
Submitted:
13 December 2024
Posted:
13 December 2024
You are already at the latest version
Abstract
Spectrum of ixodid ticks that bite people in Western Siberia significantly changed over the past two decades. In this study, we determined tick species attacking people in surroundings of Novosibirsk and the range of bacterial agents they infected with. This study included 301 ticks taken from people and 46% were Ixodes pavlovsky, followed by Ixodes persulcatus (19.6%), I. persulcatus / I. pavlovskyi interspecies hybrids (19.6%), and Dermacentor reticulatus (12.8%). Human DNA was determined in ticks, first demonstrating that all these tick species, including hybrids, were able effectively feed on humans. DNA of Borrelia spp., Rickettsia spp. and Anaplasmatacreae bacteria was detected in different tick species. Borrelia garinii prevailed in Ixodes species, being found in 8.8% ticks, whereas B. afzelii and B. bavariensis were found in single ticks. Borrelia miyamotoi was revealed in 3.7% ticks. Among Rickettsia spp., “Candidatus Rickettsia tarasevichiae” and R. raoultii were identified mainly in I. persulcatus and D reticulatus (44.8% and 26.3%, respectively), whereas Rickettsia helvetica was found only in 2.2% I. pavlovsky. The prevalence of Anaplasma phagocytophilum, Ehrlichia muris and Neoehrlichia mikurensis did not exceed 2%. The obtained results indicate a high risk for humans to be infected with agents of Lyme borreliosis, primarily B. garinii.
Keywords:
Ixodes pavlovsky
; Ixodes persulcatus
; Dermacentor reticulatus
; engorged ticks
; human blood
; Borrelia burgdorferi sensu lato (s.l.)
; B. miyamotoi
; Rickettsia spp.
; Anaplasmataceae
1. Introduction
Several species of ixodid ticks inhabit Novosibirsk province that is located in the southern part of Russian Western Siberia. Among ticks that bite humans, Ixodes persulcatus, Ixodes pavlovskyi and Dermacentor reticulatus are the most common, whereas the number of Dermacentor marginatus and Dermacentor silvarum is low (Livanova et al., 2011; Yakimenko et al., 2019).
Ixodes persulcatus has a wide distribution area in the forest part of Russia, from the North-Western to Far Eastern regions. Until the end of the last century, I. persulcatus was the predominant tick species in all examined locations of the Siberian forest zone. In contrast to I. persulcatus, I. pavlovskyi has a discontinuous distribution and inhabits the Far Eastern and Western Siberian regions (Bolotin et al., 1977; Pomerantsev, 1948; Zamoto-Niikura et al., 2023; Kovalevskiy, et al., 1975). In the last century, I. pavlovskyi was found mainly in mountain regions (Altai and Kuznetsk Alatau Mountains and Salair Ridge) of Siberia (Belyantseva et al., 1974; Sapegina et al., 1969; Kovalevskiy, et al., 1975). In the 21st century, interest in the study of I. pavlovskyi has increased sharply due to the rapid and significant spread of this tick. Nowadays, I. pavlovskyi inhabits not only foothills but also lowland biotopes; in particular, it dominates in areas around large cities, Novosibirsk and Tomsk (Livanova et al., 2011; Rar et al., 2017; Romanenko et al., 2015). Moreover, it was shown the presence of natural interspecies hybrids of I. persulcatus / I. pavlovskyi (hereinafter referred to as hybrids) in all examined locations of their simultaneous inhabitance, namely in the Republic of Altai and Novosibirsk and Tomsk provinces in Siberia as well as in Russky Island in the Far East (Kovalev et al., 2015; Rar 2019, Igolkina 2023a). Both these Ixodes species and their hybrids are infected with the same vector-borne agents, including highly pathogenic tick-borne encephalitis virus, agents of Lyme borreliosis (LB) and Borrelia miyamotoi disease (BMD) as well as several Rickettsia species and bacteria from the Anaplasmataceae family (Cleveland et al., 2022; Eisen 2020; Korenberg et al., 2010; Rar et al., 2017, 2019; Rollins et al., 2023). Notably, the prevalence of some agents varied depending on the tick species. Thus, I. persulcatus was significantly more frequently infected with Borrelia bavariensis and “Candidatus Rickettsia tarasevichiae” and less frequently with Borrelia garinii compared to I. pavlovskyi (Rar et al., 2017)
Unlike Ixodes spp., Dermacentor spp. ticks carry mainly rickettsial pathogens, most often Rickettsia raoultii and less often the highly pathogenic causative agent of Siberian tick typhus (STT), Rickettsia sibirica (Földvári et al., 2016; Kartashov et al., 2019; Rakov et al., 2023; Yakubovskij et al., 2023; Parola et al., 2013). Compared to Ixodes spp., Dermacentor reticulatus and Dermacentor marginatus inhabit drier and warmer areas located in forest-steppe and steppe zones, whereas D. silvarum inhabits forest areas; in lowland locations of Western Siberia the number of D. silvarum is low (Yakimenko et al., 2019)..
Despite the distribution of different ixodidae ticks in Western Siberia was examined in a number of studies (Livanova et al., 2011, 2015; Romanenko et al., 2015, 2017), the epidemiological significance of ticks recently invaded this region has not been sufficiently studied. To clarify this issue, in this study we examined the spectrum of ticks attacking humans in Novosibirsk province (Western Siberia), and their association with various bacterial agents.
2. Materials and Methods
2.1. Sampling
The study included ixodid ticks that were attached to people, as well as those removed from the body and clothing. The ticks were collected in diagnostic laboratories in Novosibirsk (Figure 1). In most cases, any information concerning tick species, developmental stage, degree of engorgement, and the location of sites, where ticks attacked humans, was absent.
Total DNA was extracted from ticks using “Real Best Extraction 100” (“Vector-Best”, Novosibirsk, Russian Federation), according to the protocol, and 50 µl from 400 µl of extracted DNA from each tick were used for this study.
The study was approved by the Local Medical Ethical Committee of the Institute of Chemical Biology and Fundamental Medicine SB RAS, Novosibirsk, Russia (Protocol No. 8, 03.08.2020).
2.2. Tick Species Determination
Tick species determination was conducted based on results of species-specific PCR of the mitochondrial cytochrome c oxidase subunit 1 (cox1) gene and sequencing of the nuclear multicopy internal transcribed spacer (ITS2) fragment. For all samples, cox1 gene fragments were amplified using primers specific to I. persulcatus, I. pavlovskyi and D. reticulatus (Table 1). For subsequent sequencing, ITS2 fragment was amplified in each sample from Ixodes spp. ticks and a number of Dermacentor spp. ticks using primers indicated in Table 1.
2.3. Identification of Engorged Ticks
To identify the engorged ticks and estimate the degree of engorgement, all ticks were examined for the presence of human DNA by RT-PCR with TaqMan probe targeted to the human unique TROSP gene as described previously (Horsman et al., 2006). To standardize this assay, DNA isolated from 100 µl of human blood and serially diluted 10-fold was used as positive control. The results were considered positive if cycle threshold (Ct) was < 40.
2.4. Detection and Genetic Characterization of Bacterial Agents
Identification of bacterial agents in tick specimens was carried out by genus-specific and species-specific PCR using primers specified in Table 1 and/or subsequent sequencing, as previously described (Rar et al., 2017).
Borrelia burgdorferi sensu lato (s.l.) and B. miyamotoi DNA was detected using multiplex PCR targeted to the 5S-23S rRNA intergenic spacer (IGS) of B. burgdorferi s.l. and the p66 gene of B. miyamotoi. For positive B. burgdorferi s.l. specimens, additional PCR assays with primers specific to clpA and p83/100 genes were carried out and the obtained PCR fragments were sequenced (Table 1). To determine B. burgdorferi genospecies, clpA gene sequences were analyzed using PubMLST (https://pubmlst.org/organisms/borrelia-spp) and Blastn (https://blast.ncbi.nlm.nih.gov). To determine the genospecies of samples, which could not be amplified by the clpA gene, the obtained p83/100 gene or IGS sequences were compared with available sequences using Blastn search.
Rickettsia spp. was identified in tick samples using nested PCR with primers targeting the gltA and ompB genes, as described previously (Rar et al., 2017). To determine possible mixed infection, all positive samples were independently amplified using primers RT1 and RT2 specific to “Candidatus R. tarasevichiae”, and primers RH1 and RH3 specific to spotted fever group rickettsiae (SFGR) (Table 1). The species of all SFGR were determined by sequencing of gltA or ompB gene fragments. For R. helvetica-positive samples, the sequences of a long fragment of the ompB gene with a total length of 3117 bp were determined, as previously described (Igolkina et al., 2023b).
Anaplasmataceae bacteria DNA was revealed by nested PCR using primers targeted to 16S rRNA gene (Table 1). For species determination, the obtained PCR fragments were sequenced.
2.5. Sequencing and Phylogenetic Analysis
The obtained amplicons were gel purified in 0.6% SeaKem® GTG-agarose (Lonza, Haifa, Israel). Sanger sequencing was carried out using BigDye Terminator V. 3.1 Cycling Sequencing Kit (Applied Biosystems, CA, USA). Sanger reaction products were analyzed using an ABI 3500 Genetic Analyzer (Applied Biosystems Inc.). Obtained sequences were aligned, analyzed and compared using BioEdit (http://www.mbio.ncsu.edu/BioEdit/bioedit.html). The maximum likelihood (ML) method was used to construct phylogenetic trees using MEGA 7.0 software with 1000 bootstrap replicates (Kumar et al. 2016), based on obtained sequences and known sequences from the GenBank database available on 20 November 2024.
2.6. Statistical Analysis
The Pearson's chi-square test (http://www.socscistatistics.com/tests/chisquare/) was used for statistical analysis. P < 0.05 was considered as significant.
2.7. Nucleotide Sequence Accession Numbers
Nucleotide sequences determined in the study are available in the GenBank database under accession numbers: PQ685972-PQ685977 and PQ682399-PQ682400 for Dermacentor; PQ682653-PQ682655 and PQ724397-PQ724411 for Borrelia spp.; PQ682631-PQ682652 for Rickettsia spp.
3. Results
3.1. Tick Species
A total of 301 specimens of ticks attacking humans in surroundings of Novosibirsk were examined. Tick species were determined based on analysis of the mitochondrial cox1 gene and nuclear ITS2. We failed to determine tick species in five specimens; these specimens were excluded and 296 specimens were investigated. Because of probable presence of I. persulcatus / I. pavlovskyi interspecies hybrids among examined ticks, all Ixodes spp. were genetically characterized by both mitochondrial and nuclear loci, as previously described (Rar 2019). A total of 137 I. pavlovsky, 58 I. persulcatus and 58 hybrids were identified.
Dermacentor reticulatus DNA was found in 38 specimens based on PCR using species-specific primers by the cox1 gene. For 13 specimens, ITS2 or cox1 fragments of D. reticulatus were sequenced to confirm the correctness of D. reticulatus identification using primers designed in this study. All determined ITS2 fragments were identical and had four polymorphic sites in positions, which differentiate D. reticulatus haplotypes previously identified in Eurasia (Figure 2) (Bilbija et al., 2023). The determined cox1 sequences of D. reticulatus were identical to those previously identified in Novosibirsk province (M867332) or differed from this sequence by one unique nucleotide substitution.
Based on sequences of ITS2 fragments, two D. marginatus and three D. nuttalli / D. silvarum were identified among the collected ticks. Two determined sequences of D. marginatus had five and six polymorphic sites; the location of these sites differed in these sequences. The probable haplotypes of D. marginatus corresponded to those of D. marginatus from Turkey (PP456826, etc) (Figure 2). Notably, this is the first determination of ITS2 sequences of D. marginatus collected in Russia.
The sequences of three Dermacentor spp. were closely related to available sequences of D. nuttalli and D. silvarum from Russian Siberia and China. Due to the high genetic similarity between D. nuttalli and D. silvarum, these tick species can be distinguished by morphology and distribution area rather than genetically (Figure 2). Two D. nuttalli / D. silvarum sequences (1069 bp fragments) were identical and showed the most similarity with D. nuttalli from Baikal region (KF241872), differing from it by the presence of four polymorphic sites. However, since the locations where these ticks were taken are known and correspond to the distribution area of D. silvarum, but not D. nuttalli (forest biotopes in the Republic of Altai), these ticks are probably D. silvarum. The sequence of the third tick was the most similar to those of D. nuttalli from Baikal region (KF241869) and China (OQ955291, OQ955293, etc), differing from them by three polymorphic sites (Figure 2). The determined in this study D. nuttalli / D. silvarum sequences differed between themselves by eight mismatches (seven substitutions and one indel) and seven polymorphic sites.
3.2. Determination of Human DNA in Ticks
In order to determine the portion of engorged ticks and estimate the degree of their engorgement, all specimens were tested for the presence of human DNA by Taq-man real-time PCR targeted to the unique human TROSP gene. The quantity of human DNA corresponded to the volume of human blood. Among 296 tested specimens, human DNA was found in 72 (24.3%) ticks. Notably, most of them (55 samples) contained a small amount of human DNA, equivalent to 0.4-3.5 µl of human blood and only 17 ticks contained human DNA in amounts, corresponding to 5.5-106 µl of blood. None of the ticks contained human DNA in quantities equivalent to 3.5–5.5 µl of blood. Ticks may have picked up trace amounts of human material while moving across the skin or during their attachment. Thus, to exclude false-positive results, only ticks with the amount of human DNA equivalent to > 5.5 µl of blood were considered as engorged (Table 2).
The portion of ticks containing a small amount of human DNA (<3.5 µl of blood) varied from 13.8% to 25.9% between different tick species, being the lowest for hybrids and the highest for I. persulcatus (Table 2); the difference was not significant between any of the tick species (p > 0.5). The portion of engorged ticks (> 5.5 µl of blood) also varied depending on tick species and constituted 2.3%, 3.4%, 5.8%, and 10.3% for D. reticulatus, hybrids, I. pavlovsky, and I. persulcatus, respectively. The difference between different tick species was not significant (p > 0.5).
3.3. Detection of Borrelia spp. in Ticks
Spirochetes from B. burgdorferi s.l. species complex and B. miyamotoi were detected in Ixodes spp. but not Dermacentor spp. ticks. In total, B. burgdorferi s.l. was found in 30 (10.1%) ticks, including 19 (13.9%) I. pavlovskyi, 5 (8.6%) I. persulcatus, and 6 (10.3%) hybrids (Table 3). B. garinii prevailed in all Ixodes species, being found in 17 I. pavlovskyi, 4 I. persulcatus, and 5 hybrids. B. afzelii was found only in two I. pavlovskyi, whereas B. bavariensis was detected in one I. persulcatus and a hybrid tick. I. pavlovskyi ticks were significantly more often infected with B. garinii than other B. burgdorferi s.l. genospecies (χ2 = 12.72, df = 1, P < 0.001). The difference in the pathogen prevalence among other tick species or between different tick species for the same pathogen was not significant (p > 0.5).
The identified B. burgdorferi s.l. samples were genetically characterized by the clpA gene. A total of 23 B. garinii, two B. bavariensis and one B. afzelii clpA gene fragments with lengths of 719-785 bp were successfully sequenced. Two B. bavariensis clpA sequences exactly matched to 56 and 72 clpA alleles from the PubMLST database, which were common for Russian Siberia and Asian countries. The only determined clpA sequence of B. afzelii corresponded to allele 36 from the PubMLST database, which also is typical for ticks from Siberia.
Six B. garinii sequences contained polymorphic sites and were excluded from further analysis. Among remaining 17 B. garinii sequences, 12 different sequence variants were identified. The sequences from seven ticks exactly matched clpA alleles from the PubMLST database (192, 195, 196, 211, and 326), which were previously found only in Western Siberia. The sequences from five ticks were identical to three variants of clpA gene sequences, previously identified in ticks from the Novosibirsk province (KX980253, KX980226, and KX980260) but differed from known clpA alleles. The sequence from one I. pavlovskyi (Nov21-186_Ipavl) was novel and differed by one substitution from allele 196, common for Siberia. Two ticks (I. persulcatus and I. pavlovskyi) carried B. garinii variant corresponding to the clpA allele 112, which is widespread in European countries and Western Siberia. In addition, a novel for Siberia B. garinii sequence from I. persulcatus (Nov21-43_Iper) matched to clpA allele 45, which previously was found only in European countries. Another variant unusual for Siberia was detected in one hybrid (Nov21-185_Iper/Ipav); this variant exactly matched the clpA allele 185, which was found in I. ureae collected from seabirds from Canada and Norway, one I. persulcatus in Japan (CP075232) and one I. pavlovskyi in Novosibirsk province (KX980241) (Figure 3).
Borrelia miyamotoi was found in 11 ticks, including five I. pavlovskyi, five I. persulcatus and one hybrid (Table 3). Based on p66 gene sequence analysis, the determined samples of B. miyamotoi belong to the Siberian subtype.
3.4. Detection of Rickettsia spp. in Ticks
Rickettsial DNA was detected in 49/296 (16.6%) ticks: 9/137 (6.6%) I. pavlovskyi, 26/58 (44.8%) I. persulcatus, 3/58 (5.2%) hybrids, 10/38 (26.3%) D. reticulatus, and one D. silvarum (Table 4). Three rickettsial species were identified. “Candidatus R. tarasevichiae” was found in 26/58 (44.8%) I. persulcatus, 1/137 (0.7%) I. pavlovskyi, and 3/58 (5.2%) hybrids. Rickettsia raoultii was detected in 10/38 (26.3%) D. reticulatus, 4/137 (2.9%) I. pavlovskyi, and one D. silvarum, whereas R. helvetica was found in 3/137 (2.2%) I. pavlovskyi. In addition, Rickettsia sp. not belonging to valid species was identified in one I. pavlovskyi.
Positive samples were genotyped by the gltA and ompB genes. The determined “Candidatus R. tarasevichiae” sequences from different tick species were identical and corresponded to known sequences from I. persulcatus (KM288450, OP839041).
The gltA and ompB sequences of R. raoultii isolate from a single positive D. silvarum exactly matched to the sequences of R. raoultii isolate Am-650_Ds (MG545017; MG545018) previously found in D. silvarum from the Russian Far East. The determined R. raoultii sequences from eight D. reticulatus were identical by both examined genes; the gltA gene sequences exactly matched the sequence of the R. raoultii strain Marne from France (RpA4 genotype, DQ355803) and the ompB gene sequences corresponded to the sequence of the R. raoultii strain Khabarovsk from the Russian Far East (DnS14 genotype, DQ365798). For another two R. raoultii isolates from D. reticulatus and four isolates from I. pavlovskyi, only ompB gene sequences were obtained and all these sequences differed from each other. A sequence from I. pavlovskyi was identical to that of Rickettsia raoultii strain Khabarovsk, whereas another five sequences were most similar to the sequence of R. raoultii from Western Siberia (isolate Gorno-Altai-7, PP155665), differing from it by 2-3 substitutions (Figure 4).
For two of three R. helvetica isolates, the fragments of gltA and ompB genes were amplified and sequenced. The obtained sequences of each of the gltA and ompB genes were identical within the gene and corresponded to sequences of R. helvetica from Ixodes ricinus from Netherlands (OY974080) and Ixodes apronophorus from Russian Siberia (OQ866615 and OQ866619) and differed by one substitution in the ompB gene sequence from R. helvetica strain C9P9 from I. ricinus from Switzerland (NZ_CM001467). For the third R. helvetica isolate, only a ompB gene fragment was amplified; the obtained sequence differed from the other two determined sequences by one substitution.
A new Rickettsia sp. (isolate Rickettsia sp. Nov21-51_Ipav) was characterized by the ompB gene and the obtained sequence was most similar (99.2 % similarity) to the corresponding sequence of the R. raoultii isolate Gorno-Altai-7 (PP155665), differing from it by six substitutions.
3.5. Detection of Anaplasmataceae Bacteria in Ticks
Three species from the Anaplasmataceae family were found in the examined ticks; however, their prevalence was rather low. The agent of HGA, Anaplasma phagocytophilum, was found in 3/137 (2.2%) I. pavlovskyi; Ehrlichia muris was detected in 2/137 (1.5%) I. pavlovskyi, and Neoehrlichia mikurensis was identified in 4/137 (2.9%) I. pavlovskyi and 2/58 (3.4%) I. persulcatus. Nor Dermacentor spp., nor hybrids were infected with bacteria from this family.

Table 5.
Prevalence of Anaplasmataceae bacteria in Ixodidae ticks collected from humans.
| Tick species | No. of ticks | No. (%) of ticks containing DNA of tested agents | |||
| Aph | Em | Nm | All Anaplasmataceae | ||
| I. pavlovskyi | 137 | 3 (2.2) | 2 (1.5) | 4 (2.9) | 9 (6.6) |
| I. persulcatus | 58 | 0 | 0 | 2 (3.4) | 2 (3.4) |
| Hybrids | 58 | 0 | 0 | 0 | 0 |
| Dermacentor spp. | 43 | 0 | 0 | 0 | 0 |
| All species | 296 | 3 (1.0) | 2 (0.7) | 6 (2.0) | 11 (3.7) |
Abbreviations: Aph - A. phagocytophilum; Em - E. muris; Nm - N. mikurensis.
4. Discussion
The composition of tick population in the southern regions of Western Siberia significantly changed in recent decades. In some locations, especially in the suburbs of large cities (Novosibirsk and Tomsk), I. pavlovskyi almost completely displaced I. persulcatus (Rar et al., 2017; Romanenko et al., 2015). Moreover, natural I. persulcatus / I. pavlovskyi hybrids were identified throughout the sympatric areas of I. persulcatus and I. pavlovskyi (Kovalev et al., 2015; Rar et al., 2019). The distribution area of D. reticulatus ticks also expanded in last decades and their prevalence near large cities significantly increased (Kartashov et al., 2019; Romanenko et al., 2017). The reasons for such rapid changes in the tick population are not entirely clear, but may be related to climate change or anthropogenic impact. We cannot exclude that hybridization events, including backcrossing of hybrids with parental species, also influenced the ability of I. pavlovskyi to spread rapidly.
However, the comparative epidemiological significance of different Ixodes spp. remains to be established. Unlike previous investigations (Kartashov et al., 2022; Romanenko and Kondratyeva, 2011, we genetically identified not only valid tick species but also I. persulcatus / I. pavlovskyi hybrids in this study. I. pavlovskyi was shown to be the predominant tick species in the surroundings of Novosibirsk, accounting for 46% of ticks attacking humans. The prevalence of other ticks was lower than that amounting to 20% for both I. persulcatus and hybrids and only 13% for D. reticulatus (Table 2). Dermacentor marginatus, D. nuttalli and D. silvarum were found in rare cases. The obtained results are in good agreement with the data of a recent study of Kartashov et al. (2022), in which I. pavlovskyi dominated among ticks attacking humans with a frequency of 43%. Notably, in our previous study a similar prevalence of Ixodes spp. was observed in ticks collected from vegetation; the proportion of I. pavlovskyi, I. persulcatus and hybrids was 50%, 17%, and 26%, respectively (Rar et al., 2019). This close correspondence was unexpected, since questing ticks were collected from vegetation in only five randomly selected locations in the Novosibirsk province, whereas ticks taken from humans could have inhabited anywhere in the region.
The fact that I. pavlovskyi and hybrids readily attack humans does not mean that they are able to feed on humans effectively. To compare the ability of different tick species to feed on humans, we estimated the amount of human blood in the engorged ticks that was received during feeding. Certainly, this is a relative estimation, because the DNA content in human blood varies among individuals. Another limitation is the inability to distinguish whether the small amount of human material was obtained from the skin during movement and attachment or it was obtained during blood feeding. To exclude false positive results, we did not consider ticks with small amounts of human DNA as engorged and set the threshold at a level corresponding to 5.5 µl of blood. As expected, the number of engorged ticks was small, as in most cases people removed the ticks before or immediately after attachment.
Despite all the above limitations, engorged ticks were found among all tick species, with the maximum amount of human blood being approximately 106 µl for I. pavlovskyi, 78 µl for I. persulcatus, 22 µl for hybrids, and only 9 µl for D. reticulatus. The obtained results first demonstrated that I. pavlovskyi and hybrids can effectively feed on humans. However, the prevalence of engorged hybrids (3.4%) and Dermacentor spp. (2.3%) was lower compared to I. pavlovskyi (5.8%) and I. persulcatus (10.3%) (Table 2). Although the observed difference between the species was not statistically significant, it suggests that hybrids are less adapted to feeding on humans compared to I. persulcatus, possibly because they need more time for attachment. The low number of engorged Dermacentor spp. is likely due to the larger size of these ticks, which allows people to notice them.
In Western Siberia, LB and BMD are the most common and severe bacterial tick-borne infections. The main agents of LB in Siberia are B. afzelii, B. bavariensis, and B. garinii; these genospecies were most frequently identified in Ixodes spp. ticks and clinical samples (Rar et al., 2017, Tkachev et al., 2008). Notably, a new species “Candidatus Borrelia sibirica” was recently discovered in Ixodes spp. in the neighboring Omsk province; however, the pathogenic properties of this species are unknown (Sabitova et al., 2023). Previous studies of ticks collected in various regions of Siberia have demonstrated the association of I. persulcatus with B. afzelii and B. bavariensis and I. pavlovskyi with B. garinii (Rar et al., 2017; Sabitova et al., 2023; Mukhacheva and Kovalev, 2014). Unexpectedly, in ticks tested in this study B. garinii almost completely displaced B. afzelii and B. bavariensis and was dominant not only in I. pavlovskyi but also in hybrids and I. persulcatus (Table 3). This discrepancy may be due to the significant dominance of I. pavlovskyi in the tick population and the ability of I. pavlovskyi to transmit B. garinii to other Ixodes species via infected small mammals or by co-feeding.
Borrelia garinii is a genetically variable species associated with terrestrial and marine birds, whereas B. afzelii and B. bavariensis are associated with small mammals. Because of host specificity, B. garinii can be transmitted over long distances, and different B. garinii genovariants do not cluster by geography or tick species (Rollins et al., 2023). Since the analyzed ticks could attack humans anywhere, we expected to find new Borrelia genovariants.
Indeed, we found one novel B. garinii variant and two variants, corresponded to clpA alleles, which were widespread only in I. ricinis distributive area in Europe (allele 45) or in both European countries and Western Siberia (allele 112). Another unusual for Siberia variant exactly matched the clpA allele 185, closely associated with I. ureae and seabirds from Canada and Norway (Margos et al., 2023). Despite the close association with marine birds, several B. garinii isolates, containing the clpA allele 185, were found in single I. persulcatus, I. pavlovskyi, and I. persulcatus / I.pavlovskyi hybrid in Japan and Novosibirsk province (Rar et al., 2017 and this study). These findings clearly demonstrate the adaptation of the specialized B. garinii variant to a broader host range.
In this study, Borrelia miyamotoi, a spirochete of the relapsing fever group, was detected in all Ixodes spp., including hybrids, with a prevalence of 3.7% among all examined ticks and 4.3% among Ixodes spp. (Table 3). The observed prevalence in ticks attacking humans was consistent with the B. miyamotoi prevalence in ticks collected from vegetation in the Novosibirsk province, which ranged from 3.9% to 6.7% for various Ixodes species (Fomenko et al., 2010; Rar et al., 2019). The stable and relatively high prevalence of B. miyamotoi in ticks explains the consistently high incidence of BMD in Novosibirsk province, which is only twice as rare as LB and accounts for 10% of hospitalized patients (Savel’eva et al., 2018).
In Western Siberia, rickettsioses can be caused by several Rickettsia spp.; most cases were caused by R. sibirica, followed by R. raoultii. In rare cases, “Candidatus R. tarasevichiae”, Rickettsia aeschlimannii, and Rickettsia slovaca were recorded as causative agents of infections in Novosibirsk province (Igolkina et al., 2022). For Rickettsia spp., the main route of transmission is transovarial; thus, their association with certain tick species should be more specific compared to B. burgdorferi s.l. The study of ticks taken from humans demonstrated a close association of I. persulcatus with “Candidatus R. tarasevichiae” and D. reticulatus with R. raoultii (Table 3); a similar association was shown for questing ticks collected from various locations in the Western Siberia (Rar et al., 2017, 2019; Yakubovsky et al., 2023). Despite the high prevalence of “Candidatus R. tarasevichiae” and R. raoultii in ticks removed from humans, cases of infections with these pathogens are quite rare, which can be explained by the low pathogenicity of these agents.
Both R. raoultii and R. helvetica are genetically variable species. This study demonstrated higher genetic variability of R. raoultii samples obtained from I. pavlovskyi compared to samples from D. reticulatus. These results correspond to our previous findings that R. raoultii isolates from Ixodes spp. ticks are more variable than isolates from Dermacentor spp. (Rar et al., 2017, 2019; Yakubovsky et al., 2023). Notably, Ixodes spp. ticks infected with R. raoultii were taken from humans, consistent with the high genetic variability of R. raoultii in clinical samples (Igolkina et al., 2022).
It has recently been shown that R. helvetica isolates are reliably subdivided into four genetic lineages (Igolkina et al., 2023b). The European lineage is the most numerous and includes all genotyped R. helvetica isolates from I. ricinus from European countries (Silaghi et al., 2011) and from I. persulcatus from Western Siberia (Kartashov et al., 2022; Igolkina et al., 2023b). In this study, R. helvetica isolates from I. pavlovskyi were first genotyped by a long fragment of the ompB gene, which showed that these isolates also belong to the European lineage. Despite the presence of pathogenic R. helvetica in Ixodes spp. ticks, no cases of R. helvetica infection have been registered in Novosibirsk province. This may be due to the fact that R. helvetica infection has symptoms atypical for rickettsioses (Nilsson et al., 1999, 2010) and was not properly diagnosed.
Rickettsia sibirica, which is the causative agent of widespread STT, was not detected among the examined ticks. The main vectors of R. sibirica are D. nuttalli, D. silvarum, and D. marginatus (Shpynov et al., 2006, 2009), which, as shown in this and other studies (Kartashov et al., 2022), rarely attack humans in the surroundings of Novosibirsk. Due to the high pathogenicity of R. sibirica, even rare cases of human infection with this agent can manifest as severe infection.
Three potentially pathogenic members of Anaplasmataceae family, A. phagocytophilum, E. muris and N. mikurensis, were found in ticks attacking humans; the prevalence of each species was low and did not exceed 2%. There are no confirmed cases of anaplasmosis and ehrlichiosis in humans in Siberia (Tkachev et al., 2008), so the epidemiological significance of the identified Anaplasmataceae bacteria is probably insignificant.
In conclusion, the obtained results indicated that I. pavlovskyi currently has the greatest epidemic significance for residents of Novosibirsk; these ticks attack humans more than 2-3 times as often as I. persulcatus, hybrids, and D. reticulatus and are able effectively feed on humans. It was first shown that I. persulcatus / I. pavlovskyi hybrids can readily attack humans and feed on them effectively. Ticks attacking humans were infected with three genospecies of B. burgdorferi s.l. species complex, B. miyamotoi from the relapsing fever group, three species of Rickettsia, and three species from Anaplasmataceae family. Notably, B. garinii almost completely displaced B. afzelii and B. bavariensis from tick population. The obtained results indicate a high risk of infection humans with causative agents of LB, primarily B. garinii.
Author Contributions
Conceptualization, V.R.; Methodology, V.R. and Y.I.; Formal Analysis, Y.I. and V.F.; Investigation, V.R., G.C., Y.I., V.F. and T.E.; Writing – Original Draft Preparation, V.R.; Writing – Review & Editing, N.T.; Supervision, N.T.; Funding Acquisition, V.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, research project No. 24-24-00390.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Livanova, N.N.; Livanov, S.G.; Panov, V.V. Features of the distribution of ticks Ixodes pavlovskyi on the border of the forest and forest-steppe zone of the Ob region. Parazitologiya 2011, 45, 94–102. (In Russian) [Google Scholar]
- Yakimenko, V.V.; Mal’kova, M.G.; Tul’ko, Z.S.; Tkachev, S.E.; Makenov, M.T.; Vasilenko, A.G. Vector-borne viral infections of Western Siberia (regional aspects of epidemiology, ecology of pathogens and issues of microevolution). Publishing Center KAN: Omsk, 2019; pp. 1-312. (in Russian).
- Pomerantsev, B.I. New ticks of Ixodes family (Ixodidae). Parasit. Sbornik ZIN AS USSR 1948, 9, 36–46. (In Russian) [Google Scholar]
- Kovalevskiy, Yu.V.; Kuksgauzen, N.A.; Zhmaeva, Z.M. Data on the distribution of Ixodes pavlovskyi Pom. in the Altai (Acarina, Ixodidae). Parazitologiia 1975, 9, 518–521. (In Russian) [Google Scholar]
- Bolotin, E.; Kolonin, G.; Kiselev, A.; Matjushina, O. The distribution and ecology of Ixodes pavlovskyi (Ixodidae) on Sykhote-Alin. Parazitologiya 1977, 3, 225–229. (In Russian) [Google Scholar]
- Zamoto-Niikura, A.; Saigo, A.; Sato, M.; Kobayashi, H.; Sasaki, M.; Nakao, M.; Suzuki, T.; Morikawa, S. The presence of Ixodes pavlovskyi and I. pavlovskyi-borne microorganisms in Rishiri Island: an ecological survey. mSphere 2023, 8, e0021323. [Google Scholar] [CrossRef] [PubMed]
- Belyantseva, G.I.; Okulova, N.M. Seasonal changes in the activity of ixodid ticks in the natural focus of tick-borne encephalitis in the south of the Kemerovo region. Med. Parasit Parasit Bolezni. 1974, 43, 710–715. (In Russian) [Google Scholar]
- Sapegina, V.F.; Ravkin, Yu.S. The finding of Ixodes pavlovskyi Pom. in North-East Altai. Parazitologiya 1969, 3, 22–23. (In Russian) [Google Scholar]
- Romanenko, V.; Leonovich, S. Long-term monitoring and population dynamics of ixodid ticks in Tomsk city (Western Siberia). Exp. Appl. Acarol. 2015, 66, 103–108. [Google Scholar] [CrossRef]
- Rar, V.; Livanova, N.; Tkachev, S.; Kaverina, G.; Tikunov, A.; Sabitova, Y.; Igolkina, Y.; Panov, V.; Livanov, S.; Fomenko, N.; et al. Detection and genetic characterization of a wide range of infectious agents in Ixodes pavlovskyi ticks in Western Siberia, Russia. Parasit. Vectors 2017, 10, 258. [Google Scholar] [CrossRef]
- Kovalev, S. Y.; Mikhaylishcheva, M. S.; Mukhacheva, T. A. (2015). Natural hybridization of the ticks Ixodes persulcatus and Ixodes pavlovskyi in their sympatric populations in Western Siberia. Infect. Genet. Evol. 2015, 32, 388–395. [Google Scholar] [CrossRef]
- Rar, V.; Livanova, N.; Sabitova, Y.; Igolkina, Y.; Tkachev, S.; Tikunov, A.; Babkin, I.; Golovljova, I.; Panov, V.; Tikunova, N. Ixodes persulcatus/pavlovskyi natural hybrids in Siberia: Occurrence in sympatric areas and infection by a wide range of tick-transmitted agents. Ticks Tick Borne Dis. 2019, 10, 101254. [Google Scholar] [CrossRef]
- Igolkina, Y.; Nikitin, A.; Verzhutskaya, Y.; Gordeyko, N.; Tikunov, A.; Epikhina, T.; Tikunova, N.; Rar, V. Multilocus genetic analysis indicates taxonomic status of "Candidatus Rickettsia mendelii" as a separate basal group. Ticks Tick Borne Dis. 2023, 14, 102104. [Google Scholar] [CrossRef]
- Korenberg, E.I.; Nefedova, V.V.; Romanenko, V.N.; Gorelova, N.B. The tick Ixodes pavlovskyi as a host of spirochetes pathogenic for humans and its possible role in the epizootiology and epidemiology of borrelioses. Vector Borne Zoonotic Dis. 2010, 10, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L. Vector competence studies with hard ticks and Borrelia burgdorferi sensu lato spirochetes: a review. Ticks Tick Borne Dis. 2020, 11, 101359. [Google Scholar] [CrossRef]
- Cleveland, D.W.; Anderson, C.C.; Brissette, C.A. Borrelia miyamotoi: A Comprehensive Review. Pathogens 2023, 12, 267. [Google Scholar] [CrossRef]
- Rollins, RE.; Sato, K.; Nakao, M.; Tawfeeq, MT.; Herrera-Mesías, F.; Pereira, RJ.; Kovalev, S.; Margos, G.; Fingerle, V.; Kawabata, H.; et al. Out of Asia? Expansion of Eurasian Lyme borreliosis causing genospecies display unique evolutionary trajectories. Mol. Ecol. 2023, 32, 786–799. [Google Scholar] [CrossRef]
- 1Parola, P.; Paddock, C. D.; Socolovschi, C.; Labruna, M. B.; Mediannikov, O.; Kernif, T.; Abdad, M. Y.; Stenos, J.; Bitam, I.; Fournier, P. E.; Raoult, D. Update on tick-borne rickettsioses around the world: a geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed]
- Földvári, G.; Široký, P.; Szekeres, S.; Majoros, G.; Sprong, H. Dermacentor reticulatus: a vector on the rise. Parasit. Vectors 2016, 9, 314. [Google Scholar] [CrossRef] [PubMed]
- Kartashov, M.Yu.; Mikryukova, T.P.; Krivosheina, E.I.; Kuznecov, A.I.; Romanenko, V.N.; Moskvitina, N.S.; Ternovoj, V.A.; Loktev, V.B. Genotyping of tick-borne pathogens in Dermacentor reticulatus ticks collected in urban biotopes of Tomsk. Parazitologiya 2019, 53, 355–369. (In Russian) [Google Scholar] [CrossRef]
- Rakov, A.V.; Chekanova, T.A.; Petremgvdlishvili, K.; Timonin, A.V.; Valdokhina, A.V.; Shirokostup, S.V.; Lukyanenko, N.V.; Akimkin, V.G. High Prevalence of Rickettsia raoultii Found in Dermacentor Ticks Collected in Barnaul.; Altai Krai.; Western Siberia. Pathogens 2023, 12, 914. [Google Scholar] [CrossRef] [PubMed]
- Yakubovskij, V.I.; Igolkina, Y.P.; Tikunov, A.Y. Panov V.V., Yakymenko V.V., Zhabykpayeva A.G., Epikhina T.I., Rar V.A. Genetic Diversity of Rickettsiae in Dermacentor spp. Ticks on the Territory of Western Siberia and Northern Kazakhstan. Mol. Genet. Microbiol. Virol. 2023, 38, 158–167. [Google Scholar] [CrossRef]
- Livanova, N.N.; Tikunov, A.Y.; Kurilshikov, A.M.; Livanov, S.G.; Fomenko, N.V.; Taranenko, D.E.; Kvashnina, A.E.; Tikunova, N.V. Genetic diversity of Ixodes pavlovskyi and I. persulcatus (Acari: Ixodidae) from the sympatric zone in the south of Western Siberia and Kazakhstan. Exp. Appl. Acarol 2015, 67, 441–456. [Google Scholar] [CrossRef]
- Romanenko, V.N.; Sokolenko, V.V.; Maximova, Y.V. Local Formation of High Population Density of the Ticks Dermacentor reticulatus (Parasitiformes, Ixodidae) in Tomsk. Entmol. Rev. 2017, 97, 1391–1396. [Google Scholar] [CrossRef]
- Fomenko, N.V.; Livanova, N.N.; Borgoiakov, V.Iu.; Kozlova, I.V.; Shulaĭkina, I.V.; Pukhovskaia, N.M.; Tokarevich, K.N.; Livanov, S.G.; Doroshchenko, E.K.; Ivanov, L.I. [Detection of Borrelia miyamotoi in ticks Ixodes persulcatus from Russia]. Parazitologiya 2010, 44, 201–211. (In Russian) [Google Scholar]
- Margos, G.; Gatewood, A.G.; Aanensen, D.M.; Hanincová, K.; Terekhova, D.; Vollmer, S.A.; Cornet, M.; Piesman, J.; Donaghy, M.; Bormane, A.; et al. MLST of housekeeping genes captures geographic population structure and suggests a European origin of Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA. 2008, 105, 8730–8735. [Google Scholar] [CrossRef] [PubMed]
- Igolkina, Y.; Rar, V.; Krasnova, E.; Filimonova, E.; Tikunov, A.; Epikhina, T.; Tikunova, N. Occurrence and clinical manifestations of tick-borne rickettsioses in Western Siberia: First Russian cases of Rickettsia aeschlimannii and Rickettsia slovaca infections. Ticks Tick Borne Dis. 2022, 13, 101927. [Google Scholar] [CrossRef] [PubMed]
- Horsman, K.M.; Hickey, J.A.; Cotton, R.W.; Landers, J.P.; Maddox, L.O. Development of a human-specific real-time PCR assay for the simultaneous quantitation of total genomic and male DNA. J. Forensic. Sci. 2006, 51, 758–65. [Google Scholar] [CrossRef]
- Igolkina, Y.; Yakimenko, V.; Tikunov, A.; Epikhina, T.; Tancev, A.; Tikunova, N.; Rar, V. Novel Genetic Lineages of Rickettsia helvetica Associated with Ixodes apronophorus and Ixodes trianguliceps Ticks. Microorganisms 2023, 11, 1215. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33(7), 1870–1874. [Google Scholar] [CrossRef]
- Bilbija, B.; Spitzweg, C.; Papoušek, I.; Fritz, U.; Földvári, G.; Mullett, M.; Ihlow, F.; Sprong, H.; Civáňová Křížová, K.; Anisimov, N.; et al. Dermacentor reticulatus - a tick on its way from glacial refugia to a panmictic Eurasian population. Int. J. Parasitol. 2023, 53, 91–101. [Google Scholar] [CrossRef]
- Kartashov, M.Yu.; Krivosheina, E.I.; Svirin, K.A.; Tupota, N.L.; Ternovoi, V.A.; Loktev, V.B. Genotyping of tick-borne pathogens and determination of human attacking tick species in Novosibirsk and its suburbs. Russian Journal of Infection and Immunity 2022, 12, 1103–1112. [Google Scholar] [CrossRef]
- Romanenko, V.N.; Kondratyeva, L.M. The infection of ixodid ticks collected from humans with the tick-borne encephalitis virus in Tomsk city and its suburbs. Parazitologiya 2011, 45, 3–10. (In Russian) [Google Scholar]
- Tkachev, S.E.; Fomenko, N.V.; Rar, V.A.; Igolkina, Y.P.; Kazakova, Y.V.; Chernousova, N.Y. PCR-detection and molecular-genetic analysis of tick-transmitted pathogens in patients of Novosibirsk region, Russia. International Journal of Medical Microbiology 2008, 298, 365–367. [Google Scholar] [CrossRef]
- Sabitova, Y.; Rar, V.; Tikunov, A.; Yakimenko, V.; Korallo-Vinarskaya, N.; Livanova, N.; Tikunova, N. Detection and genetic characterization of a putative novel Borrelia genospecies in Ixodes apronophorus / Ixodes persulcatus / Ixodes trianguliceps sympatric areas in Western Siberia. Ticks Tick Borne Dis. 2023, 14, 102075. [Google Scholar] [CrossRef] [PubMed]
- Mukhacheva, TA.; Kovalev, SY. Borrelia spirochetes in Russia: Genospecies differentiation by real-time PCR. Ticks Tick Borne Dis. 2014, 5, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Hofmann, M.; Casjens, S.; Dupraz, M.; Heinzinger, S.; Hartberger, C.; Hepner, S.; Schmeusser, M.; Sing, A.; Fingerle, V.; et al. Genome diversity of Borrelia garinii in marine transmission cycles does not match host associations but reflects the strains evolutionary history. Infect. Genet. Evol. 2023, 115, 105502. [Google Scholar] [CrossRef]
- Savel'eva, M.V.; Krasnova, E.I.; Khokhlova, N.I.; Filimonova, E.S.; Provorova, V.V.; Rar, V.A.; Tikunova, N.V. Clinical and laboratory characteristics of diseases caused by Borrelia spp. in the inhabitants of the Novosibirsk region in 2015–2017. Journal Infectology 2018, 10, 68–75. [Google Scholar] [CrossRef]
- Silaghi, C.; Hamel, D.; Thiel, C.; Pfister, K.; Pfeffer, M. Spotted fever group rickettsiae in ticks, Germany. Emerg. Infect. Dis. 2011, 17, 890–892. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.; Lindquist, O.; Pahlson, C. Association of Rickettsia helvetica with chronic perimyocarditis in sudden cardiac death. Lancet 1999, 354, 1169–1173. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.; Elfving, K.; Páhlson, C. Rickettsia helvetica in patient with meningitis, Sweden 2006. Emerg. Infect. Dis. 2010, 16, 490–492. [Google Scholar] [CrossRef]
- Shpynov, S.; Fournier, P. E.; Rudakov, N.; Tarasevich, I.; Raoult, D. Detection of members of the genera Rickettsia, Anaplasma, and Ehrlichia in ticks collected in the Asiatic part of Russia. Ann. N. Y. Acad. Sci. 2006, 1078, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Shpynov, S. N.; Fournier, P. E.; Rudakov, N. V.; Samoilenko, I. E.; Reshetnikova, T. A.; Yastrebov, V. K.; Schaiman, M. S.; Tarasevich, I. V.; Raoult, D. Molecular identification of a collection of spotted Fever group rickettsiae obtained from patients and ticks from Russia. Am..J Trop. Med. Hyg. 2006, 74, 440–443. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The map shows the location of tick collection.
Figure 2.
Phylogenetic trees constructed by the ML method based on nucleotide sequences of 1078 bp fragment of ITS2 of Dermacentor spp. The scale bar indicates an evolutionary distance of 0.001 nucleotide per position in the sequence. Significant bootstrapping values (>70%) are shown on the nodes. Legend: ● – D. nuttalli; ♦ – D. silvarum; ■ – D. marginatus; ▲ – D. reticulatus.
Figure 2.
Phylogenetic trees constructed by the ML method based on nucleotide sequences of 1078 bp fragment of ITS2 of Dermacentor spp. The scale bar indicates an evolutionary distance of 0.001 nucleotide per position in the sequence. Significant bootstrapping values (>70%) are shown on the nodes. Legend: ● – D. nuttalli; ♦ – D. silvarum; ■ – D. marginatus; ▲ – D. reticulatus.

Figure 3.
Phylogenetic trees constructed by the ML method based on nucleotide sequences of 579 bp fragment of clpA of Borrelia spp. The scale bar indicates an evolutionary distance of 0.001 nucleotide per position in the sequence. Significant bootstrapping values (>70%) are shown on the nodes. Legend: ■ – I. pavlovskyi; ▲ – I. persulcatus; ♦ – hybrids.
Figure 3.
Phylogenetic trees constructed by the ML method based on nucleotide sequences of 579 bp fragment of clpA of Borrelia spp. The scale bar indicates an evolutionary distance of 0.001 nucleotide per position in the sequence. Significant bootstrapping values (>70%) are shown on the nodes. Legend: ■ – I. pavlovskyi; ▲ – I. persulcatus; ♦ – hybrids.
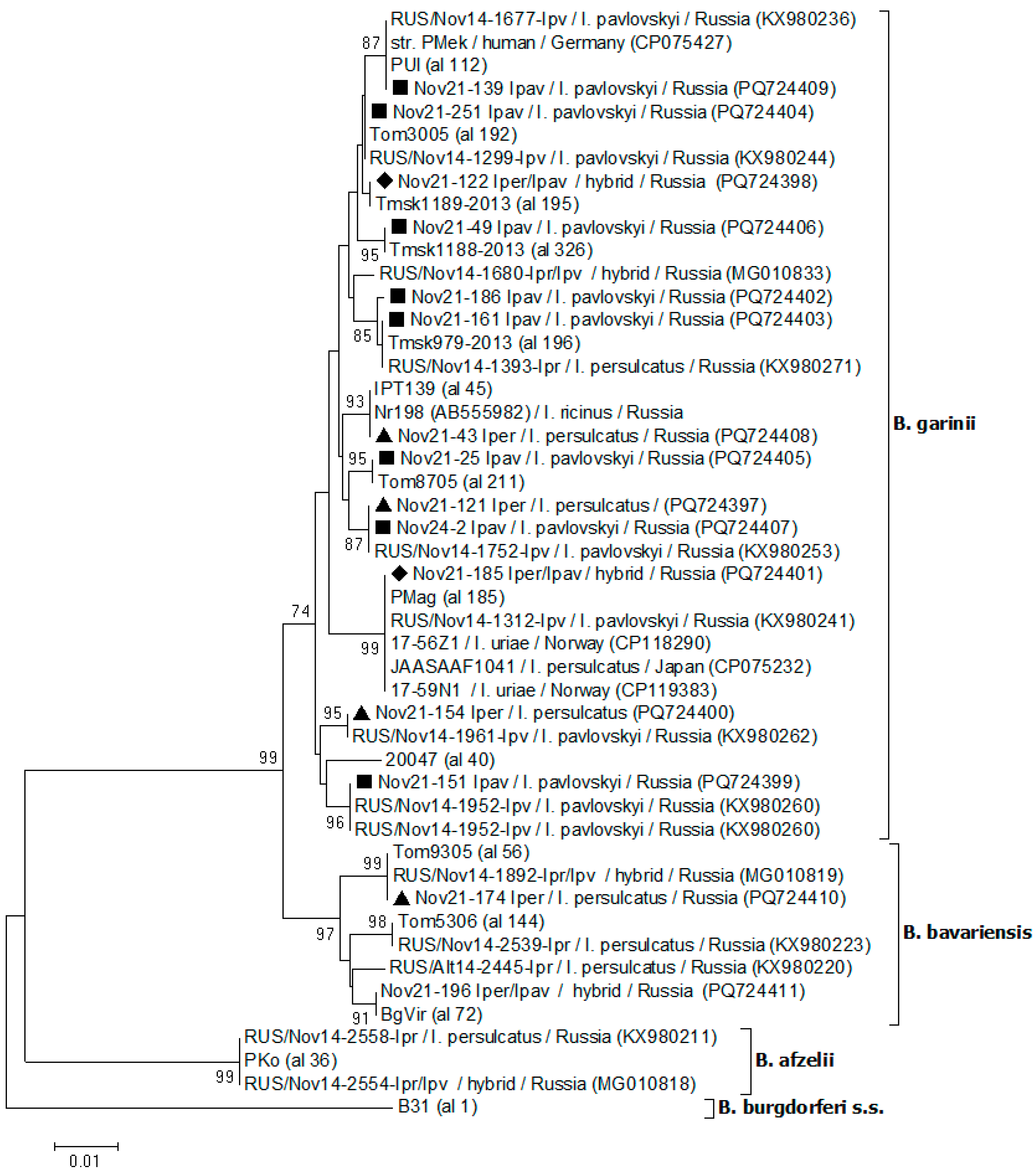
Figure 4.
Phylogenetic trees constructed by the ML method based on nucleotide sequences of A) 716 bp fragment of ompB gene of Rickettsia spp. and B) 3097 bp fragment of ompB gene of Rickettsia helvetica. The scale bar indicates an evolutionary distance of 0.001 nucleotide per position in the sequence. Significant bootstrapping values (>70%) are shown on the nodes. Legend: ○ – I. pavlovskyi; ♦ – D. silvarum; ▲ – D. reticulatus.
Figure 4.
Phylogenetic trees constructed by the ML method based on nucleotide sequences of A) 716 bp fragment of ompB gene of Rickettsia spp. and B) 3097 bp fragment of ompB gene of Rickettsia helvetica. The scale bar indicates an evolutionary distance of 0.001 nucleotide per position in the sequence. Significant bootstrapping values (>70%) are shown on the nodes. Legend: ○ – I. pavlovskyi; ♦ – D. silvarum; ▲ – D. reticulatus.
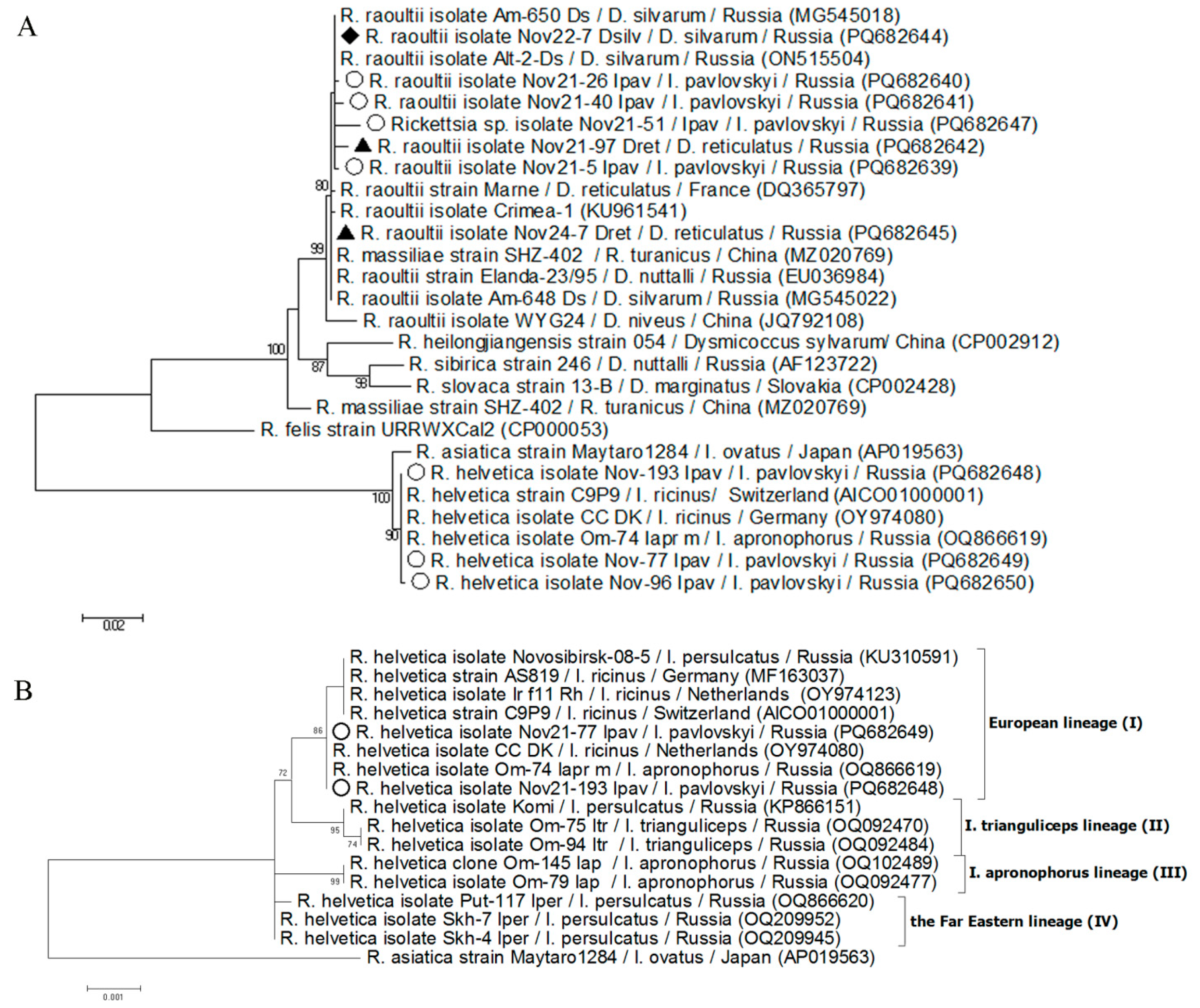
Table 1.
Primers used for identidication of tick species and bacterial agents.
| Locus | Organism | Reaction | Primer name | Primer sequences 5’-3’ | T# (°С) | References |
| ITS2 | Ixodidae | conventional | F-ITS2 | cacactgagcacttactctttg | 57 | [23] |
| R1-ITS2 | actggatggctccagtattc | |||||
| cox1 | I. persulcatus | conventional | Ixodes-F | acctgatatagctttccctcg | 55 | [10] |
| Ipers-R | ttgattcctgttggaacagc | |||||
| I. pavlovskyi | conventional | Ixodes-F | acctgatatagctttccctcg | 55 | [10] | |
| Ipav-R | taatccccgtggggacg | |||||
| Ixodidae | conventional | C1 | accacaaagacattggaactatatat | 50 | [23] | |
| C2 | aatccaggaagaataagaatatatac | |||||
| D. reticulatus | conventional | Dret -F | ctaagacaacccggaacattaattg | 60 | This study | |
| Dret-R | aaaccctaaaagaccaattgcggc | |||||
| IGS | B. burgdorferi s.l. | Primary | NC1 | cctgttatcattccgaacacag | 50 | [10] |
| NC2 | tactccattcggtaatcttggg | |||||
| Nested | NC3 | tactgcgagttcgcgggag | 50 | |||
| NC4 | cctaggcattcaccatagac | |||||
| p66 | B. miyamotoi | Primary | M3 | ttctatatttggacacatgtc | 50 | [25] |
| M4 | cagattgtttagttctaatccg | |||||
| Nested | M1 | ctaaattattaaatccaaaatcg | 50 | |||
| M2 | ggaaatgagtacctacatatg | |||||
| clpA | B. burgdorferi s.l. | Primary | clpAF1237 | aaagatagatttcttccagac | 50 | [26] |
| clpAR2218 | gaatttcatctattaaaagctttc | |||||
| Nested | clpAF1255 | gacaaagcttttgatattttag | 50 | |||
| clpAR2104 | caaaaaaaacatcaaattttctatctc | |||||
| р83/100 | B. burgdorferi s.l. | Primary | F7 | ttcaaagggatactgttagagag | 50 | [10] |
| F10 | aagaaggcttatctaatggtgatg | |||||
| Nested | F5 | acctggtgatgtaagttctcc | 54 | |||
| F12 | ctaacctcattgttgttagactt | |||||
| gltA | Rickettsia spp. | Primary | glt1 | gattgctttacttacgaccc | 52 | [10] |
| glt2 | tgcatttctttccattgtgc | |||||
| Nested | glt3 | tatagacggtgataaaggaatc | 53 | |||
| glt4 | cagaactaccgatttctttaagc | |||||
| Ca. R. tarasevichiae | Nested | RT1 | tactaaaaaagtcgctgttcattc | 56 | [10] | |
| RT2 | tgttgcaaacatcatgcgtaa | |||||
| SFGR | Nested | RH1 | gtcagtctactatcacctatatag | 54 | [10] | |
| RH3 | taaaatattcatctttaagagcga | |||||
| ompB | Rickettsia spp. | Primary | B1 | atatgcaggtatcggtact | 56 | [27] |
| B2 | ccatataccgtaagctacat | |||||
| Nested | B3 | gcaggtatcggtactataaac | 56 | |||
| B4 | aatttacgaaacgattacttccgg | |||||
| 16S rRNA | Anaplasmataceae | Primary | Ehr1 | gaacgaacgctggcggcaagc | 57 | [10] |
| Ehr2 | agtaycgraccagatagccgc | |||||
| Nested | Ehr3 | tgcataggaatctacctagtag | 60 | |||
| Ehr4 | ctaggaattccgctatcctct |
T* - annealing temperature.
Table 2.
Human DNA in different tick species determined by RT-PCR.
| Amount (µl) of human blood in a tick | The No (%) of ticks containing different amount of human blood | ||||
| I. pavlovskyi (n=137) | I. persulcatus (n=58) | Hybrids (n=58) | Dermacentor spp. (n=43) | All species (n=296) | |
| Nd (<0.4) | 106 (77.4) | 37 (63.8) | 48 (82.8) | 33 (76.7) | 224 (75.7) |
| 0.4-0.9 | 12 | 7 | 5 | 8 | 32 |
| 1.0-1.9 | 8 | 5 | 2 | 0 | 15 |
| 2.0-2.9 | 1 | 2 | 0 | 1 | 4 |
| 3.0-3.5 | 2 | 1 | 1 | 0 | 4 |
| 3.5-5.5 | 0 | 0 | 0 | 0 | 0 |
| Subtotal 0.4-5.5 | 23 (16.8) | 15 (25.9) | 8 (13.8) | 9 (20.9) | 55 (18.6) |
| 5.5-10 | 3 | 1 | 1 | 1 | 6 |
| 10-20 | 2 | 1 | 0 | 0 | 3 |
| 20-50 | 1 | 2 | 1 | 0 | 4 |
| 50-106 | 2 | 2 | 0 | 0 | 4 |
| Subtotal5.5-106 | 8 (5.8) | 6 (10.3) | 2 (3.4) | 1 (2.3) | 17 (5.7) |
Table 3.
Prevalence of Borrelia spp. in Ixodidae ticks collected from humans.
| Tick species | No. of ticks | No. (%) of ticks containing DNA of tested agents* | ||||
| Bg | Ba | Bb | all B. burgdorferi (s.l.) | Bm | ||
| I. pavlovskyi | 137 | 17 (12.4) | 2 (1.5) | 0 | 19 (13.9) | 5 (3.6) |
| I. persulcatus | 58 | 4 (6.9) | 0 | 1 (1.7) | 5 (8.6) | 5 (8.6) |
| Hybrids | 58 | 5 (8.6) | 0 | 1 (1.7) | 6 (10.3) | 1 (1.7) |
| Dermacentor spp. | 43 | 0 | 0 | 0 | 0 | 0 |
| All species | 296 | 26 (8.8) | 2 (0.7) | 2 (0.7) | 30 (10.1) | 11 (3.7) |
Abbreviations: Bg - B. garinii; Ba - B. afzelii; Bb - B. bavariensis; Bm – B. miyamotoi. *Including cases of mixed infection.
Table 4.
Prevalence of Rickettsia spp. in Ixodidae ticks collected from humans.
| Tick species | No. of ticks | No. (%) of ticks containing DNA of tested agents | ||||
| Rt | Rr | Rh | Rsp | All Rickettsia spp. | ||
| I. pavlovskyi | 137 | 1 (0.7) | 4 (2.9) | 3 (2.2) | 1 (0.7) | 9 (6.6) |
| I. persulcatus | 58 | 26 (44.8) | 0 | 0 | 0 | 26 (44.8) |
| Hybrids | 58 | 3 (5.2) | 0 | 0 | 0 | 3 (5.2) |
| D. reticulatus. | 38 | 0 | 10 (26.3) | 0 | 0 | 10 (26.3) |
| D. nuttalli / D. silvarum | 3 | 0 | 1 | 0 | 0 | 1 |
| D. marginatus | 2 | 0 | 0 | 0 | 0 | 0 |
| All species | 296 | 30 (10.1) | 15 (5.1) | 3 (1.0) | 1 (0.3) | 49 (16.6) |
Abbreviations: Rt - “Candidatus R. tarasevichiae”; Rr - R. raoultii; Rh – R. helvetica; Rsp – Rickettsia sp.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



