
Submitted:
12 August 2025
Posted:
13 August 2025
You are already at the latest version
Abstract
Mangrove ecosystems near Panama City are under increasing pressure from human activities such as urban expansion and deforestation. These habitats are crucial for pollinators like orchid bees (tribe Euglossini), yet little is known about how these bees respond to such environmental stressors. In this study, we investigated the diversity and composition of orchid bee communities in mangrove forests from two coastal are-as in Panama with contrasting levels of human disturbance: Panama Bay and Chame Bay. Over six months, we used McPhail traps baited with eucalyptus oil to sample bees across multiple zones within each site. We recorded a total of 427 individuals across 14 species from three genera. Species richness and abundance were notably lower in the more urbanized mangroves, with Eulaema nigrita dominating most collections. Statis-tical analyses (GLMM, PERMANOVA, PCoA) confirmed significant differences in community structure between sites. These findings suggest that anthropogenic pres-sures may reduce pollinator diversity in mangrove forests, with potential implications for plant reproduction and ecosystem functioning. Our results highlight the im-portance of preserving and restoring mangrove habitats to support orchid bee popula-tions and maintain biodiversity in tropical coastal ecosystems.
Keywords:
Euglossini bees
; conservation
; deforestation
; fragmentation
; mangrove forests
; perturbation
1. Introduction
Orchid bees (tribe Euglossini, Apidae: Hymenoptera) are brightly colored pollinators endemic to the Neotropics [1]. Known for their iridescent hues and perfume-collecting behavior, these bees play a vital role in pollinating a wide range of tropical plant species, especially orchids [2]. The tribe includes five genera—Aglae Lepeletier & Serville, 1828; Exaerete Hoffmannsegg, 1817; Eufriesea Cockerell, 1908; Eulaema Lepeletier, 1841; and Euglossa Latreille, 1802—encompassing approximately 250 described species distributed from sea level to elevations above 2000 meters [2,3]. Unlike other corbiculate bees, orchid bees are solitary and exhibit distinct nesting and foraging behaviors [4]. Females of Euglossa species possess a corbicula on the hind tibia, while males show a thickening in the same area. Although they do not form colonies, some species may establish communal nests in natural cavities, including abandoned nests of other bees [5].
Panama and Costa Rica are considered hotspots of Euglossini diversity, together hosting over 70 species reported for Central America and southern Mexico—representing nearly 40% of the known species in the region [2,6]. In Panama, orchid bees have been highlighted not only as key pollinators but also as bioindicators of forest health due to their sensitivity to habitat disturbance [7,8]. However, rapid urban expansion, deforestation, and pollution along Panama’s Pacific coast—particularly around Panama Bay—have significantly altered mangrove ecosystems [9,10]. These changes may reduce the availability of floral and nesting resources, ultimately impacting the composition and abundance of orchid bee communities [11,12].
Despite their ecological importance, there is a lack of research focused specifically on Euglossini communities in mangrove habitats of Panama. Given their role as sensitive indicators of environmental change, studying their response to varying degrees of anthropogenic disturbance is crucial for developing informed conservation strategies [7,13]. This study aims to compare the composition and diversity of orchid bee communities in two mangrove systems under contrasting levels of human disturbance: Panama Bay (urbanized) and Chame Bay (conserved). We hypothesize that the urbanized site will present lower species richness and evenness, with a community structure dominated by generalist or disturbance-tolerant species.
2. Materials and Methods
2.1. Study area
Panama Bay, located south of the province of the same name, is a vast body of water forming part of the Gulf of Panama. With an approximate length of 96 km and a maximum width of 35 km, its coasts include Panama City and the Pacific entrance to the Panama Canal. This ecosystem comprises estuaries, mangroves, floodplain forests, marshes, and freshwater lagoons [14]. It is recognized as a Ramsar site of international importance, serving as a habitat for one to two million migratory shorebirds, including species such as the western sandpiper (Calidris mauri) and the semipalmated plover (Charadrius semipalmatus) [15]. However, the bay faces significant environmental challenges due to urban development, pollution, and mangrove deforestation, leading to alterations in its ecological characteristics [9].
On the other hand, Chame Bay is located on the Pacific slope of Panama and is a semi-enclosed body of water with a system of shallow channels. It receives inputs from rivers and streams with low sedimentation, which has allowed the development of one of the best-preserved mangrove ecosystems on Panama’s Pacific coast [14]. This mangrove forest extends from the mouth of the Chame River to the Monte Oscuro district, covering more than 8 000 hectares [10]. Unlike Panama Bay, Chame Bay has experienced less human intervention, contributing to the preservation of its biodiversity and ecological functions. Nevertheless, recent studies have emphasized the need to monitor and manage human activities in the region to ensure the conservation of this valuable ecosystem [14].
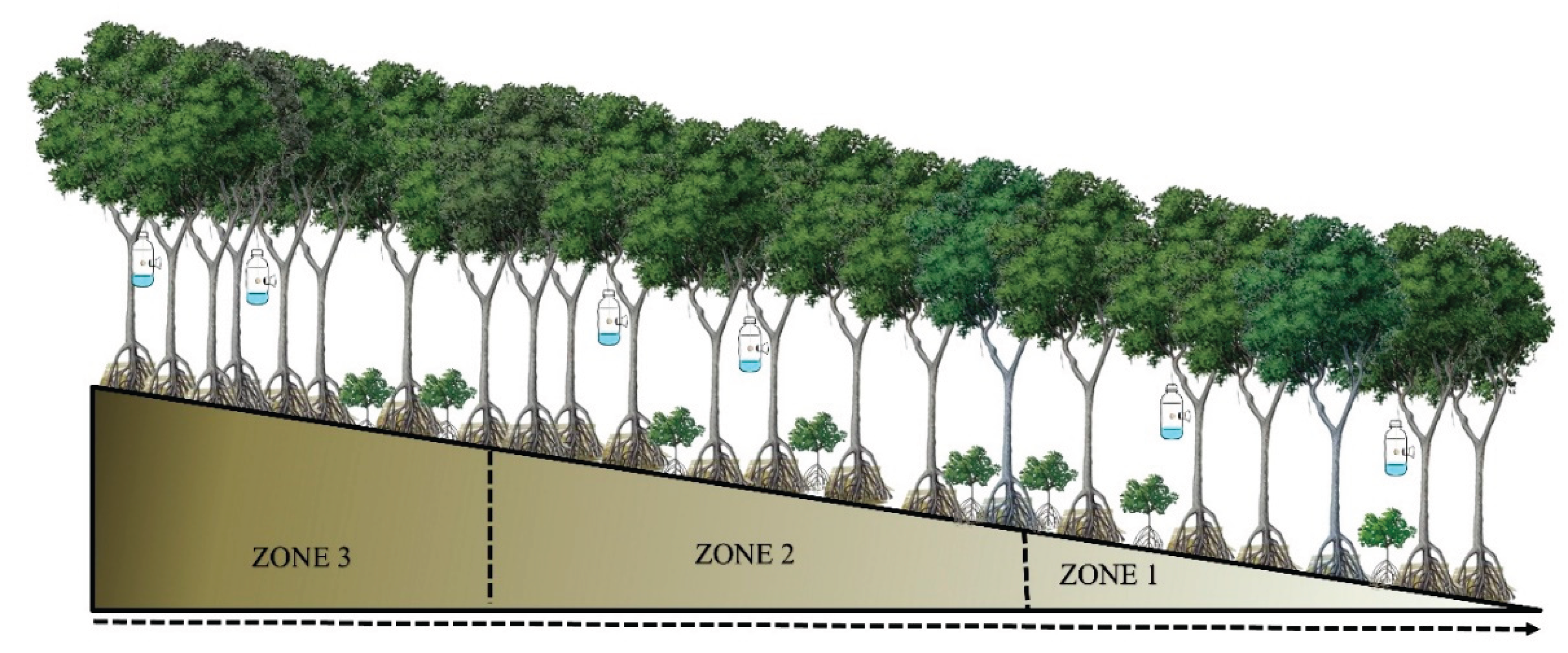
I collected bee specimens from orchids in four areas located in Panama Bay and Chame Bay: El Embarcadero, Don Bosco and Costa Sur, situated in the Panama Bay (coordinates 9.019036, -79.436975); and El Líbano, located in the Bay of Chame (coordinates 8.620592478338297, -79.80706863161527) (see Fig. 1). In both locations, three zones were outlined using plots measuring 0.4 km² (200 meters long by 200 meters wide). These plots extended from the tidal region to the mainland, divided into three zones: Zone 1 was closest to the shoreline, Zone 2 was positioned midway between the tidal area and the forest, and Zone 3 marked the boundary between the mangrove and the forest (Fig. 2).
Figure 1.
Map of the sampling area of orchid bees in Panama Bay and Chame Bay.
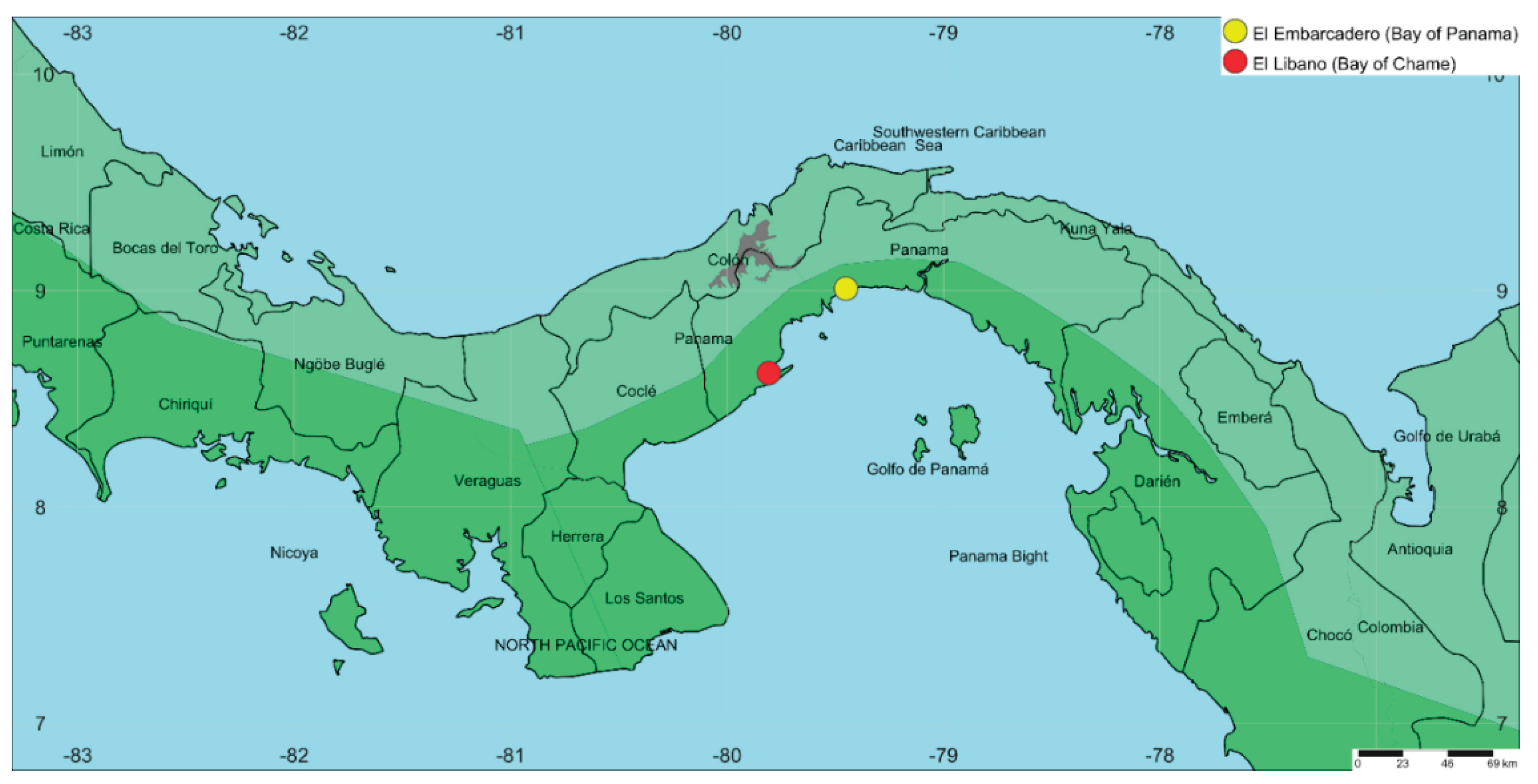
Figure 2.
Arrangement of the sampling zones inside the mangrove forest f Panama Bay and Chame Bay. Zone 1 is closest to the shoreline, zone 2 intermediate area, and zone 3 the closest to the tropical forest near the mangroves.
Figure 2.
Arrangement of the sampling zones inside the mangrove forest f Panama Bay and Chame Bay. Zone 1 is closest to the shoreline, zone 2 intermediate area, and zone 3 the closest to the tropical forest near the mangroves.
2.2. Sampling of Orchid Bees (Euglossini)
In both Panama Bay and Chame Bay, sampling was conducted over a six-month period using the same methodology. Each site included three 200 × 200 m plots, subdivided into three zones (Zone 1: closest to the shore, Zone 2: mid-mangrove, Zone 3: forest edge). Two McPhail traps were installed per zone, totaling six traps per site, with identical spatial distribution and trap height (~1.5 m above ground). Traps were baited exclusively with eucalyptus oil and remained active continuous throughout the study period. Samples were collected biweekly, resulting in a total of 12 collection events per trap at each site. This standardized sampling effort ensured comparability between sites in terms of trapping intensity, duration, and spatial coverage. The decision to use only eucalyptus oil was driven by budgetary constraints and time limitations, which prevented the incorporation of multiple chemical attractants. However, eucalyptus oil has been shown to be effective in capturing a broad range of orchid bee species in previous studies [2,16]. The traps were exposed throughout the entire project, and the collected material was retrieved biweekly. The samples were then taken to the Museo de Invertebrados G. B. Fairchild at the Universidad de Panamá (MIUP) for processing. Specimens were mounted on No. 2 pins, labeled, and identified using the taxonomic key in [2]. The specimens analyzed in this study were collected under collection permit ARB-133-2022, granted by the corresponding environmental authorities. All orchid bee specimens were preserved and deposited as voucher specimens in the entomological collection of the Museum of Invertebrates at the University of Panama (MIUP-HEU). The deposited vouchers are cataloged under the reference numbers MIUP-HEU-0001 to MIUP-HEU-0427, ensuring proper documentation and availability for future taxonomic and ecological studies. All specimens were preserved under controlled humidity and temperature conditions, following the curation protocols of the MIUP collection.
2.3. Data Analysis
All statistical analyses were conducted using R software (version 4.3.1) [17]. Alpha diversity metrics—species richness, Shannon index, Simpson index, Fisher’s alpha, and Pielou’s evenness—were calculated using the vegan and BiodiversityR packages to assess species heterogeneity within sites. To estimate and compare species richness relative to sampling effort, species accumulation and rarefaction curves were generated using the specaccum () and rarecurve () functions from the vegan package. Beta diversity was evaluated using the Bray–Curtis dissimilarity and Jaccard similarity index, calculated with the vegdist() function in vegan, to assess species turnover and community differentiation between sites. These dissimilarity matrices were used to perform a Principal Coordinates Analysis (PCoA) with the cmdscale() function, and visualizations were generated using ggplot2.
To statistically test differences in community composition between sites, we applied a Permutational Multivariate Analysis of Variance (PERMANOVA) using the adonis2() function in the vegan package with 999 permutations. To assess the effect of site on total species abundance, a Generalized Linear Mixed Model (GLMM) was fitted using the glmmTMB package. The model assumed a negative binomial distribution to account for overdispersion, with site as a fixed effect and sampling zone as a random effect. The significance of the model was assessed using Wald chi-square tests via the Anova () function from the car package. All plots were created using the ggplot2 and ggpubr packages.
3. Results
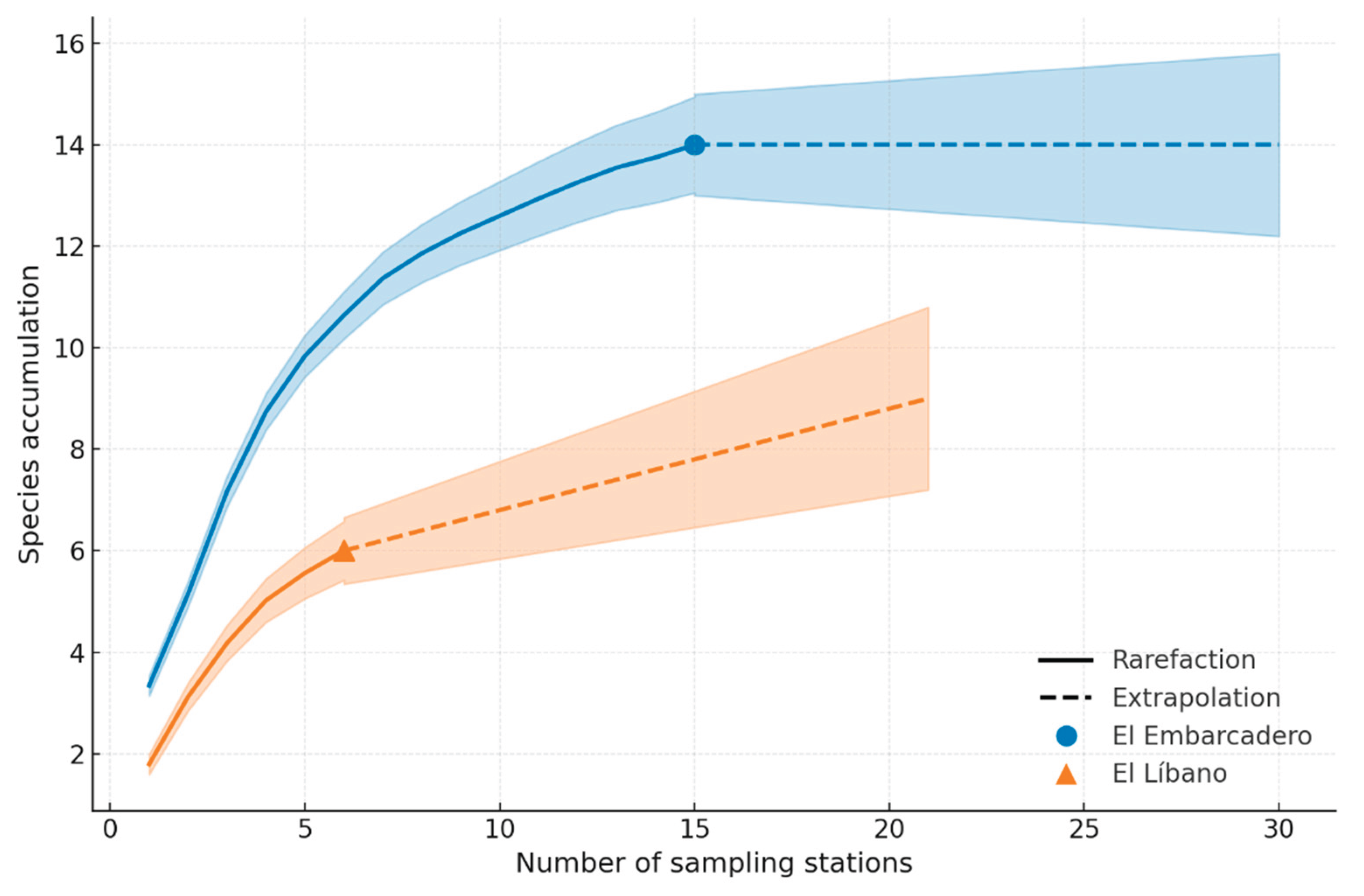
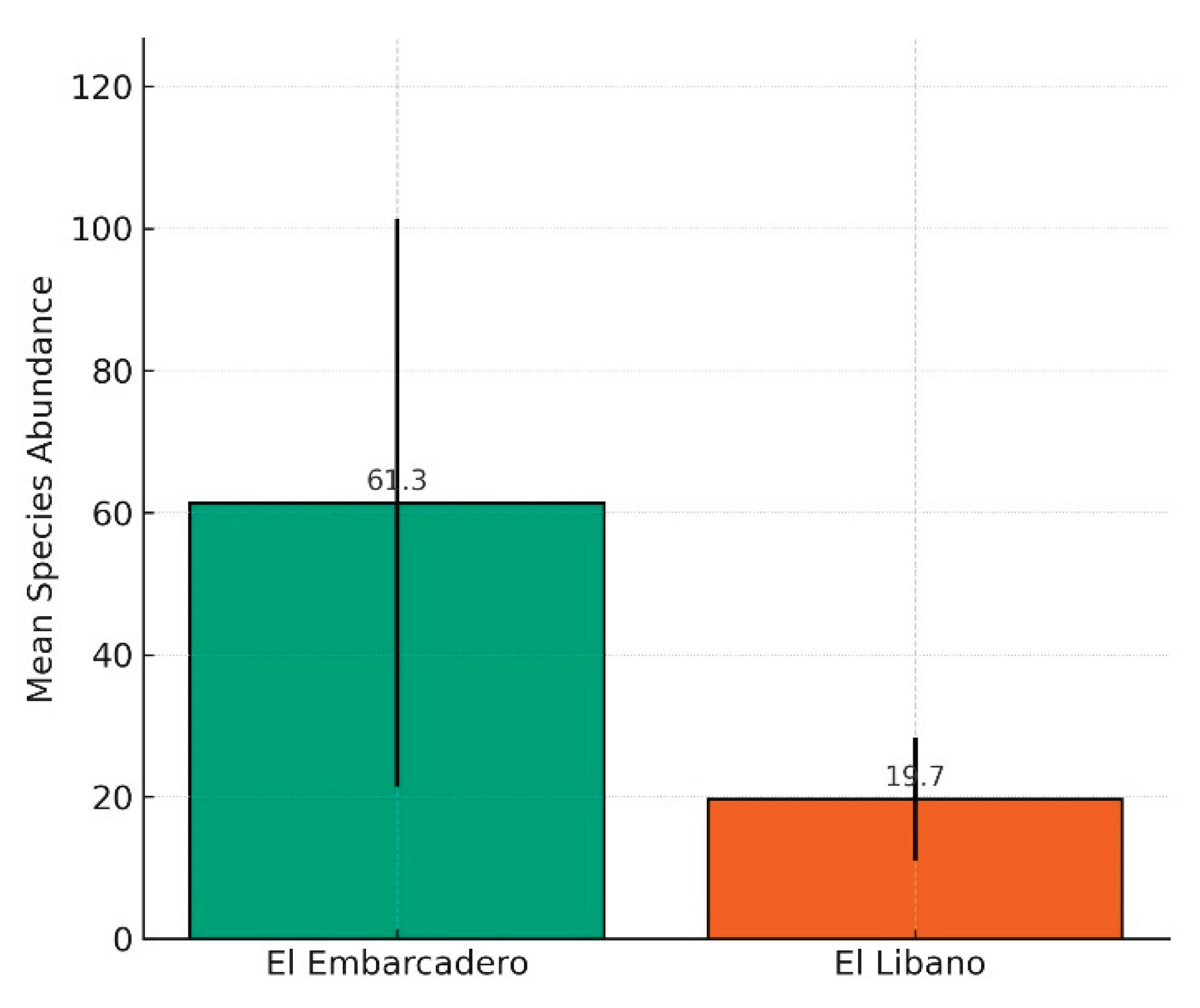
In six months of sampling in Panama and Chame Bays, a total of 427 specimens were collected, belonging to 14 species in three genera (Euglossa, Exaerete and Eulaema). The genus Euglossa presented an abundance of 117 individuals and 12 species, the genus Eulaema presented 234 individuals and one species, and the genus Exaerete presented one species and 17 individuals. Overall, Zone 1 had 106 individuals, Zone 2 had 134, and Zone 3 had 128. The species accumulation and rarefaction curves reveal key patterns in species richness at El Embarcadero and El Líbano. The species accumulation curve indicates that as the number of samples increases, the accumulated number of species also grows, although it tends to stabilize, suggesting that the sampling has captured most of the expected diversity. On the other hand, the rarefaction curve shows the estimated species richness as a function of sampling effort, highlighting differences in diversity between sites and allowing for a more equitable comparison of the number of detected species (Fig. 3). The GLMM analysis comparing species abundance between El Embarcadero and El Líbano shows a significant difference, with El Líbano having a lower species abundance compared to El Embarcadero (coefficient = -0.9832, p < 0.001). This indicates that species abundance in El Líbano is approximately 62 % lower than in El Embarcadero.
Figure 3.
Species accumulation and rarefaction curves in El Embarcadero and El Líbano; the species accumulation curve (left) shows the total number of recorded species as the number of samples increases, with a tendency to stabilize, indicating that the sampling has captured most of the expected diversity. The rarefaction curve (right) represents the estimated species richness as a function of sampling effort, allowing for a more equitable comparison of diversity between sites and highlighting differences in the number of detected species.
Figure 3.
Species accumulation and rarefaction curves in El Embarcadero and El Líbano; the species accumulation curve (left) shows the total number of recorded species as the number of samples increases, with a tendency to stabilize, indicating that the sampling has captured most of the expected diversity. The rarefaction curve (right) represents the estimated species richness as a function of sampling effort, allowing for a more equitable comparison of diversity between sites and highlighting differences in the number of detected species.
In El Líbano, Bahía de Chame, a total of 59 individuals from three genera (Euglossa, Exaerete, Eulaema) were collected, distributed across six species (Euglossa deceptrix, Euglossa imperialis, Euglossa variabilis, Eulaema meriana, Eulaema nigrita, and Exaerete smaragdina). In terms of distribution by zones, 22 individuals were recorded in Zone 1, 18 in Zone 2, and 19 in Zone 3. Species richness was highest in Zone 3 with four species, followed by Zone 2 with three species, and Zone 1 with two species. At the El Embarcadero site, a total of 319 orchid bee individuals were collected, distributed across three zones: 63 in Zone 1, 78 in Zone 2, and 85 in Zone 3. These collections allowed for the identification of 9 species, with Euglossa nigrita being the most abundant with 156 individuals. The species Euglossa imperialis and Euglossa smaragdina exhibited notable abundance with 30 and 14 individuals, respectively, while species such as Euglossa dressleri and Euglossa cybelia were present in lower numbers, with 8 and 12 individuals, respectively.
In Panama Bay, sampling was performed in three sites: El Embarcadero, Don Bosco and Costa Sur, in the months of April, May, June and July 2022; obtaining a total of 319 individuals, the area of El Embarcadero presented the most individuals with 226; Costa Sur and Don Bosco showed low abundance with 70 and 23 individuals, respectively. Regardind total richness of the sampling the highest diversity was found at El Embarcadero with nine species. In contrast, the Costa Sur and Don Bosco sites presented a diversity of six and four species, respectively (Table 1). In the three sampling sites, the distribution of individuals by zones showed interesting variations. In general, considering all the zones, Zone 2 recorded the highest abundance with a total of 124 individuals, followed by Zone 3 with 107 individuals, and Zone 1 with 88 individuals. However, this pattern was not consistent across all sites. Specifically, in Costa Sur, Zone 2 had the highest abundance of individuals, while in Don Bosco, it was Zone 1 that recorded the most individuals, and in El Embarcadero, Zone 3 presented the highest number, based on the data, a total of 10 species were recorded, with 5 species occurring exclusively in a single location —specifically, E. cognata was found only in Costa Sur, while E. allosticta, E. disimula, E. hemichlora, and E. tridentata were recorded solely in El Embarcadero— and the remaining 5 species (E. cybelia, E. dressleri, E. imperialis, E. nigrita, and E. smaragdina) were found in more than one location, indicating a balanced distribution between site-specific species and those with a broader occurrence. (Table 2).
Figure 4.
Mean species abundance (± SE) in two mangrove sampling sites in Panama: El Embarcadero (Bay of Panama) and El Líbano (Bay of Chame). Bars represent the mean number of individuals per sampling event. A generalized linear mixed model (GLMM) showed that El Líbano had significantly lower species abundance compared to El Embarcadero (coefficient = –0.9832, p < 0.001), indicating a ~62% reduction in species abundance.
Figure 4.
Mean species abundance (± SE) in two mangrove sampling sites in Panama: El Embarcadero (Bay of Panama) and El Líbano (Bay of Chame). Bars represent the mean number of individuals per sampling event. A generalized linear mixed model (GLMM) showed that El Líbano had significantly lower species abundance compared to El Embarcadero (coefficient = –0.9832, p < 0.001), indicating a ~62% reduction in species abundance.
Three genera were collected: Euglossa Latreille, 1802; Exaerete Hoffmannsegg, 1817; and Eulaema Lepeletier, Havva, and Victoria, 1841; Eulaema being the most abundant genus with 219 individuals, representing 68.6 % of the total sampling. The genus Euglossa accounted for a total of 83 individuals, and Exaerete for a total of 17 individuals. The genus with the highest species richness was Euglossa with seven species, while Eulaema and Exaerete each had one species (Table 2).
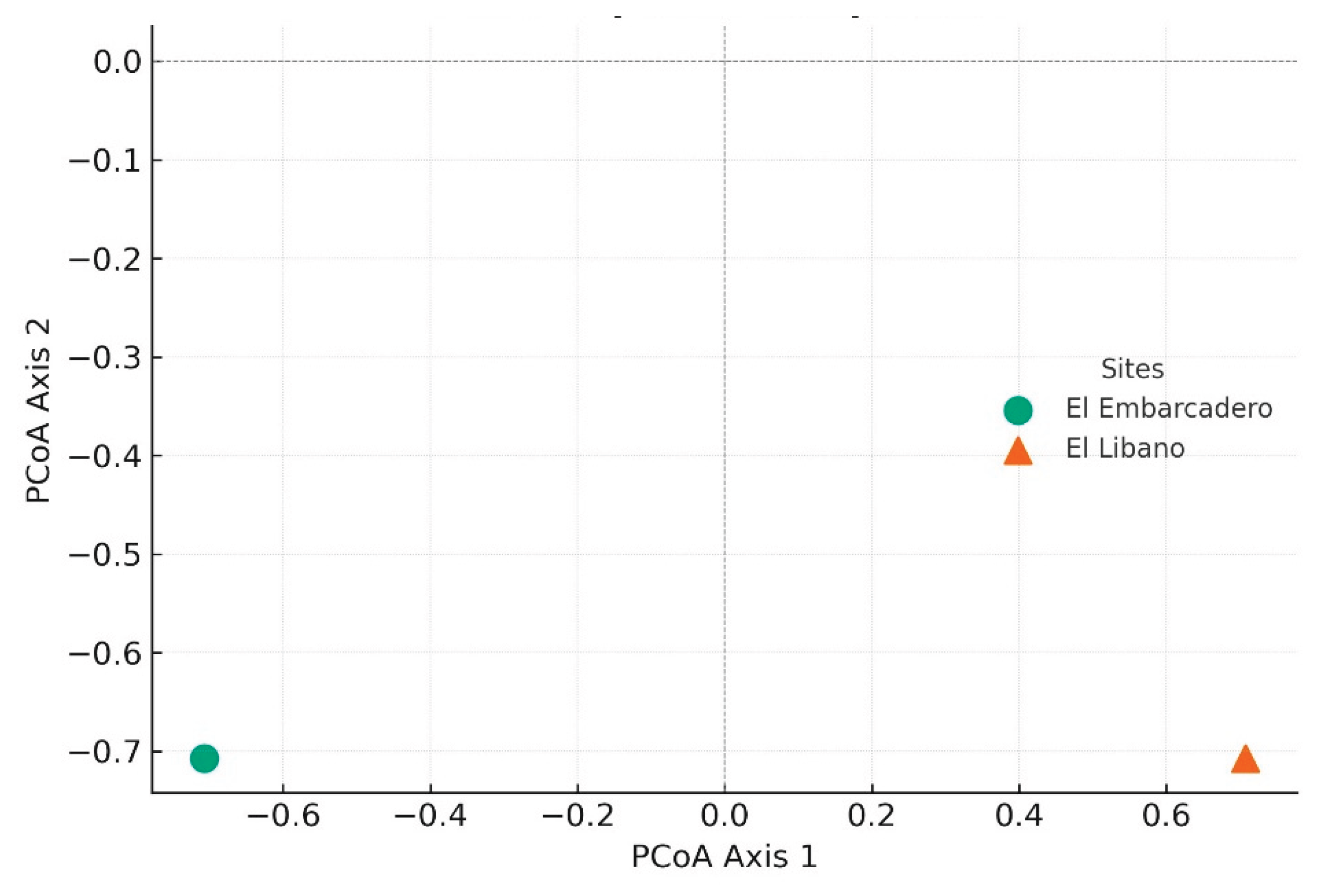
Comparatively, in the sampling sites of the Bay of Panama and the Bay of Chame during the same period, a general abundance of 108 individuals was observed. Specifically, 59 individuals were collected in El Líbano and 49 in El Embarcadero, Bay of Panama (Fig. 5). Of these, 75 belonged to the genus Euglossa, 32 to Eulaema, and one to Exaerete. The overall richness was 12 species distributed across three genera, with six species recorded in El Líbano and nine in El Embarcadero. Regarding genus distribution, nine species of Euglossa, two of Eulaema, and one of Exaerete were identified (Table 3). PCoA analysis was performed for comparison, where we can observe that El Embarcadero (circle) and El Líbano (cross) are separated in the bidimensional space, suggesting differences in species composition between both sites. However, the distance between the points is not extreme, indicating that these differences may not be significant.
Figure 5.
PCoA (Alternative PCA) Analysis of Species Composition: presents the PCoA analysis based on Jaccard distance to compare species composition between El Embarcadero (circle) and El Líbano (cross).
Figure 5.
PCoA (Alternative PCA) Analysis of Species Composition: presents the PCoA analysis based on Jaccard distance to compare species composition between El Embarcadero (circle) and El Líbano (cross).
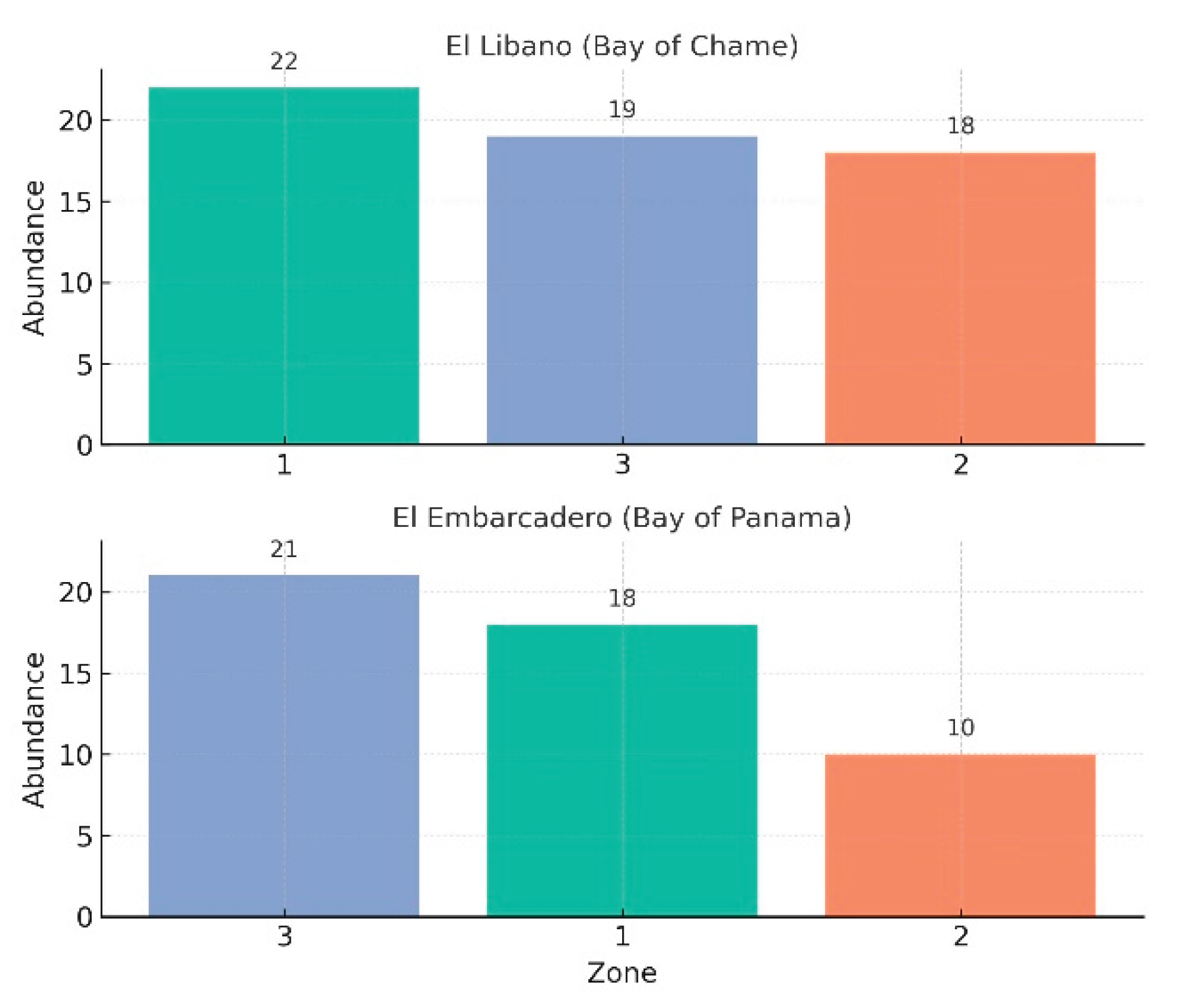
At the site of El Embarcadero, Bay of Panama, a total of nine species were collected: eight of Euglossa and one of Eulaema, with an abundance of 34 and 15 individuals, respectively. Regarding the sampling zone, the highest abundance was observed in Zone 3 with 21 individuals, followed by Zone 1 with 18, and Zone 2 with 10 individuals. In terms of species’ richness, Zone 3 had a total of seven species, while Zones 1 and 2 each had a richness of four species. For El Líbano, Bay of Chame, a total of six species were collected: three of Euglossa, one of Eulaema, and one of Exaerete, with an abundance of 41 individuals, 17 individuals, and one individual, respectively. Regarding the sampling zone, the highest abundance was recorded in Zone 1 with 22 individuals, followed by Zone 3 with 19, and finally, Zone 2 with 18 individuals (Fig. 6).
Figure 6.
Abundance per sampling zone in the mangrove forest of Bay of Panama and Bay of Chame.
Statistical analyses were performed to determine the alpha (α) and beta (β) diversity indices of the sampled zones (Table 4). The Shannon Diversity Index was found to be 1.014 for El Líbano and 1.863 for El Embarcadero. Additionally, the Bray-Curtis index was 0.7407, and the Jaccard index was 0.750 (Table 5).
4. Discussion
Despite six months of continuous sampling, only 427 Euglossini bees representing 14 species were collected across the mangrove sites, a markedly low abundance when compared to records from undisturbed tropical forests. This supports the hypothesis that anthropogenic pressures—such as urban expansion, deforestation, and habitat fragmentation—are having a negative impact on orchid bee communities in coastal ecosystems [18]. The highest richness and abundance were observed in El Embarcadero, particularly in the transition zone between mangrove and adjacent forest. In contrast, Don Bosco, the most urbanized site, exhibited the lowest diversity metrics. These patterns are consistent with previous research indicating that habitat degradation reduces the availability of floral resources and suitable nesting sites, thereby reshaping pollinator community composition [19].
The dominance of Eulaema nigrita, a generalist species known for its tolerance to disturbed environments, and the low Shannon and Simpson indices found in El Líbano (Bay of Chame) suggest a community shift toward disturbance-resilient taxa. Meanwhile, specialist species appear to be declining, likely due to the simplification of habitat structure and floral diversity. Although some variation in species composition was observed among sites, overall beta diversity was modest, indicating limited heterogeneity. These results suggest that the current environmental conditions may be insufficient to support diverse and stable orchid bee communities, potentially threatening long-term population viability [20,21].
A methodological limitation to consider is the exclusive use of eucalyptus oil as an attractant. While it is a widely used and effective lure for a broad range of Euglossini species, the lack of other scent options may have biased the sampling against species with more specialized chemical preferences. Future studies should incorporate multiple attractants to capture a more complete picture of orchid bee diversity [2,16]. These findings highlight the vulnerability of pollinator communities in urban mangrove ecosystems and the need for targeted conservation strategies. Multi-seasonal and multi-site monitoring is essential to better understand temporal trends and to guide biodiversity management in increasingly disturbed coastal habitats.
5. Conclusions
This study demonstrates that anthropogenic pressure is associated with reduced species richness and altered community composition of orchid bees in mangrove ecosystems, with a marked dominance of disturbance-tolerant species such as Eulaema nigrita. These changes may affect pollination dynamics and compromise the overall ecosystem functioning in tropical coastal habitats. Conservation strategies should prioritize habitat protection and restoration, as well as the enhancement of floral resources within mangrove areas. Future studies should incorporate multi-attractant sampling, seasonal monitoring, and broader landscape comparisons to improve our understanding of the resilience of Euglossini communities under increasing environmental stress.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, JAL, JR and ASM; methodology, JAL, JR and ASM; validation, JAL, JR, AGT and ASM; formal analysis, JAL and AGT; investigation, JAL, JR and ASM; resources, ASM; data curation, JAL; writing—original draft preparation, JAL; writing—review and editing, JAL, and AGT; visualization, JAL and AGT; supervision, ASM; project administration, JAL and ASM.; funding acquisition, ASM. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Cooperation Agreement No. 084-2021, SENACYT / City of Knowledge Foundation (FCDS), Climate Change Sentinel Mangroves Project: Monitoring, vulnerability and resilience of marine-coastal ecosystems in the Bay of Panama and Chame Bay.
Data Availability Statement
the preprocessed data that supports the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We would like to thank the Smithsonian Tropical Research Institute, Panama and the project management “Impact of Insects Herbivory on Mangroves in the Ramsar Wetland Wildlife Protected Area, Juan Diaz, Bay of Panama”. The National Secretariat of Science and Technology (SENCYT) and the City of Knowledge Foundation, with the project “Mangroves sentinels of climate change: monitoring, vulnerability and resilience of marine-coastal ecosystems in the Bay of Panama and Chame Bay”.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hanson, P. E., & Gauld, I. D. Hymenoptera de la región neotropical. Memoirs of the American Entomological Institute 2006, 77, 1–994.
- Roubik, W. D., & Hanson, P. E. Abejas de orquídeas de la América tropical: Biología y guía de campo. 2004. Instituto Nacional de Biodiversidad (INBio).
- Nemésio, A., & Silveira, F. A. Diversity and distribution of orchid bees (Hymenoptera: Apidae) with a revised checklist of species. Neotropical Entomology 2007, 36(6), 874–888. [CrossRef]
- Engel, M. S. The first fossil Euglossa and phylogeny of orchid bees (Hymenoptera, Apidae, Euglossini). American Museum Novitates 1999, 3272, 1–14.
- Hinojosa-Díaz, I. A., & Engel, M. S. A checklist of the orchid bees of Nicaragua (Hymenoptera: Apidae: Euglossini). Journal of the Kansas Entomological Society 2012, 85(2), 135–144. [CrossRef]
- Reyes-Ledezma, K. Y., Santos-Murgas, A., González, P., Gómez, I. Y., & Barrios-Vargas, A. Diversidad alpha y beta de abejas Euglossini (Hymenoptera: Apidae) en el dosel y sotobosque del Cerro Turega, provincia de Coclé, Panamá. Tecnociencia 2020, 22(2), 205–225. [CrossRef]
- Reyes-Novelo, E., Meléndez R. V., Delfín, G. H., & Ayala, R. Abejas silvestres como bioindicadores en el Neotrópico. Agroecosistemas Tropicales y Subtropicales 2009, 10(1), 1–13.
- Cane, J. H. Habitat fragmentation and native bees: A premature verdict? Conservation Ecology 2001, 5(1), 3. http://www.consecol.org/vol5/iss1/art3/.
- Ramos, M., Pérez, J., & Torres, L. Impacto del desarrollo urbano en la calidad del agua de la Bahía de Panamá. Revista Panameña de Ciencias Ambientales 2020, 15(2), 45–60.
- Guzmán, H., & Vega, A. Evaluación de ecosistemas costeros en Panamá: Manglares y áreas protegidas. 2019. Smithsonian Tropical Research Institute.
- Brosi, B. J. The effects of forest fragmentation on Euglossine bee communities (Hymenoptera: Apidae: Euglossini). Biological Conservation 2009, 142(2), 414–423. [CrossRef]
- Sánchez-Bayo, F., & Wyckhuys, K. A. Worldwide decline of the entomofauna: A review of its drivers. Biological Conservation 2019, 232, 8–27. [CrossRef]
- Somejima, H., Markuzi, M., Nagamitsu, T., & Nakasizuka, T. The effects of human disturbance on a stingless bee community in a tropical rainforest. Biological Conservation 2004, 120, 577–587. [CrossRef]
- Autoridad del Canal de Panamá. Estado ecológico y biodiversidad de la cuenca del Canal de Panamá. 2022. Autoridad del Canal de Panamá.
- Ramsar. Ficha informativa sobre los Humedales de Importancia Internacional: Bahía de Panamá. 2021. Convención Ramsar.
- Pérez-Buitrago, N., Mojica-Candela, L. J., & Agudelo-Martínez, J. C. Variación estacional de abejas euglosinas (Apidae: Euglossini) en el norte de la Orinoquia colombiana. Revista De La Academia Colombiana De Ciencias Exactas, Físicas Y Naturales 2022, 46(179), 470-481. [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria; 2023.
- Rojas, B., Vásquez, O., Santos-Murgas, A., Cobos, R., & Gómez-Robles, I. Y. Abejas de las orquídeas como bioindicadores del estado de conservación de un bosque. Manglar 2022, 19(3), 271–277. [CrossRef]
- Muñoz-Urias, A., Moya-Raygoza, G., & Vásquez-Bolaños, M. Effects of urbanization and floral diversity on the bee community in a tropical dry forest. Journal of Hymenoptera Research 2025, 98, 47–68. [CrossRef]
- Giannini, T. C., Cordeiro, G. D., Freitas, B. M., Saraiva, A. M., & Imperatriz-Fonseca, V. L. The dependence of crops for pollinators and the economic value of pollination in Brazil. Journal of Economic Entomology 2017, 108(3), 849–857. [CrossRef]
- Jaffé, R., Pope, N., Carvalho, A. T., Maia, U. M., Blochtein, B., de Carvalho, C. A. L., & Imperatriz-Fonseca, V. L. Landscape genomics to the rescue of a tropical bee threatened by habitat loss and climate change. Evolutionary Applications 2019, 12(6), 1164–1177. [CrossRef]

Table 1.
Orchid bee richness and abundance per sampling site (El Embarcadero, Costa Sur and Don Bosco).
Table 1.
Orchid bee richness and abundance per sampling site (El Embarcadero, Costa Sur and Don Bosco).
| Costa Sur | Don Bosco | El Embarcadero | |
| Richness | 6 | 4 | 9 |
| Abundance | 70 | 23 | 226 |
Table 2.
Orchid bee (Euglosiini) diversity and abundance per sampling site.
| Site/Species | Zones | Total/ Site | ||
| 1 | 2 | 3 | ||
| Costa Sur | 10 | 43 | 17 | 70 |
| E. cognata | 4 | 4 | ||
| E. cybelia | 1 | 9 | 10 | |
| E. dressleri | 2 | 3 | 5 | |
| E. imperialis | 6 | 6 | ||
| E. nigrita | 27 | 16 | 43 | |
| E. smaragdina | 1 | 1 | 2 | |
| Don Bosco | 15 | 3 | 5 | 23 |
| E. cybelia | 1 | 1 | ||
| E. dressleri | 1 | 1 | ||
| E. nigrita | 14 | 3 | 3 | 20 |
| E. smaragdina | 1 | 1 | ||
| El Embarcadero | 63 | 78 | 85 | 226 |
| E. allosticta | 1 | 2 | 3 | |
| E. cybelia | 1 | 1 | ||
| E. disimula | 1 | 1 | 2 | |
| E. dressleri | 1 | 7 | 8 | |
| E. hemichlora | 5 | 5 | 10 | |
| E. imperialis | 11 | 19 | 30 | |
| E. nigrita | 57 | 49 | 50 | 156 |
| E. smaragdina | 4 | 10 | 14 | |
| E. tridentata | 2 | 2 | ||
| Total | 88 | 124 | 107 | 319 |
Table 3.
Orchid bee richness for Bay of Chame and Bay of Panama.
| Orchid bee species | El Embarcadero (Bay of Panama) |
El Líbano (Bay of Chame) |
|---|---|---|
| Euglossa allosticta Moure, 1969 | x | - |
| Euglossa deceptrix Moure, 1968 | x | x |
| Euglossa imperialis Cockerell, 1922 | - | x |
| Euglossa disimula Dressler, 1978 | x | - |
| Euglossa dodsoni Moure, 1965 | x | - |
| Euglossa hemichlora Cockerell, 1917 | x | - |
| Euglossa heterosticta Moure, 1968 | x | - |
| Euglossa tridentata Moure, 1970 | x | - |
| Euglossa variabilis Friese, 1899 | x | x |
| Eulaema nigrita Lepeletier, 1841 | x | x |
| Eulaema meriana (Olivier, 1789) | - | x |
| Exaerete smaragdina (Guérin-Méneville, 1845) | - | x |
| x: present, -: absent. |
Table 4.
Alpha diversity indices (α) of El Líbano, Bay of Chame, and El Embarcadero, Bay of Panama.
| Alpha diversity indices | El Líbano (Bay of Chame) | El Embarcadero (Bay of Panamá) |
| Richness | 6 | 9 |
| Abundance | 59 | 49 |
| Shannon | 1.014 | 1.863 |
| Gini- Simpson | 0.5136 | 0.8038 |
| Inverse Simpson Index | 2.056 | 5.098 |
| Fisher- Alpha | 1.670 | 3.236 |
| Berger- Parker | 0.6610 | 0.3061 |
| Pielou's Evenness | 0.5660 | 0.8478 |
| E-evenness | 0.4595 | 0.7157 |
Table 5.
Beta diversity indices (β) of El Líbano, Bay of Chame, and El Embarcadero, Bay of Panama.
| Sampling sites | El Líbano (Bay of Chame) | Beta diversity indices |
| El Embarcadero (Bay of Panamá) | 0.7407 | Bray- Curtis |
| El Embarcadero (Bay of Panamá) | 0.75 | Jaccard |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



