
Submitted:
04 December 2024
Posted:
05 December 2024
You are already at the latest version
Abstract
Previous research has consistently shown that high-fat diet (HFD) consumption can lead to the development of colonic inflammation. Neohesperidin (NHP), a naturally occurring flavanone glycoside in citrus fruits, has anti-inflammatory properties. However, the efficacy and mechanism of NHP in countering prolonged HFD-induced inflammation remains unclear. In this study, rats on HFD were intragastric administered (i.g.) with NHP for 12 consecutive weeks. Results indicated that this natural compound is effective in reducing colorectal inflammation at doses of 40-80 mg/kg body weight by i.g. administration, with significant decreases in inflammation markers such as TNF-α and IL-1β levels. It also improved intestinal mucosal tissue integrity and reduced HFD-stimulated colorectal inflammation via the JAK2/STAT3 pathway. Furthermore, intestinal microbiota sequencing results showed that NHP intervention significantly downregulated the Firmicutes/Bacteroidetes ratio. This ratio is closely related to the preventive role in the context of glycolipid metabolism disorder. Compared with fecal cultures of rats from the HFD group, after 48 h in vitro fermentation, those from the NHP group had distinct microbiota composition and notably higher concentrations of SCFAs. Collectively, these observations suggest that NHP's biological activities in downregulating HFD-induced colorectal inflammation are achieved by regulating intestinal flora and promoting SCFAs formation.
Keywords:
1. Introduction
2. Results and Discussion
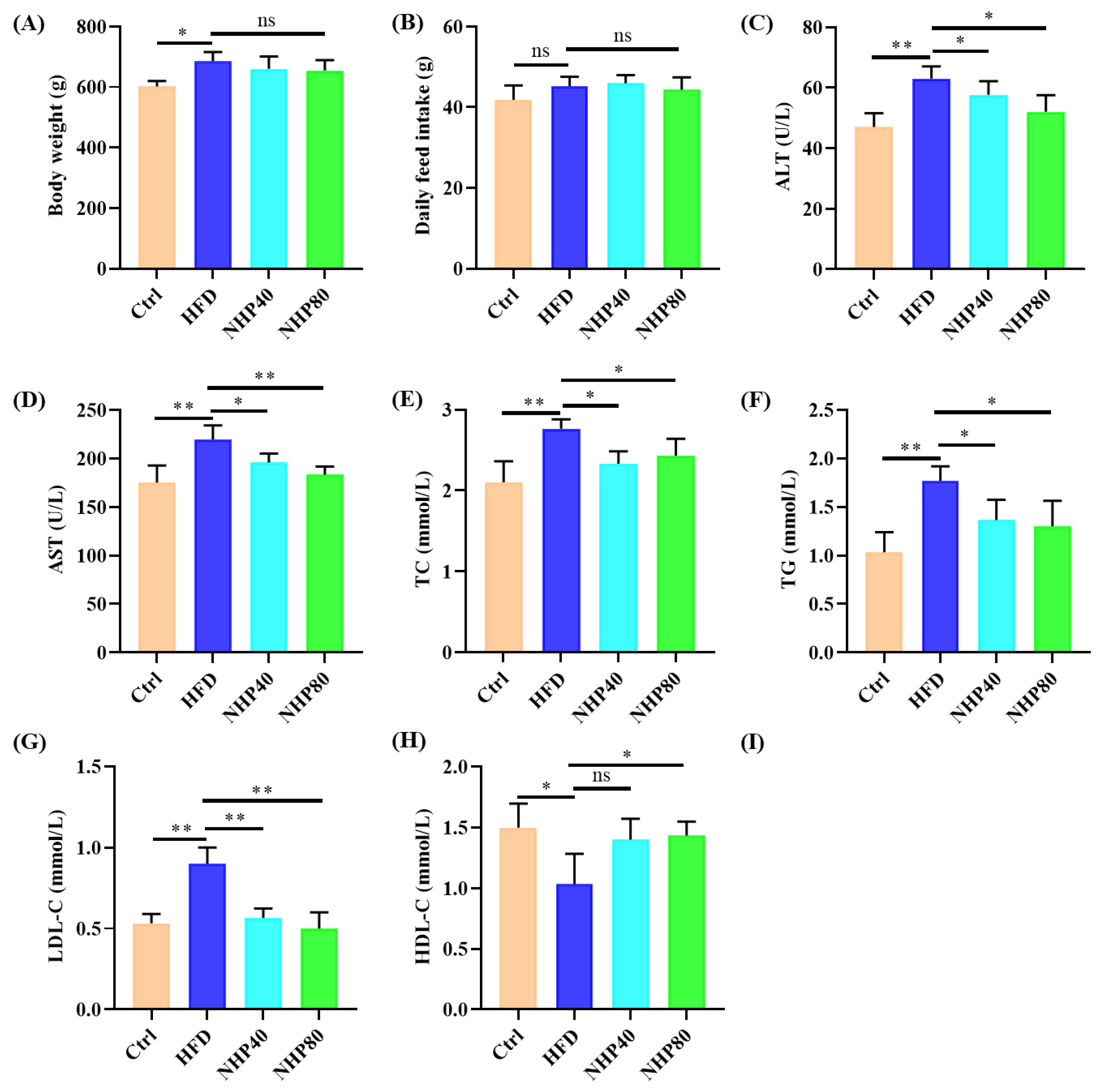
2.1. NHP Improved Lipid Metabolism Disorders in HFD Rats
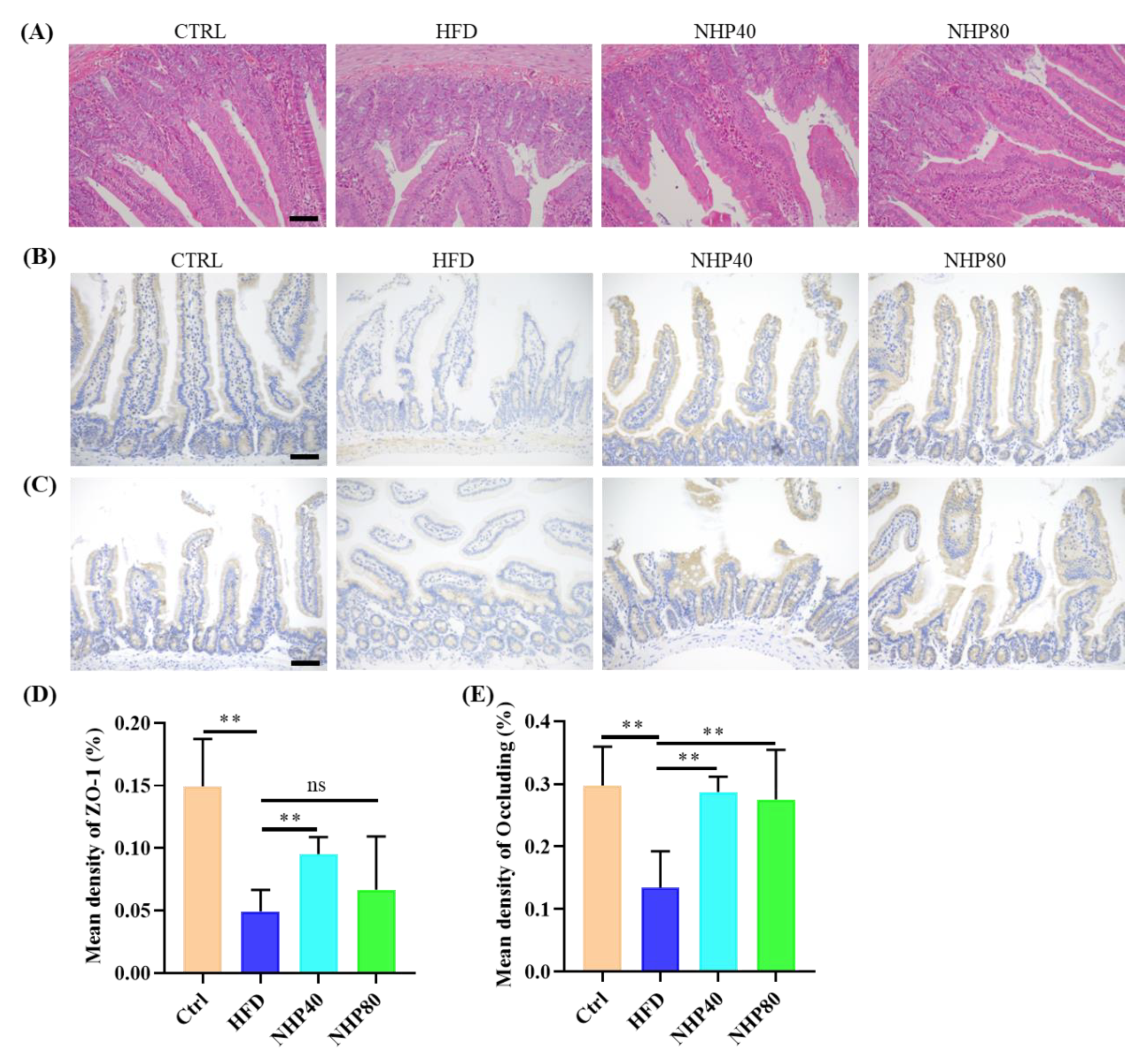
2.2. NHP Improved Pathological Ileum Injury
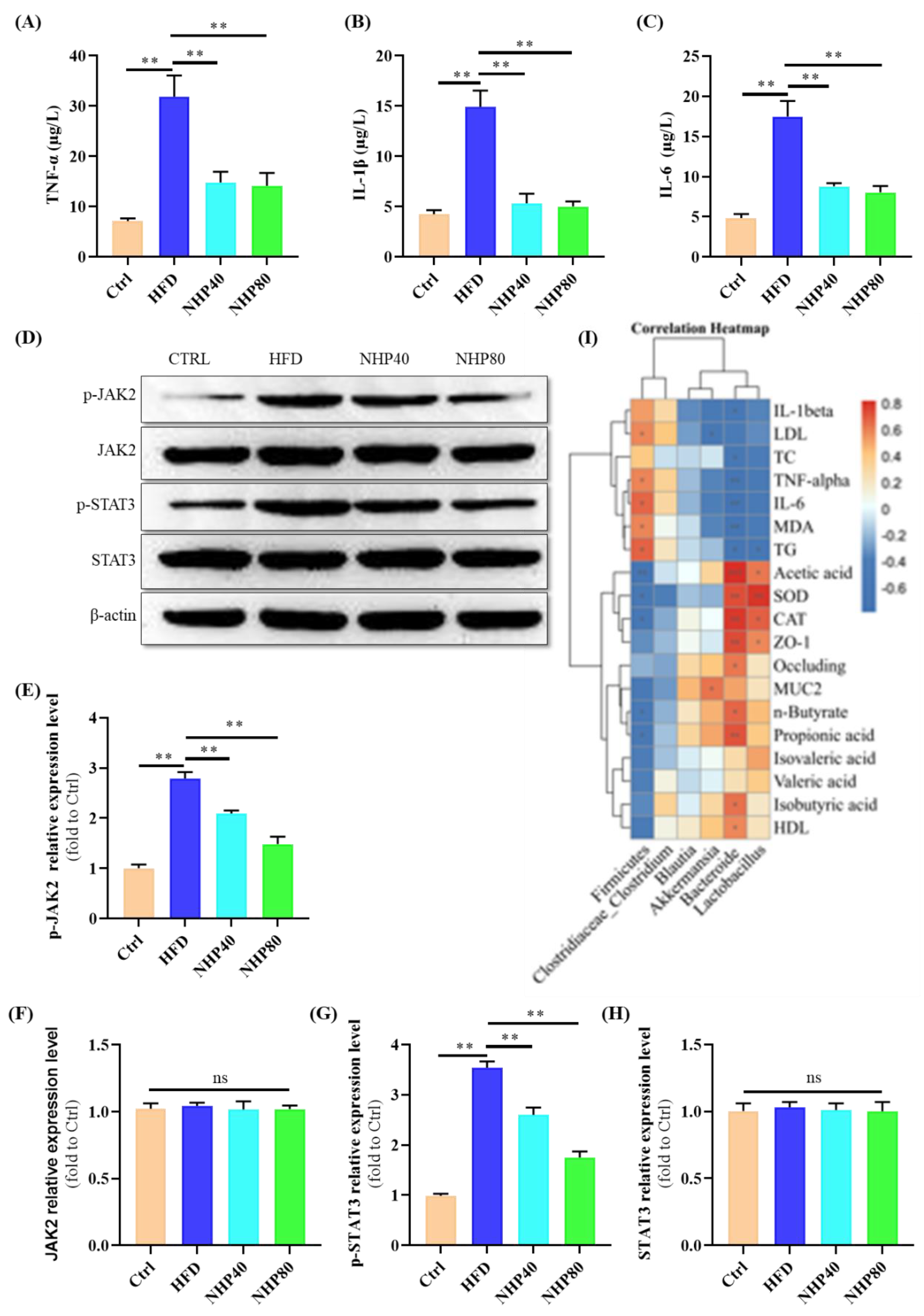
2.3. NHP Mitigated HFD-Induced Inflammation
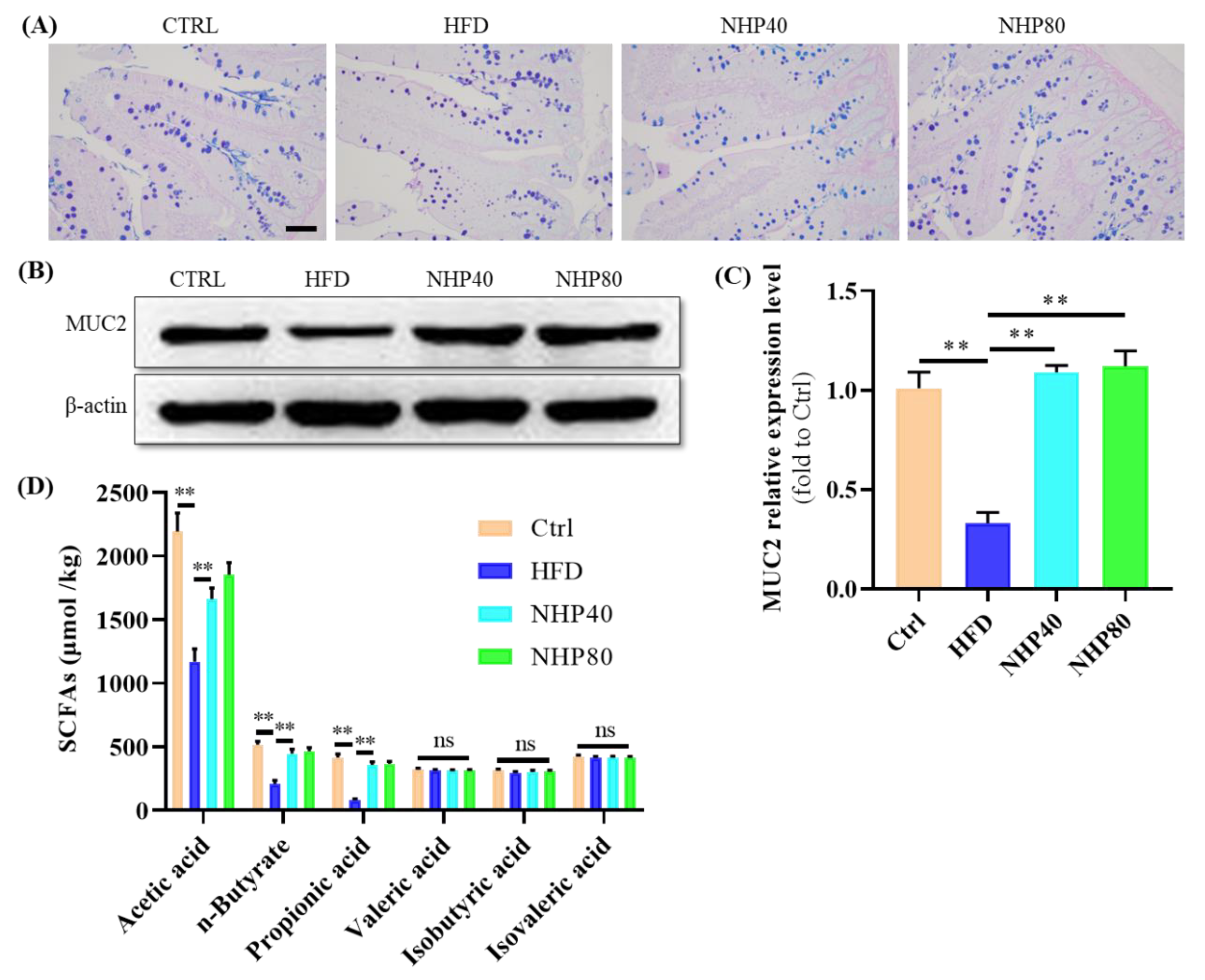
2.4. NHP Maintained the Integrity of Colonic Mucosa
2.5. NHP Elevated the Level of SCFAs
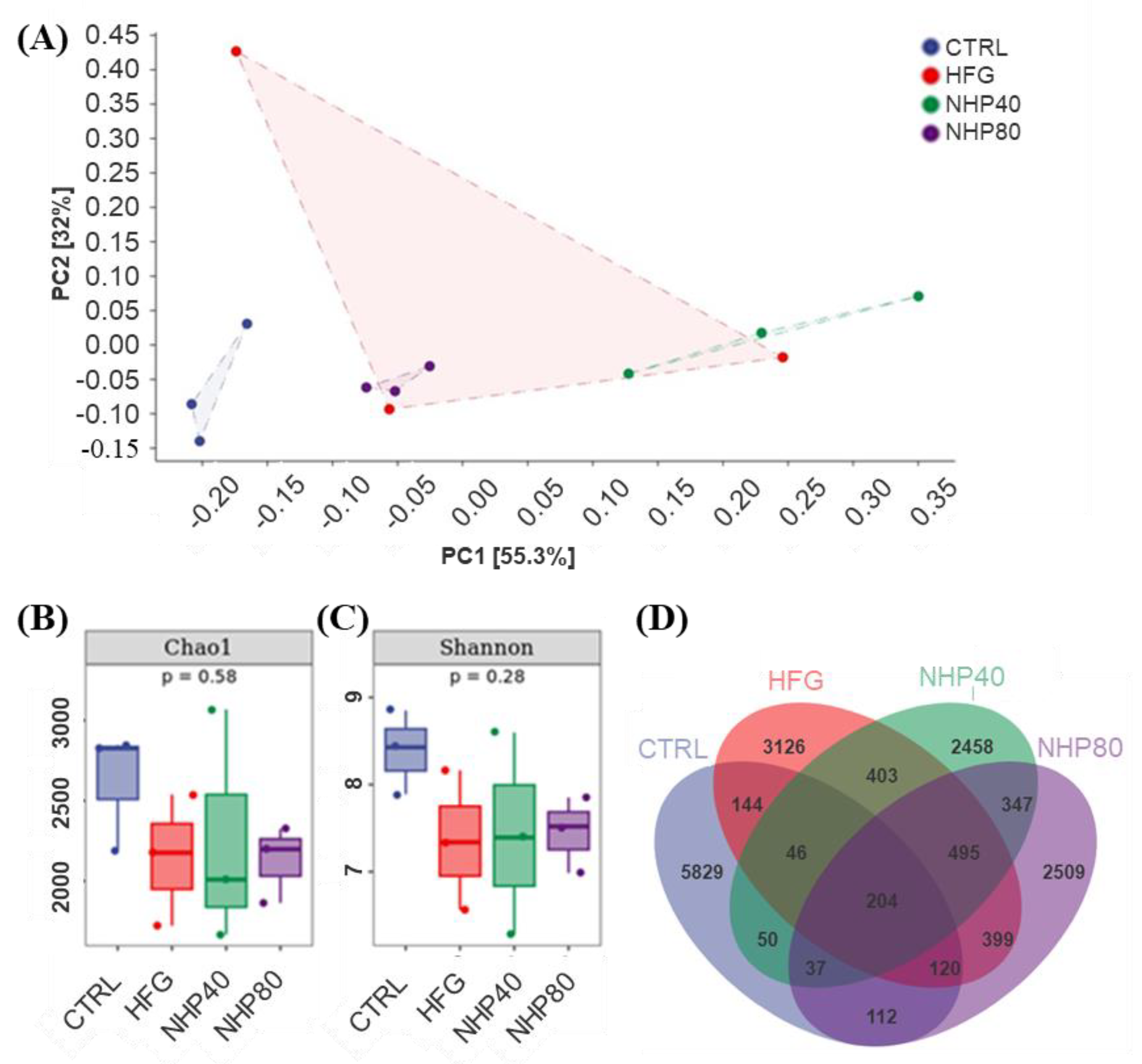
2.6. NHP Regulated Intestinal Flora Diversity
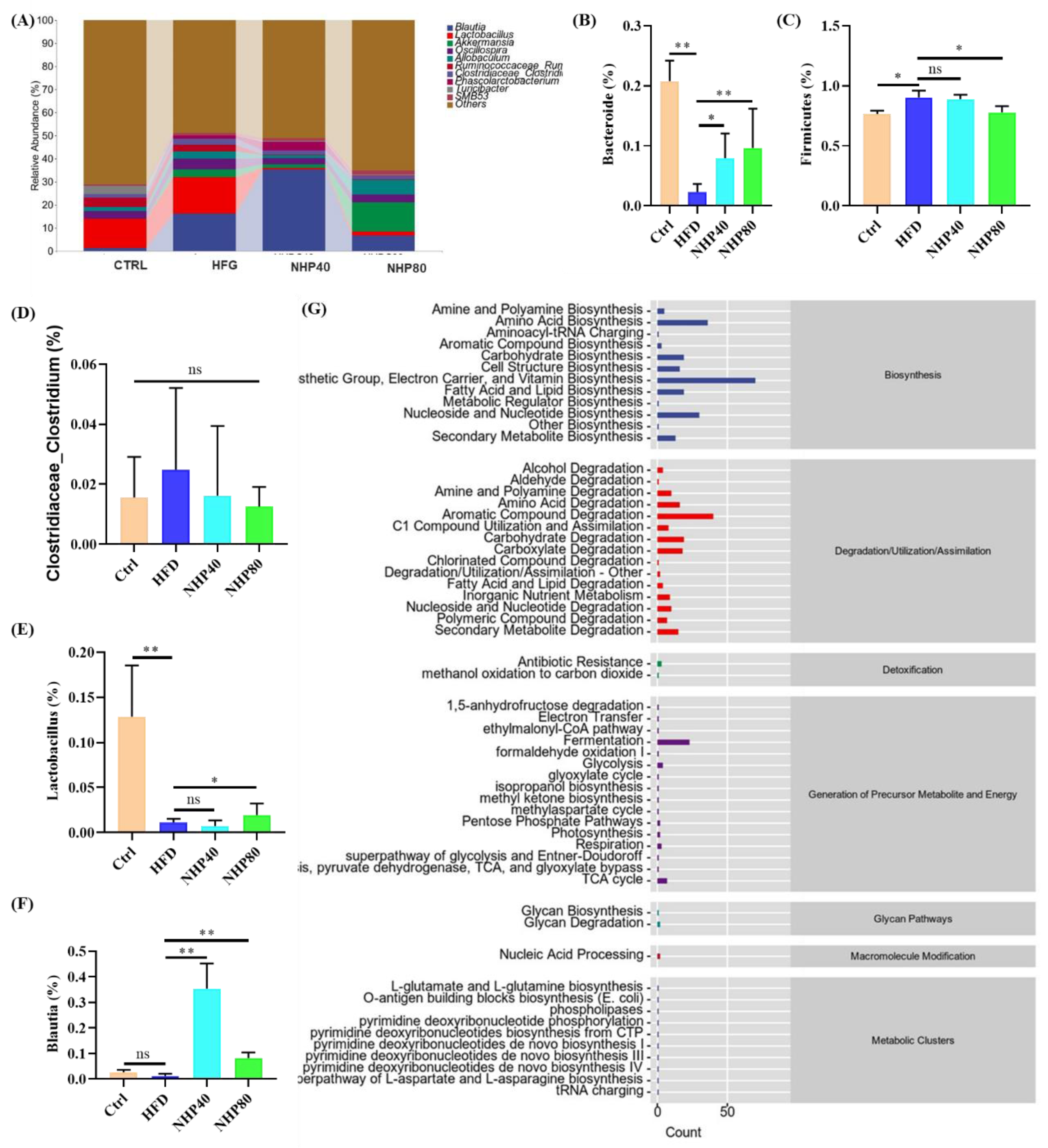
2.7. NHP Modulated the Overall Structure and Composition of Gut Microbiota
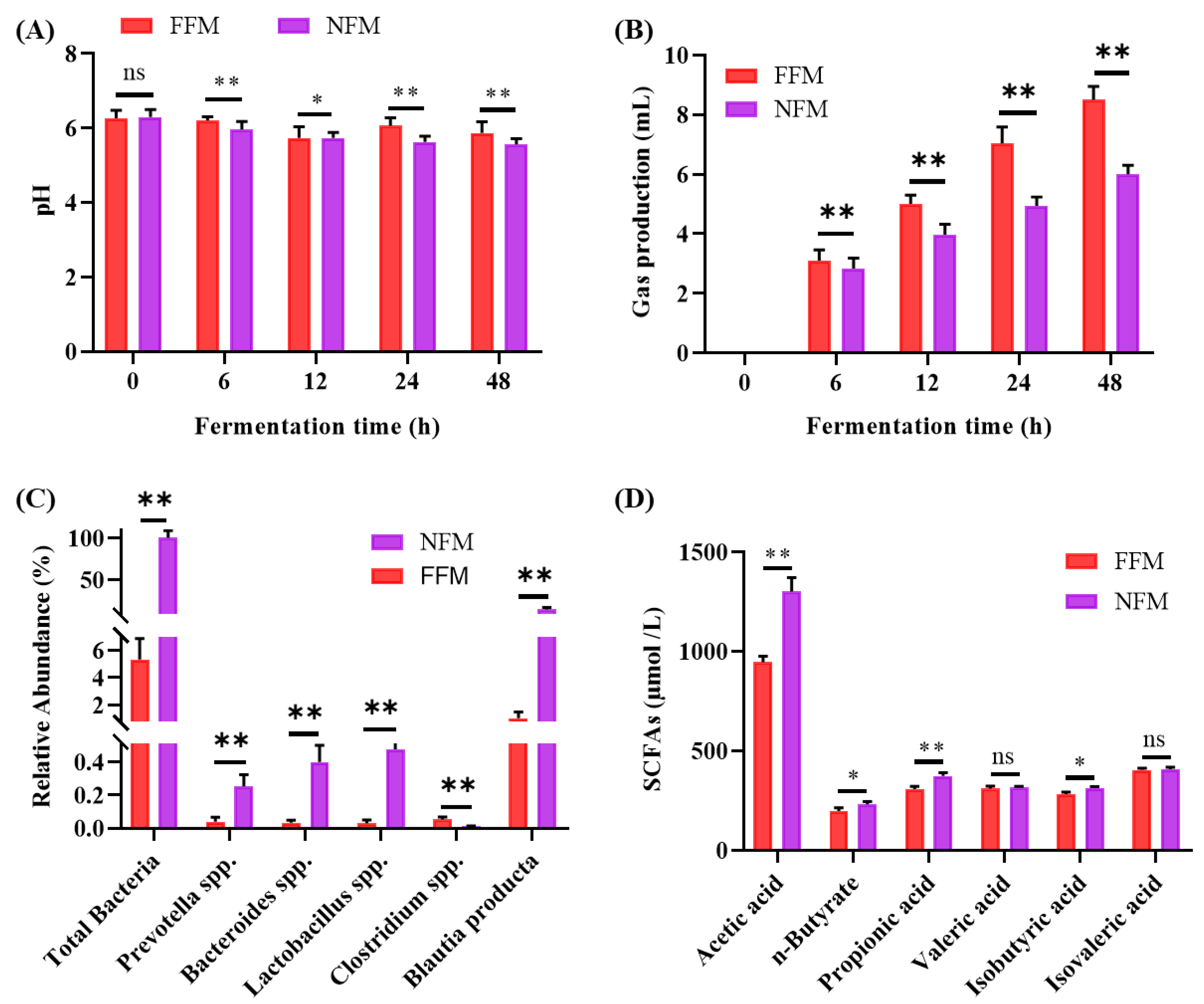
2.8. In vitro Fermentation Results
2.8.1. Fermentation pH Fluctuation
2.8.2. Fermentation Gas Production
2.8.3. Microbiota Composition and SCFAs Production
3. Materials and Methods
3.1. Experimental Materials
3.2. Experimental Animals and Treatments
3.3. Colorectal Histopathological Examination
3.4. Immunohistochemical Detection
3.5. Cytokine Analysis
3.6. Intestinal Microbiota Sequencing Procedure
3.7. Fecal Microbiota In vitro Fermentation
3.8. Measurement of pH and Gas Production
3.9. Determination of SCFAs Content
3.10. RT-qPCR Detection of Specific Bacterial Groups
3.11. Western Blot Analysis
3.12. Statistical Methods
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Ren, Q.; Sun, Q.; Fu, J. Dysfunction of autophagy in high-fat diet-induced non-alcoholic fatty liver disease. Autophagy 2024, 20, 221–241. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Gao, X.; Hong, Y.; Zhong, J.; Li, Y.; Zhu, W.; Ma, J.; Huang, W.; Li, Y.; Li, Y.; et al. High Fat Diet and High Sucrose Intake Divergently Induce Dysregulation of Glucose Homeostasis through Distinct Gut Microbiota-Derived Bile Acid Metabolism in Mice. Journal of agricultural and food chemistry 2024, 72, 230–244. [Google Scholar] [CrossRef] [PubMed]
- Hamamah, S.; Iatcu, O.C.; Covasa, M. Nutrition at the Intersection between Gut Microbiota Eubiosis and Effective Management of Type 2 Diabetes. Nutrients 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhao, Y.; Wang, X.; Kong, L.; Johnston, L.J.; Lu, L.; Ma, X. Dietary nutrients shape gut microbes and intestinal mucosa via epigenetic modifications. Critical reviews in food science and nutrition 2022, 62, 783–797. [Google Scholar] [CrossRef]
- Yang, Q.; Cai, X.; Zhu, Y.; Hu, Z.; Wei, Y.; Dang, Q.; Zhang, Y.; Zhao, X.; Jiang, X.; Yu, H. Oat β-glucan supplementation pre- and during pregnancy alleviates fetal intestinal immunity development damaged by gestational diabetes in rats. Food & function 2023, 14, 8453–8466. [Google Scholar] [CrossRef]
- Yan, W.; Luo, J.; Yu, Z.; Xu, B. A critical review on intestinal mucosal barrier protection effects of dietary polysaccharides. Food & function 2024, 15, 481–492. [Google Scholar] [CrossRef]
- Gustafsson, J.K.; Johansson, M.E.V. The role of goblet cells and mucus in intestinal homeostasis. Nature reviews. Gastroenterology & hepatology 2022, 19, 785–803. [Google Scholar] [CrossRef]
- Rana, N.; Privitera, G.; Kondolf, H.C.; Bulek, K.; Lechuga, S.; De Salvo, C.; Corridoni, D.; Antanaviciute, A.; Maywald, R.L.; Hurtado, A.M.; et al. GSDMB is increased in IBD and regulates epithelial restitution/repair independent of pyroptosis. Cell 2022, 185, 283–298.e217. [Google Scholar] [CrossRef]
- Dolinger, M.; Torres, J.; Vermeire, S. Crohn's disease. Lancet (London, England) 2024, 403, 1177–1191. [Google Scholar] [CrossRef]
- Fan, B.; Zhou, J.; Zhao, Y.; Zhu, X.; Zhu, M.; Peng, Q.; Li, J.; Chang, X.; Shi, D.; Yin, J.; et al. Identification of Cell Types and Transcriptome Landscapes of Porcine Epidemic Diarrhea Virus-Infected Porcine Small Intestine Using Single-Cell RNA Sequencing. Journal of immunology (Baltimore, Md. : 1950) 2023, 210, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.P.; Lai, M.D.; Lee, J.C.; Yen, M.C.; Weng, T.Y.; Chen, W.C.; Fang, J.H.; Chen, Y.L. Mucin 2 silencing promotes colon cancer metastasis through interleukin-6 signaling. Sci Rep 2017, 7, 5823. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Dai, W.; Dong, M.; Dai, C.; Wu, S. MUC2 and related bacterial factors: Therapeutic targets for ulcerative colitis. EBioMedicine 2021, 74, 103751. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Wu, S.; Li, L.; Hao, F.; Wu, J.; Liao, Z.; Wang, J.; Zhong, R.; Wei, H.; Fang, X. Alleviation of lipid metabolic dysfunction through regulation of intestinal bacteriophages and bacteria by green tea polyphenols in Ob/Ob mice. Food Chem 2024, 456, 139988. [Google Scholar] [CrossRef]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A.J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; et al. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat Commun 2020, 11, 4457. [Google Scholar] [CrossRef]
- Wang, M.; Zhao, H.; Wen, X.; Ho, C.T.; Li, S. Citrus flavonoids and the intestinal barrier: Interactions and effects. Compr Rev Food Sci Food Saf 2021, 20, 225–251. [Google Scholar] [CrossRef]
- Dai, H.; Jiang, Y.; Liu, S.; Li, D.; Zhang, X. Dietary flavonoids modulate the gut microbiota: A new perspective on improving autism spectrum disorder through the gut-brain axis. Food Research International 2024, 186, 114404. [Google Scholar] [CrossRef]
- Ortiz, A.C.; Fideles, S.O.M.; Reis, C.H.B.; Bellini, M.Z.; Pereira, E.; Pilon, J.P.G.; de Marchi, M.; Detregiachi, C.R.P.; Flato, U.A.P.; Trazzi, B.F.M.; et al. Therapeutic Effects of Citrus Flavonoids Neohesperidin, Hesperidin and Its Aglycone, Hesperetin on Bone Health. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- Li, J.; Rui, X.; Xu, L.; Liu, Y.; Yang, Y.; Yin, D. Enhanced therapeutic effect on colitis with powder formulations of Painong San associated with the promotion of intestinal adhesion and absorption. Journal of ethnopharmacology 2022, 289, 115030. [Google Scholar] [CrossRef]
- Chen, S.Y.; Zhou, Q.Y.; Chen, L.; Liao, X.; Li, R.; Xie, T. The Aurantii Fructus Immaturus flavonoid extract alleviates inflammation and modulate gut microbiota in DSS-induced colitis mice. Frontiers in nutrition 2022, 9, 1013899. [Google Scholar] [CrossRef]
- Kirschner, N.; Poetzl, C.; von den Driesch, P.; Wladykowski, E.; Moll, I.; Behne, M.J.; Brandner, J.M. Alteration of tight junction proteins is an early event in psoriasis: putative involvement of proinflammatory cytokines. Am J Pathol 2009, 175, 1095–1106. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Gómez, O.; García-Rodríguez, R.V.; Pérez-Gutierrez, S.; Rivero-Ramírez, N.L.; García-Martínez, Y.; Pablo-Pérez, S.S.; Pérez-Pastén-Borja, R.; Cristóbal-Luna, J.M.; Chamorro-Cevallos, G. Protective effect of the phycobiliproteins from arthrospira maxima on indomethacin-induced gastric ulcer in a rat model. Plants 2023, 12, 1586. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Zhao, Y.; Qian, L.; Jiang, S.; Tang, Y.; Han, T. DHA-enriched phosphatidylserine alleviates high fat diet-induced jejunum injury in mice by modulating gut microbiota. Food & function 2023, 14, 1415–1429. [Google Scholar] [CrossRef]
- Yang, J.; Martínez, I.; Walter, J.; Keshavarzian, A.; Rose, D.J. In vitro characterization of the impact of selected dietary fibers on fecal microbiota composition and short chain fatty acid production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef]
- Zhai, Z.; Yang, Y.; Chen, S.; Wu, Z. Long-Term Exposure to Polystyrene Microspheres and High-Fat Diet-Induced Obesity in Mice: Evaluating a Role for Microbiota Dysbiosis. Environ Health Perspect 2024, 132, 97002. [Google Scholar] [CrossRef]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate Improves Insulin Sensitivity and Increases Energy Expenditure in Mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- Al-Lahham, S.; Roelofsen, H.; Rezaee, F.; Weening, D.; Hoek, A.; Vonk, R.; Venema, K. Propionic acid affects immune status and metabolism in adipose tissue from overweight subjects. Eur J Clin Invest 2012, 42, 357–364. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Lau, H.C.; Zhang, X.; Ji, F.; Lin, Y.; Liang, W.; Li, Q.; Chen, D.; Fong, W.; Kang, X.; Liu, W.; et al. Lactobacillus acidophilus suppresses non-alcoholic fatty liver disease-associated hepatocellular carcinoma through producing valeric acid. EBioMedicine 2024, 100, 104952. [Google Scholar] [CrossRef]
- Hosomi, K.; Saito, M.; Park, J.; Murakami, H.; Shibata, N.; Ando, M.; Nagatake, T.; Konishi, K.; Ohno, H.; Tanisawa, K.; et al. Oral administration of Blautia wexlerae ameliorates obesity and type 2 diabetes via metabolic remodeling of the gut microbiota. Nature communications 2022, 13, 4477. [Google Scholar] [CrossRef]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia-a new functional genus with potential probiotic properties? Gut microbes 2021, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb Biotechnol 2019, 12, 1109–1125. [Google Scholar] [CrossRef] [PubMed]
- Gullón, B.; Gullón, P.; Tavaria, F.; Pintado, M.; Gomes, A.M.; Alonso, J.L.; Parajó, J.C. Structural features and assessment of prebiotic activity of refined arabinoxylooligosaccharides from wheat bran. Journal of Functional Foods 2014, 6, 438–449. [Google Scholar] [CrossRef]
- Kaur, A.; Rose, D.J.; Rumpagaporn, P.; Patterson, J.A.; Hamaker, B.R. In vitro batch fecal fermentation comparison of gas and short-chain fatty acid production using "slowly fermentable" dietary fibers. J Food Sci 2011, 76, H137–H142. [Google Scholar] [CrossRef]
- Li, Z.; Liang, H.; Hu, Y.; Lu, L.; Zheng, C.; Fan, Y.; Wu, B.; Zou, T.; Luo, X.; Zhang, X.; et al. Gut bacterial profiles in Parkinson's disease: A systematic review. CNS Neurosci Ther 2023, 29, 140–157. [Google Scholar] [CrossRef]
- Li, S.; Liu, W.; Li, Y.; Che, X.; Xiao, P.; Liu, S.; Ma, Y.; Ren, D.; Wu, L.; Wang, Q.; et al. Extraction, purification, structural characterization and anti-hyperlipidemia activity of fucoidan from Laminaria digitata. International journal of biological macromolecules 2024, 279, 135223. [Google Scholar] [CrossRef]
- Küçükgöz, K.; Venema, K.; Trząskowska, M. Gut microbiota modulatory capacity of fermented ketchup in a validated in vitro model of the colon. Food research international (Ottawa, Ont.) 2024, 192, 114801. [Google Scholar] [CrossRef]
- Bianchi, F.; Dall’Asta, M.; Del Rio, D.; Mangia, A.; Musci, M.; Scazzina, F. Development of a headspace solid-phase microextraction gas chromatography–mass spectrometric method for the determination of short-chain fatty acids from intestinal fermentation. Food Chemistry 2011, 129, 200–205. [Google Scholar] [CrossRef]

| Forward (5'-3') | Reverse (5'-3') | |
|---|---|---|
| Total bacteria | TCCTACGGGAGGCAGCAGTGE | TTACCGCGGCTGCTGGCACG |
| Prevotella | CGGTGAATACGTTCYCGG | GGWTACCTTGTTACGACTT |
| Bacteroides | GGTGTCGGCTTAAGTGCCAT | GCATTYCACCGCTACACATG |
| Lactobacillus | GCAGCAGTAGGGAATCTTCCA | GCATTYCACCGCTACACATG |
| Clostridium | AGAGTTTGATCCTGGCTCAG | ACGGCTACCTTGTTACGACTT |
| Blautia producta | AGCTGACGACCTGATCGAGT | TCTCGAGCTGGTACGCTTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).



