Submitted:
17 December 2024
Posted:
18 December 2024
You are already at the latest version
Abstract
Myxobolus species are Myxosporean parasites affecting various fish, causing diseases that weaken populations and result in substantial economic losses in aquaculture and fisheries. Despite extensive studies in Asia, Europe, and the Americas, there is limited research on Myxobolus in Africa, particularly in Tanzania, where fisheries and aquaculture are vital. This study investigated the prevalence and genetic characteristics of Myxobolus parasites in 384 fish samples from Dar es Salaam (Indian Ocean) and Mwanza (Lake Victoria). Samples were examined for cysts and spores using microscopy, followed by molecular characterization through PCR amplification of the 18S rDNA gene, Sanger sequencing, phylogenetic analysis, and genetic distance evaluation. The overall prevalence of Myxobolus was 12%, with a significantly higher prevalence in Mwanza (21.88%) compared to Dar es Salaam (6.25%). Statistical analysis revealed significant associations between prevalence, fish species, and locality. Phylogenetic analysis identified two genetic lineages within a monophyletic group, clustering with Myxobolus species from Israel, Egypt, and Ghana, suggesting potential novel species. Genetic distance analysis indicated greater variation in saltwater samples compared to freshwater. These findings highlight the higher prevalence of Myxobolus in freshwater and emphasize the need for targeted management strategies, continued surveillance, and research to safeguard fish populations and sustain aquaculture.
Keywords:
Myxobolus
; Prevalence
; PCR
; 18s rDNA gene
; Phylogenetic analysis
; Genetic distance
; Aquaculture
1. Introduction
Saltwater and freshwater fishes are integral in food security, where they serve as a primary protein source for millions of people worldwide. Tanzania has abundant water resources from freshwater (lakes, rivers, and dams) to Marine water (Territorial Sea, coastal line, and Exclusive Economic Zone) which is a potential source of fisheries and aquaculture production. Fish provide essential nutrients and contribute to the overall nutritional needs of the population, while also supporting the livelihoods of thousands involved in fishing, processing, and distribution [1,2]. Additionally, fish populations offer valuable ecosystem services, including water filtration, nutrient cycling, and coastal protection, which support industries like tourism, recreation, and coastal development, highlighting their multifaceted economic importance [3,4]. However, a wide range of pathogens affect fish production in different water sources, including bacteria, viruses, fungi, and parasites.
Myxobolus is a genus of parasitic organisms belonging to phylum Cnidaria, that infect various host species including fishes, amphibians, birds, elasmobranchs, reptiles, as well as mammals as intermediate hosts and annelids or bryozoans as final hosts. Myxobolus parasites in fishes have been reported around the world [5,6,7], where they are commonly found in freshwater with some other species found in marine (brackish water), and terrestrial habitats [8,9]. Myxobolus has more than 978 Myxosporean species where M. celebralis, M. pseudodispar, M. cyprini, M. squamalis and M. artus are some of the most commonly reported species [10]. Some of other species recently reported and characterized include M. grassi, M. bejeranoi, and M. dajiangensis [11,12,13,14]. In Africa, Myxobolus species have been reported in Cameroon[15,16], Benin[17], Egypt [18,19], Kenya, Uganda, and Ethiopia [20], revealing the ecological significance of these parasites in fishes and fish industries.
Myxobolus parasites produce cysts that inflict significant pathogenic effects on various organs of fish including gills, heart, liver, kidneys, spleen, gallbladder, gonads, intestine, and urethra, subsequently causing various diseases in fishes. Myxosporeans have been linked to severe diseases such as whirling disease[21], Proliferative kidney disease[22], and Gill disease, caused by several Myxobolus species[23]. Additionally, Myxobolus parasites can cause skin and muscle lesions, leading to localized inflammation, tissue damage, and reduced swimming performance[24,25,26]. The presence of Myxobolus parasites in fish populations can lead to reductions in population size due to increased mortality rates, and impaired reproduction, ultimately affecting the sustainability of fisheries, aquaculture, and the economy [27,28]. From a public health perspective, Myxobolus parasites have been reported in patients with gastrointestinal symptoms[29], in immunosuppressed patients [30], and also in human feces through ingestion of infected fishes [31]. Myxobolus parasites have been well documented in other continents including Asia, Europe, and American countries but comparably fewer studies have been documented in African countries. However, there is scarce to no information on documented and published studies of Myxobolus parasites in Tanzania despite their importance in fisheries and aquaculture sectors.
Molecular characterization techniques are employed to identify and differentiate Myxobolus species. The utilization of molecular methods provides accurate distinction between different Myxobolus species and strains. Furthermore, such genetic data may provide insights into transmission dynamics, host specificity, and potential host-switching events. Moreover, updated and additional data are needed to better understand Myxobolus divergence, interactions with fish hosts’ environment, and implications for fisheries management and aquaculture. Addressing these gaps will aid in developing effective disease control strategies and ensuring sustainable fish populations and food security. This study will provide the baseline information on molecular characteristics and assess the prevalence of Myxobolus parasites to reveal the extent of infection among fish populations and elucidate the distribution patterns of these parasites in aquatic ecosystems in saltwater and freshwater fishes from Dar es Salaam and Mwanza, Tanzania. The results are of utility in disease surveillance, taxonomic classification, and the development of targeted control measures to effectively manage Myxobolus infections and promote the health and conservation of saltwater and freshwater fish populations in the studied regions of Tanzania.
2. Results
2.1. Fish Species Morphometrics and Demographics
Three hundred and eighty-four fishes of six fish species were collected from Dar es Salaam and Mwanza in selected fish markets and or land sites. The species were Lates niloticus, Oreochromis niloticus, Synodontis victoriae, Rastrelliger kanagurta, Lethrinus nebulosus, and Caranx sexfasciatus. The fish species collected had a weight ranging from 14–288.5 g, and a length ranging from 10.1–26.5 cm. The greatest average length, mean length, mean weight, as well as weight range, was observed on fish belonging to the Lates niloticus species. Out of the 384 fishes collected, 42.97% were female (n=165) and 57.03% were male (n=219) as shown in Table 1.
2.2. Morphological and Morphometric Identification of Myxobolus Cysts and Spores
Detection of parasite infection was established through microscopic inspection of 384 fish samples to detect cysts and spores’ infestation. Cysts were white, round to ellipsoidal, and measured up to 6 mm in diameter. Myxospores were identified based on their distinctive morphology: an elongated, oval body with two equal-sized polar capsules (Figure 1). A total of 46 fish (12%) were found to be infected with Myxobolus cysts. The myxospores body measured (14.2-17 μm) in length, and (7.5–10.2 μm) in width, and the two polar capsules were equal in size and elongated, measuring (5.1–7 μm) in length and (1.2–2.2 μm) in width.
2.3. Prevalence of Myxobolus Parasites in Mwanza and Dar es Salaam
The prevalence of the parasite in the study sites was established based on the presence of Myxobolus cysts/spores’ infestation in fish. Three species of fish which are Lates niloticus, Oreochromis niloticus, and Caranx sexfasciatus were infested with Myxobolus parasites. The overall prevalence of Myxobolus parasites in fishes was 12% (46/384), being greater in Mwanza with 21.88%, (42/192) than in Dar es Salaam with 6.25%, (4/192). In selected study sites, the infestation was observed only in Mkuyuni, Kamanga, Mwaloni, Luchelele, and Ferry with 31.3% (15/48), 20.8% (10/48), 20.8% (10/48), 14.6% (7/48), 8.3% (4/48) prevalences respectively. Only fish gills were observed to be infested with multiple infestations of Myxobolus cysts. The results of this study show that host sex, length, and weight categories had no significant effect on the prevalence of Myxobolus parasites, with p-values 0.115, 0.066, and 0.105 respectively as shown in Table 2. It was found that fish species, and locality, are significant factors for Myxobolus parasites both having a p-value of 0.001(Table 2).
2.4. Polymerase Chain Reaction (PCR) Amplification of Myxobolus DNA
DNA from the 18S rDNA gene of the Myxobolus parasite was isolated from 30 positive samples (cysts and spores). These samples were selected as representative and subsequently used for nucleic acid extraction. Visualization following ethidium bromide staining and UV illumination using a gel documentation system (Vilber Lourmat, Germany) revealed successful amplification of the 18S rDNA gene from all analyzed samples, resulting in clear 1050 bp bands on the gel (Figure 2).
2.5. Sequencing and Nucleotide Sequence Analysis of the 18S rDNA Gene
Each of the sequences generated via automated dideoxynucleotide cycle sequencing was subjected to a BLASTn search to reveal homology through percentage identity, query cover, and E-value comparison. A list of the most homologous sequences to each of the samples subjected to BLASTn was identified and most sequences were similar to Myxobolus bejeranoi from Israel with a percentage identity ranging from 86.56% to 90.60%. The results are summarized in Table 3.
2.6. Phylogenetic Inference of Evolutionary Relationships
The phylogenetic analysis of 23 nucleotide sequences, including eight candidate sequences from this study with accession numbers (PQ387950, PQ387948, PQ387949, PQ387947, PQ387951, PQ387952, PQ387953, PQ387954) and 15 reference sequences obtained from NCBI's online repository, provided significant insights into the evolutionary relationships among these groups of sequences. The phylogenetic inference revealed that the candidate sequences formed two main genetic lineages within a single monophyletic group. Furthermore, these candidate sequences clustered within specific clades of the Myxobolus genus with very close to Myxobolus bejeranoi from Israel, and Myxobolus species from Egypt and Ghana. Furthermore, given that all species analyzed belong to the genus Myxobolus, the generated phylogenetic tree suggests a complex evolutionary history within this genus, characterized by multiple distinct clades representing divergent evolutionary lineages (Figure 3). Kudoa mirabilis and Kudoa thyrsites sequences were used as out-group sequences.
2.7. Genetic Divergence
The genetic distance between the candidate sequences was uneven, with the high distance observed to Dar es Salaam sequence (PQ387947) from the rest of the candidate sequences. Furthermore, the genetic distances among the sequences from the remaining samples were relatively lower (>0.1) with sequence pairs like PQ38750 and PQ38748 (0.009) as well as PQ38751 and PQ38752 (0.001) showing the lowest genetic distances (Table 4).
3. Discussion
The study established the prevalence and molecular characteristics of Myxobolus parasites in freshwater and saltwater fish from Mwanza and Dar es Salaam, Tanzania. The overall prevalence of Myxobolus parasites was 12%, with a higher occurrence in fish from freshwater ecosystems, highlighting their primary habitat. These findings align with the identification of new Myxobolus species, such as Myxobolus awadhii [36], which underscores the diversity and specificity of these parasites in freshwater fish, particularly in gill infections [37,38]. This notable difference in parasite prevalence between the two study regions is likely attributable to their respective geographical and chemical variations, suggesting that environmental factors or variations in local ecological conditions may play a critical role in influencing the prevalence of Myxobolus parasites [39].
The data indicate that Myxobolus parasites exhibit species-specific patterns, with Lates niloticus being particularly susceptible to infection. This finding aligns with observations from Cameroon, where similarly high prevalence rates were reported in this species [40,41]. This finding suggests a potential species-specific susceptibility to Myxobolus infections, as previously noted by [42]. Freshwater fish species are often more susceptible due to their life cycles and habitat preferences, which could have significant implications for fisheries targeting these species. Furthermore, the higher prevalence of these parasites in Lates niloticus can have a significant impact on its health, growth rates, and market value [38]. Given that Mwanza is the region from which the freshwater fish samples were collected, the high prevalence of Myxobolus parasites in Lates niloticus underscores the need for robust parasite monitoring and management strategies in aquaculture operations within this region. In conclusion, several studies have reported a high prevalence of Myxobolus parasites in freshwater fishes and a few species found in saltwater fishes [43,44].
Statistical analysis further supports initial findings on regional variation and species-specific prevalence of the candidate parasite. The significant variations in Myxobolus prevalence across different host species and locations (p < 0.05) highlight the need for a nuanced approach to parasite management, considering both species-specific and regional factors. Furthermore, this inferential statistical finding is in line with that provided by [45], who reported significant spatial and temporal variations in prevalence across host species and locations. In contrast, other candidate variables did not show statistically significant effects, suggesting that factors such as fish size and sex may not significantly influence parasite prevalence in this context [46]. These findings underscore the necessity for targeted health management strategies in both aquaculture and fisheries, with a particular focus on species-specific and regional factors. In the fisheries sector, such an understanding of regional disparities in Myxobolus prevalence can inform sustainable management practices and conservation strategies.
Despite robust amplification and extensive database searching, sequence similarity was insufficient for definitive species-level identification. This suggests that the amplified sequences may represent novel strains, closely related species, or potentially even new species within the Myxobolus genus. Hence, a phylogenetic analysis was conducted to refine species identification and further explore the evolutionary relationships, given the lower similarity percentages observed in the BLASTn results. However, the BLAST analysis highlights the genetic diversity within the Myxobolus genus and suggests the potential discovery of new species in the sampled regions.
Phylogenetic analysis revealed that the eight candidate sequences formed two distinct lineages within a monophyletic Myxobolus clade, indicating shared ancestry. Notably, Tanzanian Myxobolus sequences clustered with those from Israel, Egypt, and Ghana, suggesting a common evolutionary history and potential long-distance dispersal mechanisms. The clustering of these geographically distant species in the phylogeny suggests potential long-distance dispersal mechanisms, such as host migration or human activities [47]. This finding underscores significant biogeographical patterns and raises concerns about the potential dissemination of parasitic diseases among fish populations in Africa and the Middle East. The rapid spillover of an African parasite with a complex life cycle into the Middle East has been observed, potentially impacting aquaculture and wild fish populations [48].
The saltwater fish sample from Dar es Salaam exhibited the longest branch length, indicating a higher degree of genetic divergence compared to the freshwater fish samples from Mwanza. This suggests that the saltwater population has experienced older divergence events or a faster evolutionary rate. The geographical isolation of Dar es Salaam and the unique ecological pressures of the saltwater environment likely contributed to this divergence. As [49], demonstrated, distinct ecotypes can emerge due to ecological divergence, leading to localized genetic differences and potentially even taxonomic distinctions.
Genetic distance analysis revealed a significantly higher genetic distance in the saltwater sample, suggesting distinct evolutionary lineages. In contrast, the low genetic distances among freshwater sequences indicate close evolutionary relationships and limited genetic divergence, likely due to recent common ancestry or similar selective pressures. This pattern aligns with the isolation-by-distance model, where geographic distance limits gene flow and promotes genetic divergence [50,51]. Additionally, habitat variability, as exemplified by Gammarus tigrinus [52], underscores the role of environmental adaptation in driving genetic differentiation.
These findings suggest that the saltwater adaptation of the parasite may have influenced host-parasite interactions, potentially contributing to the observed genetic divergence. Furthermore, it could affect the parasite's host range and pathogenicity, with implications for disease transmission in marine fish populations. Contrary to the assumption of homogeneity in marine populations, evidence suggests significant genetic structuring and local adaptation, often driven by parasites [53,54]. These results emphasize the importance of environmental factors in shaping evolutionary pathways and highlight the need for further research on the mechanisms driving parasite evolution in diverse aquatic ecosystems.
4. Materials and Methods
4.1. Study Area
This study was conducted in Mwanza and Dar es Salaam. In Dar es Salaam, sampling sites included Kivukoni, Msasani, Ununio, and Kunduchi along the Indian Ocean, while Mwanza sites included Mkuyuni, Mwaloni, Kamanga, and Luchelele along Lake Victoria as shown in Figure 4. Dar es Salaam, situated at 6º 48ʻS, 39º 17ʻE, covers 1,393 km² and experiences a hot, humid tropical climate. It is bordered to the east by the Indian Ocean and is surrounded by the Pwani Region. Major activities in Dar es Salaam include fishing and trade. In contrast, Mwanza is located at 2.5164° S, 32.9175° E, spans 25,233 km², and lies between 1,200 and 1,400 meters above sea level. This region is bordered to the north by Lake Victoria and Kagera region, and east, south, and west by Mara, Simiyu, Shinyanga, and Geita regions respectively. Major economic activities here include fishing and agriculture.
4.1.1. Study Design, Selection of Study Area, and Sampling Strategies
A cross-sectional study design was employed to establish the prevalence and molecular characteristics of Myxobolus parasites in Mwanza along Lake Victoria (freshwater body) and Dar es Salaam along the Indian Ocean (saltwater body). Mwanza was purposively selected due to its role as a major mainland freshwater fish import and export depot, the presence of common fish species that host the targeted parasite, and the existence of prior studies on this parasite in Lake Victoria [55]. Dar es Salaam was chosen as a counterpart location due to its proximity to the Indian Ocean, a saltwater fishing hotspot, and the presence of previous studies on the parasite, which support the consideration of parasite transmission in this area [44]. Systematic sampling was employed to select aquaculture and fishery sites within each region, beginning with a random starting point at fish landing sites or markets then from the starting point, additional sites were chosen at regular intervals to ensure representative sampling. Within each selected market or landing site, fish were systematically chosen from the fish sellers and fishermen to capture variability. Sample collection occurred in the morning and evening to introduce variability, minimize sampling bias, and account for diurnal fluctuations. This approach allowed for the collection of a diverse range of fish from all selected sites from March to April of 2024. Cochran prevalence formula for estimation of sample size, N=Z2PQ/E2, where N= sample size, Z=1.96 (the number of standard deviations from the mean) corresponding to the desired confidence level, P=50% prevalence, Q= 1-P, E=5% precision error was employed in the study [56]. A total of 384 fish samples were included in the study, where 192 fish samples were obtained from each study location (Mwanza and Dar es Salaam).
4.1.2. Sample Collection and Transportation
Fish were collected from their landing sites and/or fish markets in Mwanza and Dar es Salaam for one month (March – April 2024). The Mackerel (Rastrelliger kanagurta), Spangled emperor (Lethrinus nebulosus), and Bigeye trevally (Caranx sexfasciatus) were the common fish species sampled in Dar es Salaam while the Nile perch (Lates niloticus), Nile tilapia (Oreochromis niloticus) and Lake Victoria squeaker (Synodontis victoriae) were the common fish species sampled in Mwanza. All the collected fish samples were bought from the fish markets and fishermen, and identified based on a protocol similar to that employed by [57], and then stored in a cool box containing ice packs for initial preservation. The samples were then transported to the parasitology laboratory at Sokoine University of Agriculture in Morogoro for parasite examination.
4.1.3. Identification of Features of Collected Fish Samples
The identification of fish sex was done in the parasitology laboratory, based on observation of morphological features particularly the genital papilla located posterior to the fish’s anus. Slight distinctions in the genital papilla were utilized to sort out the fish sexes where the female genital papilla was identified with the presence of an oviduct in addition to the urinary pore unlike that of the males. The weight of the fish was measured by using an electronic weighing machine model (SF-400) while the fish length was measured by using a standard measuring ruler. The location of the fish sample collection was recorded.
4.1.4. Parasites (Myxobolus Cysts/Spores) Detection, Fish Dissection and Storage
Collected fish were observed to identify the presence of the parasites (cysts) in the skin, scales, eyes, operculum, and fins with the aid of a stereomicroscope (OPTA-TECH, Warsaw, Poland) with a 4x objective lens. This was followed by fish dissection by using a sterilized surgical blade to allow examination of cysts infesting the fish’s internal organs including gills, intestines, liver, heart, kidney, spleen, and muscles. Samples (dissected organs) were placed onto Petri dishes and examined for the presence of cysts using a stereo microscope (OPTA-TECH, Warsaw, Poland) with a 4x objective lens. Detected and extracted tissue cysts were placed on the microscopic slides and crushed by using sterilized syringe needles to release spores, followed by the addition of one drop of distilled water to form a wet smear which was then covered with coverslips. The smear was then observed under the light microscope (OPTA-TECH, Warsaw, Poland) with an objective lens of 100x and oil immersion to identify and measure the spores belonging to the Myxobolus parasites based on guidelines set by [58]. Measurements were taken by using a calibrated eyepiece micrometer. The samples that tested positive upon microscopy were carefully preserved in falcon tubes containing 70% ethanol to await further molecular analysis.
4.1.5. Extraction of Myxobolus DNA
Genomic DNA was extracted using the Quick-DNA TM Universal kit (ZYMO Research, USA) following the manufacturer’s instructions. Each tissue sample in the Eppendorf tube received 95μl of nuclease-free water, 95μl of solid tissue buffer, and 10μl of proteinase K, and was then incubated at 55°C for 3 hours to ensure solubility. After incubation, the sample was centrifuged at 12,000xg for one minute, and the supernatant was carefully transferred to a clean tube. Subsequently, 400μl of genomic binding buffer was added to each 200μl volume of transferred supernatant. This mixture was then transferred to a Zymo-spinTM IIC-XL column placed in a collection tube and centrifuged at 12,000xg for 1 minute. The DNA purification process included three wash steps with various wash buffers per the protocol instructions. This involved using 400μl of DNA pre-wash buffer once, followed by two washes with 700μl and 200μl of g-DNA wash buffer, respectively. The purified DNA extracted was then eluted using 50μl of DNA elution buffer. Before storage, the concentration and quality of extracted DNA were measured using a NanoDropTM spectrophotometer. Finally, the extracted DNA was stored at -20°C for future use.
4.1.6. PCR Amplification, Gel Electrophoresis, and Amplicon Sequencing
PCR amplification was conducted targeting the 18S rDNA gene using forward (MC5F: 5’-CCT GAG AAA CGG CTA CCA CAT CCA-3’) and reverse (MC3R:5’-GAT TAG CCT GAC AGA TCA CTC CAC GA-3’) primers [59]. The Master mix prepared for the 25 μL PCR reaction per tube comprised an enzyme premix of 12.5 µl, template DNA 5 µl, nuclease-free water 4 µl, Bovine Serum Albumin (BSA) 2.5 µl, and the primers 1 µl. Amplification conditions were as follows; an initial denaturation step at 95˚C for 5 minutes, followed by 35 cycles of denaturation at 95˚C for 60 seconds, annealing temperature at 60˚C for 60 seconds, and an extension at 72˚C for 90 seconds. A final extension step at 72˚C for 5 minutes to terminate the amplification end process. The amplicons were separated by 1.5% agarose gel stained with ethidium bromide at 120 Volts for 45 minutes. Visualization and documentation were done by a gel documentation system (Vilber, Germany). A volume of 20 μL from each positive PCR product was sent to Macrogen Company (Europe, Netherlands), where the 18S rDNA gene sequences were generated via dideoxynucleotide cycle sequencing (Sanger sequencing). The Applied Biosystem’s 96 Capillary 3730xl DNA Analyzer (Thermo Fisher Scientific, Carlsbad, CA, USA) was used to sequence each sample in both forward and reverse directions preceded by purification of the PCR products.
4.2. Statistical Analysis
Collected data were coded and then entered in Microsoft Excel 2017 spreadsheets. The prevalence of Myxobolus parasites from freshwater and saltwater fish was determined by computing the proportions of fish that were infected by Myxobolus parasites within the populations. Prevalence of Myxobolus parasite infestation was hence calculated using the following formula; P = No /NT × 100 %, Where P: Prevalence, No: number of positive samples, and NT: number of total samples tested. The results were then expressed as a percentage. Descriptive statistics were employed to present distributions and relative occurrences of the parasite in tables. The SPSS software version 26, created by IBM Corporation, Armonk, NY, USA in 2017, on the other hand, was employed where the Chi-square test was used to assess the statistical significance between the prevalence of parasites and various study variables (locality, sex, length, weight, and species of fishes). Findings were considered statistically significant when the p-value was less than 0.05.
4.3. Molecular Analysis
Generated sequences were checked for quality, trimmed, edited, and assembled into consensus sequences using Bio Edit version 7.2.5. The consensus sequences of 18S rDNA in Myxobolus were then searched for similarity in the National Center for Biotechnology Information (NCBI) GenBank using the nucleotide Basic Local Alignment Search Tool (BLASTn). The consensus sequences and 15 sequences retrieved from NCBI GenBank were aligned using the ClustalW statistical algorithm built in Evolutionary Genetics Analysis (MEGA) version 11. The phylogenetic tree was constructed by using the Neighbor-Joining Method and 1000 bootstrap replications by using MEGA software version 11 to evaluate the relatedness of Myxobolus sp. isolated from this study with those from other published studies [35]. Genetic distance analysis was conducted using the Kimura 2-parameter model in MEGA 11 software where pairwise distance analysis for sequences was computed [34].
4.4. Ethical Statement
Ethical clearance for this study was approved by the directorate of research, consultancy, and technology transfer (DPRTC) of the Sokoine University of Agriculture, Morogoro Tanzania (Ref. no: DPRTC/R/186/23) issued on 21 June 2024. All protocols and procedures for sample collection and sample processing in this study complied to the direction of the Ethics Committee, Sokoine University of Agriculture.
5. Conclusions
This study provides the first baseline data on the prevalence and status of Myxobolus parasites in Tanzanian freshwater and saltwater fish. The presence of these parasites in various fish species raises concerns about potential disease outbreaks and economic impacts on the fisheries industry. Additionally, given the significant role of fish in the Tanzanian diet, there are concerns about the potential zoonotic transmission of these parasites to humans. Long-term monitoring studies are recommended to better understand the dynamics of these parasites. particularly focusing on seasonal variations in infection rates. Furthermore, research on Myxobolus parasites in annelid hosts, which serve as the final hosts, is essential, as no such studies currently exist in Tanzania. A comprehensive epidemiological investigation of these parasites and their zoonotic potential is crucial to assess the public health risks associated with these infections in the study area and throughout Tanzania.
Author Contributions
Conceptualization, J.L.M. and E.M.M.; sample collection, J.L.M.; laboratory analyses, J.L.M.; formal analysis, J.L.M. and A.A.C.; writing—original draft preparation, J.L.M.; writing—review and editing, G.G.B., A.A.C. and E.M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Mwalimu Julius K. Nyerere University of Agriculture and Technology under HEET Project.
Institutional Review Board Statement
Ethical clearance for this study was approved by the directorate of research, consultancy, and technology transfer (DPRTC) of the Sokoine University of Agriculture, Morogoro Tanzania (Ref. no: DPRTC/R/186/23) issued on 21 June 2024. All protocols and procedures for sample collection and sample processing in this study complied to the direction of the Ethics Committee, Sokoine University of Agriculture.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and/or analysed during the current study are available at the GenBank repository (https//ncbi.nlm.nih.gov/genbank) with accession numbers (PQ387950, PQ387948, PQ387949, PQ387947, PQ387951, PQ387952, PQ387953, PQ387954).
Acknowledgments
We thank Mr. Salim Omary and Mr. Mabula Msabula for their support in the parasitology lab, as well as Dr. Elisa Mwega, Ms. Gladness Manyasi, Mr. Erick Msafiri, and Mr. Yumba Nobart for their assistance during molecular laboratory activities.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Fish-Farming Value Chain Analysis: Policy Implications for Transformations and Robust Growth in Tanzania. J. Rural Community Dev. 2015, 10.
- Mbunda, A.S.; Kapinga, A.F. Mobile Phone Technology for Enhancing Small-Scale Fishing in Tanzania. A Case of Nyasa District. 2021. [Google Scholar]
- Borland, H.P.; Gilby, B.L.; Henderson, C.J.; Connolly, R.M.; Gorissen, B.; Ortodossi, N.L.; Rummell, A.J.; Pittman, S.J.; Sheaves, M.; Olds, A.D. Dredging Fundamentally Reshapes the Ecological Significance of 3D Terrain Features for Fish in Estuarine Seascapes. Landsc. Ecol. 2022, 37, 1385–1400. [Google Scholar] [CrossRef]
- Lal, D.M.; Sreekanth, G.B.; Soman, C.; Ramteke, K.K.; Kumar, R.; Abidi, Z.J. Fish community structure as an indicator of the ecological significance: A study from Ulhas River Estuary, Western coast of India. J Env. Biol 2020. [Google Scholar] [CrossRef]
- Benoit, L.F.G.; Sorel, D.N.A.; Abraham, F. Three New Species of Myxobolus (Myxosporea: Myxobolidae), Parasites of Barbus Callipterus Boulenger, 1907 in Cameroon. Asian J. Biol. Sci. 2017, 10, 110–120. [Google Scholar] [CrossRef]
- Tahir, U.B.; Guo, Q.; Zhao, D.; Liu, Y.; Gu, Z. Description of Myxobolus Xiantaoensis n. Sp. from the Fins of Yellow Catfish in China: A Species Previously Attributed to Myxobolus Physophilus Reuss, 1906 in Chinese Records. Parasitol. Res. 2019, 118, 1137–1146. [Google Scholar] [CrossRef]
- Chinh, N.N.; Tham, N.T.; Yurakhno, V.M.; Doanh, P.N.; Whipps, C.M.; Shirakashi, S. Description of Myxobolus Hoabinhensis n. Sp. (Myxosporea: Myxobolidae), Infecting the Trunk Muscles of Goldfish Carassius Auratus (Linnaeus, 1758) (Cypriniformes: Cyprinidae) in Northern Vietnam. Parasitol. Res. 2022, 121, 2495–2502. [Google Scholar] [CrossRef]
- Fiala, I.; Bartošová-Sojková, P.; Okamura, B.; Hartikainen, H. Adaptive Radiation and Evolution Within the Myxozoa. In Myxozoan Evolution, Ecology and Development; Okamura, B., Gruhl, A., Bartholomew, J.L., Eds.; Springer International Publishing: Cham, 2015. [Google Scholar] [CrossRef]
- Kodádková, A.; Bartošová-Sojková, P.; Holzer, A.S.; Fiala, I. Bipteria Vetusta n. Sp. – an Old Parasite in an Old Host: Tracing the Origin of Myxosporean Parasitism in Vertebrates. Int. J. Parasitol. 2015, 45, 269–276. [Google Scholar] [CrossRef]
- Eiras, J.C.; Cruz, C.F.; Saraiva, A.; Adriano, E.A. Synopsis of the Species of Myxobolus (Cnidaria, Myxozoa, Myxosporea) Described between 2014 and 2020. Folia Parasitol. (Praha) 2021, 68, 2021.012. [Google Scholar] [CrossRef]
- Fariya, N.; Kaur, H.; Singh, M.; Abidi, R.; El-Matbouli, M.; Kumar, G. Morphological and Molecular Characterization of a New Myxozoan, Myxobolus Grassi Sp. Nov. (Myxosporea), Infecting the Grass Carp, Ctenopharyngodon Idella in the Gomti River, India. Pathog. Basel Switz. 2022, 11, 303. [Google Scholar] [CrossRef]
- Lövy, A.; Smirnov, M.; Brekhman, V.; Ofek, T.; Lotan, T. Morphological and Molecular Characterization of a Novel Myxosporean Parasite Myxobolus Bejeranoi n. Sp. (Cnidaria: Myxosporea) from Hybrid Tilapia in Israel. Parasitol. Res. 2018, 117, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Mathews, P.D.; Mertins, O.; Milanin, T.; Espinoza, L.L.; Flores-Gonzales, A.P.; Audebert, F.; Morandini, A.C. Molecular Phylogeny and Taxonomy of a New Myxobolus Species from the Endangered Ornamental Fish, Otocinclus Cocama Endemic to Peru: A Host-Parasite Coextinction Approach. Acta Trop. 2020, 210, 105545. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, J.; Zhao, Y. Morphological Description and Molecular Identification of Myxobolus Dajiangensis n. Sp. (Myxozoa: Myxobolidae) from the Gill of Cyprinus Carpio in Southwest China. PeerJ 2022, 10, e13023. [Google Scholar] [CrossRef] [PubMed]
- Folefack, G.B.L.; Tiwa, A.E.T.; Dongmo, B.F.; Nguegang, L.M.; Fomena, A. Morphotaxonomy and Histopathology of Three Species of Myxobolus Bütschli, 1882 Parasites of Enteromius Martorelli Roman, 1971 from the Anga River in Cameroon. Int. J. Biol. Chem. Sci. 2019, 13, 1705–1719. [Google Scholar] [CrossRef]
- Georges, F.; Marc, K.K.; Timoléon, T.; Minette, T.E.; Joseph, T. Myxobolus (Myxosporea: Myxobolidae) Polyinfection Patterns in Oreochromis Niloticus in Adamawa-Cameroon. Int. J. Fish. Aquat. Stud. 2021, 9, 123–130. [Google Scholar] [CrossRef]
- Gbankoto, A.; Pampoulie, C.; Marques, A.; Sakiti, G. Occurrence of Myxosporean Parasites in the Gills of Two Tilapia Species from Lake Nokoué (Bénin, West Africa): Effect of Host Size and Sex, and Seasonal Patterns of Infection. Dis. Aquat. Organ. 2001, 44, 217–222. [Google Scholar] [CrossRef]
- Abdel-Gaber, R.; Abdel-Ghaffar, F.; Maher, S.; El-Mallah, A.-M.; Al Quraishy, S.; Mehlhorn, H. Morphological Re-Description and Phylogenetic Relationship of Five Myxosporean Species of the Family Myxobolidae Infecting Nile Tilapia. Dis. Aquat. Organ. 2017, 124, 201–214. [Google Scholar] [CrossRef]
- Abdel-Ghaffar, F.; El-Toukhy, A.; Al-Quraishy, S.; Al-Rasheid, K.; Abdel-Baki, A.S.; Hegazy, A.; Bashtar, A.-R. Five New Myxosporean Species (Myxozoa: Myxosporea) Infecting the Nile Tilapia Oreochromis Niloticus in Bahr Shebin, Nile Tributary, Nile Delta, Egypt. Parasitol. Res. 2008, 103, 1197–1205. [Google Scholar] [CrossRef]
- Florio, D.; Gustinelli, A.; Caffara, M.; Turci, F.; Quaglio, F.; Konecny, R.; Nikowitz, T.; Wathuta, E.M.; Magana, A.; Otachi, E.O.; Matolla, G.K.; Warugu, H.W.; Liti, D.; Mbaluka, R.; Thiga, B.; Munguti, J.; Akoll, P.; Mwanja, W.; Asaminew, K.; Tadesse, Z.; Letizia Fioravanti, M. Veterinary and Public Health Aspects in Tilapia (Oreochromis Niloticus Niloticus) Aquaculture in Kenya, Uganda and Ethiopia. 2009.
- Hoffman, G.L. Myxobolus Cerebralis, a Worldwide Cause of Salmonid Whirling Disease. J. Aquat. Anim. Health 1990, 2, 30–37. [Google Scholar] [CrossRef]
- Henderson, M.; Okamura, B. The Phylogeography of Salmonid Proliferative Kidney Disease in Europe and North America. Proc. Biol. Sci. 2004, 271, 1729–1736. [Google Scholar] [CrossRef]
- Yanuhar, U.; Hardiono, S.A.; Junirahma, N.S.; Caesar, N.R. Profile of Myxobolus Infection in Koi Fish (Cyprinus Carpio) Gill Tissue from Land Pond, Nglegok, Blitar Regency. IOP Conf. Ser. Earth Environ. Sci. 2021, 674, 012016. [Google Scholar] [CrossRef]
- Kaur, H.; Singh, R. Two New Species of Myxobolus (Myxozoa: Myxosporea: Bivalvulida) Infecting an Indian Major Carp and a Cat Fish in Wetlands of Punjab, India. J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2011, 35, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Longshaw, M.; Frear, P.A.; Feist, S.W. Descriptions, Development and Pathogenicity of Myxozoan (Myxozoa: Myxosporea) Parasites of Juvenile Cyprinids (Pisces: Cyprinidae). J. Fish Dis. 2005, 28, 489–508. [Google Scholar] [CrossRef]
- Molnár, K.; Eszterbauer, E.; Marton, S.; Cech, G.; Székely, C. Myxobolus Erythrophthalmi Sp. n. and Myxobolus Shaharomae Sp. n. (Myxozoa: Myxobolidae) from the Internal Organs of Rudd, Scardinius Erythrophthalmus (L.), and Bleak, Alburnus Alburnus (L.). J. Fish Dis. 2009, 32, 219–231. [Google Scholar] [CrossRef]
- Jones, S.R.M.; Bartholomew, J.L.; Zhang, J.Y. Mitigating Myxozoan Disease Impacts on Wild Fish Populations. In Myxozoan Evolution, Ecology and Development; Okamura, B., Gruhl, A., Bartholomew, J.L., Eds.; Springer International Publishing: Cham, 2015. [Google Scholar] [CrossRef]
- Timi, J.T.; Poulin, R. Why Ignoring Parasites in Fish Ecology Is a Mistake. Int. J. Parasitol. 2020, 50, 755–761. [Google Scholar] [CrossRef]
- Boreham, R.E.; Hendrick, S.; O’Donoghue, P.J.; Stenzel, D.J. Incidental Finding of Myxobolus Spores (Protozoa: Myxozoa) in Stool Samples from Patients with Gastrointestinal Symptoms. J. Clin. Microbiol. 1998, 36, 3728–3730. [Google Scholar] [CrossRef]
- Moncada, L.I.; López, M.C.; Murcia, M.I.; Nicholls, S.; León, F.; Guío, O.L.; Corredor, A. Myxobolus Sp., Another Opportunistic Parasite in Immunosuppressed Patients? J. Clin. Microbiol. 2001, 39, 1938–1940. [Google Scholar] [CrossRef]
- Reis, L.L.D.; Jesus, L.C.D.; Fernandes, O.C.C.; Barroso, D.E. First Report of Myxobolus (Cnidaria: Myxozoa) Spores in Human Feces in Brazil. Acta Amaz. 2019, 49, 162–165. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. CONFIDENCE LIMITS ON PHYLOGENIES: AN APPROACH USING THE BOOTSTRAP. Evol. Int. J. Org. Evol. 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Fariya, N.; Abidi, R.; Chauhan, U.K. Description of New Myxozoan Parasite Myxobolus Awadhii Sp. Nov from the Gills of Freshwater Catfish Clarias Batrachus Linn. J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2018, 42, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Fontes, I.; Hallett, S.L.; Mo, T.A. Comparative Epidemiology of Myxozoan Diseases. In Myxozoan Evolution, Ecology and Development; Okamura, B., Gruhl, A., Bartholomew, J.L., Eds.; Springer International Publishing: Cham, 2015. [Google Scholar] [CrossRef]
- Maor-Landaw, K.; Avidor, I.; Rostowsky, N.; Salti, B.; Smirnov, M.; Ofek-Lalzar, M.; Levin, L.; Brekhman, V.; Lotan, T. The Molecular Mechanisms Employed by the Parasite Myxobolus Bejeranoi (Cnidaria: Myxozoa) from Invasion through Sporulation for Successful Proliferation in Its Fish Host. Int. J. Mol. Sci. 2023, 24, 12824. [Google Scholar] [CrossRef]
- de la Hoz Franco, E.; Budy, P. Linking Environmental Heterogeneity to the Distribution and Prevalence of Myxobolus Cerebralis: A Comparison across Sites in a Northern Utah Watershed. Trans. Am. Fish. Soc. 2004, 133, 1176–1189. [Google Scholar] [CrossRef]
- Fonkwa, G.; Nack, J.; K, M.K.; Eyango, M.T.; Tchoumboue, J. Some Epidemiology Aspects of Myxosporean Infections in Oreochromis Niloticus (Linnaeus, 1758) and Hemichromis Fasciatus (Peters, 1857), Two Cultured Cichlid Fishes in the West - Cameroon. Int. J. Aquac. Fish. Sci. 2022, 8, 001. [Google Scholar] [CrossRef]
- Fonkwa, G.; Tchuinkam, T.; Towa, A.N.; Tchoumboue, J. Prévalences Des Myxosporidioses Chez Oreochromis Niloticus Linné, 1758 (Cichlidae) Au Barrage Réservoir de La Mapé (Adamaoua-Cameroun). J. Appl. Biosci. 2018, 123, 12332–12345. [Google Scholar] [CrossRef]
- Rocha, S.; Azevedo, C.; Oliveira, E.; Alves, Â.; Antunes, C.; Rodrigues, P.; Casal, G. Phylogeny and Comprehensive Revision of Mugiliform-Infecting Myxobolids (Myxozoa, Myxobolidae), with the Morphological and Molecular Redescription of the Cryptic Species Myxobolus Exiguus. Parasitology 2019, 146, 479–496. [Google Scholar] [CrossRef]
- Fiala, I.; Bartošová-Sojková, P.; Okamura, B.; Hartikainen, H. Adaptive Radiation and Evolution Within the Myxozoa. In Myxozoan Evolution, Ecology and Development; Okamura, B., Gruhl, A., Bartholomew, J.L., Eds.; Springer International Publishing: Cham, 2015. [Google Scholar] [CrossRef]
- MacKenzie, K.; Kalavati, C. Myxosporean Parasites of Marine Fishes: Their Distribution in the World’s Oceans. Parasitology 2014, 141, 1709–1717. [Google Scholar] [CrossRef]
- Schall, M.K.; Blazer, V.S.; Walsh, H.L.; Smith, G.D.; Wertz, T.; Wagner, T. Spatial and Temporal Variability of Myxozoan Parasite, Myxobolus Inornatus, Prevalence in Young of the Year Smallmouth Bass in the Susquehanna River Basin, Pennsylvania. J. Fish Dis. 2018, 41, 1689–1700. [Google Scholar] [CrossRef]
- Risk Factors Associated with the Presence of Parasitic Diseases in Cultured Tench (Tinca Tinca L.) from the Tormes River (NW Spain). J. Aquac. Mar. Biol. 2015, 2. [CrossRef]
- Salti, B.; Atkinson, S.D.; Brekhman, V.; Smirnov, M.; Lotan, T. Exotic Myxozoan Parasites Establish Complex Life Cycles in Farm Pond Aquaculture. J. Invertebr. Pathol. 2024, 204, 108105. [Google Scholar] [CrossRef] [PubMed]
- Davidovich, N.; Yasur-Landau, D.; Behar, A.; Pretto, T.; Scholz, T. Invasive Parasites and Global Change: Evidence for the Recent and Rapid Spillover of a Potential Pathogen of Tilapias with a Complex, Three-Host Life Cycle. Heliyon 2023, 9, e18831. [Google Scholar] [CrossRef] [PubMed]
- Panova, M.; Hollander, J.; Johannesson, K. Site-Specific Genetic Divergence in Parallel Hybrid Zones Suggests Nonallopatric Evolution of Reproductive Barriers. Mol. Ecol. 2006, 15, 4021–4031. [Google Scholar] [CrossRef]
- McLean, S.-A. Isolation by Distance and the Problem of the Twenty-First Century. Hum. Biol. 2020, 92, 167–179. [Google Scholar] [CrossRef]
- Stadler, P.F.; Geiß, M.; Schaller, D.; López Sánchez, A.; González Laffitte, M.; Valdivia, D.I.; Hellmuth, M.; Hernández Rosales, M. From Pairs of Most Similar Sequences to Phylogenetic Best Matches. Algorithms Mol. Biol. AMB 2020, 15, 5. [Google Scholar] [CrossRef]
- Kelly, D.W.; Muirhead, J.R.; Heath, D.D.; Macisaac, H.J. Contrasting Patterns in Genetic Diversity Following Multiple Invasions of Fresh and Brackish Waters. Mol. Ecol. 2006, 15, 3641–3653. [Google Scholar] [CrossRef]
- Conover, D.O.; Clarke, L.M.; Munch, S.B.; Wagner, G.N. Spatial and Temporal Scales of Adaptive Divergence in Marine Fishes and the Implications for Conservation. J. Fish Biol. 2006, 69, 21–47. [Google Scholar] [CrossRef]
- Brunner, F.S.; Anaya-Rojas, J.M.; Matthews, B.; Eizaguirre, C. Experimental Evidence That Parasites Drive Eco-Evolutionary Feedbacks. Proc. Natl. Acad. Sci. 2017, 114, 3678–3683. [Google Scholar] [CrossRef]
- Florio, D.; Gustinelli, A.; Caffara, M.; Turci, F.; Quaglio, F.; Konecny, R.; Nikowitz, T.; Wathuta, E.M.; Magana, A.; Otachi, E.O.; Matolla, G.K.; Warugu, H.W.; Liti, D.; Mbaluka, R.; Thiga, B.; Munguti, J.; Akoll, P.; Mwanja, W.; Asaminew, K.; Tadesse, Z.; Letizia Fioravanti, M. Veterinary and Public Health Aspects in Tilapia (Oreochromis Niloticus Niloticus) Aquaculture in Kenya, Uganda and Ethiopia. 2009.
- Sugden, R.A.; Smith, T.M.F.; Jones, R.P. Cochran’s Rule for Simple Random Sampling. J. R. Stat. Soc. Ser. B Stat. Methodol. 2000, 62, 787–793. [Google Scholar] [CrossRef]
- Stiassny, M.L.J.; Teugels, G.G.; Hopkins, C.D. Fresh and brackish water fishes of Lower Guinea, West-Central Africa; Institut de recherche pour le développement, 2007.
- Lom, J.; Arthur, J.R. A Guideline for the Preparation of Species Descriptions in Myxosporea. J. Fish Dis. 1989, 12, 151–156. [Google Scholar] [CrossRef]
- Molnár, K.; Eszterbauer, E.; Marton, S.; Cech, G.; Székely, C. Myxobolus Erythrophthalmi Sp. n. and Myxobolus Shaharomae Sp. n. (Myxozoa: Myxobolidae) from the Internal Organs of Rudd, Scardinius Erythrophthalmus (L.), and Bleak, Alburnus Alburnus (L.). J. Fish Dis. 2009, 32, 219–231. [Google Scholar] [CrossRef]
Figure 1.
Cysts (a) of Myxobolus parasites under a stereo microscope (OPTA-TECH, Warsaw, Poland) at 40x total magnification power and spores (b,c) of Myxobolus parasites under a light microscope (OPTA-TECH, Warsaw, Poland) with 1000x total magnification power (Source: author photography during study labwork).
Figure 1.
Cysts (a) of Myxobolus parasites under a stereo microscope (OPTA-TECH, Warsaw, Poland) at 40x total magnification power and spores (b,c) of Myxobolus parasites under a light microscope (OPTA-TECH, Warsaw, Poland) with 1000x total magnification power (Source: author photography during study labwork).

Figure 2.
Gel electrophoresis of amplified partial coding region of the 18S rDNA gene of Myxobolus spp. M is a DNA ladder while lane number 1 and 2 are positive and negative controls respectively, Lanes 3-11 are test samples of freshwater fishes, and lanes 12-13 are test samples of saltwater fishes.
Figure 2.
Gel electrophoresis of amplified partial coding region of the 18S rDNA gene of Myxobolus spp. M is a DNA ladder while lane number 1 and 2 are positive and negative controls respectively, Lanes 3-11 are test samples of freshwater fishes, and lanes 12-13 are test samples of saltwater fishes.
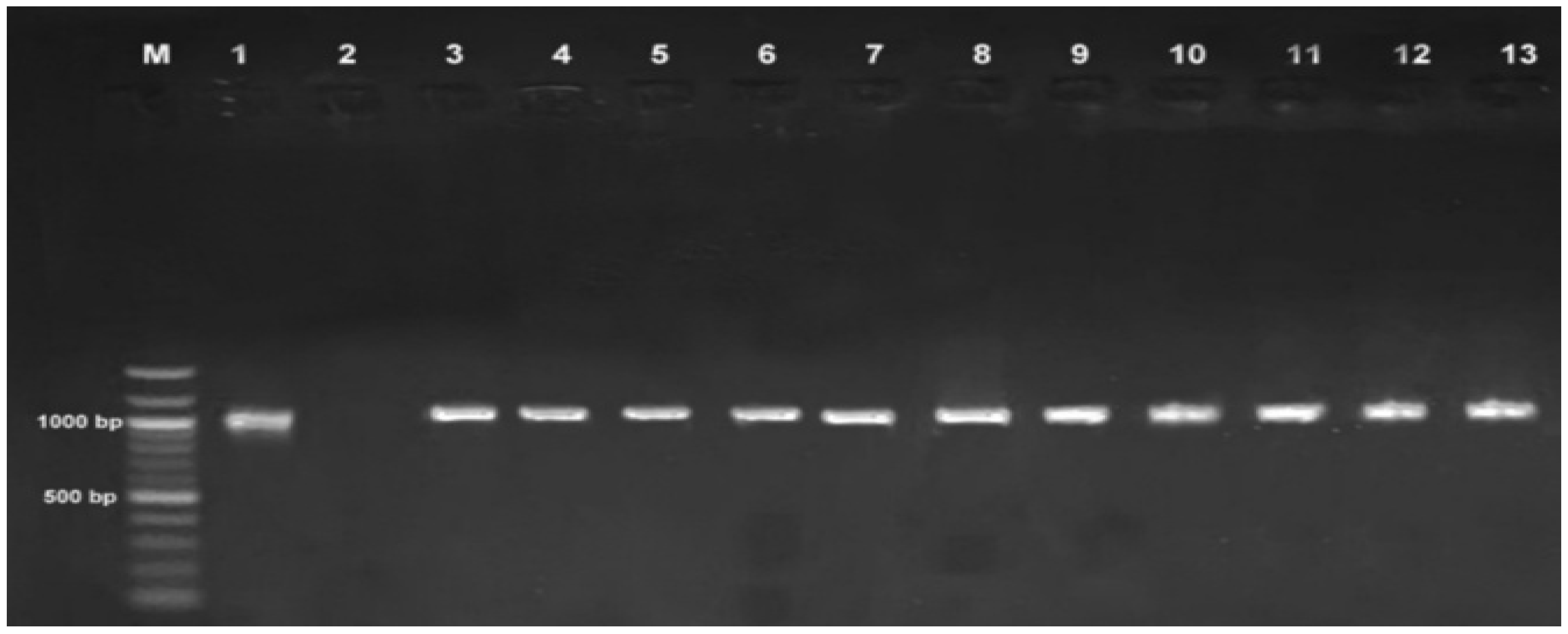
Figure 3.
The evolutionary history was inferred using the Neighbor-Joining method [32]. The optimal tree is shown. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches [33]. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Kimura 2-parameter method [34], and are in the units of the number of base substitutions per site. This analysis involved 23 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated (complete deletion option). There were a total of 187 positions in the final dataset. Evolutionary analyses were conducted in MEGA11 [35]. Candidate study sequences are denoted by red dots, and outgroup sequences are represented by black squares.
Figure 3.
The evolutionary history was inferred using the Neighbor-Joining method [32]. The optimal tree is shown. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches [33]. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Kimura 2-parameter method [34], and are in the units of the number of base substitutions per site. This analysis involved 23 nucleotide sequences. Codon positions included were 1st+2nd+3rd+Noncoding. All positions containing gaps and missing data were eliminated (complete deletion option). There were a total of 187 positions in the final dataset. Evolutionary analyses were conducted in MEGA11 [35]. Candidate study sequences are denoted by red dots, and outgroup sequences are represented by black squares.
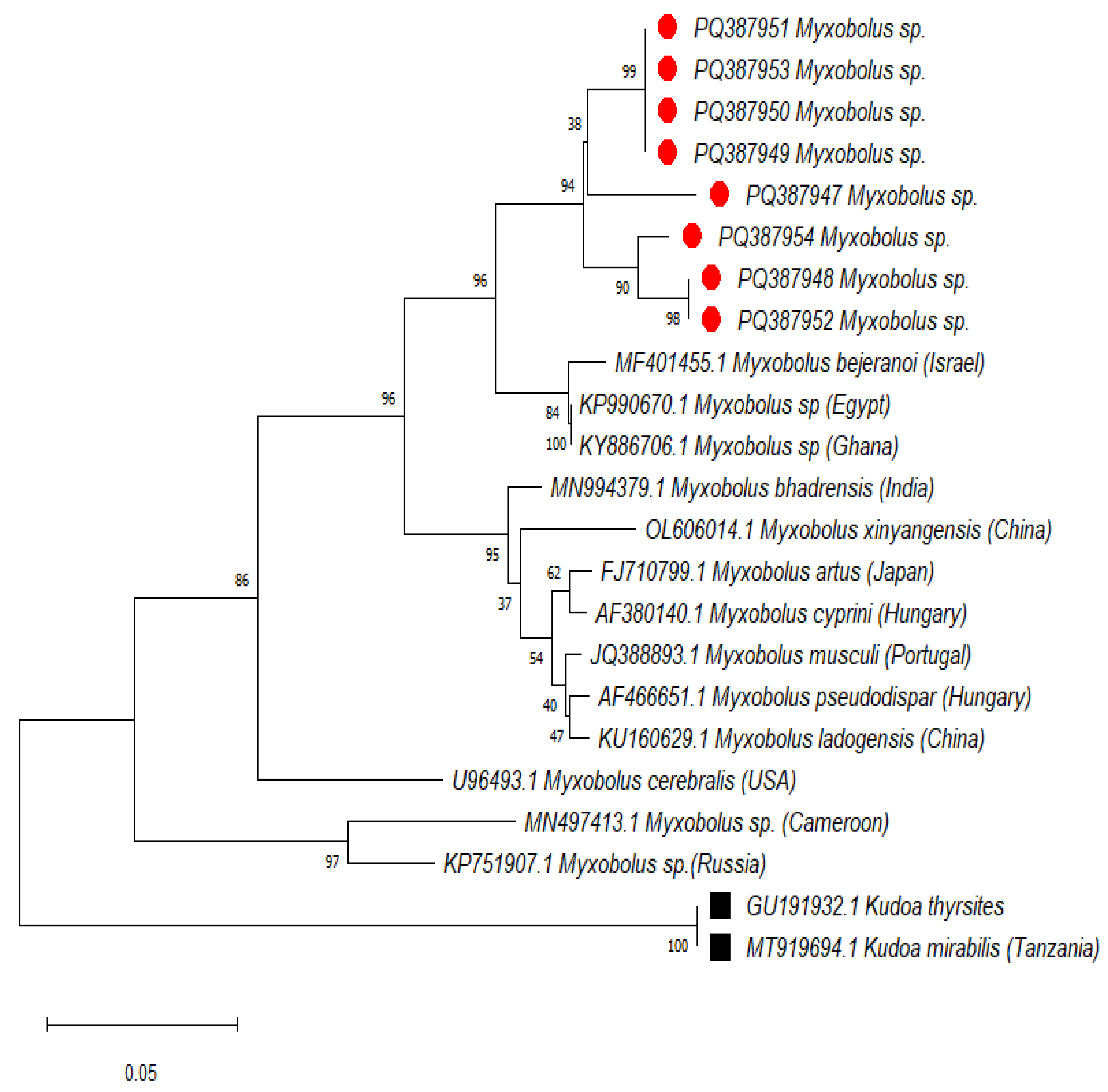
Figure 4.
A map of Mwanza and Dar es Salaam regions depicting selected study sites. The map was created using QGIS software version 3.36.1 and shape files from DIVA-GIS (retrieved on 10 May 2024).
Figure 4.
A map of Mwanza and Dar es Salaam regions depicting selected study sites. The map was created using QGIS software version 3.36.1 and shape files from DIVA-GIS (retrieved on 10 May 2024).
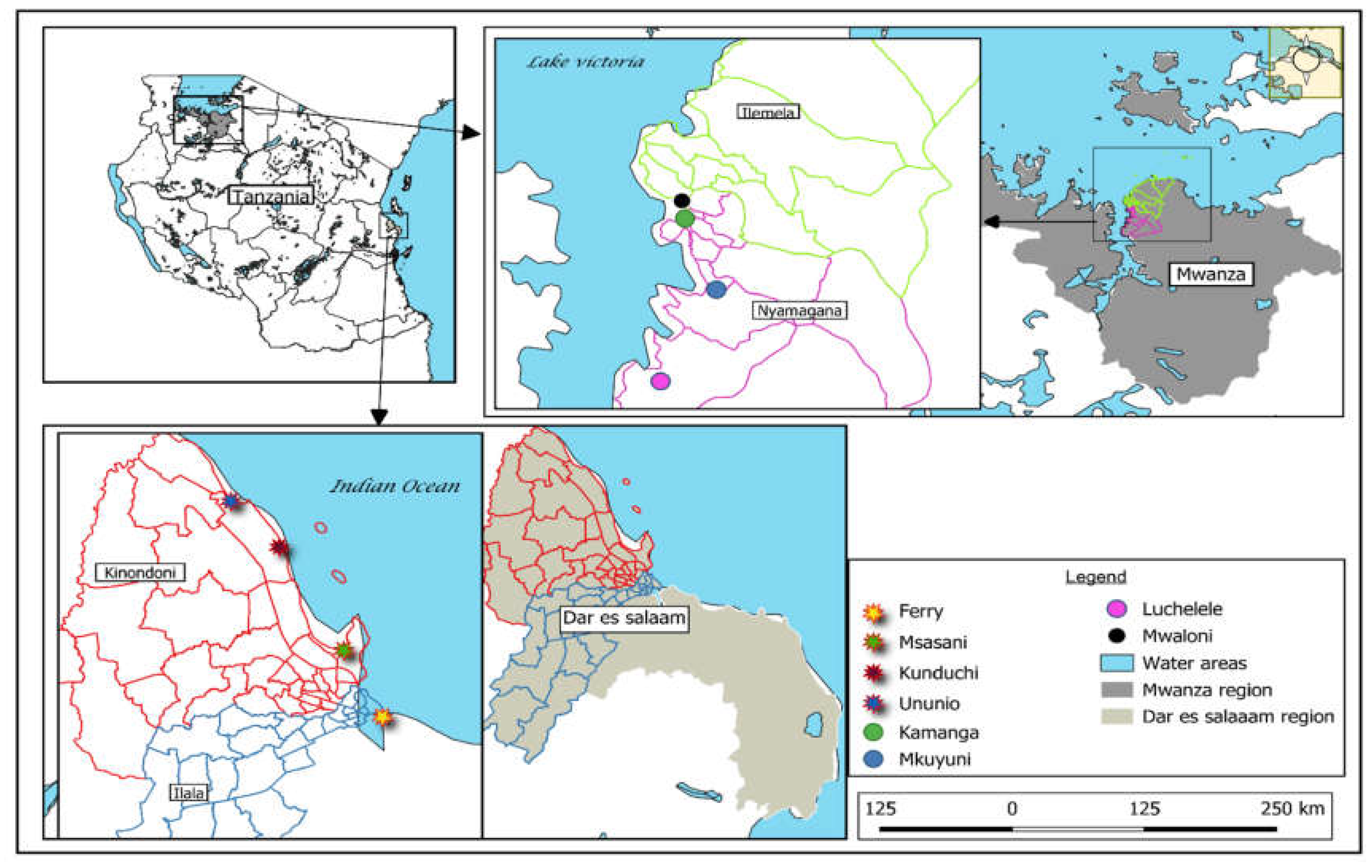

Table 1.
Morphometric and demographic characteristics of fish samples.
| Rastrelliger kanagurta | Lethrinus nebulosus | Caranx sexfasciatus | Lates niloticus | Oreochromis niloticus | Synodontis victoriae | |
|---|---|---|---|---|---|---|
| Total No. of sample | 64 | 64 | 64 | 64 | 64 | 64 |
|
Samples Sex |
Male:40 | Male:37 | Male:29 | Male:42 | Male:36 | Male:35 |
| Female:24 | Female:27 | Female:35 | Female:22 | Female:28 | Female:29 | |
| Location | Ferry:16 | Ferry:16 | Ferry:16 | Mwaloni:16 | Mwaloni:16 | Mwaloni:16 |
| Ununio:16 | Ununio:16 | Ununio:16 | Mkuyuni:16 | Mkuyuni:16 | Mkuyuni:16 | |
| Msasani:16 | Msasani:16 | Msasani:16 | Luchelele:16 | Luchelele:16 | Luchelele:16 | |
| Kunduchi:16 | Kunduchi:16 | Kunduchi:16 | Kamanga:16 | Kamanga:16 | Kamanga:16 | |
| Average length(cm)(SD) | 18.186(2.26) | 18.241(2.99) | 19.16(2.8) | 23.32(1.86) | 20.47(2.59) | 22(1.72) |
| Length Size range(cm) | 14.5-25.1 | 10.1-24.5 | 13.7-25.2 | 19-30 | 14.3-26.5 | 18-26 |
| Mean weight(g)(SD) | 62.69(27.49) | 24.91(5.209) | 96.34(41.66) | 147.311(42.8) | 140.19(54.38) | 94.81(19.19) |
| Weight range(g) | 16-139 | 14-53 | 32-224 | 80-281 | 66-288.5 | 48-126 |
Table 2.
The Chi-square (ꭓ2) association between the prevalence of Myxobolus parasites and variable parameters.
Table 2.
The Chi-square (ꭓ2) association between the prevalence of Myxobolus parasites and variable parameters.
| Variable | Categories (N=Individual Collected) | Infested(n) | Prevalence (n/N*100) |
Odds Ratio | CIa (95%, Lower Limit-Upper Limit) | ꭓ2 | p-Value |
|---|---|---|---|---|---|---|---|
| Fish species | Oreochromis niloticus(N=64) | 20 | 31.25% | 1.23 | 0.00-0.46 | 81.107 | 0.001 |
|
Lates niloticus(N=64) |
22 | 34.38% | |||||
| Caranx sexfasciatus (N=64) | 4 | 6.25% | |||||
| Rastrelliger kanagurta, (N=64) | 0 | 0 | |||||
| Lethrinus nebulosus (N=64) | 0 | 0 | |||||
| Synodontis victoriae(N=64) | 0 | 0 | |||||
| Sex | Female (N=165) | 15 | 9.09% | 0.565 | 0.04-0.19 | 2.484 | 0.115 |
| male(N=219) | 31 | 14.154% | |||||
| Location | Mwanza (N=192) Dar es salaam (N=192) |
42 4 |
21.88% 6.25% |
13.424 | 0.00-0.28 | 35.663 | 0.001 |
| Length (13.7-30cm) | Below Q1(N=94) | 6 | 5.8% | 1.304 | 0.01-0.25 | 7.201 | 0.066 |
| Q1-Q2(N=97) | 13 | 13.30% | |||||
| Q2-Q3(N=90) | 9 | 11% | |||||
| Above Q3(N=103) | 18 | 17.81% | |||||
| Weight(g) | Light(N=227) Medium(N=127) Heavy(N=30) |
21 19 6 |
9.3% 15% 20% |
0.579 | 0.05-0.35 | 4.503 | 0.105 |
Footnote 1: Prevalence is calculated as the number of infected fishes divided by the total number of fishes sampled. Footnote 2: Q1, Q2, and Q3 refer to the first, second, and third quartiles of the length distribution, respectively.
Table 3.
Comparison of 18S rDNA Gene Sequences via BLASTn.
| Query sequence | Sequence from database producing significant alignments with a query sequence | NCBI Accession number | % Similarity | E-Value | Query cover |
|---|---|---|---|---|---|
| S1 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455.1 | 89.62% | 0.0 | 84% |
| S2 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455. | 90.30% | 0.0 | 84% |
| S3 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455. | 90.60% | 0.0 | 79% |
| S4 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455. | 86.58% | 9e-159 | 99% |
| S5 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455. | 90.58% | 0.0 | 83% |
| S6 |
Myxobolus bejeranoi
small subunit
ribosomal RNA gene, partial sequence |
MF401455. | 90.52% | 0.0 | 83% |
| S7 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455. | 90.37% | 0.0 | 81% |
| S8 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455. | 90.43% | 0.0 | 82% |
| S9 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | MF401455. | 89.78% | 0.0 | 79% |
| S10 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | KP990670.1 | 86.88% | 0.0 | 88% |
| S11 | Myxobolus bejeranoi small subunit ribosomal RNA gene, partial sequence | KP990670.1 | 87.44% | 1e-134 | 87% |
Table 4.
Genetic divergence between candidates Myxobolus sp. sequences.
| PQ387950 | PQ387948 | PQ387949 | PQ387947 | PQ387951 | PQ387952 | PQ387953 | PQ387954 | |
|---|---|---|---|---|---|---|---|---|
| PQ387950 | ||||||||
| PQ387948 | 0.071 | |||||||
| PQ387949 | 0.038 | 0.061 | ||||||
| PQ387947 | 0.295 | 0.309 | 0.269 | |||||
| PQ387951 | 0.040 | 0.061 | 0.001 | 0.244 | ||||
| PQ387952 | 0.040 | 0.058 | 0.001 | 0.244 | 0.001 | |||
| PQ387953 | 0.067 | 0.009 | 0.060 | 0.341 | 0.061 | 0.061 | ||
| PQ387954 | 0.040 | 0.060 | 0.003 | 0.250 | 0.001 | 0.001 | 0.062 | |
Footnote 1: Triangular Matrix indicates pairwise genetic distance per estimate.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



