
Submitted:
03 December 2024
Posted:
04 December 2024
You are already at the latest version
Abstract
There has been increasing interest in the study of microorganisms associated with tropical plant species, particularly fungi, due to their potential applications in biological control and the pro-duction of pharmacologically active compounds. This study aimed to identify and characterize the fungal community associated with cacao (Theobroma cacao L.) fruits in three cantons of the Orellana province in the Ecuadorian Amazon. Fungi were isolated directly from cacao fruits and subjected to cultural, morphological, and molecular characterization. Fungal diversity was assessed using relative abundance and species richness metrics. A total of 464 fungal isolates were obtained, representing 56 morphotypes and 14 genera, all belonging to the phylum Ascomycota. Penicillium sp. (27.8%), Epicoccum sp. (20.5%), Lasiodiplodia sp. (10.1%), Trichoderma sp. (9.91%), and Fusarium sp. (9.70%) were the most predominant genera. La Joya de los Sachas exhibited the highest fungal richness (14 genera), as supported by a Shannon diversity index of 2.30. This study provides in-sights into the distribution and diversity of fungal communities inhabiting cacao fruits in the northern Ecuadorian Amazon, which may have implications for both disease management and the discovery of novel bioactive compounds. Further research is warranted to investigate the functional roles of these fungi, including their potential as biocontrol agents or sources of novel pharmaceuticals. Additionally, exploring the influence of environmental factors and agricultural practices on cacao fruit mycobiota could further enhance our understanding of this complex ecosystem.
Keywords:
Cacao
; Amazon
; Fungal diversity
; Biological Control
1. Introduction
Plants ubiquitously associate with diverse microorganisms [1], forming complex interactions that span the spectrum from mutualism to antagonism [2]. Fungi, in particular, play a critical role in shaping the diversity and composition of terrestrial plant communities. Despite their importance, our understanding of these intricate plant-fungal interactions remains limited. Current research has documented such ecological relationships for less than 5% of the estimated 1.5 million extant fungal species [3]. This vast and largely unexplored fungal kingdom encompasses a wide functional diversity, with some species acting as plant pathogens, capable of infecting crops at any stage of development, while others contribute to improved crop health and resilience [4]. This underscores the need for further research to elucidate the complex interplay between plants and their associated fungal communities.
Fungi specially endophytic fungi represent a remarkably diverse group of microorganisms that contribute significantly to plant health [5]. These fungi colonize the internal tissues of a wide range of plant hosts, including mosses, liverworts, ferns, conifers, and angiosperms, typically developing asymptomatically within healthy aerial tissues [6]. Endophytes exert beneficial effects on their hosts through various mechanisms, including the production of phytohormones that promote plant growth and antimicrobial compounds that defend against pathogens [1]. Furthermore, they compete with plant pathogens for resources and ecological niches [7], effectively limiting pathogen establishment and proliferation. While research on plant-associated microbes has often focused on plant pathogens and mycorrhizal fungi, particularly in temperate environments and agricultural systems, increasing attention is being directed towards the potential of endophytic fungi for biological control and the production of pharmacologically active compounds [3].
The dominant phylum among fungi is Ascomycota, while Basidiomycota and Mucoromycota are less frequently observed in this ecological group [9]. These fungi have been identified in association with an estimated 300,000 plant species (it's unlikely to be three million as there are only about 391,000 known plant species), colonizing diverse plant tissues, including leaves, flowers, petioles, roots, and fruits [10]. Coevolutionary processes between fungi and their hosts have been documented, often resulting in a reduction of fungal virulence or pathogenicity [11]. The nature of the interaction between fungi and their plant hosts can vary considerably, spanning a spectrum from symbiotic to mutualistic to pathogenic [12,13], depending on environmental conditions and the health status of the host plant [8].
Several studies have investigated the fungal communities associated with cocoa (Theobroma cacao L.). For instance identified six fungal species belonging to four taxa: Lasiodiplodia pseudotheobromae, Arthrinium rasikravindrae, Diaporthe sp., Lasiodiplodia theobromae, and Colletotrichum sp., representing four families: Botryosphaeriaceae, Apiosporaceae, Diaporthaceae, and Glomerellaceae [7]. Furthermore, some researchers [4,14] reported that various fungi, including Trichoderma, Pestalotiopsis, Curvularia, Tolypocladium, Colletotrichum sp., Botryosphaeria sp., Xylaria sp., Clonostachys sp., Fusarium spp., Acremonium sp., and Phomopsis sp., isolated from T. cacao, are environmentally acquired and contribute to plant defense mechanisms. These mechanisms include induced systemic resistance, pathogen eradication, and pathogen inhibition through the secretion of bioactive substances [1].
Beyond the previously discussed fungi associated with T. cacao, several pathogenic fungi and oomycetes cause diseases affecting the flowering, foliage, and fruit of this crop. These pathogens include Phytophthora spp., Moniliophthora roreri, Moniliophthora perniciosa, Ceratosystis cacaofunesta, and Oncobasidium theobromae. Among these, Phytophthora spp. and Moniliophthora roreri are particularly significant, causing substantial global cocoa production losses, estimated between 20% and 60% [14,15,16]. These pathogens often exhibit resilience to adverse environmental conditions, including high temperatures, excessive rainfall, and elevated relative humidity [17]. Furthermore, climate change may influence the distribution and severity of plant diseases, potentially altering host resistance [18].
While research has explored fungal communities associated with T. cacao in various regions, there remains a significant gap in our understanding of fungal diversity within cocoa fruits specifically in the northern Ecuadorian Amazon. This knowledge gap is critical because fungi can play important roles in plant health, including disease resistance and nutrient acquisition. Furthermore, the unique environmental conditions of the northern Ecuadorian Amazon may harbor novel or understudied fungal species with potential biotechnological applications. Therefore, this study aims to comprehensively characterize the diversity of fungi associated with T. cacao fruits in this region using a combined cultural, morphological, and molecular approach. This multifaceted approach will provide a robust assessment of fungal diversity, enabling a deeper understanding of the ecological roles of these fungi and their potential contributions to cocoa cultivation in the Amazon.
2. Materials and Methods
2.1. Samples Collection
A total of 59 healthy cocoa (Theobroma cacao L.) fruit samples from traditional varieties were collected for this study. These samples were obtained from cocoa producers in the northern region of the Ecuadorian Amazon. Collection sites were located within Orellana province, Ecuador, encompassing three specific locations: Loreto (0°47'28''S, 77°22'19''W, 373 m a.s.l.), Francisco de Orellana (0°37'36''S, 77°07'40''W, 292 m a.s.l.), and La Joya de los Sachas (0°45'41''S, 77°05'01''W, 330 m a.s.l.). These locations represent distinct microclimates within the region. Loreto has an average temperature of 23.6°C, 81-89% relative humidity, and 2516 mm of annual rainfall. Francisco de Orellana experiences an average temperature of 26°C, 86-89% relative humidity, and 2800-4500 mm of annual rainfall. La Joya de los Sachas is characterized by an average temperature of 25°C, 90% relative humidity, and 4534 mm of annual precipitation. Healthy cocoa fruit samples were placed in sterile plastic bags, transported in a cooler, and stored at 4-8°C in the laboratory until processing.
2.2. Isolation of Fungi Associated with Healthy Cocoa Fruits
Fungal isolation was conducted using a sterilized scalpel to excise five tissue fragments (approximately 25 mm²) from each cocoa fruit. Under sterile conditions within a laminar flow chamber, the fragments were surface-sterilized by sequential immersion in 70% alcohol for 30 s, 2% sodium hypochlorite for 2 minutes, and three rinses with sterile distilled water for 30 s each. Fragments were then transferred to sterile paper towels for drying. Subsequently, the sterilized fragments were placed on Petri dishes containing potato dextrose agar (PDA, Difco™) supplemented with chloramphenicol (25 μg/mL) and streptomycin (25 μg/mL). Plates were incubated at 26°C in darkness for 12 days [14]. Pure cultures were obtained by transferring a portion of mycelium from the initial plates to PDA plates. These plates were sealed with parafilm, incubated at 26°C in darkness for 12 days in an inverted position. Monosporic cultures were prepared [19], briefly, a small portion of mycelium from pure cultures was transferred to test tubes containing 10 mL of sterile distilled water using a sterilized toothpick. After shaking for 10 s, the resulting suspension was spread onto synthetic nutrient-poor agar in Petri dishes. Excess water was removed, and the plates were sealed, labeled, and incubated laterally at 26°C for 24 h. Germinated spores were identified using a stereoscope, and single spores were excised with a scalpel and transferred to PDA plates. These plates were sealed, labeled, and incubated at 26°C, with daily monitoring of isolate growth. All fungal isolates obtained from T. cacao fruits were preserved using the method described by Castellani [20] and stored at 4-8°C.
2.3. Morphological and Cultural Characterization
Following the adhesive strip method [21,22], a drop of lactoglycerol was applied to a microscope slide, and a transparent adhesive tape was used to lift a portion of the fungal colony, taking care to avoid the formation of air bubbles. Microscopic and cultural analyses were conducted using a Leica DML optical microscope equipped with phase contrast and/or a Leica DMR epifluorescent light microscope equipped with a 50 W mercury lamp and a 450-490 nm excitation filter. Fungal identification was primarily based on macroscopic colony characteristics and the micromorphology of reproductive structures. Cultured colonies were characterized based on mycelial color (obverse and reverse) [23], margin shape, surface texture, elevation, edge characteristics, and mycelium color, using standard colors according to the Pantone Color Matching System. Trichoderma isolates were characterized following some methods of [24,25,26,27,28]. For each fungi isolate, 10 random measurements were taken of the following structures: hyphae, conidia, conidiophores, conidiogenous cells, and pycnidia.
2.4. DNA Extraction, Amplification and Sequencing
Genomic DNA was extracted from monoconidial cultures using the Wizard® Genomic DNA Purification Kit. DNA was quantified on 1% agarose gels against a 50 ng lambda DNA molecular weight marker (Promega, Madison, WI, USA), and visualized using a Loccus Biotechnology Molecular Imaging Transilluminator. DNA quality and concentration were assessed using a Nanodrop® 2000c (Thermo Fisher Scientific, Waltham, MA, USA) spectrophotometer by measuring the 260/280 absorbance ratio. DNA was then diluted to a final concentration of 50 ng/µL. The internal transcribed spacer region of ribosomal DNA was amplified by PCR using 0.2 µM of ITS1/ITS4 primers [30], 1× Taq DNA Polymerase Master Mix (Promega, Madison, WI, USA), and approximately 50 ng of template DNA in a total reaction volume of 50 µL. PCR cycling conditions were as follows: initial denaturation at 95°C for 5 min, followed by 35 cycles of 95°C for 1 min, 56°C for 45 s, and 72°C for 90 s, with a final extension at 72°C for 10 min, using a Thermo Fisher Scientific thermocycler (Thermo Fisher Scientific, Waltham, MA, USA). Amplification success was confirmed by electrophoresis on a 1% agarose gel. Amplicons were purified using the NZYGelpure kit and sent to Macrogen for Sanger sequencing. Resulting electropherograms were analyzed using 4Peaks software, and sequences were analyzed using the BLAST algorithm at the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov).
2.5. Fungal Diversity Analysis
The analyses of taxonomic composition were performed using the relative abundance matrix of genera identified, later grouped in Operational Taxonomic Unit (OTUs), where each genus reflects one OTU. Plots with genera taxonomic composition were constructed using R 4.4.1 program (R Development Core Team) with the ggplot library. Analysis of fungal diversity was calculated using the Hill series (effective number of genera) with the library iNEXT using the relative abundance of the taxa found in the samples. The alpha diversity, which analyzes the diversity within each sample, was estimated by local (La Paz, Bajo Huino, La Belleza, San Carlos, San Jacinto and Calumeña) obtaining Richness (q = 0), Shannon diversity (q = 1), and Simpson diversity (q = 2).
3. Results
3.1. Analysis of the Diversity of Fungi Present in Cocoa Fruits
A total of 464 fungal samples were isolated from T. cacao fruits. Most of the isolates were identified by their morphological and molecular characteristics, being the most abundant fungi of the genus Penicillium sp. (27.8%), Epicoccum sp. (20.5%), Lasiodiplodia sp. (10.1%) Trichoderma sp. (9.91%) and Fusarium sp. (9.70%); and, the least frequent Nigrospora sp, Clonostachys sp., Diaporthe sp., Daldinia sp., Neopestalotiopsis sp., Alternaria sp., Purpureocillium sp., Coniochaeta sp. and Geotrichum (Table 1).
In the canton of Loreto, 13 genera were found in the two localities, seven in La Paz and six in Bajo Huino. In the two localities, four genera were found in common (Penicillium sp.: 33.33 and 36.61%, Epicoccum sp.: 22.22 and 27.78%, Lasiodiplodia sp. 18.52 and 4.17% and Fusarium sp.: 14.81 and 4.17%). In addition, in the two localities the fungal were grouped in three (Diaporthe sp.: 1.86, Alternaria sp.: 5.56 and Purpureocillium sp. 3.7%) and two (Trichoderma sp.: 18.05 and Nigrospora sp.: 9.72%) different genera, respectively. In the canton Francisco de Orellana, in the locality of La Belleza, in order of abundance, ten genera were determined: Penicillium sp. (25.53%), Lasiodiplodia sp. (21.28%), Trichoderma sp. (15.96%), Epicoccum sp. (15.95%), Diaporthe sp. (6.38%), Geotrichum sp. (4.26%), Clonotachys sp. (3.19%), Daldinia sp. (3.19%), Neopestalotiopsis sp. (2.13%) and Alternaria sp. (2.13%). In La Joya de los Sachas, 14 genera and three genera in common were identified in the three localities (Penicillium sp.: 38.14, 2.27 and 22.33%, Fusarium sp.: 4.12, 4.55 and 27.18% and Clonotachys sp.: 4.12, 4.55 and 7.77%). In San Carlos and La Calumeña, three genera were found in common (Epicoccum sp.: 25.77 and 22.34, Lasiodiplodia sp.: 9.27 and 4.85%, Nigrospora sp.: 12.37 and 5.82%). In San Carlos and San Jacinto two genera were identified in common: Coniochaeta sp. (2.6 and 9.09%) and Geotrichum sp. (2.07 and 2.27%) as well as in San Jacinto and La Calumeña, Diaporthe sp. (9.09 and 4.85%) and Daldinia sp. (6.28 and 4.85%). In addition, in San Carlos and San Jacinto, four more genera were identified (two per locality), in San Carlos the genera Alternaria sp. (1.04%) and Purpureocillium sp. (2.6%) and in San Jacinto Trichoderma sp. (40.91%) and Neopestalotiopsis sp. (20.14%). Finally, fungal not identified by genus were found in the locality La Belleza in Francisco de Orellana (4.26%) and in San Carlos in Joya de los Sachas (2.07%) (Figure 1).
The analysis of relative abundance for the group of fungal microoganisms identified in the fruits of T. cacao allowed them to be grouped in the fungal phylum Ascomycota. On the other hand, the analysis by class grouped the fungal into Sordariomycetes (39.85%), Dothidiomycetes (31.89%), Eurotiomycetes (27.8%) and Saccharomyces (0.43%). In addition, in the two localities (La Paz and Bajo Huino) of Loreto canton, in Francisco de Orellana (La Belleza) and in two localities (San Carlos and La Calumeña) in La Joya de los Sachas, three classes of fungi (Eurotiomycetes, Dothidiomycetes, Sordariomycetes) were present; and Saccharomyces was identified in two localities (San Carlos and San Jacinto) of La Joya de los Sachas.
In the localities La Paz and Bajo Huino, the fungal were grouped in Eurotiomycetes (33.33 and 36.12%), Dothidiomycetes (46.3 and 31.94%) and Sordariomycetes (20.37 and 31.94%), respectively. In the canton of Francisco de Orellana, the most abundant class was Dothidiomycetes (39.09%), followed by Sordariomycetes (35.11%) and finally Eurotiomycetes (25.53%). In the canton of La Joya de los Sachas, in San Carlos, San Jacinto and La Calumeña, Eurotiomycetes (38.14, 2.27 and 22.33%) and Sordariomycetes (24.74, 95.46 and 50.49%), respectively, were identified. Also, the class Dothidiomycetes (36.09 and 27.18%) was identified only in San Carlos and La Calumeña; and Saccharomycetes (2.27 and 1.03%) in San Carlos and San Jacinto (Figure 2).
The analysis at order level, allowed classifying the fungal in seven orders, Eurotiales and Hypocreales (26.29 and 25.86%, respectively) were present in the six localities; Pleosporales, Xylariales and Botryosphaeriales (20.46, 11.95, 9. 68%, respectively) were only found in five localities; Diaporthales, Coniochaetales and Saccharomycetales (3.70 and 1.51, 0.55%, respectively), were the lowest frequency orders and were present in four and two localities, respectively.
In addition, the greatest number of orders were present in Cantones Francisco de Orellana and La Joya de los Sachas, six for each locality. On the other hand, in Loreto, in the two localities the fungal were grouped in five orders. In La Paz and Bajo Huino there was more presence of Eurotiales (36.11 and 33.33%), Pleosporales (27.68 and 27.78%) and Hypocreales (18.52 and 22.22%) respectively. Botryosphaeriales were present in greater numbers in La Paz (18.52%) with respect to Bajo Huiruno (4.17%). the order Xylariales was only present in Bajo Huiruno (9.72%) and Diaporthales in La Paz (1.85%). In Francisco de Orellana, the fungal microorganisms were grouped into six orders, the most frequent were Eurotiales (25.53%), Botryosphaeriales (21.28%), Hypocreales (19.16%), Saccharomycetales (18.08%), Xylariales (9.57%), Diaporthales (6.38%). In La Joya de los Sachas, the abundance of microorganisms by locality was very variable, in San Carlos the orders identified were Eurotiales (38.15%), Pleosporales (26.8%), Xylariales (14.44%), Hypocreales (10.31%), Botryosphaeriales (9.27%) and Saccharomycetales (1. 03%); in San Jacinto the orders Hypocreales (50%), Hylariales (34.95%), Diaporthales (9.1%), Coniochaetales (9.09%) and Saccharomycetales (2. 27%); and, in La Calumeña Hypocreales (34.96%), Eurotiales (22.33%), Pleosporales (22.33%), Hylariales (10.68%), Botryosphaeriales (4.85%) and Diaporthales (4.85%) were present (Figure 3).
Relative abundance at the family level allowed grouping the fungal microorganisms into 15 families. In order of importance, the presence of Aspergillaceae (26.28%), Didymellaceae (19.01%), Hypocreaceae (12.49%), Botryosphaeriaceae (9 .68%), Nectria-ceae (9.13%), Apiosporaceae (4.65%), Sporocadaceae (3.76%), Diaporthaceaceaceae (3.70%), Bionectriaceae (3.27%), Hypoxylaceae (2.48%), Plesporaceae (1.46%), Coniochaetaceae (1.52%), Xylariaceae (1.06%), Ophiocordycipitaceae (0.96%) and Dipodascaceae (0.55%).
In Loreto canton, in the two localities La Paz and Bajo Huino, microorganisms were grouped into 9 families, the most abundant being Aspergillaceae (33.33 and 36.11%) and Didymellaceae (22.22 and 22.78%); in addition, two similar but less abundant families were identified in the two sites: Botryosphaeriaceae (21.28 and 4.17%) and Nectriaceae (14.81 and 4.17%). In La Paz, 3 families were identified (Plesporaceae 5.56%, Ophiocordycipitaceae 3.70% and Diaporthaceaceae 1.86%) that were not present in Bajo Huiruno; and two that were present in Bajo Huiruno (Hypocreaceae 18.05% and Apiosporaceae 9.72%) but not in La Paz. In the canton Francisco de Orellana, in La Belleza, the microorganisms were grouped in ten families, four similar to those of the canton Loreto and six different; Aspergillaceae (25.53%), Didymellaceae (15. 95%) Botryosphaeriaceae (15.95%), Hypocreaceae (15.96%), Diaporthaceaceaceae (6.38%), Coniochaetaceae (4.26%), Biocectriaceae (3.19%), Hypoxylaceae (3.19%), Plesporaceae (2.13%) and Sporocadaceae (2.13%) (Figure 4).
In the canton of La Joya de los Sachas, in San Carlos, San Jacinto and La Calumeña the microorganisms were grouped in ten, nine and eight families, respectively. In the three localities the similar families were: Aspergillaceae (38.14, 2.27 and 22.34%), Bionectriaceae (4.12, 4.55 and 7.77%) and Nectriaceae (4.12, 4.55 and 27.18%). San Carlos and La Calumeña presented three identical families: Didymellaceae (25.77 and 22.34%), Botryosphaeriaceae (9.27 and 4.85%), Apiosporaceae (12.37 and 5.82%); and, between San Carlos and San Jacinto a similar family Dipodascaceae (1.04 and 2.27%) was identified. In addition, two identical families were identified in San Jacinto and La Calumeña: Diaporthaceaceae (9.09 and 4.85%) and Hypoxylaceae (6.28 and 4.85%). Finally, three more families were identified in San Carlos (Plesporaceae: 1.04%, Xylariaceae: 2.07% and Ophiocordycipitaceae: 2.06%) as in San Jacinto: Hypocreaceae 40.91%, Sporocadaceae 20.45% and Coniochaetaceae 9.09% (Figure 4).
The highest richness of fungal microorganisms was found in La Joya de los Sachas, with 14 genera, followed by Francisco de Orellana with 9 genera and finally Loreto with 9 genera. The analysis by locality shows that in La Paz and Bajo Huino 7 and 6 genera were present, respectively. In La Belleza, 10 genera of the 14 genera were found; and in San Carlos, San Jacinto and La Calumeña, 11, 9 and 7 genera were present, respectively.
The average values recorded according to the Shannon Diversity (q=1) index in the cantones Loreto, Francisco de Orellana and La Joya de los Sachas were 1.76, 1.96 and 2.30 respectively. In La Paz, the Shannon diversity index was 1.65 and Bajo Huino was 1.52. In La Belleza, canton Francisco de Orellana, the Shannon diversity index was 1.96. Finally, in the caton La Joya de los Sachas, the diversity of the San Carlos was 1.80, San Jacinto was 1.76 and La Calumeña was 1.74 (Figure 6). According to the Shannon Diversity (q=1) index, the sample population with the highest fungal diversity value was found in La Joya de los Sachas, with 2.30.
According to the results of Simpson Diversity (q=2) index, the sample population was found 0.21, in Bajo Huino 0.25, with a higher probability of individuals of the same gender. In Francisco de Orellana and La Joya de los Sachas the Simpson Diversity (q=2) index was 0.16 and 0.12, respectively.
3.2. Cultural, Morphological and Molecular Characterization of Fungal Microorganisms
The relative abundance at the genus level of the fungal present in the fruits of T. cacao allowed the selection of the five most abundant genera in the three cantons; cultural (Table 2), morphological (Table 3) and molecular (Table 4) characterization was carried out for these genera. The cultural characterization of the fungi shows that the morphotypes presented circular, irregular and filamentous mycelial growth; with flat, curly, filamentous and effuse elevations; with smooth, rough, wavy, concentric and crenulate surfaces and colors varying from yellow, red, green, black, beige and gray on the reverse and reverse (Table 2, Figures 10 – 14 a). And, morphological characterization of morphotypes was performed based on hyphal shape and conidial shape and size (Table 3).
Finally, the isolates were identified by ITS sequences in order to determine species richness and abundance (Table 4). Amplification of the PCR products of the ITS regions of the rDNA of the fungal isolates generated fragments of 300 to 600 bp with primers ITS1 and ITS4. Such size variations were initial evidence that they corresponded to different genera. Analysis with BLASTN confirmed this observation by obtaining first hits with percentage identity values >99% for the genera presented in Table 4. The results of the BLASTN comparisons corresponded to a local alignment leading to putative identifications.
4. Discussion
Analysis of the fungal diversity present in the cocoa fruits revealed a rich mycobiome comprising 14 genera. Penicillium sp. (27.8%), Epicoccum sp. (20.5%), Lasiodiplodia sp. (10.1%), Trichoderma sp. (9.91%), and Fusarium sp. (9.70%) were the most prevalent. Other genera, including Nigrospora sp., Clonostachys sp., Diaporthe sp., Daldinia sp., Neopestalotiopsis sp., Alternaria sp., Purpureocillium sp., Coniochaeta sp., and Geotrichum sp., were detected at lower frequencies. Significantly, many of these genera, such as Epicoccum, Lasiodiplodia, Clonostachys, Fusarium, and Trichoderma, are recognized for their production of metabolites with plant-protective properties [31].
The presence of these potentially beneficial fungi within the cocoa fruit microbiome suggests a complex interplay between the plant and its associated fungi, potentially contributing to a natural defense system against phytopathogens. This observation warrants further investigation into the specific roles and interactions of these fungi within the cocoa ecosystem.
Epicoccum spp., representing a substantial portion of the identified fungi, are known for their antagonistic activity against a broad spectrum of plant pathogens. Previous studies have documented their inhibitory effects on Pythium ultimum, Aphanomyces cochlioides, and Rhizoctonia solani, as well as antibiotic activity against Escherichia coli, and multidrug-resistant strains of Bacillus subtilis and Staphylococcus aureus [32]. Their presence in cocoa fruits may contribute to a natural defense mechanism against these and other potential pathogens. Further research should explore the specific mechanisms of action employed by Epicoccum spp. in the cocoa fruit environment, including the identification and characterization of the bioactive metabolites involved.
Lasiodiplodia spp., often found as endophytes, have demonstrated biocontrol potential against two major cocoa pathogens, Moniliophthora roreri (causing frosty pod rot) and Moniliophthora perniciosa (causing witches' broom disease) [33]. This finding highlights the potential of Lasiodiplodia spp. as biocontrol agents in cocoa cultivation. Further research is needed to investigate the efficacy and feasibility of utilizing Lasiodiplodia spp. in integrated disease management strategies, including the development of appropriate formulations and application methods.
Trichoderma spp., widely recognized for their antifungal activity, are also well-represented in the cocoa fruit mycobiome. Their diverse mechanisms of action, including antibiosis, antagonism, mycoparasitism, and induced resistance, make them promising candidates for biocontrol applications. Specifically, Trichoderma asperellum has shown efficacy in suppressing Anthracnose dieback in cocoa [4]. Further research should focus on optimizing the application of Trichoderma spp. for disease management in cocoa, including the selection of effective strains, the development of suitable delivery systems, and the evaluation of their impact on the overall cocoa ecosystem.
The presence of Clonostachys sp. and Fusarium sp., both exhibiting biocontrol activity against Phytophthora palmivora (causing black pod rot) and M. roreri, further enriches the potential of the cocoa fruit mycobiome for disease suppression. While some Fusarium species are known pathogens, others exhibit beneficial properties, highlighting the complexity of fungal interactions within the cocoa ecosystem. Further research is needed to elucidate the specific roles and interactions of these fungi in cocoa disease dynamics [34].
The diverse fungal community inhabiting cocoa fruits, particularly the presence of genera known for their plant-protective properties, offers promising avenues for developing sustainable disease management strategies. Further research focusing on the specific mechanisms of action, interactions, and potential applications of these beneficial fungi will be crucial for harnessing the full potential of the cocoa fruit mycobiome for enhanced disease resistance and sustainable cocoa production.
Four classes within the Ascomycota Phylum were identified. Sordariomycetes (39.85%), Dothidiomycetes (31.89%), and Eurotiomycetes (27.8%) were dominant, while Saccharomyces represented a minor component (0.43%). The observed classes are consistent with those reported in similar studies conducted across diverse geographical regions, including China, India, Germany, Spain, the United Kingdom, Brazil, and the United States, where Sordariomycetes (41%), Dothidiomycetes (25.8%), and Eurotiomycetes (11%) were found [9].
Our study identified eight fungal orders: Eurotiales (26.29%), Hypocreales (25.86%), Pleosporales (20.46%), Xylariales (11.95%), Botryosphaeriales (9.68%), Diaporthales (3.70%), Coniochaetales (1.51%), and Saccharomycetales (0.55%). These orders have also been reported in other studies conducted across Asia, Europe, South America, and North America, albeit in different proportions (Pleosporales (15%), Hypocreales (13%), Xylariales (9%), Eurotiales (8%), Botryosphaeriales and Diaporthales (5%), and Saccharomycetales (1%)) [35]. Furthermore, the identified fungi were classified into 15 families. The observed variation in fungal diversity across the three cantons of the Orellana province likely reflects the influence of environmental factors, such as temperature and rainfall, and host-specific factors, including tissue type (leaves, inflorescences, stem, roots, and fruit), which can affect the presence and abundance of specific microorganisms [35].
The presence of a diverse fungal community within healthy cocoa fruits, including several genera recognized for their plant-protective properties, highlights the complex interplay between the cacao plant and its associated mycobiome. This intricate relationship plays a crucial role in maintaining fruit health and warrants further investigation to fully understand its dynamics and potential applications. The discovery of these beneficial fungi within the cocoa fruit microbiome offers promising avenues for developing sustainable disease management strategies, reducing reliance on synthetic chemical treatments, and promoting environmentally friendly cocoa cultivation practices.
Further research is essential to explore several key aspects of this plant-fungal interaction:
Specific Mechanisms of Action: A deeper understanding of the precise mechanisms employed by these beneficial fungi to suppress pathogens is crucial. This includes investigating the production of antifungal metabolites, competition for resources, mycoparasitism, and the induction of systemic resistance in the cacao plant. Identifying and characterizing the bioactive compounds involved will be essential for developing targeted biocontrol strategies.
Interactions within the Mycobiome: The cocoa fruit mycobiome is a complex community, and the interactions between different fungal species can influence their overall impact on plant health. Research should explore the synergistic or antagonistic relationships between beneficial fungi and potential pathogens, as well as the factors that influence community composition and stability.
Environmental Influences: Environmental factors, such as temperature, humidity, and soil conditions, can significantly impact the composition and activity of the cocoa fruit mycobiome. Understanding how these factors influence the prevalence and efficacy of beneficial fungi is crucial for developing effective biocontrol strategies.
Application Strategies: Developing practical and effective methods for applying beneficial fungi in cocoa cultivation is essential for translating research findings into real-world applications. This includes exploring different formulation methods, delivery systems, and application timings to optimize the efficacy of biocontrol agents.
Impact on Cocoa Quality: While the focus is on disease suppression, it's important to consider the potential impact of beneficial fungi on cocoa bean quality and flavor. Research should investigate whether these fungi influence the chemical composition of the beans and the resulting chocolate products.
By addressing these research questions, we can gain a comprehensive understanding of the role of beneficial fungi in cocoa fruit health and develop sustainable disease management strategies that harness the power of the cocoa fruit mycobiome. This approach offers a promising path towards environmentally friendly and economically viable cocoa production, contributing to the long-term sustainability of the cocoa industry.
5. Conclusions
Investigations into fungal communities represent a burgeoning field of research, providing insights into the diverse microorganisms inhabiting plants. This study characterized 464 fungal isolates from healthy cocoa fruits, classifying them into 56 morphotypes and 14 genera, all within the Ascomycota phylum. The most prevalent genera were Penicillium (27.8%), Epicoccum (20.5%), Lasiodiplodia (10.1%), Trichoderma (9.91%), and Fusarium (9.70%), highlighting the rich fungal diversity associated with cocoa. This diverse assemblage of fungi suggests a complex interplay between the cocoa plant and its associated mycobiome, warranting further investigation into their ecological roles and potential impact on cocoa health and disease.
This study revealed a diverse fungal community associated with cocoa fruits, emphasizing the importance of understanding the intricate interplay between these fungi and their host plant. The identified genera, including Penicillium, Epicoccum, Lasiodiplodia, Trichoderma, and Fusarium, represent a complex ecosystem with potential implications for cocoa health and disease management. The dominance of these genera suggests their adaptation to the cocoa fruit environment and raises questions about their specific roles and interactions.
The observed fungal diversity also reflects broader ecological patterns. The dominance of Ascomycota, particularly Sordariomycetes, Dothidiomycetes, and Eurotiomycetes, aligns with findings from similar studies conducted in diverse geographical regions. This suggests a conserved pattern of fungal community structure associated with cocoa across different environments. Understanding these global patterns can inform broader biocontrol strategies and contribute to a more holistic understanding of cocoa-fungal interactions.
Further research is crucial to fully explore the potential of these beneficial fungi for developing sustainable disease management strategies in cocoa cultivation. This includes investigating the specific mechanisms of action, optimizing application methods, and evaluating the long-term efficacy and environmental impact of biocontrol strategies. Reducing reliance on synthetic fungicides is a key goal, as it promotes environmentally friendly agricultural practices and minimizes potential risks to human health and the environment. By harnessing the natural protective properties of beneficial fungi, we can move towards more sustainable and resilient cocoa cultivation practices.
Author Contributions
Conceptualization and draft preparation, P.I.A.R.; methodology, L.A.H.S., S.E.S.C., A.F.T.A.F.F and D.A.R.R..; supervision and editing, P.I.A.R, L.A.H.S. and E.P.S.C.; analysis and editing, P.I.A.R, A.F.T.A.F.F and L.A.H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Escuela Superior Politécnica de Chimborazo (ESPOCH).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available upon request to the corresponding author.
Acknowledgments
We would like to thank the Escuela Superior Politécnica de Chimborazo (ESPOCH) for all the support rendered during the course of this research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Alves-Júnior, M.; de Sousa, F.O.; Silva, T.F.; Albino, U.B.; Garcia, M.G.; Moreira, S.M.C. de O.; Vieira, M.R. da S. Functional and Morphological Analysis of Isolates of Phylloplane and Rhizoplane Endophytic Bacteria Interacting in Different Cocoa Production Systems in the Amazon. Current Research in Microbial Sciences 2021, 2, 100039.
- Gilbert, G.S. Evolutionary Ecology of Plant Diseases in Natural Ecosystems. Annual Review of Phytopathology 2002, 40, 13–43. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal Endophytes Limit Pathogen Damage in a Tropical Tree. Proc Natl Acad Sci U S A 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Ospina, J.; Molina-Hernández, J.B.; Chaves-López, C.; Romanazzi, G.; Paparella, A. The Role of Fungi in the Cocoa Production Chain and the Challenge of Climate Change. Journal of Fungi 2021, 7, 202. [Google Scholar] [CrossRef]
- Rubini, M.R.; Silva-Ribeiro, R.T.; Pomella, A.W.V.; Maki, C.S.; Araújo, W.L.; dos Santos, D.R.; Azevedo, J.L. Diversity of Endophytic Fungal Community of Cacao (Theobroma Cacao L.) and Biological Control of Crinipellis Perniciosa, Causal Agent of Witches’ Broom Disease. Int J Biol Sci 2005, 1, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Berbee, M.L. The Phylogeny of Plant and Animal Pathogens in the Ascomycota. Physiological and Molecular Plant Pathology 2001, 59, 165–187. [Google Scholar] [CrossRef]
- Chaithra, M.; Vanitha, S.; Ayyasami, R.; Jegadeeshwari, V.; Rajesh, V.; Hegde, V.; Apshara, E. Morphological and Molecular Characterization of Endophytic Fungi Associated with Cocoa (Theobroma Cacao L.) in India. Current Journal of Applied Science and Technology 2020, 1–8.
- Carlsson-Granér, U.; Thrall, P.H. The Spatial Distribution of Plant Populations, Disease Dynamics and Evolution of Resistance. Oikos 2002, 97, 97–110. [Google Scholar] [CrossRef]
- Arnold, A.E. Understanding the Diversity of Foliar Endophytic Fungi: Progress, Challenges, and Frontiers. Fungal Biology Reviews 2007, 21, 51–66. [Google Scholar] [CrossRef]
- Yu, J.; Wu, Y.; He, Z.; Li, M.; Zhu, K.; Gao, B. Diversity and Antifungal Activity of Endophytic Fungi Associated with Camellia Oleifera. Mycobiology 2018, 46, 85–91. [Google Scholar] [CrossRef]
- Sieber, T.N. Endophytic Fungi in Forest Trees: Are They Mutualists? Fungal Biology Reviews 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Usuki, F.; Narisawa, K. A Mutualistic Symbiosis between a Dark Septate Endophytic Fungus, Heteroconium Chaetospira, and a Nonmycorrhizal Plant, Chinese Cabbage. Mycologia 2007, 99, 175–184. [Google Scholar] [CrossRef]
- Tellenbach, C.; Grünig, C.R.; Sieber, T.N. Negative Effects on Survival and Performance of Norway Spruce Seedlings Colonized by Dark Septate Root Endophytes Are Primarily Isolate-Dependent. Environ Microbiol 2011, 13, 2508–2517. [Google Scholar] [CrossRef] [PubMed]
- Hanada, R.E.; Pomella, A.W.V.; Costa, H.S.; Bezerra, J.L.; Loguercio, L.L.; Pereira, J.O. Endophytic Fungal Diversity in Theobroma Cacao (Cacao) and T. Grandiflorum (Cupuaçu) Trees and Their Potential for Growth Promotion and Biocontrol of Black-Pod Disease Available online: https://www.sciencedirect.com/science/article/pii/S1878614610001364 (accessed on 19 October 2023).
- Pico, J.T.; Díaz, A.E.; Vargas Tierras, Y.B.; Viera, W.F.; Caicedo V., C. P21 Evaluación de la Dispersión de Esporas de Alternaria sp. en el Cultivo de Pitahaya (Selenicereus megalanthus) en Palora.; Galápagos, EC: INIAP, Estación Experimental Central de la Amazonía, 2019, 2019; ISBN 978-9978-68-144-2.
- Pico, J.T.; Calderón Peña, E.D.; Fernández A., F.; Díaz M., A. Guía del manejo integrado de enfermedades del cultivo de cacao (Theobroma cacao L) en la amazonía Available online: http://repositorio.iniap.gob.ec/handle/41000/3752 (accessed on 29 October 2023).
- Marelli, J.-P.; Guest, D.I.; Bailey, B.A.; Evans, H.C.; Brown, J.K.; Junaid, M.; Barreto, R.W.; Lisboa, D.O.; Puig, A.S. Chocolate Under Threat from Old and New Cacao Diseases. Phytopathology® 2019, 109, 1331–1343. [Google Scholar] [CrossRef]
- Schroth, G.; Läderach, P.; Martinez-Valle, A.I.; Bunn, C.; Jassogne, L. Vulnerability to Climate Change of Cocoa in West Africa: Patterns, Opportunities and Limits to Adaptation. Science of The Total Environment 2016, 556, 231–241. [Google Scholar] [CrossRef]
- Brunner, C.; Navarro-Barranco, H.; Ayala-Zermeño, M.; Rosas, M.A.; Toriello, C. Obtención y caracterización de cultivos monospóricos de Metarhizium anisopliae (Hypocreales : Clavicipitaceae) para genotipificación.; México, November 7 2013; p. 5.
- Castellani, A. Further Researches on the Long Viability and Growth of Many Pathogenic Fungi and Some Bacteria in Sterile Distilled Water. Mycopathologia et Mycologia Applicata 1963, 20, 1–6. [Google Scholar] [CrossRef]
- Urzı̀, C.; De Leo, F. Sampling with Adhesive Tape Strips: An Easy and Rapid Method to Monitor Microbial Colonization on Monument Surfaces. Journal of Microbiological Methods 2001, 44, 1–11. [Google Scholar] [CrossRef]
- Harris, J.L. Safe, Low-Distortion Tape Touch Method for Fungal Slide Mounts. J Clin Microbiol 2000, 38, 4683–4684. [Google Scholar] [CrossRef] [PubMed]
- Pacasa-Quisbert, F.; Loza-Murguia, M.G.; Bonifacio-Flores, A.; Vino-Nina, L.; Serrano-Canaviri, T. Comunidad de hongos filamentosos en suelos del Agroecosistema de K’iphak’iphani, Comunidad Choquenaira-Viacha. Journal of the Selva Andina Research Society 2017, 8, 2–25. [Google Scholar] [CrossRef]
- Harman, G.E.; Kubicek, C.P. Trichoderma And Gliocladium. Volume 1: Basic Biology, Taxonomy and Genetics; CRC Press, 2002; ISBN 978-1-4822-9532-0.
- Chaverri, P.; Castlebury, L.A.; Overton, B.E.; Samuels, G.J. Hypocrea/Trichoderma: Species with Conidiophore Elongations and Green Conidia. Mycologia 2003, 95, 1100–1140. [Google Scholar] [CrossRef]
- Kraus, G.F.; Druzhinina, I.; Gams, W.; Bissett, J.; Zafari, D.; Szakacs, G.; Koptchinski, A.; Prillinger, H.; Zare, R.; Kubicek, C.P. Trichoderma Brevicompactum Sp. Nov. Mycologia 2004, 96, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- Samuels, G.J.; Lieckfeldt, E.; Nirenberg, H.I. Trichoderma Asperellum, a New Species with Warted Conidia, and Redescription of T. Viride. Sydowia 1999, 51, 71–88. [Google Scholar]
- Torres-De la Cruz, M.; Ortiz-García, C.F.; Bautista-Muñoz, C.; Ramírez-Pool, J.A.; Ávalos-Contreras, N.; Cappello-García, S.; De la Cruz-Pérez, A. Diversidad de Trichoderma En El Agroecosistema Cacao Del Estado de Tabasco, México. Revista Mexicana de Biodiversidad 2015, 86, 947–961. [Google Scholar] [CrossRef]
- Rosero, D.M.C.; Rosales, H.R.B.; Pérez, L.A.C.; Hernández, J.P.O. Población de macrofauna en sistemas silvopastoriles dedicados a la producción lechera:análisis preliminar. La Granja 2018, 27, 77–85. [Google Scholar] [CrossRef]
- Aguilar Doroteo, L.; Zárate Segura, P.B.; Villanueva Arce, R.; Yáñez Fernández, J.; Garín Aguilar, M.E.; Guadarrama Mendoza, P.C.; Valencia del Toro, G. Utilización de marcadores ITS e ISSR para la caracterización molecular de cepas híbridas de Pleurotus djamor. Rev Iberoam Micol 2018, 35, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Asman, A.; Baharuddin, *!!! REPLACE !!!*; Rosmana, A. Ariska Diversity of Fungal Community Associated with Cacao (Theobromae Cacao L.) Top Clones from Sulawesi, Indonesia. IOP Conf. Ser.: Earth Environ. Sci. 2020, 486, 012171. [Google Scholar] [CrossRef]
- Talontsi, F.M.; Dittrich, B.; Schüffler, A.; Sun, H.; Laatsch, H. Epicoccolides: Antimicrobial and Antifungal Polyketides from an Endophytic Fungus Epicoccum Sp. Associated with Theobroma Cacao. Eur J Org Chem 2013, 2013, 3174–3180. [Google Scholar] [CrossRef]
- Villavicencio-Vásquez, M.; Espinoza-Lozano, R.F.; Pérez-Martínez, S.; Castillo, D.S.D. FOLIAR ENDOPHYTE FUNGI AS CANDIDATE FOR BIOCONTROL AGAINST Moniliophthora Spp. OF Theobroma Cacao (Malvaceae) IN ECUADOR. Acta Biológica Colombiana 2018, 23, 235–241. [Google Scholar] [CrossRef]
- Hoopen, G.M.; Rees, R.; Aisa, P.; Stirrup, T.; Krauss, U. Population Dynamics of Epiphytic Mycoparasites of the Genera Clonostachys and Fusarium for the Biocontrol of Black Pod (Phytophthora Palmivora) and Moniliasis (Moniliophthora Roreri) on Cocoa (Theobroma Cacao). Mycological Research 2003, 107, 587–596. [Google Scholar] [CrossRef]
- Mishra, R.; J.S., K.; v, S. A Worldwide List of Endophytic Fungi with Notes on Ecology and Diversity. Mycosphere 2019, 10.
- González-Oreja, J.A. Midiendo la diversidad biológica: más allá del índice de Shannon. Acta zoológica lilloana 2012, 56, 3–14. [Google Scholar]
Figure 1.
Relative abundance at genus level of fungi isolated from T. cacao fruits in the province of Orellana.
Figure 1.
Relative abundance at genus level of fungi isolated from T. cacao fruits in the province of Orellana.
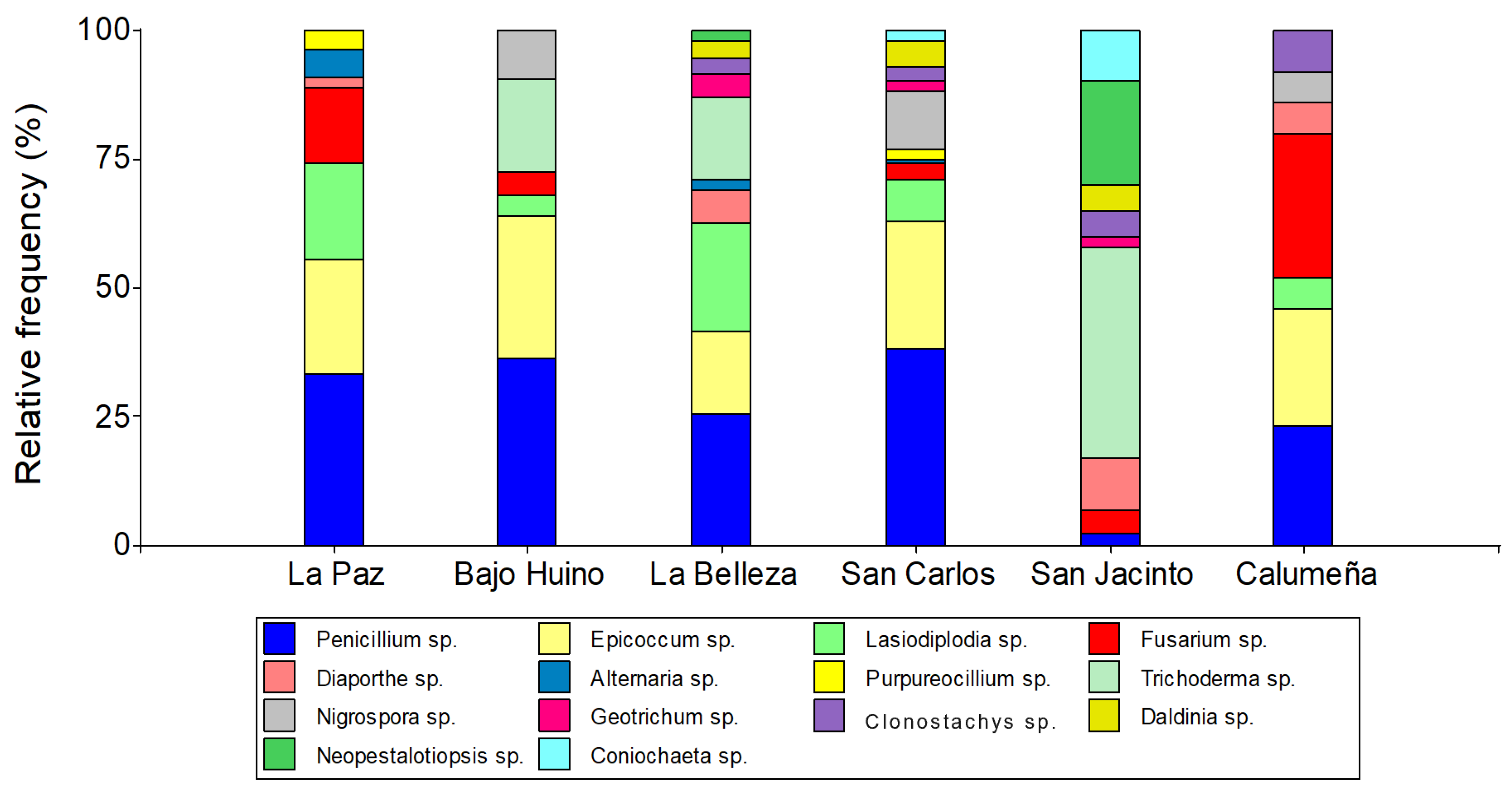
Figure 2.
Relative abundance at the class level of fungi from T. cacao fruits in the province of Orellana.
Figure 2.
Relative abundance at the class level of fungi from T. cacao fruits in the province of Orellana.
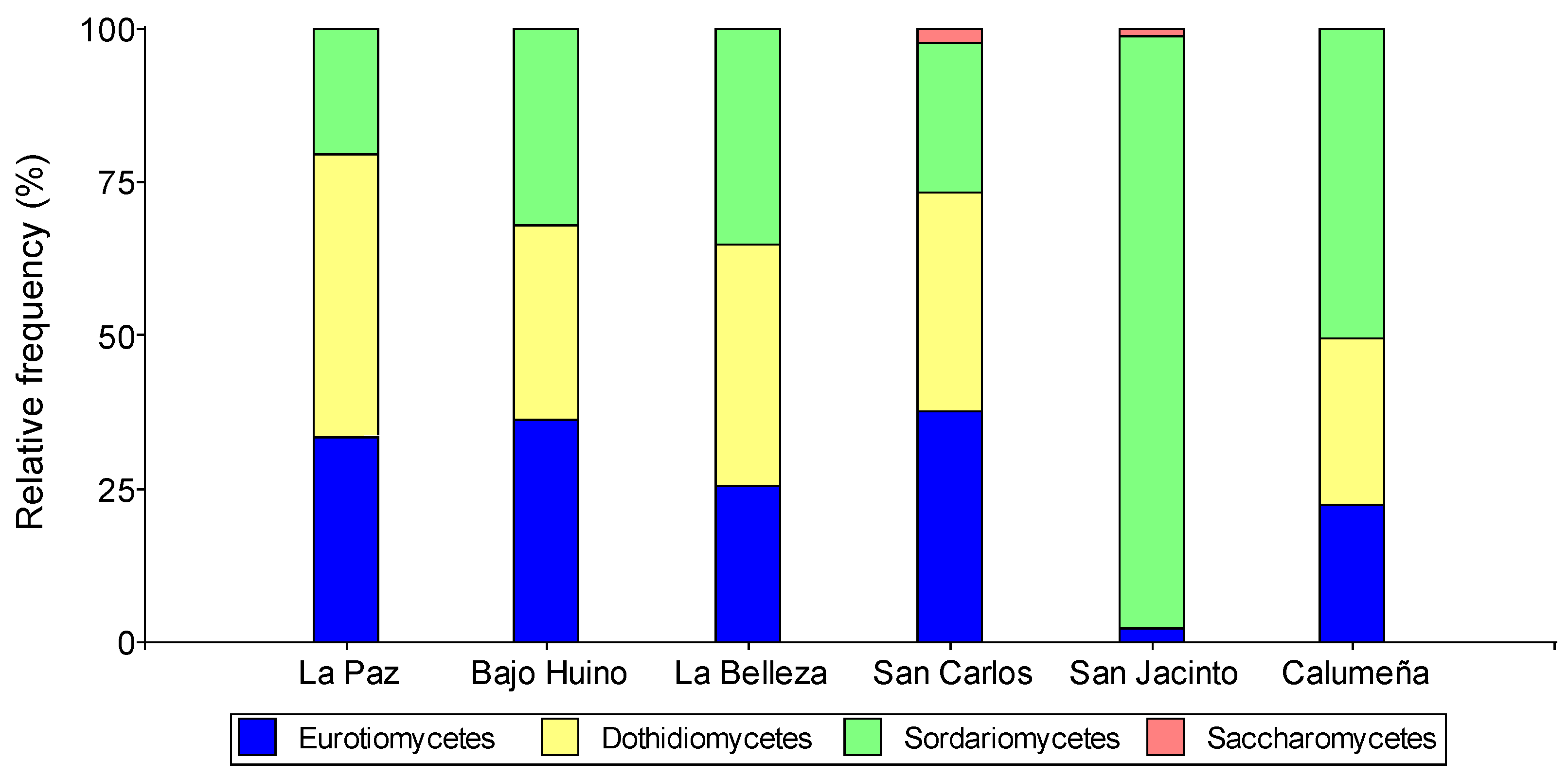
Figure 3.
Relative abundance at order level of fungi isolated from T. cacao fruits in Orellana province.
Figure 3.
Relative abundance at order level of fungi isolated from T. cacao fruits in Orellana province.
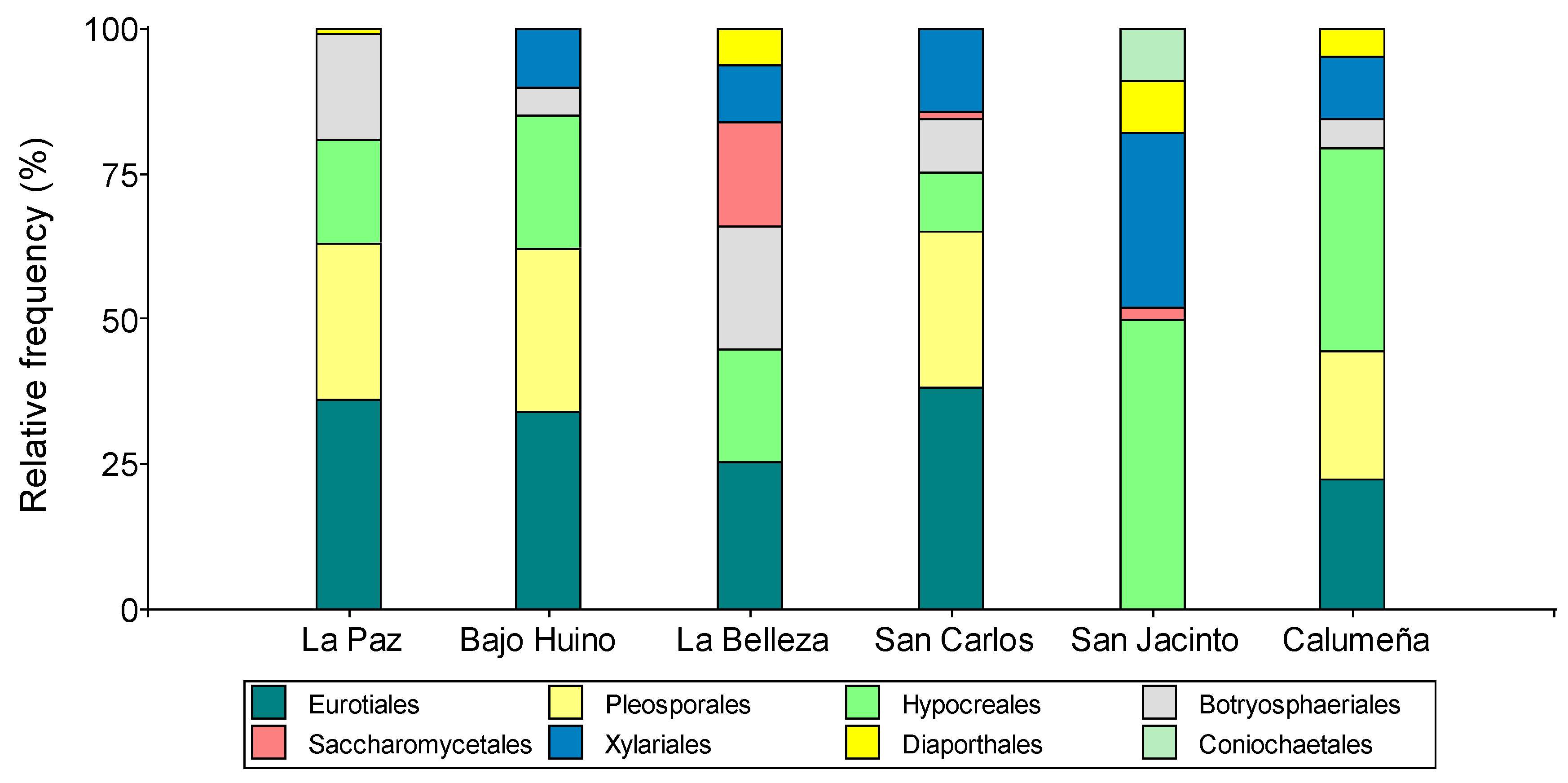
Figure 4.
Relative abundance at the family level of fungal microorganisms isolated from T. cacao fruits in the province of Orellana.
Figure 4.
Relative abundance at the family level of fungal microorganisms isolated from T. cacao fruits in the province of Orellana.
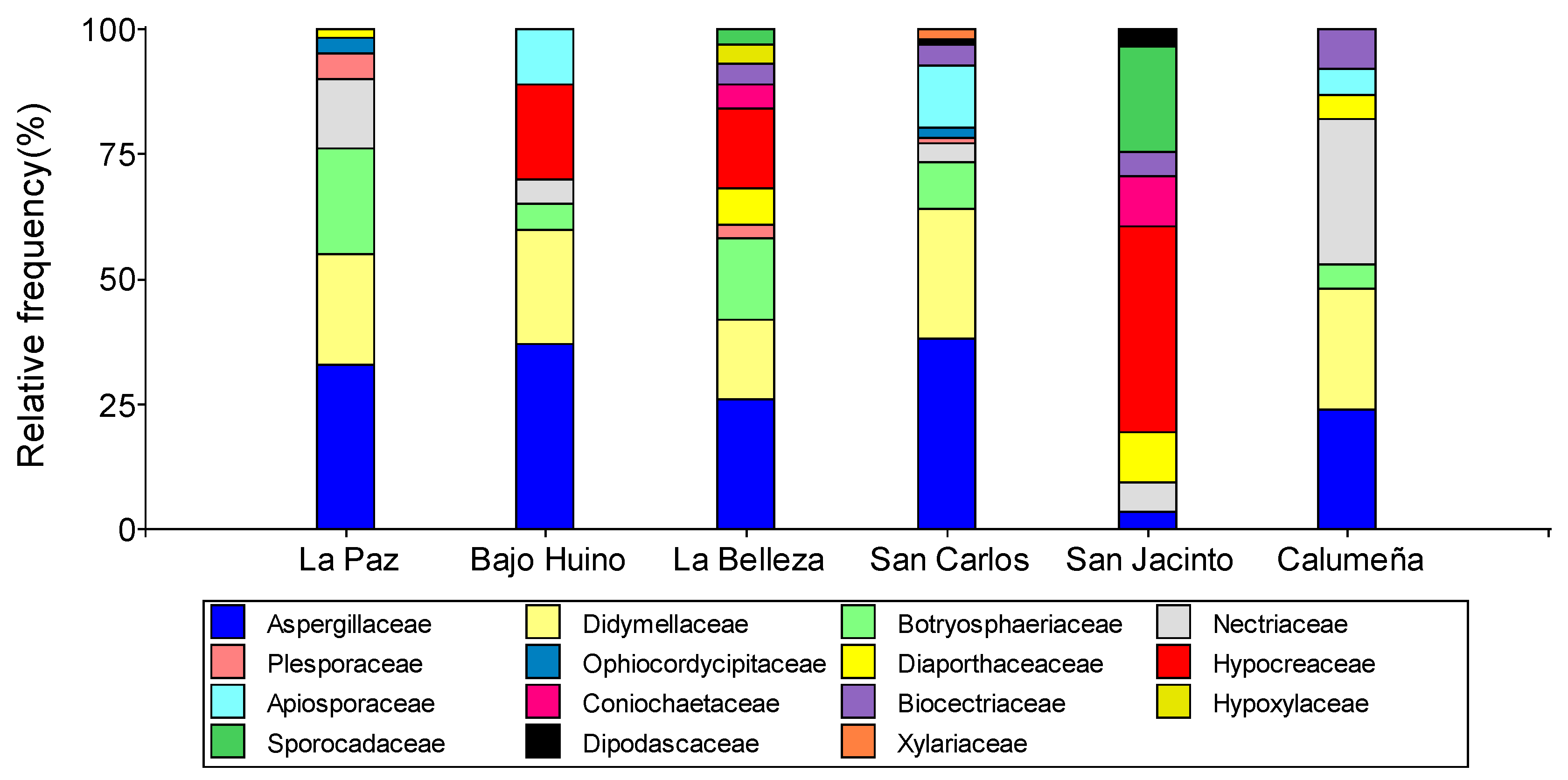
Figure 5.
Richness of fungal microorganisms isolated from T. cacao fruits in the province of Orellana.
Figure 5.
Richness of fungal microorganisms isolated from T. cacao fruits in the province of Orellana.
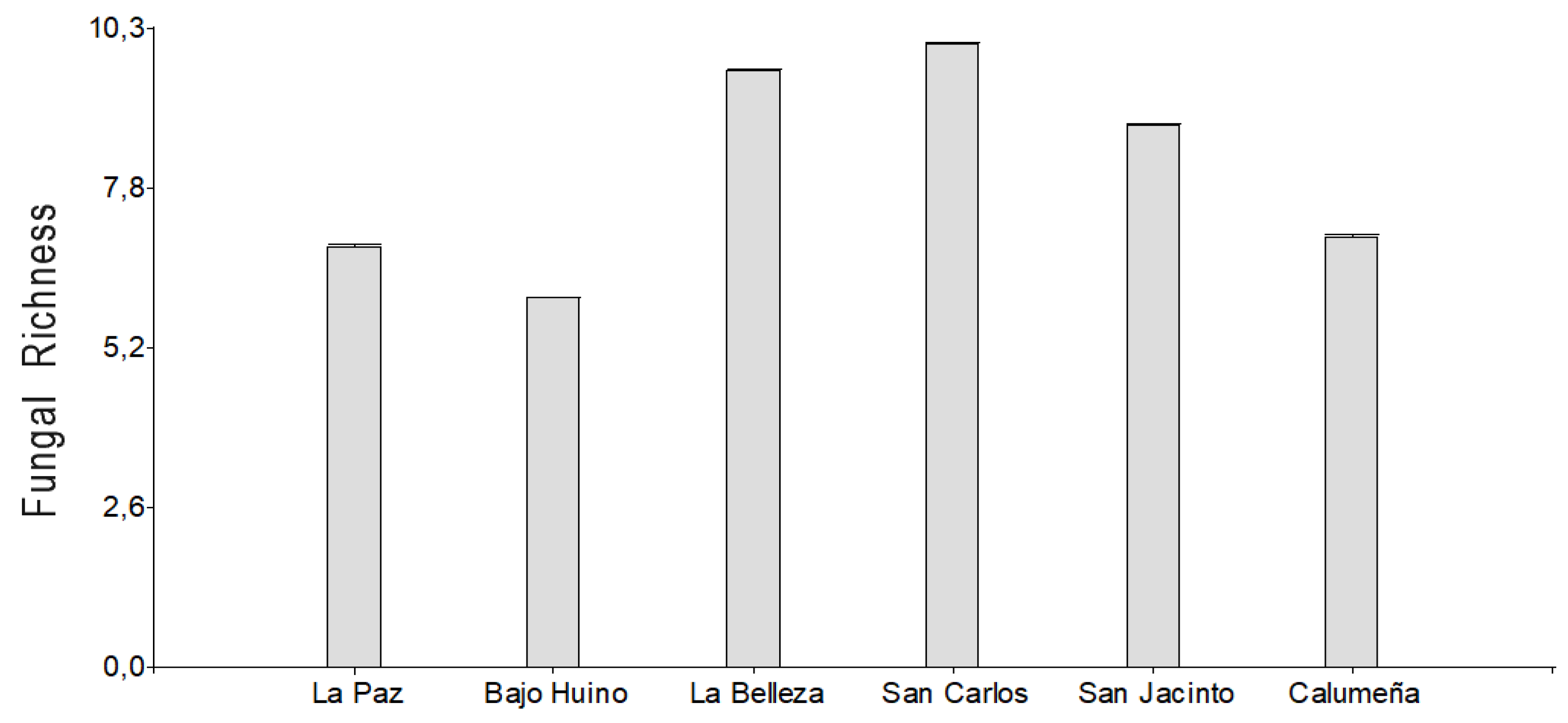
Figure 6.
Diversity indices of fungal microorganism isolated from T. cacao fruits in the province of Orellana.
Figure 6.
Diversity indices of fungal microorganism isolated from T. cacao fruits in the province of Orellana.
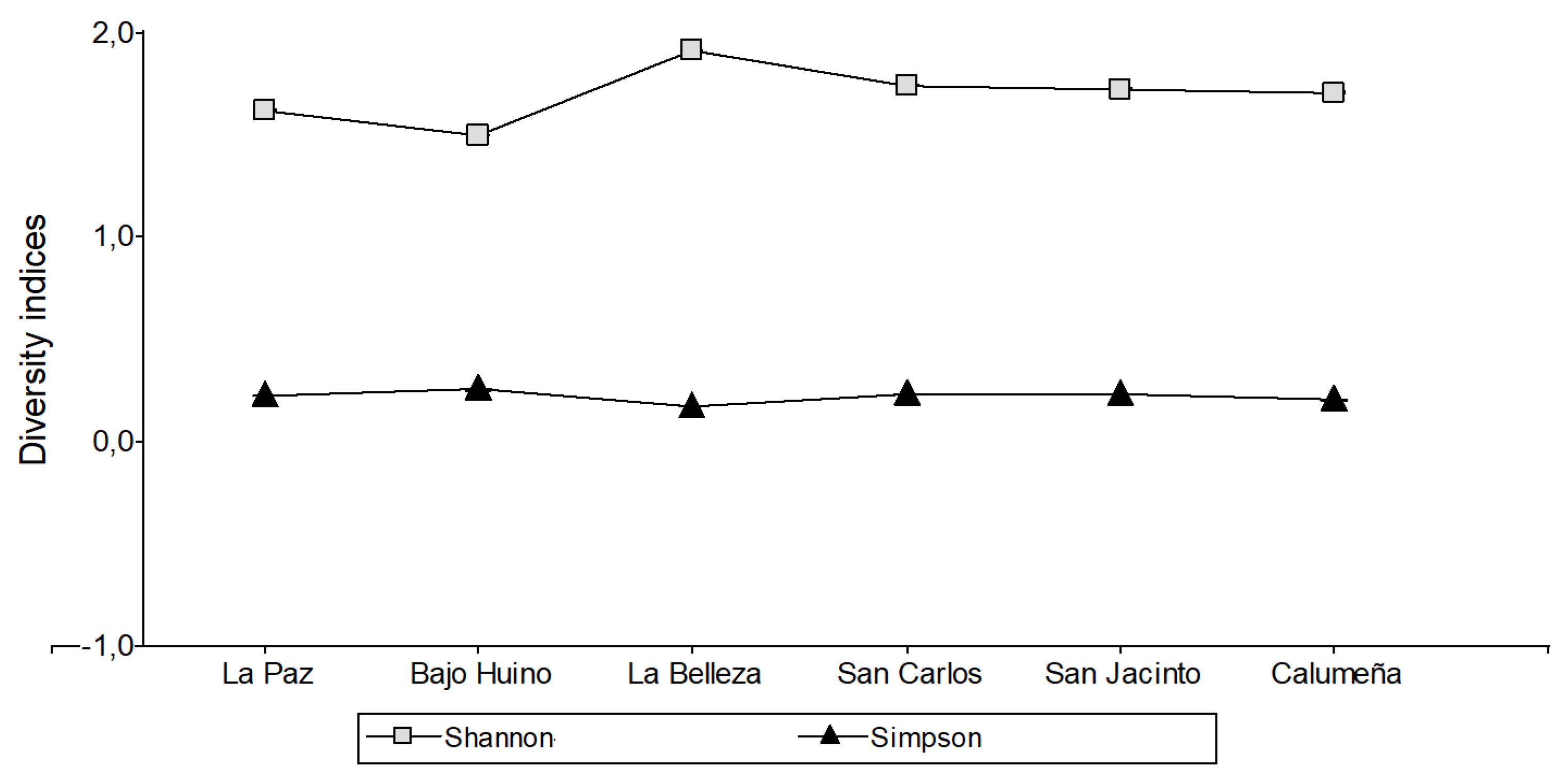

Table 1.
Fungi isolated from T. cacao healthy fruits in the province of Orellana.
| Fungal isolates | Number of isolates | Frequency (%) |
|---|---|---|
| Penicillium | 129 | 27.80 |
| Epicoccum | 95 | 20.47 |
| Lasiodiplodia | 47 | 10.13 |
| Trichoderma | 46 | 9.91 |
| Fusarium | 45 | 9.70 |
| Nigrospora | 25 | 5.39 |
| Clonostachys | 17 | 3.66 |
| Diaporthe | 16 | 3.45 |
| Daldinia | 11 | 2.37 |
| Neopestalotiopsis | 11 | 2.37 |
| Alternaria | 6 | 1.29 |
| Purpureocillium | 4 | 0.86 |
| Coniochaeta | 4 | 0.86 |
| Geotrichum | 2 | 0.55 |
| Not identified | 6 | 1.06 |
Table 2.
Cultural characterization of Fungi isolated of T. cacao fruits in the province of Orellana.
Table 2.
Cultural characterization of Fungi isolated of T. cacao fruits in the province of Orellana.
| Genera | Morphotype | Growth | Type of elevation | Surface | Adverse color | Reverse color |
|---|---|---|---|---|---|---|
| Penicillium | M.P1 M.P2 |
Irregular Circular |
Flat Curly |
Rough smooth | Dark-green | Orange Gray |
|
Epicoccum |
M.9 M.15 M.7 M.4 M.5 M.11 |
Irregular | Filamentous Curly Curly Curly Corrugated Curly |
Rugged Corrugated Corrugated Rugged Corrugated Corrugated |
Gold-yellow Yellow Cream-white Light-beige Gold-yellow Brown |
Red-mahogany Yellow Cream-black Brown Red-mahogany Black |
| Trichoderma | M.T1 M.T2 M.T5 |
Irregular Irregular Irregular |
Efusa Curly Curly |
Concentric Concentric Concentric |
Dark-green White White |
Gold-yellow Off-white Lemon-yellow |
| Lasiodiplodia | M.1 M.2 |
Filamentous | Curly | Filamentous | Gray White |
Black Linen white |
| Fusarium | M.2, M.23 M.27 | Circular Irregular Circular |
Curly Curly Curly |
Rugged Rugged Rugged |
White Pink Yellow- white |
Black Brown and pink Yellow |
Table 3.
Morphological characterization of Fungi isolated of T. cacao fruits in the province of Orellana.
Table 3.
Morphological characterization of Fungi isolated of T. cacao fruits in the province of Orellana.
| Genera | Hyphal form | Conidia form | Conidial size |
|---|---|---|---|
| Penicillium | Septate | Globose and elipsoidal | 3.5 µm long x 3.5 µm wide |
|
Epicoccum |
Septates | Muriform | 12.5 – 15.25 µm long x 10 – 13,.5 µm wide |
| Trichoderma | Septates | Oval | 2.7 – 3.5 µm long x 2.7 - .2.8 µm wide |
| Lasiodiplodia | Septates | Oval-Subovoid and elipsoidal | 5.7 µm long x 2.68 µm wide |
| Fusarium | Septates | Half moon | 20 µm long x 5,1 µm wide |
Table 4.
Most abundant fungal genera present in T. cacao fruits, identified by morphology and morphological and molecular characterization in Orellana Province.
Table 4.
Most abundant fungal genera present in T. cacao fruits, identified by morphology and morphological and molecular characterization in Orellana Province.
| Morphotypes | Morphological/molecular identification | Percentage Identity (%) |
Size (pb) | Accesion | ||
|---|---|---|---|---|---|---|
| P1 P2 |
Peniciillium sp. | 98.41 99.72 |
378 356 |
MK805460.1 MT606192.1 |
||
| M.9 M.15 M.7 M.4 M.5 M.11 |
Epicoccum nigrum Epicoccum sp. Epicoccum sp. Epicoccum sp. Epicoccum sp. Epicoccum sp. |
100.00 99.79 100.00 99.00 99.26 98.90 |
362 470 450 517 403 364 |
MH397098.1 MG832438.1 MG832438.1 MG832438.1 MG832438.1 MG832438.1 |
||
| M.T1 M.T2 M.T5 |
Trichoderma sp. Trichoderma sp. Trichoderma sp |
100.00 100.00 100.00 |
389 501 575 |
KJ783298.1 ON705517.1 KJ783298.1 |
||
| M.1 | Lasiodiplodia sp | 99.84 | 382 | MN249167.1 | ||
| M.22 M.23 M.27 |
Fusarium sp. Fusarium sp. Fusarium sp. |
100.00 100.00 100.00 |
450 465 514 |
MN602837.1 MT603307 MN602837.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



