circulation 2005, 112, 2436–2445. [Google Scholar] [CrossRef]
- Lan, V.T.T.; Ito, K.; Ohno, M.; Motoyama, T.; Ito, S.; Kawarasaki, Y. Analyzing a Dipeptide Library to Identify Human Dipeptidyl Peptidase IV Inhibitor. Food Chem 2015, 175, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.S.; Wang, F.L.; Ondetti, M.A.; Sabo, E.F.; Cushman, D.W. Binding of Peptide Substrates and Inhibitors of Angiotensin-Converting Enzyme. Importance of the COOH-Terminal Dipeptide Sequence. J Bio Chem 1980, 255, 401–407. [Google Scholar] [CrossRef]
- Dhanda, S.; Singh, J.; Singh, H. Hydrolysis of Various Bioactive Peptides by Goat Brain Dipeptidylpeptidase-III Homologue. Cell Biochem Funct 2008, 26, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Y, S.; K, W.; A, K.; S, I. Structure and Activity of Angiotensin I Converting Enzyme Inhibitory Peptides from Sake and Sake Lees. Biosci Biotechnol Biochem 1994, 58, 1767–1771. [Google Scholar] [CrossRef]
- Wang, Z.; Fan, H.; Bao, X.; Wu, J. Angiotensin-Converting Enzyme 2 Activation Is Not a Common Feature of Angiotensin-Converting Enzyme Inhibitory Peptides. J Agric Food Chem 2023, 71, 8867–8876. [Google Scholar] [CrossRef] [PubMed]
- Senadheera, T.R.L.; Hossain, A.; Dave, D.; Shahidi, F. In Silico Analysis of Bioactive Peptides Produced from Underutilized Sea Cucumber By-Products—A Bioinformatics Approach. Marine Drugs 2022, 20, 610. [Google Scholar] [CrossRef]
- Nong, N.T.P.; Hsu, J.L. Bioactive Peptides: An Understanding from Current Screening Methodology. Processes 2022, 10, 1114. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular Docking and Structure-Based Drug Design Strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef]
- Valadez-Cosmes, P.; Raftopoulou, S.; Mihalic, Z.N.; Marsche, G.; Kargl, J. Myeloperoxidase: Growing Importance in Cancer Pathogenesis and Potential Drug Target. Pharmacol Ther 2022, 236, 108052. [Google Scholar] [CrossRef] [PubMed]
- Vakhrusheva, T. V.; Sokolov, A. V.; Moroz, G.D.; Kostevich, V.A.; Gorbunov, N.P.; Smirnov, I.P.; Grafskaia, E.N.; Latsis, I.A.; Panasenko, O.M.; Lazarev, V.N. Effects of Synthetic Short Cationic Antimicrobial Peptides on the Catalytic Activity of Myeloperoxidase, Reducing Its Oxidative Capacity. Antioxidants 2022, 11, 2419–2419. [Google Scholar] [CrossRef]
- Majumder, K.; Chakrabarti, S.; Morton, J.S.; Panahi, S.; Kaufman, S.; Davidge, S.T.; Wu, J. Egg-Derived ACE-Inhibitory Peptides IQW and LKP Reduce Blood Pressure in Spontaneously Hypertensive Rats. J Funct Foods 2015, 13, 50–60. [Google Scholar] [CrossRef]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural Requirements of Angiotensin I-Converting Enzyme Inhibitory Peptides: Quantitative Structure-Activity Relationship Modeling of Peptides Containing 4-10 Amino Acid Residues. QSAR Comb Sci 2006, 25, 873–880. [Google Scholar] [CrossRef]
- Natesh, R.; Schwager, S.L.U.; Sturrock, E.D.; Acharya, K.R. Crystal Structure of the Human Angiotensin-Converting Enzyme-Lisinopril Complex. Nature 2003, 421, 551–554. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Mora, L.; Hayes, M. In Vitro and in Silico Approaches to Generating and Identifying Angiotensin-Converting Enzyme I Inhibitory Peptides from Green Macroalga Ulva Lactuca. Mar Drugs 2019, 17, 204. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Graphical representation of the workflow followed in the present study.
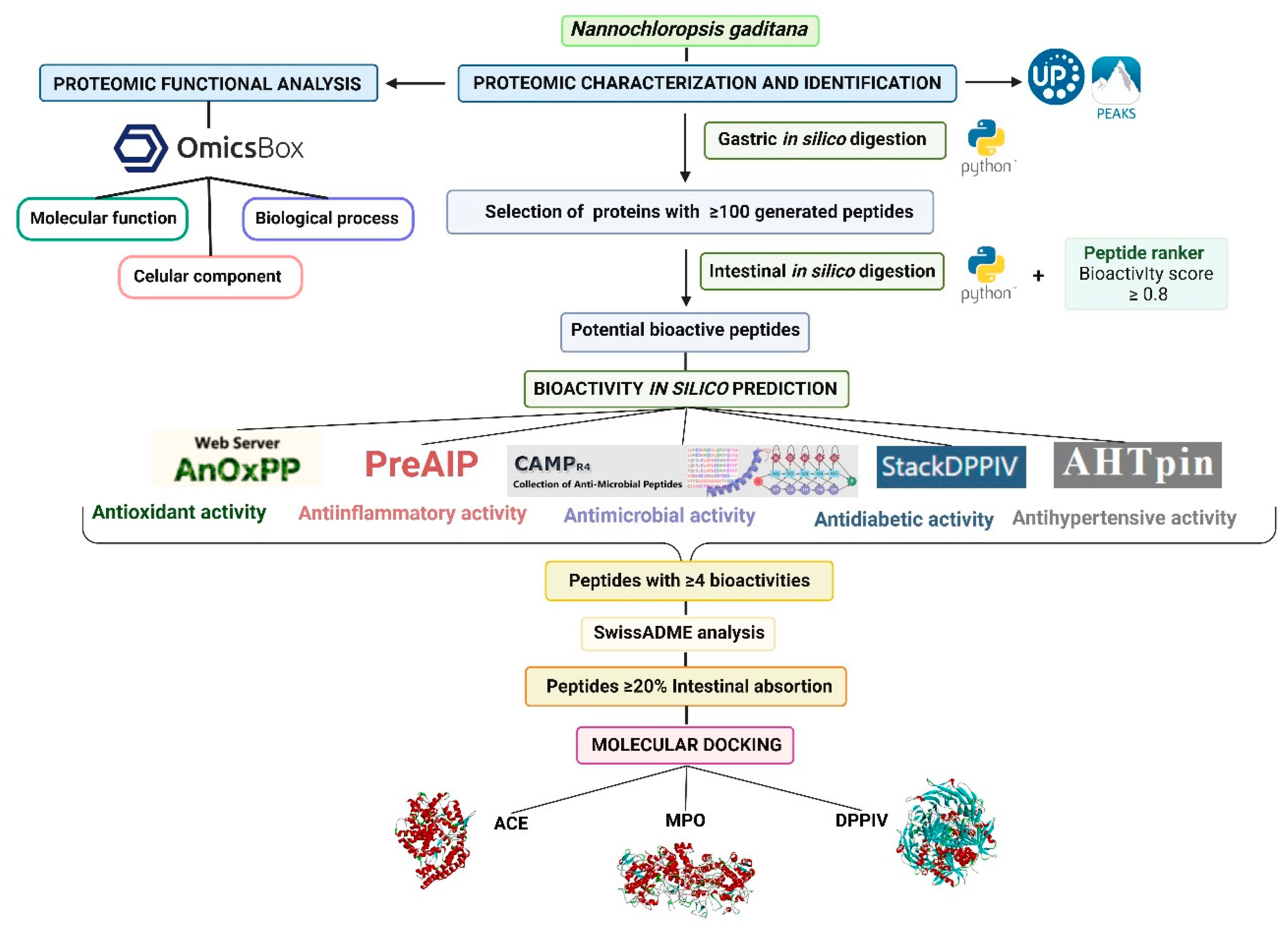
Figure 2.
Proteomic functional distribution of identified proteins from Nannochloropsis gaditana biomass using gene ontology (GO). A) Proteomic percentage distribution at GO level 1 classification. B) Cellular component GO level 2 distribution. C) Biological process GO level 2 distribution. D) Molecular function GO level 2 distribution.
Figure 2.
Proteomic functional distribution of identified proteins from Nannochloropsis gaditana biomass using gene ontology (GO). A) Proteomic percentage distribution at GO level 1 classification. B) Cellular component GO level 2 distribution. C) Biological process GO level 2 distribution. D) Molecular function GO level 2 distribution.
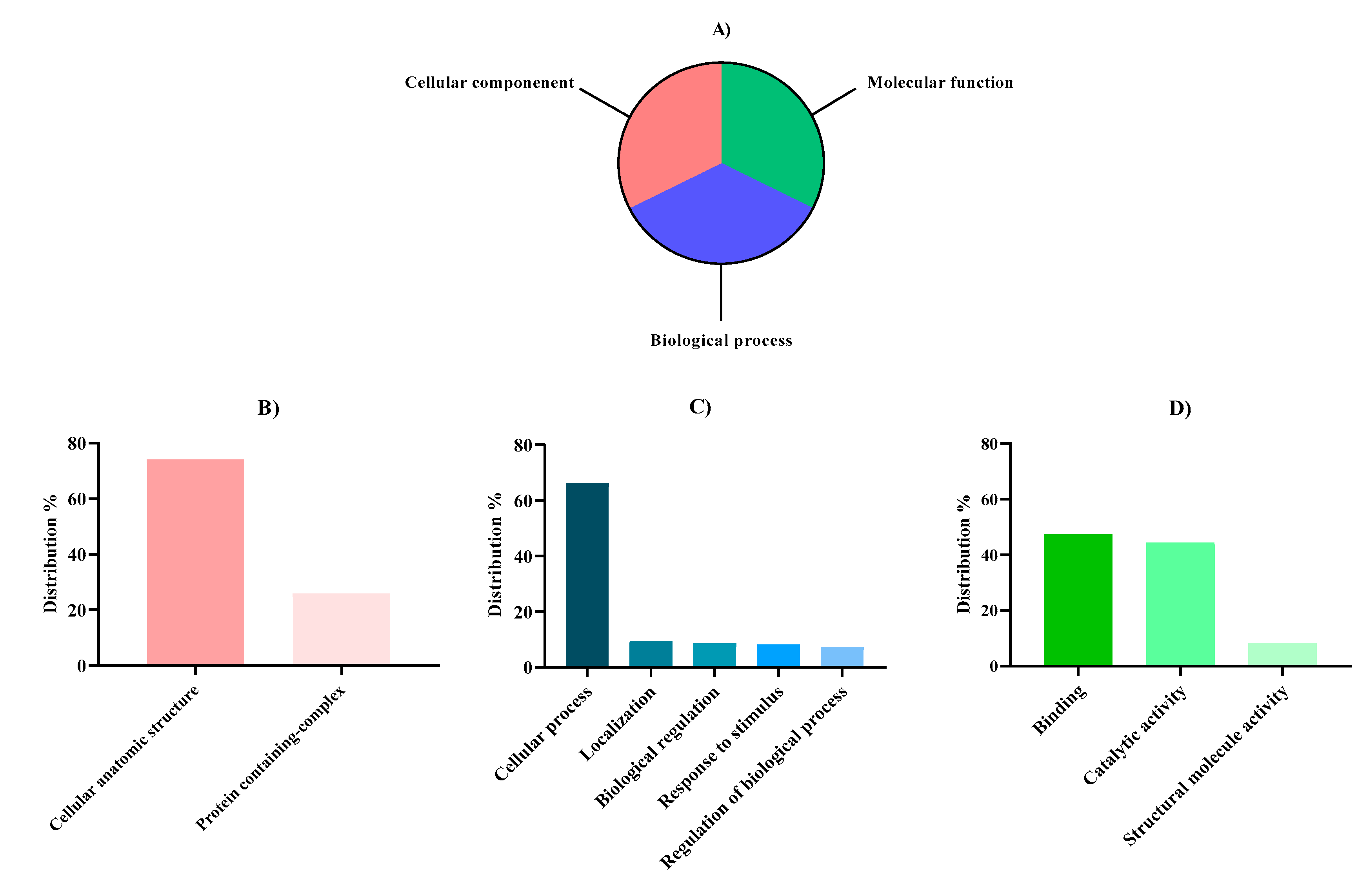

Table 1.
Amino acid profile (g/100 g protein and g/100 g biomass) of Nannochloropsis gaditana.
| Aminoacid | Content | FAO recommendation (g/100 g protein) |
|
|---|---|---|---|
| g/100 g protein | g/100 g biomass | ||
| Essential | |||
| Lysine (K) | 4.53 ± 0.09 | 2.01 ± 0.04 | 5.20 |
| Tryptophan (W) | n.d. | n.d. | 0.70 |
| Phenylalanine (F) | 3.62 ± 0.17 | 1.61 ± 0.08 | 4.60a |
| Tyrosine (Y) | 2.42 ± 0.07 | 1.07 ± 0.03 | |
| Methionine (M) | 1.57 ± 0.01 | 0.70 ± 0.01 | 2.60b |
| Cysteine (C) | 0.87 ± 0.12 | 0.38 ± 0.05 | |
| Threonine (T) | 3.40± 0.15 | 1.51 ± 0.06 | 2.70 |
| Leucine (L) | 5.97 ± 0.01 | 2.65 ± 0.00 | 6.30 |
| Isoleucine (I) | 2.53 ± 0.03 | 1.12 ± 0.01 | 3.10 |
| Valine (V) | 3.40 ± 0.02 | 1.51 ± 001 | 4.20 |
| Non-essential | |||
| Aspartic acid + Asparragine (D + N) | 6.44 ± 0.00 | 2.85 ± 0.00 | |
| Glutamic acid + Glutamine (E + Q) | 8.81 ± 0.09 | 3.91 ± 0.04 | |
| Serine (S) | 3.29 ± 0.15 | 1.46 ± 0.07 | |
| Histidine (H) | 1.38 ± 0.01 | 0.61 ± 0.00 | |
| Arginine (R) | 4.17 ± 0.00 | 1.85 ± 0.00 | |
| Alanine (A) | 5.02 ± 0,01 | 2.22 ± 0.01 | |
| Proline (P) | 6.44 ± 0,01 | 2.85 ± 0.00 | |
| Glycine (G) | 3.55 ± 0.02 | 1.57 ± 0.01 | |
| EAA | 28.31 | 14.13 | |
| NEAA | 39.01 | 17.28 | |
| TAA | 67.41 | 31.41 | |
| EAA*100/TAA (%) | 42.00 | ||
| EAA*100/NEAA (%) | 72.57 | ||
| HAA*100/TAA (%) | 47.23 | ||
| AAA*100/TAA (%) | 8.96 | ||
n.d. not detected a: F + Y; b: M + C; EAA: essential amino acids; NEAA: non-essential amino acids; TAA: total amino acids; HAA: hydrophobic amino acids (A + V + I + L + Y + F + W + M + P + C); AAA: aromatic amino acids (F + W + Y).
Table 2.
Nannochloropsis gaditana proteins that generated more than 100 peptides/protein after in silico digestion with pepsin. a Accession number of the protein as obtained from the FASTA database. b PEAKS protein score calculated as the weighted sum of the −10logP scores of the proteins supporting peptides. c Description of proteins as obtained from the FASTA database.
Table 2.
Nannochloropsis gaditana proteins that generated more than 100 peptides/protein after in silico digestion with pepsin. a Accession number of the protein as obtained from the FASTA database. b PEAKS protein score calculated as the weighted sum of the −10logP scores of the proteins supporting peptides. c Description of proteins as obtained from the FASTA database.
| Accession a | -10logP b | Description c | Average mass (KDa) |
Peptides generated after in silico gastric digestion |
| I2CQP5 | 426.87 | Acetyl-CoA carboxylase | 235,196 | 207 |
| W7U8G3 | 380.26 | ATP-citrate synthase | 120,317 | 102 |
| W7TN63 | 348.14 | Choline dehydrogenase | 138,839 | 113 |
| K8YSL1 | 287.43 | Aminopeptidase N | 137,581 | 102 |
| W7TQD1 | 264.44 | Pyruvate dehydrogenase E1 component subunit alpha | 120,314 | 103 |
| W7TTR4 | 192.91 | P-type atpase | 129,122 | 103 |
| W7UC18 | 192.19 | Phosphoribosylformylglycinamidine synthase | 145,158 | 113 |
| W7TNH0 | 182.55 | Pyruvate carboxilase | 134,055 | 102 |
| K8Z9A0 | 174.11 | Uncharacterized protein | 123,187 | 107 |
| W7TRK7 | 118.17 | Coatomer subunit alpha | 141,192 | 119 |
| W7UBG5 | 106.82 | Ubiquitin-activating enzyme e1 | 136,993 | 111 |
| W7U8K2 | 105.69 | Clathrin heavy chain | 196,113 | 163 |
| W7TM9 | 93.66 | Pentatricopeptide repeat containing protein | 171,849 | 132 |
| W7UBN0 | 87.72 | WD40-repeat-containing protein | 138,346 | 110 |
| W7U2D5 | 80.11 | Peptidase M16 | 143,228 | 109 |
| W7U0Z1 | 79.24 | Hydantoin utilization protein | 146,652 | 128 |
| W7U5E4 | 65.46 | Carbamoyl-phosphate synthase | 168,186 | 140 |
| W7U2B9 | 62.39 | Zinc finger. ZZ-type | 552,820 | 376 |
| K8YRI3 | 61.47 | DUF2428 domain-containing protein (Fragment) | 124,306 | 102 |
| W7TVB1 | 56.86 | Bromodomain containing 1 | 243,436 | 169 |
| W7TYI7 | 46.56 | Cytochrome p450 | 118,277 | 117 |
| W7U1T9 | 46.41 | Nuclear receptor corepressor 1-like protein | 164,110 | 122 |
| W7U7L8 | 45.19 | Protease-associated domain PA | 142,381 | 101 |
| W7TPR4 | 44.49 | Tubulin-specific chaperone d | 147,094 | 109 |
| TOTAL | 3160 |
Table 3.
Physicochemical and pharmacokinetic properties of peptides released from in silico gastrointestinal digestion of Nannochloropsis gaditana and predicted to have 4 or more bioactivities.
Table 3.
Physicochemical and pharmacokinetic properties of peptides released from in silico gastrointestinal digestion of Nannochloropsis gaditana and predicted to have 4 or more bioactivities.
| Peptide | Molecular weight (Da) | Lipophilicity (MLogP)a | Bioavailability scoreb | Water solubility (log mol/L)c |
% Intestinal absorptiond | AMES toxicitye |
|---|---|---|---|---|---|---|
| FK FR GF RF ARF1 DPMP FHPR FSPR HPKF MPPR VPGF APMRP FIPGL1, 2 GPGCG KAPPF KSPGW1 PCMIR PFGNR PRPMR RRCLF1 WWGGV YLPPR2 AVMPIF EFPMIR FARPGL1 FGPQGG1 FLPPAL1 |
293.36 | 0.6 | 0.55 | -2.818 | 35.3 | No |
| 321.37 | 0.15 | 0.55 | -2.643 | 21.61 | No | |
| 222.24 | 0.34 | 0.55 | -1.85 | 41.89 | No | |
| 321.37 | 0.14 | 0.55 | -2.617 | 21.43 | No | |
| 60.06 | -1.6 | 0.55 | 0.824 | 71.496 | No | |
| 458.53 | -1.11 | 0.11 | -2.27 | 0 | No | |
| 555.63 | -1.46 | 0.17 | -2.875 | 6.011 | No | |
| 505.57 | -1.56 | 0.17 | -2.835 | 0 | No | |
| 527.62 | -1.11 | 0.17 | -2.815 | 17.04 | No | |
| 499.63 | -1.01 | 0.17 | -2784 | 6.78 | No | |
| 418.49 | -0.1 | 0.55 | -2.524 | 28.49 | No | |
| 570.71 | -1.53 | 0.17 | -2.922 | 0 | No | |
| 545.67 | 0.15 | 0.17 | -3.174 | 21.02 | No | |
| 389.43 | -2.73 | 0.55 | -2.516 | 2.96 | No | |
| 558.67 | -0.6 | 0.17 | -2.833 | 16.56 | No | |
| 573.64 | -2.17 | 0.17 | -2.862 | 0.134 | No | |
| 618.81 | -1.32 | 0.17 | -2.889 | 0 | No | |
| 589.64 | -2.49 | 0.17 | -2.889 | 0 | No | |
| 655.81 | -1.98 | 0.17 | -2.889 | 0 | No | |
| 693.86 | -1.22 | 0.17 | -2.894 | 0 | No | |
| 603.67 | -0.74 | 0.17 | -3.015 | 19.76 | No | |
| 644.76 | -0.84 | 0.17 | -2.904 | 19.333 | No | |
| 676.87 | -0.07 | 0.17 | -3.012 | 11.265 | No | |
| 791.96 | -1.31 | 0.17 | -2.894 | 0 | No | |
| 659.78 | -1.51 | 0.17 | -2.922 | 0 | No | |
| 561.59 | -2.77 | 0.17 | -2.977 | 0.112 | No | |
| 656.81 | 0.12 | 0.17 | -3.29 | 17.79 | No |
a Lipophilicity expressed as the partition coefficient between octanol and water [74,75,76]. b Bioavailability score expressed as the probability of F > 10% in rat [77]. c Solubility of the molecule in water at 25ºC. d Percentage of the molecules that would be absorbed through the human intestine [78]. e Mutagenic potential of the molecule in bacteria [78]. 1 Peptides resistant to in silico intestinal digestion. 2 Peptides predicted to have all 5 tested bioactivities.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



