Submitted:
24 April 2024
Posted:
26 April 2024
You are already at the latest version
Abstract
Temporal changes of population densities and species richness of three main pollinator groups: moths and butterflies (Lepidoptera), bees, wasps and sawflies (Hymenoptera) and hoverflies, horse-flies, tachninids and bee flies (Diptera) were investigated in the Carpathian Basin. Maintaining pollinator divesrity is a crucial factor for preserving our biodiversity and ecosystems, furthermore several pollinator species has strong economic role in maintaning crop and fruit cultures. Our conclusions are based on our three or four decades of faunistic surveys in various regions of the Carpathian Basin. Analysing and comparing our data with the historical data of the last 50 years, we concluded, densities of some pollinators were declined during the past decade and the half (Symphyta, hoverflies), although populations of several species of Mediterranean-origin were grown (Aculeata) and even new species were migrated from the warmer regions. In numerous cases, this decrease was dramatic: more than 90% decrease of certain butterfly species were detected. On the other hand, the composition of pollinator fauna significantly changed due to the disappearance of some montaneous or mesophyle species. The main reason of decrease of pollinator communities is partly the climatic change and partly anthropogenic factors. Our conclusion: in our region, the pollinator crisis is present, but moderate; however, there is clear sign of the gradual transition of our pollinator fauna towards the Mediterranean type.
Keywords:
pollinators
; hymenoptera
; Diptera
; lepidoptera
; carpathian basin
; population density
; species richness
; climate change
; global warming
1. Introduction
“The apple trees were coming into bloom but no bees droned among the blossoms, so there was no pollination and there would be no fruit. The roadsides, once so attractive, were now lined with browned and withered vegetation as though swept by fire. These, too, were silent, deserted by all living things. Even the streams were now lifeless.”(Rachel Carson: Silent Spring)
In 1962, Carson predicted the future quiet spring, when neither the noise of bumblebees nor the song of birds would disturb the peaceful growth of green vegetation. The very beginnings of social environmental movements are linked to this iconic work. Today, after 62 years, we have reached a point where the nightmare of silent spring has come within reach. The gradual disappearance of pollinators threatens to become an ecological catastrophe, since pollinators play fundamental role in maintaining our ecosystem. The reproduction the vast majority of cultivated and wild plants depends on the pollination activity of various animals (mainly insects). In recent years, we have seen a significant decline of various pollinator groups. Although the causes of this decline are not yet fully understood, the consequences are commonly referred as a pollination crisis [1]. This pollination crisis goes far beyond the reproductive biology of plants, as Rhodes [2] writes: “The decline in the health and abundance of pollinators may significantly threaten the integrity of global biodiversity, the inclusion of food webs, and risks to the health of humans and other animals”. Ultimately, the pollination crisis may cause a global food crisis and a social crisis at the same time. It draws attention to the vulnerability of food chains. Here in our region, we can see this, particularly in the decline of several songbirds (e.g., swallows), since pollinators play role not only in pollination, but they are essential food source for many bird species through their biomass [3]. According to the IPBES report [4], pollinator-dependent plants contribute to 35 per cent of global crop production. According to Dicks et al. [5], the pollination crisis primarily affects the Global South. The risk of pollination crisis in our region is moderate, but at the same time, the decline in diversity of certain groups seems to be truly tragic, as our results show. Of the 8 reasons analyzed, they cited changes in land use, climate change, pollution and the spread of invasive species. Other scientists [6,7], also emphasize habitat fragmentation and degradation, as well as the excessive and inappropriate use of pesticides and herbicides as the main causes of the pollinator crisis. All other reasons are listed as ‘other factors’.
The 2021 EU directive on the protection of pollinators, entitled: “Protecting pollinators in the EU” [8], is a response to the current situation, which includes the development of action plans and monitoring programs, the provision of resources, and extension of the ban on certain pesticides such as imidacloprid and all neonicotinoid-based pesticides containing clothianidin and thiamethoxam.
In our work, we monitor the changes in population densities and species richness of different groups of wild pollinators in Hymenoptera, Diptera and Lepidoptera during the last half century. We also study pollinators at the species level either. In this study, only those species were selected that were once common or widespread, but now, they are less common or sporadic; or conversely, those species that are winners of the changes and whose populations are increasing due to recent climatic circumstances. We have also compiled a list of sporadic species of several groups that we had not been able to collect in the last 20 years, even though, they were not rare at all earlier. We have also included species that their populations were increased in the recent decades. These newly spreading species may have arisen due to the expansion of their range (as a result of global warming) or introduced by humans from distant regions of the Earth.
2. Materials and Methods
Our results are based on regular faunistic surveys carried out over the past 50 years in diverse areas of the Carpathian Basin (Pannonian Basin). This kind of tradition of faunistic investigation can be documented back to the time of Scopoli (Observartionis Zoologicae). This time, we compare quantitative and qualitative faunistic data from the 1970s and 1980s with our own recent and sub-recent data from the last 2-4 decades. Statistical analysis of these data determine various trends. These trends either confirm or reject our initial hypothesis regarding the pollination crisis and as a final result, figures out the direction of pollinator changes and point out those pollinator groups and species that suffer from the recent changes or have benefit from these. Finally, we attempt the explanation for these changes based on our field experiences.
Data Selection
Our present work has two main sources: unpublished databases from diverse areas of the Carpathian Basin spanning 4 or 5 decades. Voucher specimens are deposited in various natural science collections of the region (Zoological Institute at Bratislava, Rippli-Rónai Museum at Kaposvár, Natural History Museums at Zirc and Budapest). The second source is the published faunistic papers.
Majority of our data are original, results of 50 or 40 years of continuous collections covering the Pannonian biogeographical region. These databases contain circa 100 000 Aculeata data, 45,000 Diptera data, and 40,000 Lepidoptera data. These databases, wherever possible (Hymenoptera, Diptera), are results of the work done by one-one specialist over several decades (Dr. Sándor Tóth, Zsolt Józan, Dr. Ladislav Roller and Dr. Attila Haris), consistently using the same methods. To supplement these databases, we used the following publications dominantly from the precvious authors: Symphyta: Zombori [9,10,11,12,13], Haris [14,15,16,17,18,19,20,21,22,23], Haris et al. [24] and Roller [25,26]; Aculeata: Józan [27,28,29,30,31,32,33,34,35,36,37,38,39,40]; Diptera: Tóth [41,42,43,44]; Lepidoptera: Ábrahám [45,46,47,48], Ábrahám et. al. [49], Ábrahám and Uherkovich [50], Uherkovich [51], Pillich [52], Sáfián1 [53], Ács et al. [54], Čanády [55], Dietzel [56], Sarvašová [57], Gergely [58], Gór [59], Schmidt [60], Németh [61], Szabóky et al., 2014[62], Hudák [63], Varga et al. [64], Árnyas et al. [65] and Kovács [66].
We selected those data only (in Hymenoptera and Diptera) which were results by comparable and regular field recordings, 30-35 field days per year, and we omitted results of non regular collections and scattered data, as these data cannot be processed statistically. The analyzed species were selected according to the following criteria: they should have important role in pollination (i.e., rare species were excluded) and we highlighted those species whose change proved to be the strongest. In case of light trap data series, we checked the beginning and the end of time series to see which species occurred in the largest number in the beginning and at the end of the studied 50 years period from 1970 till 2022. After this, we analyzed those species, that provided the most significant changes over the past 50 years. Since the scope of our work does not allow the analysis of circa 2,500 Hymenoptera, Diptera and Lepidoptera species, and due to lack of space and opportunity, we restricted it to 15–40 species from each group, especially for those species where the changes were outstanding or for those which we found characteristic for the total group or play important role in pollination due to their population densities.
Changes in Methods and Their Statistical Balancing
In nocturnal lepidoptera, methodological changes had to be taken into account. From 2014, UV Led light trap used which replaced the black light UV 20 W (between 1990 and 2010) and Jermy type light trap with 125W mercury vapor lamp (used between 1970 and 2010). These different light sources had different selectivity according to Infusino et al., and Pan et al. [67,68]. This is the reason, that we calculated trends from 1970 and a separate trend from 2014 to draw reliable conclusions. From light-trap data, we selected only those data-series, which span 6 months of intensive, daily collections generally from April to October, in few cases from May to November. Light-traps worked continuously during these periods. Other, shorter or incomplete light trap data were excluded from the analysis.
Regarding diurnal lepidoptera, we have minimal original data. Regular butterfly monitoring started only in the last decade in our region, Therefore we have to rely on processing of previously published datasets. We analyzed the earlier commonest pollinator species (but not all), where, (according to our field experiences), we noticed the greatest changes. In absence of reliable quantitative data, we used the method followed by Ábrahám [47] as follows: 0: the species is not present or has disappeared:, 1: rare, 2: sporadic, 3: occasionally frequent, 4: generally frequent, 5: common. Other methods, finally, had to be rejected: database of the European Butterfly Monitoring Scheme - eBMS contains different amounts of observations from various years. For example, we have approximately 600 observations from Transdanubia in 2018, and 2 280 from the same region in 2023. We also attempted to compare the amount of diurnal butterflies collected during an average collecting day. These data have high standard deviation, without showing any trend, e.g., Csombárd 2015: [60] 330 individuals per day on average, Zvolen between 2009 and 2011: 143 individuals per day on average [57], Tapolca between 1977 and 1990 approximately 28 individuals per field-day [61]. In other words, these butterfly surveys proved to be completely useless. In this way, the firstly mentioned approach with its many subjective elements remained the only one possibility for us.
Data processing
At Hymenoptera and Diptera, we formed our data into 5 or 6 years batches (providing even time-intervals). After plotting these data-groups on bar graph, we added trend lines including trend equation and coefficient of determination (r2 value). In nocturnal Lepidoptera, instead of creating batches, we analyzed separate data-series of each location (5,000-15,000 specimens per location per 1 year) throughout the various ecosystems of the Carpathian Basin. These data series are dating back to 50 years except 90s (period of changing the political system in this region) when the light trap network was suspended and we have 10 years gap this time. After arranging these data series in chronological order and creating the bar charts, trend-lines wer drawn (mostly linear trends) and the trend-parameters were recorded. These parameters are the slope of trend (linear x coefficient) and the coefficient of determination. The linear x coefficient indicates the direction of the trend (minus: decrease, plus increase), and its value indicates the slope of the trend line, i.e., the intensity of change (for some Syrphidae species, this trend is exponential). Coefficient of determination indicates how strong is the trend and in how our data fit to the trend-line i.e., in case of high proportion of continuous temporal changes (real trend: high r2 value) and r2 value is low, when spatial or cyclical variables has main role in the changes of populations or species richness.
Sampling Sites
Diptera and Hymenoptera (except Symphyta) collected from approximately 1 200 various locations from all parts of the Pannonian biogeographic region. These are smaller samples (1-50 specimens) from ech location.
Lepidoptera and Symphyta specimens are from separate and significantly lower number of locations (circa 50 locations sampled in different years), as listed below. These are larger samples, between 500 and 30 000 specimen from each location (Symphyta samples are smaller, between 500 - 4 000 per location, Lepidoptera samples are larger, 5 000 - 30 000 specimen per location per 1 year).
Collecting sites: Symphyta: Nagykovácsi and its surroundings, Aggtelek National Park, Fertő-Hanság National Park, Zselic Hills, Keszthelyi Hills, Cserhát Hills, Vértes Hills, Southern Transdanubia at River Drava, North Somogy, Börzsöny Mountains, Szeged and its surroundings, South Somogy, Ivanka pri Dunaji, Javorina, Mošovce, Pernek, Devin, Hriňová, Stefanova, Horša, Bokroš, Tvrdošovce, Virt and Malacky regions. Applied methods were sweeping net and Malaise trap.
Collecting sites:: Lepidoptera: Felsőtárkány, Tompa, Gilvánfa, Magyarszombat, Mike, Vásárosbéc, Almamallék, Palé, Lipótfa, Aggtelek, Tompa, Répáshuta, Sopron, Plain of River Dráva, Bakonynána, Boronka, Tapolca, Keszthelyi Hills, Aggtelek National Park, Biatorbágy, Pomáz, Székesfehérvár, Őrség, Csombárd, a Sliač in Zvolenská kotlina and surroundings of Košice. Applied methods were light trap and sweping net.
3. Results
Symphyta
Of the 797 species living in the Carpathian Basin, 376 are rare, 277 sporadic and 144 frequent or common. The last 3 categories (sporadic, frequent and common) can be considered as important pollinators. Similar to Aculeata species, we can observe an increase of species are predominant in the Mediterranean region (Table 1 and Table 2). On the other hand, we found significant decrease in most of sawfly species, which is clearly evident from the Malaise trap data (Table 2 and Figure 1). In two countries, we measured changes in individual numbers and species richness over 5 and 3 decades, respectively, using two different methods. We have Malaise trap data from Slovakia and sweeping net data from Hungary, which are shown in Table 1 and Table 2 and in Figure 1. Although there are differences between the two areas and the two methods, the similarities are: increase of Mediterranean sawfly species and gradual decline in number of individuals of most sawfly species. Species that are tolerant to climatic changes are: Tenthredo distinguenda (Stein, 1885), Arge nigripes (Retzius, 1783), Arge ochropus (Gmelin, 1790), Arge cyanocrocea (Forster, 1771). These species mainly dominate the fauna of the Mediterranean region and Anatolia. Strongest decline has been observed in moisture-loving species such as Tenthredo mesomela Linné, 1758. However, as suborder Symphyta reaches its maximum species richness and abundance in northern Europe, the negative trends appear realistic and are indication of the Mediterranean transformation of our pollinator fauna. As a matter sporadic species (regularly collected, but in low 1-3 number per year), we have listed in Table 3 those species that have not been collected in the Carpathian Basin in the last 20 years. These species were not rare in the middle of the 20th century (until the 1970s), but despite our efforts, we have not succeeded to collect them in the area of the Pannonian biogeographical region. It seems, these species haven’t been disappeared but their populations have fallen below the detection limit. Of the 41 species listed, 9 were not found in the entire Carpathian Basin, while 32 species were found in the high-altitude regions (alpine and subalpine) of Slovakia. The list shows that Tenthredo species and several Nematinae species (genera Pristiphora and Pteronidea) have become rare. The decline of the northern Nematinae subfamily is explained by global warming. The reasons for the rarity and disappearance of the Tenthredo species are still unclear. As for the Tenthredo species, similar observations were made by Goulet in Canada [69]. We can say: sporadic species listed in Table 3 are now rare. So far, we have not detected any species spreading from south to north that have appeared in recent decades, although the occurrence of some southern species such as Macrophya superba Tischbein, 1852 can be expected in the near future. In addition, the population densities of Tenthredo bifasciata ssp. bifasciata O. F. Müller, 1776 and Tenthredo costata Klug, 1817 are expected to increase. M. superba reaches its distribution area at the very southern border of the Carpathian Basin [70]. T. costata has the northernmost limit of its distribution range in our region, this is the reason, that its populations are subject of fluctuation in our region. Several specimens of T. costata were captured in the 70s and after long disappearance, it was captured again 2 years ago in Nagybajom, Southern Transdanubia. T. bifasciata ssp. bifasciata was recently recorded in Slovakia [70]. It reaches its highest density in the Anatolian region [71], we can expect its increase in our region as well. In the fauna of the Carpathian Basin, an invasive species, Aproceros leucopoda Takeuchi, 1939, appeared in the early 2000s. Overall, based on our Malaise trap data series, we have observed a strong decline in sawflies over the last 3 decades, which is not surprising if one knows the ecological needs of this group (Table 2 and Figure 1).
Aculeata
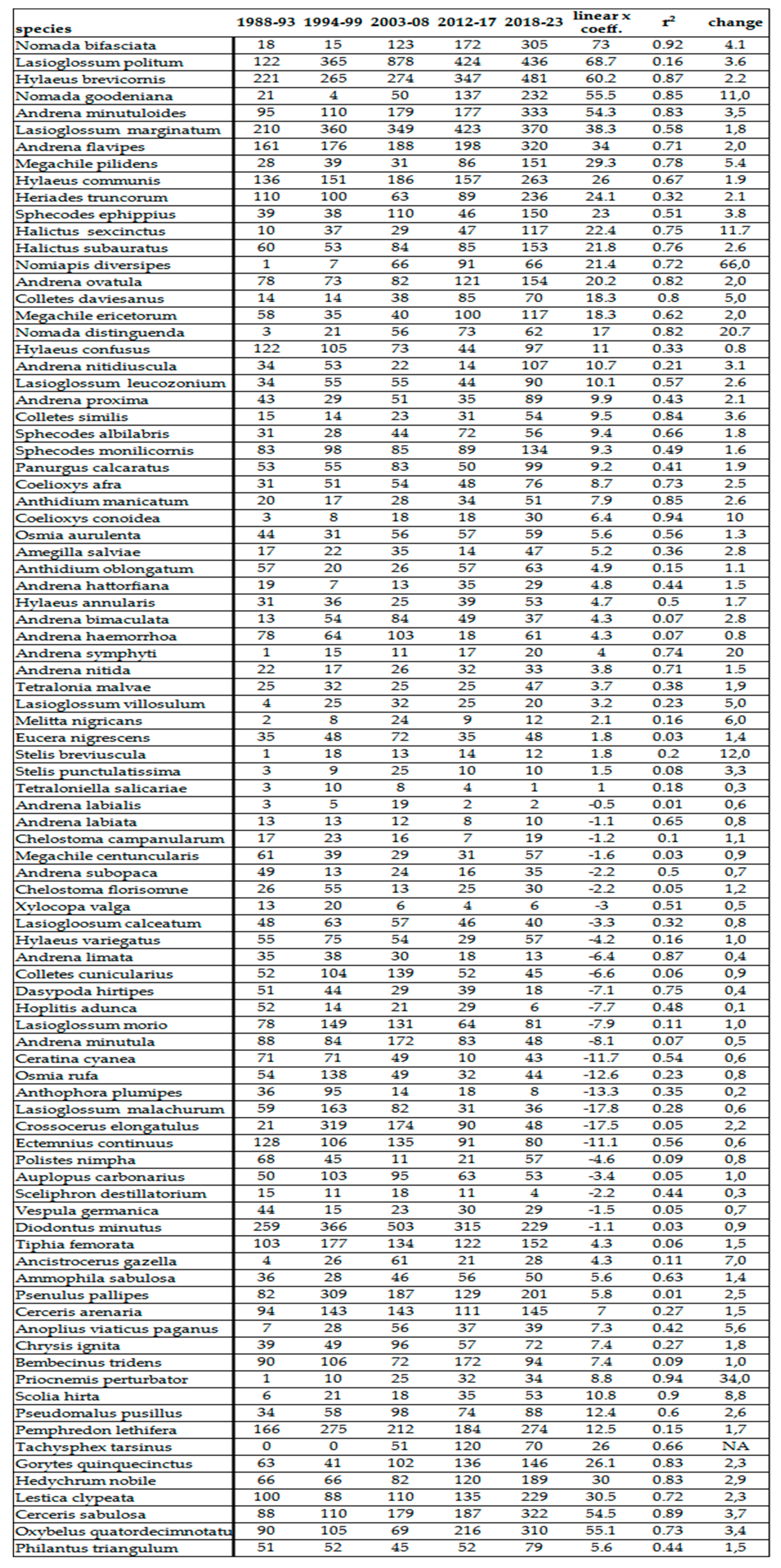
Population densities of most Aculeata groups have intensively increased in the last 40 years (Figure 2). In Table 4, we listed those species which have changed the most in number of individuals. In wild bees, the most intense increase was observed at Nomiapis diversipes (Latreille, 1806), Nomada distinguenda Morawitz, 1874 and Halictus sexcinctus (Fabricius, 1775) (Table 6). Since the main distribution area of these species is in the Mediterranean biogeographic region, they are excellent indicators of global warming. In other groups of Aculeata, Priocnemis perturbator (Harris,1780), Scolia hirta (Schrank 1781) and Ancistrocerus gazella (Panzer, 1798) also produced outstanding growth (Table 6).
In terms of proportions, the average population-increase of these species compared to the first half of the 80s are between 1.6 and 3.8x. In the Carpathian Basin, Nomiapis diversipes (Latreille, 1806), Nomada distinguenda Morawitz, 1874, Coelioxys conoidea (Illiger,1806), Andrena symphyti Schmiedeknecht, 1883, Lasioglossum villosulum (Kirby,1802), Melitta nigricans Alfken 1905 and Stelis breviuscula (Nylander,1848) were rare till the 1980s,,but now, they are sporadic or even frequent species.
In contrast, certain moisture-loving or mountainous species became particularly rare, or their populations are decreased from frequent to sporadic such as Anthophora plumipes (Pallas,1772), Ceratina cyanea (Kirby, 1802), Dasypoda hirtipes (Fabricius,1793) and Andrena limata Smith, 1853.
Decline of bumble bees are discussed separately in the next entry. In other non bee groups of Aculeata (Crabronidae, Philanthidae, Scoliidae, Pompilidae, Chrysididae, etc.), this increase was between 1.5 and 3.4x. Some species of Crabronidae associated with the wet conditions of marshy meadows, such as Ectemnius continuus (Fabricius,1804), or saprophilic Hymenoptera which are associated with old trees and forests, like Xylocopa valga Gerstaecker 1872 or Crossocerus elongatulus (Vander Linden,1829), became significantly rarer.
At generic level, we have obviously experienced similar changes. The most striking increase (3-4x) was observed in genera Priocnemis, Oxybelus, Gorytes, Cerceris, Sceliphron and Megascolia (Table 4). Decline of the moisture-loving and saproxyl species observed only at species level.
According to our observations, certain xerotolerant, Mediterranean species are gradually spreading north. These species are: Cerceris rubida (Jurine, 1807) (Philantidae), Chrysis taczanovskii Radoszkowski, 1876 (Chrysididae), Megascolia maculata (Drury, 1773) (Scoliidae), Colletes hederae Schmidt & Westrich , 1993 (Colletidae). Nomiapis bispinosa (Brullé, 1832) (Halictidae) Pasites maculatus Jurine, 1807, Scolia galbula (Pallas, 1771), and Scolia hirta (Schrank 1781) (Scoliidae) (Table 5). The last one was recently discovered in Slovakia. Until now the northern border of its distribution was Hungary, inside the Carpathian Basin. Furthermore, 2 species with originally Mediterranean-distribution, namely Lasioglossum griseolum. (Morawitz 1872) and Heriades rubicola Pérez, 1890 were subrecently captured in Slovakia [80,82] in 2014 and 2009.
Non native species are also enriched our pollinator fauna. This enrichment has 2 ways: one is the expansion of the area of certain species due to climatic change or introducing species from distant regions by human activity. The following species reached the Carpathian Basin from the Mediterranean region (Table 5): Sceliphron madraspatanum (Fabricius, 1781) (Sphecidae), Diodontus brevilabris Beaumont, 1967 (Pemphedronidae), and Chelostoma styriacum M.Schwarz & Gusenleitner, 1999 (Megachilidae). On the other had, Sceliphron curvatum (F. Smith 1870), Sceliphron caementarium (Drury, 1773), Isodontia mexicana (Saussure, 1867) (Sphecidae), Megachile sculpturalis Smith, 1853 (Megacjilidae) and Vespa velutina (Lepeletier, 1836) (Vespidae) were introduced from distant regions of the Earth (Table 5). Among these, Sceliphron curvatum (F. Smith 1870), Sceliphron caementarium (Drury, 1773) and Isodontia mexicana (Saussure, 1867) were succeed to be frequent (Table 5). We managed to collect only 4 specimens of Megachile sculpturalis near Harta (Bács-Kiskun County). These invasive species expanded quickly in the southern and moderately quickly in the northern area of the Panonian biogeographic region (comapre Hungarian and Slovak data in Table 5). As a matter of the rest, only the voucher specimens were captured.

Table 6.
Number of individuals of various Aculeata species between 1988 and 2023.
Bumblebees (Bombus spp.)
Trend of population densities are strikingly different from the majority of other Aculeata species, this is the reason that we discuss separately the true bumblebees and also their social parasites the cuckoo bumblebees (Table 7). Their trends are opposite to other Aculeate species. Almost all bumblebee species have experienced significant decline. According to our data, only Bombus argillaceus (Scopoli, 1763) went through a moderate and Bombus haematurus Kriechbaumer, 1870 an intensive increase in populations. Numerous bumblebee species such as Bombus confusus Schenck, 1859, Bombus subterraneus (Linnés, 1758) and Bombus (Thoracobombus) pomorum fall below the detection limit in the Pannonian biogeographic region. Particularly interesting are those species that were common in the middle of the last century, but showed considerable decrease in frequency in the last 20 years: Bombus lapidarius (Linné, 1758), Bombus pascuorum (Scopoli, 1763), Bombus hortorum (Linné,1761) and Bombus ruderarius (Mueller,1776) suffered the strongest decline. Data from longer time window (1980 – 2023) provide more realistic, and even optimistic picture about the above named bumble bees (Table 7, sum of historical and more recent data). Here the trend in overal abundance increase is observable. As a conclusion, we may say that the population of these common and widespread species increased.
Especially interesting species are Bombus haematurus Kriechbaumer, 1870 and Bombus argillaceus Scopoli, 1763. These two species were historically very sporadic [27,99,100,101,102]. Due to the changed climatic condition, they are spreading to newer territories situated North-West of their original area of distribution [94,103,104,105,106]. The center of distribution of these 2 species is the Western Palearctic, Ponto-Mediterranean region. Their increase is strong indication of ongoing climatic change. According to projections of Rasmont et al. [106], these species can benefit from the climate change and potentially enlarge their current distribution in Europe in the upcoming decades. Cuckoo bumblebees were never been frequent but for now, they became even rarer as they were before. They didn’t disappear (since time by time they are observed by various entomologists and reporting the observation on the net) but they fall below our detection limit (Table 7).
Diptera
Syrphidae (Hoverflies)
Being a moisture-loving group of insects, it is not surprising that their number shows decreasing trend Table 8 and Table 12). The surprising is the intensity of this trend. At family level, the decrease compared to the beginning of the 80s is about 80%. Some species, namely Sphaerophoria scripta (Linné, 1758), Cheilosia variabilis (Panzer, 1798) and Syrphus torvus (Osten Sacken, 1875) suffered drastic decrease in numbers (96-97%) (Table 8). We didn’t find any species in the family whose population density would have been positively affected by the climatic conditions of recent decades. No invasive species were detected in this group. The northern expansion of a Mediterranean species, Chalcosyrphus pannonicus (Ooldengberg, 1916), was detected in Poland and Slovakia in 2010 and in 2011: Poland, Carpathians, Lower Beskid, Magura NP, Żydowskie, 530 ma.s.l., 24. 07. 2011, 1 male; Slovakia, Carpathians, Lower Beskids, Ondavskie Foothills (Slov. Ondavská vrchovina), district. Chalcosyrphus pannonicus (Ooldenberg, 1916) is a rare species, so far it has been caught in Croatia, Romania, Bulgaria, Greece and the Caucasus [83]. According to our experience, the strong decline of hoverflies is independent of their ecological type: whether the insect is eurytherm or mesophyl. This decline also affects all hoverfly groups independently of their lifestyle: if the species develops in water, compost, plant parts, fungi or even aphidophage or nest parasite, all of them have suffered serious decline in their population density. Decline of some species is so strong, that itcan be modeled by exponential trend (ln x coeff. value) instead of linear trend, Table 8). Fifty, earlier regularly collected, sporadic species became so rare, that they fell below the detection limit in the last 2 decades (Table 13).
Tabanidae (Horse-flies)
At family level, the number of individuals shows an increase about 1.89x compared to the beginning of the 80s (Table 12). Certain xerotolerant, warm-loving species, namely Therioplectes gigas (Herbst, 1787), Chrysops caecutiens (Linné, 1758), Haematopota italica Meigen, 1804 and Tabanus bovinus Linnaeus, 1758 increased remarkably, taking the early 1980s as a base, this increase is 3-5x (Table 9). We have (so far) caught neither Mediterranean newcomers nor invasive species. Decline of Haematopota pluvialis (Linné, 1758) moorland and silvícole species and Atylotus rusticus (Linné, 1761) mesophile species shade the overall picture (Table 9). Only the Mediterranean Pangonius pyritosus (Loew, 1859) can be assumed to have been able to expand to the north due to climate change. In 1991, it appeared for the first time in the Carpathian Basin near Homorúd [84], but its population density hasn’t expanded so far. Till this time, invasive species hasn’t been detected either.
Bombyliidae (Bee Flies)
We experienced intensive increase in population densities in family (Table 12) and in species level as well (Table 10). Taking the beginning of the 80’s as the base period, this increase is about 80%. It makes this group one of the winners of climate change. The following species produced outstanding growth: Bombylius discolor Mikan, 1796, Conophorus virescens (Fabricius, 1787), Bombylius fimbriatus Meigen, 1820, Bombylius cinerascens Mikan, 1796, Villa hottentotta (Linné, 1758), Bombylius canescens Mikan, 1796, Bombylius fulvescens Meigen & Wiedemann, 1820, Bombylius major Linné, 1758, Anthrax anthrax (Schrank, 1781), Anthrax leucogaster Meigen & Wiedemann, 1820, Bombylius pictus Panzer, 1794 and Hemipenthes morio (Linné,1758) (Table 10). The extreme 10-18x increase of density of Exoprosopa jacchus (Fabricius, 1805), Lomatia sabaea (Fabricius 1781) and Bombylius medius Linné, 1758 is associated with the intensive expansion of these species. The reasons for this expansion is unknown, it is likely their hosts are warm-loving insect (for instance antlions [85,86], and other xerotolerant gorups like Acrididae, Tenebrionidae, Aculeata etc. [87]). We haven’t detected any invasive species so far.
Tachinidae (Tachinids)
They suffered significant decline similar to that of Syrphidae (Table 11 and Table 12). However, this decrease is not strong trend, the average r2 value is around 0.2. For hoverflies, it is 0.5. Probably, thanks to their endoparasitoids way of life, they are less exposed to external influences than moisture-loving Syrphidae. No species has population density increased, however some previously common species, such as Phasia pusilla Meigen, 1824 or Gymnosoma dolycoridis Dupuis 1961, have suffered so strong decline in populations that their numbers have fallen below detection limit. We have not detected any invasive species till this time. Twenty-seven, earlier regularly collected, sporadic species became so rare, that they fell below the detection limit in the last 2 decades (Table 13).
Table 11.
Number of individuals of various Tachinid species between 1980 and 2014.
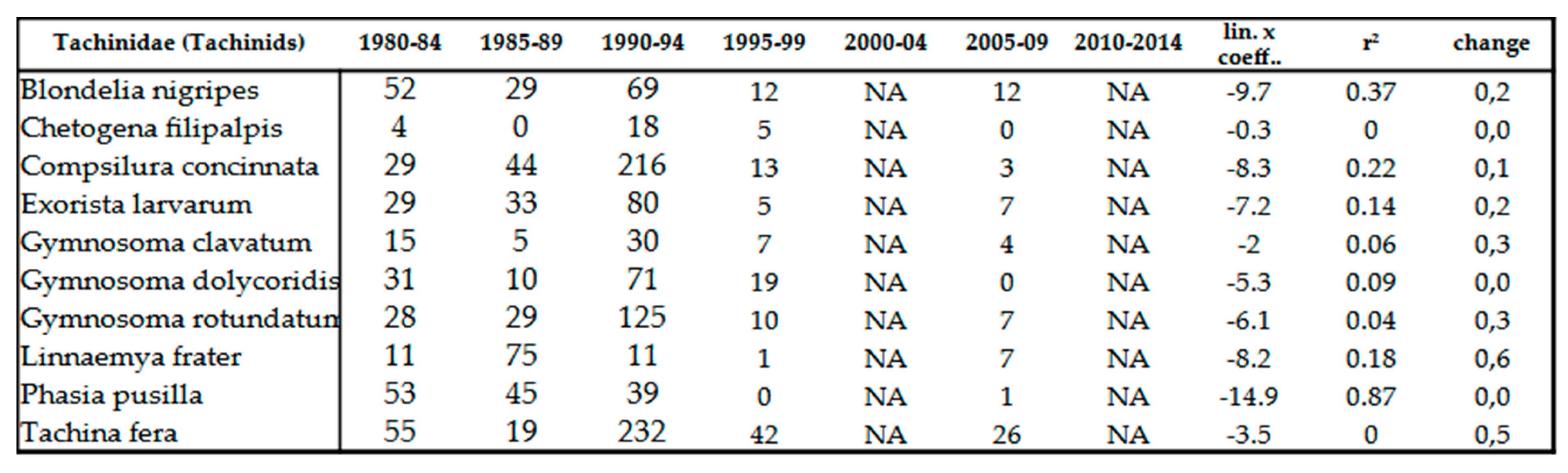
Table 12.
Changes of frequency of vEarious Diptera families between 1980 and 2019.
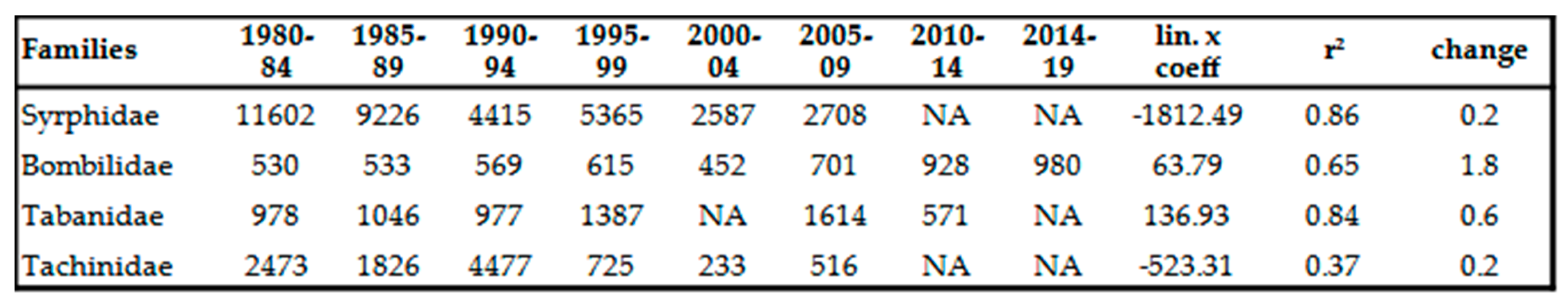
Table 13.
Earlier sporadic Diptera species not colelcted in the last 20 years.

Lepidoptera
Nocturnal macrolepidoptera
Considering the beginning of the 70’s as the base period, population-decrease of moths is relatively strong (Figure 3), but with low determination coefficient, which means the long term, tendentious changes (for example: climate change, habitat changes), are only partly the cause of their decline; other cyclical temporal variables, and their spatial distribution pattern are also significant and strongly influence their populations. According to our observations and our available data, decline-trend lasted till the 2010s. After this time, the trend reversed and we may consider a certain increase in the number of individuals: (linear x coefficient from -157 up to 147 , Table 15, moths total). The low r2 value (0,04) indicates some influence of different method of light trapping (see methods and material part). Taking a closer look at the various groups, owlet moths, sphinx moths and Drepanids suffered the most drastic changes. In family level, the trend is continuously declining in these groups. Nolidae and Notodontidae species show an increasing trend in number of individuals during the last decade (only UV LED portable light traps were applied in this decade). In terms of species richness, negative trend is experienced. In each year, light traps catch fewer and fewer species, till 2014 when this trend stopped (Table 16 and Figure 4). At Geometridae, we observed the strongest decline in species diversity. This decrease in species richness of this group has not stopped till this day (Table 16), while in other families, this trend has stopped and turned into slight increase (Table 16). During this phase, there was no change of methodology. In last decade, we noticed some changes in the order of the 10 most frequent species: relative proportion of Mythimna turca, Athetis furvula and Mythimna pallens declined and proportion of Eilema lurideola and Colocasia coryli increased. For details, see Table 14.
Our macrolepidoptera fauna is enriched by two imported species (Antheraea yamamai. Guérin-Méneville, 1861 and Tarachidia candefacta (Hübner, 1831)), one accidentally introduced species (Hyphantria cunea (Drury, 1773)) and one expansive species (Helicoverpa armigera (Hübner, 1808)). Hyphantria cunea (Drury, 1773) is very likely not a pollinator and this species has strong tendency to gradation, so its occasional erruptions (such as in the 70s) makes difficult to determine its population trend. Our data, show declining trend, which can easily be overwritten by a population eruption (gradation) at any time. The trend of the other 3 species shows slight increase (Table 17).
Butterflies (Rhopalocera)
Scheme 60. but in some groups even up to 90% (compared to 1970s). The estimated relative changes in populations of some important butterflies are displayed in Table 18. Certain species, especially Aglais urticae (Linné, 1758), were once among the most common species, but for now, almost completely disappeared. Species of Pieridae, Lycaenidae, Nymphalidae and Hesperiidae show strong decrease either. Species of family Paplionidae seem to be stable. Population densities of some species tend to opposite direction. Increase of Iphiclides podalirius (Linnaeus, 1758) is the strongest. Our data in Table 18, indicate the expansion of Euphydryas aurinia Rottemburg, 1775 and Libythea celtis (Laicharting, 1782) either.
Recent re-investigation of the 40 years or even earlier researched areas were carried out in three regions: Dráva Plain (border region between Hungary and Croatia), Bátorliget Nature Reserve (NE Hungary, close to Ukraine) and the area around Simonfa town. Decline in species richness in these regions shows strong change. Compare data of Table 19.
4. Discussion
Symphyta
We have only small amount of literature data on the changes in population densities of sawflies. In the Pannonian biogeographic region, the decline in the diversity of certain localities and the number of individuals of the Nematinae group is logical, as this subfamily typically reaches its maximum diversity and number of individuals in Scandinavia and the north [70]. The decline of Tenthredo species is somewhat incomprehensible. Goulet [69], found the same trend in Canada. Goulet attributes the decline in Tenthredo species to the use of pesticides. However, the spread of beekeeping is also having an unfavorable effect on the this group [72].
To date, the negative effects of climate change on the Symphyta group have been studied in Andalusia [73]: “While some species were frequently found in the same areas as 50 years ago, climate changes affected the vertical displacement of other studied sawfly species to higher altitudes. The main results of this study showed that in the 21st century, four species (Megalodontes bucephalus, Macrophya militaris, Strongylogaster multifasciata, Dolerus (Poodolerus) puncticollis) were not observed in any location or sampling area, which means that these species (important specialized pollinators) have disappeared from the Andalusia region.” This “vertical shift” can be observed in the higher areas of the Carpathian Basin either (Table 3). High altitudes, like surrounding mountains of the Carpathian Basin, help us to save the diversity of the sawfly fauna for a while longer.
Aculeata
Scientific papers don’t discuss the change of Aculeata in total, however our results are supported by scientific publications on specific genera and species. These results are consistent with our experiences. Olszewski et al. [74] reported the increase of population density of Philanthus triangulum (Fabricius,1775) and Eickermann et al. [75] wrote about the increasing population of Polistes spp. in Europe. Similar tendencies are took place in North America either, as indicated by the proliferation of several Sphex species [76]. Also, in South America, from where, the northern expansion of Centris nigrescens Lepeletier, 1841 was reported in connection with global warming [77]. Zimmerman et al. [78] published their similar experiences about the gradual increase and expansion of wild bee populations in Eastern Austria: “Among the newly recorded species, Ceratina nigrolabiata, Icteranthidium laterale, Lithurgus chrysurus, L. cornutus, Osmia bidentata, O. spinulosa, Pseudapis diversipes, and its parasite Pasites maculatus currently expand their distribution from warmer, more southern and eastern regions to Austria, probably as a response to climatic warming”. Data on the decline of Ectemnius and Crossocerus species are provided by Bogusch and Jakub, also Pearce-Higgins et al. [79,80]. Reasons for their decline, one is the general decline of saprophilic Hymenoptera due to the disappearance of old forests (energy crisis) and the climatic change: some of Ectemnius species are mesophile, and they are characteristic species of marshy meadows. Finally, these marshy meadow are threatened not only by the global warming but also by the non native Golden rod (Solidago spp.) expansion which kill the original vegetation.
Bumblebees (Bombus spp.)
Data of international scientific papers confirm the same that we experienced in the Pannonian Basin: slighter or similar decrease in the population densities and diversity was observed in many other European countries [88]. At the same time, in Western Europe, originally common species (e.g., B. lapidarius, B. pascuorum, B. terrestris) became rare [89]. In England, this decline was so strong in the 1980s that only six bumblebee species were collected in those regions where 19 species had been captured before 1960 [90].
According to Plowright et al. [91]: between 1977 and 1994, Bombus muscorum (Linné,1758) disappeared from many habitats in northern England and they were replaced by B. pascuorum. Our observation may confirm this conclusion, although, in our region, the Mediterranean B. haematurus has more significant role in replacing other bumblebee species. Regarding Bombus terrestis, B. hortorum, B. lapidarius, B. pascuorum, B. haematurus, B. ruderarius and B. argillaceus our long term data confirm the results of Jakab et. al. [92]. Our data from period of 2000 – 2003 are well in line with the results of Sárospataki et. al. [93], According to this paper: ‘36% of the bumblebee fauna can be considered rare and 24% moderately rare, i.e., over half of the total number of species can be classified into these two categories. Almost half (47%) of the species still living in the Pannonian biogeographic region of the Carpathian Basin, showing decreasing trend starting from the 1950s and 1960s’. However, montaneous regions of Western Carpathians situated north of Pannonicum (Carpaticum occidentale and Carpaticum orientale), could show different trends. Bumble bee communities of Outer and Inner Western Carpathians are apparently still rich in diversity and relative abundance. Rare and infrequent species, such as e.g., Bombus distinguendus, B. subterraneus, B. pomorum, B. confusus, B. veteranus, B. quadricolor and B. norvegicus are still present in this area [96,97,98]. Moreover, sub recently, a new species, Bombus semenoviellus. Skorikov, 1910, appeared in our region [95]. For the more precise knowledge on status of the bumble bee fauna of the Pannonian basin, we would strongly advice to continue in systematic and frequent monitoring of all members of bumble bee fauna in the entire area of interest.
Diptera
We have limited information available from the scientific papers therefore our research brings original and new results. Available literature published in the neighboring regions agrees that certain groups, especially hoverflies, suffer significant decline in numbers of individuals similar to our results [107]. According to an IUCN study [108]: “Hoverflies generally ensure better pollination than bees at higher altitudes, under Nordic climatic conditions, or in cool microclimate or weather situations.”. Sommagio et al., [109] in addition to the catastrophic decline in number of hoverflies, mention that mountainous regions are able to provide shelter for them, but this is not true for bumble bees: “The two taxa show different distribution patterns: hoverflies have a unimodal distribution (richness and abundance) with peak at middle altitude (1500 m), while bees have a monotonic decline (richness and abundance) with increasing altitude.”
The experienced increase of populations of horse-flies is explained by their need for warmth. Herczeg et al. [110] did not manage to collect even a single Tabanid specimen below 18 °C. At the same time, they note that variability in moisture requirements per species is high, which explains the often opposite trends in populations of various species, similar to our experiences. Also interesting the opposite trend of 2 similar and closely related large horse-fly species, namely Tabanus bromius Linné, 1758 and Tabanus bovinus Linné, 1758 (Table 9). Probably there is niche competition between these 2 species as Dörge et al. [111] write:”Tabanus bovinus and T. bromius have similarly large niches which are mostly overlapping.” In case of Tachinids, climatic factors have strong, but indirect and very diverse effect. The most important effect is the optimization of the synchronicity between the presence of host and the parasitoid’s egg-laying time: an optimally developed host animal larva should be available at the time of reproduction. Climatic conditions can improve this, but it can also shift it in an unfavorable direction [112], which could also be the reason of the fluctuation shown in Table 11 and Table 12. For Bombyliidae, Boesi et al., [113] provide good explanation for their increased reproduction:”Bee flies (Diptera: Bombyliidae) have a virtually cosmopolitan distribution and are commonly found in warm arid to semi-arid habitats, where they can form a conspicuous part of the flower-visiting insect fauna [113].
Lepidoptera
Nocturnal macrolepidoptera
Only 3 papers study and discuss temporal changes of various moths species during the last 3-4 decades. Far the most important comparative study available is the PhD. thesis of Fox [114]. Fox investigated changes of about 600 moth species between 1970 and 2010. Our trend for many species is opposite to the tendencies set in Great Britain by Fox. These are Phragmatobia fuliginosa (Linnaeus, 1758), Xestia c-nigrum (Linnaeus, 1758), Eilema lurideola (Zincken, 1817), Spilosoma lubricipeda (Linnaeus, 1758), Lacanobia oleracea (Linnaeus, 1758) and Mythimna turca (Linnaeus, 1761) (Table 14). Their decrease in the south and their expansion in the north may indicate that they find better living conditions in the northern and humid Atlantic areas due to the gradually warming climatic conditions in their original habitats. Major proportion of the declined or rare species from the South (including the Carpathian Basin) are usually mesophile, silvicol species. Their typical habitats are meadows, swamps, tall sedges, mesophile forests, groves, and alder forests. The opposite movement can be observed at warm-loving species: their populations in England declined, while in the Carpathian Basin, according to our data, their individual density increased, for instance Paracolax tristalis (Fabricius, 1794). Other xerotolerant, warm-loving species, like Macdunnoughia confusa (Stephens, 1850), Drymonia obliterata (Esper, 1785), Athetis furvula (Hübner, 1808), Zanclognatha lunalis (Scopoli, 1763) and Earias vernana (Fabricius, 1787).are not on the British list, since they are Mediterranean species. These species expand their territories and population densities in the Carpathian Basin These are eurytherm and/or polyphagous species, adapted better to more extreme conditions. Those species which prefer cold and moisture ecosystems are strongly declined like Diachrysia and Abrostola species (Table 14).
Otherpapers, like Conrad et. al., [115] treating 337 species from Britain and Mikkola’s work [116] discussing 54 species from Finnland, howver those species which are discussed in these monographs are hardly overlap the fauna of the Carpathian Basin, therefore we don’t discuss these works in details.
The reasons behind the above described trends can be traced back to many influencing factors that act in very complex way: global warming, frequent extreme temperature maximums, changed temporal distribution of precipitation: droughts, torrential rains, improper forest management, degradation of habitats, large-scale clear-cutting that affect microclimate, intensive lawn management, fertilization of lawns, incorrect selection of lawn-mowing dates. groundwater depletion, underground piping etc. Internal factors also may influence population densities: just an example: according to Hill et.al .[117], Xestia c-nigrum (Linné, 1758) can produce a so called heat shock protein, Hsp70: “Another molecular marker that is likely to be important in the response to climate change is the heat shock protein (Hsp70). Hsp70 genes play a critical role in helping insects survive exposure to extreme temperatures by increasing heat tolerance”. In this aspect, Xestia c-nigrum population shall increase, however: “We found that grazing and mowing/fertilization (in Germany) had largely opposing effects on the moth assemblages: species characteristic for meadows such as Agriphila straminella, Agriphila tristella and Crambus perellus declined under more intense livestock grazing and were replaced by other species such as Anerastia lotella, Mythimna pallens and Xestia c-nigrum” [118]. In our region, livestock grazing has declined and the trend is opposite than in Germany. Finally, there is still an open discussion, which moths, or even which insects are pollinators [119]. We may say, pollinators are those moths (and even those animals), which have at least minimal parts of their life-cycle is temporarily or regularly connected to flowers and in this way, they transport pollen, helping the fertilization of plants. These insects could be predators: hunting for their prey on flowers, insects attracted to various colors and wavelengths emitted by flowers, insects attracted to various odors and pheromone-like chemicals of flowers, or attracted to special appearance of flowers, animals feeding on nectar or consuming various parts of flowers, or those which find temporary shelter or place for warming up themselves on the surfaces of flowers etc.
Butterflies (Rhopalocera)
Population increase of Euphydryas aurinia is described by Dietzel, Ábrahám and Ács et al. [48,54,56]. Euphydryas aurinia has two ecotypes. Wet meadow ecotype has been drastically decreased, while the dry meadow ecotype has been spreading since the 90s. Other results confirm the expansion of Libythea celtis (Laicharting, 1782) [56,120]. Libythea celtis is a migratory species, reaches the Carpathian Basin from South. Its population is increasing step by step as a result of global warming. Also, its food plant (Celtis occidentalis) is planting in parks and also in forests. Bury et al., [121] observed the population increase of Iphiclides podalirius (Linné, 1758) in Poland, the same that we experienced in the Carpathian basin (Table 18). Its gradual population increase caused by the expansion of blackthorn (Prunus spinosa) in the abandoned areas. Neptis hylas (Linné, 1758) was frequent in Southern and Western Transdanubia only. In recent years, it has appeared in areas where it hasn’t bred before. Fuirthermore this species started to feed on locust tree (Robinia pseudoacacia). Till recently, only Lathyrus spp. were its host-plants [56,120]. Decline of populations of other rare butterfly species like Nymphalis antiopa. (Linné, 1758), Apatura and Maculinea spp. is rather a nature conservation problem.
In terms of population-densities of butterflies, Hill et al., [117] came to a similar conclusion:: “In most cases, oligo- or polyphagous species (diet generalists) have an advantage because they have a wider breadth of host plants, allowing for easy colonization of new sites during range expansion. By contrast, diet and habitat specialists typically have poor dispersal ability and may not be able to track environmental changes when suitable habitat patches are reduced and fragmented, resulting in local extinctions and range declines.”.
5. Conclusions
Our results support the conclusion of Dicks et al. [5] ”Europe was the region where human well-being was considered at the lowest risk from pollinator declines overall (mean risk score = 19.6), with no ‘high’ risks, and only two ‘serious’ risks (pollination deficit and wild pollinator diversity).”
Beyond the above mentioned pollinator decline, most important is the gradual transition of our Continental type pollinator towards Mediterranean type.
Population densities of warm-loving and drought-tolerant species, and species groups are increasing, while those of northern, silvicole species are declining.
Butterflies (Rhopalocera), hoverflies (Syrphidae), tachinids (Tachinidae), Symphyta and bumblebees (Bombus spp.) suffered decline in the last 2 decades. Meanwhile, population densities of Aculeata, Bombyylidae, Tabanidae are increasing.
High altitudes may provide shelter and help to keep diversity for some moisture-loving group. We provided evidence for this at sawflies (Table 3). Probably it is true for other pollinator groups which prefer moderate climatic and moisture conditions.
Decline of bumblebees started around 2015, Syrphidae around 2000, Tachinidae around 1995 and butterflies around 2000 (the latest is indicated by disappearance of the so far common Aglais urticae).
The influx of Mediterranean species into the Carpathian Basin was the strongest at Aculeata. Introduced species also enriched local pollinator fauna, especially in Aculeata and Lepidoptera (in Symphyta only 1 species are introduced recently).
The Mediterranean transformation of our pollinator fauna is a response to the gradual aridification of the Carpathian Basin [122] .
Author Contributions
Symphyta and Lepidoptera, Attila Haris; Symphyta Ladislav Roller; Aculeata, Zsolt Józan.; Diptera, Sándor Tóth.; bumblebees, and partly invasive Aculeata, Peter Šima.; statistical analysis, Attila Haris; resources, All authors.; data curation, All authors; writing—original draft preparation, All authors; All authors have read and agreed to the published version of the manuscript.
Funding
This study was partly financed by the project No. 2/0070/23 of the Slovakian Funding Agency VEGA.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
We encourage all authors of articles published in MDPI journals to share their research data. In this section, please provide details regarding where data supporting reported results can be found, including links to publicly archived datasets analyzed or generated during the study. Where no new data were created, or where data is unavailable due to privacy or ethical restrictions, a statement is still required. Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
Authors express their grateful thanks to Vladimir Smetana (Tekov Museum, Levice, Slovak Republic), Levente Ábrahám (Rippl-Rónai Museum, Kaposvár, Hungary), Ákos Uherkovich, (Janus Pannonius Museum Pécs, Hungary), György Csóka and Anikó Hirka (University of Sopron, Forest Research Institute, Hungary) for their generous supports, data provisions and advises.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Molnár, A.V.; Takács, A. Megporzási válság. A pollináció mint természeti szolgáltatás. Ökológia. Természettudományi közlöny, 2016, 147, 303–305. [Google Scholar]
- Rhodes, C.J. Pollinator decline – an ecological calamity in the making? Sci. Prog. 2018, 101, 121–160. [Google Scholar] [CrossRef]
- Halmos, G.; A hosszútávú vonuló énekesmadár fajok állományváltozásának és populációdinamikájának vizsgálata. Doctoral thesis. ELTE Department of Systematic Zoology and Ecology, 2009, 117. Available online: https://teo.elte.hu/minosites/tezis2009/halmos_g.pdf (accessed on 18 March 2024).
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A. , Hill, R.; Settele, J. and Vanbergen, A.J. Summary for policymakers of the assessment report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on pollinators, pollination and food production. Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, Bonn, Germany. 2016; 36 pp.
- Dicks, L.V.; Breeze, T.D.; Ngo, H.T.; Senapathi, D.; Jiandong, A.; Aizen, M.A.; Basu, P.; Buchori, D.; Galetto, L.; Garibaldi, L.A.; et al. A global-scale expert assessment of drivers and risks associated with pollinator decline. Nat. Ecol. Evol. 2021, 5, 1453–1461. [Google Scholar] [CrossRef]
- Kluser, S.; Peduzzi, P. Global Pollinator Decline: A Literature Review. A scientific report about the current situation, recent findings and potential solution to shed light on the global pollinator crisis. UNEP/DEWA/GRID-Europe, 2007; 10 pp.
- Marks, R. Native Pollinators. Fish Wildl. Habitat Manag. Leafl. 2005, 34, 1–10. [Google Scholar]
- EPRS | European Parliamentary Research Service Protecting pollinators in the EU. European Parliament Briefing. 2021, 8 pp.
- Zombori, L. Adatok Nagykovácsi levéldarázsfaunájához I. (Hymenoptera, Symphyta). Folia Ent. Hung. 1973, 26, 217–224. [Google Scholar]
- Zombori, L. Jegyzetek Nagykovácsi levéldarázs faunájáról (Hymenoptera: Symphyta) II. Folia Ent. Hung. 1975, 28, 223–229. [Google Scholar]
- Zombori, L. Adatok Nagykovácsi levéldarázs-faunájához (Hymenoptera: Symphyta) III.-IV. Folia Ent. Hung. 1975, 28, 369–381. [Google Scholar]
- Zombori, L. Sawflies from the Agtelek National Park (Hymenoptera: Symphyta). In The fauna of the Aggtelek National Park, II. Mahunka, S. Ed., Hungarian Natural History Museum, Budapest. 1999, pp. 573–580.
- Zombori, L. Sawflies form Fertő-Hanság National Park (Hymenoptera: Symphyta). In The fauna of the Fertő-Hanság National Park, Mahunka, S. Ed., Hungarian Natural History Museum, Budapest. 2002, pp. 545–552.
- Haris, A. Sawflies of the Zselic Hills, SW Hungary Hymenoptera: Symphyta. Nat. Som. 2009, 15, 127–158. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Vértes Mountains Hymenoptera: Symphyta. Nat. Som. 2010, 17, 209–238. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Börzsöny Mountains North Hungary Hymenoptera: Symphyta. Nat. Som. 2011, 19, 149–176. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of Belső-Somogy (Hymenoptera: Symphyta). Nat. Som. 2012, 22, 141–162. [Google Scholar] [CrossRef]
- Haris, A. Second contribution to the sawflies of Belső Somogy Hymenoptera: Symphyta. Nat. Som. 2018, 31, 45–62. [Google Scholar] [CrossRef]
- Haris, A. Sawflies from Külső-Somogy, South-West Hungary (Hymenoptera: Symphyta). Nat. Som. 2018, 32, 147–164. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Keszthely Hills and its surroundings. Nat. Som. 2019, 33, 107–128. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of Southern part of Somogy county Hymenoptera: Symphyta. Nat. Som. 2020, 35, 51–70. [Google Scholar] [CrossRef]
- Haris, A. Sawflies of the Cserhát Mountains Hymenoptera: Symphyta. Nat. Som. 2021, 37, 25–42. [Google Scholar] [CrossRef]
- Haris, A. Second contribution to the knowledge of sawflies of the Zselic Hills (Hymenoptera: Symphyta). A Kaposvári Rippl-Rónai Múzeum Közleményei (Commun. Rippl-Rónai Mus. Kaposvár) 2022, 08, 65–80. [Google Scholar] [CrossRef]
- Haris, A.; Vidlička, L.; Majzlan, O.; Roller, L. Effectiveness of Malaise trap and sweep net sampling in sawfly research (Hymenoptera: Symphyta). Biologia, 2024; online. [Google Scholar] [CrossRef]
- Roller, L. Sawfly (Hymenoptera, Symphyta) community in the Devínska. Kobyla Nat Reserve Biol. Bratisl. 1998, 53, 213–221. [Google Scholar]
- Roller, L. Seasonal flight activity of sawflies Hymenoptera, Symphyta in submontane region of the West Carpathians, Central Slovakia. Biol. Bratisl. 2006, 61, 193–205. [Google Scholar] [CrossRef]
- Józan, Z. A Zselic méhszerű (Hymenoptera, Apoidea) faunájának alapvetése. The Apoidea (Hymenoptea) fauna of the Zselic Downs. Janus Pann. Múz. Évk. 1989, 34, 81–92. [Google Scholar]
- Józan, Z. A Zselic darázsfaunájának (Hymenoptera, Aculeata) állatföldrajzi és ökofaunisztikai vizsgálata. Zoologeographic and ecofaunistic study of the Aculeata fauna (Hymenoptera, Aculeata) of Zselic. Somogyi Múzeumok Közleményei 1992, 9, 279–292. [Google Scholar]
- Józan, Z. A Béda-Karapancsa Tájvédelmi Körzet fullánkos hártyásszárnyú (Hymenoptera, Aculeata) faunájának alapvetése. Aculeata (Hymenoptera, Aculeata) fauna of the Béda-Karapancsa Landscape Protection Area. Dun. Dolg. Term. Tud. Sor. 1992, 6, 219–246. [Google Scholar]
- Józan, Z. A Boronka-melléki Tájvédelmi Körzet fullánkos hártyásszárnyú (Hymenoptera, Aculeata) faunájának alapvetése. Aculeata (Hymenoptera, Aculeata) fauna of the Boronka Landscape Protection Area. Dun. Dolg. Term. Tud. Sor. 1992, 7, 163–210. [Google Scholar]
- Józan, Z. Adatok a tervezett Duna-Dráva Nemzeti Park fullánkos hártyásszárnyú (Hymenoptera, Aculeata) faunájának ismeretéhez. Data to the knowledge of the Aculeata (Hymenoptera, Aculeata) fauna of the planned Danube-Dráva National Park. Dun. Dolg. Term. Tud. Sor. 1995, 8, 99–115. [Google Scholar]
- Józan, Z. A Mecsek méhszerű faunája (Hymenoptera, Apoidea). The Apoidea (Hymenoptera) fauna of the Mecsek Mountains (Hungary: South Transdanubia). Janus Pann. Múz. Évk. 1995; 40, 29–43. [Google Scholar]
- Józan, Z. A Baláta környék fullánkos hártyásszárnyú faunájának (Hym., Aculeata) alapvetése. Aculeata fauna of Lake Baláta (Hym., Aculeata). Somogyi Múzeumok Közleményei 1996, 12, 271–297. [Google Scholar]
- Józan, Z. A Duna-Dráva Nemzeti Park fullánkos hártyásszárnyú (Hymenoptera, Aculeata) faunája. – The Aculeata fauna of the Duna-Dráva National Park, Hungary (Hymenoptera, Aculeata). Dun. Dolg. Term. Tud. Sor. 1998, 9, 291–327. [Google Scholar]
- Józan, Z. A Villányi-hegység fullánkos hártyásszárnyú (Hymenoptera, Aculeata) faunája. The Aculeata (Hymenoptera) fauna of the Villány Hills, South Hungary. Dun. Dolg. Term. Tud. Sor. 2000, 10, 267–283. [Google Scholar]
- Józan, Z. A Mecsek kaparódarázs faunájának (Hymenoptera: Sphecoidea) faunisztikai, állatföldrajzi és ökofunisztikai vizsgálata. Faunistical, zoogeographical and ecofaunistical investigation on the Sphecoids fauna of the Mecsek Montains (Hymenoptera, Sphecoidea). Nat. Som. 2002, 3, 45–56. [Google Scholar]
- Józan, Z. Az Őrség és környéke fullánkos hártyásszárnyú faunájának alapvetése (Hymenoptera, Aculeata). Aculeata (Hymenoptera, Aculeata) fauna of Őrség and its surroundings. Praenorica Fol. Hist.-nat. 2002, 6, 59–96. [Google Scholar]
- Józan, Z. A Mecsek fullánkos hártyásszárnyú faunája (Hymenoptera, Aculeata). Aculeata fauna of Mecsek Hills (Hymenoptera, Aculeata). Folia Comloensis, 2006, 15, 219–238. [Google Scholar]
- Józan, Z. Új kaparódarázs fajok (Hymenoptera, Sphecidae) Magyarország faunájában. New sphecid wasps (Hymenoptera, Sphecidae) in the fauna of Hungary. Somogyi Múzeumok Közleményei 2008, 18, 81–83. [Google Scholar]
- Józan, Z. A Barcsi borókás fullánkos faunája, III. (Hymenoptera: Aculeata). Aculeata fauna of Barcs Juniper Woodland (Hymenoptera: Aculeata). Nat. Som. 2015, 26, 95–108. [Google Scholar]
- Tóth, S. Angaben zur Kenntnis der Schwebfliegen-Fauna der Slowakei (Diptera: Syrphidae). Fol. Mus. Hist.-Nat. Bakony. 1990, 9, 91–108. [Google Scholar]
- Tóth, S. Magyarország zengőlégy faunája (Diptera: Syrphidae). Hoverflies of Hungary (Diptera: Syrphidae). e-Acta Naturalia Pannonica, 2011, Suppl. 1., 1–408.
- Tóth, S. Magyarország fürkészlégy faunája (Diptera: Tachinidae). Tachinid flies of Hungary (Diptera: Tachinidae). e-Acta Naturalia Pannonica, 2013, 5, Suppl. 1, 1–321.
- Tóth, S. A hazai bögölyök nyomában (Diptera: Tabanidae). Horse-flies of Hungary (Diptera: Tabanidae). e-Acta Nat. Pannonica 2023, Suppl. 4, 1–124.
- Ábrahám, L. Biomonitoring of the buttetfly fauna in the Drava region {Lepidopteia: Diurna). Nat. Som. 2005, 7, 63–74. [Google Scholar]
- Ábrahám, L. A Boronka-melléki Tájvédelmi Körzet nagylepke faunájának természetvédelmi értékelése I. (Lepidoptera). Nature conservation evaluation of the macrolepidoptera fauna of Boronka Landscape Protected Area (Lepidoptera). Dun. Dolg. Term. Tud. Ser. 1992, 7, 241–271. [Google Scholar]
- Ábrahám, L. Bakonynána és környéke nagylepke faunája (Lepidoptera) Macrolepidoptera fauna of Bakonynána region. Fol. Mus. Hist.-Nat. Bakonyensis 1991, 10, 85–104. [Google Scholar]
- Ábrahám, L. Lápi tarkalepke Euphydryas aurinia (Rottembtirg, 1775). Marsh fritillary Euphydryas aurinia (Rottembtirg, 1775). In Natura 2000 fajok és élőhelyek Magyarországon. Ed. Haraszthy, L., Pro Vértes Közalapítvány, Csákvár 2014, 323-326.
- Ábrahám, L.; Herczig, B.; Bürgés, G. Faunisztikai adatok a Keszthelyi-hegység nagylepke faunájának ismeretéhez (Lepidoptera: Macrolepidoptera}. Data to the Macrolepidoptera fauna of Keszthely Hills (Lepidoptera: Macrolepidoptera}. Nat. Som. 2007, 10, 303–330. [Google Scholar] [CrossRef]
- Ábrahám, L.; Uherkovich, À. 1993: A Zselic nagylepkéi (Lepidoptera) 1. Bevezetés és faunisztikai alapvetés. Macrolepidoptera fauna of Zselic (Lepidoptera) Introduction and biomonitoring. Janus Pann. Múz. Évk. 1993, 38, 47–59. [Google Scholar]
- Uherkovich, Á. Long-term monitoring of biodiversity by the study of butterflies and larger moths (Lepidoptera) in Sellye region (South Hungary, co. Baranya) in the years 1967–2022. Nat. Som. 2022, 9, 95–138. [Google Scholar] [CrossRef]
- Pillich, F. Aus der Arthropodenwelt Simontornya’s, Pillich, F. private edition, Simontornya, Hungary, 1914. 172 pp.
- Sáfián, Sz. Butterflies of Kercaszomor (Őrség), Western Hungary (Lepidoptera: Papilionoidea and Hesperioidea). Nat. Som. 2011, 19, 251–262. [Google Scholar]
- Ács, E. ; Zs. Bálint, Ronkay, G.; Ronkay, L.; Szabóky, Cs.;Varga, Z.; Vojnits, A. The Lepidoptera of the Bátorliget Nature Conservation areas. The Bátorliget Nature Reserves – after forty years Vol. 2. Mahunka, S. Ed. Hungarian Natural History Museum, Budapest, 1991, pp. 505–540.
- Čanády, A, Príspevok k poznaniu výskytu denných motýľov (Rhopalocera) v urbánnom prostredí Košíc. (Slovensko). Contribution to the knowledge of the occurrence of butterflies (Rhopalocera) in the urban environment of Košice. (Slovakia). Folia Faun. Slovaca 2014, 19, 235–241. [Google Scholar]
- Dietzel, G. A Bakony nappali lepkéi. Butterflies of Bakony Mountains. In: A Bakony természettudományi kutatásának eredményei 21. Bakonyi Természettudományi Múzeum, Bakony NaturalHistory Museum Ed. Zirc. 1997, pp. 1–212.
- Sarvašová, L. Denné motýle (lepidoptera, papilionoidea) lúk kúpeľov sliač a okolia (Slovensko). Butterflies (lepidoptera, papilionoidea) meadow of spas sliač and surroundings (Slovakia). Folia Faun. Slovaca 2016, 21, 63–71. [Google Scholar]
- Gergely, P. A pomázi Majdán-fennsík nappali lepkéinek megfigyelései 2000 és 2020 között (Lepidoptera: Rhopalocera) Observations of butterflies in the Majdan plateau of Pomáz (Hungary) between 2000 and 2020 (Lepidoptera: Rhopalocera). Lepidopterol. Hung. 2021, 17(2), 99–107. [Google Scholar] [CrossRef]
- Gór, Á. 2018: Lepkefaunisztikai kutatások Biatorbágyon és környékén (Lepidoptera). Lepidoptera survey in Biatorbágy (Hungary) and its surrounding areas. Eacta Nat. Pannonica 2018, 16, 55–70. [Google Scholar] [CrossRef]
- Schmidt, P. A Csombárdi-rét Természetvédelmi Terület nappali lepkéinek alapállapot felmérése (Lepiodptera). The basic survey of the butterfies in the Csombárd-meadow Nature Conservation Area (Lepidoptera). Nat. Som. 2017, 30, 179–192. [Google Scholar] [CrossRef]
- Németh, L. Data to the knowledge of the Macrolepidoptera-fauna of the Tapolca basin. Fol. Mus. Hist.-Nat. Bakonyensis 1991, 10, 105–136. [Google Scholar]
- Szabóky, Cs.; Samu, F.; Szeőke, K.; Petrányi, G. Simontornya lepkevilágáról (Lepidoptera). Moths and butterflies of Simontornya. In: Simontornya ízeltlábúi. Arthropods of Simontornya. Magyar Biodiverzitás-kutató Társaság, Hungarian Biodiversity Research Society Ed., Budapest, 2014, pp. 143–186.
- Hudák, T. A nappali lepkefauna vizsgálata Székesfehérváron (Lepidoptera: Rhopalocera). Investigation on the butterfly fauna of Székesfehérvár (Lepidoptera: Rhopalocera). Nat. Som. 2018, 31, 113–136. [Google Scholar] [CrossRef]
- Varga, J.; Korompai, T.; Horokán, K.; Hirka, A.; Gáspár, Cs.; Kozma, P.; Csóka, G.; Csuzdi, Cs. 2022: Analysis of the Macrolepidoptera fauna in Répáshuta based on the catches of a light-trap between 2014-2019. Acta Univ. Esterházy, Sect. Biol. 2022, 47, 59–75. [Google Scholar]
- Árnyas, E.; Szabó, S.; Tóthmérész, B.; Varga, Z. Lepkefaunisztikai vizsgálatok fénycsapdás gyűjtéssel az Aggteleki Nemzeti Parkban. Moth fauna studies with light trap collection in the Aggtelek National Park. Természetvédelmi Közl. 2004, 11, 34–42. [Google Scholar]
- Kovács, L. Bátorliget nagylepke-faunája. Macrolepidoptera fauna of Bátorliget. pp. 326–380, 483–484. In: Bátorliget élővilága. (Die Tier- und Pflanzenwelt desNaturschutzgebietes von Bátorliget und seiner Umgebung). Székessy, V. ed., Akadémiai kiadó, Budapest, Hungary; 1953, 486 pp.
- Infusino, M.; Brehm, G.; Di Marco, C.; Scalercio, S. Assessing the efficiency of UV LEDs as light sources for sampling the diversity of macro-moths (Lepidoptera). Eur. J. Entomol. 2017, 114, 25–33. [Google Scholar] [CrossRef]
- Pan, H.; Liang, G.; Lu, Y. Response of Different Insect Groups to Various Wavelengths of Light under Field Conditions. Insects 2021, 12, 427–438. [Google Scholar] [CrossRef]
- Goulet, H. Herbicides, beetles, and the decline of insectivorous birds. Canada’s oldest field naturalist club. 2023, 35 pp. https://ofnc.ca/wp–content/uploads/2018/01/Herbicides_beetles_birds.pdf.
- Liston, A.D. Compendium of European Sawflies. List of species, modern nomenclature, distribution, foodplants, identification literature. Chalastos Forestry, Gottfrieding, Germany, 1995, pp. 1–190.
- Roller, L.; Haris, A. Sawflies of the Carpathian Basin, History and Current Research. In Nat. Som. 2008, 11, 1–261. [Google Scholar] [CrossRef]
- Kaplan, E.; Haris, A. Contribution to the knowledge of the sawflies (Hymenoptera: Symphyta) from Turkey. Nat. Som. 2021, 37, 25–38. [Google Scholar] [CrossRef]
- Barbir, J.; Martín, L.O.; Lloveras, X.R. Impact of Climate Change on Sawfly (Suborder: Symphyta) Polinators in Andalusia Region, Spain. 93-111. In Handbook of Climate Change and Biodiversity, , ; Walter Leal Filho, W.L., Barbir, J., Preziosi, R., Eds.; In series: Climate Change Management; Springer Nature Switzerland AG.: Cham, Switzerland, 2018; p. 323. [Google Scholar] [CrossRef]
- Olszewski, P. , Dyderski, M.K.; Dylewski, L., Bogusch, P., Schmid-Egger, Ch..; Ljubomirov, T.; Zimmermann, D.; Le Divelec, R.; Wiśniowski, B.; Twerd, L.; Pawlikowski, T.; Mei, M.; Florina Popa, A.; Szczypek, J.; Sparks, T.; Puchałka, R. European beewolf (Philanthus triangulum) will expand its geographic range as a result of climate warming. Reg. Environ. Change 2022, 22, 129. [Google Scholar] [CrossRef]
- Eickermann, M.; Junk, J.; Rapisarda, C. Climate Change and Insects. Insects 2023, 14, 672–678. [Google Scholar] [CrossRef]
- Lewis, J.H. Sphex ichneumoneus and Sphex pensylvanicus (Hymenoptera: Sphecidae) in Atlantic Canada: evidence of recent range expansion into the region. Can. Field-Nat. 2020, 134, 52–55. [Google Scholar] [CrossRef]
- Vélez, D.; Silva, D.P.; Vivallo, F. Where could Centris nigrescens (Hymenoptera: Apidae) go under climate change? J. Apic. Res., 2023, 62, 1082–1090. [Google Scholar] [CrossRef]
- Zimmermann, D.; Schoder, S.; Zettel, H.; Hainz-Renetzeder, Ch.; Kratschmer, S. Changes in the wild bee community (Hymenoptera: Apoidea) over 100 years in relation to land use: a case study in a protected steppe habitat in Eastern Austria. J. Insect Conserv. 2023, 27, 625–641. [Google Scholar] [CrossRef]
- Bogusch, P.; Horák, J. Saproxylic bees and wasps. In: Saproxylic insects: diversity, ecology and conservation, Ulyshen, M. Ed., Springer Books, Bern, Switzerland, 2018, pp. 217–235. [CrossRef]
- Pearce-Higgins, J.W.; Beale, C.M.; Oliver, T.H.; August, T.A.; Carroll, M.; Massimino, D.; Ockendon, N.; Savage, J.; Wheatley, C.J.; Ausden, M.A.; et al. A national-scale assessment of climate change impacts on species: assessing the balance of risks and opportunities for multiple taxa. Biol. Conserv. 2017, 213, 124–134. [Google Scholar] [CrossRef]
- Smetana, V.; Šima, P.; Bogusch, P.; Erhart, J.; Holý, K.; Macek, J.; Roller, L.; Straka, J. Hymenoptera of the selected localities in the environs of Levice and Kremnica towns. Acta Musei Tekovensis Levice 2015, 10, 44–68. [Google Scholar]
- Šima, P.; Straka, J. First records of Heriades rubicola Pérez, 1890 (Hymenoptera: Megachilidae) and Nomada moeschleri Alfken, 1913 (Hymenoptera: Apidae) from Slovakia. Entomofauna Carpathica 2016, 28, 14–18. [Google Scholar]
- Mielczarek, L. The first records of Chalcosyrphus pannonicus (Ooldenberg, 1916) (Diptera: Syrphidae) in Poland and Slovakia. Dipteron 2014, 30, 50–54. [Google Scholar]
- Tóth, S. A Dél-Dunántúl bögöly faunájáról (Diptera: Tabanidae). Nat. Som. 2016, 28, 5–16. [Google Scholar]
- Nartshuk, E.P.; Krivokhatsky, V.; Evenhuis, N. First record of a bee fly (Diptera: Bombyliidae) parasitic on antlions (Myrmeleontidae) in Russia. Russ. Entomol. J. 2019, 28, 189–191. [Google Scholar] [CrossRef]
- Ábrahám, L. Micomitra stupida (Diptera, Bombyliidae): a new parasite of Euroleon nostras (Neuroptera, Myrmeleontidae). Dun. Dolg. Term. Tud. Sor. 1998, 9, 421–422. [Google Scholar]
- Yeates, D.K. The evolutionary pattern of host use in the Bombyliidae (Diptera): a diverse family of parasitoid flies. Biol. J. Linn. Soc. Lond. 1997, 60, 149–185. [Google Scholar] [CrossRef]
- Kells, A.R. , Goulson, D. Preferred nesting sites of bumblebee queens (Hymenoptera: Apidae) in agroecosystems in the UK. Biol. Conserv. 2003, 109, 165–174. [Google Scholar] [CrossRef]
- Day, M.C. Towards the conservation of aculeate Hymenoptera in Europe. – Council of Europe Press, Strasbourg, France, 1991; 44 pp.
- Williams, P.H. The distribution and decline of British bumblebees (Bombus Latr.). J. Apic. Res. 1982, 121, 236–245. [Google Scholar] [CrossRef]
- Plowright, C.M.S.; Plowright, R.C.; Williams, P.H. Replacement of Bombus muscorum by Bombus pascuorum in northern Britain? Can. Entomol. 1997, 129, 985–990. [Google Scholar] [CrossRef]
- Jakab D, Tóth M, Szarukán I, Szanyi S, Józan Z, Sárospataki M, Nagy A (2023) Long-term changes in the composition and distribution of the Hungarian bumble bee fauna (Hymenoptera, Apidae, Bombus). J. Hymenopt. Res. 2023, 96, 207–237. [CrossRef]
- Sárospataki, M.; Novák, J.; Molnár, V. Hazai poszméhfajok (Bombus spp.) veszélyeztetettsége és védelmük szükségessége. Bumblebees of Hungary (Bombus spp.) and their conservation. Természetvédelmi Közlemények 2004, 11, 299–307. [Google Scholar]
- Šima, P.; Smetana, V.; Čížek, J. Records of Bombus (Megabombus) argillaceus (Hymenoptera: Apidae: Bombini) from Slovakia and notes on the record from the Czech Republic. Klapalekiana 2014, 50, 101–106. [Google Scholar]
- Šima, P.; Smetana, V. Bombus (Cullumanobombus) semenoviellus (Hymenoptera: Apidae: Bombini) new species for the bumble bee fauna of Slovakia. Klapalekiana 2012, 48, 141–147. [Google Scholar]
- Smetana, V. Čmele a spoločenské osy (Hymenoptera: Bombini, Polistinae et Vespinae) na vybraných lokalitách v národnom parku Veľká Fatra. Acta Mus. Tek. Levice 2008, 7, 23–33. [Google Scholar]
- Smetana, V. Výsledky výskumu čmeľovitých (Hymenoptera: Bombidae) na vybraných lokalitách v Čergove, Bachurni a v bradlovom pásme Spišsko-šarišského medzihoria. Nat. Carp. 2000, 41, 59–66. [Google Scholar]
- Šima, P.; Smetana, V. Čmele (Hymenoptera: Bombidae) na vybraných lokalitách Popradskej kotliny. Nat. Tut. 2019, 23, 169–180. [Google Scholar]
- Tkalců, B. Vier für die Slowakei neu festgestellte Bienenarten (Hymenoptera, Apoidea). Biológia (Bratisl. ) 1973, 28, 679–687. [Google Scholar]
- Móczár, M. Magyarország és a környező területek dongóméheinek (Bombus Latr.) rendszere és ökológiája. Taxonomy and ecology of bumblebees (Bombus Latr.) of Hungary and the surrounding areas. Annls hist. -nat. Mus. natn. (Ser. Nov.) 1953, 4, 131–159. [Google Scholar]
- Móczár; M. A dongóméhek (Bombus Latr.) faunakatalógusa (Cat. Hym. IV). Faunistic catalogue of Bumblebees (Cat. Hym. IV). Folia ent. hung. (Ser. Nov.) 1953, 6, 197–228. [Google Scholar]
- Tanács; L. The Apoidea (Hymenoptera) of the Tisza–dam. Tiscia 1975, 10, 55–66. [Google Scholar]
- Biella, P.; Ćetković, A.; Gogala, A.; Neumayer, J.; Sárospataki, M.; Šima, P.; Smetana, V. Northwestward range expansion of the bumblebee Bombus haematurus into Central Europe is associated with warmer winters and niche conservatism. Insect Sci. 28(3), 861–872. [CrossRef] [PubMed]
- Šima, P. , Smetana, V. Quo vadis Bombus haematurus? An outline of the ecology and biology of a species expanding in Slovakia. Acta Mus. Tek. Levice 2018, 11, 41–65. [Google Scholar]
- Biella, P.; Galimberti, A. The spread of Bombus haematurus in Italy and its first DNA barcode reference sequence. Fragm. Entomol. 2020, 52, 1–67. [Google Scholar] [CrossRef]
- Rasmont, P.; Franzén, M.; Lecocq, T.; Harpke, A.; Roberts, S.P.M.; Biesmeijer, J.C.; Castro, L.; Cederberg, B.; Dvořák, L.; Fitzpatrick, Ú.; et al. Climatic Risk and Distribution Atlas of European Bumblebees. Biorisk 2015, 10, 1–236. [Google Scholar] [CrossRef]
- Miličić, M.; Vujić, A.; Cardoso, P. Effects of climate change on the distribution of hoverflyspecies (Diptera: Syrphidae) in Southeast Europe. Biodivers. Conserv. 2018, 27, 1173–1187. [Google Scholar] [CrossRef]
- IUCN SSC HSG/CPSG European Hoverflies: Moving from Assessment to Conservation Planning. A report to the European Commission by the IUCN SSC Conservation Planning Specialist Group (CPSG) and the IUCN SSC Hoverfly Specialist Group (HSG). Conservation Planning Specialist Group, Apple Valley, MN, USA. 2022, 84 pp.
- Sommaggio, D.; Zanotelli, L.; Vettorazzo, E.; Burgio, G.; Fontana, P. Different Distribution Patterns of Hoverflies (Diptera: Syrphidae) and Bees (Hymenoptera: Anthophila) along Altitudinal Gradients in Dolomiti Bellunesi National Park (Italy). Insects 2022, 13, 293. [Google Scholar] [CrossRef]
- Herczeg, T.; Száz, D.; Blahó, M.; Barta, A. , Gyurkovszky, M.; Farkas, R.; Horváth, G. 2015: The effect of weather variables on the flight activity of horseflies (Diptera: Tabanidae) in the continental climate of Hungary. Parasitol. Res. 2015, 114, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Dörge, D.D.; Sarah Cunze, S.; Klimpel, S. Incompletely observed: niche estimation for six frequent European horsefly species (Diptera, Tabanoidea, Tabanidae). Parasit Vectors. 2020, 13, 461–471. [Google Scholar] [CrossRef]
- Régnière, J.; Thireau, J.C.; Saint-Amant, R.; Martel, V. Modeling Climatic Influences on Three Parasitoids of Low-Density Spruce Budworm Populations. Part 3: Actia interrupta (Diptera: Tachinidae). Forests 2021, 12, 1471–1487. [Google Scholar] [CrossRef]
- Boesi, R.; Polidori, C.; Andrietti, F. Searching for the Right Target: Oviposition and Feeding Behavior in Bombylius Bee Flies (Diptera: Bombyliidae). Zool. Stud. 2009, 48, 141–150. [Google Scholar]
- Fox., R.J.P. Fox., R.J.P. Citizen science and Lepidoptera biodiversity change in Great Britain. PhD. Thesis. University of Exeter 2020, 297 pp. https://ore.exeter.ac.uk/repository/bitstream/handle/10871/120925/FoxR.pdf?sequence=1 (accessed on 18. 03. 2024).
- Conrad, K.F.; Warren, M.S.; Fox, R.; Parsons, M.S.; Woiwod, I.P. Rapid declines of common, widespread British moths provide evidence of an insect biodiversity crisis. Biol. Conserv. 2006, 132, 279–291. [Google Scholar] [CrossRef]
- Mikkola, K. Population trends of Finnish Lepidoptera during 1961-1996. Entomol. Fenn. 1997, 8, 121–143. [Google Scholar] [CrossRef]
- Hill, G.M.; Kawahara, A.Y.; Daniels, J.C.; Bateman, C.C.; Scheffers, B.R. Climate change effects on animal ecology: butterflies and moths as a case study. Biol. Rev. 2021, 96, 2113–2126. [Google Scholar] [CrossRef] [PubMed]
- Mangels, J.K. Land use and climate change: Anthropogenic effects on arthropod communities and functional traits. PhD. Thesis. Department of Biology, Technical University of Darmstadt (Germany), Darmstadt, 2017, pp. 169. https://tuprints.ulb.tu-darmstadt.de/7156/1/Dissertation_Mangels.pdf (accessed on 18. 03. 2024).
- Van Zandt, P.A.; Johnson, D.D.; Hartley, Ch.; LeCroy, K.A.; Shew, H.W.; Davis, T.B.; Lehnert, M.S. Which Moths Might be Pollinators? Approaches in the Search for the Flower-Visiting Needles in the Lepidopteran Haystack. Ecol. Entomol. 2019, 45, 13–25. [Google Scholar] [CrossRef]
- Turčáni, M.; Kulfan, J.; Minďáš, J. Penetration of the south European butterfly Libythea celtis (Laicharting 1782) northwards: Indication of global man made environmental changes? Ekológia 2003, 33, 28–41. [Google Scholar]
- Bury, J.; Maslo, D.; Obszarny, M.; Paluch, F. Expansion of Iphiclides podalirius (Linnaeus, 1758) (Lepidoptera: Papilionidae) on Podkarpacie Region (SE Poland) in 2010–2014. Park. Nar. I Rezerwaty Przyr. 2015, 34, 3–17. [Google Scholar]
- Gavrilov, M.B. , Radakovič, M.G., Sipos, Gy., Mezősi, G., Gavrilov, G., Lukić, T., Basarin, B., Benyhe, B., Fiala, K., Kozák, P. ; Perič, Z.M.; et al. Aridity in the central and southern Pannonian basin. Atmosphere 2020, 11–12, 1269–1287. [Google Scholar] [CrossRef]
Table 1.
Number of individuals of sawflies between 1971 and 2022, netsweeping method.
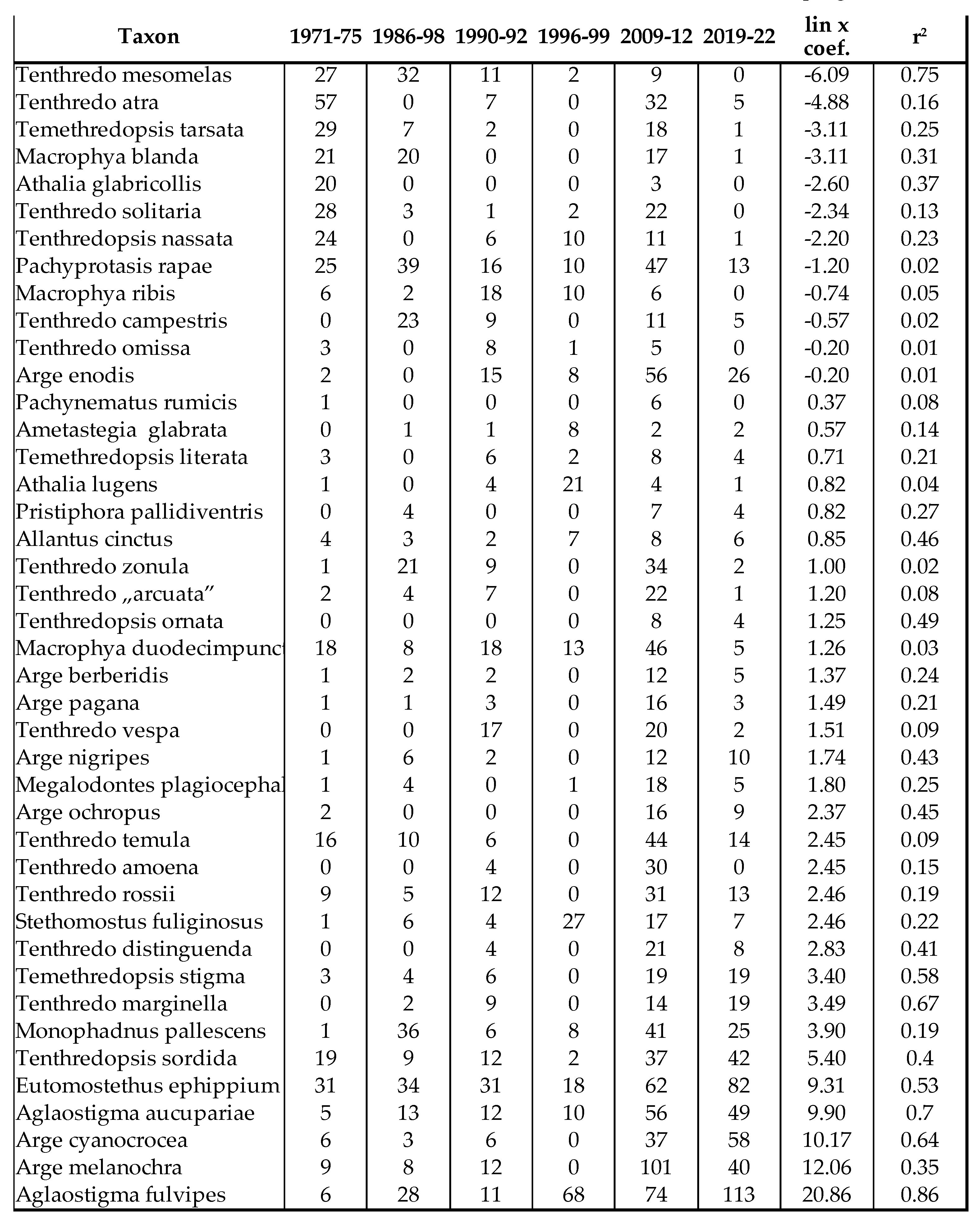
Table 2.
Population densities of sawflies between 1991 and 2021, Malaise trap.
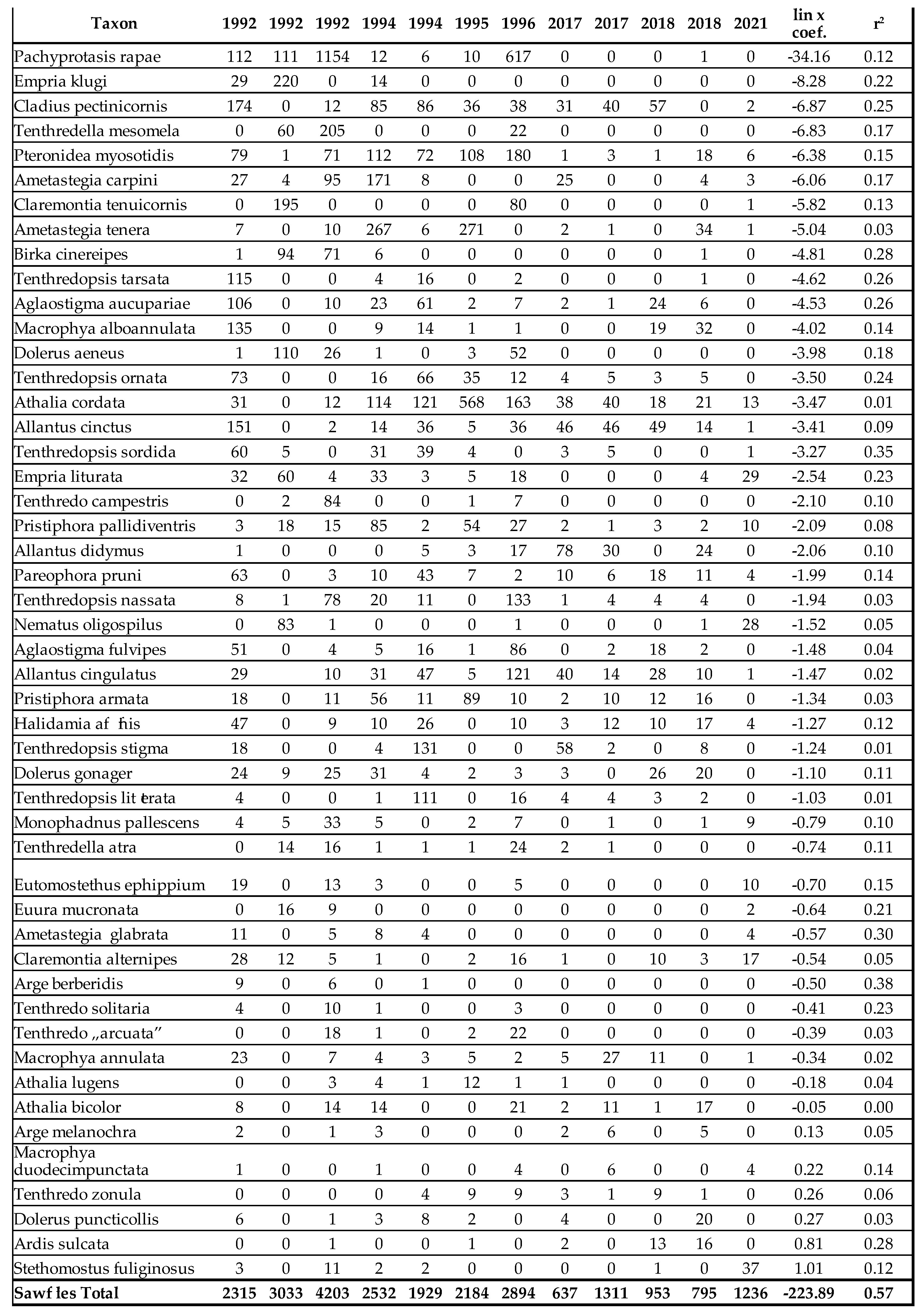
Table 3.
Earlier sporadic species, not recorded in the Pannonian biogeographic region in the last 20 years.
Table 3.
Earlier sporadic species, not recorded in the Pannonian biogeographic region in the last 20 years.
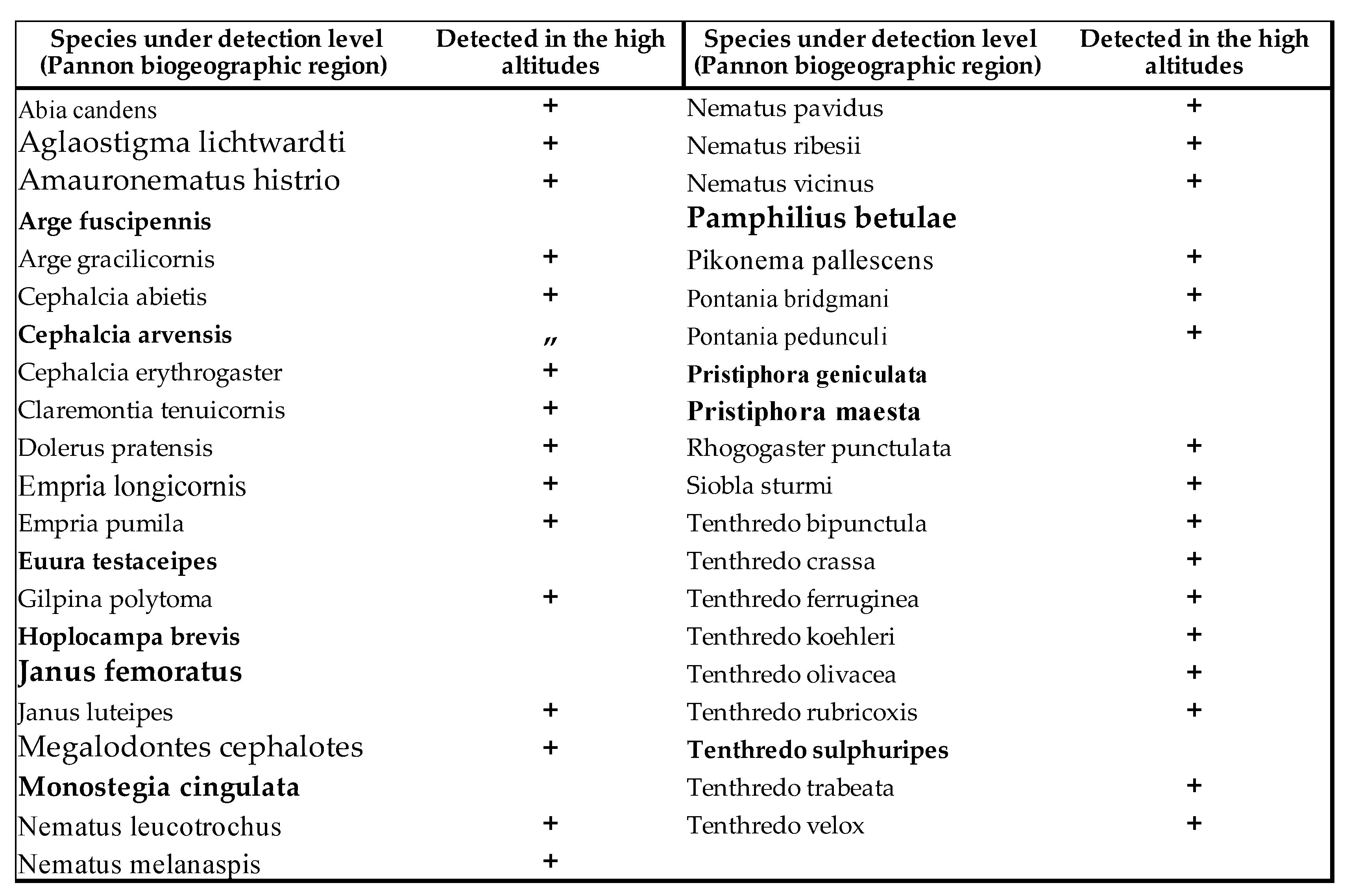
Table 4.
Number of individuals of various Aculeata genera and families collected between 1988 and 2023.
Table 4.
Number of individuals of various Aculeata genera and families collected between 1988 and 2023.
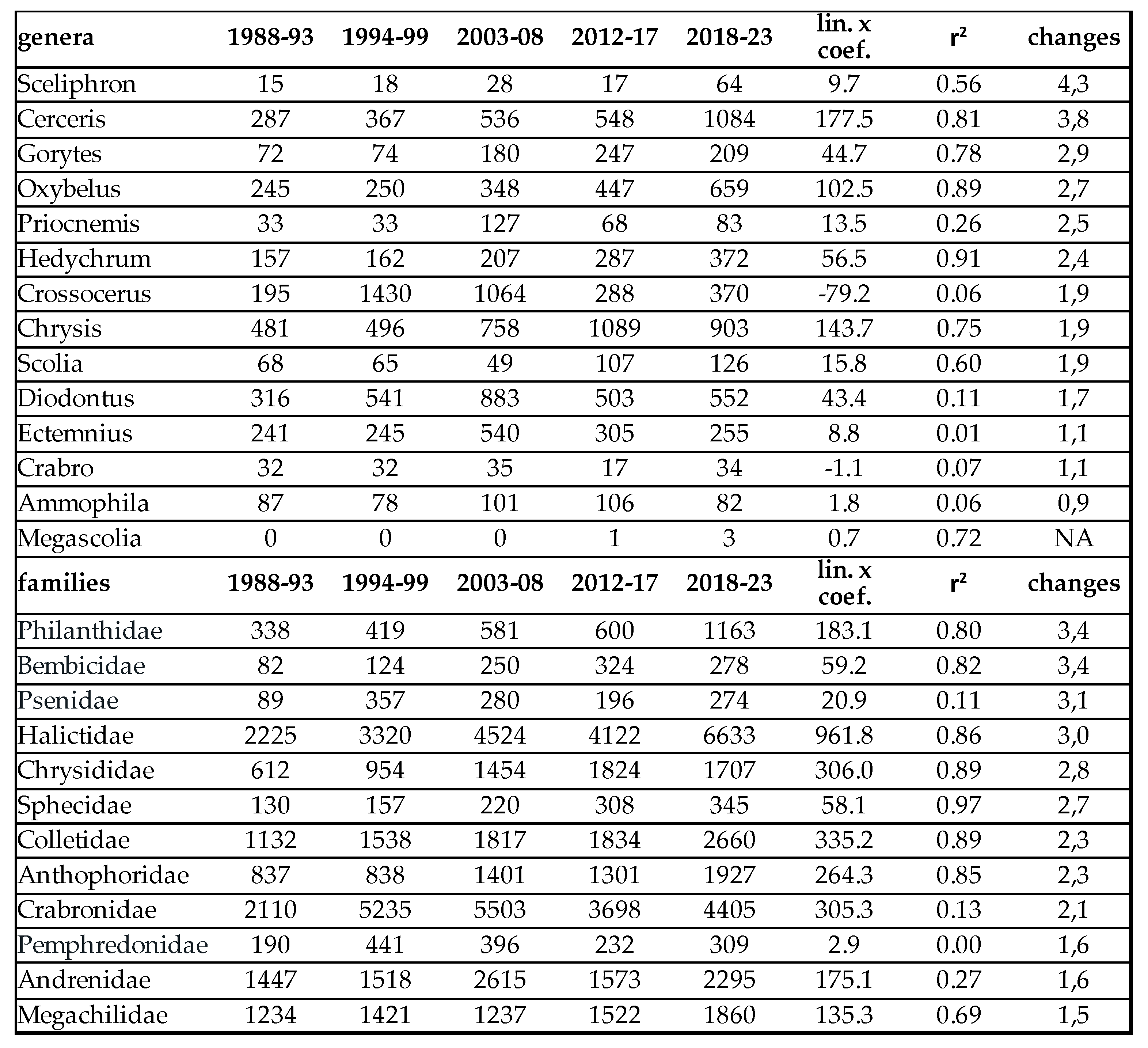
Table 5.
Invasive and recently appeared expansive Aculeata species between 1988 and 2023 (Hungarian data above, Slovak data below).
Table 5.
Invasive and recently appeared expansive Aculeata species between 1988 and 2023 (Hungarian data above, Slovak data below).
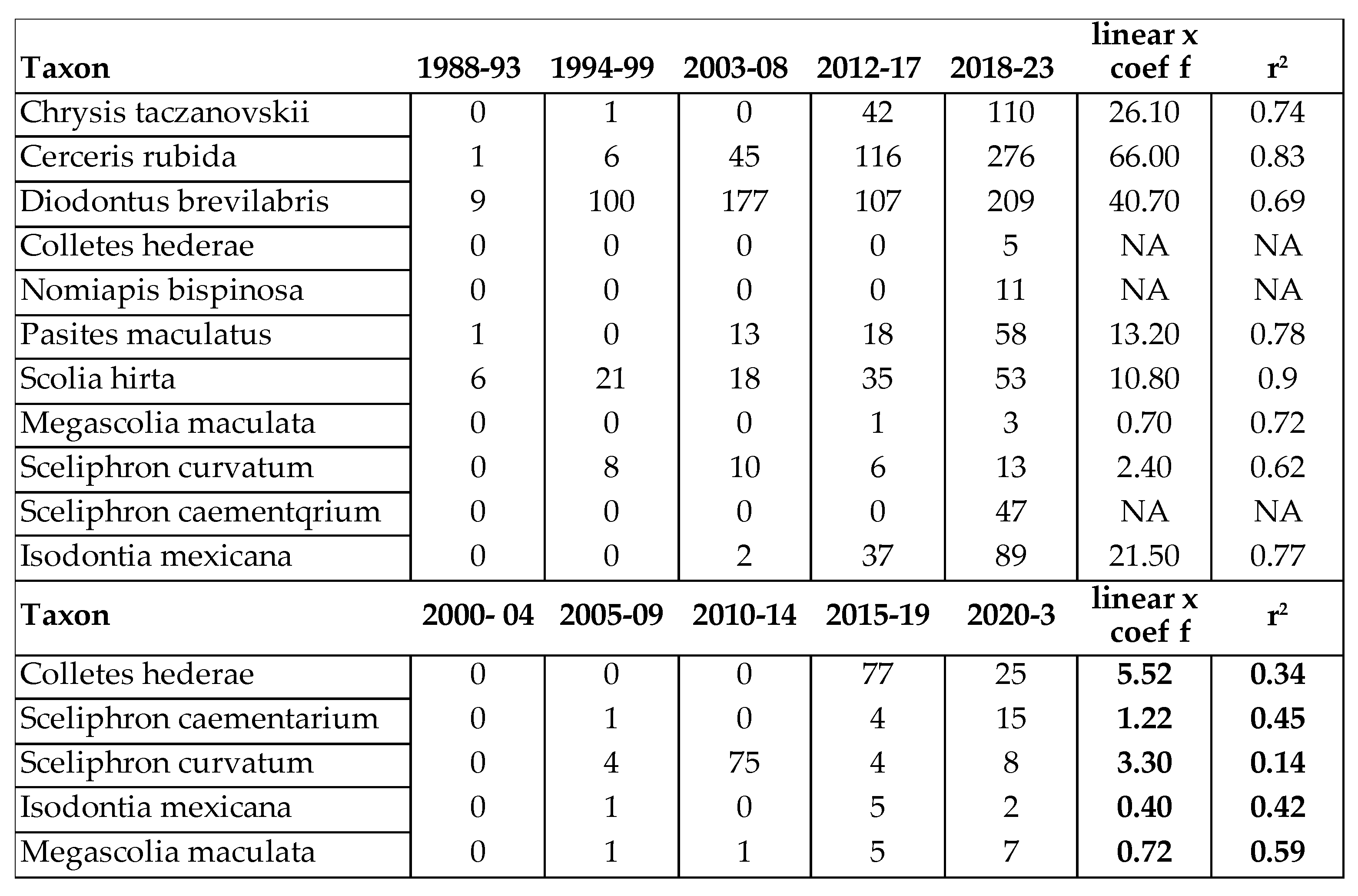
Table 7.
Number of individuals of various bumblebee and cuckoo bumblebee species between 1980 and 2023.
Table 7.
Number of individuals of various bumblebee and cuckoo bumblebee species between 1980 and 2023.
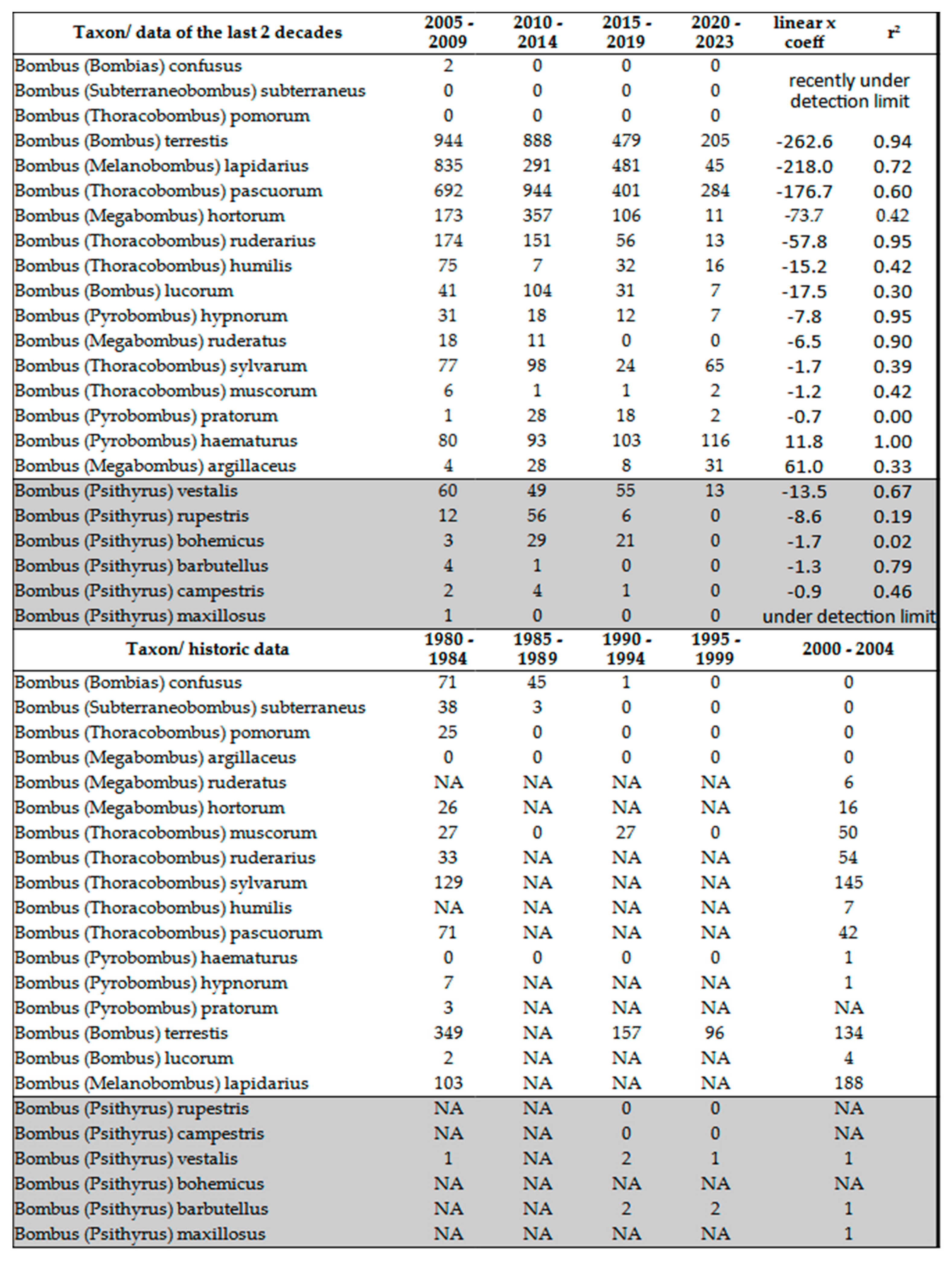
Table 8.
Number of individuals of various hoverfly species between 1980 and 2010.
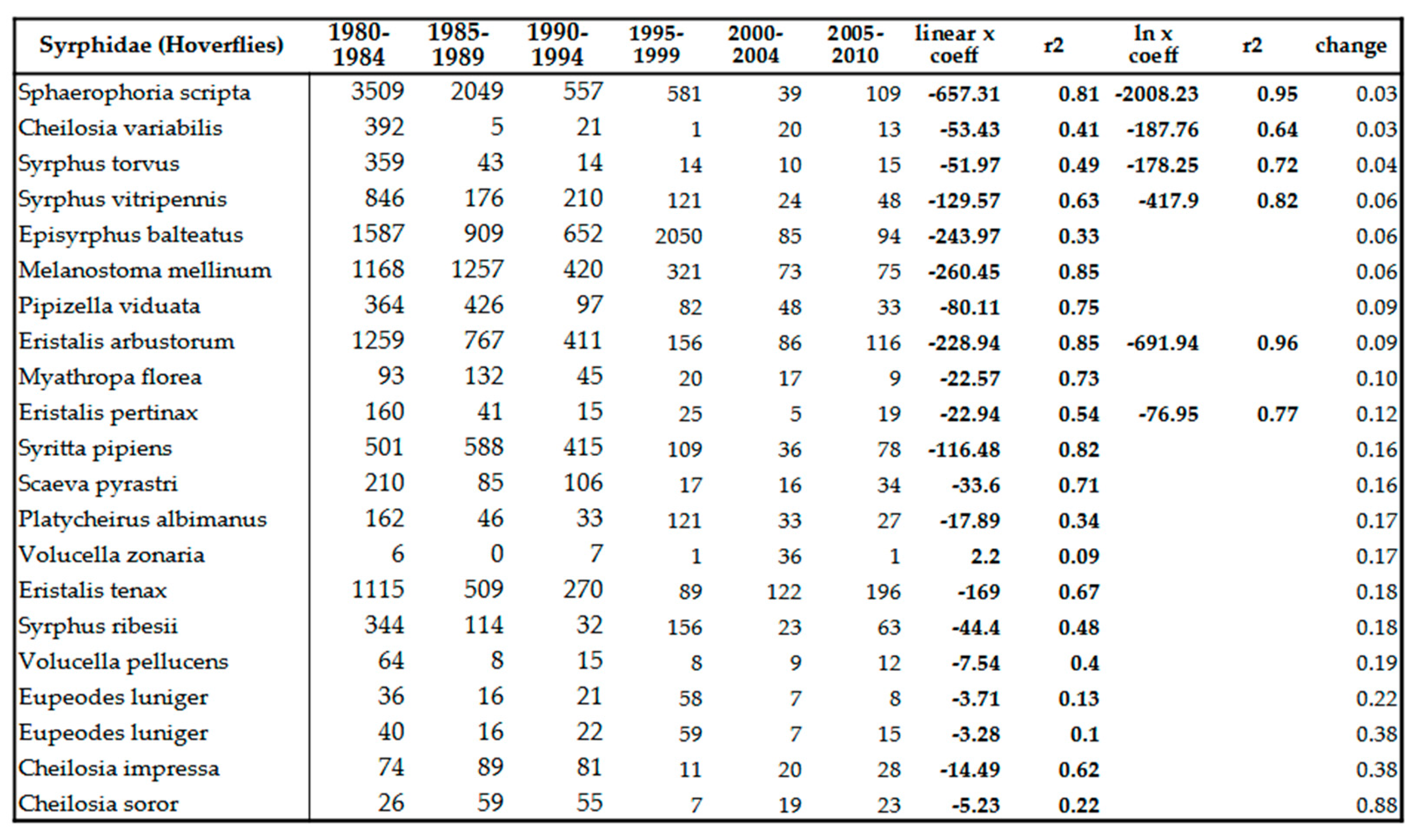
Table 9.
Number of individuals of various horse-fly species between 1980 and 2014.
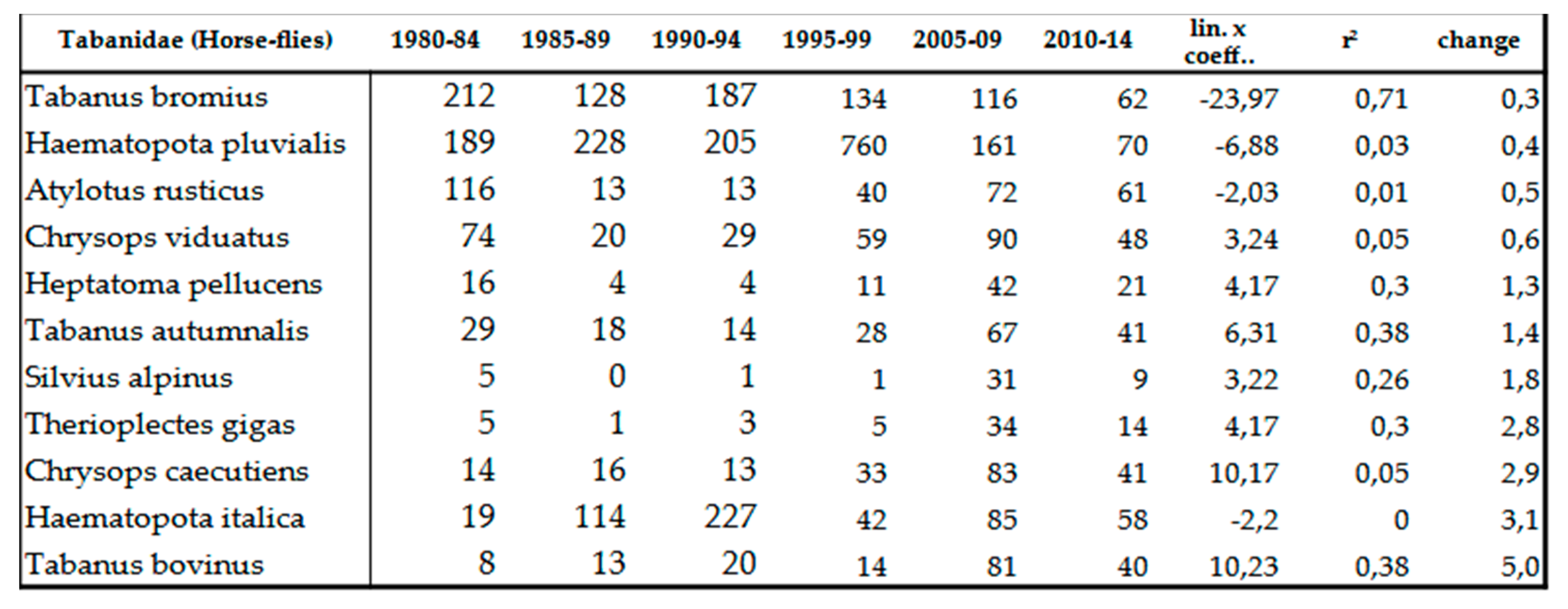
Table 10.
Number of individuals of various bee fly species between 1980 and 2020.
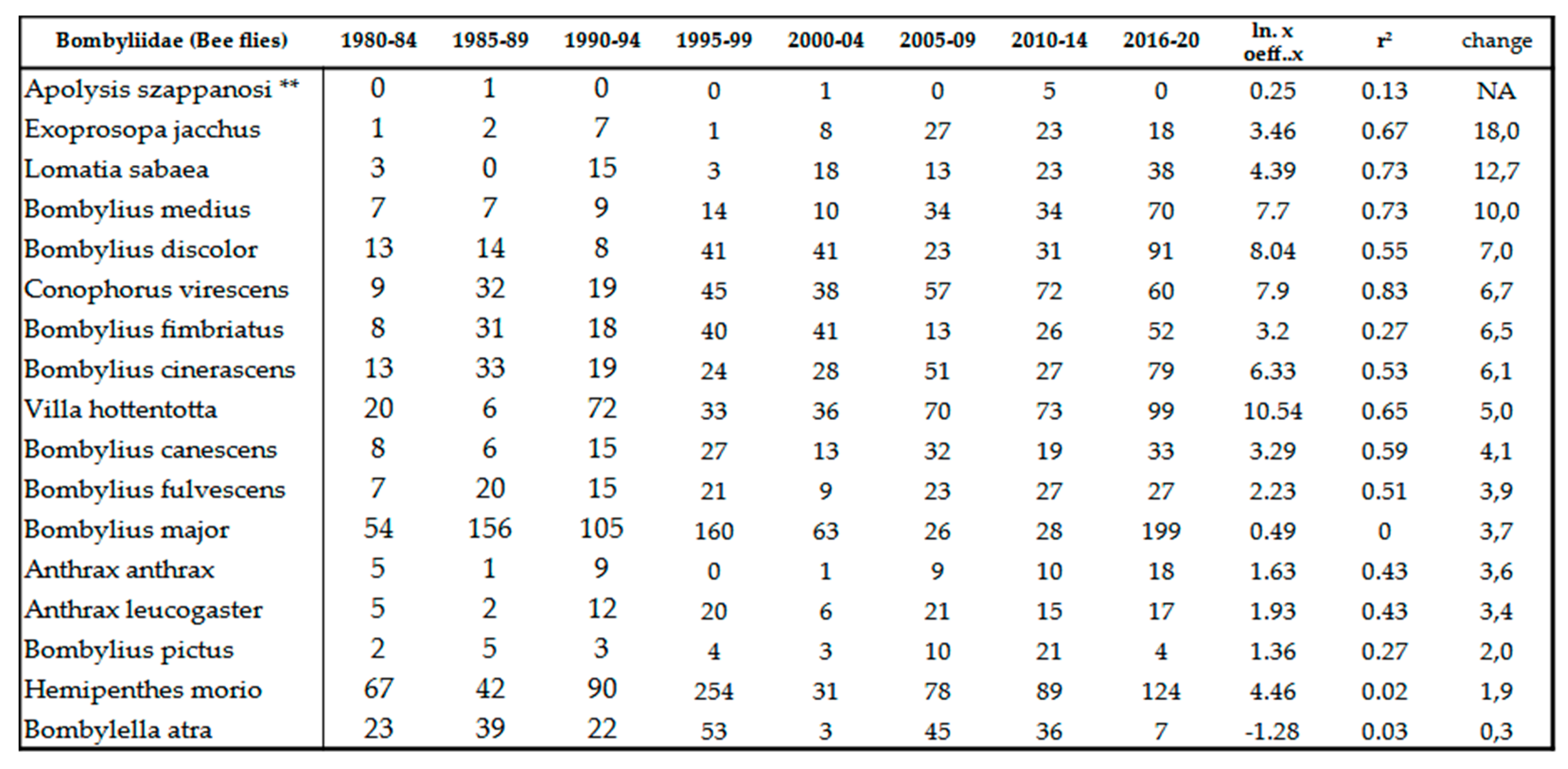
Table 14.
Changes in frequency of various moths in the last 50 years.

Table 15.
Changes in frequency of various moths families in the last 50 years.
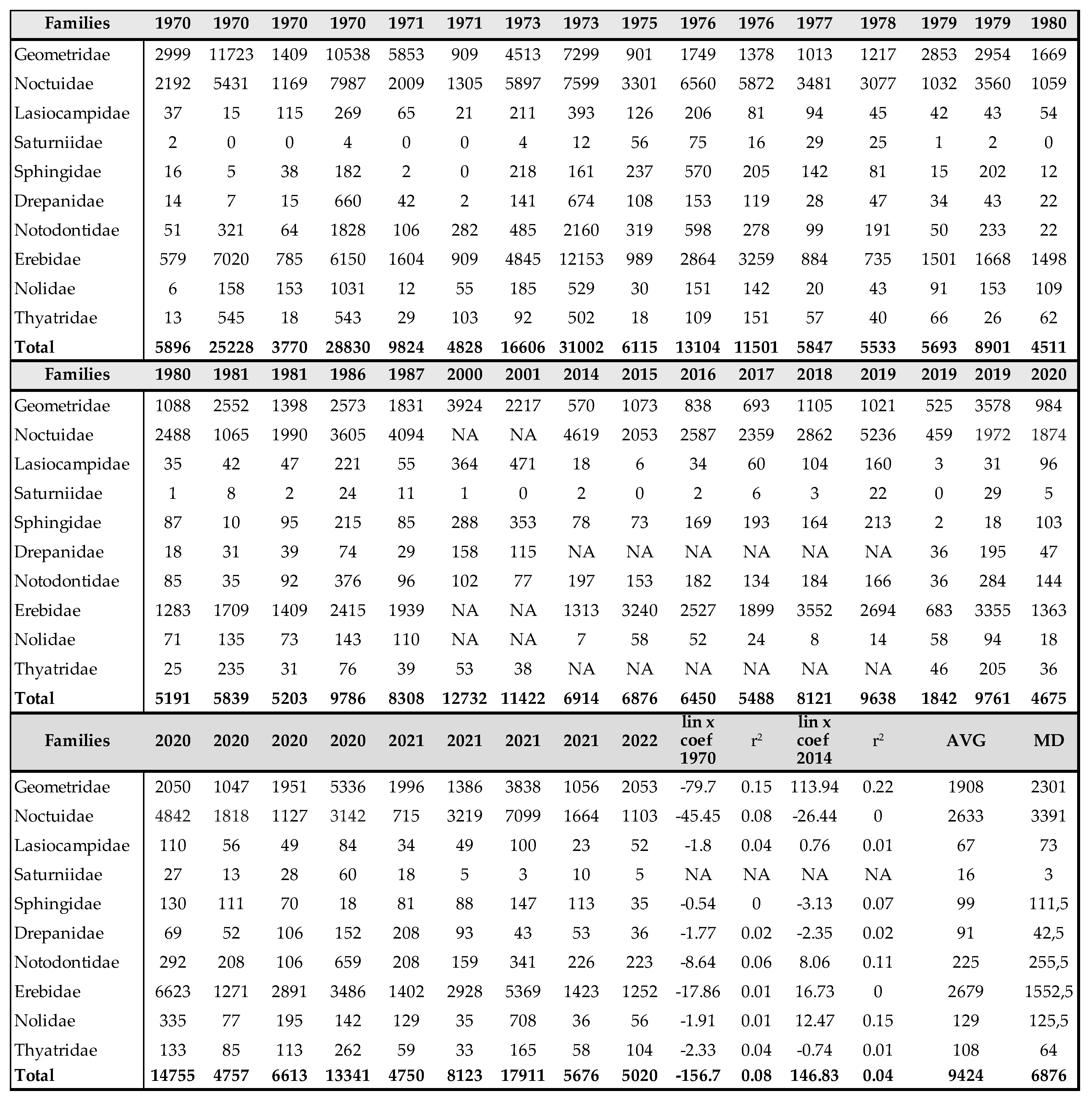
Table 16.
Changes in species richness of various moths families in the last 50 years.
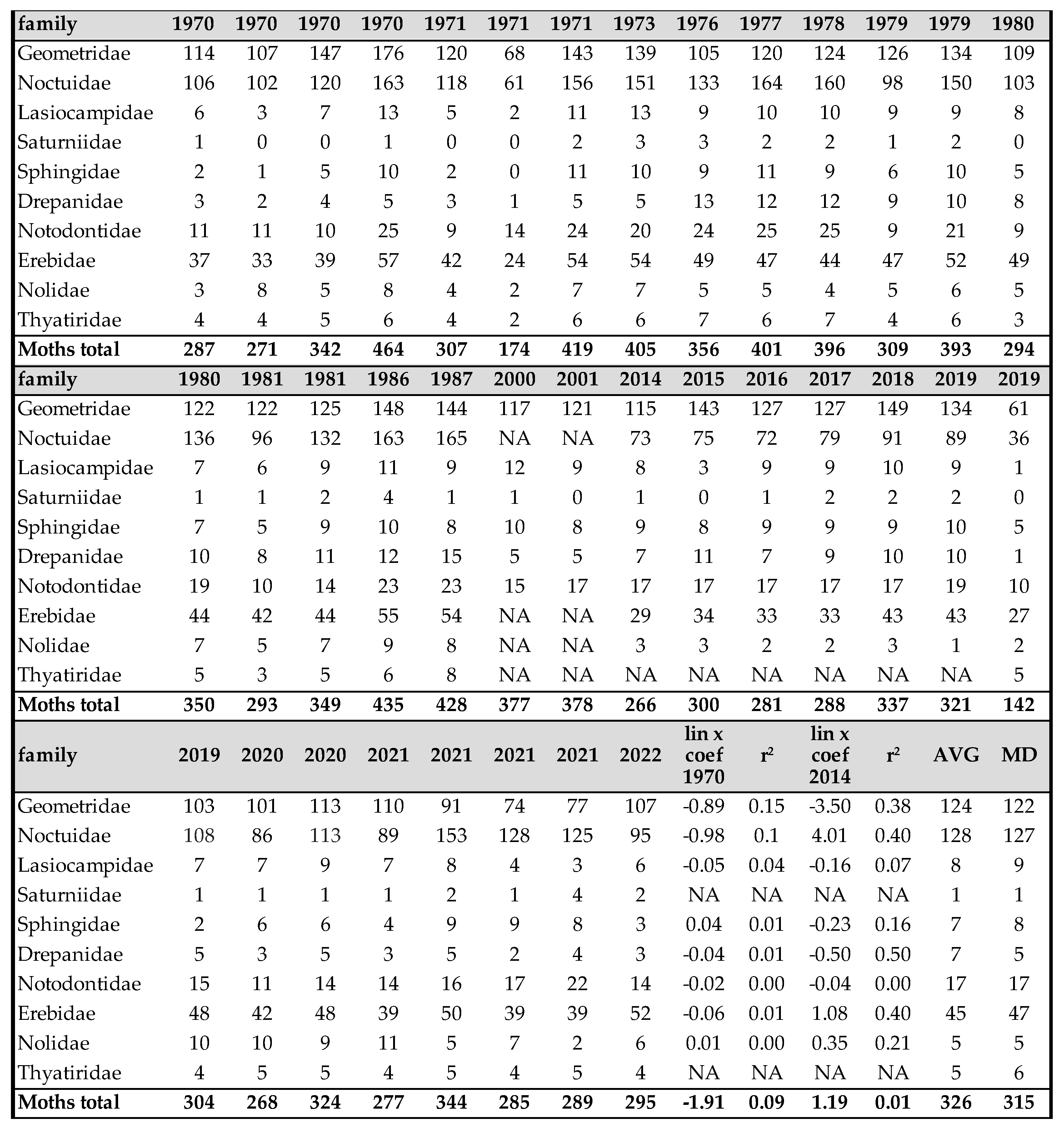
Table 17.
Invasive, expansive and introduced moths species in the last 50 years.
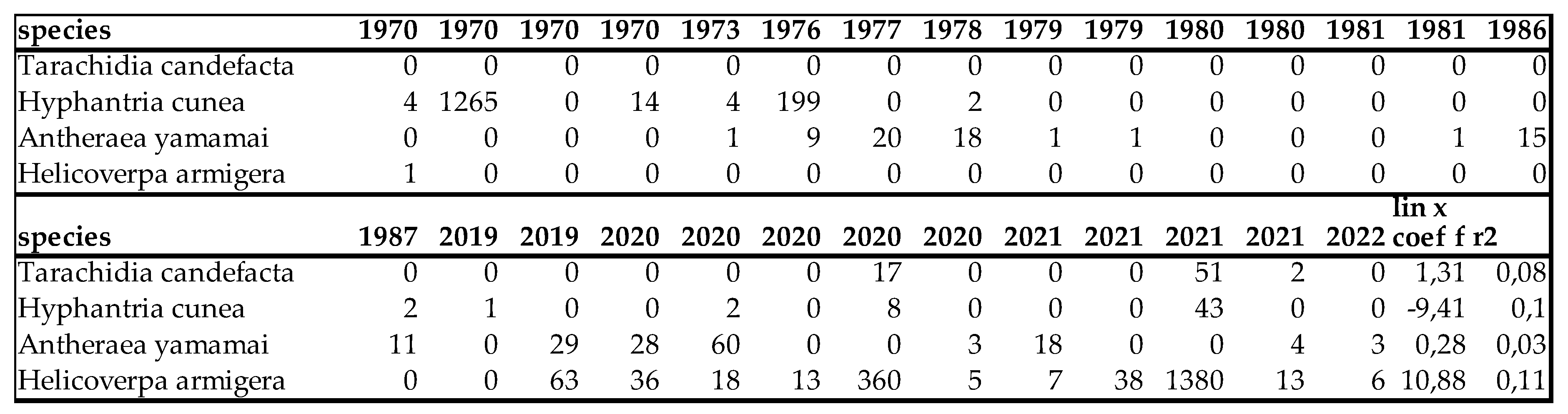
Table 18.
Relative frequency of some butterfly species in the last 40 years.
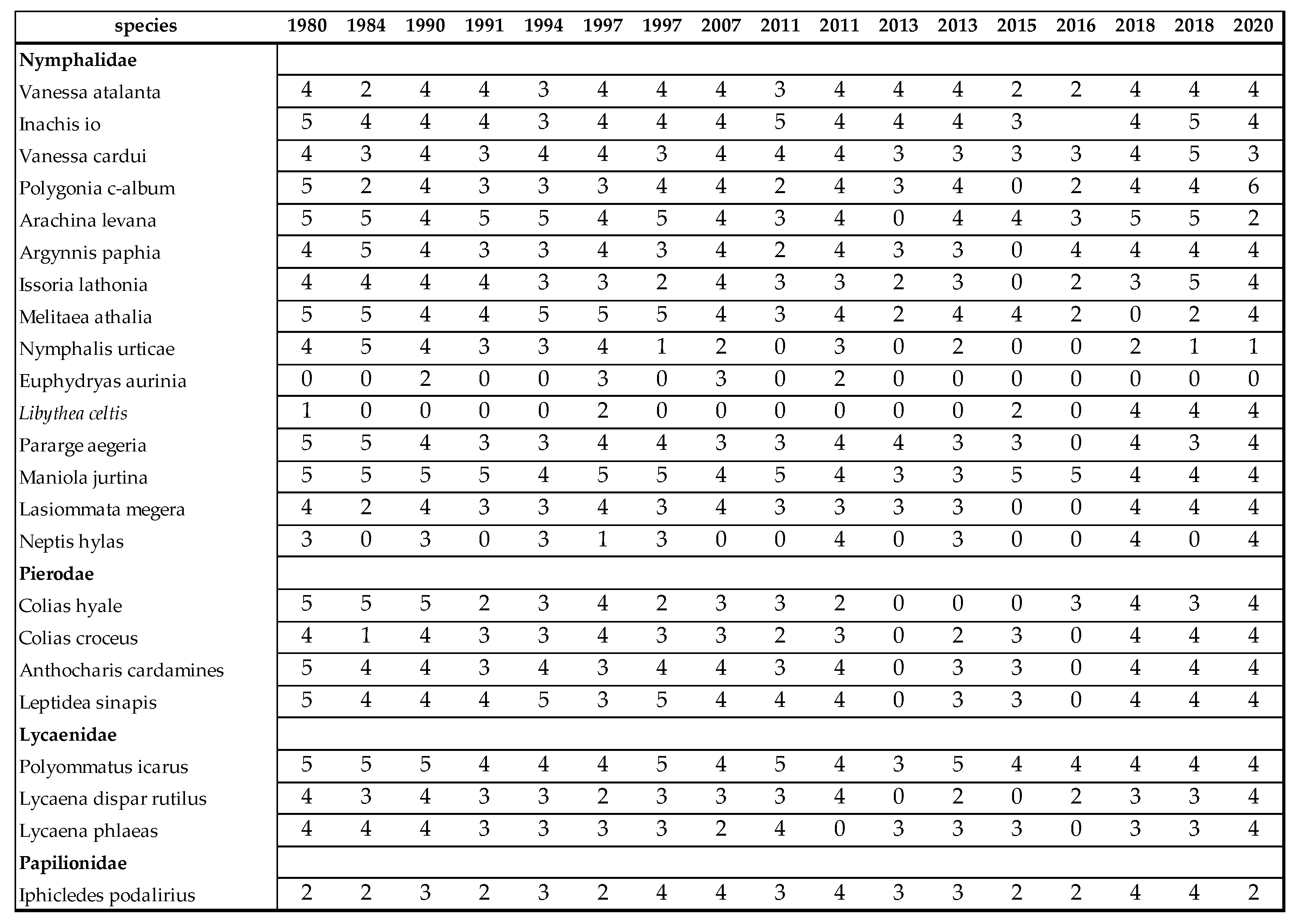
Table 19.
Changes of butterfly species richness in various regions after 40, 50 and 100 years.
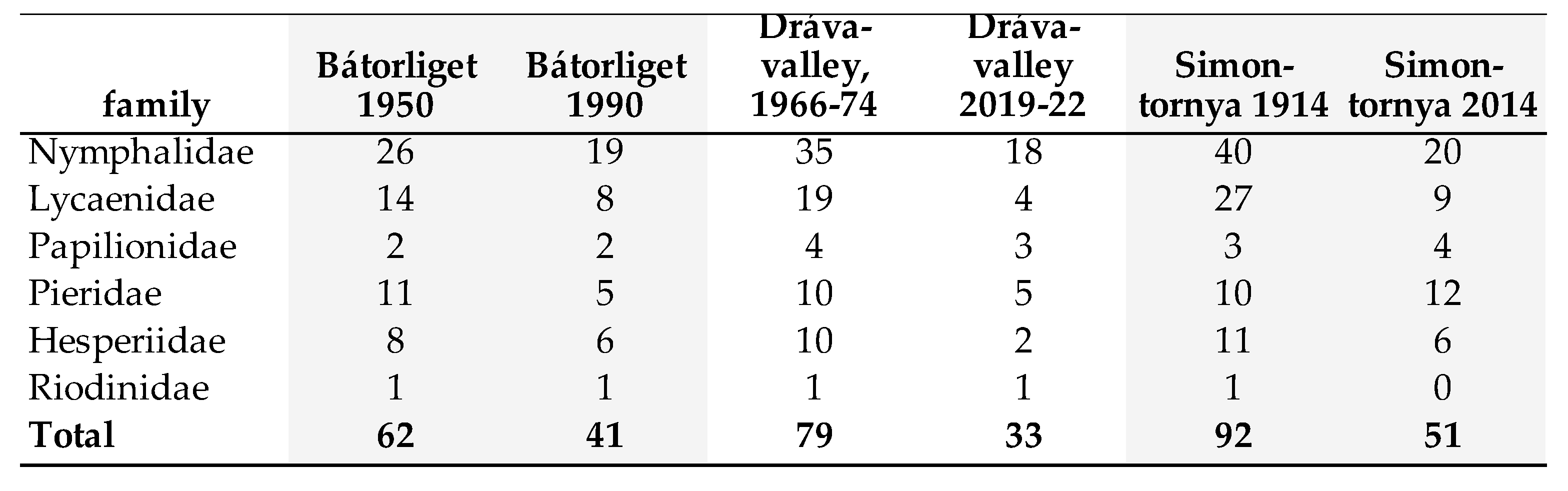
Figure 1.
Changes of sawfly populations between 1991 and 2021 with trend line and equation (based on Table 2).
Figure 1.
Changes of sawfly populations between 1991 and 2021 with trend line and equation (based on Table 2).
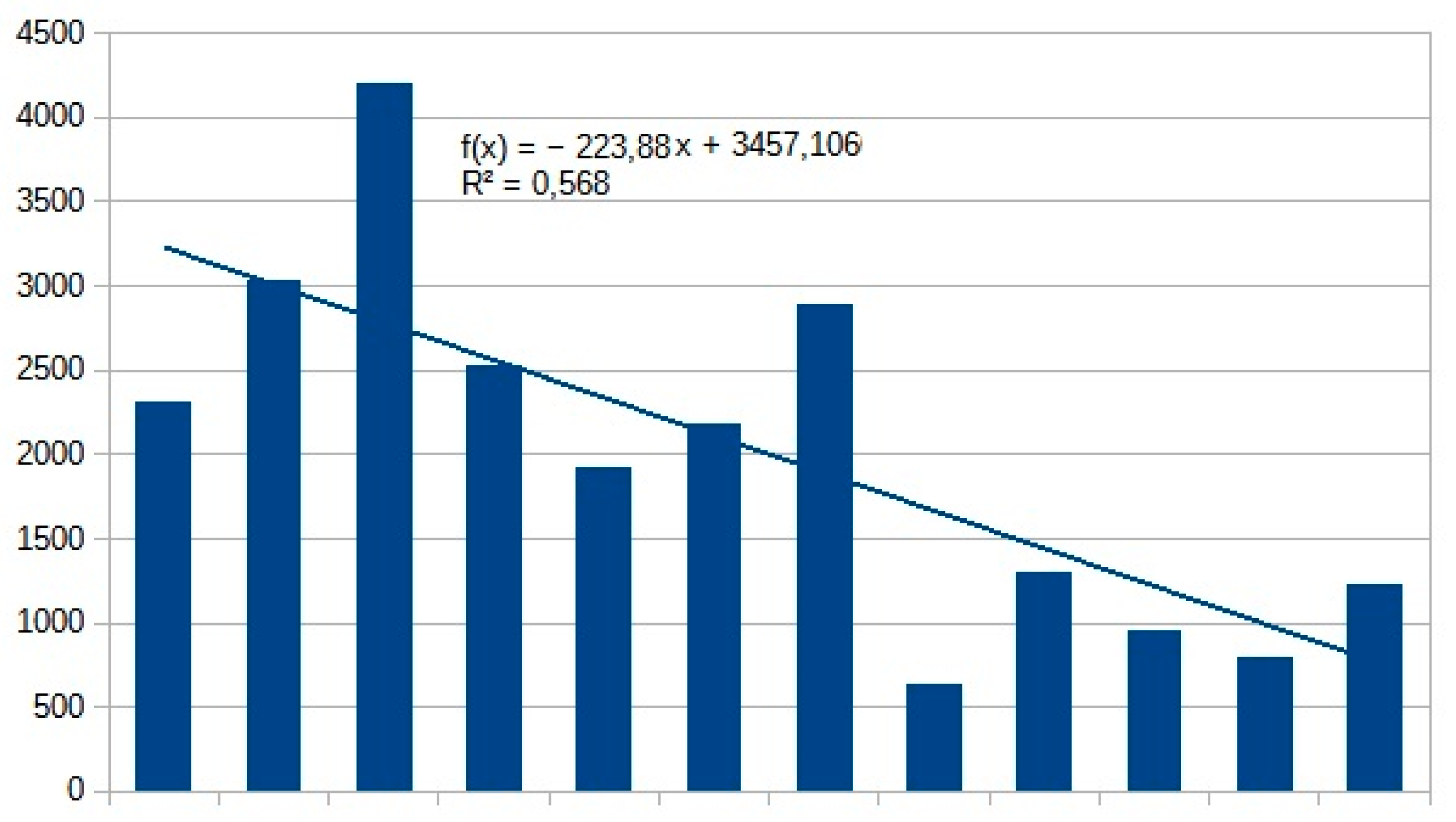
Figure 2.
Changes of Aculeata populations between 1988 and 2023 with trend line and equation (based on Table 4).
Figure 2.
Changes of Aculeata populations between 1988 and 2023 with trend line and equation (based on Table 4).
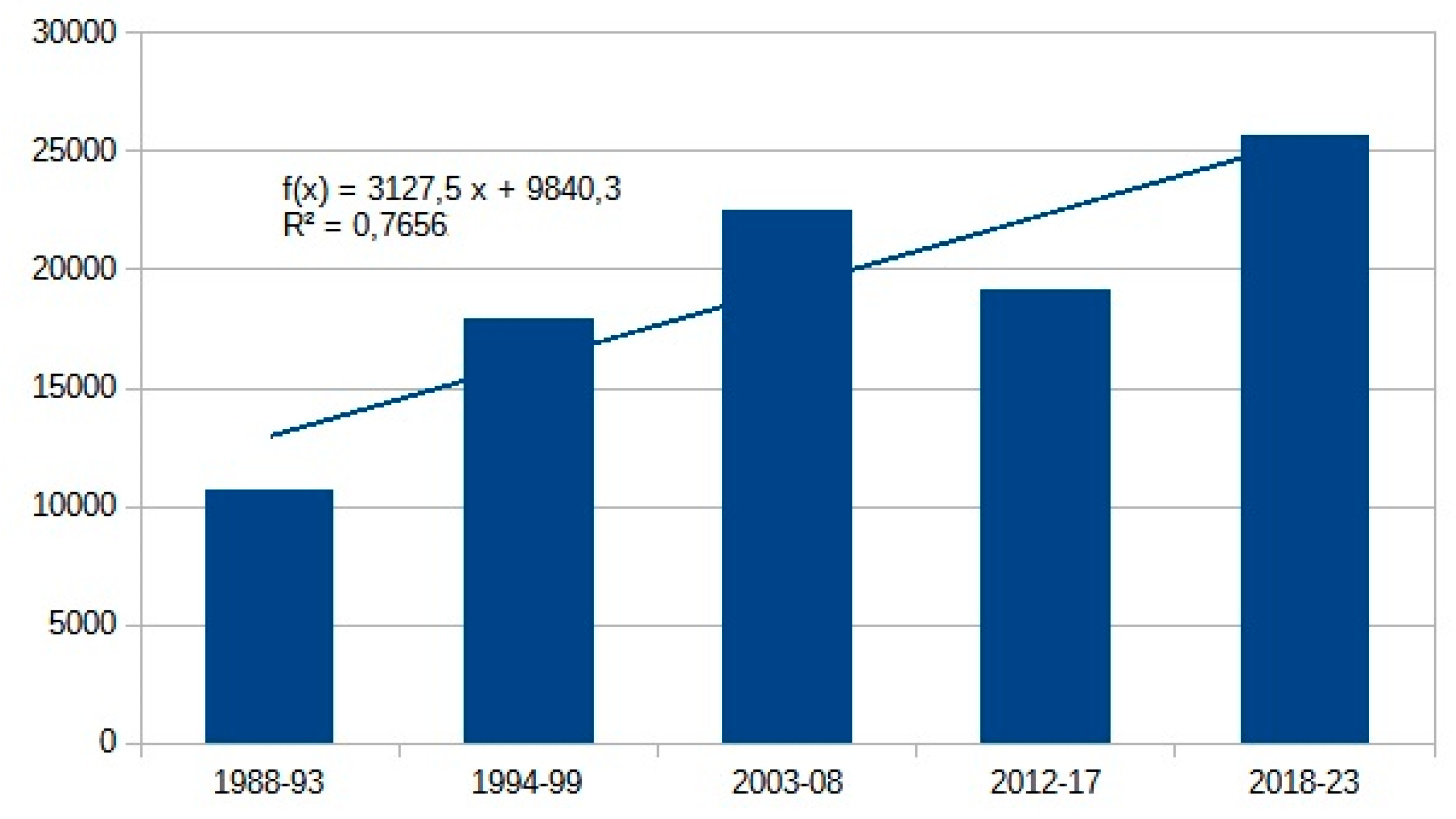
Figure 3.
Changes of moth populations between 1970 and 2023 with trend line and equation (based on Table 15).
Figure 3.
Changes of moth populations between 1970 and 2023 with trend line and equation (based on Table 15).
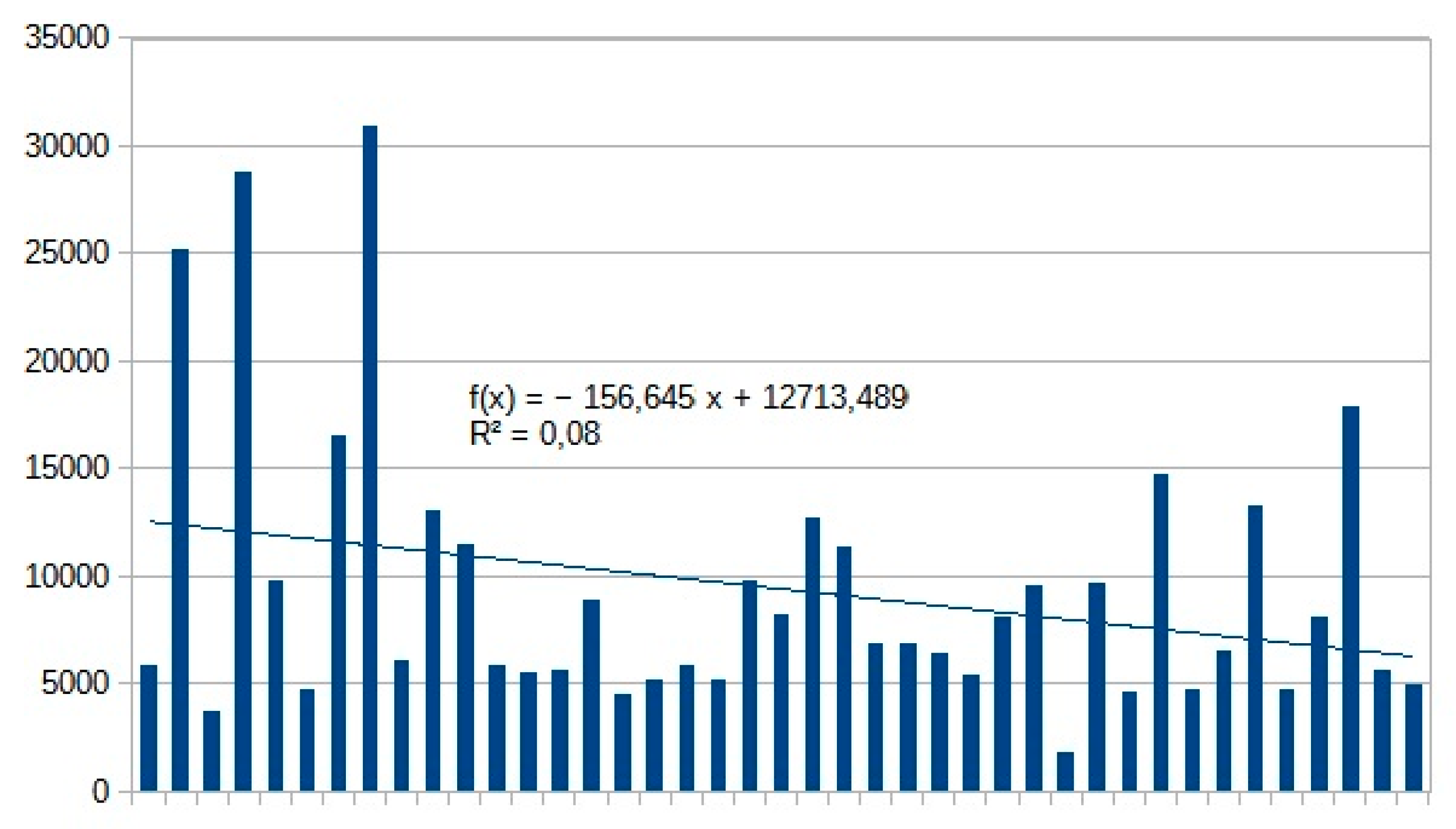
Figure 4.
Changes of moth species richness 1970 and 2023 with trend line and equation (based on Table 16).
Figure 4.
Changes of moth species richness 1970 and 2023 with trend line and equation (based on Table 16).
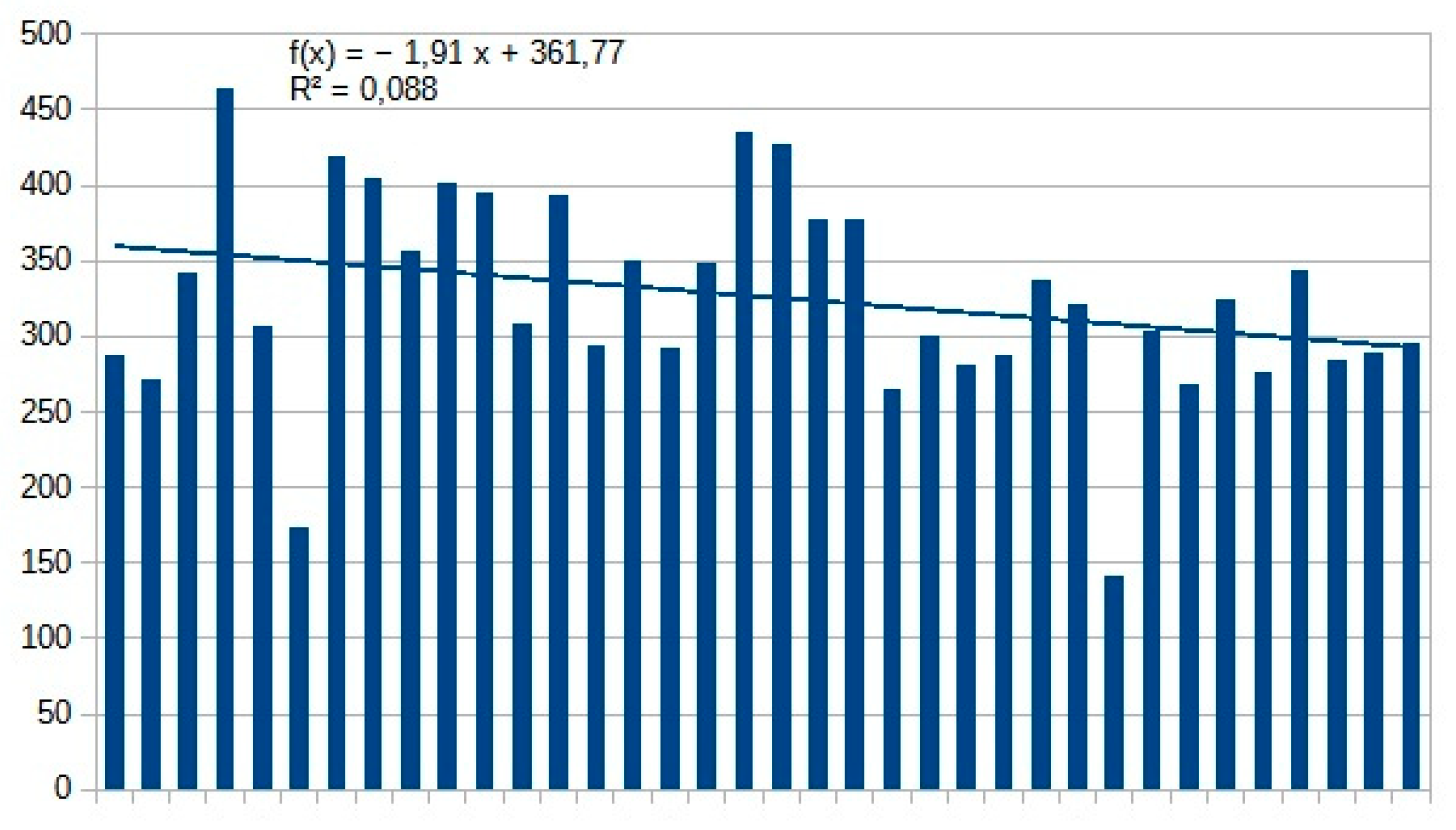
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



