
Submitted:
05 December 2024
Posted:
06 December 2024
You are already at the latest version
Abstract
The Central European fauna, in the last decades, has been undergoing a strong transformation due to four main factors: the retreat of species that require cool and wet habitats, the proliferation of species that thrive in warm and dry conditions, the northward migration of Mediterranean species, and the gradual establishment of species with tropical origins. In this study, we detail the changes in the orders Hymenoptera, Diptera, and Lepidoptera, and analyze the establishment of non native insects and the northward migration of Mediterranean species. The transformation towards a Mediterranean-type fauna is prominently indicated by the population increase of xerothermic Aculeata, bee flies (Bombyliidae), and horse flies (Tabanidae). Additionally, groups that require cool and wet ecological conditions, particularly hoverflies (Syrphidae), but also sawflies (Tenthredinidae) and tachinid flies (Tachinidae), have shown notable decline. In nocturnal moths, we observe a decrease in species richness in certain areas, as well as frequent outbreaks in populations of some less climate-sensitive species. Some species of butterflies are less sensitive to the current extent of climate change, exhibiting significant population growth under protected conditions. However, most of the previously sporadic and rare butterfly species have proven to be climate-sensitive, unable to achieve significant population growth even under strict nature conservation. In recent decades, the influx from Mediterranean regions and the establishment of tropical non native insect species have turned into exponential rate. We have reviewed the presence of alien species, recording 803 alien insect species in our region, 298 of them have arrived in the past quarter-century, with a significant proportion (54%) originating from tropical and Mediterranean regions.
Keywords:
climate change
; insects
; alien species
; Hymenoptera
; Diptera
; Lepidoptera
; Mediterranean influx
; Central Europe
1. Introduction
"If you put a frog in a pot and slowly turn up the heat, it won’t jump out. Instead, it will enjoy a nice warm bath until it is cooked to death. We humans seem to be doing pretty much the same thing."
Jeff Goodell (The Water Will Come, 2010)
Jeff Goodell's renowned quote aptly illustrates the gradual yet destructive process of climate change. Global warming, a critical aspect of climate change, profoundly impacts biodiversity and ecosystems worldwide. Insects, which play crucial roles in various ecological processes such as pollination, decomposition, and serving as food sources for other animals, are particularly affected. This paper focuses on the changes in the fauna of three major insect orders: Lepidoptera, Hymenoptera, and Diptera in Central Europe, driven by the ongoing shifts in climate patterns, additionally we discuss all groups of the alien, introduced insects (Figure 1) and the Mediterranean influx.
Previous studies have demonstrated that climate change can lead to shifts in species richness and population densities of insects. For instance, research on European butterflies has documented northward range shifts and changes in abundance patterns. Parmesan and his colleagues [1] found that 63% of 35 non-migratory European butterfly species had shifted their ranges northward by 35-240 km over the 20th century. This shift is attributed to rising temperatures, which create more favorable conditions in previously cooler regions. Similarly, the phenology of many insect species is being altered by climate change. For example, the flight periods of butterflies and moths are occurring earlier in the year. Roy and Sparks [2] observed that the first appearance dates of 35 British butterfly species advanced by an average of 2.3 days per decade from 1976 to 1998. These phenological changes can lead to mismatches between insects and their host plants or prey, potentially affecting their survival and reproduction.
Diptera, including many species of flies, are also experiencing changes in their populations due to climate change. For example, the distribution of the common housefly (Musca domestica Linnaeus, 1758) has been influenced by temperature changes, with populations increasing in regions that have become warmer. Additionally, climate change can affect the prevalence of disease vectors such as mosquitoes [3,4].. Researchers suggested that rising temperatures and altered precipitation patterns could expand the range of mosquitoes that transmit diseases like malaria and dengue fever. In Hymenoptera, a similar phenomenon may be observed. Ornithologists have noted that European bee-eaters (Merops apiaster Linnaeus, 1758) are expanding northward following their prey animals [5]. Central Europe, with its diverse habitats ranging from lowland forests to alpine meadows, provides a unique setting to study these changes. The region has already experienced noticeable shifts in climate, with increasing average temperatures and altered precipitation patterns. These climatic changes are expected to continue, posing challenges for the conservation of insect biodiversity.
The present work is a continuation of our monograph titled "Changes in Population Densities and Species Richness of Pollinators in the Carpathian Basin"[6]. In this study, we concluded that although a pollination crisis is moderately present in our region, particularly for certain groups, the more concerning issue is the gradual Mediterranean-type transformation of the fauna. In this article, we examine this transformation in detail. We will discuss the adaptation of individual species and their established ecotypes, examining how each ecotype responds to changing environmental conditions. We also explore the transformation of fauna in terms of agricultural plant protection, forest protection, and nature conservation.
The study area is Central Europe, with the Carpathian Basin at its heart (Figure 2). Of the 11 biogeographic regions in Europe, three are present here: the Pannonian, Continental, and Alpine regions [7].
Global warming is merely one aspect of the broader set of contemporary issues termed "polycrisis." This includes climate change, habitat destruction, loss of natural vegetation and fauna, overpopulation, degradation of agricultural land leading to food crises, dwindling potable water supplies triggering global migration, energy crises, and the depletion of raw materials critical to the economy. Collectively, these issues herald a global collapse process, threatening economic, commercial, political, social, and ultimately cultural stability.
Fauna is fundamentally determined by vegetation; hence, changes in fauna are driven by changes in vegetation through ecological networks. The interaction between climate change and vegetation is complex. Climate change and warming favor the expansion of the steppe zone while adversely affecting montane beech and other woodlands. Concurrently, afforestation mitigates the impact of climate change. These processes have been studied extensively in our region [8,9]: researchers mapped the changes in vegetation under climate change, and by using REMO model, they concluded that forest restoration could reduce surface temperatures by up to 0.7 °C and may increase precipitation by more than 10%.
Key aspects of faunal change include the decline of species adapted to cool climates, the expansion of species adapted to warm, dry, or occasionally warm-humid climates, the northward spread of Mediterranean species, and the promotion of invasive species. Insects have short generation times and high reproductive rates, making them more likely to respond rapidly to climate change compared to plants and vertebrates. Possible responses include changes in phenological patterns, habitat selection, and the expansion and/or contraction of geographic ranges. These rapid responses can trigger processes with potentially devastating consequences for crop production and food security [10]. . Karuppaiah and Sujayanad classified these responses into changes in insect population dynamics, migration, development and reproductive biology (phenological changes, nutrient cycling, etc.), diapause, stage survival, growth rate, and species voltinism [11].
The indirect effects of climate change are examined in a UFZ study [12], including the re-emergence of mosquito-borne diseases like dengue fever or malaria, the occurrence of multiple generations of pests due to extended reproductive periods, increasing damage, and the likelihood of forest fires in boreal areas such as Canada. The increased prevalence of certain insect species can also serve as an indicator of climate change [13].
The temporal movements of different insect groups at varying altitudes exhibit diverse patterns depending on climate change and the ecological needs of specific groups [14]. In the present work, we will also focus on these patterns, using the Carpathian Basin as a representative sample area for Central Europe.
1.2. Climate Change in Central Europe
The climate of Central and Eastern Europe is characterized by a general rise in temperature (Figure 3). Temperatures have risen significantly during this period, by approximately 1.2°C. Seasonal temperature increases are noted as follows: 1.1°C in spring, 1.3°C in summer, 1.2°C in autumn, and 1.3°C in winter. However, the frequency of cold days and nights has decreased over the past 40 years. There is no overall trend in precipitation across the region, but regional specificities must be considered. For instance, precipitation increased by about 9% in the northwest, while it decreased similarly in the southeast. In Hungary, precipitation has generally decreased by 11% since 1901, particularly since the 1970s [15]. Romania, due to its Balkan territory, and Hungary, due to its low-lying position in the Carpathian Basin, are particularly affected by a negative water balance. The Tisza, the region's second-largest river, has been strictly regulated since 1846. Consequently, the river acts as a drainage channel for the vast volumes of water from the surrounding mountains, further drying out the Carpathian Basin. In 2019, the volume of water flowing into the lower areas of the Carpathian Basin was 112 km³, while the volume flowing out was 117 km³. Global warming is disrupting this balance, leading to increased droughts in recent years [16,17]. In Central and Eastern Europe, the mean annual temperature is projected to increase between 1°C and 3°C by mid-century and up to 5°C by the end of the century [15]. However, one-way projections do not account for the phenomena of action-reaction and climatic feedback.
Our paper synthesizes current research on the impacts of global warming on Lepidoptera, Hymenoptera, and Diptera in Central Europe. By examining changes in distribution, phenology, and population dynamics, we aim to understand the broader ecological consequences and inform conservation strategies. Particular attention is given to the influx of species from the south, as well as the introduction and establishment of alien species, often referred to as insect globalization, within the context of the current warming climate. The findings underscore the urgency of addressing climate change to preserve the intricate web of life that sustains our ecosystems.
2. Materials and Methods
2.1. Hymenoptera, Diptera
Our research is based on 3 to 5 decades of systematic collection results from the 1970s and 1980s onwards. The collecting area covers the Carpathian Basin (Pannonian Basin) region, the core region of Central Europe. Approximately 4,000 collection points were utilized, with 30-35 collecting days per year, depending on the weather conditions. The collected material is partly stored in the Natural History Museum in Zirc, the insect collection of the Rippl-Ronai Museum in Kaposvár, and the Hungarian Natural History Museum. The collection method used was grass netting. For the analysis of individual trends, changes in the number and species richness of the groups defined by each ecotype were examined. The ecotypes of Aculeate were defined as euryecious intermediate, euryecious eremophilic, euryecious hylophilic, stenoeceous eremophilic, and stenoeceous hylophilic following Pittioni [19,20]. The ecotypes of Diptera were defined following Hagen [21]. Bumblebees were categorized according to their climatic sensitivity based on the monograph by Rasmont and colleagues ("Climatic Risk and Distribution Atlas of European Bumblebees") [22] and compared with our own recording experience. We provided a list of species that have not been collected in recent centuries, i.e., those whose numbers have fallen below the detection level. These species were studied in parallel in the lower parts of the Carpathian Basin and at high altitudes of 1200-1800 m above sea level (Tatras, Carpathians, Slovakia, Romania) to determine whether high altitudes can provide suitable habitats for their survival [23,24,25,26,27,28,29,30,31,32,33]. While the moth data are unpublished, Hymenoptera and Diptera data are published in part [34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57].
2.2. Nocturnal Macrolepidoptera
The species richness and number of nocturnal moths were analyzed using light trap material from faunistic research conducted in the Carpathian Basin. Notably, this includes material from the national forest light trap network in Hungary. Light trap material was collected from 41 sites across the Carpathian Basin and identified by experts such as Csaba Szabóky, Csaba Gáspár, Lajos Kovács, Katalin Leskó, Levente Ábrahám, Ákos Uherkovich, Zoltán Varga, and Péter Schmidt, among others. A single trapping point was consistently included over several decades, and changes in light sources were also considered.
Methodological changes were necessary for nocturnal butterfly studies. Since 2014, UV LED light traps have been used, replacing the black light UV 20W (used between 1990 and 2010) and the Jermy-type light trap with a 125W mercury vapor lamp (used till 1990 circa). These different light sources had varying selectivities [58,59,60]. Consequently, we calculated a long-term trend from 1970 and a separate trend from 2014 to draw reliable conclusions. Additionally, an apparent trend reversal in the 1980s was observed and calculated separately for each species as a control. Specific analysis of Hymnoptera, Lepidoptera and alien insects are in Annexes A, B and C.
From the light trap data, we selected only those series that covered six months of intensive daily collection, typically from April to October, and in some cases from May to November. The traps operated continuously during these periods. Other shorter or incomplete trap data were excluded from the analysis. The number of sampling points was 24, and several locations (such as the Drava Plain or the Zselic Mountains) were resampled after 40 years. To differentiate surveys from various locations in the same year, letters of the alphabet were used as follows: Felsötárkány: 1970a, 1971a, 2020a, 2021b, 2022c Tompa: 1970b, 2020b, 2021c, 2022b Sopron: 1970c, 202c, 2021d, Gilvánfa: 1970d, 1973b, Sellye: 1971c, Magyarszombatfa: 1975, 1976a, Kisvaszar: 1973a, Mike: 1976b, Vásárosbéc: 1977, 1978a, Bőszénfa: 1978b, 1979c, 1980c, Almamellék: 1979a, 1980a, Palé: 1979b, 1980b, 1981b, Lipótfa 1986, 1987, Jósvafő: 1990, 1998, 2005, Aggtelek: 2000, 2001, Répáshuta: 2015, 2016, 2017, 2018, 2019a, Sellye: 2019b, 2020d, 2021a, 2022a, Ropoly: 2019c, 2020e (Annex B).
The ecotypes of moths were discussed partially following the work of Sage and Utschick [61], where the authors ranked different habitats on a scale from 0 to 10. Additionally, the classification by Kanarskyi, Y. et al. [62] was employed, assigning ecotypes as follows: U (ubiquists), M1 (grassland mesophiles), M2 (seminemoral mesophiles), M3 (nemoral mesophiles), H2 (nemoral hygrophiles), X1 (grassland xerothermophiles), and X2 (seminemoral xerothermophiles).
2.3. Butterflies, Rhopalocera
The two primary sources of data on butterflies are the butterfly collection of the Rippl-Rónai Museum in Kaposvár and the Danube-Ipoly National Park. The museum's collection consists of approximately 50,000 specimens from nearly 300 sites across the Carpathian Basin, regularly collected with 30-35 field days annually. However, as the collection is selective, it does not provide reliable data on common species. Levente Ábrahám systematically collected specimens of 33 species over 30 years, offering a reliable picture of these species' changes over time. The ecological usability and limitations of museum butterfly data are discussed by several authors [63,64,65,66,67,68,69]. They consider museum collections indispensable historical and ecological resources for comparing climate change and present conditions. However, Davis [64] argues, "Museum specimens were not collected with the purpose of estimating population trends and thus can exhibit spatiotemporal and collector-specific biases that can impose severe limitations to using NHC data for evaluating population trajectories." With the help of collectors, rare and sporadic species were systematically collected and only those were analyzed for quantitative data. The trend of decline for common species cannot be described from this collection data alone.
The Danube-Ipoly National Park, covering about 600 km² in the middle of the Carpathian Basin, is an excellent source of butterfly data for analyzing these species. As a nature reserve, it minimizes disturbances such as pesticide use (partly banned or restricted in some areas), creation of ecological corridors between core areas instead of habitat fragmentation, systematic eradication of invasive plants, and adaptation of the mowing regime to phenological stages. These measures, allowed for studying the impact of climate change by eliminating most disturbances. The increase in the butterfly population was not only expected but logical. By filtering out factors causing the decline in butterflies, it became clearer which species were sensitive to climate change and which were more adaptive.
Annual data collections ran from early March to the end of October, usually twice a month, in the first and third weeks. On average collection days, a net of three hours was spent in the field, not including travel time between sites. Regular survey areas included Csomád: Magas-hegy, Fót: Somlyó-hegy and Csömör: Ősláp, upon the request of the Danube-Ipoly National Park. This 10-year data set from regular surveys covering all butterfly species is unique in Central Europe and an outstanding source for ecological-historical studies, allowing tracking changes even in common butterfly species.
Here, we employed the Pollard transect method, periodically cross-checked using the MRR (Mark-Release-Recapture) method to reduce the possibility of double-counting specimens. The procedure followed that of Nowicki et al. [70], but in our case, the duration of the Pollard transect was extended to 60 minutes instead of 30 minutes. Additionally, avoiding double counting was more feasible for species that protect their territories (e.g., swallowtail butterfly - Papilio machaon) or where distinct individual lesions were visible during sightings.
The proportion of captured and mounted butterflies was 10%. These were species that were difficult to identify, and several specimens were photographed from multiple angles (wing back, color) for accurate identification, allowing the relative proportions of similar species to be determined. Annual results showed considerable fluctuation depending on weather factors; thus, pooling several years of data in the analysis helped mitigate the impact of annual weather variations. The climatic sensitivity categorization of each species was based on the monograph by Settele and colleagues (Climatic Risk Atlas of European Butterflies) [71] and compared with our own recording experience. Values of Species Temperature Index (STI), Species Precipitation Index (SPI), Habitat Suitability Index (HSI) are based on the papers of Schweiger. et al.; Mora, A.;et al. and Kudrna, et al. [72,73,74] .
2.4. Mathematical Methods and Data Standardization
Over the years, a significant amount of data has been collected: approximately 300,000 specimens of nocturnal moths, 50,000 specimens of butterflies, 40,000 specimens of Diptera, and 60,000 specimens of Hymenoptera. For Hymenoptera, Diptera, and butterflies, data from individual years were aggregated into 3, 5, and 6-year groups to smooth out population fluctuations caused by annual weather variations. Nocturnal moths were the exception, with annual data series being sequentially analyzed. We employed trend analysis to track changes, modeling most changes using linear trends and exponential trends. We plotted the trend line and the trend equation, determining the coefficient of determination (r² value). The coefficient of determination in the trend equation indicates the trend's steepness, expressing the rate of change and its direction (positive or negative). The coefficient of determination (r²) reveals whether our data follows a trend or is dominated by non-trend factors, potentially indicating random changes. The r² value ranges from 0 to 1, with higher values indicating a stronger trend influence, such as climate change or gradual habitat loss and degradation. We calculated the r² using the sum of squares method. The dominance relationships at the beginning and end of the studied period were determined using the Ranking Method or Borda Count method, eliminating the overwhelming effect of outbreaks while appropriately calculating their magnitude and frequency relative to the locations where the ocasional outbreaks occurred.
2.5. Non Native and Invasive Species, Mediterranean Influx
With climate change, species that were previously not members of our natural fauna are appearing. Additionally, species introduced by humans, either intentionally or unintentionally, are becoming more prevalent. Mediterranean newcomers and alien species are treated separately. The northward spread of Mediterranean species is directly attributable to climate change, while the establishment, spread, and reproduction of adventitious species from warmer regions may be facilitated by mild winters.
As a starting point, we utilized the monograph titled "Alien Terrestrial Arthropods of Europe" [75] which lists 330 species from the Carpathian Basin. This number was more than doubled by processing additional works. Insects of alien origin, whether deliberately or accidentally introduced by humans, and species that have migrated northward from the Mediterranean to Central Europe in the last two decades, were analyzed separately. The temporal appearance, taxonomic composition and zoogeographic division of non-native species were depicted on graphs.
Looking back over the last 10,000 years, we evaluated changes that occurred in each historical period, comparing them with the rate of change of the fauna in our present time, and derived mathematical relationships. We looked back 10,000 years, this time, at the end of the Ice Age, [76] the climate had stabilized and present conditions gradually emerged. Therefore, the first three human-mediated introductions of Phthiraptera species and subspecies in the Palaeolithic and earlier are omitted, as subsequent insect movements induced by climate changes completely redrew the fauna during the interglacials. It is challenging to determine the precise introduction date of alien species. Thus, we rely on the date of the first sighting, which can only be indicative. For large, conspicuous species (e.g., Antheraea yamamai), this is a reliable guide. In other cases, there may be a preceding period of incubation or breeding, or the problem may be due to a lack of specialists. Currently, the published date of first sightings is constantly being revised by the growing GBIF database [77], which contains numerous unpublished old records for a wide variety of species. Additionally, archaeology has recently played a significant role in detecting the establishment dates of invasive species, extending these dates back to centuries or millennia [78]. Our detailed data on the invasive and non-native insects of the Carpathian Basin can be found in Appendix C, which is based partly on our own published investigations and partly on papers of various experts from around the world [79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157].
Predicting the emergence and expansion of non-native species or the contraction or vertical movement of native species northward in response to climate change is currently not possible. Numerous predictions exist (for example. [22,71]) based on specific climatic models [158], which predict rapid or slow warming depending on atmospheric pollution. More recent predictions, based on the impact-versus-impact principle with a very high uncertainty factor (e.g., cooling due to the collaps of Atlantic Meridional Overturning Circulation (AMOC) [159,160], have emerged. It must be acknowledged that interference with the climate triggers chaotic processes, creating extreme and highly variable conditions, rendering these predictions outdated. Extreme weather variability is at least as burdensome, if not more so, on wildlife than continuous warming.
The definition of non-native, alien, and invasive species is based on the 1999 US Executive Order #13112 quoted by Ziska et al. [161], which defines "invasive" species and "alien" species for federal agencies as follows: "Alien" species means, with respect to a particular ecosystem, any species, including its seeds, eggs, spores, or other biological material capable of propagating that species, that is not native to that ecosystem. This term also includes non-indigenous or exotic species. In contrast, "invasive or noxious species" means an alien species whose introduction does or is likely to cause economic or environmental harm or harm to human or animal health. We may add: an animal may become an invasive species through deliberate introduction, e.g., the harlequin lady beetle (Harmonia axyridis Pallas, 1773).
3. Results
3.1. Hymenoptera
3.1.1. Bumblebees
Upon preliminary observation, it is evident that bumblebee populations are experiencing a significant and marked decline across both the lower regions of the Carpathian Basin and mountainous areas, with high coefficient of determination (r²) values from 2005 (Table 1). Buff-tailed Bumblebee: Bombus terrestris (Linnaeus, 1758) and Red-tailed Bumblebee: Bombus lapidarius (Linnaeus, 1758) dominate the lower elevations, while White-tailed Bumblebee: Bombus lucorum (Linnaeus, 1761) prevails at higher elevations. Cuckoo bumblebees are rare at lower altitudes, whereas they are primarily found in mountainous areas. As cuckoo bumblebees are parasites of social bumblebees, it is evident that their numbers also decline alongside them. At lower elevations, two Mediterranean-origin bumblebee species, Blood-tailed Bumblebee: Bombus haematurus Kriechbaumer, 1870 and Bombus argillaceus (Scopoli, 1763), are replacing the native fauna. Bombus haematurus may eventually become the dominant species at lower altitudes. Bombus argillaceus did not show significant abundance at higher altitudes. In general, while the Bombus fauna is more abundant in high mountains, a similar declining trend is occurring, albeit with some delay, in the low mountain regions. In high mountainous regions, highly specialized species, such as Bombus gerstaeckeri Morawitz, 1881—already a rare sight in the Carpathians—are particularly vulnerable to the impacts of climate change, which can negatively affect the habitats of their preferred plant species.
Climatic Risk Categories align with the observed decline in bumblebee populations. Species that have declined below detection levels are categorized as extreme high risk (HHHR) at low latitudes, where the impacts of climate change are most pronounced. Conversely, the two species exhibiting population increases are classified as high risk (HR) and low risk (LR) in the literature [22]. A different trend is observed at high altitudes: species experiencing the most severe declines do not fall into the most vulnerable categories. These findings underscore that while climate change significantly impacts bumblebee population dynamics, it is not the sole influencing factor.
In the Carpathians, we have surveyed bumblebees over the past decades. In the Eastern Carpathians, certain species, such as Bombus soroeensis (Fabricius, 1776), appear to be thriving in the more humid glades within forested areas. Conversely, species like Bombus wurflenii Radoszkowski, 1860, which typically inhabit the transition zone between the upper coniferous forest and the subalpine region, have become less common, likely due to increased exposure to direct climatic fluctuations. Additionally, Bombus pyrenaeus Pérez, 1879, has not been observed in recent years, despite extensive efforts to locate it.
Meteorological data for the Eastern Carpathians indicate a significant rise in temperature and solar radiation, accompanied by a marked decline in humidity at higher altitudes, particularly in the subalpine zone. These climatic changes are likely driving the observed shifts in Bombus species distribution and abundance. At high altitudes, after a general decline since 2005, B. lucorum appears to have stable populations. Another species exhibiting notable success is the Common Carder Bumblebee: Bombus pascuorum (Scopoli, 1763), which can frequently be found up to the subalpine zone in the Ceahlău Mountains, after its decline in the last two decades. Alongside B. pascuorum and B. soroeensis, Bombus campestris (Panzer, 1801) is also commonly encountered at elevations reaching up to 1750 m. However, the impact of climate change is evident on oligolectic species such as B. gerstaeckeri, which forages primarily on Aconitum spp., plants that thrive in shaded, cool, and moist environments. Despite extensive surveys, we have not been able to locate this species in recent years, suggesting a potential decline in its population linked to habitat changes driven by climate fluctuations. Some less frequent species, such as Bombus confusus Schenck, 1861, Apple Bumblebee: Bombus pomorum (Panzer, 1805), and the Large Garden Bumblebee: Bombus ruderatus (Fabricius, 1775), tend to expand or persist in very small populations in lower mountainous areas (though this should be closely monitored in the coming years to ensure it is not accidental).
Of particular interest is the adaptability of B. haematurus, which appears more inclined to spread along forest margins, parks, and dry open areas, compared to B. argillaceus. Based on our observations, B. argillaceus is found predominantly within deciduous forests and parks, suggesting differing ecological tolerances between these two species. The maximum altitude for B. haematurus in Romania in recent years was 780–800 m (Valea Uzei – Râmeț) in the Apuseni Mountains [162]. For B. argillaceus, the highest recorded altitude was 852 m (around Gheorghieni, Harghita) [162]. Additionally, B. argillaceus was observed in a valley north of the Făgăraș Mountains (Sâmbăta de Sus) at approximately 770 m. In the Eastern Carpathians, it was recorded at a lower altitude of around 680 m (Ticoș, Neamț County) either. These observed population trends highlight the need for continued and focused monitoring in the coming years to better understand the long-term impacts of climate change and habitat modifications on Bombus species.
3.1.2. Aculeata
The two main groups under examination display radical ecological differences. The suborder Symphyta typically exhibits higher species richness in northern Europe, while the Aculeata group reaches its peak species richness in southern regions. For instance, Finland records 690 Aculeata species and 769 Symphyta species, a ratio of approximately 1:1. In contrast, Hungary reports 1,400 Symphyta species and 615 Aculeata species, yielding a ratio of about 2:1. Turkey presents an even more pronounced disparity with 370 Symphyta species compared to 3,382 Aculeate species, a ratio nearing 10:1 [163,164,165,166,167,168]. Consequently, the Mediterranean transformation of Central European fauna is most evident through the proliferation of Aculeata species and the gradual decline of Symphyta species. The subsequent figures (Figure 4, Figure 5, Figure 6 and Figure 7) illustrate this process, partially based on our previous paper [169].
The .Appendix A summarizes the changes in the 241 most common species within the deeper areas of the Carpathian Basin, categorized by their ecotypes: eurytherm xerophile, stenotherm hygrophile (sh), eurytherm intermediate (ei), eurytherm hygrophile (eh), and stenotherm xerophile (sh). Given that approximately 85% of these species fall into the intermediate and xerophilic groups, it follows that gradual warming would lead to a significant increase in individual numbers. Several groups, however, have experienced substantial declines in abundance. One such group includes the cool and humid-adapted bees, primarily from the genera Megachile and Osmia, and to a lesser extent Andrena. Another affected group is the bumblebees (Bombus spp.), which warrant separate discussion due to their economic importance and unique responses. Additionally, numerous Crabronidae species inhabiting wet marshes or wooded areas are also in decline. Ilyen módon az Aculeata csoportcsaládjai és nemzetségei különböző képpen regálnak a klímaváltozásra. We can say that up to the current extent of warming, the reaction is positive, except for two genera, namely Crabro Fabricius, 1775 and Crossocerus Lepeletier & Brullé, 1834. (Table 2).
Of the approximately 1,480 species that once inhabited the lower Carpathian Basin, 209 species have not been recorded for 20 years or more. These have been categorized into three groups. Species that have found refuge in the higher elevations of the Tatras or the Carpathians are indicated in grey in Table 3. Species absent due to rarity have been excluded from our list. For the remaining species, those that have disappeared from our region but have been recorded in Scandinavia, the northern parts of Western Europe, the Atlantic region, or the Alps. The primary distribution of these species serves as an indicator of climate sensitivity, suggesting that these species may have shifted their southern range limit further north, thereby reducing their abundance below the detection limit in our region.
3.1.3. Sawflies, Symphyta
Due to their northern distribution, it is not surprising that climate change affects this group significantly. Using Malaise trap methods, we have successfully demonstrated this decline [6], as reflected in the Figure 7. Regular data from sweep net collections have been available since the 1960s. Although this method collects few specimens (300-700 per year). Thus, the decline in collected specimens is masked by variations in different habitats, yet the decreasing trend in species that were once common is still noticeable only (Table 4).
Most Tenthredo species show a strong declining trend, as does the once-common Pachyprotasis rapae (Linnaeus, 1767). It is not surprising that the number of species in the Argidae family, which is primarily characteristic of the Mediterranean (and also the Afrotropical region), is increasing. Among Symphyta, tseveral agricultural and horticultural pests belong to this family. Our data indicate that the current rate of warming has positively impacted the Turnip Sawfly: Athalia rosae (Linnaeus, 1758), and two Dolerus species that occasionally damage cereals and grass crops. The drastic decline in the number of Athalia ancilla Serville, 1823 began before climate change became noticeable. Additionally, it appears that climate change is also unfavorable for the Wheat Stem Sawfly: Cephus pygmeus (Linnaeus, 1767).
Migration within the Carpathian Basin has also been observed in this group. The northernmost distribution of Pseudocephaleia praeteritorum (Semenov, 1934) was previously Hungary, but it was found in Slovakia in 2019 [170]. The mountainous regions surrounding the Carpathian Basin still provide refuge for previously sporadic species [6].
3.2. Diptera
3.2.1. Horse-Flies, Tabanidae
Horse-flies are primarily xerothermic. Therefore, the Tabanidae population has shown an increase (Table 5 and Figure 8). We saw significant decline only at the Band-eyed Brown Horse-fly: Tabanus bromius Linnaeus, 1758. Moderately signifcant decline was experienced at the Common Horse-fly: Haematopota pluvialis (Linnaeus, 1758). Among these, Haematopota pluvialis is the most common hylophilous species in our region. When examining trends in detail, the vast majority of species have shown an increase, which is not surprising given that most members of this group are xerothermic.
In the long term, since the larvae require aquatic or semi-aquatic environments for development, species richness and population density are expected to decrease as these environments become more scarce. In the last part of the time series, these signs are already visible. Thus far, many species in this group have benefited from climate change.
While there is a close correlation between the ecotypes of individual species, it is not absolute. We can say that species showing stronger growth trends, with one exception, all have xerothermic ecological requirements in their adult form. The number of species exhibiting a decline is very small, with only two species showing moderate or strong decreasing trends. One of the commonest species, the hylophilous Haematopota pluvialis(, also showed strong increase till the end of 1990s, after this, the population density of this species dropped.
Overall, the response of Tabanidae species to climate change is twofold: while warming has favored xerothermic species in the adult stage, it is also evident that the larvae are dependent on water. Seven species have not been collected in the Carpathian Basin for 20 years or more. Among these, three species appear to find refuge in higher elevations of the region, such as the Tatras or the Carpathians. These species are Pangonius pyritosus (Loew, 1859), Hybomitra aterrima (Meigen, 1820), and Hybomitra montana (Meigen, 1820).
3.2.2. Hoverflies, Syrphidae
This group can be characterized by a significant decline in numbers, regardless of ecotypes (Table 6 and Figure 9). Among the studied Diptera families, this group shows the strongest decline. The likely reason is the moisture-dependent lifestyle of the larvae. However, this explanation is not entirely satisfactory, as the larvae of Tabanidae also require aquatic or semi-aquatic environments but do not exhibit the same intense decline as hoverflies.
The overall picture becomes even more interesting and disheartening when we examine whether the mountainous regions of the Carpathian Basin provide refuge for species that have disappeared from lower-lying areas or have fallen below detectable levels. A significant number of Syrphidae species have not been collected in this century, although they were once sporadically found throughout the Carpathian Basin. Among the four studied groups, there are 89 species disappeared in the last 2 decades, with the vast majority, 50 species, belonging to the Syrphidae. We investigated whether high-altitude habitats serve as refuges for these species. Of the 89 Diptera species, only 14 have been found in the lst 24 years in the Tatras or the Carpathians, representing a mere 16%.
Previously established ecotypes closely correlate with population changes. Almost all ecotypes have seen a rapid decline in numbers. However, we observed a strong decline in the number of silvicol and intermediate species, while the decline in xerothermic species was moderate or showed a balance. Interestingly, the only species showing a significant increase is Anasimyia contracta Claussen & Torp, 1980, an Atlantic moisture-loving species. This indicates that while climate change greatly influences population dynamics, it is not the sole factor, and we are facing highly complex processes.
3.2.3. Bee Flies, Bombyliidae
We examined all common or frequent species within the approximately 65 species of Bombyliidae. With few exceptions almost all species have shown an intense increase in numbers or have remained stable over the past 40 years (Table 7 and Figure 10). Some species likely includes montane, forest species, which are among the few within the family that are sensitive to climate change. Their distribution supports this, as they are most common between the latitudes of 48-52 degrees in Western Europe (GBIF, [77]).
Three Bombyliidae species have not been detected in the Carpathian Basin for 20 years or more: Bombylius trichurus Wiedemann, 1818; Bombylius quadrifarius Loew, 1855; and Bombylius ambustus Pallas & Wiedemann, 1818. The disappearance of these southern species is certainly not due to climate change.
3.2.4. Tachinidae
No growth trends were observed for any species (Table 8 and Figure 11). However, the decline is not as pronounced as in the Syrphidae species (for comparision see also Table 9). On the other hand, several species, including Linnaemya frater (Rondani, 1859), Meigenia dorsalis (Meigen, 1824), Exorista larvarum (Linnaeus, 1758), and Tachina fera (Linnaeus, 1761), have shown specific trends: the general trend is decline but displayed significant increase in the last five years of investigation.
These species primarily have silvicol (forest-dwelling, cooler microclimate-preferring) ecological requirements (12 species) or are intermediate in terms of temperature (6 species), with only one xerothermic species (Table 8). Based on this, most Central European Tachinidae species are sensitive to climate change. Among the 27 species not collected for over 20 years, only seven are known to still exist in the high mountains surrounding the Carpathian Basin (and 17 of the 50 hoverfly species) (Table 10). These include Cadurciella tritaeniata (Rondani, 1859), Catharosia albisquama (Villeneuve, 1932), Ceranthia tristella (Herting, 1966), Chetoptilia puella (Rondani, 1862), Eloceria delecta (Meigen, 1824), Ligeriella aristata (Villeneuve, 1911), and Winthemia bohemani (Zetterstedt, 1844), primarily of Scandinavian distribution. The species Amelibaea tultschensis (Brauer & Bergenstamm, 1891) and Bithia acanthophora (Rondani, 1861) are rare mountainous species, with records from the Italian Alps. The species Anthomyiopsis plagioderae (Mesnil, 1972) is of Atlantic distribution, while Vibrissina debilitata (Pandellé, 1896) is of Scandinavian and Atlantic distribution. The species Aphria xyphias (Pandellé, 1896), Campylocheta latigena (Mesnil, 1974), Conogaster pruinosa (Meigen, 1824), Phytomyptera abnormis (Stein, 1924), Estheria acuta (Portschinsky, 1881), Gonia bimaculata (Wiedemann, 1819), Heraultia albipennis (Villeneuve, 1920), Minthodes pictipennis (Brauer & Bergenstamm, 1889), and Psalidoxena transsylvanica (Villeneuve, 1929) have always been extremely rare in our region (GBIF database, [77]).
3.3. Lepidoptera
3.3.1. Butterflies, Rhopalocera
The Table 11 and Table 12 show the changes in the number of individuals of certain species of butterflies between 1980 and 2023. The correlation between the Species Temperature Index (STI), Species Precipitation Index (SPI), Habitat Suitability Index (HSI) values, and changes in the number of individuals of each butterfly species is very weak for HSI (0.075), STI (0.038), and SPI (0.058), but stronger for Climate Risk Category (CRC) values. When the CRC (1980-2009) values are ranked from 1 to 6 (1: HHHR - extreme high risk, 6: PR - potential risk), and the most resistant species are assigned a value of 6, the correlation coefficient of 0.35 indicates sginificant connection (Table 11). Conversely, the correlation between the calculated CRC (2016-2023) values (Table 12) and the strength of the change in the number of individuals (lin x correlation coefficient) in the national park is only 0.034, indicating no real correlation. This suggests that the relationship between the decrease in the number of butterfly species and climate change exists and, in our case, is of medium strength (Table 11 ). On the other hand, the increase in more or less climate-resistant species is a result of nature conservation measures rather than climate change (Table 12 and Table 13).
Where we have observed a strong increase in certain species, we can infer that these species have shown resilience to the extent of climate change to date, but other conditions were necessary for their increase, such as protected habitat, a network of core areas and ecological corridors, prohibited or strictly controlled pesticide use, and mowing regimes adapted to the species' lifestyle. Therefore, sensitivity to climatic conditions is becoming more pronounced over time and is strongly reflected in the numbers of many butterfly species. Furthermore, the reproduction of species less sensitive to climatic variation requires the aforementioned conditions.
These findings are described in more detail below. Less climate-sensitive species have experienced a strong increase in the last 10 years listed on Table 11 and Table 14. In contrast, the populations of sensitive species have declined since the mid-1980s (e.g., Meleager's Blue: Polyommatus daphnis (Denis & Schiffermüller, 1776), Purple-edged copper: Lycaena hippothoe (Linnaeus, 1761); or mostly from the early 90s ( Southern Festoon: Zerynthia polyxena (Denis & Schiffermüller, 1775), False Heath Fritillary: Melitaea diamina (Lang, 1789), Large Chequered Skipper: Heteropterus morpheus (Pallas, 1771), Purple Emperor: Apatura iris (Linnaeus, 1758), Camberwell Beauty: Nymphalis antiopa (Linnaeus, 1758), and Small Tortoiseshell: Aglais urticae (Linnaeus, 1758) or have suffered from an intense decline in numbers from the second half of the 1990s onwards (e.g.,Twin-spot Fritillary: Brenthis hecate (Denis & Schiffermüller, 1775)) (Table 12, Table 13 and Table 15). Among the listed species, climate change plays a major role in the decline of almost all, with the exception of N. antiopa, classified as LR (low risk) and only slightly sensitive to climate change.
The other group includes species that have been resistant to climate change thus far, showing a significant increase in density since the early 2020s (Table 12 and Table 13). This increase occurred only under conservation protection, excluding other factors affecting their natural abundance but not mitigating the impact of climate change. Species such as Meadow Brown: Maniola jurtina (Linnaeus, 1758), Dryad: Minois dryas (Scopoli, 1763), Marbled White: Melanargia galathea (Linnaeus, 1758), Great Banded Grayling: Brintesia circe (Fabricius, 1775) (Figure 12), Common Blue: Polyommatus icarus (Rottemburg, 1775), Brown Argus: Aricia agestis (Denis & Schiffermüller, 1775) and Chestnut Heath: Coenonympha glycerion Borkhausen, 1788 were notable for both the intensity of the increase in numbers (slope of the trend line) and the strength of the trend (r²), alongside several other species (see Table 12). The species that declined in the 1990s and 2000s have not been able to increase their numbers despite conservation efforts, suggesting that climate change has most likely played a critical role in their decline. Aglais urticae, once common until the late 1980s and then extremely rare by the early 1990s, is now absent from most records. Since the early 2000s, it has been largely absent from surveys, now a distinct rarity in the deeper areas of the Carpathian Basin.
At the family level, gossamer-winged butterflies (Lycaenidae) have been the most affected by climate change: our studies show that most species are below detection levels, their population density compensated by some common species, while populations of rare and sporadic species have drastically reduced. A similar pattern is seen in the Nymphalidae family, where the decline of the vast majority of species was offset by a few species formerly belonging to the Satyridae, listed above.
Overall, climate change has had a negative impact from a conservation perspective for a significant proportion of species. Rare or sporadic species have drastically declined since the 1990s. Conservation measures have ensured that, with the exception of a few species, butterflies have not yet disappeared from the Carpathian Basin. In protected habitats, significant reproduction has been achieved, but this increase predominantly affects species that were already common. The vast majority of species that were once rare or sporadic are expected to fall victim to climate change in Central Europe, and conservation measures alone will not be sufficient to protect them.
Under optimal, protected conditions, only certain species from the families Papilionidae, Nymphalidae, and Lycaenidae were able to increase their numbers, ensuring an upward trend for these families (Figure 13). However, families like Hesperidae and Pieridae did not show a positive trend even under these conditions (Table 12, Table 14 and Table 15). The number of species collected in each period showed a slight downward trend (.Table 13 and Table 14). This decrease is not only local. In the last 20 years, seven species have disappeared (or fallen below the detection limit), while another three species are considered historically extinct, even before climate change. These are: Marbled Skipper: Carcharodus lavatherae (Esper, 1783), Large Heath: Coenonympha tullia (Müller, 1764) and Esper's Marbled White: Melanargia russiae (Esper, 1783)). Nine of the 10 species find habitat in the mountain ranges surrounding the Carpathian Mountains (Tatras, Carpathians) up to a given extent of climate change (Table 16). The species Damon blue: Polyommatus damon (Denis & Schiffermüller, 1775) has not been collected in the lower regions of the Carpathian Basin for 10 years, so it, like the previous species, persists in higher regions in our region.
3.3.2. Nocturnal Macrolepidoptera
Similarly to the vast majority of local faunistic data and regional reports, we found a decline in numbers, but it was so slight that it was not significant, and no significant r² value was measured (Table 17 and Figure 14). Compared to the 1970s, that slight decrease in the number of specimens caught, turned into a slight increase from 2014. At the same time, the decline in species richness (number of species below the detection limit in a given area) is decreasing slightly but showing a real trend (r²=0.14) (Table 17 and.Figure 15). By family, the Noctuidae (owlet moths) are the most sensitive in terms of numbers, and the decline in this group has not stopped even in the last decade. Other families, especially the Geometridae, have experienced a less significant decline in numbers compared to 1970.
In terms of species captured by individual traps (Figure 16), local species depletion has been most pronounced in the Geometridae family. Meanwhile, in the Noctuidae and Erebidae families, species diversity captured by individual traps has increased (Table 18). A strong increase in the total number of species studied was observed in Willow beauty: Peribatodes rhomboidaria (Denis & Sch., 1775), Large Yellow Underwing: Noctua pronuba (Linnaeus, 1758), Aspen Lappet: Phyllodesma tremulifolia (Hübner, 1810), Clay Fan-foot: Paracolax tristalis (Fabricius, 1794), Pine Hawk-moth: Hyloicus pinastri (Linnaeus, 1758), Jubilee Fan-foot: Zanclognatha lunalis (Scopoli, 1763), Cloaked Carpet: Euphyia biangulata (Haworth, 1809), Catocala hymenaea (Denis & Schiffermüller, 1775), Orange Footman: Eilema sororcula (Hufnagel, 1766) and Pale Oak Beauty: Hypomecis punctinalis (Scopoli, 1763). These are believed to be the most adaptive to the extent of climate change to date. The most sensitive species were Latticed Heath: Chiasmia clathrata (Linnaeus, 1758), Eyed Hawk-moth: Smerinthus ocellata (Linnaeus, 1758), Ruby Tiger: Phragmatobia fuliginosa (Linnaeus, 1758), Shining Marbled: Pseudeustrotia candidula (Denis & Schiffermüller], 1775), White Ermine: Spilosoma lubricipeda (Linnaeus, 1758), Giant Looper: Ascotis selenaria (Denis & Schiff., 1775), Straw Dot: Rivula sericealis (Scopoli, 1763), Cabbage Moth: Mamestra brassicae (Linnaeus, 1758) and Garden Tiger Moth: Arctia caja (Linnaeus, 1758) (Appendix B).
From a practical point of view, certain agricultural pests (Common Cutworm: Agrotis segetum (Denis & Schiffermüller) 1775, Heart and Dart: Agrotis exclamationis (Linnaeus, 1758), Bright-line Brown-eye: Lacanobia oleracea Linnaeus, 1758) have not been sensitive to climate change to date and have been found to be tolerant of climate change. Population fluctuations of these agricultural pests were close to equilibrium. Moderately significant declines were observed in populations of the Setaceous Hebrew Character: Xestia c-nigrum (Linnaeus, 1758), and the Fall Webworm: Hyphantria cunea (Drury, 1773), significant declines in populations of the Cabbage Moth: Mamestra brassicae (Linnaeus, 1758) and moderate increases in populations of the Silver Y: Autographa gamma (Linné, 1758) and the Cotton Bollworm: Helicoverpa armigera (Hübner, 1808). The latter two species are migrant butterflies from the south (Appendix B).
Among forest pests, an increase was observed in the Swallow Prominent: Pheosia tremula (Clerck, 1759) and the Pine Beauty: Panolis flammea Denis & Schiffermüller, 1775. The Eastern Nycteoline: Nycteola asiatica (Krulikowsky, 1904), the Archer's Dart: Agrotis vestigialis (Hufnagel, 1766), the Bordered White: Bupalus piniaria (Linnaeus, 1758), the White Satin Moth: Leucoma salicis Linnaeus, 1758, the Poplar Tip Moth: Clostera anastomosis (Linnaeus, 1758), the Brown-tail Moth: Euproctis chrysorrhoea (Linnaeus, 1758), the Oak Processionary: Thaumetopoea processionea (Linné, 1758), Lackey Moth: Malacosoma neustria (Linnaeus, 1758) and the Gypsy Moth: Lymantria dispar (Linné, 1758) did not show any trend-like variation in positive or negative directions, i.e. they were not affected by the warming up to date. In practice, this means that climate change has had a positive or neutral effect on these forest pests. But, as local researchers write [182,183,184,185], forest pests need to be monitored, as these species, but a significant proportion of nocturnal moths in general, are more susceptible to outbreaks due to gradual warming. The increasing prevalence of outbreaks can be examined in .Appendix B...
In the beginning and end of the 50 years studied, all but 5 of the top 20 species with the highest abundance were replaced, which means that climate change has rearranged the dominance of each species, almost completely (Table 16). If we take the 20 most common species, the Rosy Footman: Miltochrista miniata (Forster, 1771), the Small Quaker: Orthosia cruda Lempke, 1964, the Four-spotted Footman: Lithosia quadra (Linnaeus, 1758), the Orange Footman: Eilema sororcula (Hufnagel, 1766) and the Litter Moth: Polypogon tentacularia (Linnaeus, 1758) have remained in the dominant 20 species, as they did in the 1970s and 1980s as 30-50 years later in the 2010s and 2020s.
Codes of Table 19: 0 = xerothermophilic species (burning, semi-dry grassland, dry fallow land, heaths, gravel pits, quarries); 1 = ubiquitous open land species and cultural followers in fields, meadows, gardens, parks and on forest edges; caterpillars almost exclusively on grasses and herbs; 2 = ubiquitous species and cultural followers in open land and forest biotopes; caterpillars on grasses, herbs or shrubs; 3 = species mostly in tree-covered areas; common in forests, but also in gardens or parks with good tree cover; caterpillars predominantly on grasses, herbs or shrubs; 4 = like group 3, but caterpillars predominantly on hardwood and coniferous wood; 5 = like group 3, but more oriented towards softwood floodplains; caterpillars predominantly on softwood; 6 = forest species; caterpillars mostly on wood, rarely on herbs or grasses; 6a = ubiquitous forest species, 6b = coniferous forest species, 6c = deciduous forest species; 7 = species of moist forests and open wetlands; caterpillars predominantly on algae, lichens and liverworts; 8 = as in group 7, but caterpillars predominantly on deciduous wood (mainly hygrophilous floodplain, marsh and moor forest inhabitants; 9 = as in group 7, but caterpillars on grasses and herbs (mainly sedge meadows and reed beds inhabitants); 10 = typical floodplain forest species of larger river valleys; caterpillars predominantly on soft deciduous wood or clematis)
Regarding changes in dominant species, we observed the following: no extremely xerophilic species dominated in any period. The extreme hygophilous species, the Straw Dot: Rivula sericealis (Scopoli, 1763) and the Mottled Rustic: Caradrina morpheus (Hufnagel, 1766) have been eliminated from the top 20 species. The proportion of species native to open grasslands has declined, indicating their sensitivity to climate change. Their place has understandably been taken by other species in more closed habitats, leading to a shift in favor of species living in habitats more protected from climate exposure.
The Table 16 lists species that have been missing for 20 years or more. Species marked with an X are still found in the higher regions of the Carpathian Basin (Tatras, Carpathians). Most nocturnal butterfly species from low-lying regions disappeared several decades ago, before the 1980s. Only three species from the list are suspected to have been affected by climate change: Cosmotriche lobulina (Denis & Schiffermüller, 1775), Eublemma pannonica (Freyer, 1840), and Syngrapha ain (Hochenwarth, 1785).
3.4. Non Native Species, Mediterranean Influx
3.4.1. Non Native Species
The most comprehensive work on our region and Europe to date is the Biorisk series monograph titled "Alien Terrestrial Arthropods of Europe" [76] which lists about 330 species from the studied region. Through deep research, we have expanded this list to 803 species by carefully reviewing faunistic reports and the literature. Our chart shows that the influx of alien species is accelerating, and the dynamics of the influx of established alien insects can be described by a exponential equation (Figure 17) .
The first alien insect species likely appeared in the Carpathian Basin with Homo erectus, as parasites living on the human body. The first major invasion wave occurred in the Neolithic era when humans brought various livestock, such as sheep or goats, along with the first invasive arthropods of veterinary importance. The introduction of agriculture also brought the first storage pests from the Eastern Mediterranean [78]. During antiquity, with Roman conquests, the first exotic animals, like peacocks and pheasants, likely introduced new arthropods (their lice, ectoparasits); also those associated with the European rabbit, which was bred by the Romans.
Medieval records of invasive insects primarily document locust swarms, with the earliest record from 1191. The 1346 plague, which reached the Carpathian Basin in 1349, is often attributed to invasive rats and their fleas. Transoceanic insect invasions likely began with maritime exploration and geographic discoveries, bringing exotic animals (like guinea pigs) and food and fodder plants (tomatoes, peppers, corn, tobacco, etc.). However, early entomological literature, such as the richly illustrated manuscript "Mira Calligraphiae Monumenta" from 1590, did not depict any alien insect species.
After Linné, researchers in our region turned their focus towards insects. The first description of invasive species from our area, published by Matyás Piller and Lajos Mitterpacher, was the Monochrome Spider Beetle: Epauloecus unicolor (Piller & Mitterpacher, 1783), a foreign warehouse pest from the Dermestidae family. As international trade increased, so did the number of reports and descriptions of new alien species. This accumulation has become exponential over the past quarter-century, with climate change playing a significant role, according to the literature.
In terms of distribution, aaproximately half of the non-native insect species came from the Eastern Palearctic (144 species, 18%) and North American regions (Nearctic, 135 species, 17%). We will not discuss these in more detail, as their spread was already ensured earlier due to climatic similarities, and the main driver here is the increasing logistics. The milder winters have contributed to the influx of Mediterranean, tropical, and subtropical species, accelerating their invasion, and allowing many species to survive and proliferate to detectable levels.
The majority of incoming species, approximately 38%, belong to the order Hemiptera (true bugs, cicadas, aphids, planthoppers, leafhoppers, assassin bugs, scale insects, bed bugs, and shield bugs), with scale insects constituting the majority (83 species). Due to their small size (especially as overwintering larvae), they can easily evade plant health checks, whether on propagating material, ornamental plants, or the skins of imported fruits. Some have only been able to reproduce in greenhouses until now, but increasingly, thanks to milder weather, they can survive outdoors, such as Ceroplastes ceriferus (Fabricius, 1798), which has been detected in the open field. Whether it can overwinter in our region remains to be seen, but it is likely. Originally, the Southern Green Stink Bug: Nezara viridula (Linnaeus, 1758) was also occured exclusively in greenhouses, but it has now become a significant outdoor pest.
The second largest group is beetles, due to their very high diversity in nature. Many beetle species are warehouse pests and are spread worldwide through transportation, making the original habitats of many species difficult to trace. An example of an invasive beetle is the Asian Lady Beetle: Harmonia axyridis (Pallas, 1773), introduced as a biological control agent, which is now displacing native ladybird species. Another alien ladybird species, the Mealybug Destroyer: Cryptolaemus montrouzieri Mulsant, 1853, is polyphagous and may threaten non-damaging non-target fauna.
Other beetle species associated with crops and seeds include the Asian Seed Beetle: Megabruchidius dorsalis (Fahraeus, 1839), the East Asian Seed Beetle: Megabruchidius tonkineus (Pic, 1904), the Minute Beetle: Murmidius ovalis (Beck, 1817), and Bruchidius siliquastri Delobel, 2007. Beetles associated with woody plants include the Brown Powderpost Beetle: Lyctus brunneus (Stephens, 1830), the Red Palm Weevil: Rhynchophorus ferrugineus (A. G. Olivier, 1791), the Black Timber Bark Beetle: Xylosandrus germanus (Blandford, 1894), the Black Borer: Apate monachus Fabricius, 1775, and the Date Stone Beetle: Coccotrypes dactyliperda (Fabricius, 1802). These beetles have been introduced into European ports and easily enter the Carpathian Basin, acting as forest or crop pests. The Black Larder Beetle: Dermestes haemorrhoidalis Küster, 1852, is known for infesting mummified human corpses.
Certain tropical and subtropical species, such as the Tomato Leafminer: Tuta absoluta (Meyrick, 1917), Hipoepa fractalis (Guenée, 1854), Monema flavescens Walker, 1855, the Potato Tuber Moth: Phthorimaea operculella (Zeller, 1873), the Egyptian Cotton Leafworm: Spodoptera littoralis (Boisduval, 1833), the Locust bean moth: Apomyelois ceratoniae (Zeller, 1839), and the Geranium Bronze: Cacyreus marshalli Butler, 1897, may have increased in prevalence due to climate change. According to internet sources, Cacyreus marshalli has already reached Zagreb. This non-native butterfly lives on Pelargonium and is a synanthropic species, similar to many invasive organisms that spread during the Neolithic period. Notably, the colonization of the Palm Moth: Paysandisia archon (Burmeister, 1879), a member of the Castniidae family (Lepidoptera) previously unknown in our region, is significant. The larvae of this large moth of South American origin can overwinter inside the ornamental plants, and some of its host plants can survive outdoors in the Carpathian Basin climate.
In the 2000s, two heat-demanding tropical and northern Mediterranean cockroach species, Periplaneta australasiae (Fabricius, 1775) and Planuncus tingitanus (Bolívar, 1914), became widespread in our region.
Economically, the Southeast Asian Mantis: Hierodula tenuidentata Saussure, 1869, poses a minimal threat but may carries nature conservation risks in Transylvania. Dragonflies, due to their excellent flying ability, can naturally colonize northern areas from the Mediterranean. However, one species, the Red-veined Dropwing: Trithemis arteriosa (Burmeister, 1839), was introduced by humans from Cyprus through the import of aquatic plants, likely arriving with dragonfly larvae.
Among the alien species, Xenylla uniseta Da Gama, 1963, was the first springtail recorded in the Carpathian Basin. Within the primitive insect groups, the South-East Asian Long-Tailed Silverfish Ctenolepisma longicaudatum (Escherich, 1905), a relative of the European silverfish, Lepisma saccharinum Linnaeus, 1758, is noteworthy.
Since 2004, alien hymenopteran species permitted for agricultural use (Ministerial Regulation on Biological Control, No 89/2004, FVM Decree) have been spreading. Some species, such as Torymus sinensis Kamijo, 1982, introduced for biological control of the chestnut gall wasp (Dryocosmus kuriphilus Yasumatsu, 1951), are now found in the wild (Kriston et al., 2016).
Among Diptera, larvae of the black soldier fly: Hermetia illucens (Linnaeus, 1758) have been used in organic waste processing, with potential genetic modifications to expand their waste consumption range. Rhynchomicropteron nudiventer Papp, 1982, described by Hungarian researcher László Papp from India, was caught in Hungary over three decades after its description. Clogmia albipunctatus (Williston, 1893) became a mass species after 2012, frequently found in restrooms and sanitary facilities, gaining hygienic significance. In hospitals, as a passive vector of various bacteria, it poses a risk to patients when colonizing operating theaters.
The Tiger Mosquito: Aedes albopictus (Skuse, 1894) is a vector of arboviral pathogens such as West Nile virus, yellow fever virus, encephalitis, dengue fever, Chikungunya fever, Zika virus, and several nematodes (e.g., Dirofilaria immitis).
3.4.2. Mediterranean Influx
According to our results, of the 803 alien insect species counted so far (Appendix C), 22% are of Mediterranean origin, and 21% are from one of the tropical regions. The quantity of species imported from the Nearctic (17%) and temperate Asian (Eastern Palaearctic) region (18%) is also significant (Figure 19). The most difficult thing to decide about Mediterranean invasive species is whether they have arrived in Central Europe naturally, as a result of climate change (we are neighbours of the Mediterranean region), or whether they have been affected by human intervention. There are a few cases where human conscious or accidental transport is documented. For example, several Mediterranean species have been introduced and spread for biological control purposes, such as the Hunter Fly: Coenosia attenuata Stein, 1903, Diptera, Muscidae, (predator fly); Orius laevigatus (Fieber, 1860) Heteroptera, Anthocoridae (predator bug).
But it is quite likely that it has been introduced by the Juniper Ermine Moth: Argyresthia trifasciata Staudinger, 1871, Pyralidae, (with dried food) or Phyllonoricter millierella Staudinger, 1871, Gracillariidae (with propagules of the Celtis australis). It is not surprising then that there are species that have been introduced both naturally and artificially, i.e. more than once, such as the Cypress Jewel Beetle: Ovalisia festiva (Linnaeus, 1767).
Here we focus mainly on those species, from the last quarter of a century alone, that may have spread without human intervention, simply following weather trends.
In this group, there is so-called internal dispersal, where the range of a species is limited to the Carpathian Basin, and within this range it can naturally spread from the southern countries to the northern countries. The Pine Processionary: Thaumetopoea pityocampa (Denis & Schiffermüller, 1775) (Notodontidae) is recorded from Hungary as early as 1977, but it reached Slovakia by 2011. The northern limit of the distribution of the Mediterranean Pseudocephaleia praeteritorum (Semenov, 1934) (Pamphiilidae) has been Hungary, from where it has been known for a long time, but was collected in Slovakia for the first time in 2019. The Bat Hawkmoth: Hyles vespertilio (Esper, 1779) first entered our region as a migrant in 2020 and has been observed several times since then. It has been recorded in 2020 and has been observed since then. In such cases, the area boundary fluctuates depending on climatic conditions. A good example is the Fig-tree Skeletonizer Moth: Choreutis nemorana (Hübner, 1799), (Choreutidae), which feeds on fig leaves, appeared in 1955 in the southern Carpathian Basin, receded and reappeared in 2011 due to warming. But it also includes Tenthredo costata Klug, 1817 (Tenthredinidae), a species common in the Mediterranean and Anatolian regions, sometimes dominant in some places, which occurred sporadically in Hungary in the 1970s, then disappeared and was recaptured on 8 June 2016 at the Nagybajom woodcutter. These kind of species can be identified by the fact that they are common or even common in the Mediterranean, while they are rare in our country and absent in areas further north, since we are on the border of their distribution area, sometimes they disappeare sometimes appeare as the “border” of their area is fluctuating. There is relatively little information on the species that move northwards, partly because they do not receive the same attention as agricultural and forest pests, and partly because there are very few specialists collecting, identifying and publishing them. Based on the available data for the Carpathian Basin, the following can be said about the last quarter of a century. Ectobius vittiventris (Costa, 1847) is a species of cockroach that is distributed northwards and is otherwise harmless. Of the two species of praying mantis, Ameles spallanzania (Rossi, 1792) has reached the central parts of the Carpathian Basin, while Iris oratoria (Linnaeus, 1758) has conquered the eastern part of the Carpathian Basin. A Neuroptera, Sisyra iridipennis Costa, 1884 and three Mediterranean dragonfly species, the Goblet-marked Damselfly: Erythromma lindenii (Selys, 1840), the Black Pennant: Selysiothemis nigra (Vander Linden, 1825) and the Violet Dropwing: Trithemis annulata (Palisot de Beauvois, 1807), also reached the north. The host plants of three scale insects of Mediterranean origin Aonidia lauri (Bouché, 1833), Carulaspis minima (Signoret, 1869), and Dynaspidiotus britannicus (Newstead, 1896) ) can also survive in the wild and may have spread spontaneously or been introduced by plants. The same is true for Livilla variegata (Löw, 1881), Trioza alacris (Flor, 1861) and Uroleucon telekiae (Holman, 1965). The Mediterranean Acrosternum heegeri Fieber, 1861, which is a polyphagous bug, has been on the path to invasion, and its introduction into the Carpathian Basin is likely to have been by introduction, but spontaneous colonisation is not excluded. Entomobrya unostrigata Stach, 1930 is a springtail of Mediterranean origin. The Black Fig Fly: Silba adipata McAlpine, 1956, has been introduced into our region as a pest of figs this year, as its food plant has recently been able to ripen to sweetness in increasing areas throughout the Carpathian Basin. The Egyptian locust: Anacridium aegyptium (Linnaeus, 1764), has been introduced into our region only occasionally, as stray specimens, but has been seen from time to time for a few years. The two leafroller moths, the Juniper Piercer: Cydia interscindana (Möschler, 1866) and the Carnation Tortrix: Cacoecimorpha pronubana (Hübner, 1799), may have been introduced directly from the Mediterranean region or even by plant detour from elsewhere. The Cotton Bollworm: Helicoverpa armigera (Hübner, 1808), for example, was extremely rare in Hungary until the mid-1980s. In a quarter of a century, the more than 20 traps of the Forestry Light Trap Network caught only 4 specimens. After that, it became increasingly common, with some traps now catching thousands of the species annually.
The Cypress Jewel Beetle: Ovalisia festiva (Linnaeus, 1767) (Buprestidae) has been introduced to Hungary twice (Figure 21). A natural immigration into the Barcs Juniper Woodland Nature Conservation Area, followed about 10 years later by the destruction of evergreen ornamental plants by another population introduced by Italian and Dutch imports. While several mud-dauber wasps species of tropical origin and one Mediterranean species (Sphecidae: Sceliphron madraspatanum (Fabricius, 1781)) invaded the Carpathian-Basin, the native species, including the most common Sceliphron destillatorium (Illiger, 1807), were largely suppressed. (Appendix C). The accelerating rate of influx of insect species of Mediterranean origin is shown in Figure 22.
4. Discussion
4.1. Hymenoptera
4.1.1. Bumblebees
A remarkable observation was made by Biella, et al. [175], explained why, contrary to expectations, bumblebee numbers are decreasing in high-mountain areas: "in high-mountain areas, climate is changing faster than the global average. This warming is harmful because it accelerates the metabolisms of ectothermic organisms and increases the activity of harmful fungi and parasites, impacting survival and fecundity in different taxa including cold-adapted bumblebees." In the Italian Alps, an increase was observed [176] in bumblebee numbers at altitudes of 1400-2000 m a.s.l. from 1994 onwards. They studied four species: the Alpine Bumblebee: Bombus alpinus (Linnaeus, 1758), Bombus mendax Gerstaecker, 1869, the Bilberry Bumblebee: Bombus monticola Smith, 1849, and Bombus mucidus Gerstaecker, 1869, none of which are typical of our region. This may explain why our results align with those of Biella et al [175] .
In Hymenoptera, such as bees and wasps, climate change has been linked to changes in population densities and community composition. Bumblebee populations in North America and Europe have been declining, partly due to climate-induced habitat loss and changes in floral resources [177,178]. Kerr and colleagues [177] highlighted that bumblebee species are failing to track warming temperatures, leading to range contractions at their southern limits without corresponding expansions at their northern limits. Arnóczkyné et al. [179] reported a general decline in Hungary, citing an increase in 7 species compared to 1953. From the 1980s onwards, we observed an increase in 6 species [6] but since 2005, only two Mediterranean species have shown an upward trend: the Blood-tailed Bumblebee: Bombus haematurus Kriechbaumer, 1870, and the Clay Bumblebee: Bombus argillaceus (Scopoli, 1763). The trend of these species is observed by other researchers from our region either [180,181,182,183,184,185].
The drastic decline of bumblebees in both number and diversity in Central Europe is described in detail by Kosior et al. [186]. The typical mountain species of the Carpathians were also detected by Šima and Smetana [187] from Slovakia, similarly from our own results from the Carpathians, but the only 527 specimens (20 species) observed during the 12-year research period (2007-2019), suggest that the bumblebee population has significantly decreased not only in the Romanian Carpathians, but also in the northern mountains of the Carpathian Basin, although high altitudes still hold a rich bumblebee fauna in relatively higher density than frequently found in the low altitudes, this is especially true for cuckoo bumblebees. From Slovakia, we have a report on the spreading of Bombus semenoviellus Skorikov, 1910. Apparently, the East-West spread of insects is not attributed to the effects of global warming. However, as the authors [188] write, certain adaptations to relatively dry and warm biotopes have been observed in this species, showing a degree of plasticity in ecological adaptations.
4.1.2. Aculeata
Similar data are scarce in the literature. In the absence of long-term data series, various models are employed to understand the relationship between Hymenoptera and climate change. These models are based on findings such as: differential species responses, the necessity for wild bees to shift to higher latitudes due to temperature increases, and the importance of examining climate change effects on habitat quality [189]. Predictions are often derived from data sets of few species, obtained from the GBIF database using the SDM software package. However, these models have limitations, as they use baselines such as RCP 4.5 and RCP 8.5 (Representative Concentration Pathways) which assume continuous warming without accounting for counter-effects like the potential shutdown of the Gulf Stream leading to drastic cooling. Therefore, computer-based projections of Hymenoptera expansions are not always accurate, as climate change is inherently chaotic, with numerous feedback mechanisms and effects.
In the absence of extensive data, the impact of climate change on Hymenoptera can be estimated through direct observation of their activity. For example, Raider et al. [190] studied 20 species of American wild bees, comparing their temperature-dependent activity to that of Apis mellifica, thereby establishing a heat tolerance ranking for some wild bee species.
To our knowledge, only Belgium has produced a similar dataset [191]. Despite the different climatic conditions (Belgium - Atlantic, Central Europe - continental), the dynamics of wild bee populations in both regions show a similar upward trend (Compare Figure 4 and Figure 23).
Megachilid species such as Megachile flavipes Spinola, 1838, the Sicilian Mortar Bee: Megachile sicula (Rossi, 1792), the Red-tailed Cuckoo Leafcutter: Coelioxys coturnix Pérez, 1884, Megachile minutissima Radoszkowski, 1876, and Osmia submicans Morawitz, 1870, have been studied [192], It is recognised that many species within the family Megachilidae are sensitive to warming, including several euryecious hylophyllous species. However, predicting the distribution of these species 80 years in advance is precarious, as climate change processes are not unidirectional. This complexity is why the term "climate change" is now preferred over "global warming," reflecting the intricate feedbacks that challenge our ability to make reliable long-term predictions. On the other hand, the observation of the authors well agrees our results with hylophilous Megachilid bees. The work of Tryjanowski, et al. [193] is a crucial complementary publication, as it addresses common species that are underrepresented in our data sets. They found no significant trend in the abundance of German wasp: Vespula germanica (Fabricius, 1793). Climate change has not had an impact on the species' populations so far. This species is appearantly resistent to the climate change up to this date. Eusocial Vespidae were studied by Pawlikowski and Pawlikowski [194] between 1981 and 2000. Overall, the wasp populations proved to be stable in this period. Specifically, the Common Wasp: Vespula vulgaris (Linnaeus, 1758) showed a significant increase over 20 years, from 713 to 1,556 individuals; the Red Wasp: Vespula rufa (Linnaeus, 1758) increased from 8 to 23; and the Median Wasp: Dolichovespula media (Retzius, 1783) from 0 to 12. The Tree Wasp: Dolichovespula sylvestris (Scopoli, 1763) showed a decrease from 11 to 6, and the European Hornet: Vespa crabro Linnaeus, 1758 from 212 to 182. Vespula germanica numbers stagnated at 11,540 versus 11,352 specimens. However, European hornet has definitely increased its populations in deciduous forests, particularly oak mixt forests, in Romania, based on our observations in the Moldova and Dobrogea regions. Overall, apart from a few forest species, climate change has had a beneficial or neutral impact on social wasps.
The observed increase in cuckoo wasps may be attributed more to the rise in their hosts than to a direct response to climate change. This is supported by Verheyde, [195], who stated that after more than 80 years of the vespid wasp Euodynerus dantici (Rossi, 1790) being present, its main parasitoid, the cuckoo wasp Chrysis sexdentata (Christ, 1791), was reported for the first time in both the Netherlands (2019) and Belgium (2022). They mentioned that the main reason for these unexpected reports was the densification of available host populations driven by climate change over the past decade.
The increase in the number of Aculeata shows trends opposite to those observed in several Western European countries, particularly Germany [196]. While warming is evidently favorable for most Aculeata, it does not account for the drastic decline in the German Hymenoptera population. This discrepancy may be explained by the fact that, for instance, in Hungary, pesticide use reached 7 kg of active ingredients per hectare of agricultural area by 1989. Following the political turn, pesticide use dropped to just 1.4 kg in 1995 [197].
4.1.3. Sawflies, Symphyta
Overall, sawflies have suffered a significant decline in our region in the last decades, being mainly a northern, Scandinavian group, with their greatest species richness in the Scandinavian areas. It can also be observed that species of the Carpathian Basin, once sporadic in the more marshy regions, now below the detection limit, find shelter in high altitudes, in contrast to species of the Diptera and Aculeata, where vertical dispersal is more limited [6]. Among the genera that produce growth, there are mainly those that reach the highest species numbers in the tropics and Mediterranean regions, these are species of the family Argidae [198]. In the Mediterranean and Anatolian regions, species of the family Megalodontesidae and Tenthredo distinguenda (R. Stein, 1885) are also the commonest in the Mediterranean region with an Anatolian distribution either [199]. The most common Dolerus species are warm-loving to some extent, but are beginning to decline or even disappear in the Mediterranean region [200]. A large part of the literature deals with the climatic sensitivity of species of the family Diprionidae and Pamphiliidae [201,202]. Species of these families are not present in our region in numbers sufficient to allow their population increase or decline to be assessed statistically. Their outbreaks have been reported from time to time, but we have not observed this during our field works.
4.2. Diptera
4.2.1. Bombyliidae
In the literature, decades-old Bombyliiidae data are not available; however, the available ecological observations support our findings. Bombyliid species are typically and most frequently encountered in arid areas and constitute a high percentage of the diversity of flies in the more or less desertic regions of the earth [203]. Adult bee flies are frequently the primary pollinators of a wide variety of flowering plants, particularly in arid environments [204]. Their hosts are typically insects from warm, dry areas such as Acrididae, Formica spp., Myrmeleontidae, Tenebrionidae, Sphecidae, as well as Noctuidae, Pyralidae, Gelechiidae, Scarabaeidae, etc. 63% of the world's species, including the most common ones, parasitize xertolerant hosts like bees and wasps, locusts, and grasshoppers [205]. Since these animals mostly have xerothermic ecological requirements, we have not found a decrease in their population numbers according to the current extent of climate change. At present, they are among the winners of climate change. No similar data sets have been published by others. Herrera studied the population dynamics of pollinators in the Mediterranean for over 20 years [206]. Among the observed species, a Bombyliidae genus, Hemipenthes (Bombyliidae), exhibited a significant increase in populations.
4.2.2. Horse-Flies, Tabanidae
There is no time-series analysis available in the literature for this group, yet the work of Havkenberget and colleagues [207] is interesting from this perspective because they describe and prove that both species richness and individual density are dependent on different ecotypes. Studying the Tabanid fauna on both sides of the Velika Kapela mountain range (Croatia), they found that species occupied either the southern slope (Mediterranean zone) or the northern slope (Continental zone) according to their ecotype, and similarly occupied different altitudinal zones. In our study, the Tabanids responded differently to changing climatic conditions rather than to slopes and altitudinal zones, depending on the ecotypes they belong to. Herczeg, T. and colleagues [208] also examined ecological and climatic factors in terms of Tabanid distribution and proliferation. They studied six species and observed that the 31–35°C range was when flight activity was most vigorous, i.e., the traps captured the most individuals. As warmth-loving species, we also observed increasing captures in most species following the warming trend. Haematopota subcylindrica Pandellé, 1883 is a moisture-loving forest species [36]. In our case, the population size of this species showed a downward trend, while other xerothermic species showed a decline according to their ecological requirements. Climatic changes do not justify the decrease in the population size of Tabanus bromius Linnaeus, 1758. This decline was first observed in Great Britain, where it was found that it is now a local species in Britain which has declined substantially, possibly as a result of the widespread ploughing and destruction of meadows [209].
4.2.3. Hoverflies, Syrphidae
Hallmann et al. [210] found a decrease of the number of individuals of hoverflies of 89 % and a decrease in species richness of 23 % over a 26-year period (1989-2014) for a river valley in Germany. Barendregt et al. [211] reported similar results, a decrease of 80 % of the number of individuals of hoverflies over 40 years (1982 – 2021) and 44 % of the species over 43 years (1979–2021) for hoverflies in a forest in the Netherlands. Gatter et al. [212] report for migratory hoverflies a decline of individuals of 90–97 % over 40–50 years (based on transect counts 1970–2019 resp. Malaise traps 1980 -2019). Finally, comparing the period 2008-2022 with 1900–1969, Reemer et al. [213] found as many species of hoverflies with a declining trend (147 species) as with an increasing trend (146 species).These studies describe strong decline of both the number and the species richness of hoverflies since 1980 in western Europe. The current (1993–2017) extinction rate for hoverflies is estimated to be nearly five times higher than the historical rate (1942–1992) [214]. Longer warmer and drier periods will harm most hoverflies, but especially the aphidophagous larvae of zoophagous species, because aphids are highly sensitive to drier conditions. In addition, there may be a disconnect in phenological timing between aphid and hoverfly life-cycles. Warmer winters can make it harder to overwinter successfully in a dormant state, particularly if breaking diapause requires a cold spell. Also, potentially, warmer winters could promote fungal infections or increase predation during the winter [215].
4.2.4. Tachinid Flies, Tachinidae
Zeegers [216] examined six Tachinidae species in the Netherlands for climate sensitivity. 'The tachinids Brullaea ocypteroidea Robineau-Desvoidy, 1863, Entomophaga exoleta (Meigen, 1824), Istocheta longicornis (Fallén, 1810), Phasia aurigera (Egger, 1860), Senometopia intermedia (Herting, 1960), and Thecocarcelia acutangulata (Macquart, 1850) are recorded for the first time for the Netherlands, bringing the number of Tachinid species recorded from the country to 331. Most of these species have a southern distribution in Europe, recently extending northwards due to climate change. Similarly, many Tachinid species exhibit this northward spread: 'Four of these species show recent expansions in their range. The discovery of Leucostoma abbreviatum Herting, 1971, near Templin provides the first records for this species in the province of Brandenburg, and Elomya lateralis (Meigen, 1824) has been found for the first time in Sachsen-Anhalt, Brandenburg, and Mecklenburg-Vorpommern. Both species were known only from the south and have now been found in northeast Germany, more than 600 km north of the previous limit of their distribution. Two other bug parasitoids, Phasia barbifrons (Girschner, 1887) and Phasia aurigera (Egger, 1860), are currently expanding their range towards the north from southern and central Germany [217] . Many species of Tachinid flies possess relatively large host ranges, and the frequency of parasitism by tachinids exhibits no relationship with climatic variability among the data sets that estimated their frequency. Thus, the population changes of Tachinids are unpredictable [218]. Our findings also support this uncertainty. While many species show a steady decline, we observed a significant increase at the end of the study period compared to the beginning in species such as Linnaemya frater (Rondani, 1859), Meigenia dorsalis (Meigen, 1824), and Exorista larvarum (Linnaeus, 1758). The family should not be treated as a single ecological unit. Each species has different ecological needs, and various ecotypes are present within the family. Despite the declining trend, we see that most species have been able to adapt to the changed environmental conditions. We must also acknowledge that under changing conditions, the population size of Tachinidae species is indeed unpredictable.(Table 8 and Table 9). Thus, our detailed data sets reinforce the opinion of Stireman and colleagues [218].
4.3. Lepidoptera
4.3.1. Butterflies, Rhopalocera
Parmesan and his colleagues [1] studied the northward shift in the areal distribution of butterfly species in the British Isles. Our results are challenging to compare due to the minimal number of common species. Northward shifts have been observed in the Clouded Apollo: Parnassius mnemosyne (Linnaeus, 1758), the Green-underside Blue: Glaucopsyche alexis (Poda, 1761), the Small Pearl-bordered Fritillary: Boloria selene (Denis & Schiffermüller, 1775), the Niobe Fritillary: Argynnis niobe (Linnaeus, 1761), and the Purple Emperor: Apatura iris (Linnaeus, 1758). The distributions of the Scarce Copper: Lycaena virgaureae (Linnaeus, 1758) and the Southern White Admiral: Limenitis reducta (Staudinger, 1901) have not significantly changed. The Lesser Marbled Fritillary: Brenthis ino (von Rottemburg, 1775), however, has spread southwards. In Central European conditions (Table 11 and Table 12), the population of the Small Pearl-bordered Fritillary increased significantly. The abundance of the Niobe Fritillary is relatively stable but fluctuating. In contrast, the Green-underside Blue, the Lesser Marbled Fritillary, and the Purple Emperor have significantly declined, aligning with Parmesan's findings in Central Europe [1]. The populations of the Scarce Copper and the Southern White Admiral showed stability. Our results corroborate Parmesan et al.'s conclusions [1] regarding the climate sensitivity of butterfly species. Exceptions include the Small Pearl-bordered Fritillary, which is classified in the lowest climate sensitivity category (PR) [71], and species of the Papilionidae family, including the Clouded Apollo (Figure 24), which appear stable in our region, with some species showing an increase.
According to the Environmental Statistics and Reporting team at the Department for Environment, Food & Rural Affairs [219], of the 26 butterfly species surveyed, 7 species increased, 12 decreased, and the rest remained statistically unchanged. Similarly, in our results, of the 33 species surveyed, 6 species increased, 15 decreased, and the rest remained unchanged (Table 11). The two studies had few common species due to differences in zoogeography. Among the 5 common species, the Small Pearl-bordered Fritillary exhibited opposite trends, increasing in the Carpathian Basin and decreasing in the British Isles. The Purple Emperor decreased in the Carpathian Basin, while their numbers increased dramatically in the British Isles. The Small Tortoiseshell: Aglais urticae (Linnaeus, 1758) declined in all areas, drastically in the Carpathian Basin. The Old World Swallowtail: Papilio machaon (Linnaeus, 1758) remained stable everywhere. The Marsh Fritillary: Euphydryas aurinia (Rottemburg, 1775) increased in the Carpathian Basin and remained stable in the British Isles. In 2008, Panigaj and Panigaj [220] re-surveyed the butterfly fauna of Temnosmrečinská dolina in the High Tatras. According to their results, fifteen of the originally recorded species were not found; however, the occurrence of six new species was recorded. Similar studies are reported by our team [6] based on the files studies of the Hungarian natural History Museum and independent authors. A 35-68% decrease in species abundance was detected in different areas (Simontornya, Bátorliget, Drávasík). The problem with these studies is that the collection method and frequency were different but in that time there was no other sources. In the Kaposvár collection, we have regular records of 33 species made by the same collector, Dr. Levente Ábrahám, over three decades (Table 11). Here, we have seen that out of the initial 33 species, the density of 6 species has fallen below the detection level. At the level of total Carpathian Basin, 9 species have fallen below the detection level in the last 24 years, and no detections of these species have been made in the lower parts of the Carpathian Basin since then.
Settele et al., in their monograph Climatic Risk Atlas of European Butterflies [71], predict a decline of various butterfly species by 2050 and 2080, respectively, and a shift to northern areas based on the situation in the year 2000. We are now halfway through the first period. Obviously, the decline of a large number of butterfly species seems to be the realization of the authors' predictions. As stated in Parmesan, C.; Ryrholm, N.; Stefanescu, C. et al.: “Mean global temperatures have risen this century, and further warming is predicted to continue for the next 50–100 years [1].” The basic assumption of these and hundreds of similar predictions is that climate change equals global warming. However, climate change does not imply a general rise in temperature but rather weather disturbances, strong warming in some places, strong cooling in others, and ultimately the collapse of global ocean transport systems. Rather, abrupt regional cooling and gradual global warming may occur simultaneously. In fact, greenhouse warming is one of the destabilizing factors that makes abrupt climate change more likely. According to a 2002 report by the US National Academy of Sciences (NAS), available evidence suggests that abrupt climate changes are not only possible but likely in the future, with potentially large impacts on ecosystems and societies. In extreme cases, indefinite warming could turn into the opposite (glacial) due to a breakdown of thermohaline circulation [159,160].
Numerous papers study the changes in the population of particular butterfly species. Birch, R. J.; Markl, G.; and Gottschalk [221] discuss the adaptation and population growth of the Great Banded Grayling: Brintesia circe (Fabricius, 1775) (Figure 12) in the context of climate change. The increase in the Dryad: Minois dryas (Scopoli, 1763) under suitable conditions (nature conservation) is also well documented [222]. We have observed similar trends in the Danube-Ipoly National Park area. Hudák [223] noted an increase in the following species: the Black Hairstreak: Satyrium pruni (Linnaeus, 1758), the Scarce Large Blue: Maculinea teleius (Bergstrasser, 1779), the Brown Argus: Aricia agestis (Denis & Schiffermüller, 1775), the European Beak: Libythea celtis (Füssly, 1782), the Knapweed Fritillary: Melitaea phoebe (Denis & Schiffermüller, 1775), the Lesser Purple Emperor: Apatura ilia (Denis & Schiffermüller, 1775), the Pearly Heath: Coenonympha arcania (Linnaeus, 1761), the Dryad: Minois dryas (Scopoli, 1763) , and the Great Banded Grayling: Brintesia circe (Fabricius, 1775). This largely aligns with our observations, but other species with strong population growth in our area are not included in his work, as they were the most common species even in the past. Consequently, their population changes are rarely investigated.
Fox et al. [224] examined 58 Butterfly species, from two aspects: the change in abundance and the change in distribution. We only examined the change in abundance. On the one hand, we examined 33 selected species similarly to the method of British authors) (Table 11). On the other hand, the total butterfly fauna of a given area (National Park) which is 111 species was investigated by our team as well. In this case, the area was limited, but the number of species covered the total species richness of a given area (about 600 km2). Our data are not comparable, because while the aim of British researchers is to determine the status and change of the entire butterfly fauna, i.e. to examine the influence of all environmental factors. In contrast, we tried to eliminate all other factors influencing the number of individuals (like farming, pesticides, landscape destruction, isolation of habitats, etc.) and focus only for the effect of climate change. This is the reason that our results are different. Instead of the general decline of abundance, we observed outbreak of common and frequent butterflies (these butterflies proved to be more or less climate change tolerant), and mostly decline of rare and sporadic butterflies which are sensitive for climate changes.
4.3.2. Moths, Nocturnal Macrolepidoptera
According to: the prediction of Uhland colleagues [225], climate change, with hot summers might exceed the larval temperature optima and drought reduces food plant quality. Increasing frequency and severity of temperature and drought extremes due to climate change, therefore, might amplify insect decline in the future. Therefore, summer-developing larvae seem particularly sensitive. Changes in insect diversity caused by climate change might not be relevant in the Mediterranean area only, where summer heat might exceed the species’ physiological optima but also in Central Europe where optima will likely be exceeded in the future.
Our results also show some decline in Central Europe, but this decline is not significant at this time just becuse the decline of numerous species is compensated by increased frequency of outbreaks of some common species, also due to the gradually warming climate (see Table 17 and Figure 14). As for future projections, we are cautious to make any statement. Although the trends are clear, we should not forget the counter-effects, such as the possible shutdown of the Atlantic Meridional Overturning Circulation (AMOC) or Gulf Stream, which could lead to significant cooling. In Sweden, Betzholtz and colleagues [226] studied Catocala nupta Linnaeus, 1767, Noctua interjecta Hübner, 1803, Noctua interposita (Hübner, 1790), Mythimna albipuncta Gaede, 1916, Eucarta virgo (Treitschke, 1835), Watsonalla binaria (Hufnagel, 1769), Pseudeustrotia candidula (Denis & Schiffermüller, 1775) and Eupithecia pulchellata Stephens, 1831. According to their results, the populations of range-expanding moths in southeastern Sweden have increased both in abundance and species richness over 16 years and that their potential for rapid population growth seems to be favoured by a warming climate. We studied two of these species in Central Europe, with stable populations of Catocala nupta (Linnaeus, 1767) and a sharp decline of Pseudeustrotia candidula (Denis & Schiffermüller], 1775), apparently due to climatic differences between these 2 regions.
According to Sparks et al.,[227] the number of migratory Lepidoptera (moths and butterflies) species reported annually at a site in the south of the UK has been steadily increasing. This rise is strongly linked to rising temperatures in Southwestern Europe. It is anticipated that further climate warming in Europe will increase the number of migratory Lepidoptera reaching the UK, necessitating urgent attention to the consequences of this invasion. In our region, as it is not on the border of the distribution of these species, they do not show migratory behaviour. However, we have studied the variation in the number of individuals of certain migratory species. In our area, the increase in the number of species is not very pronounced, but the expansion of one species of migrant moth, namely Helicoverpa armigera and its increase causes significant damage to agricultural crops and here it is the main concern among migrant moths. Other migratory moth species such as Hyles livornica Esper, 1780, Acherontia atropos Linnaeus, 1758 did not show an increasing trend. For Daphnis nerii Linnaeus, 1758, there is a definite positive trend (number of observations: 2014: 1 specimen, 2018: 8 specimens, 2019: 8 specimens, 2020: 13 specimens, 2021: 3 specimens, 2023: 23 specimens, 2024: 25 specimens) [228].
On the other hand, the increased population density predicted by the British authors [227], in the form of densifying gradients (Table 17, Figure 14 and Figure 15), have already arrived to our region in form of reduced species diversity and increased frequency of population outbreaks.
In Central-Europe, Szabó,, Árnyas, and Varga [171] measured an increasing trend in the number of individuals between 1997 and 2004, according to their conclusion: arid weather conditions were favorable for the abundance of species tend to outbreaks and living in deciduous forests. The highest species richness was measured in the last year of their study series. Varga and colleagues [172] from 2019 to 2022, showed a decreasing trend in both species number and number of individuals. Furthermore Szentkirályi et al. from Várgesztes experimental site [173], between 1962 and 1999, published a significant decrease in both parameters and observed outbreaks of species susceptible to outbreak in dry, drought years. According to the observation of Uherkovich [229] there were a significant decline in species diversity, which was a clear consequence of the rapid climate change of the last decades. There was also a decrease in the overall abundance of macrolepidoptera, which was clearly seen through his recent samplings from the 2020s. In the Subcarpathian region, there was not visible impoverishment of the general species diversity within 100-year retrospective [62]. Szentkirályi and his colleagues [62,171,172,173,174] reviewed the results of a 50-year light trap network. Their results proved that there was a definitive, significant decreasing trend of moth assemblages in time series at certain trap stations but not at all stations .
Hufnagel and Sipkay [230] analyzed data from 8 light traps between 1962 and 2006. When examining the total number of species and individuals per year, they found that the number of species captured per year remained relatively stable, despite significant decline in the number of individuals. We extended this examination, including a significant number of additional traps, covering the period between 1970 and 2022. The key difference was that the decline in the number of individuals was hardly observable and not statistically significant, (although our data detected the same decline in the early 2000s). Concurrently, the number of species caught by each trap decreased (Table 18 and Figure 16). Post-2006, the warming trend persisted, leading to the disappearance of some species from the light traps (their numbers fell below the detection limit locally). Meanwhile, for other species, as described by [171,172,173,174], the outbrakes of some moth species became more frequent and intense.
In our results, in the lower Carpathian Basin over 50 years contain the main conclusions of the above articles. There is a decline in numbers, but this decline is not significant in the long term (r2=0.01), as it is more or less compensated by the outbreaks of some common species due to the increasing frequency of dry periods since the beginning of the warming period in the 1980s, (r2=0.13). This increase in the number of individuals is unfortunately associated with a decrease in species diversity per light trap. At the same time, the number of species that disappeared from the entire Carpathian Basin during the last 20 years is small (Table 16).
In addition, we have long data series from Great Britain. According to the Butterfly Conservation Organisation [231]: The total abundance of larger moths caught in the RIS light-trap network in Britain decreased by 33% over 50 years (1968–2017). Losses were greater in the southern half of Britain (39% decrease) than in the northern half (22%). It is very different from the situation in Central Europe. Our experience is confirm the results of the Ukrainian and Hungarian authors [62,171,172,173,174], as more frequent outbreaks during the warming period, compensate for the decrease in abundance furthermore the largest number of moth-capture in 50 years, was taken in 2022 (Table 17 and Figure 14 and Figure 15). We also detected regional species impoverishment per trap..(Table 18 and Figure 16), but this was not associated with species impoverishment across the whole region (Table 16). On the other hand, the replacement of dominant species was the strongest indicator of climate change (Table 19).
4.4. Alien and Invasive Species, Mediterranean Influx
The most comprehensive study to date on invasive species in our region (and Europe) is the monograph "Alien Terrestrial Arthropods of Europe" by Roques and colleagues [76], published in the Biorisk series. This study analyzes approximately 330 species from our area, the Carpathian Basin, up to 2009. Building on this work and thoroughly reviewing the relevant literature, we expanded this to 803 species. One part of the remaining 473 species have been introduced or invaded since 2009, and the other part were identified through a more in-depth examination of the literature, having been omitted from the mentioned work. We compared our findings with similar research results from nearby and distant regions.
Solarz and colleagues [232] in Poland, assessed 60 non-native plant species and 58 animal species. For seventy-nine species, they evaluated climate change as a factor that increases the likelihood of future introduction, establishment, and spread. Climate change increased the number of high-risk invasive alien species from 38 to 63. Species from warmer parts of the world were the most sensitive to climate change. We examined 803 species based on their origin and time of appearance, with a particular focus on the climate change situation. Of the 803 incoming species, 174 are from tropical areas. Among these, 74 tropical species (43%), entered the Carpathian Basin in the last quarter-century (ábra) , along with 86 Mediterranean-origin species (ábra). This high rate of influx and establishment in recent decades is largely driven by climate warming. This is supported by Gutierez and colleagues' monograph on the influx of alien species and its interpretation from an agricultural perspective, stating: whether the introduction of a new species becomes invasive or not partly depends on the biological and physical characteristics of the habitat into which it is introduced. These habitat characteristics are largely influenced by climate, which is altered by human activities [233].
Gutierrez and Ponti [233] report a phenomenon in North America similar to the Mediterranean influx we experience, using examples of four pests: the New World Screwworm Fly: Cochliomyia hominivorax (Coquerel 1858), the Pink Bollworm: Pectinophora gossypiella (Saunders, 1844), the Mediterranean Fruit Fly: Ceratitis capitata (Wiedemann, 1824), and the Olive Fruit Fly: Bactrocera oleae (Rossi, 1790). These species first appeared in California and are spreading northward and southward towards Mexico due to temperature changes. Their findings suggest that the primary driver of northward spread is temperature increase, while southward spread is facilitated by adequate moisture levels. Consequently, their forecasts indicate that the arid regions of Arizona and northern Mexico are more protected, whereas the subtropical areas of California and western and southern Mexico are more susceptible to infestations. Since desertification in Central Europe is currently limited to small areas but increasing (Kiskunság: Homokhátság in Hungary), future studies on the effects of desertification would be important. However, due to a lack of financial resources, these studies are stopped for a while (Fülöpháza, Kiskunság National Park, light trap project).
According to the IUCN's forecast [234], the number of alien species will increase by 36% between 2005 and 2050. However, in our region, we have surpassed this prediction, with the number of insect species alone increasing by 54% over the past 24 years compared to all previous years. Of these, 86 are Mediterranean and 74 are tropical species (figures references). Thus, in this century, out of the 298 incoming species, 160 (54%) have arrived from warm (Mediterranean, tropical) regions, indicating that climate change is a major driver of this influx.
Out of the 1,390 alien insect species established in Europe [76], our current findings show that 803 live in the Carpathian Basin, which is 58% of the 1,390 species reported from Europe. Specifically, this includes 164 of the 398 non-native Coleoptera species (41%) established in Europe, 303 of the 318 Hemiptera species (95%), 88 of the 297 Hymenoptera species (30%), 99 of the 97 recorded Lepidoptera species (102%), and 65 of the 98 Diptera species (66%). The total insect influx in Europe can be described by the curve 15.40 exp(0.50x) r²=0.87, while in our region it is not so steep: 5.86 exp(0.49x) r²=0.84. With a linear approximation, the intensity of alien species influx in Europe is 67.75 (lin. x coefficient, slope of the line) r²=0.86, whereas it is more moderate in the Carpathian Basin, with an intensity of 36.82 (lin. x coefficient) r²=0.77.
From a biogeographic perspective, the two studies are not fully comparable because we separately treated the Mediterranean-origin species, as different drivers (including climate change-induced influx) primarily govern them. Additionally, instead of Asia, we separately considered the Eastern Palaearctic and Oriental regions, as these are distinct units from climatic and zoogeographic point of view. It is evident that for a more accurate assessment of the influx rate, comparing 2024 data with European data from 2010 is not advisable. It indicates the necessitates a deeper analysis of the European situation and the update of the data of the European monograph.
The influx of alien species in other regions also currently shows an exponential trend. In Eastern Russia, the equation for the influx trend is quadratic: AS = exp (-5.73 + 0.000002536* Years^2) or e−5.73+0.000002536×Years2 (xsquared) [235].
Similar data are reported from China [236], where we observe an exponential increase again. The influx of alien species correlates with China's GDP. This correlation is also evident when it is split up by provinces, as the increase in freight transport and GDP are closely linked. In Central Europe, this is not necessarily the case, as the region does not show similar economic growth compared to China; however, it is still highly susceptible to the influx of alien species (Appendix C). It is evident that alien species enter through the ports and rail yards of economically stronger EU states and then spread rapidly across the continent. Therefore, in Europe, there is no such strong correlation between economic development and the influx of alien species. The ranking of European countries by non-native species as follows: Italy, France, Great Britain, Spain, and the Czech Republic [76]. Globally, the most heavily infested countries and regions by infection of non native species are Cuba, Great Britain, France, Taiwan, Florida, Texas, California, Hawaii (USA), Queensland, and Victoria (Australia) [237].
Another significant driver of the establishment of alien insects is the influx of alien plant species. Plants that have established or been introduced in Europe show an exponential trend similar to that of insects [238]. In this context, our region is positioned in the middle of the ranking (at the boundary of the upper and lower 50%).
5. Conclusions
The wildlife and vegetation of Central Europe, including the Carpathian Basin, is undergoing transformation due to four main factors: the retreat of species that require cool and wet habitats, the proliferation of species that thrive in warm and dry conditions, the northward migration of Mediterranean species, and the gradual establishment of species introduced from the tropics.The proportion of species arriving from tropical and Mediterranean regions has increased significantly, with their numbers rising by 58% over the past 24 years compared to all previous years. Of the 298 species that have arrived in this century, 160 (54%) are from warm regions (Mediterranean and tropical), including 86 Mediterranean and 74 tropical species. This indicates that climate change is a major driver of transformation.
Among native insect species, hoverflies (Syrphidae) have suffered the most significant population decline, consistent with European and global trends. The population of Tachinidae species has also decreased, although less markedly than hoverflies. Conversely, species from xerothermic families, such as beeflies (Bombyliidae) and horse-flies (Tabanidae), have shown population increases up to the current extent of warming. However, since horsefly larvae are associated with wet or aquatic environments, this trend is expected to stop if warming continues.
The warming climate has proven to be favorable for most of the Aculeata Hymenoptera, which is logical given that they achieve their highest species diversity and population numbers in the Mediterranean region. Alongside horse-flies and bee flies, increase of this group clearly demonstrate the Mediterranean transformation. However, several groups have shown significant declines, including bumblebees, Megachilid bees, and certain Crabronidae species that require cool and wet habitats (forests, marsh meadows). The population decline of bumblebee species is more complex and likely only partially caused by warming. This issue requires further investigation.
In the case of butterflies, a decline in species richness is observable in certain areas. Some previously common species have proven resistant to the current extent of climate change, showing intense reproduction under protected conditions. However, most previously sporadic and rare species have been unable to adapt to the changed climate conditions, resulting decline in their population numbers frequently even falling below the detection level regionally.
The population of nocturnal moths has entered a growth phase over the past 10 years. This growth phase has been accompanied by a decrease in species diversity. This population growths are caused by the more frequent outbreaks of some common species.
Only a smaller proportion of species find refuge in higher regions. According to our current findings, this includes 20 out of 87 Diptera species, 32 out of 71 Aculeata Hymenoptera species, and 15 out of 23 Lepidoptera species, which have not been collected in the lower regions for over 20 years. We do not and cannot attempt to make forecasts, as several predictions suggest that current trends may reverse, and our climate may become even more chaotic if ocean currents potentially collapse. Finally, we, as individuals, when we can do nothing more, should never lose hope in the cohesive power of communities, even in such challenging circumstances either.
6. Patents
This section is not mandatory but may be added if there are patents resulting from the work reported in this manuscript.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Hymenoptera, Diptera, Lepidoptera, non-native species, analysis Attila Haris; Aculeata, Zsolt Józan.; Diptera, Sándor Tóth.; bumblebees, Aculeata Peter Šima.; bumblebees, Aculeata, Dipetra in part Bogdan Tomozei; Lepidoptera, Peter Scmidt; Lepidoptera, Gábor Glemba; Invaziv and non native species, Lepidoptera, György Csóka; Invaziv and non native species, Lepidoptera, Anikó Hirka; statistical analysis, Attila Haris; resources, All authors.; data curation, All authors; writing—original draft preparation, Attila Haris; All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
Authors express their grateful thanks to Dr. Vladimír Smetana (Tekov Museum, Levice, Slovak Republic), Dr. Levente Ábrahám (Rippl-Rónai Museum, Kaposvár, Hungary), Dr. Ákos Uherkovich, (Janus Pannonius Museum Pécs, Hungary), Attila Huber (Aggtelek National Park, Aggtelek, Hungary), Prof. Dr. Zoltán Varga (University of Debrecen, Debrecen, Hungary). for their generous supports, data provisions and advises.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature. [CrossRef]
- Roy, D. B.; Sparks, T. H. Phenology of British butterflies and climate change. Global Change Biology 2000, 6(4):407-416.
- Shuman, E. K. Global Climate Change and Infectious Diseases. Int. J. Occup. Environ. Med, 1: 2(1).
- Patz, J. A.; Epstein, P. R.; Burke, T. A.; Balbus, J. M. Global climate change and emerging infectious diseases. Jama 1996, 275(3), 217–223. [Google Scholar] [CrossRef] [PubMed]
- Team eBird European Bee-eaters Expand Their Range Northwards. Last modified June 10, 2022. Available online: https://ebird.org/news/ebird-impacts-european-bee-eaters-expand-their-range-northwards. (accessed on 11. 10. 2024).
- Haris, A.; Józan, Z.; Roller, L.; Šima, P.; Tóth, S. Changes in Population Densities and Species Richness of Pollinators in the Carpathian Basin during the Last 50 Years (Hymenoptera, Diptera, Lepidoptera). Diversity 2024, 16, 328. [Google Scholar] [CrossRef]
- European Environment Agency (EEA) Biogeographical regions in Europe. Available online: https://www.eea.europa.eu/data-and-maps/figures/biogeographical-regions-in-europe-2 (accessed on 19. 11. 2024).
- Führer, E. A klímaváltozáshoz alkalmazkodó erdõgazdálkodás kihívásai — III. ( Challenges for forest management adapting to climate change – III.). Erdészeti Lapok.
- Gálos, B.; Jacob, D.; Mátyás, Cs. . Effects of Simulated Forest Cover Change on Projected Climate Change – a Case Study of Hungary. Acta Silv. Lign. Hung, 7.
- Sharma, H. C. Biological Consequences of Climate Change on Arthropod Biodiversity and Pest Management. In: International Conference on Insect Science, Bangalore, India, 14-. 17 February.
- Karuppaiah, V.; Sujayanad, G. K. Impact of Climate Change on Population Dynamics of Insect Pests. World Journal of Agr. Sci. 2012, 8(3), 240–246. [Google Scholar]
- Helmholtz-Zentrum für Umweltforschung – UFZ Klimawandel und Biodiversität. Available online: https://www.ufz.de/index.php?de=37140 (accessed on 12. 10. 2024).
- Bauer, T.; Wiblishauser, M.; Gerlach, T. (2022): Wärmeliebende Insekten als Zeiger des Klimawandels – Beispiele und Potenziale bürgerwissenschaftlicher Arterfassungen. Anleigen Natur 2022, 44(1), 141–148. [Google Scholar]
- Bebber, D.; Ramotowski, M.; Gurr, S. Crop pests and pathogens move polewards in a warming world. Nature Clim. Change. [CrossRef]
- Rannow, S.; Neubert, M. Managing Protected Areas in Central and Eastern Europe Under Climate Change. In Ser. Advances in Global Change Research, S: Beniston, M., Publisher, 2013. [Google Scholar]
- Szabics, A. Hová folynak el vizeink – a vízhiány okai Magyarországon (Where does our water go – the causes of water shortages in Hungary). Available online: https://www.mnb.hu/letoltes/szabics-andras-zsolt-hova-folynak-el-vizeink-a-vizhiany-okai-magyarorszagon.pdf (accessed on 14. 10. 2024).
- Reich, Gy. Nemzeti Vízstratégia (Kvassay Jenő terv) (National Water Strategy, J. Kvassay Plan). Ludovika University of Public Service, Budapest, Hungary, 2019, pp. 1-57.
- World Bank Group (WBG) Climate Change Knowledge Portal. Available online: https://climateknowledgeportal.worldbank.org/ (accessed on 24. 10. 2024).
- Pittioni, B. Die klimaökologische Formel als Hilfsmittel der biogeographischen Forschung. Wetter u. Leben, 1949, 2, 161–167. [Google Scholar]
- Pittioni, B. Das Problem der Formenbildung. Ein Deutungsversuch mit Hilfe der klimaökologischen Formel. Bonner zool. Beitr, -2.
- Hagen, M. D. Zur Schwebfliegenfauna des Raumes Hagen (Diptera, Syrphidae). Abh. Westfäl. Prov.-Mus. Naturk.
- Rasmont, P. , Franzén M., Lecocq T., Harpke A., Roberts S. P. M., Biesmeijer, J. C., Castro, L.; Cederberg, B.; Dvořák, L.; Fitzpatrick Ú.; Gonseth, Y.; Haubruge, E.; Mahé, G.; Manino, A.; Michez, D.; Neumayer, J.; Ødegaard, F.; Paukkunen, J.; Pawlikowski, T.; Potts, S. G.; Reemer, M.; Settele, J.; Straka, J.; Schweiger, O. Climatic Risk and Distribution Atlas of European Bumblebees. Biorisk.
- Lukáš, J.; Tyrner, P. Zlatěnky (Hymenoptera: Chrysididae) Státní přírodní reservace Devínská Kobyla. Klapalekiana 2000, 36, 113–123. [Google Scholar]
- Vepřek, D. První doplněk Check list of Czechoslovak Insects III (Hymenoptera: Sphecidae). Sbor. Pri. Kubu Uher. Hrad. 2000, 5, 233–239. [Google Scholar]
- Bogusch, J.; Straka, J.; Kment, P. Annotated checklist of the Aculeata (Hymenoptera) of the Czech Republic and Slovakia. Acta Ent. Mus Nat Pragae. Suppl. 2007, 11, 1–300. [Google Scholar]
- Deván, P. Kutavky (Sphecidae), hrabavky (Pompilidae), zlatenky (Chrysididae) NPR Tematínska lesostep na lokalite Lúka a v PR Kňaží vrch (Považský Inovec, západné Slovensko) získané Malaiseho pascou v rokoch 1999 - 2000. Nat. Tut. 2004, 8, 143–151. [Google Scholar]
- Smetana, V. Čmele a spoločenské osy (Hymenoptera: Bombini, Polistinae et Vespinae) v poľnohospodárskej krajine Poľany a Podpoľania. Nat. Tut. 2009, 13(1), 107–114. [Google Scholar]
- Smetana, V. Výsledky prieskumu vybraných skupín blanokrídlovcov (Hymenoptera: Aculeata) na Ramsarskej lokalite Poiplie. Acta Mus. Tek. 2010, 8, 71–77. [Google Scholar]
- Smetana, V.; Roller, L.; Beneš, K.; Bogusch, P.; Dvořák, L.; Holý, K.; Karas, Z.; Macek, J.; Straka, J.; Šima, P.; Tyrner, P.; Veprěk, D.; Zeman, V. Blanokrídlovce (Hymenoptera) na vybraných,lokalitách Borskej nížiny. Acta Mus. Tek. 2010, 8, 78–111. [Google Scholar]
- Smetana, V.; Šima, P.; Bogusch, P.; Erhart, J.; Holý, K.; Macek, J.; Roller, L.; Straka, J. Blanokrídlovce (Hymenoptera) na vybraných lokalitách v okolí Levíc a Kremnice. Act. Mus. Tek. 2015, 10, 44–68. [Google Scholar]
- Šima, P.; Straka, J. First records of Heriades rubicola Pérez, 1890 (Hymenoptera: Megachilidae) and Nomada moeschleri Alfken, 1913 (Hymenoptera: Apidae) from Slovakia. Ent. Carpath. 2016, 28(1), 14–18. [Google Scholar]
- Tyrner, P.; Majzlan, O. Zlatěnkovití (Hymenoptera: Chrysididae) Pohoří Burda a jeho okolí. Ochr. pr., Banská B, 27.
- Smetana, V.; Roller, L.; et al. Blanokrídlovce (Hymenoptera) na vybraných lokalitách v CHKO Malé Karpaty. Act. Mus. Tek. 2020, 12, 75–141. [Google Scholar]
- Tóth, S. Magyarország zengőlégy faunája (Diptera, Syrphidae). Hoverflies of Hungary (Diptera, Syrphidae). e-Acta Nat. Pan.
- Tóth, S. Magyarország fürkészlégy faunája (Diptera, Tachinidae). Tachinid flies of Hungary (Diptera, Tachinidae). e-Acta Nat. Pan.
- Fazekas, I.; Tóth, S. A hazai bögölyök nyomában (Diptera, Tabanidae). Horse-flies of Hungary (Diptera, Tabanidae). Pannon Intézet, Pécs, Hungary 2023, pp. 1-124.
- Zombori, L. Sawflies from Fertő-Hanság National Park (Hymenoptera, Symphyta). In The Fauna of the Fertő-Hanság National Park; Mahunka, S. Ed.; Publisher, Hungarian Natural History Museum, Budapest, Hungary 2002; pp. 545-552.
- Haris, A. Sawflies of the Zselic Hills, SW Hungary Hymenoptera, Symphyta. Nat. Som. 2009, 15, 127–158. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Vértes Mountains Hymenoptera, Symphyta. Nat. Som. 2010, 17, 209–238. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Börzsöny Mountains North Hungary Hymenoptera, Symphyta. Nat. Som. 2011, 19, 149–176. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of Belső-Somogy (Hymenoptera, Symphyta). Nat. Som. 2012, 22, 141–162. [Google Scholar] [CrossRef]
- Haris, A. Second contribution to the sawflies of Belső Somogy Hymenoptera, Symphyta. Nat. Som. 2018, 31, 45–62. [Google Scholar] [CrossRef]
- Haris, A. Sawflies from Külső-Somogy, South-West Hungary (Hymenoptera, Symphyta). Nat. Som. 2018, 32, 147–164. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Keszthely Hills and its surroundings. Nat. Som. 2019, 33, 107–128. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of Southern part of Somogy county Hymenoptera, Symphyta. Nat. Som. 2020, 35, 51–70. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Cserhát Mountains Hymenoptera, Symphyta. Nat. Som. 2021, 37, 25–42. [Google Scholar] [CrossRef]
- Haris, A. Second contribution to the knowledge of sawflies of the Zselic Hills (Hymenoptera, Symphyta). A Kaposvári Rippl-Rónai Múzeum Közleményei (Commun. Rippl-Rónai Mus. Kaposvár). [CrossRef]
- Haris, A.; Vidlička, L.; Majzlan, O.; Roller, L. Effectiveness of Malaise trap and sweep net sampling in sawfly research (Hymenoptera, Symphyta). Biologia 2024, 79, 1705–1714. [Google Scholar] [CrossRef]
- Zombori, L. A Bakonyi Természettudományi Múzeum levéldarázs-gyűjteménye (Hymenoptera, Symphyta) I. (The collection of sawflies (Hymenoptera, Symphyta) of the Bakony Natural History Museum I.). Veszprém m. múz. közl.
- Zombori, L. A Bakonyi Természettudományi Múzeum levéldarázs-gyűjteménye (Hymenoptera, Symphyta) II. (The collection of sawflies (Hymenoptera, Symphyta) of the Bakony Natural History Museum II.). Veszprém m. múz. Közl, 14.
- Zombori, L. A Bakonyi Természettudományi Múzeum levéldarázs-gyűjteménye (Hymenoptera, Symphyta) III. (The collection of sawflies (Hymenoptera, Symphyta) of the Bakony Natural History Museum III.). Veszprém m. múz. Közl, 15.
- Zombori, L. A Bakonyi Természettudományi Múzeum levéldarázs-gyűjteménye (Hymenoptera, Symphyta) IV. (The collection of sawflies (Hymenoptera, Symphyta) of the Bakony Natural History Museum IV.). Fol. Mus. Hist. Nat. Bakonyiensis.
- Józan, Z. A Zselic darázsfaunájának (Hymenoptera, Aculeata) állatföldrajzi és ökofaunisztikai vizsgálata. Zoologeographic and ecofaunistic study of the Aculeata fauna (Hymenoptera, Aculeata) of Zselic. Somogyi Múzeumok Közleményei.
- Józan, Z. A Béda-Karapancsa Tájvédelmi Körzet fullánkos hártyásszárnyú (Hymenoptera, Aculeata) faunájának alapvetése. Aculeata (Hymenoptera, Aculeata) fauna of the Béda-Karapancsa Landscape Protection Area. Dunántúli Dolg. Természettudományi Sor, 6.
- Józan, Z. A Baláta környék fullánkos hártyásszárnyú faunájának (Hym., Aculeata) alapvetése. Aculeata fauna of Lake Baláta (Hym., Aculeata). Somogyi Múzeumok Közleményei.
- Józan, Z. A Mecsek kaparódarázs faunájának (Hymenoptera, Sphecoidea) faunisztikai, állatföldrajzi és ökofunisztikai vizsgálata. Faunistical, zoogeographical and ecofaunistical investigation on the Sphecoids fauna of the Mecsek Montains (Hymenoptera, Sphecoidea). Nat. Som, 3.
- Józan, Z. A Barcsi borókás fullánkos faunája, III. (Hymenoptera, Aculeata). Aculeata fauna of Barcs Juniper Woodland (Hymenoptera, Aculeata). Nat. Som, 26.
- Infusino, M.; Brehm, G.; Di Marco, C.; Scalercio, S. Assessing the efficiency of UV LEDs as light sources for sampling the diversity of macro-moths (Lepidoptera). Eur. J. Entomol. 2017, 114, 25–33. [Google Scholar] [CrossRef]
- Pan, H.; Liang, G.; Lu, Y. Response of Different Insect Groups to Various Wavelengths of Light under Field Conditions. Insects 2021, 12, 427. [Google Scholar] [CrossRef]
- van Langevelde, F.; Ettema, J. A.; Donners, M. Wallis De Vries, M. F.; Groenendijk, D. Effect of spectral composition of artificial light on the attraction of moths. Biol. Cons. 2011, 144(9), 2274-2281.
- Sage, W.; Utschick, H. Nachtfalter (Lepidoptera, Macroheterocera) im NSG "Untere Alz" und ihre Bedeutung für die Pflege- und Entwicklungsplanung. Berichte der Anl. 1997, 21, 149–177. [Google Scholar]
- Kanarskyi, Y.; Geriak, Y.; Lashenko, E. Ecogeographic structure of the Moth fauna in upper Tisa River basin. Transylv. Rev. Syst. Ecol. Res. 2011, 11, 143–168. [Google Scholar]
- Moore, J.L.; McCollin, D. The value of museum and other uncollated data in reconstructing the decline of the chequered skipper butterfly Carterocephalus palaemon (Pallas, 1771). J. Nat. Sci. Coll. 2022, 10, 31–44. [Google Scholar]
- Davis, C. L.; Guralnick, R. P.; Zipkin, E. F. Challenges and opportunities for using natural history collections to estimate insect population trends. J. Anim Ecol. 2023, 92(2), 237–249. [Google Scholar] [CrossRef] [PubMed]
- McGie, H.; Hancock, G.; Reilly, M.; Robinson, J.; Riddington, K.; Brown, C.; Machin, R.; et al. Museum collections and biodiversity conservation. Publisher: Curating Tomorrow, Liverpool, UK, 2019; pp. 1-104.
- McDermott, A. To understand the plight of insects, entomologists look to the past. PNAS News Feature 2020, 118(2), e2018499117. [Google Scholar] [CrossRef] [PubMed]
- Jeppsson, T.; Lindhe, A.; Gardenfors, U.; Forslund, P. The use of historical collections to estimate population trends, A case study using Swedish longhorn beetles (Coleoptera, Cerambycidae). Biol. Cons. 2010, 143, 1940–1950. [Google Scholar] [CrossRef]
- McCarthy, M. A. Identifying declining and threatened species with museum data. Biol. Cons. 1998, 83(1), 9–17. [Google Scholar] [CrossRef]
- Williams, K. A. Museum Collections - Resources for Biological Monitoring. frican Invertebrates. [CrossRef]
- Nowicki, P.; Settele, J.; Henry, P. Y; Woyciechowskia, M. Butterfly Monitoring Methods, The ideal and the Real World. Isr. J. Ecol. Evol. 2008, 54, 69–88. [Google Scholar] [CrossRef]
- Settele, J.; Kudrna, O.; Harpke, A.; Kühn, I.; van Swaay, Ch.; Verovnik, R.; Warren, M.; Wiemers, M.; Hanspach, J.; Hickler, Th.; Kühn, E.; Halder, I.; Veling, K.; Vliegenthart, A.; Wynhoff, I.; Schweiger, O. Climatic Risk Atlas of European Butterflies. Biorisk.
- Schweiger, O.; Harpke, A.; Wiemers, M.; Settele, J. Climber, climatic niche characteristics of the butterflies in Europe. ZooKeys 2014, 367, 65–84. [Google Scholar] [CrossRef]
- Mora, A.; Wilby, A.; Menéndez, R. South European mountain butterflies at a high risk from land abandonment and amplified effects of climate change. Insect Cons. and Div. 2023, 16(6), 838–852. [Google Scholar] [CrossRef]
- Kudrna, O.; Harpke, A.; Lux, K. , Pennerstorfer, J.; Schweiger, O., Ed.; Settele, J. et al. Distribution Atlas of Butterflies in Europe. Publisher: Gesellschaft für Schmetterlingschutz, Halle, Germany, 2011; pp. 1–576. [Google Scholar]
- Roques, A; Marc Kenis, M.; Lees, D.; Lopez-Vaamonde, C.; Rabitsch, W.; Rasplus, J-Y.; Roy, D. B. Alien terrestrial arthropods of Europe. Biorisk, 1028.
- Brewer, S. ; Cheddadi,R.; de Beaulieu, J. L.; Reille, M. The spread of deciduous Quercus throughout Europe since the last glacial period. For. Ecol. Manage.
- Global Biodiversity Information Facility (GBIF) Available online:. Available online: https://www.gbif.org (accessed on 26. 11. 2024).
- Panagiotakopulu, E.; Buckland, P. C. Early invaders - Farmers, the granary weevil and other uninvited guests in the Neolithic. Biological Invasions. [CrossRef]
- Ripka, G. Checklist of the Aphidoidea and Phylloxeroidea of Hungary (Hemiptera, Sternorrhyncha). Folia ent. Hung. 2008, 69, 19–157. [Google Scholar]
- Zahradnik, P. A Check-list of Ptinidae (Coleoptera, Bostrichoidea) of the Balkan Peninsula. Fol. Heyrovskyana, 2016. [Google Scholar]
- Soós, A. Über die Sepsiden, Piophiliden und Drosophiliden des Karpatenbeckens. Fragm. faun. Hung. 1945, 8(1-4), 18-29.
- Ripka, G. Jövevény kártevő ízeltlábúak áttekintése Magyarországon I. (Overview of non-native insect pest arthropods in Hungary I.) Növényvédelem 2010, 46(2), 45-58.
- Skuhrava, M.; Skuhrava, V. Gall midges (Diptera, Cecidomyiidae) of Hungary. Annls hist.-nat. Mus. natn. Hung, 91.
- Lienhard, Ch. Beitrag zur Kenntnis der Psocopteren-Fauna Ungarns (Insecta). Ann. Hist.-Nat. Mus. Natl. Hung, 78.
- Fazekas, I. Dr. Kuthy Béla entomológiai gyűjteménye II. Microlepidoptera (Lepidoptera). Nat. Som. [CrossRef]
- Jenser, G. Behurcolt kártevő Thysanoptera fajok. (Introduced Thysanoptera pests). Növényvédelem.
- László, M.; Katona, G. P.; Péntek, L. A.; Nagy, A. V. Spread of the invasive Giant Asian Mantis Hierodula tenuidentata Saussure, 1869 (Mantodea, Mantidae) in Europe with new Hungarian data. Bonn zool. Bull. 2023, 72(1), 133–144. [Google Scholar]
- Teodorescu, I. Contribution to Database of Alien/Invasive Homoptera Insects in Romania. Rom. J.. Biol. Zool.
- Kóbor, P. Magyarország invaziv cimerespoloskái (Heteroptera, Pentatomidae). (Invasive stink bugs of Hungary (Heteroptera, Pentatomidae)). Növényvédelem.
- Jelinek, J.; Audisio, P.; Hajek, J.; Baviera, C. .; Moncourtier, B.; Barnouin, Th. Epuraea imperialis (Reitter, 1877). New invasive species of (Coleoptera) in Europe, with a checklist of sap beetles introduced to Europe and Mediterranean areas. Atti. Accad. Pelorit. Pericol. Cl. Sci. Fis. Mat. Nat, (2.
- Szinetár, Cs.; Kenyeres, Z. Introducing of Ameles spallanzania (Rossi, 1792) (Insecta, Mantodea) to Hungary raising questions of fauna-changes. Nat. Som. 2020, 35, 133–138. [Google Scholar] [CrossRef]
- Nitzui, E.; Dobrin, I.; Dumbrava, M.; Gutue, M. The Range Expansion of Ovalisia festiva (Linnaeus, 1767) (Coleoptera, Buprestidae) in Eastern Europe and Its Damaging Potential for Cupressaceae. Trav. du Mus. Nat. Hist. Grigore Antipa, 58. [CrossRef]
- Merkl, O.; Hegyessy, G. Negyvenkét bogárcsalád adatai a sátoraljaújhelyi PIM – Kazinczy Ferenc Múzeum gyűjteményéből (Coleoptera, Polyphaga, Bostrichiformia, Cucujiformia, Elateriformia, Staphyliniformia). (Data on forty-two beetle families from the collection of the PIM – Kazinczy Ferenc Museum at Sátoraljaújhely) Folia hist. Nat. Mus. Matraensis 2022, 46, 45–134.
- Vas, Z.; Rékási, J. , Rózsa, L. A checklist of lice of Hungary (Insecta, Phthiraptera). Ann. Hist.-Nat. Mus. Nat. Hung.
- Adam, C.; Constantinescu, I.C.; Drăghici A., C.; Fusu M., M.; Gheoca, V.; Iancu, L.; Iorgu I., Ș.; Irimia A., G.; Maican, S.; Manu, M.; Petrescu A., M.; Popa A., F.; Rădac I., A.; Ruști D., M.; Sahlean C., T.; Szekely, L.; Șerban, C.; Tăușan, I. Lista preliminară a speciilor alogene invazive și potențial invazive de nevertebrate terestre din România (Preliminary list of invasive and potentially invasive alien species of terrestrial invertebrates in Romania). Ministerul Mediului, Apelor şi Pădurilor & Universitatea din Bucureşti.Bucureşti, Romania, 2020; pp. 1-40.
- Bednár, F.; Hemala, V.; Čejka, T. First records of two new silverfish species (Ctenolepisma longicaudatum and Ctenolepisma calvum) in Slovakia, with checklist and identification key of Slovak Zygentoma. Biologia. [CrossRef]
- Papp, L.; Pecsenye, K. Drosophilidae (Diptera) of Hungary. Acta Biol. Debrecina, 1986. [Google Scholar]
- Schwarz, Ch. J.; Ehrmann, R. Invasive Mantodea species in Europe. Articulata 2018, 33, 73–90. [Google Scholar]
- Tóth, J. A fajspektrum változásának lehetséges okai. Behurcolt új erdészeti kártevõk Magyarországon. ( Possible causes of the change in the species spectrum. New introduced forest pests in Hungary.) Mag, Kutatás, Termesztés, Kereskedelem 2001, 6, 9-17.
- Kontschán, J.; Kiss, E.; Ripka, G. Új adatok a hazai levélbolhák (Insecta, Psylloidea) előfordulásaihoz III. (New Data to Occurrences of the Hungarian Jumping Plant Lice (Insecta, Psylloidea) III.). Növényvédelem.
- Kontschán, J.; Ripka, G. Új adatok a hazai levélbolhák (Insecta, Psylloidea) előfordulásaihoz II.(New Data to Occurrences of the Hungarian Jumping Plant Lice (Insecta, Psylloidea) II.). Növényvédelem.
- CABI, Centre for Agriculture and Bioscience International, Exotic Insect Biocontrol Agents Released in Europe. Available online: https://cabidigitallibrary.org by 80.98.245.128 (accessed on 16.10. 2024).
- Szeőke, K.; Csóka, Gy. Jövevény kártevő ízeltlábúak Magyarországon – Lepkék (Lepidoptera). (An overview of the alien arthropods in Hungary Lepidoptera). Növényvédelem.
- Merkl, O. New beetle species in the Hungarian fauna (Coleoptera). Folia ent. Hung. 2006, 67, 19–36. [Google Scholar]
- Kinál, F.; Puskás, G. Occurrence of Ectobius vittiventris (Costa, 1847) (Blattellidae, Ectobiinae) in Hungary. Állattani Közlemények. [CrossRef]
- Vas, Z.; Kőszegi, K.; Takács, A. First record of the Nearctic blue mud-dauber wasp Chalybion californicum (de Saussure, 1867) from Hungary (Hymenoptera, Sphecidae). Folia ent. Hung. 2024, 85, 35–39. [Google Scholar] [CrossRef]
- OEPP/EPPO, PM 6/3 (5) Biological control agents safely used in the EPPO region. OEPP/EPPO, Luxembourg, 2021; pp.
- Schlitt, B. P.; Lajtár, L.; Orosz, A. New grape-feeding leafhoppers in Hungary – first records of Erasmoneura vulnerata (Fitch, 1851) and Arboridia kakogawana (Matsumura, 1931) (Hemiptera, Clypeorrhyncha, Cicadellidae). Folia ent. Hung. 2024, 85, 41–52. [Google Scholar] [CrossRef]
- Kondorosy, E. Adventív poloskafajok Magyarországon (Invasive alien bugs (Heteroptera) in Hungary). Növényvédelem.
- Kohútová, M; Obona, J. Príspevok k poznaniu inváznych druhov hmyzu z územia Slovenska. Contribution to the knowledge of invasive insect species from Slovakia. Folia Oec.
- Groot, M. An overview of alien Diptera in Slovenia. Acta ent. Slovenica 2013, 21(1), 5–15. [Google Scholar]
- Kontschán, J.; Bodnár, D.; Ripka, G. Új adatok a hazai levélbolhák (Insecta, Psylloidea) előfordulásaihoz III.(New Data to Occurrences of the Hungarian Jumping Plant Lice (Insecta, Psylloidea) III.). Növényvédelem.
- Kozár, F.; Benedicty, Zs.; Fetykó, K.; Kiss, B.; Szita, É. An annotated update of the scale insect checklist of Hungary (Hemiptera, Coccoidea). ZooKeys 2013, 309, 49–66. [Google Scholar] [CrossRef]
- Józan, Z. Új kaparódarázs fajok (Hymenoptera, Sphecidae) Magyarország faunájában. New sphecid wasps (Hymenoptera, Sphecidae) in the fauna of Hungary. Somogyi Múzeumok Közleményei.
- Báldi, A.; Csányi, B.; Csorba, G.; Erős, T.; Hornung, E. , Merkl, O.; Orosz, A., Papp, L.; Szinetár, Cs., Varga, A.; Vas, Z.; Vétek, G. Ronkay, L.; Samu, F.; Soltész, Z.; Szép, T.; Vörös, J.; Zöldi, V.; Zsuga, K. Behurcolt és invazív állatok Magyarországon. (Introduced and invasive animals in Hungary). Magyar Tudomány.
- Radac, I. A. Expansion of some native or alien species of seed beetles and true bugs in Romania (Insecta, Coleoptera, Heteroptera). PhD. Thesis, Babeș-Bolyai University, Faculty of Biology and Geology, Kolozsvár, Romania, 2022; pp. 1–146. [Google Scholar]
- Kiss, A. A sáskajárások néhány területi és tájtörténeti vonatkozása a Kárpát-medencében. (Some territorial and landscape historical aspects of locust migrations in the Carpathian Basin). 9th Conference on landscape history, Keszthely, Hungary, 21. 06. 2012.
- Merkl, O. Hívatlan bogárvendégek Magyarországon. (Uninvited beetle guests in Hungary). Természettudományi Közlöny.
- Havasréti, B. A filoxérától a kígyóaknás szőlőmolyig. Few Words about the Phylloxera and the Grape Leaf Miner Moth. -Légkör, 2016, 61(4), 161-163.
- Zikic, V.; Stankovic, S.; Ilic, M.; Kavallieratos, N. G. Braconid parasitoids (Hymenoptera, Braconidae) on poplars and aspen (Populus spp.) in Serbia and Montenegro. North-West. J. Zool.
- Hári, K.; Fail, J.; Streito J., C.; Fetykó K., G.; Szita, É. , Haltrich A.; Vikár D.; Radácsi P.; Vétek G. First record of Aleuroclava aucubae (Hemiptera Aleyrodidae) in Hungary, with a checklist of whiteflies occurring in the country. Redia, 1926. [Google Scholar]
- Csősz, S.; Báthori, F.; Gallé, L.; Lőrinczi, G.; Maák, I.; Tartally, A.; Kovács, É.; Somogyi, A. Á.; Markó, B. The Myrmecofauna (Hymenoptera, Formicidae) of Hungary, Survey of Ant Species with an Annotated Synonymic Inventory. Insects 2021, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Tóth, J. Behurcolt és új erdészeti kártevők Magyarországon. (Introduced and new forest pests in Hungary). Erdészeti lapok.
- Csóka, Gy. Recent Invasions of Five Species of Leafmining Lepidoptera in Hungary. 31-36. In Proceedings, integrated management and dynamics of forest insects. Liebhold, A. M.; McManus, M. L.; Otvos, I. S.; Fosbroke, S. L. C. (Eds.); Publisher: Department of Agriculture, Forest Service, Northeastern Research Station, Madison, U. S. 167 pp. [CrossRef]
- Szőke, V. First records of two spongillafly species from Hungary (Neuroptera, Sisyridae). Folia ent. Hung. 2024, 85, 1–6. [Google Scholar] [CrossRef]
- Winkler, D.; Korda, M.; Traser, Gy. Two species of Collembola new for the fauna of Hungary. Opusc. Zool. 2011, 42(2), 199–206. [Google Scholar]
- Szabóky, Cs. New data to the Microlepidoptera fauna of Hungary, part XX (Lepidoptera, Autostichidae, Batrachedridae, Elachistidae, Sesiidae, Tineidae, Tortricidae). Folia ent. Hung. 2023, 84, 113–119. [Google Scholar] [CrossRef]
- Szeőke, K.; Avar, K. Athetis hospes (Freyer, 1831) Nyugat-Magyarországon (Lepidoptera, Noctuidae). (Athetis hospes (Freyer, 1831) in West-Hungary (Lepidoptera, Noctuidae). Nat. Som, 33. [CrossRef]
- Kóbor, P. Platycranus metriorrhynchus, A new Mediterranean plant bug in Hungary (Heteroptera, Cimicomorpha, Miridae, Orthotylinae). Acta Phytopathol. Entomol. Hung. 2023, 58(2), 131–138. [Google Scholar] [CrossRef]
- Koczor, S.; Kiss, B.; Szita, É.; Fetykó, K. Two Leafhopper Species New o the Fauna of Hungary (Hemiptera, Cicadomorpha, Cicadellidae). Acta Phytopathol. Entomol. Hung. 2012, 47(1), 71–76. [Google Scholar] [CrossRef]
- Balázs, K.; Rédey, D. Acrosternum heegeri Fieber, 1861 (Hemiptera, Heteroptera, Pentatomidae), another Mediterranean bug expanding to the north. Zootaxa 2017, 4347(2), 392–400. [Google Scholar] [CrossRef]
- Katona, G.; Balázs, S.; Dombi, O.; Tóth, B. First record of Spodoptera littoralis in Hungary (Lepidoptera, Noctuidae). Folia ent. Hung. 2020, 81, 119–122. [Google Scholar] [CrossRef]
- Nagy, Z. First record of Geocoris pallidipennis from Hungary (Hemiptera, Geocoridae). Folia ent. Hung, 1711; 81. [Google Scholar]
- Móra, A.; Sebteoui, K. Trithemis arteriosa (Burmeister, 1839) (Odonata, Libellulidae) in Hungary, can aquarium trade speed up the area expansion of Mediterranean species? North-West. J. Zool. 2020, 16(2), 237–238. [Google Scholar]
- Torma, A. Three new and a rare true bug species in the Hungarian fauna (Heteroptera, Dipsocoridae, Reduviidae, Lygaeidae). 2005, Folia ent. Hung. 66, 35-38.
- Takács, A.; Kőszegi, K. New records of Coleophoridae and Crambidae from Hungary (Lepidoptera). Folia ent. Hung. 2024, 85, 93–100. [Google Scholar] [CrossRef]
- Fiam, J.; Németh, T. An uninvited guest, the fifth species of silverfish in Hungary (Zygentoma, Lepismatidae). Folia ent. Hung. 2024, 85, 53–60. [Google Scholar] [CrossRef]
- De Ketelaere, A.; Magyar, B. First report of Ecclitura primoris Kokujev, 1902 in Hungary (Hymenoptera, Braconidae). Folia ent. Hung. 2024, 85, 81–84. [Google Scholar] [CrossRef]
- Schlitt B., P.; Székely, Á.; Orosz, A. First records of Ziczacella heptapotamica (Kusnezov, 1928) and Asymmetrasca decedens (Paoli, 1932) from Hungary (Hemiptera, Clypeorrhyncha, Cicadellidae). Folia ent. Hung. 2024, 85, 85–92. [Google Scholar] [CrossRef]
- Lukátsi, M.; Horváth, D. Natural enemy of Ambrosia artemisiifolia L. is spreading in Central Europe, first records of the ragweed leaf beetle (Ophraella communa LeSage, 1986) from Austria and Slovakia (Coleoptera, Chrysomelidae). Folia ent. Hung, 85. [CrossRef]
- Tóth, B.; Dombi, O.; Takács, A. Coleophora texanella Chambers, 1878, a new alien species in Hungary (Lepidoptera, Coleophoridae). Folia ent. Hung. 2024, 85, 107–114. [Google Scholar] [CrossRef]
- Tóth, B.; Balogh, B. Occurrence of Grammodes bifasciata (Petagna, 1786) in Hungary (Lepidoptera, Erebidae). Folia ent. Hung. 2024, 85, 133–136. [Google Scholar] [CrossRef]
- Balogh, B.; Tóth, B. First occurrence of Hypena lividalis (Hübner, 1796) in Hungary (Lepidoptera, Erebidae). Folia ent. Hung. 2024, 85, 147–151. [Google Scholar] [CrossRef]
- Schlitt B., P.; Horváth, Á.; Orosz, A. A Mediterranean gatecrasher, Neoaliturus inscriptus (Haupt, 1927) new to the Carpathian Basin (Hemiptera, Auchenorrhyncha, Cicadellidae). Folia ent. Hung. 2024, 85, 159–165. [Google Scholar] [CrossRef]
- Bertelsmeier, C.; Bonnamour, A.; Brockerhoff, E. G.; Pyšek, P.; Skuhrovec, J.; Richardson, D. M.; Liebhold, A. M. Global proliferation of nonnative plants is a major driver of insect invasions. BioScience 2024, 74(11), 770–781. [Google Scholar] [CrossRef] [PubMed]
- Csóka, Gy. A gyapottok bagolylepke (Helicoverpa armigera) terjedése Magyarországon. (The spread of the cotton bollworm (Helicoverpa armigera) in Hungary). In, Magyarország környezeti állapota 2015. Riesz, L. (Ed.); Publisher: Hermannn Ottó Nonprofit Intézet, Budapest, Hungary, 2016; pp. 62–64. [Google Scholar]
- Csóka, Gy.; Hirka, A.; Szőcs, L. Rovarglobalizáció a magyar erdőkben. (Insect globalization in Hungarian forests). Erdészettudományi Közlemények.
- Csóka, Gy.; Stone, G. N.; Melika, G. Non-native gall-inducing insects on forest trees, a global review. Biological Invasions 2017, 19, 3161–3181. [Google Scholar] [CrossRef]
- Kriston, É.; Bozsó, M.; Krizbai, L.; Csóka, Gy. és Melika, G. Klasszikus biológiai védekezés Magyarországon a szelídgesztenye gubacsdarázs, Dryocosmus kuriphilus (Yasumatsu, 1951) ellen, előzetes eredmények. (Classical biological control against the chestnut gall wasp, Dryocosmus kuriphilus (Yasumatsu, 1951), preliminary results). Növényvédelem.
- Duhay, G. Behurcolt bogárfajok Magyarországon. (Introduced beetles in Hungary). In Tények könyve 1998 A.Kereszty Ed.; Publisher: Greger-Delacroix Kiadó, Budapest, Hungary, 1998. [Google Scholar]
- García-Morales, M.; Denno, B. D.; Miller, D. R.; Miller, G,L.; Ben-Dov, Y.; Hardy, N. B. ScaleNet, A literature-based model of scale insect biology and systematics. Database. 2016. Available online: http://scalenet.info (accessed on day month year). [CrossRef]
- Ellis, W. N. Plant parasites of Europe, leafminers, galls and fungi. Available online: https://bladmineerders.nl (accessed on day month year).
- Bourgoin, Th. FLOW (Fulgoromorpha Lists on The Web), a world knowledge base dedicated to Fulgoromorpha. Version 8, Available online:. Available online: http://www.hemiptera-databases.org/flow (accessed on day month year).
- Blackman, R.; Eastop, V. Blackman & Eastop's Aphids on the World's Plants. Available online: https://aphidsonworldsplants.info/ (accessed on day month year).
- Szénási, V. Two new weevil species in Hungary (Coleoptera, Curculionidae, Entiminae). Folia ent. Hung. 2016, 77, 53–55. [Google Scholar] [CrossRef]
- European Commission: Joint Research Centre. European Alien Species Information Network (EASIN) Available online:. Available online: https://easin.jrc.ec.europa.eu/ (accessed on day month year).
- Sáfián, Sz.; Katona, G.; Tóth, B. First report of the palm borer moth, Paysandisia archon (Burmeister, 1879), in Hungary (Lepidoptera, Castniidae). Folia ent. Hung. 2023, 84, 137–143. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A. ; Connors, S.L. et al. IPCC, 2021: Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Publisher: Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2021; pp. 1-2391. [CrossRef]
- Ditlevsen, P.; Ditlevsen, S. Warning of a forthcoming collapse of the Atlantic meridional overturning circulation. Nature, Nat. Com, 4254; 14. [Google Scholar]
- Gagosian, R. B. Abrupt climate change. Woods Hole Oceanographic Institution. World Economic Forum, Davos, Switzerland, , 2003, Retrieved from https://www.whoi. 27 January.
- Ziska, L. H.; Blumenthal, D. M.; Runion, G. B.; Raymond Hunt, R. E. Diaz-Soltero, H. Invasive species and climate change, an agronomic perspective. Climatic Change, 1007. [Google Scholar]
- iNaturalist Available online:. Available online: https://www.inaturalist.org/places/romania (accessed on 27.11. 2024).
- Calmasur, Ö. New records and some new distribution data for the Turkish Nematinae (Hymenoptera: Symphyta: Tenthredinidae) fauna. Türk. entomol. Derg, 4: 44 (3). [CrossRef]
- Strumia F, Yildirim E (2007). Contribution to the knowledge of Chrysididae fauna of Turkey (Hymenoptera, Aculeata). Frust. Ent, 30.
- John Ascher, John Pickering: World Bee Diversity Interactive checklists of world bees by country. Available online: https://www.discoverlife.org/nh/cl/counts/Apoidea_species.html (accessed on 07. 11. 2024).
- Juho Paukkunen Sawflies, wasps, ants and bees – Hymenoptera https://laji.fi/en/taxon/MX.43122 (accessed on 03.08.2024). (accessed on 03.08.2024).
- Yildirim, E. The distribution and biogeography of Pompilidae in Turkey (Hymenoptera: Aculeata). Entomologie faunistique. Faun. Ent. 2011, 63(1), 23–34. [Google Scholar]
- Yildirim, E.; Özbek, H. An Evaluation on The Fauna of Vespoidea (Hymenoptera, Aculeata) of Turkey, Along with New Records and New Localities for Some Species. Turk. J. Zool. 1999, 23(6), 591–604. [Google Scholar]
- Haris, A.; Vidlička, L.; Majzlan, O.; Roller, L. Effectiveness of Malaise trap and sweep net sampling in sawfly research (Hymenoptera: Symphyta). Biologia 2024, 79, 1705–1714. [Google Scholar] [CrossRef]
- Balázs, A.; Haris, A. Sawflies (Hymenoptera: Symphyta) of Cerová vrchovina Upland (South Slovakia). Nat. Som, 6: 33. [CrossRef]
- Szabó, S.; Árnyas, E.; Varga, Z. Long-term light trap study on the macro-moth (Lepidoptera: Macroheterocera) fauna of the Aggtelek National Park. Acta zool. Acad. Sci. Hung. 2007, 53(3), 257–269. [Google Scholar]
- Varga, J.; Korompai, T.; Horokán, K.; Hirka, A.; Gáspár, C.; Kozma, P.; Csóka, G.; Csuzdi, C. Analysis of the Macrolepidoptera fauna in Répáshuta based on the catches of a light-trap between 2014–2019. Acta Univ. Esterházy Sect. Biol, 47.
- Szentkirályi, F.; Leskó, K.; Kádár, F. Hosszú távú rovarmonitorozás a várgesztesi erdészeti fénycsapdával. 2. A nagylepke együttes diverzitási mintázatának változásai. (Long-term insect monitoring with forestry light trap of Várgesztes. 2. Changes of pattern of species diversity of Macrolepidopteran assemblages). Erdészeti Kutatások, 91.
- Szentkirályi, F.; Leskó, K.; Kádár, F. Climatic effects on long-term fluctuations in species richness and abundance level of forest macrolepidopteran assesmblages in a Hungarian mountainous region. Carpth. J. Earth Environ. Sci. 2007, 2, 73–82. [Google Scholar]
- Biella, P. ; Cornalba,M.; Rasmont, P., Neumayer, J., Mei, M., Eds.; Brambilla, M. Climate tracking by mountain bumblebees across a century: Distribution retreats, small refugia and elevational shifts. Global Ecol. Cons. 2023, 54, 03163. [Google Scholar] [CrossRef]
- Manino, A.; Patetta, A.; Porporato, M.; Quaranta, M.; Intoppa, F.; Piazza, M. G.; Frilli, F. Bumblebee (Bombus Latreille, 1802) distribution in high mountains and global warming. Redia 2007, 90, 90,125–129. [Google Scholar]
- Kerr, J.T.; Pindar, A.; Galpern, P.; Packer, L.; et al. Climate change impacts on bumblebees converge across continents. Science, 6244. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G. C.; Darvill, B. Decline and conservation of bumble bees. Ann. Rev. Ent. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Arnóczkyné Jakab, D.; Tóth, M.; Szarukán, I.; Szanyi, S.; Józan, Z.; Sárospataki, M.; Nagy, A. Long-term changes in the composition and distribution of the Hungarian bumble bee fauna (Hymenoptera, Apidae, Bombus). J. Hym. Res, 2: 96. [CrossRef]
- Ban-Calefariu, C.; Sárospataki, M. Contributions to the knowledge of Bombus and Psithyrus Genera (Apoidea: Apidae) in Romania. Trav. Mus. Nat. Hist. Grigore Antipa 2007, 50, 239–258. [Google Scholar]
- Šima, P.; Smetana, V. ; Current distribution of the bumble bee Bombus haematurus (Hymenoptera: Apidae, Bombini) in Slovakia. Klapalekiana 2009, 45, 209–212. [Google Scholar]
- Šima, P.; Smetana, V. Quo vadis Bombus haematurus? An outline of the ecology and biology of a species expanding in Slovakia. Act. Mus. Tek. 2018, 11, 41–65. [Google Scholar]
- Biella, P.; Ćetković, A.; Gogala, A.; Neumayer, J.; Sárospataki, M.; Šima, P.; Smetana, V. Northwestward range expansion of the bumblebee Bombus haematurus into Central Europe is associated with warmer winters and niche conservatism. Insect Sci. 2021, 28, 861–872. [Google Scholar] [CrossRef]
- Manole, T. Biodiversity of insect populations from Apoidea superfamily in agricultural ecosystems. Rom. J. Plant Prot. 2014, 7, 111–128. [Google Scholar]
- Czyżewski, S.; Kierat, J.; Zapotoczny, K. First record of Bombus haematurus Kriechbaumer, 1870 (Hymenoptera: Apidae) in Poland. Act. zool.a crac, 7: 67.
- Kosior, A.; Celary, W.; Olejniczak, P.; Fijał, J.; Kro´l, W.; Solarz, W.; Płonka, P. The decline of the bumble bees and cuckoo bees (Hymenoptera: Apidae: Bombini) of Western and Central Europe. Oryx. [CrossRef]
- Šima, P. & Smetana, V. Čmele (Hymenoptera: Bombini) Liptovskej kotliny a Tatranského podhoria. (Bumble bees (Hymenoptera: Bombini) at selected localities of the Liptovská kotlina basin and the Tatranské podhorie foothill). Nat. Tut.
- Šima, P. & Smetana, V. Bombus (Cullumanobombus) semenoviellus (Hymenoptera: Apidae: Bombini) new species for the bumble bee fauna of Slovakia. Klapalekiana.
- Rahimi, E.; Barghjelveh, S.; Dong, P. Estimating potential range shift of some wild bees in response to climate change scenarios in northwestern regions of Iran. J. Ecol Environ. 1: 2021, 45, 2021. [Google Scholar] [CrossRef]
- Raider, R.; Reilly, J.; Bartomeus, I.; Winfree, R. Native bees buffer the negative impact of climate warming on honey bee pollination of watermelon crops. Global Change Biology 2013, 19, 3103–3110. [Google Scholar] [CrossRef]
- Duchenne, E.; Thébault, E.; Michez, D.; Gérard, M.; Celine Devaux, C.; et al. Long-term effects of global change on occupancy and flight period of wild bees in Belgium. Global Change Biology 2020, 26(12), 6753–6766. [Google Scholar] [CrossRef] [PubMed]
- Okely, M.; Engel, M.S.; Shebl, M.A. Climate Change Influence on the Potential Distribution of Some Cavity-Nesting Bees (Hymenoptera: Megachilidae). Diversity 2023, 15, 1172. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Pawlikowski, T.; Pawlikowski, K.; Banaszak-Cibicka, W.; Sparks, T. H. Does climate influence phenological trends in social wasps (Hymenoptera: Vespinae) in Poland? Eur. J. Entomol, 2: 107.
- Pawlikowski, T.; Pawlikowski, K. Phenology of Social Wasps (Hymenoptera: Vespinae) in the Kujawy Region (Northern Poland) under the the influence of climatic changes 1981–2000. Bull. Geog. Ser. Phys. Geogr. 2009, 1, 125–134. [Google Scholar] [CrossRef]
- Verheyde, F.; Vertommen, W.; Rhebergen, F.; De Ketelaere, A.; Doggen, K. & van Loon, M. Notes on the population dynamics of Euodynerus dantici (Rossi, 1790) and first records of its associated parasitoid Chrysis sexdentata Christ, 1791 in the Low Countries. Bull. ann. Soc. r. belge entomolog.
- Hallmann, C. A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Laczó, F. Pesticide use, issues and how to promote sustainable agriculture in Hungary. PAN Germany & Center for Environmental Studies Foundation, CES.,Brussels, Belgium, 2004; pp. 1-4. Retrieved from https://www.pan-germany.org/download/fs_hu_eng.pdf (accessed on 15. 11. 2024). (accessed on 15. 11. 2024).
- Goulet, H.; Huber, J. T. Hymenoptera of the world: an identification guide to families. Canada Communication Group, Ottawa, Canada 1993; pp. 668.
- Benson, R. B. 1968: Hymenoptera from Turkey, Symphyta. Bull. Brit. Mus. Nat. Hist. Ser. Ent. 1968, 22(4), 111–207. [Google Scholar]
- Barbir, J.; Martín, L.O.; Lloveras, X.R. Impact of Climate Change on Sawfly (Suborder: Symphyta) Polinators in Andalusia Region, Spain. In Handbook of Climate Change and Biodiversity; Filho, W. L., Barbir, J., Preziosi, R., Eds.; Climate Change Management; Springer Nature Switzerland AG: Cham, Switzerland, 2018; 323p. [Google Scholar]
- Sánchez-Martínez, G.; González-Gaona, E.; López-Martínez, V.; Espinosa-Zaragoza, S.; López-Baez, O.; Sanzón-Gómez, D.; Pérez-De la O, N.B. Climatic Suitability and Distribution Overlap of Sawflies (Hymenoptera: Diprionidae) and Threatened Populations of Pinaceae. Forests 2022, 13, 1067. [Google Scholar] [CrossRef]
- Nemer, N. Forest Pests Outbreaks and Climate Change. Second Thematic Workshop on Climate Change and Preparedness for Pandemic Situations, Beirut, Lebanon, 28. 09. 2021. Retrieved from https://civil-protection-knowledge-network.europa.eu/system/files/2022-08/Forest-Insects-and-Climate-Change-Sept-28.pdf (accessed on 21. 10. 2024). (accessed on 21. 10. 2024).
- Evenhuis, N.L.; Greathead, D. J. World Catalog of Bee Flies (Diptera: Bombyliidae). Backhuys Publishers, Leiden. The Netherlands, 1999; pp.
- Roberts, H.; El-Hawagry, M. S. A. New records of bee flies (Bombyliidae, Diptera) from the United Arab Emirates. Egypt. J. Biol. Pest Contr. 2024, 34(29), 1–11. [Google Scholar] [CrossRef]
- Yeates, D. K. The evolutionary pattern of host use in the Bombyliidae (Diptera): a diverse family of parasitoid flies. Biol J Linn Soc Lond. 1997, 60, 149–185. [Google Scholar] [CrossRef]
- Herrera, C. M. Complex long-term dynamics of pollinator abundance in undisturbed Mediterranean montane habitats over two decades. Ecological Monographs.
- Havkenberget, K.; Jaric, D.; Krcmar, S. Distribution of Tabanids (Diptera: Tabanidae) Along a Two-Sided Altitudinal Transect. Environ. Entomol. 2009, 38(6), 1600–1607. [Google Scholar] [CrossRef]
- Herczeg, T.; Száz, D.; Blahó, M.; Barta, A.; Gyurkovszky, M.; Farkas, R.; Horváth, G. The effect of weather variables on the flight activity of horseflies (Diptera: Tabanidae) in the continental climate of Hungary. Parasitol. Res, ,: 114, 1087. [Google Scholar]
- Dransfield, B.; Brightwell, B. InfluentialPoints.com Biology, images, analysis, design. https://influentialpoints.com/Gallery/Tabanus_bromius_band-eyed_brown_horsefly.htm (accessed on 23. 09. 2024). (accessed on 23. 09. 2024).
- Hallmann, C.A.; Ssymank, A.; Sorg, M.; de Kroon, H.; Jongejans, E. Insect biomass decline scaled to species diversity: General patterns derived from a hoverfly community. PNAS, 2021, 118(2). e2002554117. [CrossRef]
- Barendregt, A.; Zeegers, Th.; van Steenis, W.; Jongejans, E. Forest hoverfly community collapse: Abundance and species richness drop over four decades. Insect. Conserv. Divers. 2022, 15(5), 510–521. [Google Scholar] [CrossRef]
- Gatter, W.; Ebenhöhm, H.; Kimam, R.; Scherer, F. 50-jährige Unterschungen an migrierenden Schwebfliegen, Waffenfliegen und Schlupfwespen belegen extreme Rückgänge (Diptera: Syrphidae, Stratiomyidae; Hymenoptera: Ichneumonidae). Ent. Zeit. 2020, 130(3), 131–142. [Google Scholar]
- Reemer, M.; Smit, J. T.; Zeegers, Th.; Basisrapport voor de Rode Lijst Zweefvliegen. EIS Kenniscentrum Insecten. EIS 2024–03. Available online: https://www.eis-nederland.nl/rapporten (accessed on 19. 11. 2024).
- Zeegers, Th.; van Steenis, W.; Reemer, M.; Smit, J. T. Drastic acceleration of the extinction rate of hoverflies (Diptera: Syrphidae) in the Netherlands in recent decades, contrary to wild bees (Hymenoptera: Anthophila). J. van Syrphidae 2024, 3(1), 1–11. [Google Scholar] [CrossRef]
- IUCN SSC HSG/CPSG European Hoverflies: Moving from Assessment to Conservation Planning. A report to the European Commission by the IUCN SSC Conservation Planning Specialist Group (CPSG) and the IUCN SSC Hoverfly Specialist Group (HSG). Conservation Planning Specialist Group, Apple Valley, MN, USA. 2022, pp. 1-84.
- Zeegers, Th. Second addition to the checklist of Dutch tachinid flies (Diptera: Tachinidae). Nederl. Faun. Meded, 5: 34.
- Ziegler, J. Recent range extensions of tachinid flies (Diptera: Tachinidae, Phasiinae) in north-east Germany and a review of the overall distribution of five species. Stud. Dipt.
- Stireman, J. O.; Dyer, L. A.; Janzen, D. H.; et al. Climatic unpredictability and parasitism of caterpillars: Implications of global warming. PNAS Biol. Sci, 1738. [Google Scholar] [CrossRef]
- Environmental Statistics and Reporting team, Department for Environment Food & Rural Affairs: Butterflies in the United Kingdom and in England: 2023. Available online: https://www.gov.uk/government/statistics/butterflies-in-the-wider-countryside-uk (accessed on 19. 09. 2024).
- Panigaj, L.; Panigaj, M. Changes in lepidopteran assemblages in Temnosmrečinská dolina valley (the High Tatra Mts, Slovakia) over the last 55 years. Biologia 2008, 63(4), 582–587. [Google Scholar] [CrossRef]
- Birch, R. J.; Markl, G.; Gottschalk, T. K. Aestivation as a response to climate change: the Great Banded Grayling Brintesia circe in Central Europe. Ecol. Ent. 2021, 46, 1342–1352. [Google Scholar] [CrossRef]
- Schwarz, C.; Fartmann, Th. Conservation of a strongly declining butterfy species depends on traditionally managed grasslands. J. Ins. Cons. [CrossRef]
- Hudák, T. A nappali lepkefauna vizsgálata Székesfehérváron (Lepidoptera: Rhopalocera). Investigation on the butterfly fauna of Székesfehérvár (Lepidoptera: Rhopalocera). Nat. Som, 31.
- Fox, R.; Dennis, E. B.; Purdy, K.; Middlebrook, I.; Roy, D. B.; Noble, D.; Bothham, M. S.; Bourn, N. A. D. The State of the UK's Butterflies 2022. Publisher: Butterfly Conservation, Wareham, UK, 2023; pp. 1-28. Retrieved from https://eidc.ac.uk/ (accessed on 22. 10. 2024). (accessed on 22. 10. 2024).
- Uhl, B.; Wölfing, M.; Bässler, C. Mediterranean moth diversity is sensitive to increasing temperatures and drought under climate change. Sci. Rep. 2022, 12, 14473. [Google Scholar] [CrossRef] [PubMed]
- Betzholtz, P.-E.; Forsman, A.; Franzén, M. Associations of 16-Year Population Dynamics in Range-Expanding Moths with Temperature and Years since Establishment. Insects 2023, 14, 55. [Google Scholar] [CrossRef]
- Sparks, T. H.; Dennis, L. R. H.; Croxton, Ph. J.; Cade, M. Increased migration of Lepidoptera linked to climate change. Eur. J. Entomol. 2007, 104, 139–143. [Google Scholar] [CrossRef]
- Közösen a Természetért Alapítvány (Together for the Nature Foundation):Ízeltlábúak (Arthropodes). Available online: https://www.izeltlabuak.hu/ (accessed on day month year).
- Uherkovich, Á. Long-term monitoring of biodiversity by the study of butterflies and larger moths (Lepidoptera) in Sellye region (South Hungary, co. Baranya) in the years 1967–2022. Nat. Som, 9.
- Hufnagel, L.; Sipkay, Cs. A klímaváltozás hatása ökológiai folyamatokra és közösségekre. (The impact of climate change on ecological processes and communities). Budapesti Corvinus University, Budapest, Hungary, 2012; pp.
- Fox, R.; Dennis, E. B.; Harrower, C. A.; Blumgart, D. ; Bell. J. R.; Cook, P.; Davis, A. M.; Evans-Hill, L. J.; Haynes, F.; Hill, D.; Isaac, N. J. B,; Parsons, M. S.; Pocock, M. J. O.; Prescott, T.; Randle, Z.; Shortall, C. R.; Tordoff, G. M.; Tuson, D.; Bourn, N. A. D. The State of Britain’s Larger Moths 2021, 2021. [Google Scholar]
- Solarz, W.; Najberek, K.; Tokarska-Guzik, B.; Pietrzyk-Kaszyńska, A. Climate change as a factor enhancing the invasiveness of alien species. Environ. Socio.-econ. Stud, 11. [CrossRef]
- Gutierrez, A.P.; Ponti, L. Analysis of invasive insects: links to climate change. In Invasive Species and Global Climate Change, Ziska L. H., Dukes J.S., (Eds.); CABI Publishing, Wallingford, UK, 2014, pp. 45-61. DOI 10.1079/9781780641645. 0045. [Google Scholar]
- IUCN (International Union for Conservation of Nature), Invasive Alien Species and Climate Change. Available online: https://iucn.org/resources/issues-brief/invasive-alien-species-and-climate-change (accessed on 12. 09. 2024).
- Petrosyan, V.; Osipov, F.; Feniova, I.; Dergunova, N.; Warshavsky, A.; Khlyap, L.; Dzialowski, A. The TOP100 most dangerous invasive alien species in Northern Eurasia: invasion trends and species distribution modelling. NeoBiota 2023, 82, 23–56. [Google Scholar] [CrossRef]
- Lin, W.; Zhou, G.; Cheng, X.; Xu, R. Fast Economic Development Accelerates Biological Invasions in China. Fast Economic Development Accelerates Biological Invasions in China. PLoS ONE 2007, 2(11), e1208. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P. E.; Simberloff, E. et. al. Scientists’ warning on invasive alien species. Biol. Rev, 1511; 95. [Google Scholar] [CrossRef]
- Lambdon, P. W.; Pyšek, P.; Basnou, C.; Hejda, M.; et al. Alien flora of Europe: species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. [Google Scholar]
Figure 1.
The Rhododendron Leafhopper (Graphocephala fennahi Young, 1977), native to North America, were first found on the leaves of Rhododendron catawbiense in Hungary in 2012 (photo: György Csóka).
Figure 1.
The Rhododendron Leafhopper (Graphocephala fennahi Young, 1977), native to North America, were first found on the leaves of Rhododendron catawbiense in Hungary in 2012 (photo: György Csóka).

Figure 2.
Biogeographic map of the Carpathian Basin Pannonian (orange), Continental (green), and Alpine biogeographic regions (violet). (source: EEA [7]).
Figure 2.
Biogeographic map of the Carpathian Basin Pannonian (orange), Continental (green), and Alpine biogeographic regions (violet). (source: EEA [7]).
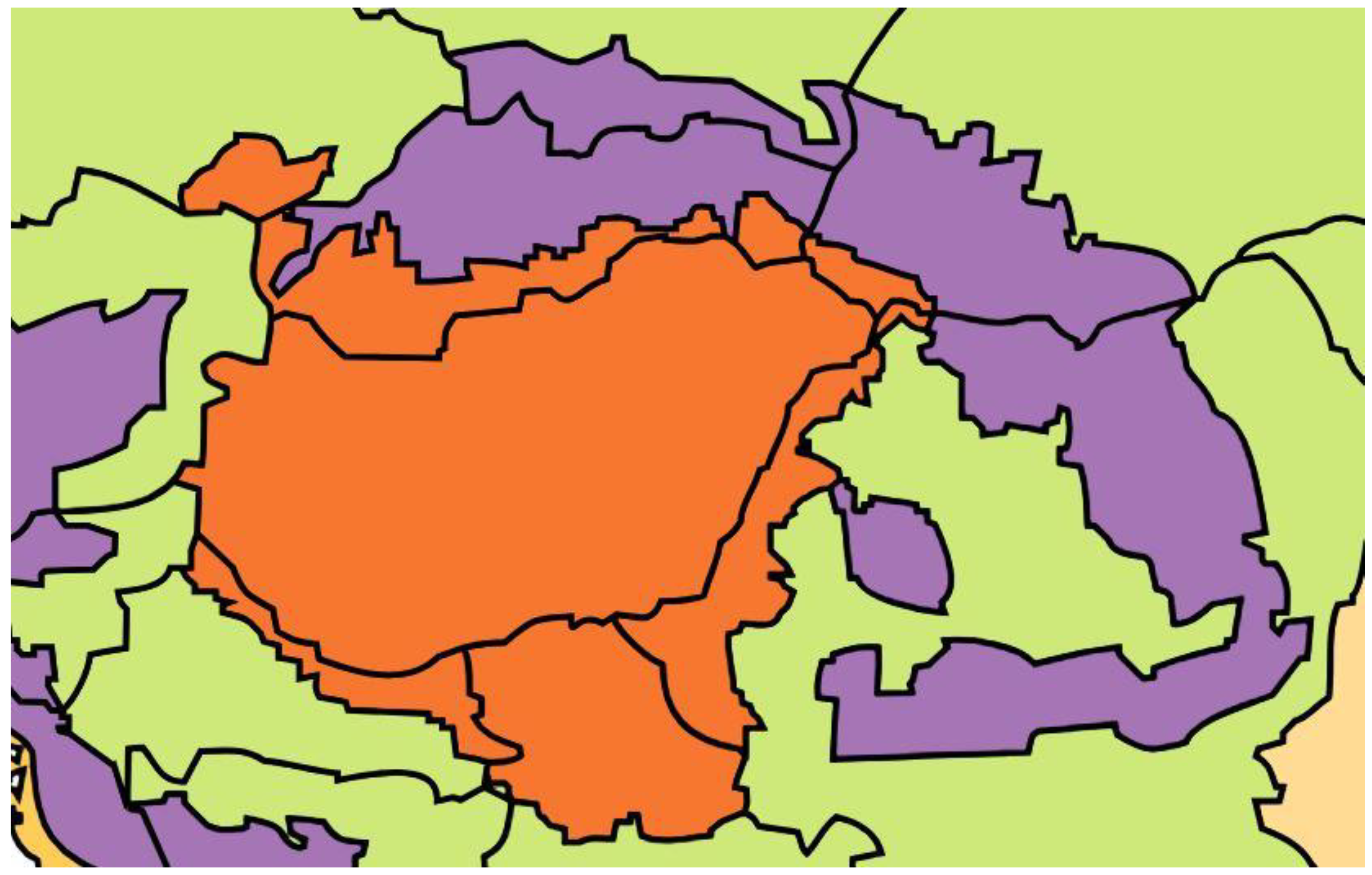
Figure 3.
Average annual temperatures of three Central European countries between 1970 and 2023. Green: Hungary, orange: Romania, red: Slovakia. Climatic data from World Bank Portal [18].
Figure 3.
Average annual temperatures of three Central European countries between 1970 and 2023. Green: Hungary, orange: Romania, red: Slovakia. Climatic data from World Bank Portal [18].
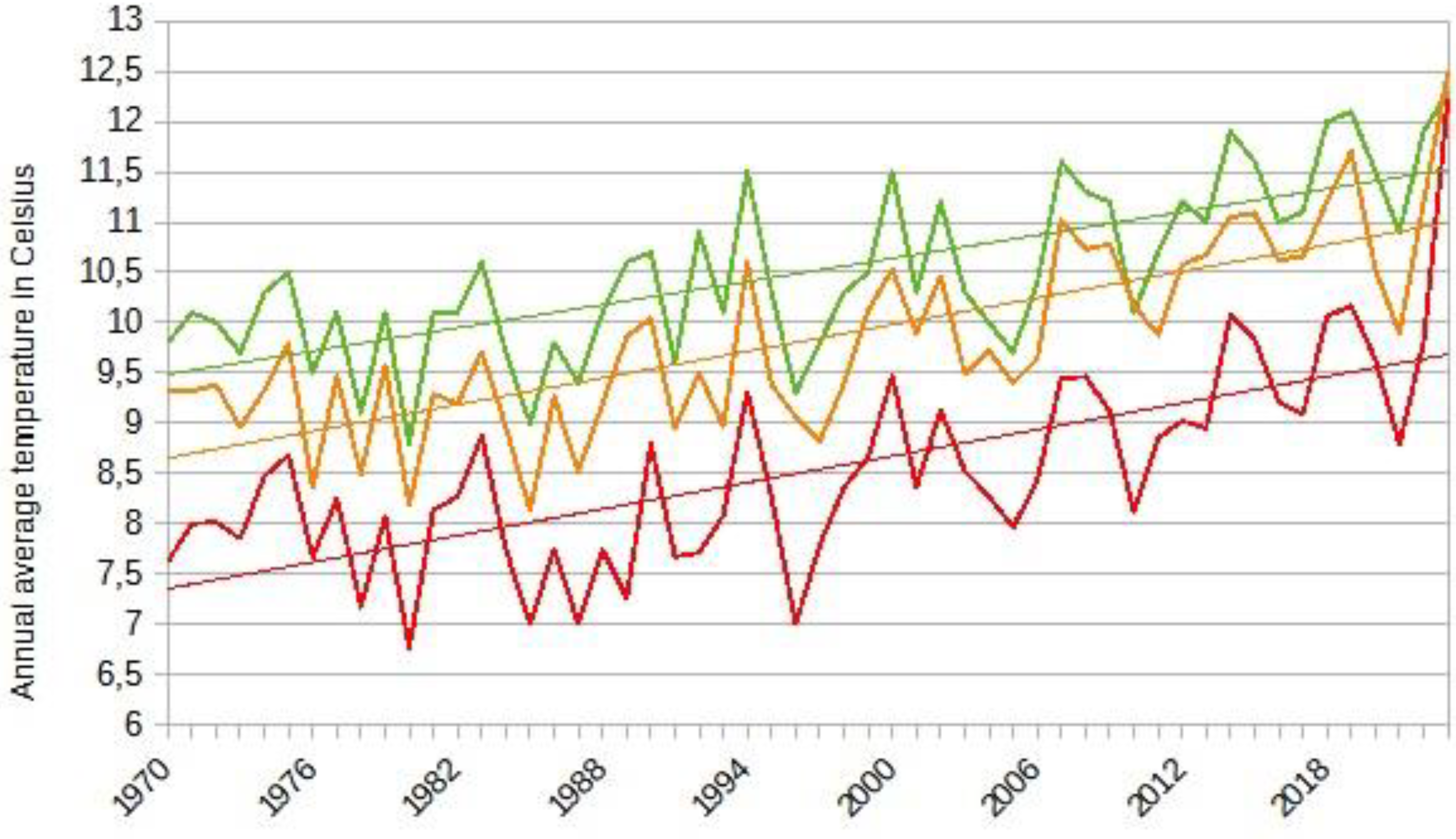
Figure 4.
Long term trend of wild bee families in the Carpathian Basin.
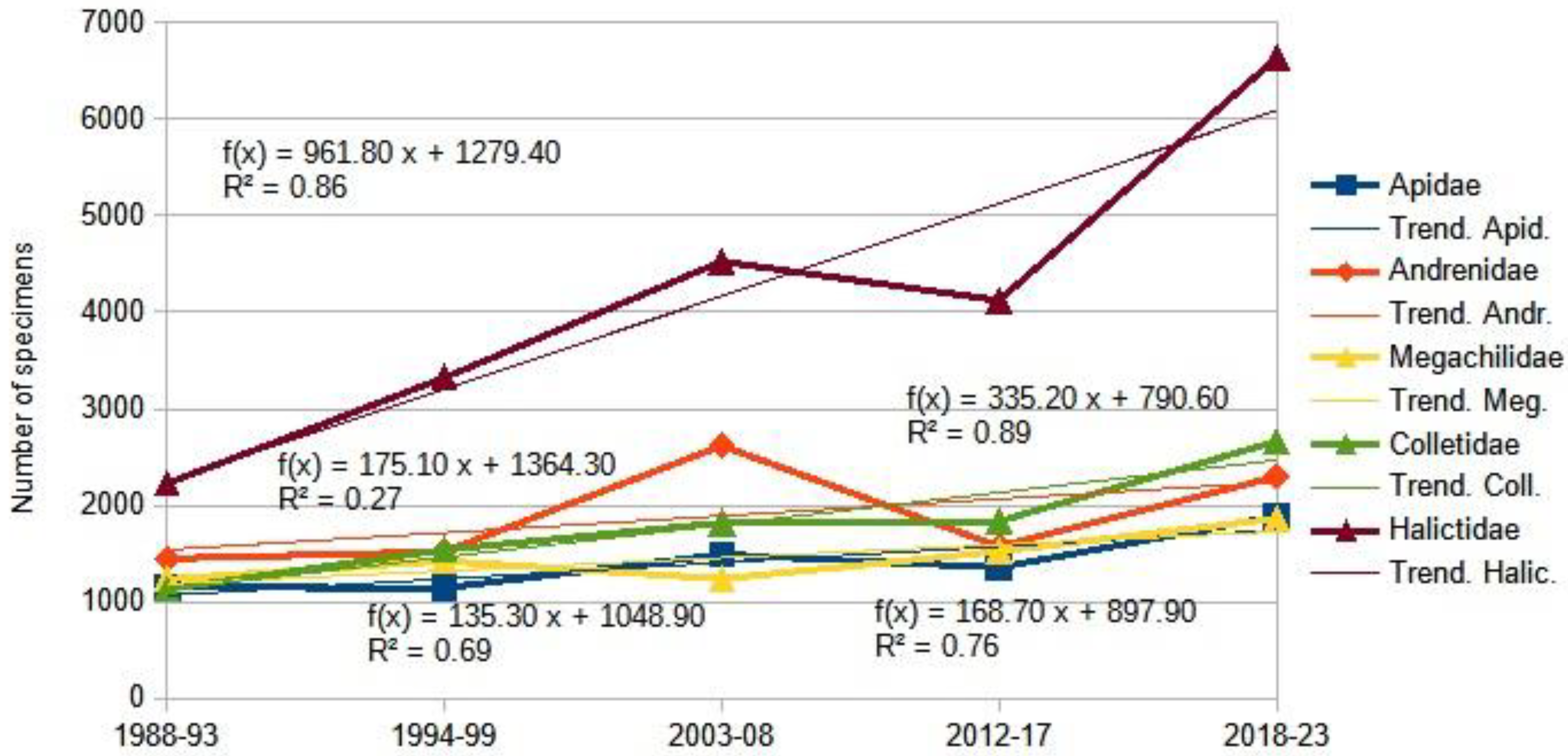
Figure 5.
Long term trend of Bembecidae, Psenidae, Sphecidae and Pemphedronidae in the Carpathian Basint.
Figure 5.
Long term trend of Bembecidae, Psenidae, Sphecidae and Pemphedronidae in the Carpathian Basint.
Figure 6.
Long term trend of Chrysididae, Philantidae and Crabronidae in the Carpathian Basin.
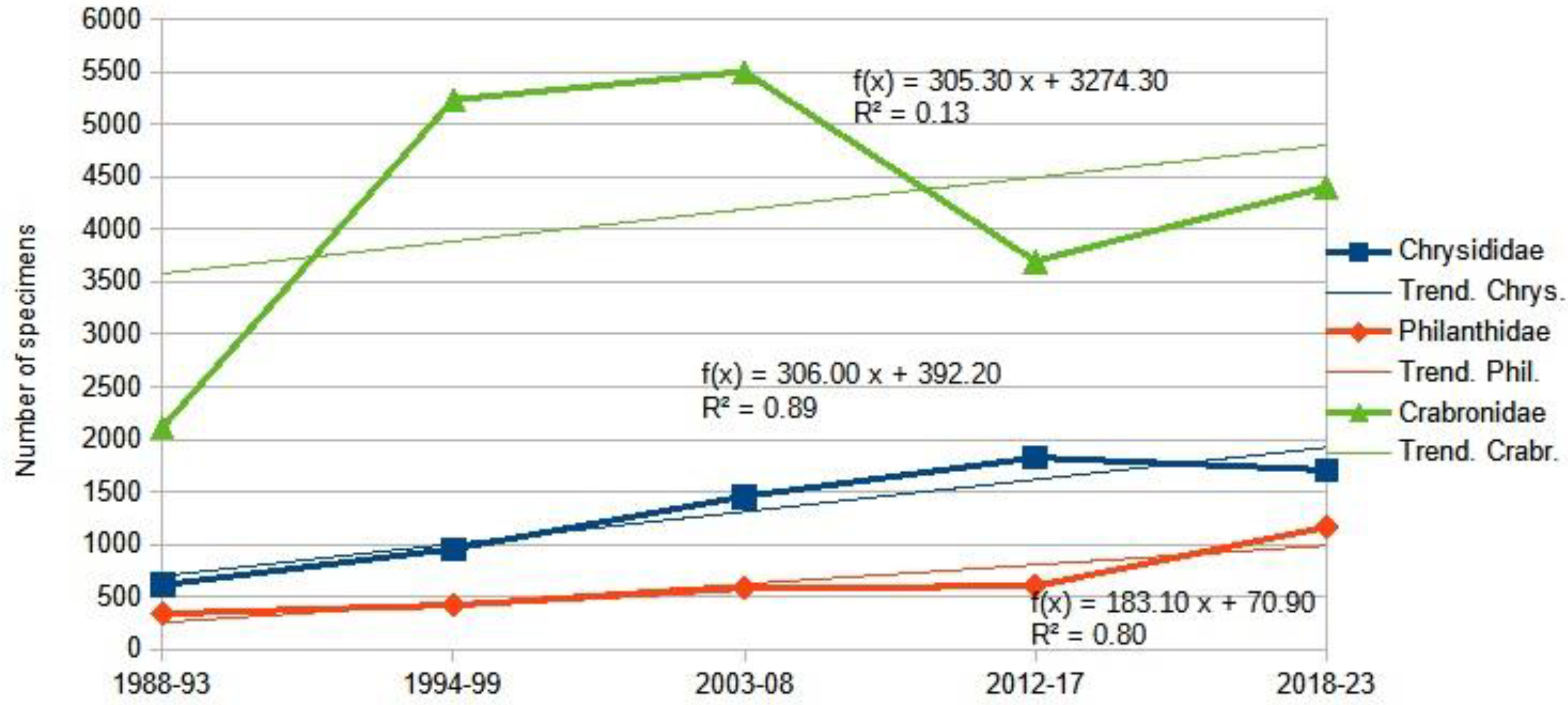
Figure 7.
Long term trend of sawflies (Symphyta) based on Haris et al., 2024 [6].
Figure 7.
Long term trend of sawflies (Symphyta) based on Haris et al., 2024 [6].
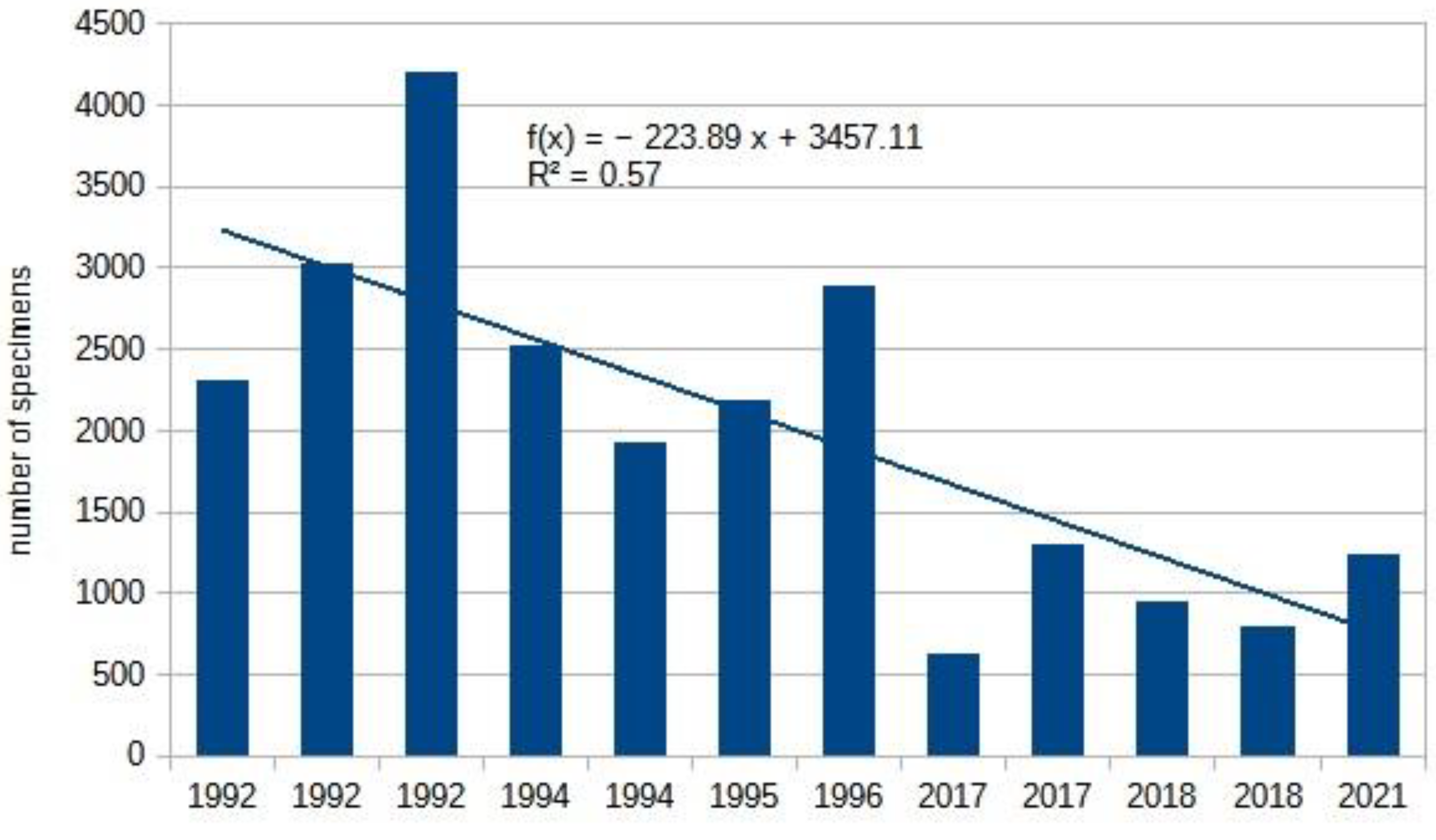
Figure 8.
Long term trend of horse-flies (Tabanidae) in the Carpathian Basin.
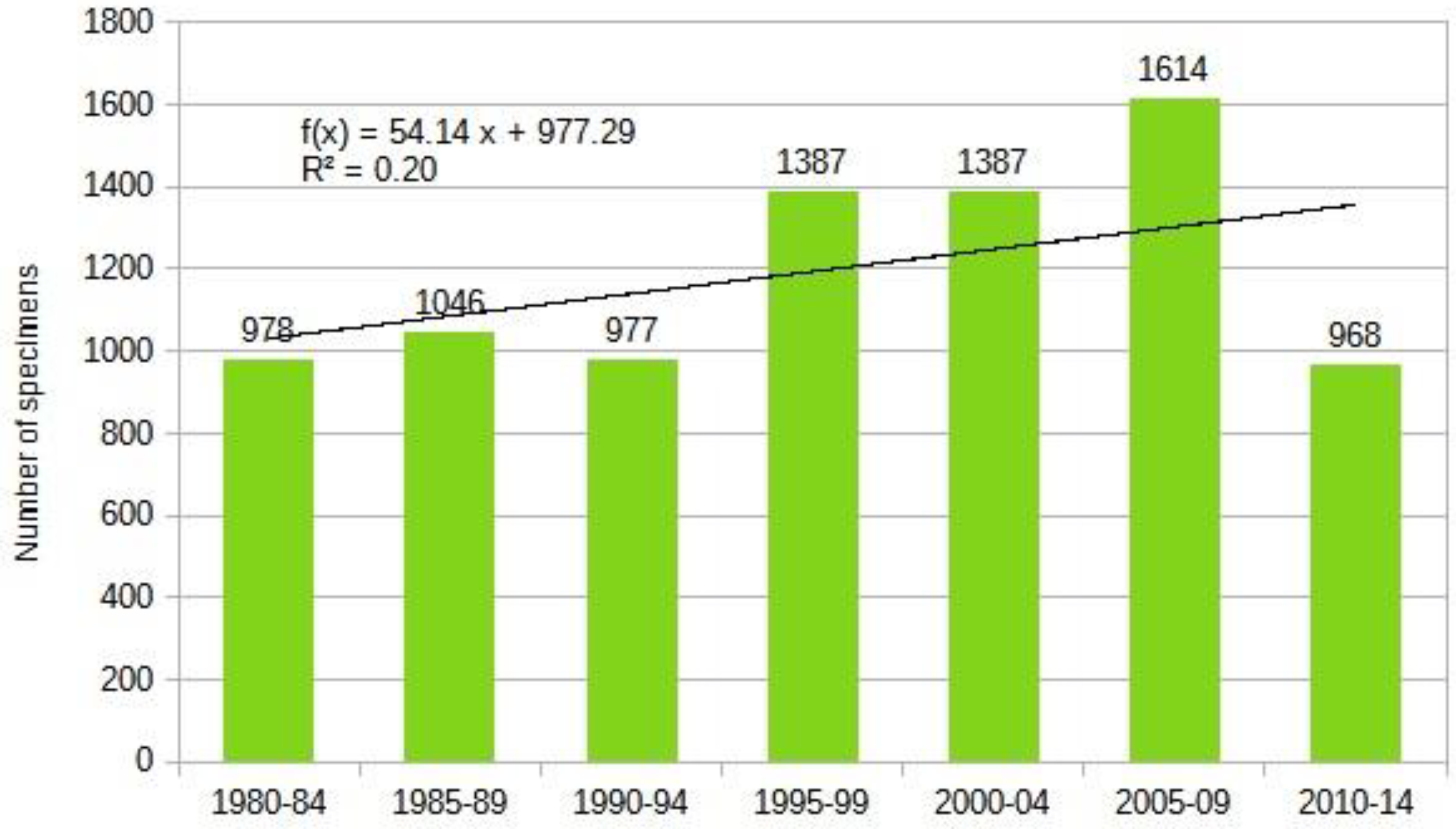
Figure 9.
Long term trend of hoverflies (Syrphidae) in the Carpathian Basin.
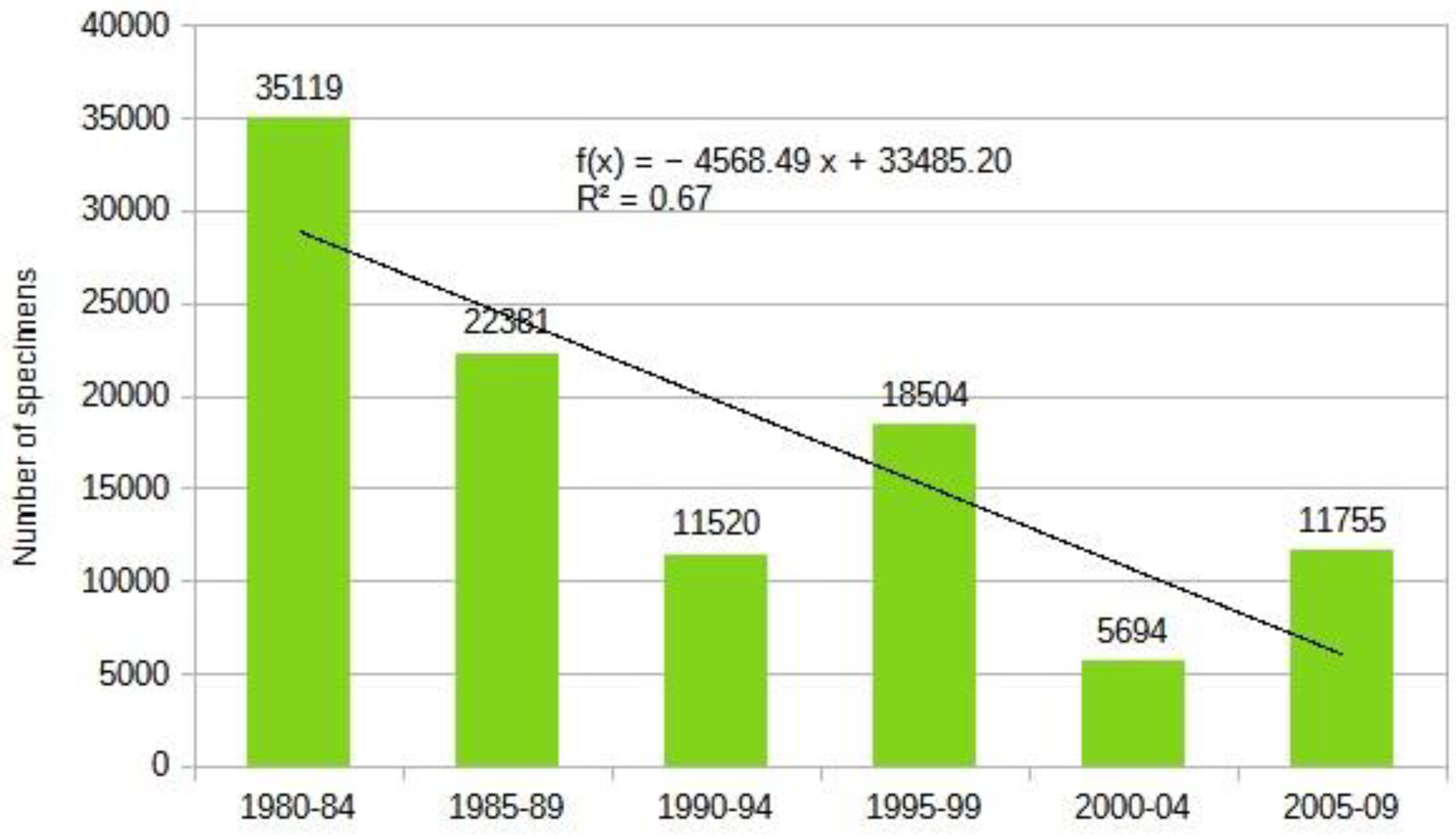
Figure 10.
Long term trend of bee flies (Bombyliidae) in the Carpathian Basin.
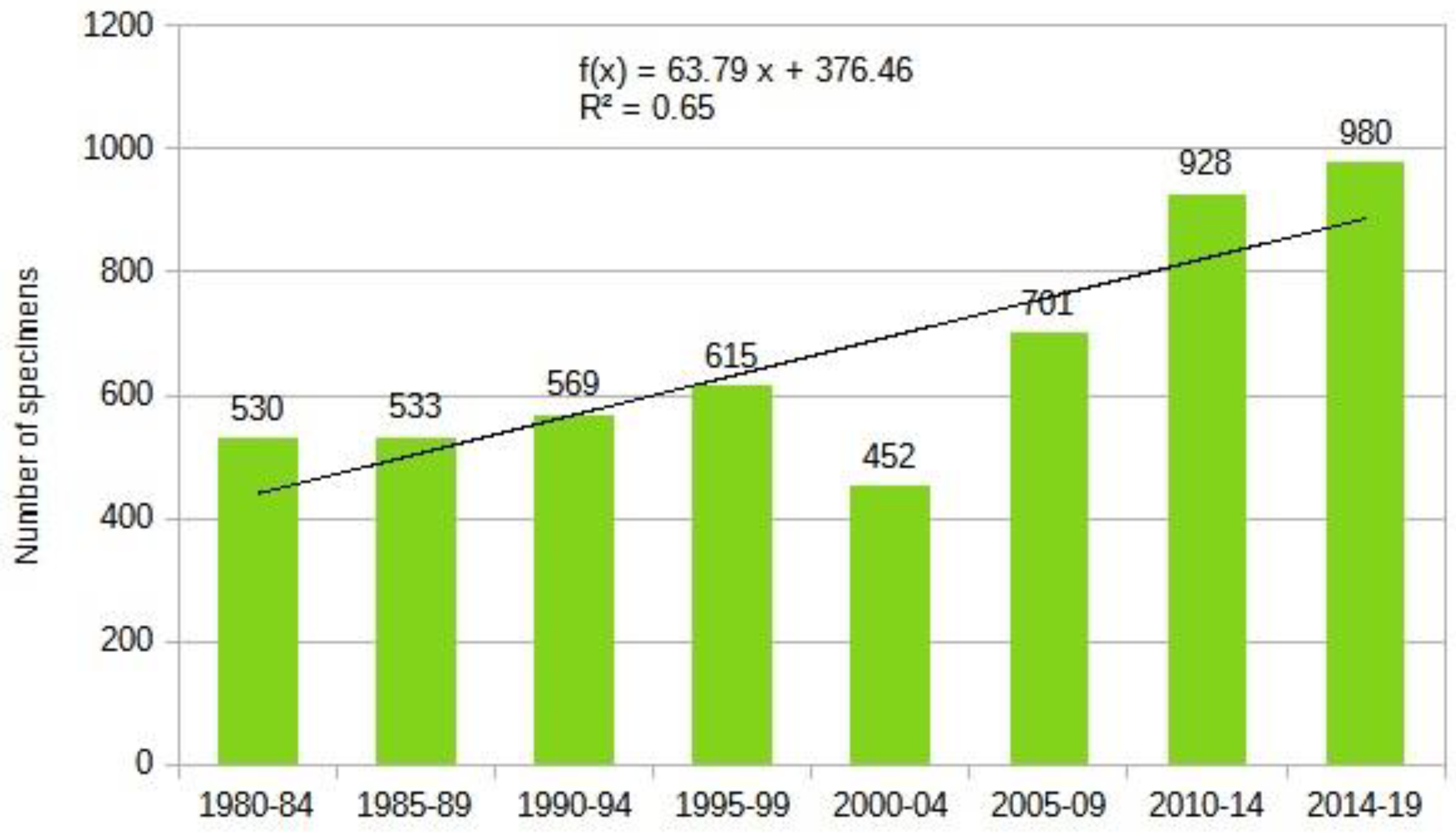
Figure 11.
Long term trend of tachinid flies (Tachinidae) in the Carpathian Basin.
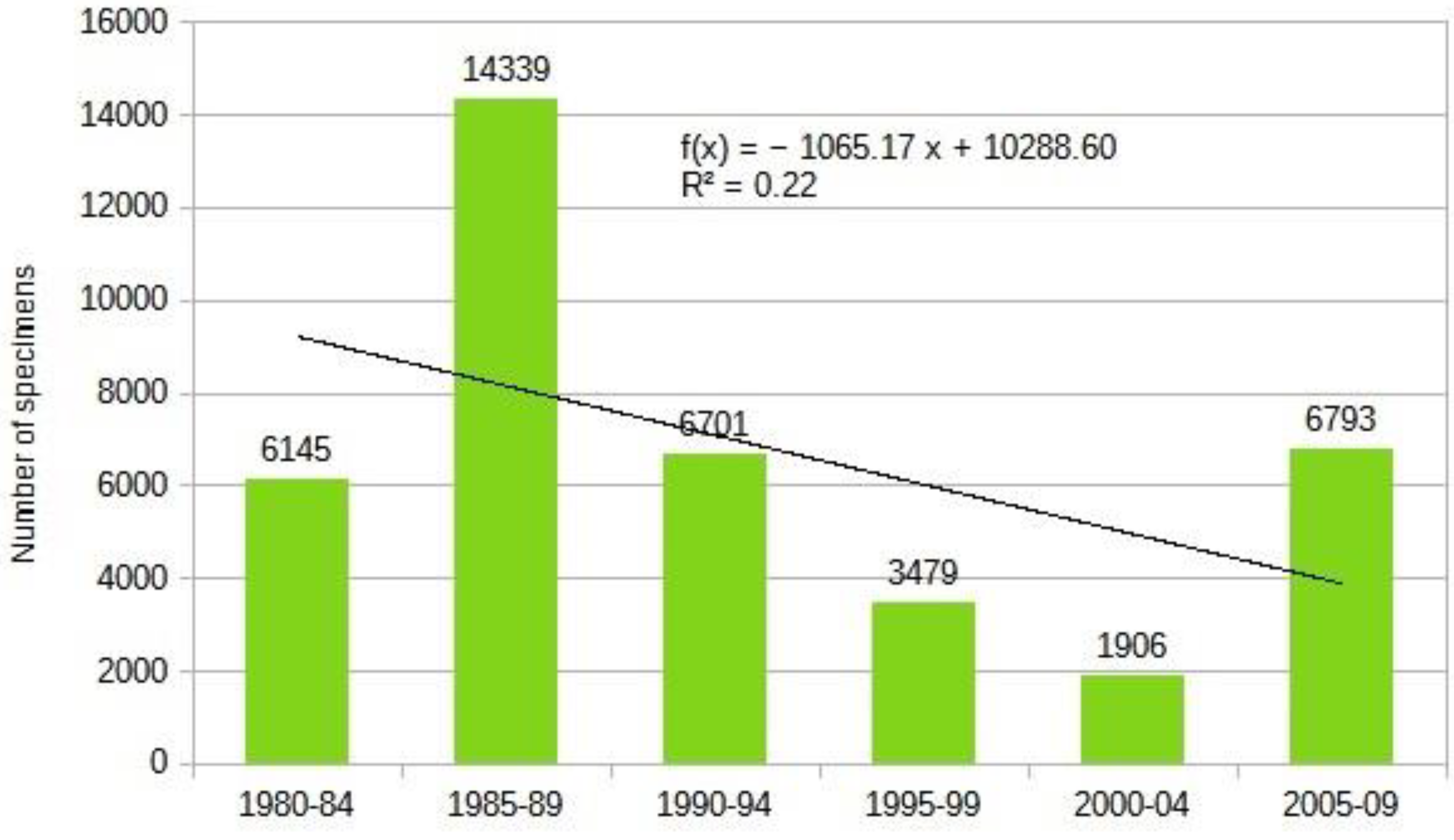
Figure 12.
Great Banded Grayling: Brintesia circe (Fabricius, 1775) has successfully resisted climate change so far (photo: Péter Schmidt).
Figure 12.
Great Banded Grayling: Brintesia circe (Fabricius, 1775) has successfully resisted climate change so far (photo: Péter Schmidt).

Figure 13.
Some butterflies, like the Scarce Swallowtail: Iphiclides podalirius (Linnaeus, 1758), have even been able to increase their population in Central Europe (Photo: Gábor Glemba).
Figure 13.
Some butterflies, like the Scarce Swallowtail: Iphiclides podalirius (Linnaeus, 1758), have even been able to increase their population in Central Europe (Photo: Gábor Glemba).

Figure 14.
Changes in nocturnal macrolepidoptera populations between 1970 and 2022.
Figure 15.
Changes in nocturnal macrolepidoptera populations between 2014 and 2022.
Figure 16.
Changes in species richness of nocturnal macrolepidoptera between 1970 and 2022.
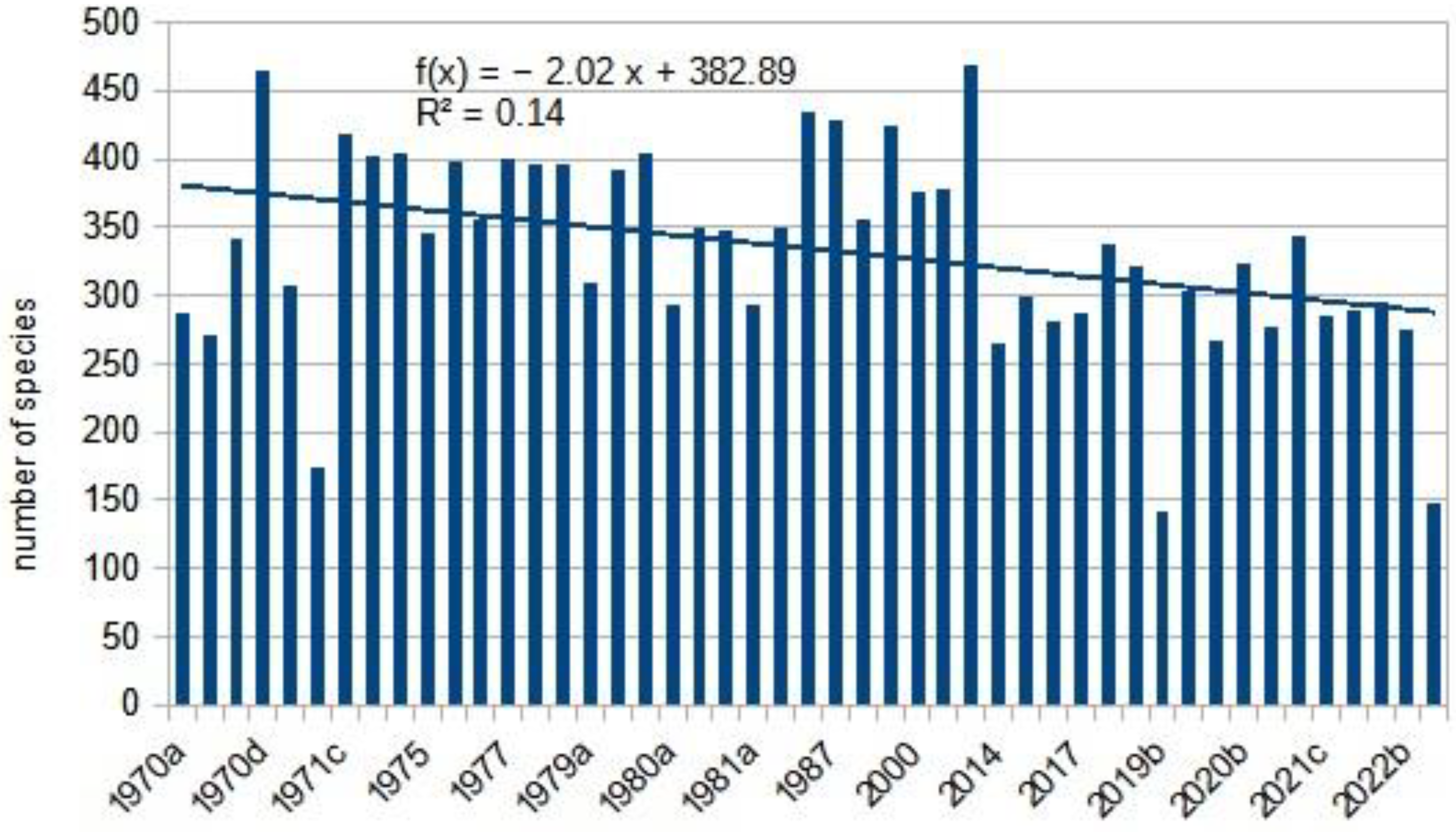
Figure 17.
Timeline of non-native insects in the Carpathian Basin from the Neolithic period till 2024.
Figure 17.
Timeline of non-native insects in the Carpathian Basin from the Neolithic period till 2024.
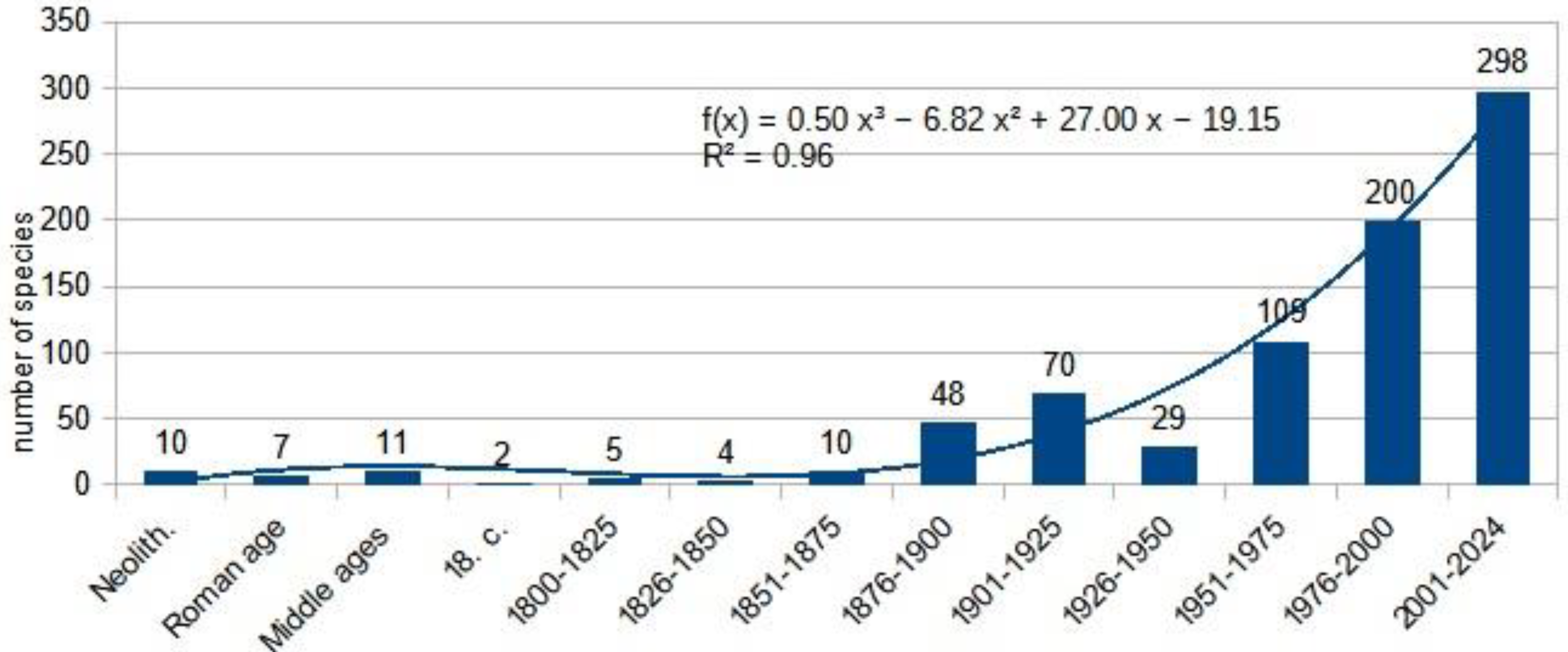
Figure 18.
Timeline of the introduction of tropical insect species.
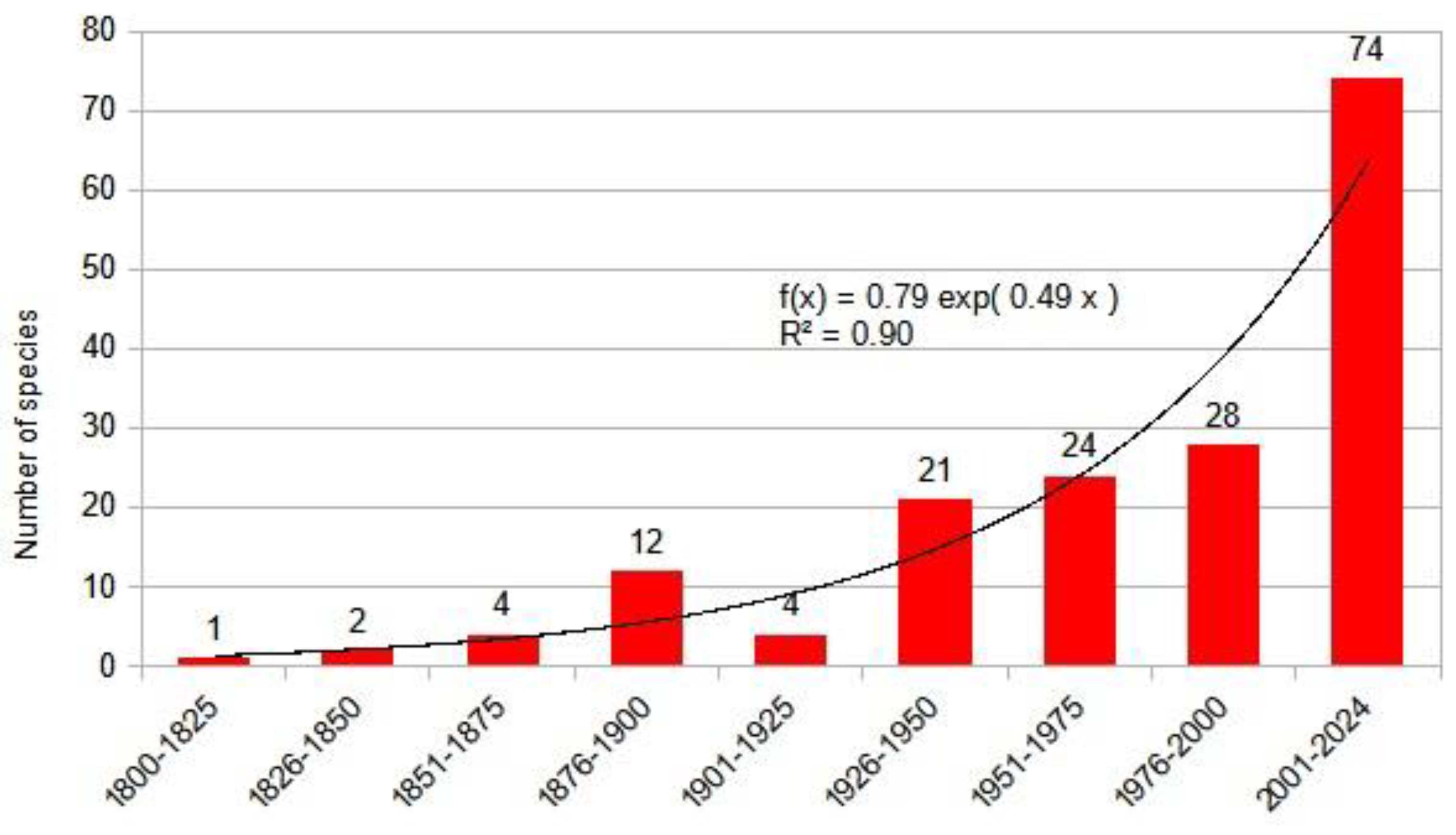
Figure 19.
Division of non-native insects according to their taxonomic groups.
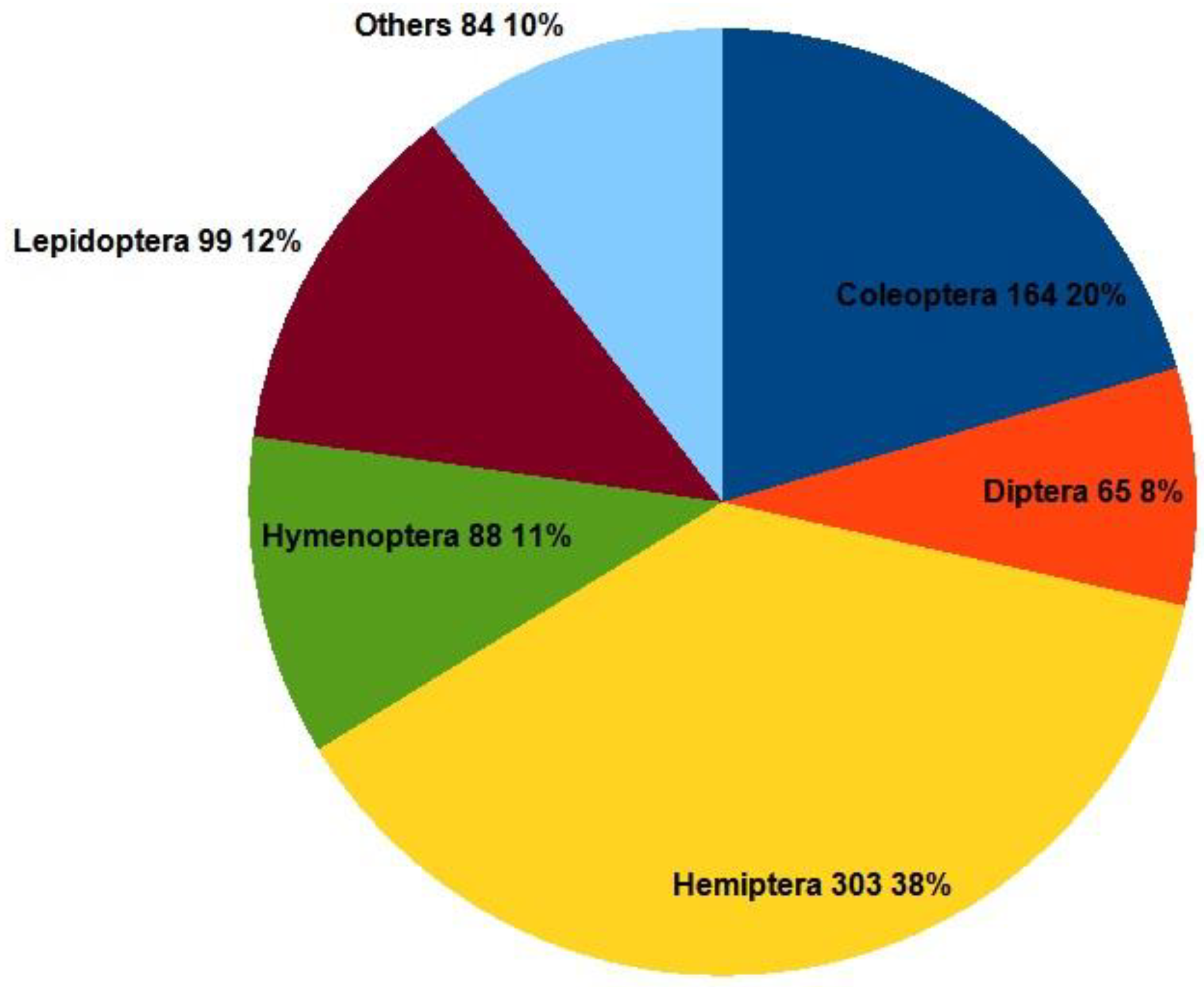
Figure 20.
Division of non-native insects according to their origin.
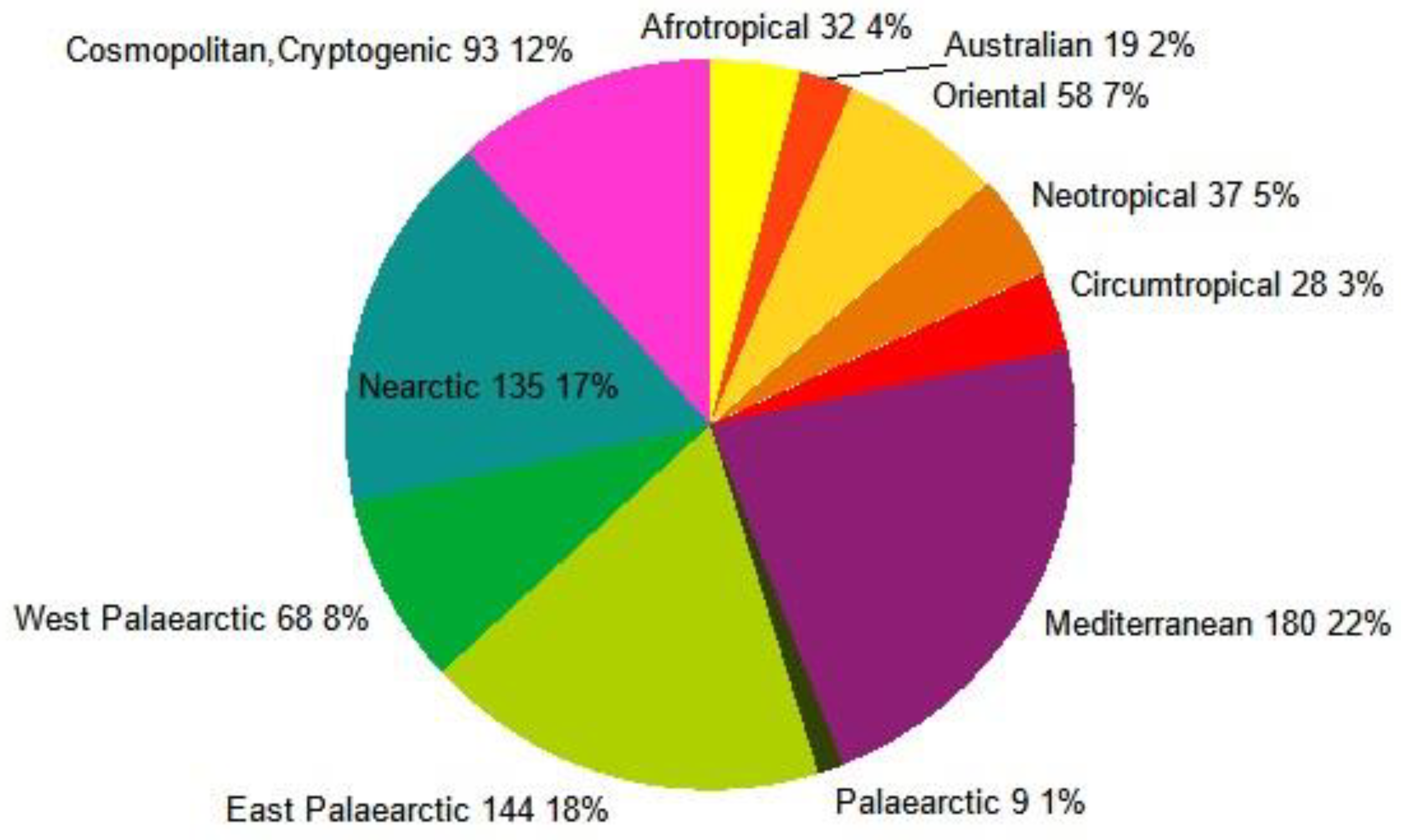
Figure 21.
The Cypress Jewel Beetle: Ovalisia festiva (Linnaeus, 1767) had double colonisation: natural and introduction with ornamental plants (photo: György Csóka).
Figure 21.
The Cypress Jewel Beetle: Ovalisia festiva (Linnaeus, 1767) had double colonisation: natural and introduction with ornamental plants (photo: György Csóka).

Figure 22.
Timeline of the influx of Mediterranean insect species.
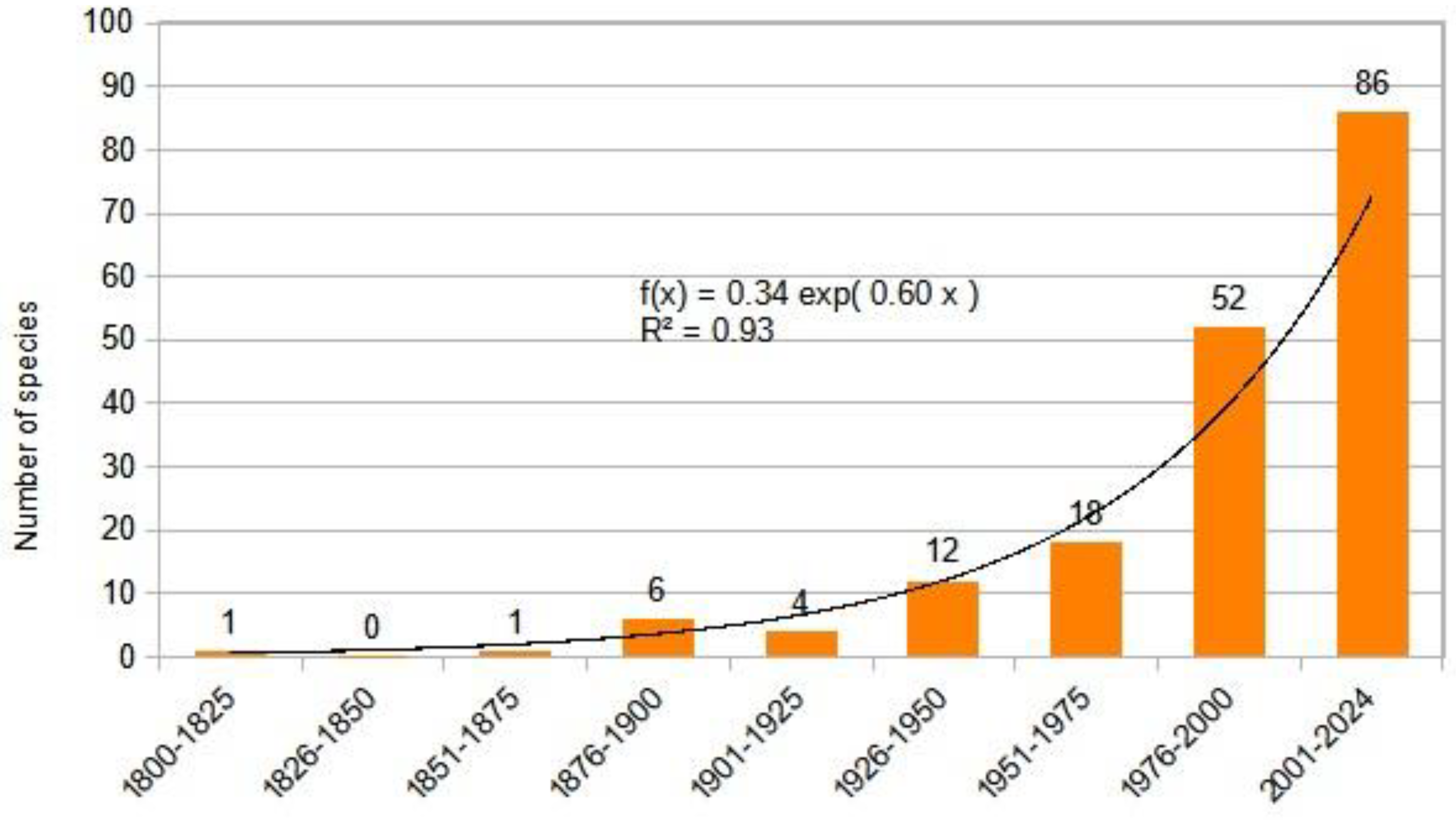
Figure 23.
Long term trend of wild bees in Belgium (after Duchenne et al., [191]).
Figure 23.
Long term trend of wild bees in Belgium (after Duchenne et al., [191]).
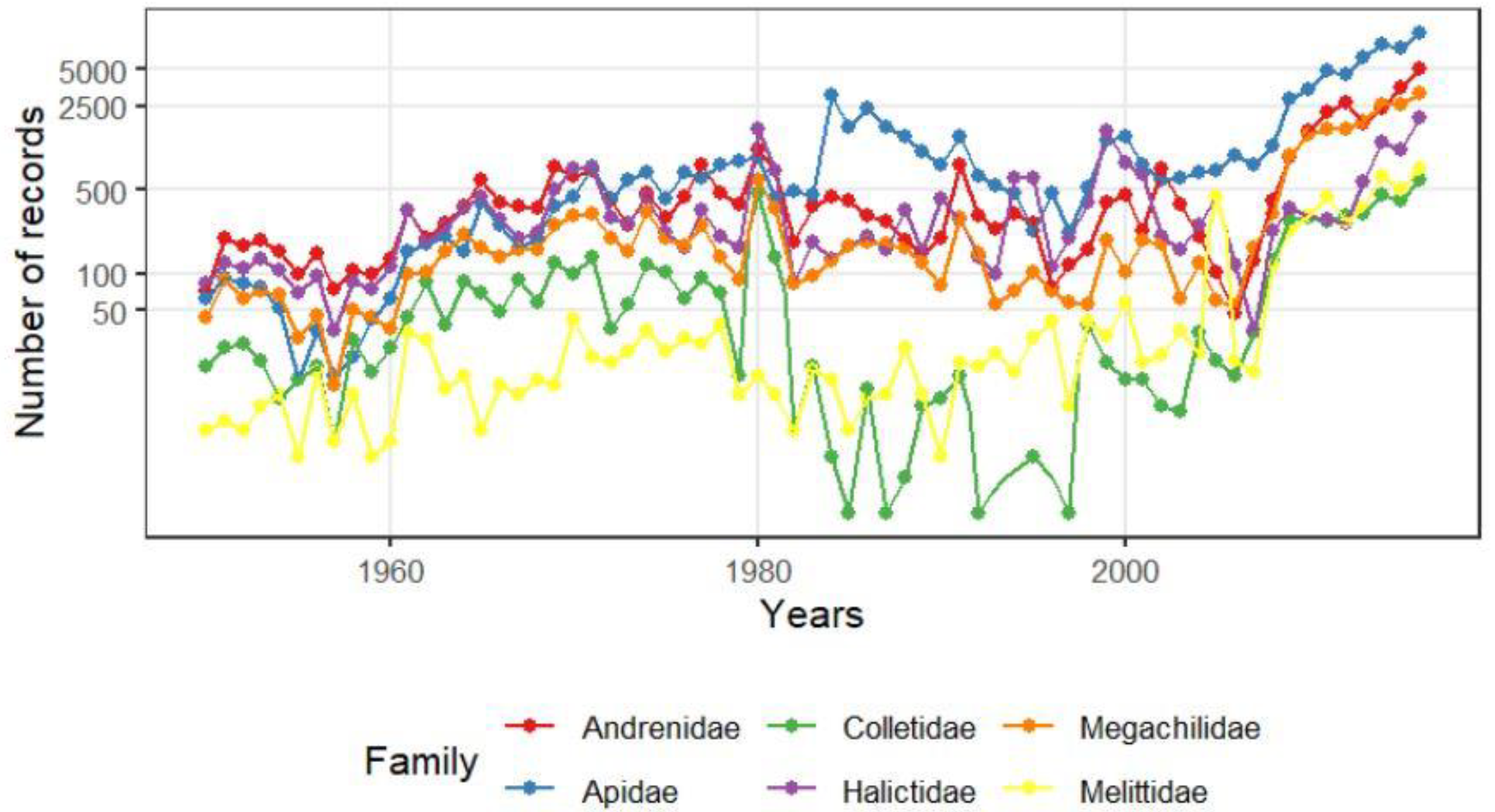
Figure 24.
The Clouded Apollo: Parnassius mnemosyne (Linnaeus, 1758), appears stable in our region (photo: Gábor Glemba).
Figure 24.
The Clouded Apollo: Parnassius mnemosyne (Linnaeus, 1758), appears stable in our region (photo: Gábor Glemba).


Table 1.
Number of individuals of various bumblebee and cuckoo bumblebee species between 2005 and 2023 in the low and high altitudes of the Carpathian Basin (CRC: Climate Risk Category).
Table 1.
Number of individuals of various bumblebee and cuckoo bumblebee species between 2005 and 2023 in the low and high altitudes of the Carpathian Basin (CRC: Climate Risk Category).
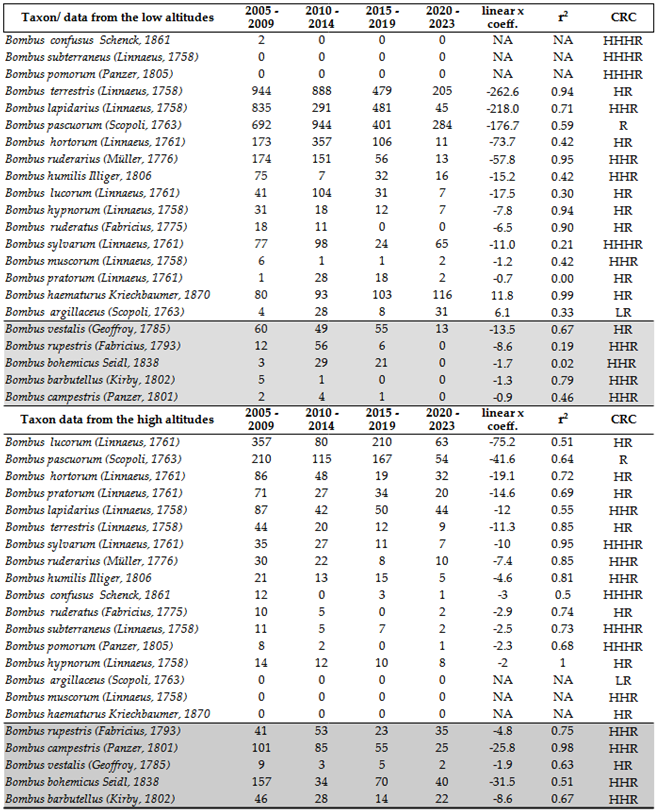 |
Table 2.
Long term trends of various Aculeata genera and families between 1988 and 2023
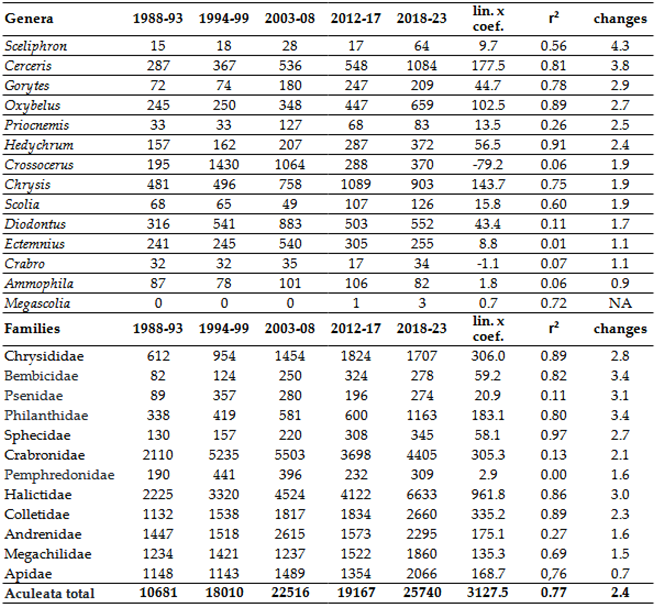 |
Table 3.
Previously sporadically occurring Aculeata species, but not collected for at least 20 years, in the low-lying regions of the Carpathian Basin. Gray background and x-mark: species that still occur in the higher regions and in the northern parts.
Table 3.
Previously sporadically occurring Aculeata species, but not collected for at least 20 years, in the low-lying regions of the Carpathian Basin. Gray background and x-mark: species that still occur in the higher regions and in the northern parts.
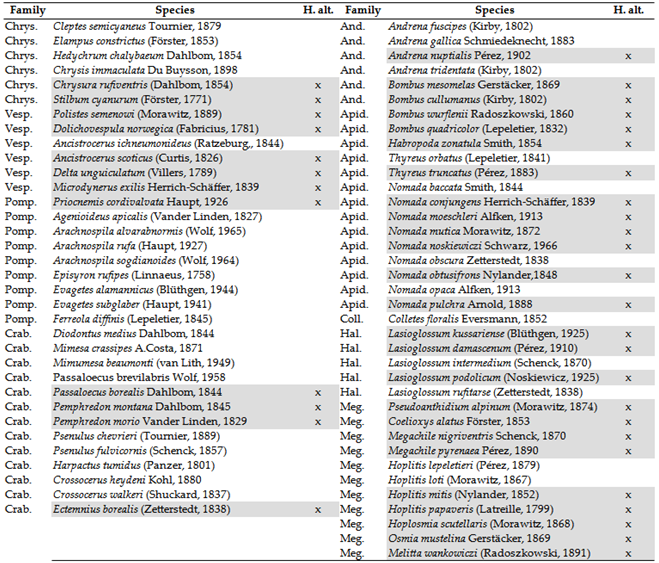 |
Table 4.
Long term trend of sawfly species between 1960 and 2022
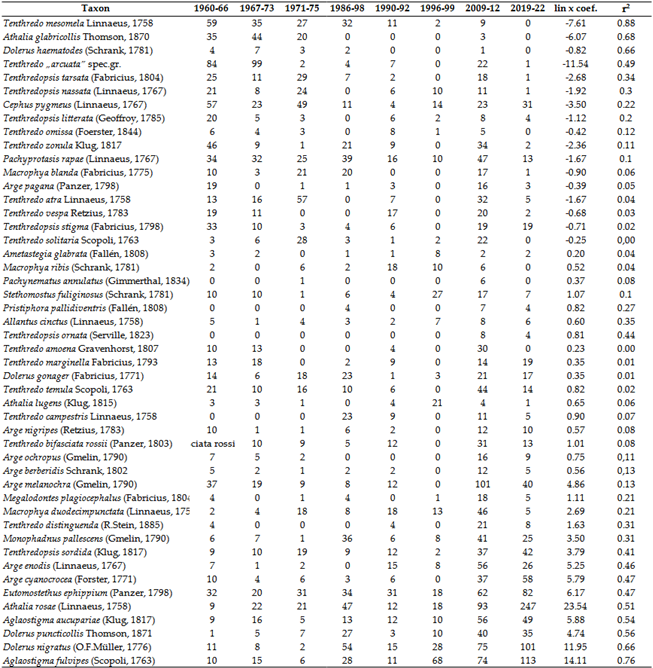 |
Table 5.
Long term trend of horse-fly (Tabanidae) species
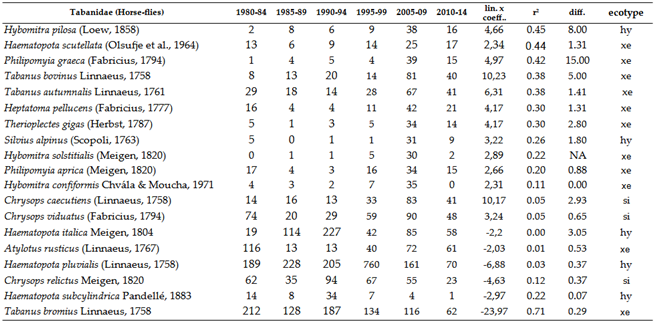 |
Table 6.
Long term trend of hoverfly (Syrphidae) species
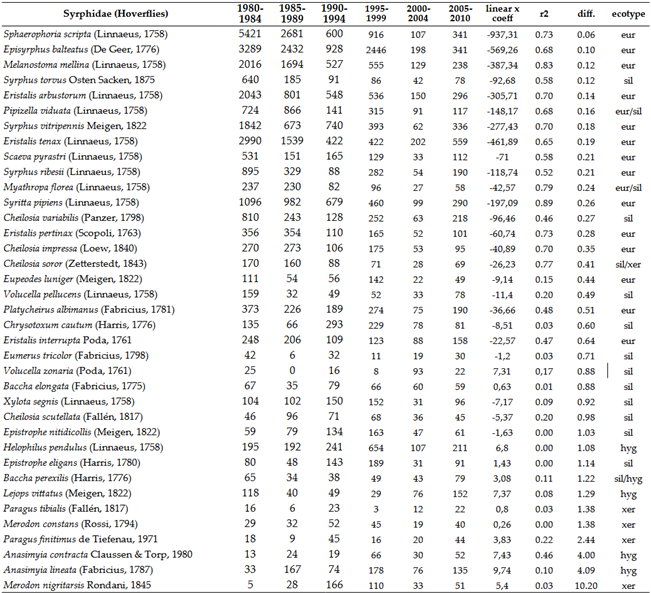 |
Table 7.
Long term trend of bee-fly (Bombyliidae) species
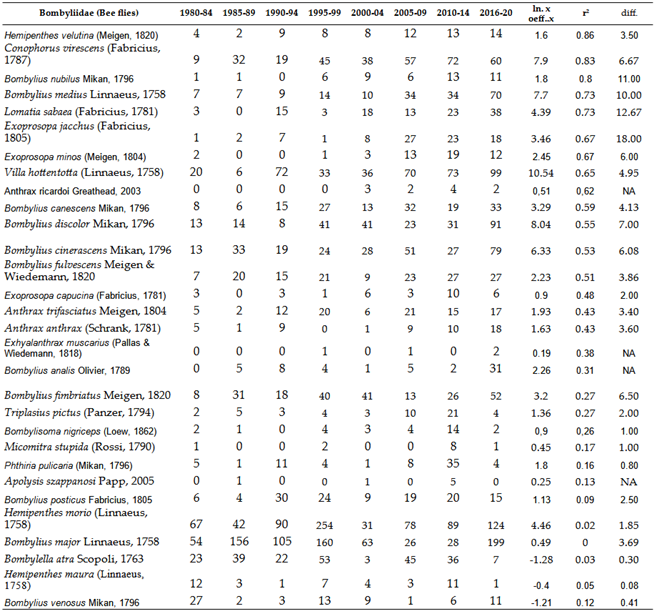 |
Table 8.
Long term trend of tachinid fly (Tachinidae) species
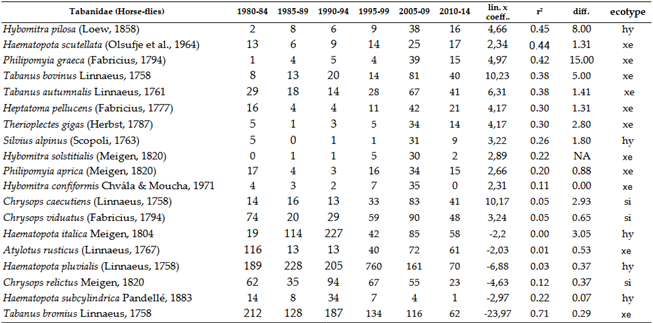 |
Table 9.
Long term trend of various Diptera families
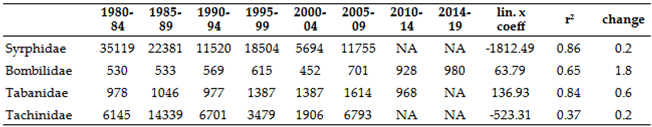 |
Table 10.
Previously sporadically occurring Diptera species, but not collected for at least 20 years, in the low-lying regions of the Carpathian Basin. Gray background and x-mark: species that still occur in the higher regions and in the northern parts
Table 10.
Previously sporadically occurring Diptera species, but not collected for at least 20 years, in the low-lying regions of the Carpathian Basin. Gray background and x-mark: species that still occur in the higher regions and in the northern parts
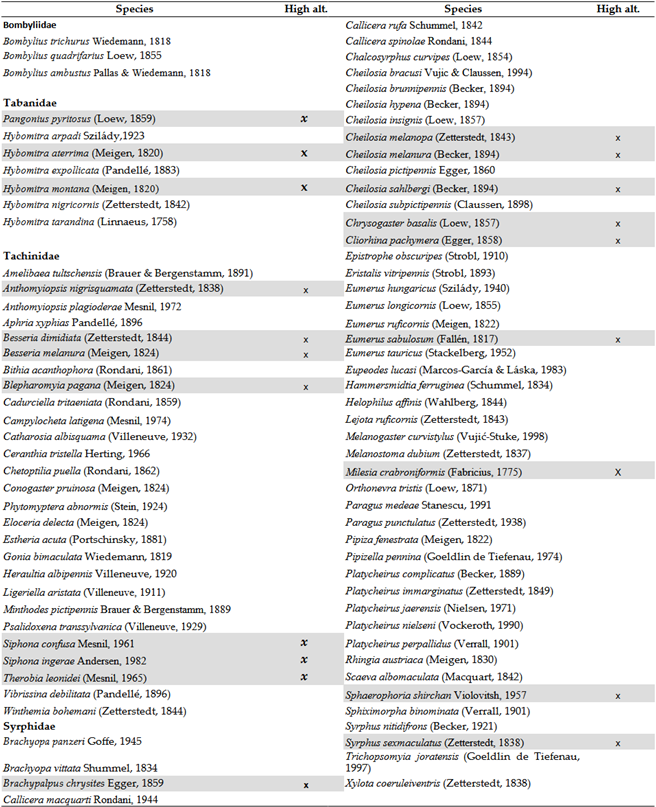 |
Table 11.
Long term trend of selected butterfly species between 1980 and 2009
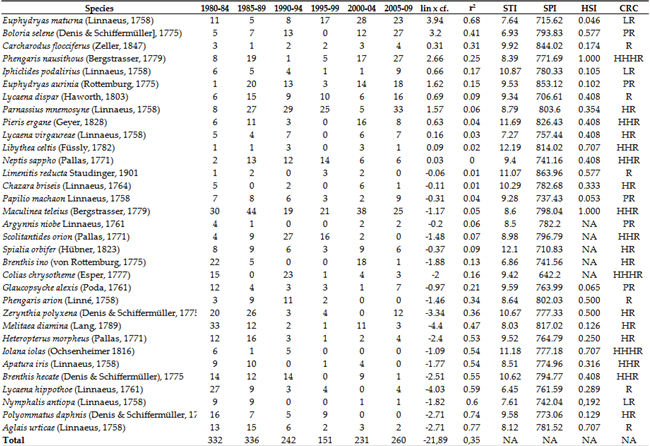 |
Table 12.
Long term trend of selected butterfly species between 2016 and 2023 (Duna-Ipoly National Park)
Table 12.
Long term trend of selected butterfly species between 2016 and 2023 (Duna-Ipoly National Park)
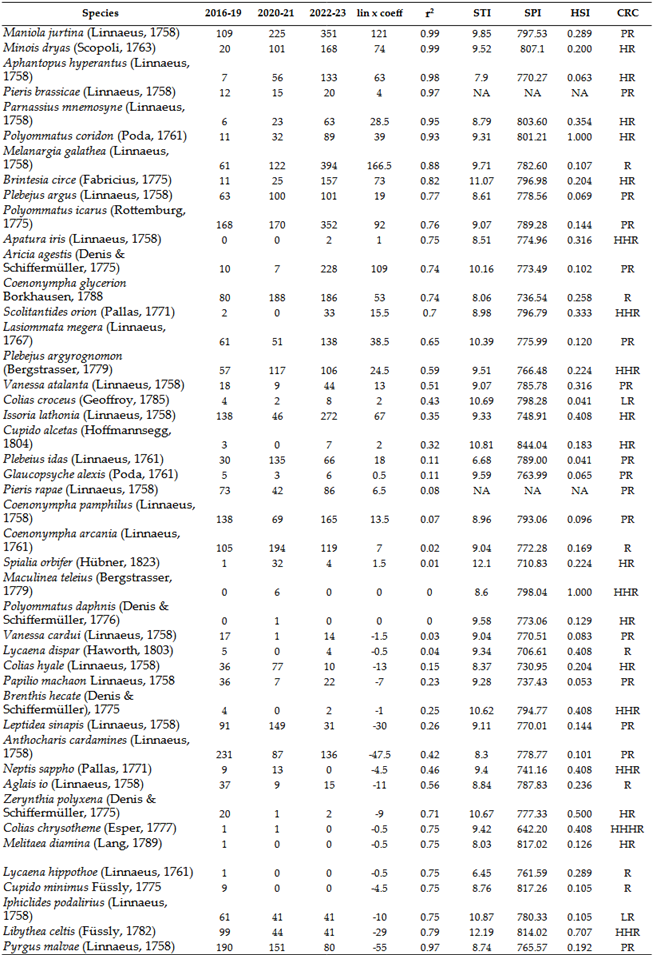 |
Table 13.
Long term trend of number of individuals of butterfly families between 2013 and 2023 (Duna-Ipoly National Park).
Table 13.
Long term trend of number of individuals of butterfly families between 2013 and 2023 (Duna-Ipoly National Park).
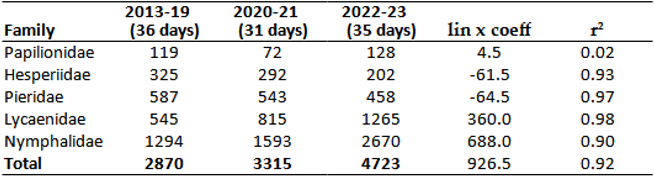 |
Table 14.
Long term trend of number of species richness of butterfly families between 1980 and 2009 low-lying region of the Carpathian Basin.
Table 14.
Long term trend of number of species richness of butterfly families between 1980 and 2009 low-lying region of the Carpathian Basin.
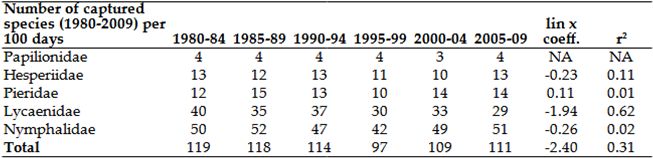 |
Table 15.
Long term trend of number of species richness of butterfly families 2016 and 2023 (Duna-Ipoly National Park).
Table 15.
Long term trend of number of species richness of butterfly families 2016 and 2023 (Duna-Ipoly National Park).
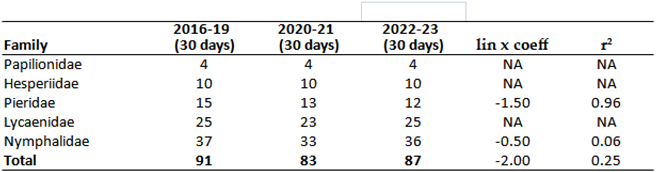 |
Table 16.
Previously sporadically occurring Lepidoptera species, but not collected for at least 20 years, in the low-lying regions of the Carpathian Basin. X-mark: species that still occur in the higher regions.
Table 16.
Previously sporadically occurring Lepidoptera species, but not collected for at least 20 years, in the low-lying regions of the Carpathian Basin. X-mark: species that still occur in the higher regions.
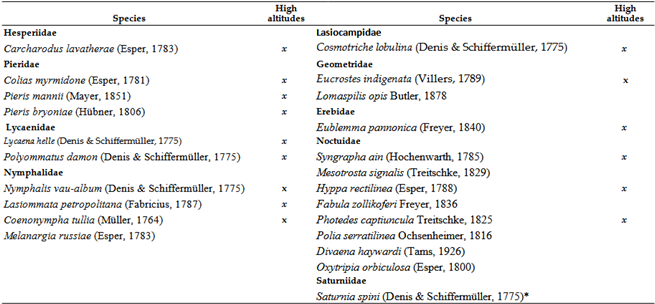 |
Table 17.
Long term trend of number of individuals of nocturnal macrolepidoptera families between 1970 and 2020.
Table 17.
Long term trend of number of individuals of nocturnal macrolepidoptera families between 1970 and 2020.
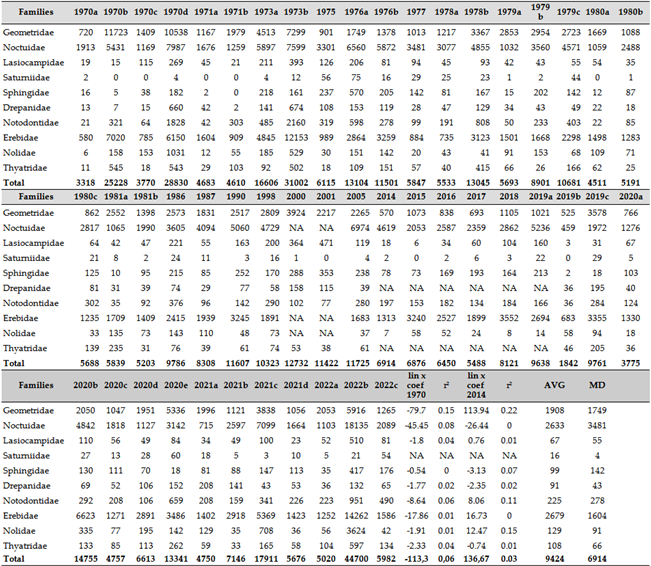 |
Table 18.
Long term trend of species richness of nocturnal macrolepidoptera families between 1970 and 2020.
Table 18.
Long term trend of species richness of nocturnal macrolepidoptera families between 1970 and 2020.
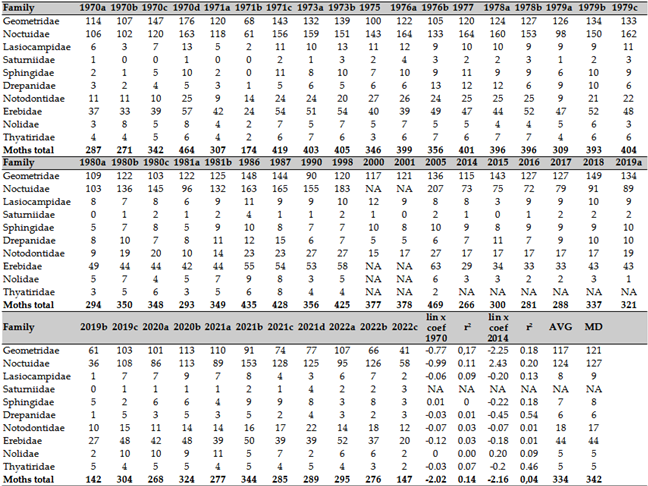 |
Table 19.
Changes in dominant Lepidoptera species over a 50-year period.
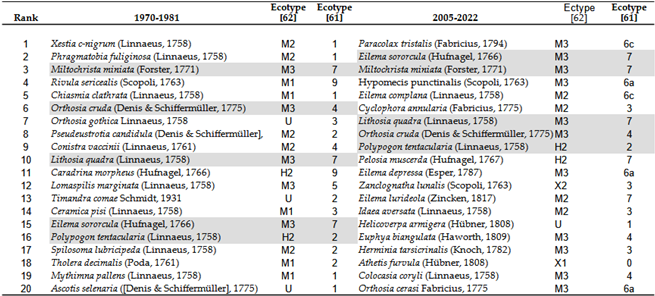 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



