
Submitted:
26 November 2024
Posted:
27 November 2024
You are already at the latest version
Abstract
Fusarium circinatum (anamorph of Gibberella circinata) is the etiological agent of resinous canker in conifers, especially within the genus Pinus. This pathogen can impact all phases of tree development. Infections caused by F. circinatum in pine forests have been catastrophic in numerous countries globally, underscoring the necessity of understanding the geographical distribution of this fungus to enhance ecological comprehension and inform effective control measures. This study aims to report the existence of F. circinatum as a potential pathogen of pine trees in Honduras. Samples were obtained from mature trees and tree nurseries of Pinus oocarpa in three departments of Honduras, exhibiting symptoms such as chlorosis, needle wilting, and progressive mortality. Samples underwent microbiological and molecular identification. The isolates were identified using phenotypic approaches focused on micro and macro morphology, followed by the amplification and sequencing of three genes: ITS, EF-1α, and Beta-tubulin. Three isolates of the fungus were identified as F. circinatum. No genetic diversity was seen among the three isolates; nevertheless, some diversity was evident in comparison to sequences deposited in databases from different geographic regions. This is the first report and molecular characterization of F. circinatum isolated from Pinus oocarpa in Honduras.
Keywords:
Pinus oocarpa
; Fusarium circinatum
; Honduras
; forest health
1. Introduction
The genus Pinus, member of the Pinaceae family, encompasses approximately 100 species found across the northern hemisphere, including areas of Europe, Asia, North Africa, North America, and extending to Central America [1]. Pine forests in these regions serve crucial roles in local and global ecosystems, affecting meteorological events, controlling the hydrological cycle, safeguarding watersheds and their flora, and supporting a wide array of species across various domains [2,3]. Also, as one of the most significant timber trees globally, pines constitute a substantial source of financial resources for the economies of various countries, especially those classified as medium and low income [4,5]. Pine forests provide numerous benefits; however, they are subjected to various threats from both biotic and abiotic agents that compromise their health [6,7].
Fungi are significant biotic agents responsible for diseases in forests, compromising biodiversity and impacting aesthetic, commercial, and recreational values [8]. Pine infections have intensified recently due to climate change and human activities, resulting in a notable deterioration of global forest health [9,10].
Fungi are heterotrophic eukaryotes that play a crucial role in the composition of forest ecosystems [11,12]. Although certain fungal species contribute significantly to ecological balance, numerous others exert detrimental impacts on forest ecosystems [13]. Fusarium spp. represents a primary genus of endophytic fungi associated with pine species [13,14,15]. Key species linked to pine disease include Fusarium oxysporum, F. solani, and F. circinatum [16,17]. Fusarium spp. are primarily associated with diseases such as wilting and root rot in nurseries, leading to inadequate seedling establishment in external plantations [18]. This genus has also been associated with diseases in mature trees, including pine resin canker [18,19]. Resin canker is a significant disease impacting nurseries and pine plantations globally, leading to forest losses, hindering the establishment of pine trees across various ecosystems, and affecting national economies [20,21,22,23,24,25,26]. This pathogen has been demonstrated to impact all stages of pine development, including seeds, seedlings, and mature trees [27]. Advanced stages of the disease are characterized by the presence of resinous cankers on the main stem and branches, along with treetop dieback [27,28].
F. circinatum has been documented in 14 countries across 4 continents, with its presence on the American continent limited to 7 countries [29,30]. Honduras, situated in the central region of the Central American isthmus (15°00′N 86°30′W), encompasses an area of 112,777 km² and exhibits significant biological diversity, including seven species of pine, with Pinus oocarpa being the most notable due to its wide distribution and significant economic and environmental relevance [31,32]. Pine forests constitute 30.9% of the nation’s total forest cover, which spans 6,314,815 hectares, primarily located in the central, eastern, and western regions [32]. Recognized as part of the Mesoamerican biodiversity hotspot [33,34], Honduras emphasizes the safeguarding of its forest ecosystems, which are crucial for biodiversity and climate change mitigation, in accordance with Sustainable Development Goal (SDG) 15 [35].
This report details the presence of F. circinatum in Honduras, within the framework of the epidemiological surveillance program of the National Plan for Protection Against Forest Pests and Diseases, managed by the National Institute of Forest Conservation and Development, Protected Areas and Wildlife (ICF). This represents the first detection of F. circinatum in the country utilizing a molecular approach and aims to enhance the comprehension of infections induced by this pathogen in pine trees and to contribute to the global understanding of its effects.
2. Materials and Methods
2.1. Sample Collection
Samples were collected from pine trees exhibiting symptoms including needle chlorosis, root rot, and dieback as part of a national surveillance program. The samples were collected from three geographical regions in Honduras, the departments of Comayagua, Francisco Morazán, and El Paraíso. The affected plant tissues were dispatched to the Center for Research on Infectious and Zoonotic Agents (CIAIZ) at the National Autonomous University of Honduras for examination.
For the processing of the samples, the methodology previously described by Stone et al., 2004 was followed [36]. Briefly, some twigs were selected in which the transition zone between healthy and diseased plant tissue was identified. Three 1 cm samples were randomly selected from each sampling site. The exterior of the plant tissue was sterilized with 5% sodium hypochlorite solution for 3 min, then rinsed with sterile distilled water and immersed in 70% alcohol for 3 minutes, and then rinsed three times with sterile distilled water. Once the plant tissue was disinfected, it was cultured on PDA (Potato Dextrose Agar). The Petri dishes were incubated at 25 °C and observed daily for 10 days to detect the appearance of colonies, then all isolates were purified by hyphal tip or single spore method. Subsequently, with the aim of identifying the genus of the fungus, the isolates were evaluated microscopically, by staining cotton blue with lactophenol.
2.2. DNA Extraction
DNA extraction was performed following a previously described protocol [37]. Briefly, the mycelium of each strain was inoculated into 3 ml of GYEP broth (2% glucose, 0.3% yeast extract, 1% peptone). Then, all cultures were incubated for 7 days at 26°C. The mycelium mats were subsequently recovered and subjected to a protocol based on organic solvents. The mycelium was resuspended in 500 μL of lysis buffer composed of 10 mM Tris, pH 8; 1 mM EDTA, pH 8; and 100 mM NaCl. This suspension was heated at 100 °C for 1 min in a water bath and then shaken three times in the Disruptor Genie System (Scientific Industries, Inc. USA) with 0.1 mm glass beads for 2 min with rest intervals of 1 min between each shake. 400 μL of phenol-chloroform-isoamyl alcohol (25:24:1) was added, mixed by vortex, and centrifuged at 13,000 rpm for 15 min. The supernatant was recovered and one volume of chloroform-isoamyl alcohol (24:1) was added. The solution was mixed with vortex and centrifuged at 13,000 rpm for 5 min. The aqueous phase was transferred to a new vial and precipitated with one volume of cold absolute ethanol and 1/10 volume of sodium acetate (3 M, pH 5.2), mixed by inversion, and centrifuged at 13,000 rpm for 10 min. The supernatant was removed and subsequently washed with 70% ethanol. The dried pellet was resuspended in 50 μL of nuclease-free water. The DNA concentration was calculated with a NanoDrop spectrophotometer (Thermo Fisher Scientific Inc. Waltham, MA, USA), and diluted to a final concentration of 40 ng/μL and stored at −20◦C until further use.
2.3. Molecular Identification
The identification of all isolates was carried out through the amplification and sequencing of three polymorphic molecular markers: the internal transcribed spacer (ITS) of the nuclear ribosomal DNA gene, the elongation factor 1 alpha (EF-1α), and the beta tubulin (BT) gene, using the primers proposed by White et al. (1990) [38], O’Donnell et al., 1998 [39], and Glass et al., 1995 [40] respectively.
2.3.1. Amplification of ITS Regions
The amplifications were carried out in a 50 μL volume: 25 μL of PCR Master Mix (Promega Corp. Madison, WI, USA), 1 μL of 10 μM ITS1 and ITS4 primers: 5′-TCC GTA GGT GAA CCT GCG G -3′ / 5′-TCC TCC GCT TAT TGA TAT GC—3′, and 1 μL of DNA (40 ng/ μL). Reactions were carried out with an initial denaturation step at 95 °C for 5 min, 37 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s, with a final extension at 72 °C for 10 min.
2.3.2. Amplification of the Elongation Factor 1 Alpha
Amplifications were performed in a volume of 50 μL: 25 PCR Master Mix (PromegaCorp. Madison, WI, USA), using 1 μL of 10 μM primers EF1 and EF2: 5′-ATG GGT AAG GAR GAC AAG AC -3′ / 5′- GGA RGT ACC AGT SAT CAT GTT—3′ and 1 μL of DNA (40 ng/μL). The reactions were carried out with an initial denaturation step at 95 °C for 5 min, 35 cycles of 95 °C for 1 min 30s, 52 °C for 1 min and 72 °C for 2 min, with a final extension at 72 °C for 10 min.
2.3.3. Amplification of Beta Tubulin
Likewise, for partial amplification of the BT gene, 25 μL of Master Mix (Promega Corp. Madison, WI, USA) and 1 μL of the 10 μM primers Bt1a’ and Bt1b: 5’- TTC CCC CGT CTC CAC TTC TTC ATG—3 / 5’- GAC GAG ATC GTT CAT GTT GAA CTC -3’. The amplification conditions were as follows. An initial denaturation at 95 °C for 5 min, 35 cycles of 95 °C for 1 min, 56 °C for 1 min and 72 °C for 1 min, with a final extension at 72 °C for 5 min. All amplicons were visualized in 1.5% agarose gel electrophoresis with ethidium bromide.
2.4. Sequencing
The amplified products were sequenced on both flanks with their same primers used for the PCR reactions, and according to Psomagen company protocols (https://lims.psomagen.com/). The quality of the sequences was analyzed in the Geneious® software v. 2023.1.2. The sequences were curated and then aligned to search for nucleotide polymorphisms (SNP) among the isolates. Subsequently, the sequences were imported and analyzed using the BLAST tool in the NCBI database (https://www.ncbi.nlm.nih.gov/), where they were compared with the available sequences and subsequently the highest similarity index was recorded.
To elucidate the phylogeny of the isolates, 30 homologous sequences corresponding to the ITS marker and 29 sequences of the EF-1α gene were retrieved from GenBank, based on reports of F. circinatum isolated from various geographic regions worldwide. The downloaded sequences and those obtained in this study were aligned using the Geneious alignment tool of the Geneious software. The aligned sequences were edited to produce a homogeneous length of 461 and 471 bp for the ITS and BT regions respectively. Similarly, the EF-1α marker sequences were delimited to a length of 600 bp. With the alignments, an unrooted dendrogram for EF-1α was constructed using the Neighbor-Joining algorithm and Tamura Nei’s genetic distance model with 1000 Bootstrap replicates with a homologous sequence from Fusarium oxysporum as outgroup.
3. Results
This study evaluated samples of Pinus oocarpa specimens exhibiting symptoms compatible with resinous canker, and collected from three departments in central Honduras, as illustrated in Figure 1. Samples were collected from nursery seedlings in the departments of Francisco Morazán and El Paraíso while madure pine samples were collected from natural forests in the Comayagua department. Samples were collected between July and September 2023, coinciding with the rainy season in Honduras. Table 1 presents the sampling sites along with the observed symptoms and signs.
3.1. Isolation and Phenotypic Identification
Following a 10-day incubation period in PDA at 25 °C, colonies exhibiting a velvety white to purple appearance were noted on the obverse (Figure 2A), accompanied by the production of purple pigment on the reverse of the plates. Staining with blue lactophenol revealed an abundance of fusiform macroconidia (Figure 2B). The fungus was morphologically identified as Fusarium spp. based on the taxonomic keys established by O’Donnell et al., 2000 [41].
3.2. Molecular Identification of the Isolates
The isolates were identified at the species level through the amplification and sequencing of three markers (ITS, EF-1α, and BT). The PCR products for EF-1α showed a length of 710 bp, while the lengths for the ITS and BT regions were 570 bp and 530 bp, respectively (Figure 3A). The sequences were aligned, revealing no size or sequence polymorphisms (SNP) in any of the markers across the three isolates examined in this study (Figure 3B). In contrast, a comparison of the sequences obtained in this study with those previously deposited in GenBank for both ITS and EF-1α revealed that the three isolates were identified as Fusarium circinatum, exhibiting 100% similarity for each gene. The lack of annotated sequences for the BT gene in GenBank hindered the verification of this fungal identification using this molecular marker.
A phylogenetic tree was constructed by comparing the EF-1α sequences obtained in this study with 29 sequences of F. circinatum from various geographical regions worldwide. The isolates derived from F. circinatum in Honduras demonstrated significant similarity with most previously documented sequences for each molecular marker. Three clusters were identified: the first cluster comprised two sequences from Korea, exhibiting a bootstrap value of 99.57. The second cluster, with a bootstrap value of 81.99, included nine sequences from North America, Japan, Brazil, Guatemala, and South Africa. The third cluster contained the largest number of sequences, incorporating sequences from Honduras and 18 sequences from North America, Nicaragua, Colombia, the Mediterranean, India, and South Africa (Figure 4). Thus, clustering based on the geographical origin of the isolates of this species does not seem to be evident when analyzed using the EF-1α marker.
In addition, the comparison of the three ITS sequences obtained in this study with 30 sequences from various geographical regions indicated that the F. circinatum isolates from Honduras exhibited high similarity, clustering into a homogeneous group with the other sequences. Thus, no phylogenetic clustering pattern based on geographic origin was identified for ITS (Supplementary Material 1). A sequence for each of the evaluated molecular markers was deposited in GenBank, with the following accession numbers: ITS (PP453646.1), EF-1α (PQ368253), and BT (PQ368252).
4. Discussion
Resin canker, caused by F. circinatum, is recognized as one of the most detrimental diseases affecting Pinus spp. in various countries globally [42]. The primary mode of propagation for this fungus is through seeds and seedlings, which has been suggested to facilitate its introduction into new countries [27,42,43,44]. In addition, the spread of F. circinatun is also usually facilitated by wind, fires and insects [27,42,44,45,46,47], particularly when plants present wounds or suffer water stress, which in turn coincide with optimal environmental conditions such as humidity and temperature that favor the colonization of the fungus [27,42].
The first report of F. circinatum as a pine pathogen was made in North Carolina in the United States [48]. Subsequently, this fungus has been documented in 14 countries across 4 continents, with no reports of its presence in Oceania to date [29,30]. The pathogen has been reported in France, Italy, Portugal, Spain, Japan, South Korea, and South Africa [29,30]. In the Americas, it has been reported in the USA, Mexico, Colombia, Brazil, Haiti, Chile, and Uruguay, with the USA exhibiting the highest incidence globally [29,30]. Figure 5 illustrates the global distribution of F. circinatum, utilizing data from the EPPO Global Database along with the analyses conducted by Drenkhan et al. (2020) and Vettraino et al. (2018) [29,30,42].
Currently, there are no documented reports of F. circinatum in Honduras. Our findings indicate that two accession numbers, KX364413.1 and KX364414.1, are registered in GenBank for the EF-1α gene from isolates of F. circinatum in Nicaragua and Guatemala. These records, however, are deficient in epidemiological and ecological information, including the exact location of isolation and the species of conifer from which they were derived. Consequently, this would be the first formal report of the presence of this fungus in Central America.
This fungus has been documented in 24 species of pine, with P. radiata, P. taeda, P. echinata, and P. patula being the most affected species [29]. To our knowledge, this would be the first report of F. circinatum isolated from P. oocarpa. Dvorak et al. (2009) [49] highlighted the significant resistance levels of P. oocarpa to F. circinatum. Consequently, our findings prompt several inquiries regarding the presence of this pathogen in the pine species and the associated symptoms and signs observed. Therefore, additional research is required to elucidate the development of infections in P. oocarpa caused by F. circinatum and to identify the factors that facilitate the establishment of this fungus in the conifer species.
A hypothesis regarding the presence of F. circinatum as a potential infectious agent in P. oocarpa in Honduras suggests that climate change and human activities contribute to rising global temperatures [50,51,52]. This increase affects drought frequency and water availability, resulting in forest stress and heightened susceptibility to infections [53,54]. McDonald et al. (1994) demonstrated a positive correlation between the distribution, establishment, and development of F. circinatum and elevated temperature and humidity. Temperatures ranging from 14 to 26 °C promote the growth of this fungus, whereas temperatures at or below 10 °C inhibit its development [55].
In Honduras, as in other regions globally, temperatures have risen during this century [54], maintaining an average temperature of 32 °C throughout the national territory [56], which could have favored the adaptation and development of F. circinatum in this pine species. The rise in average temperatures, coupled with the epidemic caused by the beetle Dendroctonus frontalis from 2015 to 2017, resulted in a loss of approximately 23% of the forest cover in Honduras’ pine forests [52,57,58], whose outbreaks have coincided with long periods of drought, which are factors that are strongly associated with infections by F. circinatum [59,60]. Another argument that could explain the presence of this pathogen at least in the mature forest is that this and other microorganisms could act as secondary pathogens in forests previously attacked by D. frontalis.
Furthermore, anthropogenic factors, especially forest fires, contribute to forest stress, resulting in wildlife and biodiversity loss. This also leads to soil weakening and erosion, habitat fragmentation, and a reduction in forest cover, which further diminishes trees’ resistance to biotic and abiotic pests [61]. Likewise, it has been shown that smoke from forest fires could act as a vector of fungal propagules [46,47]. Between 2023 and 2024, Honduras has averaged 2,800 fires, predominantly located in the central eastern region of the country. This phenomenon may influence the prevalence of this and other pathogens in Honduras’ forests [62].
Resin canker caused by Fusarium circinatum has been recognized as one of the most devastating diseases in Pinus spp. worldwide [42]. The European Union has designated this pathogen as a quarantine organism, categorizing it on list A2 of the EPPO (European and Mediterranean Plant Protection Organization) [42]. England has also developed a contingency plan to address potential outbreaks of F. circinatum in its coniferous forests [63]. The International Plant Protection Convention (IPPC), through its protocol DP:22 Fusarium circinatum, establishes a standard protocol for the detection and diagnosis of this pathogen in phytopathology laboratories and crop inspection systems. It emphasizes that molecular-level EF-1α sequencing is adequate for confirming the identification of this fungus [64].
Honduras currently lacks an action plan or specific legislation to prevent or manage outbreaks of F. circinatum, despite the associated risks to the nation’s coniferous ecosystems and forest economy. The lack of concrete measures underscores the necessity of developing national strategies that encompass diagnosis, monitoring, and control protocols to prevent the introduction and/or dissemination of F. circinatum to new locations or pine species within the country. By demonstrating the presence of F. circinatum in Honduras, this information aims to assist decision makers in identifying areas requiring prompt intervention and to guide future efforts in enhancing the understanding of this fungus within the country.
5. Conclusions
F. circinatum is a virulent pathogen of pine, linked to seedling wilt and resinous canker in mature trees. This is the first report of F. circumatum isolated from P. oocarpa in Honduras. This description will enhance the understanding of infections in pine trees caused by this pathogen. The molecular approach employed in this study, encompassing the amplification and sequencing of the ITS, EF-1α, and BT genetic regions, enabled precise confirmation of the pathogen’s identification. This underscores the significance of these tools for early and reliable diagnosis in future research and management strategies within the region.
6. Limitations
This study did not include infection assays due to the necessity for controlled conditions, appropriate physical space, and specialized equipment. While assessing the pathogenic potential of F. circinatum would have been advantageous, our emphasis was on detecting and confirming the fungus’s presence in Honduras. This focus remains pertinent, as our findings enhance the understanding of the epidemiology of this fungus globally.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Alignment of 30 Fusarium circinatum ITS sequences from various regions revealed no polymorphisms when compared to three sequences from Honduras.
Author Contributions
B.O, K.M, L.E conceptualized the study; Y.Y-E, K.A-F, I.U-V; obtained the samples, B.O, K.M, K.A performed the mycological diagnosis, B.O, K.M, K.A performed the experiments, B.O, K.M, K.A organized and cured the data; B.O, G.F writing and original draft preparation, All the authors contributed with writing, review, and editing the manuscript; supervision, project administration, and funding acquisition, B.O, K.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Banco Interamericano de Desarrollo, Instituto Nacional de Conservación Forestal, Areas Protegidas y Vida Silvestre (ICF), grant number BID 5878/BL-HO” and “The APC was funded by Dirección de Investigación Científica, Humanística y Tecnológica (DICIHT), Universidad Nacional Autonoma de Honduras”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Burley, J.; Barnes, R.D. TROPICAL ECOSYSTEMS | Tropical Pine Ecosystems and Genetic Resources. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Oxford, 2004; pp. 1728–1740. [Google Scholar]
- Food and Agriculture Organization of the United Nations FAO. El estado de los bosques del mundo 2020, Los bosques, la biodiversidad y las personas, Roma. 2020. [CrossRef]
- Thompson, I.; Mackey, B.; McNulty, S.; Mosseler, A. Forest resilience, biodiversity, and climate change. In Proceedings of the Secretariat of the Convention on Biological Diversity, Montreal. Technical Series no. 43. 1-67., 2009; pp. 1-67.
- Küçüker, D.; Baskent, E. State of stone pine (Pinus pinea) forests in Turkey and their economic importance for rural development. Mediterranean pine nuts from forests and plantations 2017, 122, 111–117. [Google Scholar]
- Moctezuma López, G.; Flores, A. Economic importance of pine (Pinus spp.) as a natural resource in Mexico. Revista mexicana de ciencias forestales 2020, 11, 161–185. [Google Scholar] [CrossRef]
- Muñoz, J. Regeneración Natural: Una revisión de los aspectos ecológicos en el bosque tropical de montaña del sur del Ecuador. Bosques Latitud Cero 2017, 7. [Google Scholar]
- Gea-Izquierdo, G.; Férriz, M.; García-Garrido, S.; Aguín, O.; Elvira-Recuenco, M.; Hernandez-Escribano, L.; Martin-Benito, D.; Raposo, R. Synergistic abiotic and biotic stressors explain widespread decline of Pinus pinaster in a mixed forest. Science of the Total Environment 2019, 685, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Dussi, M.C.; Flores, L.B. Visión multidimensional de la agroecología como estrategia ante el cambio climático. Inter disciplina 2018, 6, 129–153. [Google Scholar] [CrossRef]
- Armenteras, D.; González, T.M.; Vargas Ríos, O.; Meza Elizalde, M.C.; Oliveras, I. Incendios en ecosistemas del norte de Suramérica: avances en la ecología del fuego tropical en Colombia, Ecuador y Perú. Caldasia 2020, 42, 1–16. [Google Scholar] [CrossRef]
- Callan, B. Introduction to forest diseases. 2001.
- Ghelardini, L.; Pepori, A.L.; Luchi, N.; Capretti, P.; Santini, A. Drivers of emerging fungal diseases of forest trees. Forest Ecology and Management 2016, 381, 235–246. [Google Scholar] [CrossRef]
- Gomdola, D.; Bhunjun, C.; Hyde, K.; Jeewon, R.; Pem, D.; Jayawardena, R. Ten important forest fungal pathogens: a review on their emergence and biology. 2022. [CrossRef]
- Rashmi, M.; Kushveer, J.; Sarma, V. A worldwide list of endophytic fungi with notes on ecology and diversity. Mycosphere 2019, 10, 798–1079. [Google Scholar] [CrossRef]
- Wang, L.; Ren, L.; Li, C.; Gao, C.; Liu, X.; Wang, M.; Luo, Y. Effects of endophytic fungi diversity in different coniferous species on the colonization of Sirex noctilio (Hymenoptera: Siricidae). Scientific reports 2019, 9, 5077. [Google Scholar] [CrossRef]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annual review of phytopathology 1996, 34, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, R.S.; Hyde, K.D.; de Farias, A.R.G.; Bhunjun, C.S.; Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Herath, I.S.; Thambugala, K.M.; Manawasinghe, I.S. What is a species in fungal plant pathogens? Fungal Diversity 2021, 109, 239–266. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 fungal pathogens in molecular plant pathology. Molecular plant pathology 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Akgül, D.S.; Önder, S.; Savaş, N.G.; Yıldız, M.; Bülbül, İ.; Özarslandan, M. Molecular Identification and Pathogenicity of Fusarium Species Associated with Wood Canker, Root and Basal Rot in Turkish Grapevine Nurseries. Journal of Fungi 2024, 10, 444. [Google Scholar] [CrossRef] [PubMed]
- Dwinell, L.D.; Barrows-Broaddus, J.B.; Kuhlman, E.G. Pitch canker: a disease complex. Plant Dis 1985, 69, 270–276. [Google Scholar] [CrossRef]
- Kuhlman, E.; Dianis, S.; Smith, T. Epidemiology of pitch canker disease in a loblolly pine seed orchard in North Carolina. Phytopathology 1982, 72, 1212–1216. [Google Scholar] [CrossRef]
- McCain, A.; Koehler, C.; Tjosvold, S. Pitch canker threatens California pines. California agriculture 1987, 41, 22–23. [Google Scholar]
- Muramoto, M.; Dwinell, L. Pitch canker of Pinus luchuensis in Japan. 1990.
- Guerra-Santos, J. Pitch canker on Monterey pine in Mexico. Current and potencial impacts of pitch canker in radiata pine. In Proceedings of the Proceedings of the Impact Monterey Workshop. Monterey, California, CSIRO, Collingwood, Victoria, Australia, 1999.
- Viljoen, A.; Marasas, W.; Wingfield, M.; Viljoen, C. Characterization of Fusarium subglutinans f. sp. pini causing root disease of Pinus patula seedlings in South Africa. Mycological Research 1997, 101, 437–445. [Google Scholar] [CrossRef]
- Landeras, E.; García, P.; Fernández, Y.; Braña, M.; Fernández-Alonso, O.; Méndez-Lodos, S.; Pérez-Sierra, A.; León, M.; Abad-Campos, P.; Berbegal, M. Outbreak of pitch canker caused by Fusarium circinatum on Pinus spp. in northern Spain. Plant Disease 2005, 89, 1015. [Google Scholar] [CrossRef]
- Wingfield, M.; Hammerbacher, A.; Ganley, R.; Steenkamp, E.; Gordon, T.; Wingfield, B.; Coutinho, T. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australasian Plant Pathology 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Storer, A.J.; Wood, D.L.; Gordon, T.R. The epidemiology of pitch canker of Monterey pine in California. Forest Science 2002, 48, 694–700. [Google Scholar] [CrossRef]
- Drenkhan, R.; Ganley, B.; Martín-García, J.; Vahalík, P.; Adamson, K.; Adamčíková, K.; Ahumada, R.; Blank, L.; Bragança, H.; Capretti, P. Global geographic distribution and host range of Fusarium circinatum, the causal agent of pine pitch canker. Forests 2020, 11, 724. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization (EPPO). Distribution Fusarium circinatum. Available online: https://gd.eppo.int/taxon/GIBBCI/distribution (accessed on October 28).
- Organización de las Naciones Unidas para la Alimentación y la Agricultura. Dirección de recursos forestales. Estado de la diversidad biológica de los árboles y bosques de Honduras. Available online: https://www.fao.org/4/j0607s/j0607s00.htm#TopOfPage (accessed on 27 September 2024).
- Instituto de Conservación Forestal. Anuario Estadistico Forestal de Honduras, 3era edición. Centro de Información y Patrimonio Forestal, Unidad de Estadisticas Forestales. Available online: https://icf.gob.hn/?portfolio=cipf-2 (accessed on 27 September 2024).
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Coates, A.G. Paseo Pantera: una historia de la naturaleza y cultura de Centroamérica. (No Title) 2003.
- Naciones Unidas. La Agenda 2030 y los Objetivos de Desarrollo Sostenible: una oportunidad para América Latina y el Caribe (LC/G.2681-P/Rev.3), Santiago. Available online: https://repositorio.cepal.org/server/api/core/bitstreams/cb30a4de-7d87-4e79-8e7a-ad5279038718/content (accessed on 27 September 2024).
- Stone, J.K.; Polishook, J.D.; White, J.F. Endophytic fungi. Biodiversity of fungi: inventory and monitoring methods 2004, 241, 270. [Google Scholar]
- Ortiz, B.; Enríquez, L.; Mejía, K.; Yanez, Y.; Sorto, Y.; Guzman, S.; Aguilar, K.; Fontecha, G. Molecular characterization of endophytic fungi from pine (Pinus oocarpa) in Honduras. Revis Bionatura 2022; 7 (3) 13. 2022. [CrossRef]
- Tj, W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols: a guide to methods and applications 1990.
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Sciences 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Applied and environmental microbiology 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Nirenberg, H.I.; O’Donnell, K. New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia 1998, 90, 434–458. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Potting, R.; Raposo, R. EU legislation on forest plant health: An overview with a focus on Fusarium circinatum. Forests 2018, 9, 568. [Google Scholar] [CrossRef]
- Evira-Recuenco, M.; Iturritxa, E.; Raposo, R. Impact of seed transmission on the infection and development of pitch canker disease in Pinus radiata. Forests 2015, 6, 3353–3368. [Google Scholar] [CrossRef]
- Serrano, Y.; Iturritxa, E.; Elvira-Recuenco, M.; Raposo, R. Survival of Fusarium circinatum in soil and Pinus radiata needle and branch segments. Plant Pathology 2017, 66, 934–940. [Google Scholar] [CrossRef]
- Dvořák, M.; Janoš, P.; Botella, L.; Rotková, G.; Zas, R. Spore dispersal patterns of Fusarium circinatum on an infested Monterey pine forest in North-Western Spain. Forests 2017, 8, 432. [Google Scholar] [CrossRef]
- Camacho, I.; Góis, A.; Camacho, R.; Nóbrega, V.; Fernandez. The impact of urban and forest fires on the airborne fungal spore aerobiology. Aerobiologia 2018, 34, 585–592. [Google Scholar] [CrossRef]
- Kobziar, L.N.; Thompson III, G.R. Wildfire smoke, a potential infectious agent. Science 2020, 370, 1408–1410. [Google Scholar] [CrossRef]
- Hepting, G.H.; Roth, E.R. Pitch canker, a new disease of some southern pines. 1946.
- Dvorak, W.; Potter, K.; Hipkins, V.; Hodge, G. Genetic diversity and gene exchange in Pinus oocarpa, a Mesoamerican pine with resistance to the pitch canker fungus (Fusarium circinatum). International Journal of Plant Sciences 2009, 170, 609–626. [Google Scholar] [CrossRef]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nature Reviews Microbiology 2023, 21, 640–656. [Google Scholar] [CrossRef] [PubMed]
- Park Williams, A.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D. Temperature as a potent driver of regional forest drought stress and tree mortality. Nature climate change 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Rojas, M.R.; Bocatelli, B.; Billings, R. Cambio climático y eventos epidémicos del gorgojo descortezador del pino” Dendroctonus frontalis” en Honduras. Forest systems 2010, 19, 70–76. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest ecology and management 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Hansen, J.; Sato, M.; Ruedy, R.; Lo, K.; Lea, D.W.; Medina-Elizade, M. Global temperature change. Proceedings of the National Academy of Sciences 2006, 103, 14288–14293. [Google Scholar] [CrossRef]
- McDonald, M.J. Temperature: effects on Fusarium subglutinans f. sp. pini infection on juvenile Pinus radiata (Monterey pine) and influence on growth of Fusarium subglutinans f. sp. pini isolates from California and Florida; San Jose State University: 1994.
- Instituto Nacional de Estadística de Honduras. Boletín temperatura atmosférica 2017–2021. Available online: https://ine.gob.hn/v4/wp-content/uploads/2023/07/Temperatura-2017-2021.pdf (accessed on 25 September 2024).
- Instituto Nacional de Conservación Forestal. Areas Protegidas y Vida Silvestre. (ICF). Norma técnica para el manejo de insectos descortezadores del pino (Dendroctonus spp. e Ips spp.). Available online: https://icf.gob.hn/wp-content/uploads/2024/05/Norma-T%E2%80%9Acnica-MIDP_-1.pdf (accessed on 24 September 2024).
- Instituto Nacional de Conservación Forestal. Areas Protegidas y Vida Silvestre (ICF). Informe de episodio de ataque del gorgojo descortezador del pino dendroctonus frontalis en Honduras 2014-2017. Available online: https://icf.gob.hn/wp-content/uploads/2024/05/Informe-Episodio-de-Plaga-2014-2017.pdf (accessed on 25 September 2024).
- Billings, R.; Clarke, S.; Espino-Mendoza, V.; Cordón Cabrera, P.; Meléndez Figueroa, B.; Ramón Campos, J.; Baeza, G. Bark beetle outbreaks and fire: a devastating combination for Central America’s pine forests. Unasylva 2004, 55, 15–21. [Google Scholar]
- Singh, V.V.; Naseer, A.; Mogilicherla, K.; Trubin, A.; Zabihi, K.; Roy, A.; Jakuš, R.; Erbilgin, N. Understanding bark beetle outbreaks: exploring the impact of changing temperature regimes, droughts, forest structure, and prospects for future forest pest management. Reviews in Environmental Science and Bio/Technology 2024, 1-34. [CrossRef]
- Gajendiran, K.; Kandasamy, S.; Narayanan, M. Influences of wildfire on the forest ecosystem and climate change: A comprehensive study. Environmental Research 2024, 240, 117537. [Google Scholar] [CrossRef] [PubMed]
- Instituto de Conservación y Desarrollo Forestal Áreas Protegidas y Vida Silvestre (ICF Honduras). Estadisticas Incendios Forestales Reportados en Honduras. Available online: https://sigmof.icf.gob.hn/reportes/incendios-forestales-2/ (accessed on 17 November 2024).
- Forestry Commission. Contingency plan pitch canker of pine. Available online: https://cdn.forestresearch.gov.uk/2022/02/contingency-plan-pitch-canker-of-pine-published-_sept-05-2016.pdf (accessed on 17 November 2024).
- International Plant Protection Convention (IPPC). Diagnostic protocols for regulated pests, DP 22: Fusarium circinatum. Available online: https://openknowledge.fao.org/server/api/core/bitstreams/30e5bc8f-4c8e-491d-9fc6-7e1b72a5dbd0/content (accessed on 17 November 2024).
Figure 1.
A. Map of Honduras showing sample collection sites in three departments. B. Flagging of branches on adult trees of P. oocarpa. C. Chlorosis observed in Pinus oocarpa seedlings in the nursery. D. A seedling of P. oocarpa with wilting and dieback symptoms.
Figure 1.
A. Map of Honduras showing sample collection sites in three departments. B. Flagging of branches on adult trees of P. oocarpa. C. Chlorosis observed in Pinus oocarpa seedlings in the nursery. D. A seedling of P. oocarpa with wilting and dieback symptoms.
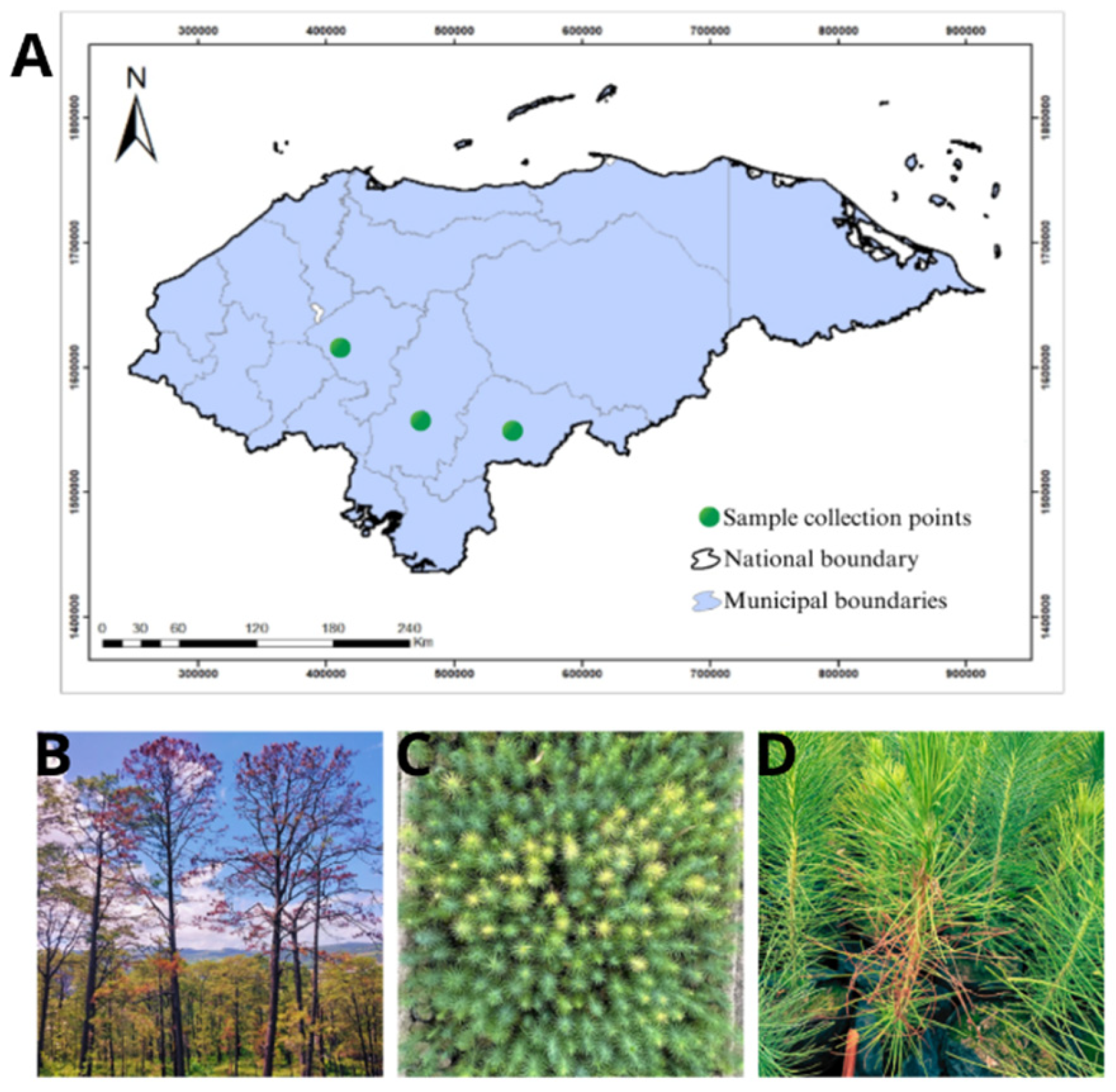
Figure 2.
A. Fungal growth from pine needles B. Lactophenol cotton blue stain showing fusiform macroconidia, suggestive of Fusarium spp.
Figure 2.
A. Fungal growth from pine needles B. Lactophenol cotton blue stain showing fusiform macroconidia, suggestive of Fusarium spp.
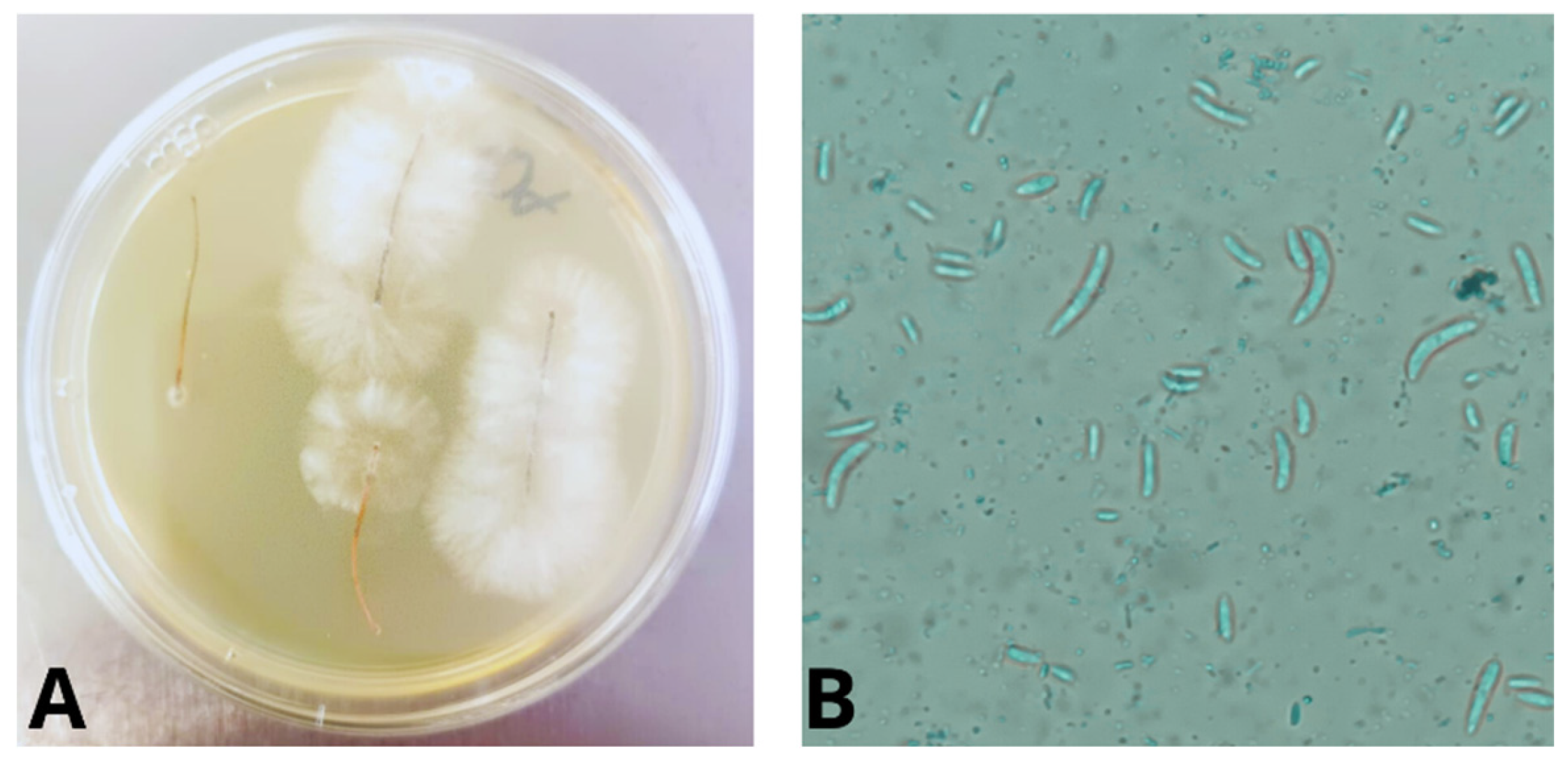
Figure 3.
A. Agarose gel electrophoresis. Lanes 1-3 show the amplification products of the internal transcribed spacer (ITS) of the ribosomal region of F. circinatum; lanes 5-7 show the amplification products of the elongation factor 1-alpha (EF-1α); lanes 9- 11 show the amplification products of β-tubulin gene. (C -) Negative control. B-D. Alignment of the three sequences for ITS, EF-1α, and β-tubulin obtained in this study showing the absence of size or sequence polymorphism.
Figure 3.
A. Agarose gel electrophoresis. Lanes 1-3 show the amplification products of the internal transcribed spacer (ITS) of the ribosomal region of F. circinatum; lanes 5-7 show the amplification products of the elongation factor 1-alpha (EF-1α); lanes 9- 11 show the amplification products of β-tubulin gene. (C -) Negative control. B-D. Alignment of the three sequences for ITS, EF-1α, and β-tubulin obtained in this study showing the absence of size or sequence polymorphism.
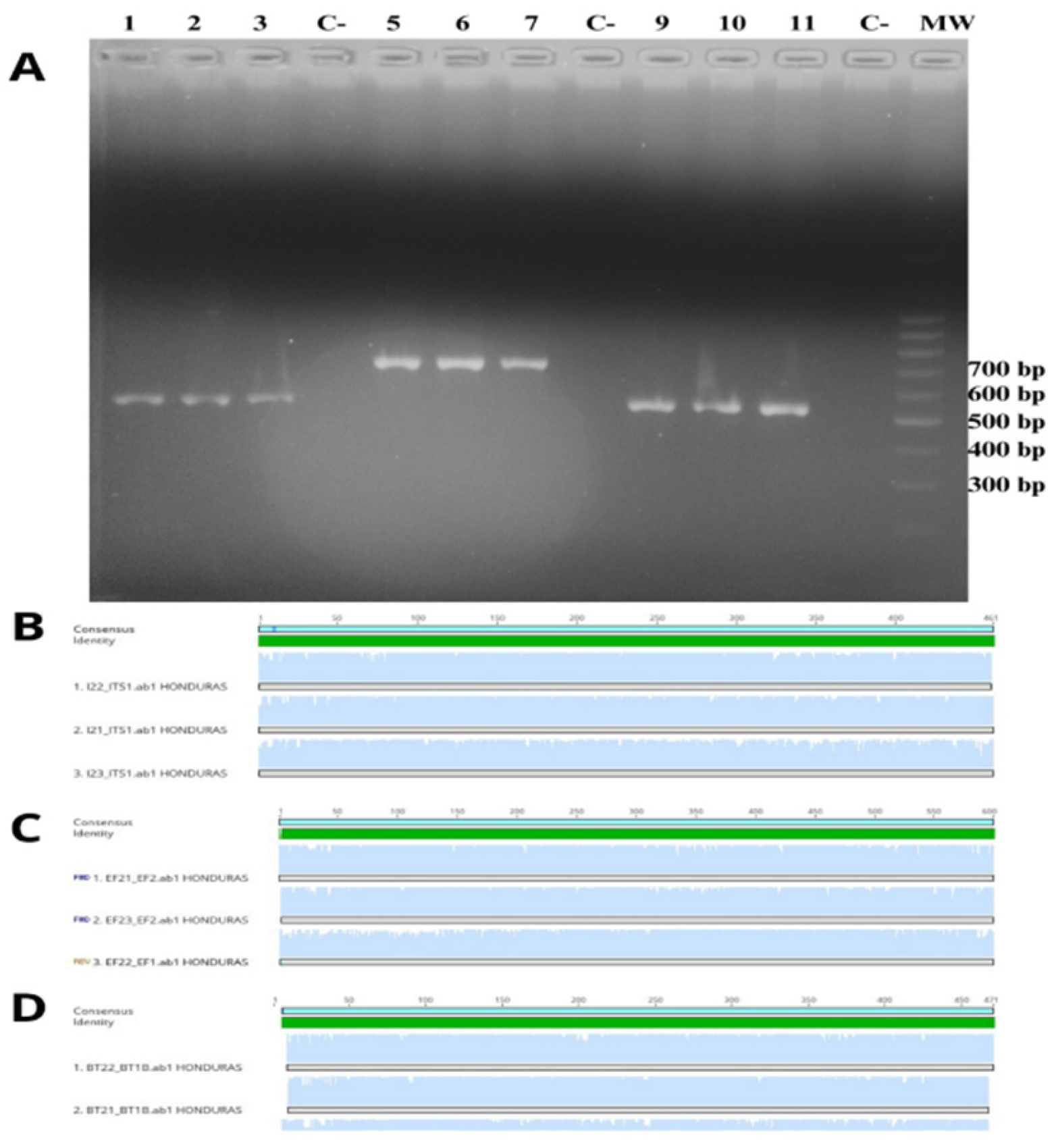
Figure 4.
Phylogenetic analysis of F. circinatum utilizing EF-1α from isolates across various geographical regions. The Tamura–Nei genetic distance model and the Neighbor-Joining method, utilizing a bootstrap of 1000 replicates, were employed to construct a cladogram. The sequences obtained in this study are presented within the light blue box.
Figure 4.
Phylogenetic analysis of F. circinatum utilizing EF-1α from isolates across various geographical regions. The Tamura–Nei genetic distance model and the Neighbor-Joining method, utilizing a bootstrap of 1000 replicates, were employed to construct a cladogram. The sequences obtained in this study are presented within the light blue box.
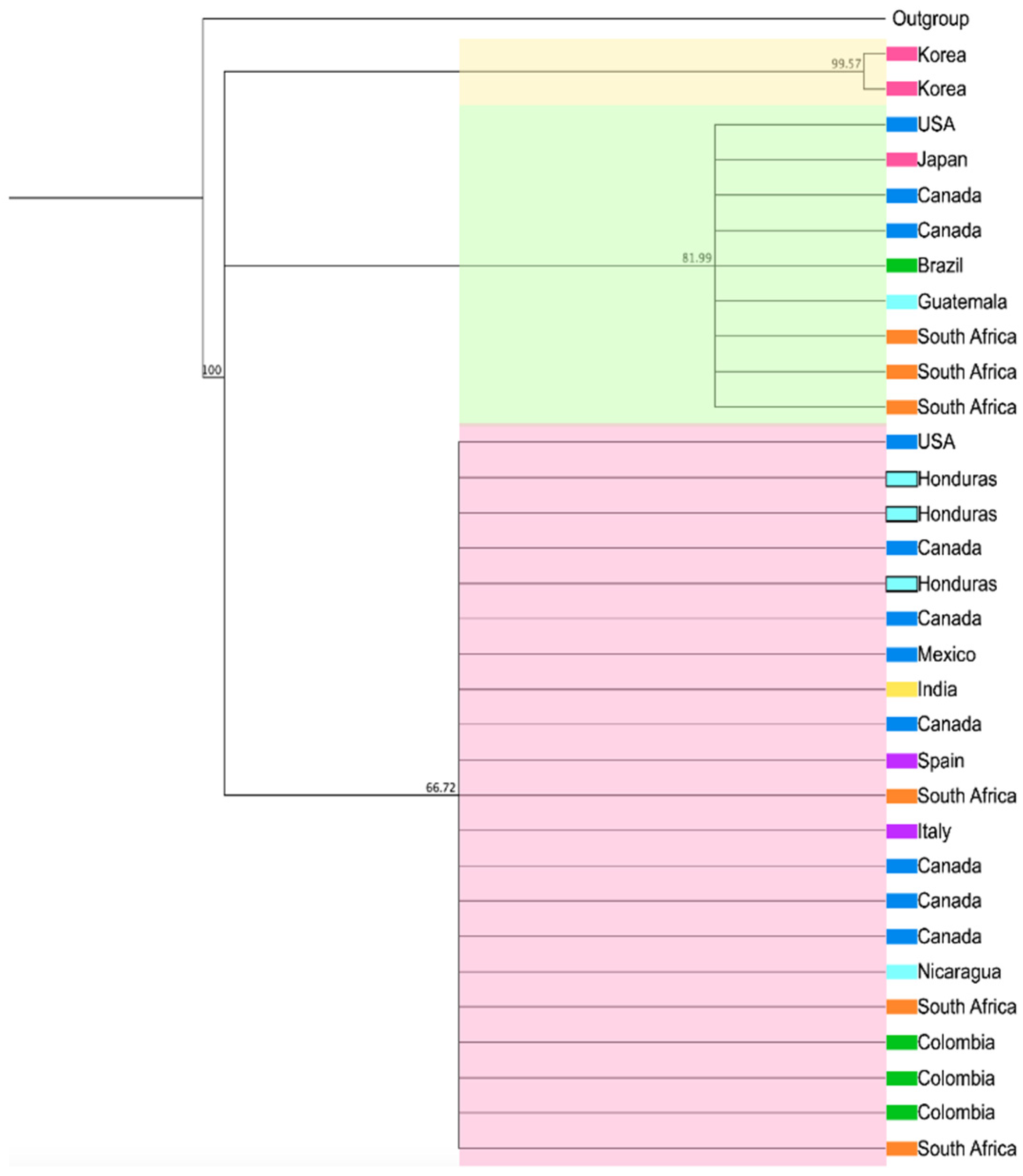
Figure 5.
World map of the current distribution of Fusarium circinatum represented in yellow.
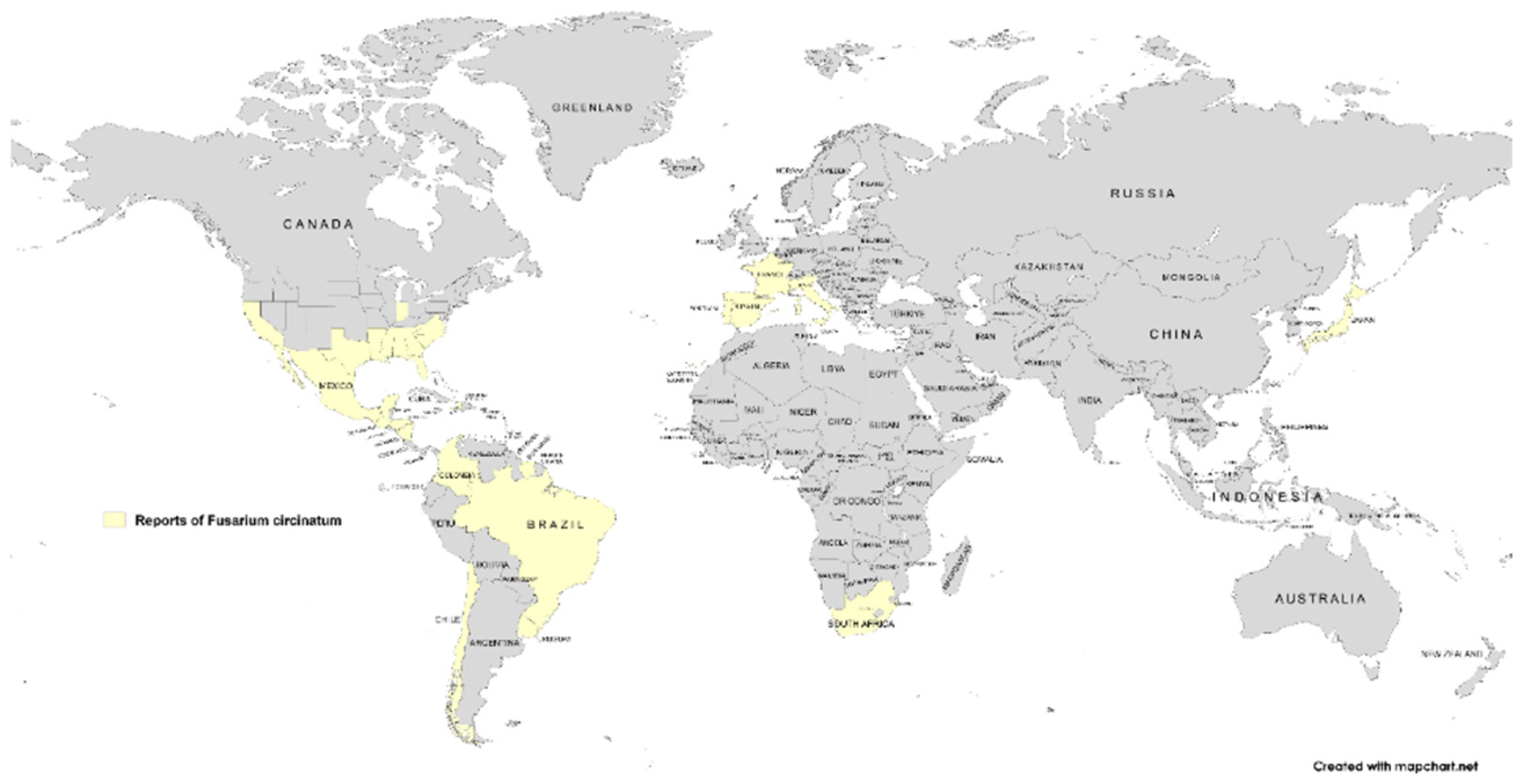

Table 1.
Details regarding the collection sites and the symptoms noted in the plant material.
| Code | Collection season | Department | Collection coordinates | Sample | Symptoms |
|---|---|---|---|---|---|
| 01-2023 | June 20 | Comayagua | X: 409008; Y: 1614567; A: 1133 | Tree in natural forest | Needles with chlorosis, treetop dieback, resin exudation |
| 02-2023 | July 18 | Francisco Morazán | X: 475338; Y: 1559190; A:1082 | Nursery seedling | Needles with yellow and reddish color |
| 09-2023 2 | September 19 | El Paraíso | X: 545270; Y: 1550795; A: 745 | Nursery seedling | Lack of growth, discoloration and wilting in the needles |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



