
Submitted:
28 November 2024
Posted:
28 November 2024
You are already at the latest version
Abstract
Actinomycetes are a group of bacteria that have been reported as potential controllers of several plant pathogens and as plant growth promoting agents. This study focused on the isolation of 60 actinobacterial strains from the Cuatro Cienegas Basin, Coahuila, Mexico, with an emphasis on evaluating their potential as biocontrol agents against plant pathogens. Among the strains tested, the isolate PR69 exhibited significant in vitro antagonistic activity against eight plant pathogens, with inhibition rates ranging from 44% to 73%, including Phytophthora capsici. The genome of PR69 was sequenced and assembled, confirming its classification as a member of the genus Streptomyces through a phylogenetic analysis based on concatenated multiple sequence alignments of 81 core bacterial genes. Additionally, volatile compounds produced by PR69 enhanced the growth of Arabidopsis thaliana seedlings in vitro, increasing seedling weight, primary root length, and the number of secondary roots. Furthermore, the soil treated with Streptomyces sp. PR69 effectively controlled the infection caused by the pathogen P. capsici in bell pepper plants, reducing disease symptoms by 50% compared to plants inoculated solely with the pathogen. PR69-treated plants also showed an increase in fresh weight compared to untreated controls. These findings suggest that Streptomyces sp. PR69 holds promise as a bioinoculant for promoting pepper plant growth and controlling P. capsici populations.
Keywords:
Antifungal activity
; Phytopathogen
; Biocontrol
; Streptomyces
; Capsicum annum
; Plant Growth Promotion
1. Introduction
Phytophthora capsici is an oomycete pathogen that affects numerous economically important crops worldwide, particularly members of the Solanaceae and Cucurbitaceae families. It is considered one of the most significant pathogens of bell pepper. Disease management primarily relies on fungicide application; however, under environmental conditions favorable to the pathogen, no currently available fungicide achieves complete disease control [1].
Over the years, numerous fungicides have been developed to prevent and control pepper blight, with metalaxyl being one of the most widely used. However, Phytophthora spp. is highly susceptible to mutations and has developed resistance to such fungicides, which typically have a single site of action on pathogens [2]. Cases of resistance have also been reported with other fungicides, such as mefenoxam, fluopicolide, cyazofamid, and oxathiapiprolin, particularly in regions where intensive, fungicide-based disease management programs are practiced [3].
Pesticide resistance is one of the primary causes driving the increased use of chemical pesticides. Extensive pesticide application has been associated to various human diseases and health disorders, as well as environmental damage. Consequently, environmentally friendly pest control methods, such as biological control, have been promoted to minimize chemical pesticide usage [4].
The use of living organisms for pest control is considered a more successful alternative for pest management. Recent studies have investigated antagonistic bacteria for the control of fungal and bacterial pathogens, with particular attention to species from the genera Pseudomonas spp., Bacillus spp, and Streptomyces spp, some of which are already available as commercial products [5].
Streptomyces are an important group of soil-dwelling bacteria within the Actinomycetaceae family, distinguished by their high G+C content, Gram-positive nature, and filamentous structure similar to that of fungi. These bacteria produce approximately 75% of currently used antibiotics and various bioactive metabolites with antifungal, antiviral, immunosuppressive, anticancer, and antioxidant activities [6].
Streptomyces strains from unexplored environments are considered potential sources of novel natural compounds, particularly those from desert habitats. Microorganisms in these extreme environments endure desiccation, extreme temperatures, intense radiation, low nutrient levels, low water availability, and high salinity, which may enhance their production of unique bioactive compounds [7].
The Cuatro Cienegas Basin, an oasis in the middle of the Chihuahuan Desert, contains a network of pools with exceptional microbial diversity and endemism. Its highly oligotrophic conditions make it a promising area for the discovery of microbial agents with antagonistic potential [8].
The objectives of this study were to (i) identify and characterize a strain isolated from Cuatro Cienegas, (ii) evaluate the antifungal activity of selected isolates against soil-borne fungal pathogens in vitro, and (iii) determine the efficacy of the selected isolate as a growth promoter and biological control agent against P. capsici in bell pepper (Capsicum annuum) plants.
2. Materials and Methods
2.1. Bacterial Strain, Fungal Plant Pathogens and Seeds
Sixty strains of actinomycetes used in this study (Table S1) were isolated in a previous investigation at different points in Cuatro Cienegas, Coahuila, Mexico and were reactivated and grown at 28°C/10 days on International Streptomyces Project 2 (ISP2) culture medium [8].
Phytopathogens used in this study (Phytophthora capsici, Fusarium solani, Fusarium oxysporum, Fusarium lateritium, Macrophomina phaseolina, Colletotrichum coccodes, Botryosphaeria rhodina, Botrytis cinerea and Sclerotium rolfsii), were obtained from culture collections from Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) General Terán; Universidad Autónoma de Querétaro (UAQ), and Centro de Investigación en Alimentación y Desarrollo (CIAD) Cuauhtémoc, Chihuahua. All pathogens were cultured at 28°C in Petri dishes with potato dextrose agar (PDA) for 10 days.
Bell pepper seeds (Capsicum annuum cv. California Wonder) from the 'Rancho Los Molinos' brand, or Arabidopsis thaliana ecotype Columbia, were used for all growth promotion and biocontrol assays.
2.2. Screening of Actinobacterias with Antagonistic Properties
Sixty strains of actinobacteria were evaluated for their ability to inhibit the growth of seven phytopathogens using the dual culture technique. A mycelial disk (7 mm diameter) of each pathogen was placed at the center of a Petri dish with ISP4 solid medium and a different actinobacteria strain was inoculated at four points equidistant from the fungal inoculum. Control plates contained only the pathogen inoculum. After 10 days of incubation at 28°C, the actinobacteria that showed inhibition halos against the pathogen were recorded. The actinobacteria with the highest number of antagonisms against phytopathogens was selected for a second round of selection using the dual culture technique against 9 phytopathogens.
2.4. Antifungal Activity In Vitro
Spores of the selected strain PR69 were streaked in two parallel lines using a bacteriological loop, each line placed 1.5 cm from the edge of a Petri dish containing ISP4 agar medium, and incubated for 3 days at 28°C. Then, a mycelial disk from a pathogen was placed in the center of a Petri dish containing ISP4 medium, between the parallel lines of actinomycetes. Control plates contained only the pathogen inoculum. After incubation for 10 days at 28°C, the radial fungal growth of each pathogen was measured and the reduction in colony radius was calculated and compared to control samples using the formula: % Inhibition = [(D-d)x100]/D, where D is the mycelial diameter (mm) in control plates and d is the mycelial diameter grown opposite to actinobacteria [9]. The experiment was conducted in triplicate.
2.5. VOCs-Mediated Antifungal Activity
The effect of PR69 volatile organic compounds (VOCs) on pathogens was investigated using the sealed plate method as previously described [10]. Strain PR69 was streaked by covering the entire ISP4 medium in a Petri dish. Then, a 7mm mycelial disk from a pathogen was placed in the center of another Petri dish with ISP4 medium. The two plates (without lids) were sealed using parafilm to obtain a double-plate chamber where the plate with phytopathogen was above and the plate with actinomycete below. Double plates were incubated at 27°C for 10 days. As a control, the Petri dish containing pathogens was exposed to a Petri dish containing only ISP4. The experiment was performed in triplicate and the radial growth of the pathogen was measured to calculate the percentage of inhibition.
2.3. Morphological and Culture Characterization
Strain PR69 was stained with Gram staining to determine the thickness of its peptidoglycan wall and then was grown in 11 different liquid and solid media and incubated at 27°C for 14 days: ISP1 (5 g hydrolyzed casein, 3 g yeast extract); ISP2 (25 g Kent marine reef salt mix, 10 g malt extract, 5 g yeast extract, 4 g dextrose,); ISP3 (20 g oat meal, 0.001 g MgCl2, 0.001 g ZnSO4, 0.001 g Fe2(SO4)3); ISP4 (10 g starch, 2 g CaCO3, 2 g (NH₄)₂SO₄, 1 g NaCl, 1 g K2HPO4, 1 g MgSO₄·7H₂O, 0.001 g ZnSO4, 0.001 g MgCl2, 0.001 g FeSO4·7H2O); ISP5 (1 g L-asparagine, 1 g K2HPO4, 0.001 g FeSO4, 0.001 g MnCl₂, 0.001 g ZnSO₄ and 10 mL glycerol); ISP6 (15 g peptone, 5 g protease peptone, 1 g yeast extract, 0.5 g C6H11FeNO7, 1 g K2HPO4, 0.08 g Na2S2O3); ISP7 (1 g L-Asparagine, 0.05 g L-Tyrosine, 0.05 g NaCl, 0.05 g K2HPO4, 0.05 g MgSO₄·7H₂O, 2.850 mg H3BO3, 1.8 mg MgCl₂·4H2O, 1.77 mg C4H4Na2O6, 1.360 mg FeSO4·7H2O, 0.027 mg CuCl2·2H2O, 0.04 mg CoCl2, 0.02 mg ZnCl₂, 0.025 mg Na2MoO4·2H2O, 15 mL Glycerol 100%); SFM (20 g mannitol, 20 g soy flour, 25 g Kent marine reef salt mix); M1 (50 mL salt stock, 5 g mannitol, 5 g glycerol); M3 (8 g peptone, 1 g starch) and Chitin (25 g Kent marine reef salt mix and 10 g chitin). Microscopic observations were performed to record the color of aerial and substrate mycelium and the capabilities to produce diffusible pigments and spore production.
2.6. Genome Sequencing, Assembly, and Bioinformatics Analysis
Genomic DNA was extracted using a modified phenol/chloroform method described previously [8]. The Actinobacteria mycelium was scraped from agar plates and transferred into Eppendorf tubes. The mycelium was then washed with 1 mL of 10% sucrose solution and resuspended in 400 μL of QTP extraction buffer, containing 4% Triton X-100, 20% SDS, 5 M NaCl, 2 M Tris-HCl (pH 8), and 500 mM EDTA (pH 8). Following resuspension, 400 μL of a 1:1 phenol/chloroform solution was added. To facilitate lysis, 0.1 mm glass beads were included, and the mixture was mechanically disrupted for 2 minutes. The lysates were centrifuged at 12,000 rpm for 15 minutes, and DNA in the aqueous phase was precipitated with 2 volumes of ethanol and 1/10 volume of 3 M sodium acetate. After incubation at -20°C, the DNA was pelleted by centrifugation at 12,000 rpm for 10 minutes at 4°C. The pellet was washed twice with 70% ethanol and then resuspended in Milli-Q water containing RNase. Sequencing was carried out by whole genome sequencing using Illumina Mi-Seq 2*300 at CINVESTAV-LANGEBIO (Irapuato, Mexico).
Resulting sequences were trimmed to remove low-quality reads using Trimgalore v0.6.6 (version 0.12.5), then assembled with Unicycler v0.4. After assembly, partial 16S rDNA was compared to the NCBI GenBank database using BLASTn (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to determine the genus. Phylogenetic analysis was performed based on 81 core genes from genomic sequences using the UBCG2 tool [11]. Comparison of isolate PR69 with the most closely related strain was performed using average nucleotide identity (ANI) using EzBioCloud ANI calculator (https://www.ezbiocloud.net/tools/ani) [12] and Kostas Lab ANI (http://enve-omics.ce.gatech.edu/ani/index) [13]. DSMZ GGDC Genome-to-Genome Distance Calculator 3.0 (https://ggdc.dsmz.de/ggdc.php#) was used for digital DNA-DNA hybridization (dDDH) [14]. The assembled genome was annotated with RAST v2.0 (Rapid Annotation using Subsystem Technology) (https://rast.nmpdr.org/rast.cgi) [15]. Genome mining for potential secondary metabolites was performed using antiSMASH bacterial version 7.0 (https://antismash.secondarymetabolites.org/#!/start) [16].
2.7. Production of Extracellular Enzymes and Biochemical Characterization
The bacterial isolates were qualitatively screened for the production of enzymes such as cellulase, protease, lipase, and chitinase. Additionally, their growth-promoting characteristics, including siderophore production, phosphate solubilization, and nitrogen fixation, were evaluated.
Cellulase: Carboxymethyl cellulose agar (CMC) (10 g CMC carboxymethylcellulose, 2 g K2HPO4, 0.35 g (NH4)2SO4, 0.0748 g urea, 25 g Kent marine reef salt mix) with pH 6.8 to 7.0. Cellulolytic activity was evidenced by qualitative evaluation with the 1% Congo Red reagent (w/v).
Lipase: Rhodamine B agar (250 μL Tween 20%, 30 mL vegetable oil (extra virgin), 20 mL Rhodamine B 0.02% w/v). The production of lipase enzymes was observed by UV light (350 nm) [17].
Protease: Nutrient Agar supplemented with 20% (w/v) non-fat milk. The stock solution of 20% (w/v) of the milk was mixed with nutrient agar to a final concentration of 2% (w/v). The results were visualized by a clear halo around the colony [18].
Chitin: Colloidal chitin medium (10 g chitin with 25 g Kent marine reef salt mix). Colloidal chitin substrate was prepared using the method reported previously [19] modified by adding H₃PO₄ to dissolve the chitin and allowed to stir for 24 hours. Subsequently, cold 98% ethanol was added and centrifuged, followed by washing with distilled water until reaching pH 7.0. A clear halo around the colonies showed the capacity for chitinase production.
Siderophore Production: Chrome-Azurol S (CAS) agar was used to determine siderophore production according to the method reported previously [20]. Chrome Azurol S solution was prepared with 60.5 mg chrome azurol S, 72.9 mg HDTMA, 30.29 g PIPES and agarose 0.9% filled with MiliQ water. After incubation for 1 hour at room temperature in the dark, the change of color around the colony (from blue to orange) indicated the siderophore production.
Phosphate Solubilization: Pikovskaya’s agar (PVK) was used to measure phosphate solubilizing (Ca3(PO4)2) activity. Solubilization was observed by the appearance of a clear zone around the colony [21].
Nitrogen fixation: Nitrogen fixation capacity was determined by the presence or absence of bacterial colony growth in NFB medium (Nitrogen-Free medium: 15 g sucrose, 5 g malic acid, 0.2 g MgSO4·H2O, 0.5 g K2HPO4, 0.1 g NaCl, 0.2 g CaCl2·H2O, 0.4 g KOH, 4 mL FeCl3·7H2O, 2 mL bromothymol blue 1%, 2 mL macronutrient solution, 1 mL vitamin solution, 15 g bacteriological agar). The macronutrient solution contains 0.2 g Na2MoO4·2H2O, 0.28 g H3BO3, 0.235 g MnSO4, 0.008 g CuSO4·5H2O and the vitamin solution was prepared with 0.1 g biotin, 0.2 g pyridoxal-HCl. Each colony was inoculated onto NFB agar medium and then incubated at 27°C for 10 days.
2.8. VOC-Mediated Plant Growth Promotion
Arabidopsis thaliana seeds were surface-sterilized using 70% ethanol for 1 minute, followed by 20% sodium hypochlorite for 2 minutes, and then rinsed three times with sterile, double-distilled water. Eight seeds were sown in 90x15 mm Petri dishes containing 50% MS medium, composed of 2.21 g of MS salts, 15 g of sucrose, and 15 g of bacteriological agar. After 4 days of growth, the seedlings were exposed to either bacterial volatiles or an agar medium control.
The experimental setup involved placing a 90x15 mm Petri dish with the A. thaliana seedlings (without its lid) inside a larger 150x20 mm glass Petri dish. Below it, a small 35x10 mm Petri dish containing a 7-day-old culture of Streptomyces sp. PR69 grown in ISP4 medium (or an agar medium for the negative control) was positioned, also without a lid. The glass plate was sealed with plastic wrap to ensure the volatile compounds from the Streptomyces isolate could interact directly with the plants. The seedlings were then allowed to grow for 7 days at 23°C, under a 16-hour light/8-hour dark cycle. The effects of the treatment were assessed by measuring the number of secondary roots, the root length and the fresh weight.
2.9. Inoculation of Bell Pepper Plants with Streptomyces sp. PR69 and Phytophthora capsici
Streptomyces sp. PR69 was grown in SFM medium with shaking at 125 rpm for 10 days at 30°C. The biomass around the flask was collected and resuspended in sterile distilled water (sdw) to prepare an inoculum with 10⁶ cfu/mL. Bell pepper seeds were sterilized with 97% alcohol, 15% sodium hypochlorite and several washes with sdw. The seeds were sown in seedling trays filled with a soil mix (Sunshine mix 3) inoculated with 1 mL of spore suspension of Streptomyces sp. PR69. Germinated seeds were sown in pots (9cm x 6.5cm x 6.5cm) containing soil treated with 10 mL of Streptomyces sp. PR69 suspension and kept at 27–30 °C with a relative humidity level of 40–60%. One week after inoculation, each pot was inoculated with 10 mL zoospore suspension (approximately 1x10⁶ zoospores/mL) of P. capsici. The zoospore suspension was prepared using cultures of P. capsici grown on potato dextrose agar(PDA). Sterile, bidistilled water was added to the culture, which were then left at room temperature for 24 hours. Following this, the cultures were refrigerated at 4 °C for 1 hour and subsequently incubated at 30°C for 1 hour to induce zoospore release. The supernatant was collected from the plates, and the zoospore concentration was adjusted to 1×10⁶ zoospores/mL using a hemocytometer. One week after pathogen inoculation, each pot was additionally inoculated with 10 mL of Streptomyces sp. PR69 suspension. Each treatment, consisting of 10 plants, was replicated three times and observed daily for 20 days post-inoculation. Seedlings inoculated only with P. capsici or only with Streptomyces sp. PR69 served as controls.
2.10. Statistical Analysis
All experiments were performed in triplicate, and statistical significance was set at p < 0.05. The results were analyzed using a Student's t-test or ANOVA, with means compared using Tukey's test.
3. Results
3.1. Antifungal Activity In Vitro
A total of 60 actinomycete isolates were tested for their inhibitory effects against various phytopathogens. Of these, 29 strains demonstrated inhibitory activity against at least one plant pathogen (Figure S1). Notably, strain PR69 exhibited significant growth inhibition in direct confrontation assays against several fungal pathogens, including the soilborne pathogens Phytophthora capsici, Colletotrichum coccodes, Macrophomina phaseolina, Fusarium lateritium, Fusarium solani, and Fusarium oxysporum, as well as foliar and floral pathogens Botryosphaeria rhodina and Botrytis cinerea (Figure 1). The strongest inhibitory effect was observed against C. coccodes (Table 1). However, isolate PR69 did not inhibit pathogen growth through volatile compound-mediated mechanisms (Figure S2)
3.2. Media-Dependent Growth and Biomass Optimization of Strain PR69
The strain PR69 was preliminarily identified as an actinomycete by Gram staining (Figure S3), and then was incubated in 11 different media (Figure S4). In solid media the strain was able to grow in the 11 types of culture media tested (ISP1 to ISP7, M1, M2, SFM and Chitin). Colonies were circular, raised, and irregular, it was also observed that the morphology of the colony does not differ with the medium (Table S2). In liquid it was observed that they excrete dark brown pigments, melanin or melanoid, in ISP1, ISP6, ISP7 and M3, light brown in ISP2 and SFM, also yellow pigment in Chitin media. In the SFM medium it showed greater production of biomass and spores for which this medium was selected for future tests (Table S3).
3.3. Genome Sequencing, Assembly, and Bioinformatics Analysis
The genome of Streptomyces sp. PR69 was successfully sequenced and assembled. The final assembly consisted of 36 contigs, encompassing a total genomic length of 6,533,639 base pairs (bp). The N50 value of the assembly was calculated to be 6,393,861, indicating a high quality of genome assembly. The genome displayed a high G+C content of 71.52%. In terms of genomic features, the Streptomyces sp. PR69 genome contains 5,909 protein coding sequences (CDS), alongside 63 transfer RNA (tRNA) genes, and 4 ribosomal RNA (rRNA) genes (Table 2). No plasmid sequences were found. The genomic sequence of Streptomyces sp. PR69 was deposited in the NCBI GenBank database under BioProject ID PRJNA889045, with BioSample accession number SAMN31231225. The genome assembly is available by searching the NCBI database as GCA_026420845 to facilitate future research and comparative analysis with other microbial genomes.
The 16S rRNA gene sequence comparison using the BLASTn tool confirmed that PR69 belongs to the Streptomyces genus, showing the highest sequence identity of 98.97% with Streptomyces sp. strain HNM0663. In the phylogenetic tree constructed with 95 Streptomyces genomes (Table S4), Streptomyces sp. PR69 forms a clade with Streptomyces spongiicola HNM0071 and Streptomyces tirandamycinicus HNM0039 (Figure 2). To further clarify the phylogenetic relationship, digital DNA-DNA hybridization (dDDH) and Average Nucleotide Identity (ANI) values were calculated. The dDDH value between Streptomyces sp. PR69 and S. tirandamycinicus HNM0039 was 24.70%, which is below the 70% threshold generally used for species delineation. Similarly, the ANI values, calculated using ANI calculator and OrthoANIu tool, were 82.79% and 80.45% respectively, both below the standard species delimitation threshold of 95-96% (Table S5). The genome was further analyzed for secondary metabolite biosynthetic potential using the antiSMASH version 7.0, revealing a total of 28 gene clusters associated with secondary metabolite biosynthesis. These clusters corresponded to seven different types of secondary metabolites. Remarkably, seven clusters showed 100% similarity to known clusters, while two clusters did not show similarity to any entries in the current database (Table 3).
3.4. Production of Extracellular Enzymes and Biochemical Characterization
To assess the potential of Streptomyces sp. PR69 as a biological control agent, a series of qualitative enzymatic tests were conducted (Table 4). These tests revealed notable lipolytic activity, indicating the strain's ability to hydrolyze lipids (Figure S5).
Further investigations were carried out to evaluate the plant growth-promoting attributes of Streptomyces sp. PR69. The strain was tested for several key attributes, including phosphate solubilization, siderophore production and in vitro nitrogen fixation. Using a colorimetric method demonstrated the ability to produce siderophores. Additionally, the PR69 strain confirmed the ability to fix nitrogen. However, when tested on Pikovskaya agar it did not show phosphate solubilization activity (Figure S6).
3.5. Effect of Streptomyces sp. PR69 on Arabidopsis thaliana Growth and Root Development
The influence of volatile organic compounds (VOCs) emitted by Streptomyces sp. PR69 on the growth of Arabidopsis thaliana was evaluated in vitro. Notably, A. thaliana plants exposed to the VOCs from Streptomyces sp. PR69 exhibited significant growth enhancement compared to the control group. These plants were observed to be larger with more developed secondary root systems. The results of the Student t-test confirmed that the differences in fresh weight, root length and secondary root development between the treated and control groups were statistically significant. This suggests that exposure to Streptomyces sp. PR69 VOCs positively influences the growth and root development of A. thaliana under the conditions tested (Figure 3).
3.6. Biocontrol of P. capsici in Bell Pepper Plants by Streptomyces sp. PR69
To evaluate the potential of PR69 to control P. capsici infection in bell pepper plants, an experiment was conducted with four plant groups: an untreated control group (light blue bar), a group treated only with PR69 (yellow bar), a group treated with both PR69 spore suspension and the pathogen P. capsici (dark blue bar), and a group inoculated solely with the pathogen (red bar). After inoculating the bell pepper plants, the progression of disease symptoms was monitored. Stem rot symptoms, attributable to P. capsici infection, began to appear around day 10 post-inoculation. By day 20, plants treated with Streptomyces sp. PR69 exhibited a significant reduction in infection incidence compared to plants treated only with the pathogen (Figure 4). At the end of the experiment, approximately 40% of the bell pepper plants treated only with the pathogen (red bar) survived, whereas those treated with both the pathogen and PR69 (dark blue bar) reached a 67% survival rate. This result demonstrated that treatment with spore suspension of Streptomyces sp. PR69 increased bell pepper plant survival by 40% when grown in soil inoculated with a P. capsici suspension (Figure 5). Additionally, a significant difference was observed in the fresh weight of bell pepper plants between the treatment group that received only Streptomyces sp. PR69 and the untreated control group (Figure 6). This finding suggests that, beyond its role in disease suppression, Streptomyces sp. PR69 may positively influence plant growth parameters.
4. Discussion
Actinobacteria represent the largest group of microorganisms that produce antibiotics and other bioactive substances. Recent research has focused on discovering new organisms in extreme and underexplored environments, aiming to identify microorganisms with novel secondary metabolic pathways, thereby increasing the likelihood of uncovering new microbial products. These environments—such as deep oceans, ice sheets, deserts, and volcanoes—often harbor unique organisms that have evolved specialized survival mechanisms. One such environment is Cuatro Ciénegas, known for its exceptional nutrient conditions, which resemble those of the Precambrian Ocean, making it a distinctive and scientifically valuable site [22].
The Cuatro Ciénegas Valley, located in the Chihuahuan Desert, is one of Mexico’s most important wetlands and has been internationally recognized as a RAMSAR site (Wetlands of International Importance). A recent study conducted in this area isolated and identified strains of Actinobacteria, with Streptomyces emerging as the most abundant genus [8]. The researchers also identified potential new species, highlighting the high diversity and endemism of the site. These findings increase the biotechnological interest in this region due to the potential industrial applications of the unique microorganisms found there.
The objective of this study was to evaluate the strain Streptomyces sp. PR69, isolated from Cuatro Ciénegas, Coahuila, as a biofungicide against the phytopathogen Phytophthora capsici in bell pepper plants. In preliminary in vitro confrontation tests, 50% of the actinomycetes isolated from Cuatro Ciénegas exhibited antifungal activity against at least one of the seven pathogens tested (Figure S1). Among the actinomycetes evaluated, the strain Streptomyces sp. PR69 demonstrated significant antifungal activity, inhibiting the growth of eight of the nine phytopathogens tested: Botrytis cinerea, Botryosphaeria rhodina, Colletotrichum coccodes, Macrophomina phaseolina, Phytophthora capsici, Fusarium lateritium, Fusarium solani, and Fusarium oxysporum.
The percentage of inhibition in vitro against Phytophthora capsici was 71.09%, which is higher than previously reported values for other Streptomyces strains. For instance, Streptomyces griseus H7602 showed 53.33% inhibition of P. capsici growth [23], while Streptomyces rochei IT20 and Streptomyces vinaceusdrappus SS14 demonstrated 69% and 63% inhibition, respectively [24]. Additionally, Streptomyces sp. ABV38 and ABV45 exhibited inhibition rates of 51% and 50%, respectively, against P. capsici CH11[25].
Due to its in vitro antagonistic capacity, the strain PR69 was selected for further morphological and genomic analysis. PR69 has a linear genome of 6,570,163 base pairs, with a G+C content of 71.51%. The genome size obtained is smaller compared to the average size of another Streptomyces, which ranges between 6.7 and 10.1 Mbp. However, it is much larger than Streptomyces xiamenensis 318 (isolated from mangrove soil), whose genome length is 5.96 Mb, being the smallest found so far [26],[27]. Streptomyces albus which is another of the smallest Streptomyces genomes, with 6.8Mb, but with the highest known G+C content 73.3% [28]. Streptomyces genomes typically have a high G+C content (70% to 74%) and many repetitive sequences, which influence their genetic stability and the production of secondary metabolites but makes it difficult to fully assemble them from short reads. Consequently, most available genomes are only in draft status [29]. High genomic G+C content is associated with certain environments such as soil and aerobic environments, which induce relatively high rates of DNA damage in the form of double-strand breaks (DSBs) requiring repair. This suggests that DNA damage may be a fundamental factor of G+C content and that it may be a selective adaptation to particular habitats [30]. Despite limited studies on marine adaptations, recent comparative genomics has revealed that Streptomyces of marine origin generally have smaller genome sizes and slightly higher G+C content compared to Streptomyces from other environments. Marine Streptomyces isolates have been shown to produce unique bioactive compounds not found in their terrestrial counterparts, highlighting their metabolic capabilities in marine environments [31]. After analyzing the 16S rRNA sequence and considering the morphological characteristics, strain PR69 was identified as a member of the Streptomyces genus. Further phylogenetic analysis using the complete genome showed that Streptomyces sp. PR69 is closely related to Streptomyces tirandamycinicus HNM0039. This strain of actinobacteria was isolated from a marine sponge, and its crude extract demonstrated strong antibacterial activity against Streptococcus agalactiae [32]. It is also related to Streptomyces spongiicola HNM0071, another actinobacteria isolated from a marine sponge that produces antitumor metabolites such as staurosporine and echinomycin [33]. Streptomyces sp. PR69 when compared with related species showed an ANI of 80% and a dDDH of 20%, indicating that it is a distinct strain. The relationship of strain PR69 to marine bacteria is not surprising given its source of isolation. Cuatro Cienegas is recognized for its unique aquatic ecosystems, with unusual and extreme stoichiometric ratios. These conditions mimic those found in the ancient seas of the late Precambrian, which host diverse microbial communities. Previous research on bacterial biodiversity in Cuatro Cienegas has reported the isolation of genomes of some Bacillus [22] and Actinobacteria [8] closely related to strains of marine origin, indicating their ecological importance in these ecosystems.
Once the complete genome of Streptomyces sp. PR69 was available, 28 putative biosynthetic gene clusters (BGC) were identified. In actinomycetes, the average BGC covers 1.64 Mbp and encodes 35 secondary metabolites. Certain strains, such as Kutzneria albida, Streptomyces bingchenggensis, and Streptomyces rapamycinicus, are found to encode the greatest number of natural products, dedicating 2.5 to 3.09 Mbp to encode between 48 and 53 secondary metabolites [34]. In the strain Streptomyces sp. PR69, 28 potential Biosynthetic Gene Clusters (BGCs) were identified (Table 3). Among these, 7 BGCs exhibit a similarity above 80%, while the remaining 21 BGCs show a very low similarity, indicating a small percentage of matching genes with the closest known compounds. Unfortunately, even though Streptomyces harbors numerous gene clusters responsible for producing natural products, most of these BGCs do not get expressed under laboratory conditions [35]. Although the presence of 28 gene clusters encoding secondary metabolites was identified, only 6 clusters could be identified that contained 100% of the known cluster genes for the biosynthetic clusters of ectoine, melanin, geosmin, naringenin, SapB and pristinol.
Ectoine (1,4,5,6-tetrahydro-2-methyl-4-pyrimidinecarboxylic acid) is a natural compound found in halophilic bacteria such as Ectothiorhodospira halochloris. It assists microorganisms in maintaining osmotic balance within cells without disrupting cellular metabolic processes. Due to its robust stabilizing ability, it often functions as a thermal protector against extreme freezing, drying, and heating conditions [36]. Streptomyces commonly produces ectoines in response to osmotic stress conditions [37]. Ectoine may be a key compound that helps Streptomyces sp. PR69 survive under conditions of extremely imbalanced nutrient stoichiometry, sulfur and magnesium minerals such as those found in Cuatro Cienegas [22]. Another compound that may help Streptomyces sp. PR69 in its survival in extreme environments is melanin. Melanin protects microorganisms from environmental stressors such as ultraviolet radiation, toxic heavy metals, and oxidative stress. They can also influence bacterial interactions with other organisms and are crucial for bacterial survival and pathogenesis in diverse environments [38]. In growth tests in different culture media, Streptomyces sp. PR69 showed great pigment production. The diverse properties of melanins have driven many applications and recent efforts have been made to produce the pigment on a biotechnologically relevant scale. One more metabolite found in the genome is SapB which is a hydrophobic peptide that plays a crucial role in the process of aerial hyphae formation and sporulation in Streptomyces species. It is thought to act as a surfactant during aerial morphogenesis, reducing surface tension and allowing hyphae to grow upwards [39]. These developmental processes are key in Streptomyces that led to the production of spores, to colonize new habitats and survive adverse conditions. A common metabolite found in Streptomyces is geosmin. Geosmin is a volatile compound known for its distinct earthy odor. It is produced by various microorganisms, including most Streptomyces, as well as several species of cyanobacteria, myxobacteria, and fungi [40]. The gene responsible for producing geosmin is present in almost all sequenced Streptomyces genomes. Geosmin plays a crucial role in mediating interactions between microorganisms and their environment, particularly in influencing the composition and dynamics of the soil microbial community. This, in turn, can impact nutrient cycling, soil structure, and resource availability for surrounding organisms [41]. Some studies suggest that the production of geosmin and other volatile organic compounds may be linked to defense or offense mechanisms against other microorganisms. The widespread conservation of the gene cluster layout suggests its significant role in regulating environmental adaptation. However, the precise function of geosmin and the benefits of this group of genes in bacteria are still not fully understood [42]. In in vitro assays, it was found that Streptomyces sp. PR69, which produces the earthy smell geosmin typical of most Streptomyces, also releases other volatile compounds. When tested with A. thaliana, this strain of Streptomyces sp. PR69 significantly boosted the total fresh weight of the seedlings by 186%, mainly due to an increase in the number of secondary roots. These findings align with previous studies show that Streptomyces can produce volatile compounds that enhance secondary root growth in A. thaliana. For example, Pérez-Corra et al. [43] demonstrated significant improvement in the in vitro growth of A. thaliana seedlings with their Streptomyces strains, particularly increasing fresh root weight by 94-300%. Additionally, Cordovez et al. [10] reported an increase in the fresh weight of A. thaliana seedlings exposed to VOCs from Streptomyces lividans 1326, mentioning acetoin (3-hydroxy-2-butanone) as a possible responsible. Another study showed that Arabidopsis thaliana Col-0 seedlings exposed to volatile compounds emitted by Streptomyces coelicolor M145 experienced significant morphological changes. In that experiment, it was identified that the volatile bacterial 3-octanone was the bioactive component responsible for this effect. Specifically, the weight of both roots (65%) and shoots (63%) significantly increased with the highest exposure to the volatile component [44]. The growth of secondary roots may play an important role in acquiring and transporting soil resources, interacting with soil organisms, and competing with other plants under stressful environmental conditions [45].
Besides producing volatile compounds, growth-promoting bacteria have multiple mechanisms to enhance plant growth, including producing phytohormones (auxins), improving nutrient acquisition (nitrogen fixation), and suppressing plant diseases (siderophores) [46]. The strain Streptomyces sp. PR69 demonstrated the ability to grow in a nitrogen-free medium, indicating its capability to obtain nitrogen from alternative sources. Nitrogen is essential for the formation of amino acids and proteins and plays a crucial role in all living organisms. Actinobacteria are recognized as plant growth-promoting bacteria (PGPB) because they can help metabolize nitrogen, converting it into compounds that plants can absorb [47]. Plants do not have the machinery to obtain nitrogen from the air on their own, but instead rely primarily on free-living bacteria in the soil or bacteria that live symbiotically in nodules on the roots that supply the combined nitrogen in the form of ammonia or nitrates, resulting from nitrogen fixation [48]. In addition to nitrogen fixation, Streptomyces sp. PR69 showed siderophore production activity in the Chrome Azurol S (CAS) assay, where the presence of siderophores is indicated by a color change from blue to orange. The chromium azurol S (CAS) assay is a universal colorimetric method that detects siderophores regardless of their structure [49]. Siderophore compounds produced by bacteria can promote plant growth and suppress diseases. Numerous studies have demonstrated that siderophore-producing bacteria can increase the yield of various plant species by enhancing iron uptake and act as antagonists by preventing iron availability to pathogens [50]. In experiments using bell pepper plants, the application of Streptomyces sp. PR69 resulted in a significant difference in the fresh weight of pepper plants compared to the non-inoculated negative control group. Similar results were observed in tests with A. thaliana. It was found that Streptomyces sp. PR69 produces volatile compounds that increase root length and the number of lateral roots, which may explain the increase in fresh weight of bell pepper plants. This suggests that PR69 has the potential to serve as a growth promoter, but further evaluations are required to assess its effectiveness and the mechanisms through which it promotes growth. The application of Streptomyces sp. PR69 in bell pepper plants led to an increase in the fresh weight of the plant and also increased the survival rate of plants inoculated with P. capcisi by 40%. It has been reported that bell pepper plants (C. annuum L. cv elmas) treated with Streptomyces rochei IT20 and Streptomyces vinaceus drappus SS14 reduced disease symptoms caused by P. capsici by 40% and 60%. When these two strains were co-cultured, the inhibition of the disease increased to 75%. [24]. Interestingly, it has been reported that soil treated with the supernatant of microorganism can show greater antifungal activity than soil treatment with the microorganism itself. As demonstrated by Chen et al. [51], in tests on pepper plants, the culture medium of Streptomyces plicatus B4-7, added three hours after inoculation with P. capsici, resulted in a 75% reduction in root rot. Trinidad-Cruz et al. [25] conducted an in vivo evaluation, demonstrating that the culture supernatants of Streptomyces strains ABV38 and ABV45 exhibited a wilting-suppressing effect in pepper plants, achieving survival rates of 77% and 94%, respectively. Although our results are lower than those reported in the literature, further trials are necessary to identify the specific compound responsible for the observed antifungal activity.
Additionally, a biosynthetic gene cluster (BGC) with 100% similarity to naringenin was identified in the genome of Streptomyces sp. PR69. Naringenin is a well-known flavonoid metabolite commonly found in plants; however, it has also been recently detected in Streptomyces clavuligerus [52]. Plant-derived naringenin is recognized for its numerous beneficial properties, including antioxidant, anti-inflammatory, antiviral, and anticancer effects [53]. Moreover, it exhibits antifungal activity against Candida albicans [54]. The isolation and characterization of this compound from Streptomyces could potentially lead to more effective antifungal treatments.
While extensive research has been conducted on microorganisms that suppress the growth of P. capsici, including Streptomyces spp., Bacillus spp., Paenibacillus spp., Trichoderma sp., Clitocybe nuda, and Aspergillus sp. [52], field application is limited due to the unstable and poor antimicrobial activity in the soil. Therefore, it remains important to discover highly efficient, broad-spectrum antagonistic Streptomyces strains for the development of better biocontrol agents [55]. The lack of efficacy during field application may be due to various factors that can affect the effectiveness of the microorganism. These factors include variability from plant to plant, crop to crop, year to year, and interactions with other microorganisms. The discovery of microorganisms with potential antifungal activity opens interesting possibilities for advances in agriculture. Researchers are optimistic about the potential application of these microorganisms in controlling pathogenic fungi that affect crops, which could lead to increased agricultural yields and reduced reliance on chemical fungicides. For example, the strain Streptomyces sp. PR69 has shown significant antifungal activity against several plant pathogens in both in vitro and pot tests. Future research aims to explore the full potential of these bacteria, including their application in various crops and environmental conditions. Furthermore, future field trials are necessary to further evaluate their effectiveness. These tests will involve evaluating the performance of the strain in various environmental settings, understanding its interactions with other microorganisms, and assessing its impact on non-target species. Successful results could contribute to the management of plant diseases, offering an environmentally friendly and efficient solution.
5. Conclusions
In conclusion, out of the 60 isolates obtained from Cuatro Cienegas Coahuila, the PR69 strain stands out with potential as a biocontrol agent for Phytophthora capsici in bell pepper plants. Its efficacy as a growth promoter has been demonstrated both in vitro with Arabidopsis thaliana and in vivo with pepper plants. Morphological characterization and analysis of its genome revealed the identity of the isolate as belonging to the Streptomyces genus. Genome sequencing further suggested the presence of several secondary metabolites with potential plant growth-promoting, antimicrobial, and antifungal properties. Future research should focus on identifying the specific metabolites responsible for its biocontrol activity and on developing different formulations for field trials on a broader range of pathogens and crops to validate its efficacy.
6. Patents
No patented.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Number of actinomycetes that showed inhibition against phytopathogens Fl: Fusarium lateritium, Mp: Macrophomina phaseolina, Cc: Colletotrichum coccodes, Pc: Phytophthora capsici, Fo: Fusarium oxysporum, Fs: Fusarium solani and Sr: Sclerotium rolfsii. Figure S2. Effect of volatile compounds produced by Streptomyces sp. PR69 on phytopathogens using the sealed plate method. A) Colletotrichum coccodes, B) Phytophthora capsica, C) Fusarium oxysporum, D) Sclerotium rolfsii E) Fusarium solani F) Macrophomina phaseolina. Figure S3. Gram staining of isolate PR69 under 100x magnification. Figure S4. Macroscopic morphological characteristics of isolate PR69 cultured in different solid and liquid media. A) ISP1, B) ISP2, C) ISP3, D) ISP4, E) ISP5, F) ISP6, G) ISP7, H) M1, I) M3, J) SFM and K) Chitin. Figure S5. Qualitative screening of enzyme production of Streptomyces sp. PR69 A) Cellulase, B) Chitinase, C) Protease, and D) Lipase activity. Figure S6. Qualitative evaluation of Streptomyces sp. PR69 for siderophore production, phosphate solubilization and nitrogen fixation. A) Chrome-Azurol S Assay B) Pikovskaya agar medium C) Nitrogen-free medium. Table S1. Evaluation of the antagonistic activity of 60 actinomycete isolates against seven phytopathogens. Symbols indicate inhibition zones as follows: - no inhibition zone; + inhibition zone less than 2 mm; ++ inhibition zone greater than 2 mm. Table S2. Culture characteristics of Streptomyces sp. PR69 in 11 different solid media. Table S3. Culture characteristics of Streptomyces sp. PR69 in 11 different liquid media. Table S4. Streptomyces genomes used for phylogenetic analysis. Table S5. Genomic relationship of Streptomyces sp. PR69 with other Streptomyces by DNA-DNA hybridization (DDH) and average nucleotide identity (ANI).
Author Contributions
PKLR, STZ, MMCG and HAA contributed with methodology, experimental design and experiments. PKLR and LJGW performed the statistical analyses. PKLR and HAA wrote the original draft manuscript. PKLR, HAA and STZ contributed with writing—review and editing. HAA and LJGW contributed with funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research project was supported by grants from The Program for Scientific and Technological Research from Universidad Autónoma de Nuevo León, PAICyT-UANL (CT729-19, CT-1196-20 and CT1571-21), PROACTI (67-CA-2023 and 112-CAAFE-2024). Perla Karina López-Reyes (924454) and María Mercedes Cortés-González (927958) were supported by postgraduate scholarships granted by The National Council of Science and Technology (CONACYT, México). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability Statement
The Streptomyces sp. PR69 sequence was deposited at NCBI under GenBank accession number GCA_026420845.
Acknowledgments
We would like to thank Dr. Raul Rodríguez Guerra (INIFAP) for providing the pathogen strains, and Cynthia Damaris Correa Oviedo (UANL) for her assistance during the microbiological work with the Streptomyces collection. We also extend our gratitude to Max Mizraím Apolinar Hernández and Janneth Escudero Agudelo for their valuable support and expertise.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Saltos, L.A.; Corozo-Quiñones, L.; Pacheco-Coello, R.; Santos-Ordóñez, E.; Monteros-Altamirano, Á.; Garcés-Fiallos, F.R. Tissue specific colonization of Phytophthora capsici in Capsicum spp.: molecular insights over plant-pathogen interaction. Phytoparasitica 2021, 49, 113-122. [CrossRef]
- Wang, W.; Liu, X.; Han, T.; Li, K.; Qu, Y.; Gao, Z. Differential Potential of Phytophthora capsici Resistance Mechanisms to the Fungicide Metalaxyl in Peppers. Microorganisms 2020, 8, 278. [CrossRef]
- Siegenthaler, T.B.; Hansen, Z.R. Sensitivity of Phytophthora capsici from Tennessee to Mefenoxam, Fluopicolide, Oxathiapiprolin, Dimethomorph, Mandipropamid, and Cyazofamid. Plant Disease 2021, 105, 3000-3007. [CrossRef]
- Hu, Z. What Socio-Economic and Political Factors Lead to Global Pesticide Dependence? A Critical Review from a Social Science Perspective. International Journal of Environmental Research and Public Health 2020, 17. [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Francés, J.; Roselló, G.; Montesinos, E. Bacteria as Biological Control Agents of Plant Diseases. Microorganisms 2022, 10, 1759. [CrossRef]
- Sharma, V.; Sharma, A.; Malannavar, A.B.; Salwan, R. Molecular aspects of biocontrol species of Streptomyces in agricultural crops. In Molecular Aspects of Plant Beneficial Microbes in Agriculture, Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Elsevier: 2020; pp. 89-109.
- Sivakala, K.K.; Gutiérrez-García, K.; Jose, P.A.; Thinesh, T.; Anandham, R.; Barona-Gómez, F.; Sivakumar, N. Desert Environments Facilitate Unique Evolution of Biosynthetic Potential in Streptomyces. Molecules 2021, 26, 588. [CrossRef]
- Arocha-Garza, H.F.; Canales-Del Castillo, R.; Eguiarte, L.E.; Souza, V.; De la Torre-Zavala, S. High diversity and suggested endemicity of culturable Actinobacteria in an extremely oligotrophic desert oasis. PeerJ 2017, 5, e3247. [CrossRef]
- Liotti, R.G.; da Silva Figueiredo, M.I.; Soares, M.A. Streptomyces griseocarneus R132 controls phytopathogens and promotes growth of pepper (Capsicum annuum). Biological Control 2019, 138, 104065. [CrossRef]
- Cordovez, V.; Carrion, V.J.; Etalo, D.W.; Mumm, R.; Zhu, H.; van Wezel, G.P.; Raaijmakers, J.M. Diversity and functions of volatile organic compounds produced by Streptomyces from a disease-suppressive soil. Frontiers in Microbiology 2015, 6, 1-13. [CrossRef]
- Kim, J.; Na, S.-I.; Kim, D.; Chun, J. UBCG2: Up-to-date bacterial core genes and pipeline for phylogenomic analysis. Journal of Microbiology 2021, 59, 609-615. [CrossRef]
- Yoon, S.-H.; Ha, S.-m.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie van Leeuwenhoek 2017, 110, 1281-1286. [CrossRef]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: a toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Preprints 2016, 4, e1900v1901. [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: a database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Research 2022, 50, D801-D807. [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genomics 2008, 9, 75. [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: new and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Research 2023, 51, W46-W50. [CrossRef]
- Ortiz Lechuga, E.G.; Quintero Zapata, I.; Arévalo Niño, K. Detection of extracellular enzymatic activity in microorganisms isolated from waste vegetable oil contaminated soil using plate methodologies. African Journal of Biotechnology 2016, 15, 408-416. [CrossRef]
- Mun, B.G.; Lee, W.H.; Kang, S.M.; Lee, S.U.; Lee, S.M.; Lee, D.Y.; Shahid, M.; Yun, B.W.; Lee, I.J. Streptomyces sp. LH 4 promotes plant growth and resistance against Sclerotinia sclerotiorum in cucumber via modulation of enzymatic and defense pathways. Plant and Soil 2020, 448, 87-103. [CrossRef]
- Joe, S.; Sarojini, S. An Efficient Method of Production of Colloidal Chitin for Enumeration of Chitinase Producing Bacteria. Mapana - Journal of Sciences 2017, 16, 37-45. [CrossRef]
- Hu, Q.-P.; Xu, J.-G. A simple double-layered chrome azurol S agar (SD-CASA) plate assay to optimize the production of siderophores by a potential biocontrol agent Bacillus. African Journal of Microbiology Research 2011, 5. [CrossRef]
- Abbasi, S.; Safaie, N.; Sadeghi, A.; Shamsbakhsh, M. Streptomyces Strains Induce Resistance to Fusarium oxysporum f. Sp. Lycopersici Race 3 in Tomato through Different Molecular Mechanisms. Frontiers in Microbiology 2019, 10. [CrossRef]
- Souza, V.; Moreno-Letelier, A.; Travisano, M.; Alcaraz, L.D.; Olmedo, G.; Eguiarte, L.E. The lost world of cuatro ciénegas basin, a relictual bacterial niche in a desert oasis. eLife 2018, 7, e38278. [CrossRef]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Tindwa, H.; Lee, G.H.; Jeong, B.K.; Ro, H.M.; Kim, S.J.; Jung, W.J.; Kim, K.Y. Biocontrol potential of streptomyces griseus H7602 against root rot disease (Phytophthora capsici) in pepper. Plant Pathology Journal 2012, 28, 282-289. [CrossRef]
- Abbasi, S.; Safaie, N.; Sadeghi, A.; Shamsbakhsh, M. Tissue-specific synergistic bio-priming of pepper by two streptomyces species against phytophthora capsici. PLoS ONE 2020, 15, e0230531. [CrossRef]
- Trinidad-Cruz, J.R.; Rincón-Enríquez, G.; Evangelista-Martínez, Z.; Quiñones-Aguilar, E.E. Control biorracional de Phytophthora capsici en plantas de chile mediante Streptomyces spp. Revista Chapingo Serie Horticultura 2021, 27, 85-99. [CrossRef]
- Xu, M.J.; Wang, J.H.; Bu, X.L.; Yu, H.L.; Li, P.; Ou, H.Y.; He, Y.; Xu, F.D.; Hu, X.Y.; Zhu, X.M.; et al. Deciphering the streamlined genome of Streptomyces xiamenensis 318 as the producer of the anti-fibrotic drug candidate xiamenmycin. Scientific Reports 2016, 6, 18977. [CrossRef]
- Caicedo-Montoya, C.; Manzo-Ruiz, M.; Ríos-Estepa, R. Pan-Genome of the Genus Streptomyces and Prioritization of Biosynthetic Gene Clusters With Potential to Produce Antibiotic Compounds. Frontiers in Microbiology 2021, 12. [CrossRef]
- Zaburannyi, N.; Rabyk, M.; Ostash, B.; Fedorenko, V.; Luzhetskyy, A. Insights into naturally minimised Streptomyces albus J1074 genome. BMC Genomics 2014, 15, 97. [CrossRef]
- Sharma, V.; Kaur, R.; Salwan, R. Streptomyces: host for refactoring of diverse bioactive secondary metabolites. 3 Biotech 2021, 11, 340. [CrossRef]
- Weissman, J.L.; Fagan, W.F.; Johnson, P.L.F. Linking high GC content to the repair of double strand breaks in prokaryotic genomes. PLoS Genetics 2019, 15, e1008493. [CrossRef]
- Lacey, H.J.; Rutledge, P.J. Recently Discovered Secondary Metabolites from Streptomyces Species. Molecules 2022, 27, 887. [CrossRef]
- Huang, X.; Kong, F.; Zhou, S.; Huang, D.; Zheng, J.; Zhu, W. Streptomyces tirandamycinicus sp. Nov., a novel marine sponge-derived actinobacterium with antibacterial potential against streptococcus agalactiae. Frontiers in Microbiology 2019, 10. [CrossRef]
- Zhou, S.; Xiao, K.; Huang, D.; Wu, W.; Xu, Y.; Xia, W.; Huang, X. Complete genome sequence of Streptomyces spongiicola HNM0071 T , a marine sponge-associated actinomycete producing staurosporine and echinomycin. Marine Genomics 2019, 43, 61-64. [CrossRef]
- Jakubiec-Krzesniak, K.; Rajnisz-Mateusiak, A.; Guspiel, A.; Ziemska, J.; Solecka, J. Secondary metabolites of actinomycetes and their antibacterial, antifungal and antiviral properties. Polish Journal of Microbiology 2018, 67, 259-272. [CrossRef]
- Liu, Z.; Zhao, Y.; Huang, C.; Luo, Y. Recent Advances in Silent Gene Cluster Activation in Streptomyces. Frontiers in Bioengineering and Biotechnology 2021, 9. [CrossRef]
- Ng, H.S.; Wan, P.K.; Kondo, A.; Chang, J.S.; Lan, J.C.W. Production and Recovery of Ectoine: A Review of Current State and Future Prospects. Processes 2023, 11, 339. [CrossRef]
- Sadeghi, A.; Soltani, B.M.; Nekouei, M.K.; Jouzani, G.S.; Mirzaei, H.H.; Sadeghizadeh, M. Diversity of the ectoines biosynthesis genes in the salt tolerant Streptomyces and evidence for inductive effect of ectoines on their accumulation. Microbiological Research 2014, 169, 699-708. [CrossRef]
- Pavan, M.E.; López, N.I.; Pettinari, M.J. Melanin biosynthesis in bacteria, regulation and production perspectives. Applied Microbiology and Biotechnology 2020, 104, 1357-1370. [CrossRef]
- McCormick, J.R.; Flärdh, K. Signals and regulators that govern Streptomyces development. FEMS Microbiology Reviews 2012, 36, 206-231. [CrossRef]
- Jiang, J.; He, X.; Cane, D.E. Biosynthesis of the earthy odorant geosmin by a bifunctional Streptomyces coelicolor enzyme. Nature Chemical Biology 2007, 3, 711-715. [CrossRef]
- Becher, P.G.; Verschut, V.; Bibb, M.J.; Bush, M.J.; Molnár, B.P.; Barane, E.; Al-Bassam, M.M.; Chandra, G.; Song, L.; Challis, G.L.; et al. Developmentally regulated volatiles geosmin and 2-methylisoborneol attract a soil arthropod to Streptomyces bacteria promoting spore dispersal. Nature Microbiology 2020, 5, 821-829. [CrossRef]
- Churro, C.; Semedo-Aguiar, A.P.; Silva, A.D.; Pereira-Leal, J.B.; Leite, R.B. A novel cyanobacterial geosmin producer, revising GeoA distribution and dispersion patterns in Bacteria. Scientific Reports 2020, 10, 8679. [CrossRef]
- Pérez-Corral, D.A.; Ornelas-Paz, J.d.J.; Olivas, G.I.; Acosta-Muñiz, C.H.; Salas-Marina, M.Á.; Berlanga-Reyes, D.I.; Sepulveda, D.R.; de León, Y.M.P.; Rios-Velasco, C. Growth Promotion of Phaseolus vulgaris and Arabidopsis thaliana Seedlings by Streptomycetes Volatile Compounds. Plants 2022, 11, 875. [CrossRef]
- Dotson, B.R.; Verschut, V.; Flärdh, K.; Becher, P.G.; Rasmusson, A.G. The Streptomyces volatile 3-octanone alters auxin/cytokinin and growth in Arabidopsis thaliana via the gene family KISS ME DEADLY. bioRxiv 2020. [CrossRef]
- Strock, C.F.; Lynch, J.P. Root secondary growth: an unexplored component of soil resource acquisition. Annals of Botany 2020, 126, 205-218. [CrossRef]
- Sousa, J.A.d.J.; Olivares, F.L. Plant growth promotion by streptomycetes: ecophysiology, mechanisms and applications. Chemical and Biological Technologies in Agriculture 2016, 3, 24. [CrossRef]
- Kumar, M.; Kumar, P.; Das, P.; Solanki, R.; Kapur, M.K. Proactive role of Streptomyces spp. in plant growth stimulation and management of chemical pesticides and fertilizers. International Journal of Environmental Science and Technology 2022, 19, 10457-10476. [CrossRef]
- Cocking, E.C. Helping plants get more nitrogen from the air. European Review 2000, 8, 193-200. [CrossRef]
- Timofeeva, A.M.; Galyamova, M.R.; Sedykh, S.E. Bacterial Siderophores: Classification, Biosynthesis, Perspectives of Use in Agriculture. Plants 2022, 11, 3065. [CrossRef]
- Swarnalatha, G.V.; Goudar, V.; Gari Surendranatha Reddy, E.C.R.; Al Tawaha, A.R.M.; Sayyed, R.Z. Siderophores and Their Applications in Sustainable Management of Plant Diseases. In Secondary Metabolites and Volatiles of PGPR in Plant-Growth Promotion, Sayyed, R.Z., Uarrota, V.G., Eds.; Springer International Publishing: Cham, 2022; pp. 289-302.
- Chen, Y.Y.; Chen, P.C.; Tsay, T.T. The biocontrol efficacy and antibiotic activity of Streptomyces plicatus on the oomycete Phytophthora capsici. Biological Control 2016, 98, 34-42. [CrossRef]
- Álvarez-Álvarez, R.; Botas, A.; Albillos, S.M.; Rumbero, A.; Martín, J.F.; Liras, P. Molecular genetics of naringenin biosynthesis, a typical plant secondary metabolite produced by Streptomyces clavuligerus. Microbial Cell Factories 2015, 14, 178. [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of naringenin: A review of clinical trials. Pharmaceuticals 2019, 12, 11. [CrossRef]
- Soberón, J.R.; Sgariglia, M.A.; Carabajal Torrez, J.A.; Aguilar, F.A.; Pero, E.J.I.; Sampietro, D.A.; Fernández de Luco, J.; Labadie, G.R. Antifungal activity and toxicity studies of flavanones isolated from Tessaria dodoneifolia aerial parts. Heliyon 2020, 6, e05174. [CrossRef]
- Zhang, L.; Zhang, H.; Huang, Y.; Peng, J.; Xie, J.; Wang, W. Isolation and Evaluation of Rhizosphere Actinomycetes With Potential Application for Biocontrolling Fusarium Wilt of Banana Caused by Fusarium oxysporum f. sp. cubense Tropical Race 4. Frontiers in Microbiology 2021, 12. [CrossRef]
Figure 1.
Antifungal activity of Streptomyces sp. PR69 against different phytopathogens A) Botrytis cinerea, B) Botryosphaeria rhodina, C) Colletotrichum coccodes, D) Macrophomina phaseolina, E) Phytophthora capsici, F) Fusarium lateritium G) Fusarium solani, and H) Fusarium oxysporum.
Figure 1.
Antifungal activity of Streptomyces sp. PR69 against different phytopathogens A) Botrytis cinerea, B) Botryosphaeria rhodina, C) Colletotrichum coccodes, D) Macrophomina phaseolina, E) Phytophthora capsici, F) Fusarium lateritium G) Fusarium solani, and H) Fusarium oxysporum.
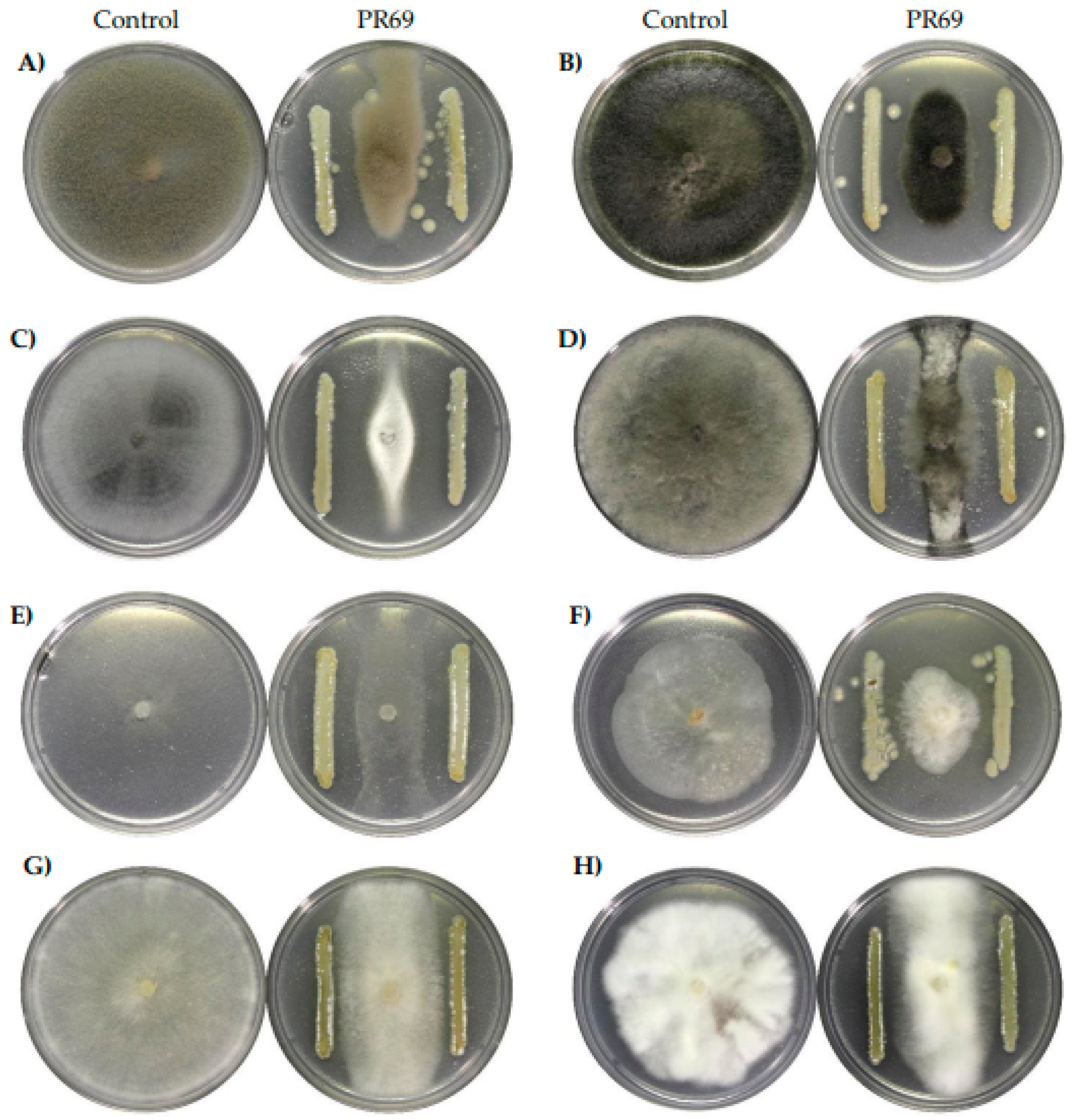
Figure 2.
Phylogenetic tree inferred by the maximum likelihood method based on 81 core genes obtained with UBCG2, showing the taxonomic position of the strain Streptmomyces sp. PR69. Phylogenetic analysis used the GTR+GAMMA+I model. The circles indicate that the value supports 100%.
Figure 2.
Phylogenetic tree inferred by the maximum likelihood method based on 81 core genes obtained with UBCG2, showing the taxonomic position of the strain Streptmomyces sp. PR69. Phylogenetic analysis used the GTR+GAMMA+I model. The circles indicate that the value supports 100%.
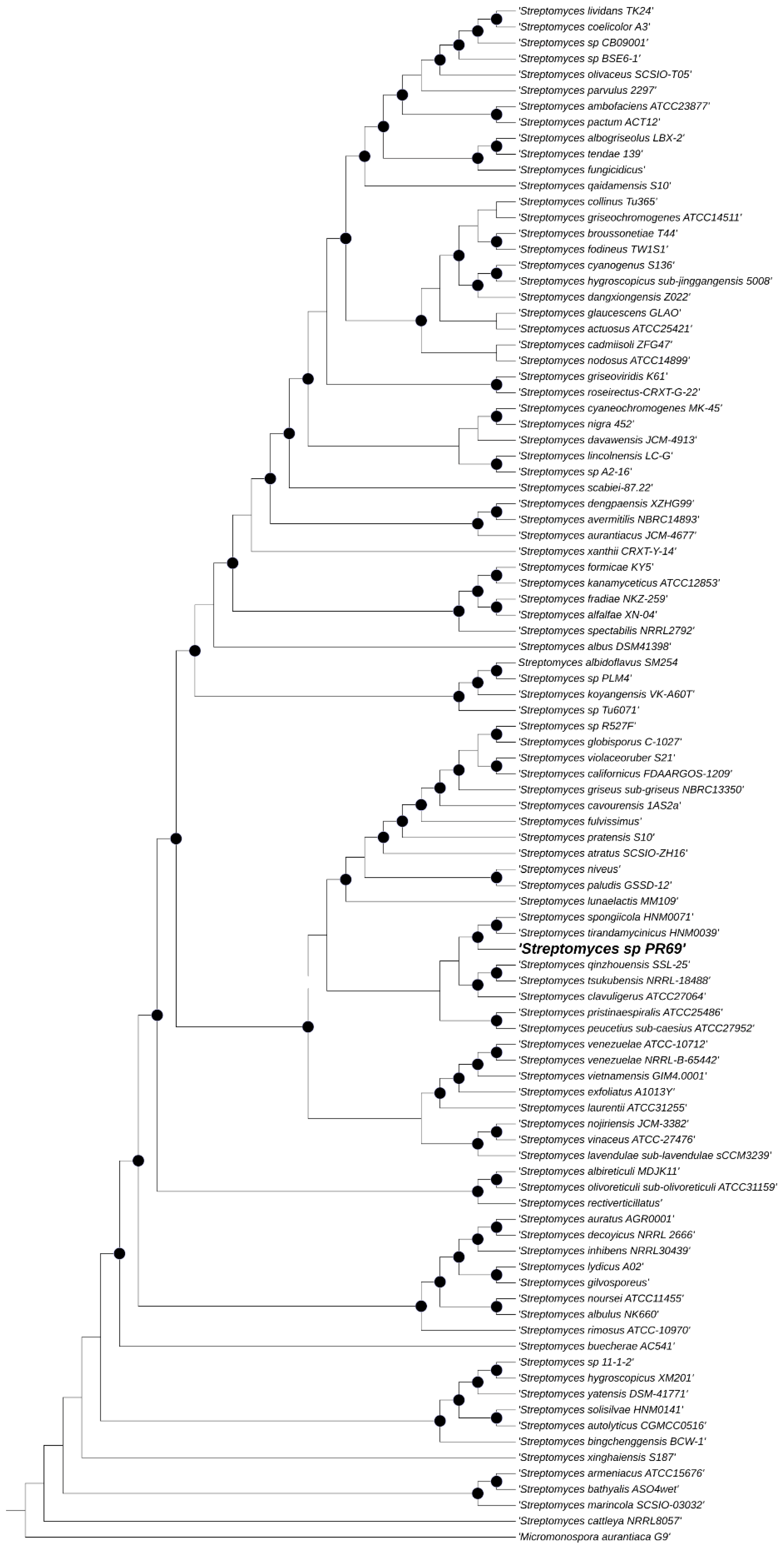
Figure 3.
Arabidopsis thaliana plants in an atmosphere shared set with Streptomyces sp. PR69 A) Control plants without actinomycete and plants exposed to Streptomyces sp. PR69 VOCs. Comparison of B) Secondary roots, C) Fresh weight and D) Root length. Error bars indicated standard errors of the means from three repeated experiments. Different letters indicate a statistical difference compared to controls using Student's t-test (p < 0.05).
Figure 3.
Arabidopsis thaliana plants in an atmosphere shared set with Streptomyces sp. PR69 A) Control plants without actinomycete and plants exposed to Streptomyces sp. PR69 VOCs. Comparison of B) Secondary roots, C) Fresh weight and D) Root length. Error bars indicated standard errors of the means from three repeated experiments. Different letters indicate a statistical difference compared to controls using Student's t-test (p < 0.05).
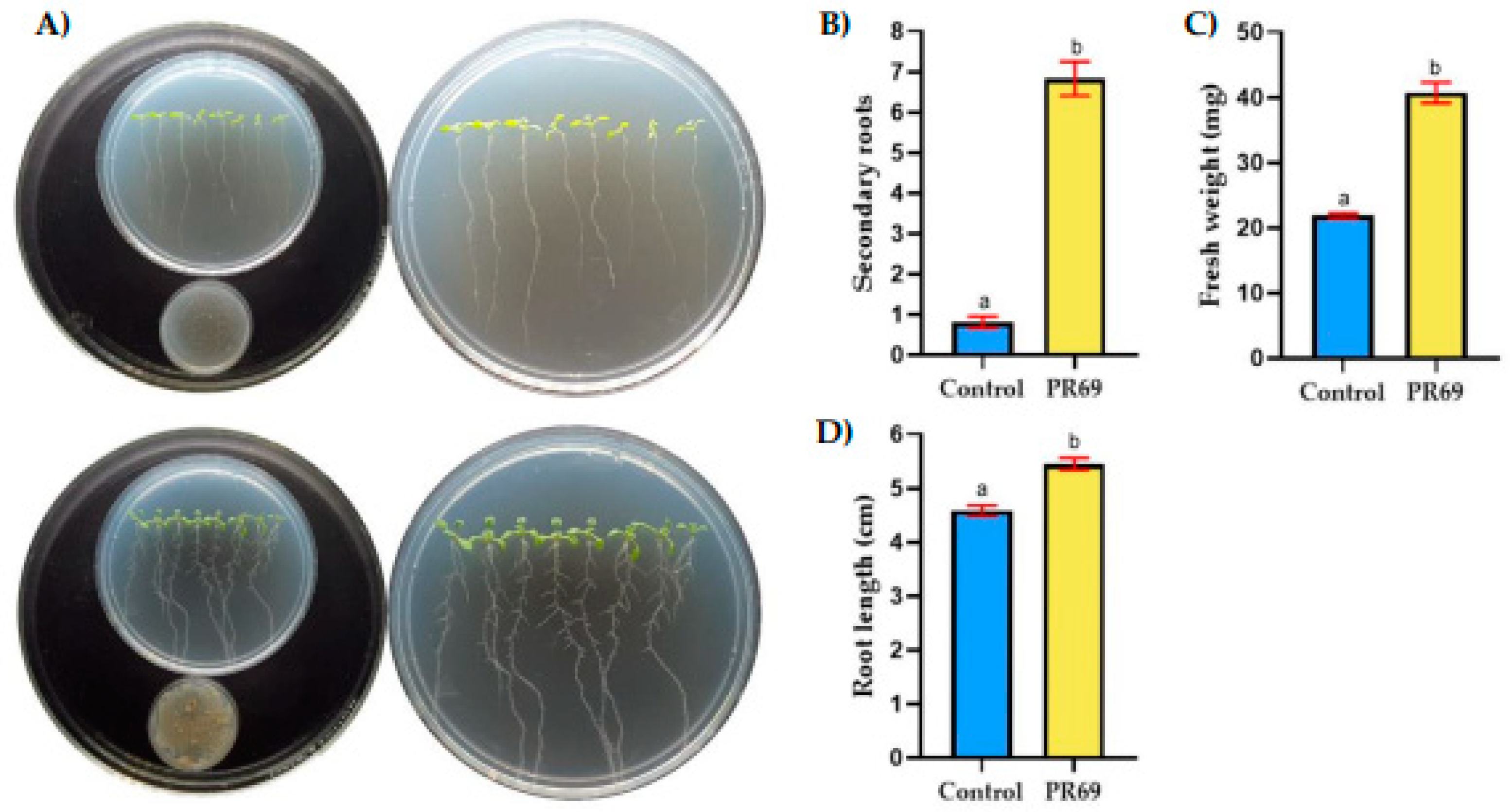
Figure 4.
Bell pepper plants (Capsicum annuum) were treated as follows: A) inoculation with Streptomyces sp. PR69 and Phytophthora capsici suspension, and B) inoculation with only P. capsici suspension (pathogen).
Figure 4.
Bell pepper plants (Capsicum annuum) were treated as follows: A) inoculation with Streptomyces sp. PR69 and Phytophthora capsici suspension, and B) inoculation with only P. capsici suspension (pathogen).

Figure 5.
Survival percentage of bell pepper plants for the four treatments. Control, untreated plants. Pc, Phytophthora capsici. Means ± standard errors for each treatment labeled with different letters indicate significant differences determined by ANOVA and a Tukey´s test.
Figure 5.
Survival percentage of bell pepper plants for the four treatments. Control, untreated plants. Pc, Phytophthora capsici. Means ± standard errors for each treatment labeled with different letters indicate significant differences determined by ANOVA and a Tukey´s test.
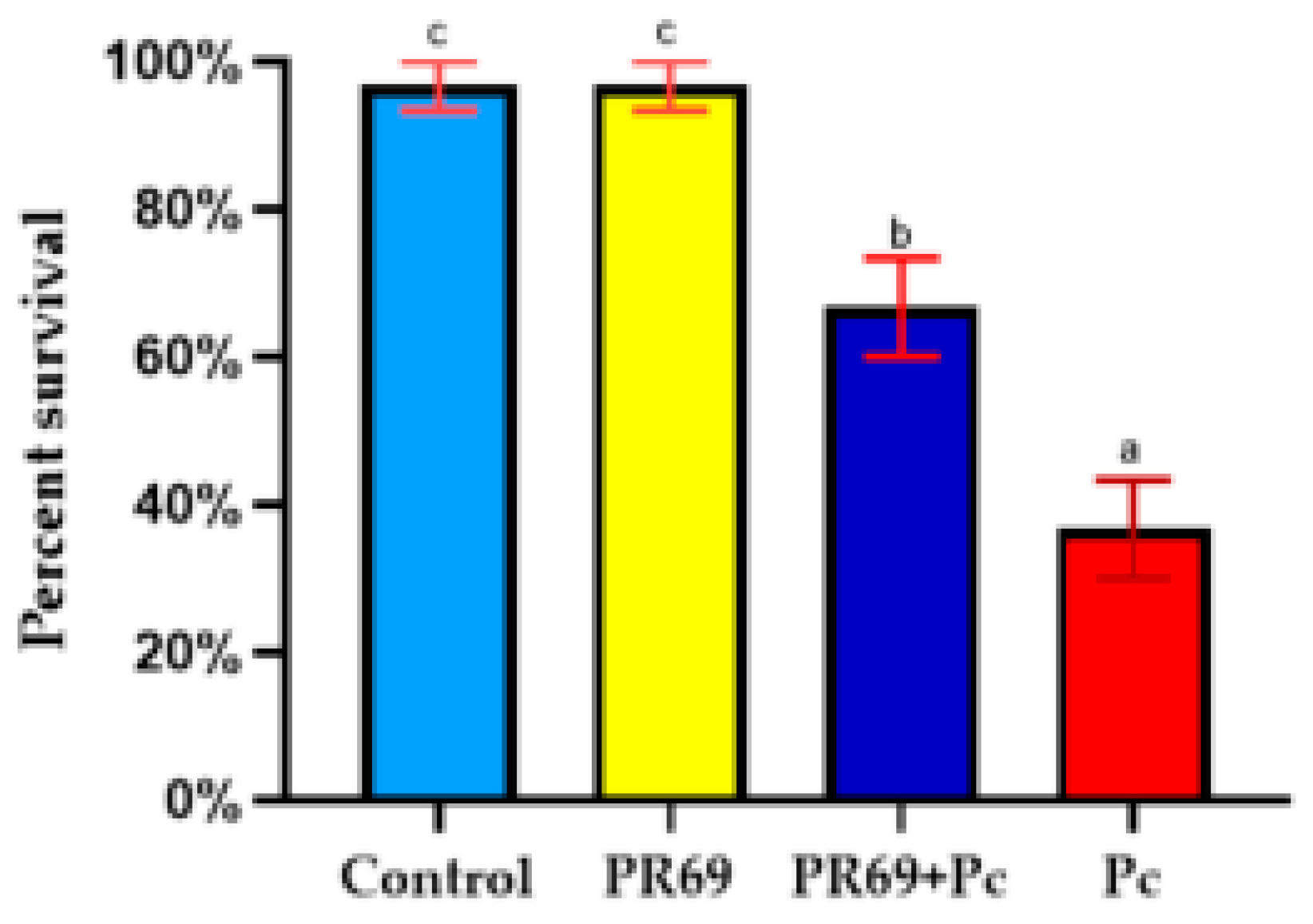
Figure 6.
Effect of Streptomyces sp. PR69 on the fresh weight of bell pepper plants. Error bars represent the standard error of the mean from three independent experiments. Different letters denote statistically significant differences, as determined by the Student's t-test.
Figure 6.
Effect of Streptomyces sp. PR69 on the fresh weight of bell pepper plants. Error bars represent the standard error of the mean from three independent experiments. Different letters denote statistically significant differences, as determined by the Student's t-test.


Table 1.
Antifungal activity of Streptomyces sp. PR69 against different phytopathogens.
| % Inhibition | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pc1 | Cc2 | Mp3 | Fl4 | Br5 | Fs6 | Bc7 | Fo8 | ||
| Isolate PR69 |
71.09±0.18 | 73.63±0.16 | 63.92±0.19 | 52.59±0.12 | 65.46±0.10 | 50.43±0.31 | 69.76±0.13 | 44.85±0.29 | |
Data are expressed as means ± standard errors of three replicates. 1Phytophthora capsici, 2Colletotrichum coccodes, 3Macrophomina phaseolina, 4Fusarium lateritium, 5Botryosphaeria rhodina, 6Fusarium solani, 7Botrytis cinerea, and 8Fusarium oxysporum.
Table 2.
Genome assembly and annotation data Streptomyces sp. PR69.
| Feature | |
|---|---|
| Contigs | 105 |
| Genome lenght | 6,570,163bp |
| G+C % | 71.51 |
| Contig L50 | 12 |
| ContigN50 | 186,895 |
| CDS | 5,956 |
| tRNA | 63 |
| rRNA | 4 |
| Protein with functional assignments | 3,898 |
| Antibiotic Resistance (source CARD, NDARO,PATRIC) |
43 |
Table 3.
Potential Biosynthetic Gene clusters regions found using antiSMASH (Version 7.0).
| Type | From-To (location) |
Most similar known cluster | Similarity% | ||
|---|---|---|---|---|---|
| Component | Type | ||||
| Ectoine | 324,766 - 335,164 | Ectoine | Other | 100 | |
| Melanin | 85,744 - 96,241 | Melanin | Other | 100 | |
| Terpene | 223,129 - 244,059 | Geosmin | Terpene | 100 | |
| T3PKS | 1 - 39,280 | Naringenin | Polyketide:Type III polyketide | 100 | |
| Lanthipeptide-class-iii | 28,816 - 51,518 | SapB | RiPP:Lanthipeptide | 100 | |
| Terpene | 1 - 16,602 | Pristinol | Terpene | 100 | |
| NRPS-like,NRPS | 1 - 28,694 | Antipain | NRP | 83 | |
| NRP-metallophore,NRPS,redox-cofactor | 278,062 - 337,959 | Mirubactin | NRP | 78 | |
| Terpene | 263,573 - 290,198 | Hopene | Terpene | 76 | |
| Melanin | 101,546 - 111,953 | Grixazone A | Terpene | 61 | |
| NRP-metallophore,NRPS,T1PKS | 160,886 - 250,379 | Peucechelin | NRP | 55 | |
| T1PKS,T2PKS,RiPP-like | 160,514 - 249,283 | Xantholipin | Polyketide | 51 | |
| NRPS | 1 - 37,748 | Netropsin | NRP | 40 | |
| Phenazine | 20,316 - 40,780 | Endophenazine A/endophenazine B | Other:Phenazine | 33 | |
| Melanin | 170,575 - 180,985 | Melanin | Other | 28 | |
| T3PKS,NRPS | 79,735 - 109,902 | Totopotensamide A/totopotensamide B | NRP+Polyketide | 28 | |
| NRPS,NRPS-like | 1 - 36,960 | Disgocidine/distamycin/congocidine | NRP | 28 | |
| NRPS-like | 12,279 - 53,110 | Lipstatin | NRP | 21 | |
| Terpene | 24,797 - 46,026 | Legonindolizidine A6 | NRP+Alkaloid | 12 | |
| NI-siderophore | 132,640 - 147,396 | Synechobactin C9/ C11/ 13/ 14/ 16/ A/ B/ C | Other | 9 | |
| NRPS-like | 43,673 - 85,050 | Chejuenolide A/chejuenolide B | Polyketide | 7 | |
| RiPP-like | 151,678 - 160,095 | Hexacosalactone A | Other | 4 | |
| NRPS-like | 109,671 - 132,257 | Sanglifehrin A | NRP+Polyketide | 4 | |
| Thioamitides | 1 - 13,147 | Prejadomycin/rabelomycin/gauDimycin C/gaudimycin D/UWM6/gaudimycin A | Polyketide:Type II polyketide+Saccharide:Hybrid/tailoring saccharide | 4 | |
| Other | 90,844 - 113,876 | A-503083 A/A-503083 B/A-503083 E/A-503083 F | NRP | 3 | |
| Terpene | 54,693 - 71,392 | Bombyxamycin A/bombyxamycin B | Polyketide | 3 | |
| CDPS | 66,910 - 87,659 | ||||
| Indole | 1 -19,239 | ||||
NRPS, Nonribosomal peptide synthetase cluster; PKS, polyketide synthase cluster; T3PKS, Type 3 polyketide synthase; RiPP, ribosomally synthesized and post-translational modified peptides.
Table 4.
Characteristics of extracellular enzymes and growth promotion.
| Characteristics | Streptomyces sp. PR69 |
|---|---|
| Cellulase | - |
| Protease | - |
| Chitinase | - |
| Lipase | + |
| Siderophores | + |
| Nitrogen fixation | + |
| Phosphate solubilization | - |
+ positive, − negative
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



