
Submitted:
23 November 2024
Posted:
25 November 2024
You are already at the latest version
Abstract
In much of neurobiological research, senescence and death are regarded as negative outcomes of aging. In the lives of animals, however, death may occur in many different ways, including as a natural result of reproduction, a common mode of death in invertebrates. This review examines the intersection of the nervous system, behavior, and reproductive death in three study systems that span the diversity of neural forms and life history strategies in invertebrates. In C. elegans, hermaphrodites experience a shortened lifespan as a result of extreme physiological changes caused by mating. In octopuses, signaling from the neuroendocrine center causes females to deteriorate over the duration of egg-brooding, dying before their offspring hatch. Lastly, in bumblebees, the entire colony perishes due to behavioral changes induced by the interplay of signaling between queens and her workers. Each model offers unique insights into the neuroendocrine control of reproductive death.
Keywords:
Reproductive death
; semelparity
; insulin/IGF-1 signaling
; juvenile hormone
Introduction
Invertebrate animals exhibit an incredible diversity of neural forms and life history strategies that underlie their evolutionary success in nearly every biome on earth. Only three invertebrate lineages have evolved brains, defined as a centralized collection of neural centers with distributed and hierarchical functions: annelids, cephalopod molluscs, and arthropods [1]. Animals in other invertebrate lineages have diffuse nerve nets or, as in the case of the well-studied nematode worm C. elegans, cerebral ganglia [1,2]. Reproductive strategies in invertebrates range from semelparity, classically defined as reproducing once before death, to iteroparity, or reproducing multiple times before death [3,4,5]. Here, we highlight research on three invertebrates that span a range of reproductive strategies and neural structures: the nematode worm, the octopus, and the bumblebee. Each study system provides important mechanistic insights into the neuroendocrine systems that regulate reproductively-triggered senescence and death, revealing shared mechanisms and lineage-specific adaptations.
C. elegans
Short generation times, well-established genomic tools, and visual markers of organismal fitness make C. elegans a useful model organism for probing mechanisms of reproduction-triggered senescence and longevity [6]. C. elegans also displays a noteworthy survival strategy; under stressful environmental conditions, larvae can develop into dauers, a specialized state that promotes survival and dispersal (Figure 1a) [7]. Formation of this alternate life stage is controlled by the nuclear hormone receptors, such as DAF-12, and their ligands, including the dafachronic acids [8,9,10]. Most C. elegans worms are self-fertilizing hermaphrodites (Figure 1a). Male C. elegans are rare, constituting less than 0.1% of the population [11,12]. Reproduction with males is necessary for genetic diversity, yielding larger and healthier clutches compared to self-fertilization, but it comes at a cost (Figure 1b). Exogenous sperm triggers development of severe senescent phenotypes in hermaphrodites, including uterine tumors, shrinking, pools of yolk lipoproteins in the body cavity, intestinal atrophy, and shortened lifespan. This is mediated through perturbations of the insulin/insulin-like growth factor (IGF1) signaling pathway (IIS), an evolutionarily-conserved modulator of lifespan [13,14,15,16,17,18,19]. IIS in C. elegans is mediated through DAF-2, the sole receptor for insulin-like peptides [20]. DAF-2 mutants, which exhibit reduced IIS signaling, experience increased lifespans [21]. Both DAF-2 and IIS play key roles in reproduction and life stage transitions, such as dauer formation.
Mating increases IIS and leads to downstream physiological changes in hermaphrodite worms. DAF-2 mediated inhibition of the DAF-16 transcription factor causes increased production of yolk protein through autophagy of the gut [13,16]. As eggs develop, the gut fills with pools of yolk protein. This extra yolk is expelled along with the fertilized eggs and is subsequently consumed by developing larvae [13,22]. Larvae that consume this yolk experience increased overall fitness, however, yolk overproduction lead to immediate detrimental outcomes in mated hermaphrodite worms (Figure 1b) [13,22]. Intestinal atrophy can lead to shortened lifespan, and the overproduction of yolk and depletion of self-sperm lead to teratoma-like uterine tumors [23,24]. Worms that lack functional DAF-2 or DAF-16 in the intestines exhibit less intestinal atrophy, reduced accumulation of yolk protein, and lower mortality rates as compared to mated wildtype worms [13]. Taken together, the increase in IIS caused by reproduction leads directly to gut autophagy and senescent phenotypes.
Mating also decreases lifespan through modulation of the DAF-9/DAF-12 and DAF-16 pathways [15]. Sperm decreases activity of DAF-9, a cytochrome P450 oxygenase, leading to a reduction in dafachronic acids [15,25]. Despite the decrease in dafachronic acid production, mated worms show increased activity of DAF-12, a nuclear receptor involved in osmotic regulation and longevity pathways [15,26]. Dysregulation of osmotic adaptability leads to shrinking due to water loss and shortened lifespan (Figure 1b) [15]. DAF-2 signaling leads to the cytoplasmic localization of DAF-16, causing reduction in the activity of the transcription factor and leading to a loss of fat, possibly through yolk overproduction. Loss of fat causes shrinking and is associated with reduced lifespan. The decreased DAF-9 and DAF-16 activity and increased DAF-12 activity after mating lead to fat loss, osmotic stress susceptibility, whole body shrinking, and reduced lifespan [15].
In C. elegans, reproduction presents a tradeoff between decreased longevity with increased offspring fitness. The dafachronic acid and IIS pathways that are essential in development also cause senescent phenotypes after reproduction [17,20,27]. Our understanding of how mating in worms recruits these same molecular pathways to accelerate the transition from adulthood to death provides a useful template for studying similar phenomena in other invertebrates.
Octopuses
Of the invertebrates, octopuses have one of the most complex nervous systems, with almost 40 lobes in the central brain and a brain to body mass ratio comparable to that of fish and reptiles [28]. The octopus’s many morphological and neural novelties make them advantageous for comparative neurobiological research. The optic glands, small spherical structures located between the optic lobes and central brain mass, are particularly noteworthy. Nicknamed the “self-destruct” system [29], the optic glands are the principal neuroendocrine signaling center of the octopus. Functionally analogous to the vertebrate anterior pituitary gland, the optic glands control reproductive maturation and, ultimately, reproduction-triggered death [29,30,31].
Almost all species of octopuses are semelparous: females die while brooding their singular clutch of eggs [32]. During the maternal brooding period, the female transitions from feeding to fasting, then undergoes rapid organismal decline [29,30]. Senescent females lose the ability to heal from wounds and exhibit signs of a compromised vestibulo-ocular reflex. In the period preceding death, the female may engage in self-injury behaviors, rubbing her skin off and eating the tips of her arms [29,30]. All physiological changes and maternal behaviors are controlled by secretions from the optic gland. Senescence is abolished if the gland is removed, enabling the octopus to reproduce again and live up to 6 months longer than intact counterparts [29].
Multiomics studies in Octopus bimaculoides demonstrate that mating triggers sweeping changes in optic gland neuroendocrinology. Mating upregulates the daf-36 gene, which remains elevated through the end of life; daf-36 transcript is the most abundant transcript found in the optic glands of mated females [30,33]. The daf-36 gene encodes a cholesterol 7-desaturase that converts cholesterol to 7-dehydrocholesterol (7-DHC) (Figure 2) [33,34,35]. In arthropods and nematodes, this is the first committed step in the synthesis of bioactive steroid hormones from cholesterol [34,35]. Mammals lack a homolog to daf-36 [35]. In humans, the accumulation of 7-DHC is toxic and can lead to death [36,37]. In octopuses, it is possible that 7-DHC itself, rather than its downstream metabolites, is used as a signaling molecule at the end of life. In addition, high levels of 7-DHC may be lethal, as it is in humans, contributing to the behavioral and physiological changes that lead to death. These findings demonstrate that DAF-36 and 7-DHC share neuroendocrine functions across distantly-related animals.
As in C. elegans and other animals, the IIS pathway plays an important role in mediating reproduction-triggered death in octopuses (Figure 2). When maternal fasting begins, the optic glands upregulate the production of ImpL2, a homolog to insulin-like growth factor binding protein [30]. In Drosophila, increased ImpL2 production occurs under starvation conditions and suppresses IIS activity, ultimately promoting survivorship [38]. However, in octopuses, increased ImpL2 is found only in fasting, senescent females. Non-senescent females under experimentally-induced starvation conditions have comparable ImpL2 expression as females who are fed normally. These findings suggest that different mechanisms control octopus IIS pathways under starvation and senescent conditions [39]. Increased ImpL2 also causes whole body tissue wasting in Drosophila cancer models [40,41]. This cachexia-like wasting resembles the physiological markers of senescence in octopuses, strongly suggesting that high ImpL2 promotes tissue loss at the end of life (Figure 2). Remarkably, studies of the optic gland reveal that daf-36 and insulin signaling pathways are critical to the control of lifespan after reproduction in both octopuses and C. elegans.
Although most octopuses are semelparous, Octopus chierchiae is iteroparous: males and females reproduce multiple times, and females can brood multiple clutches [42,43]. The maternal behavior of O. chierchiae offers insight into the evolution of reproduction-triggered death. O. chierchiae mothers feed for most of the brooding period. They stop eating when their eggs begin to hatch, but resume feeding again 2-3 days before hatching is complete. During this time, the female eats any young remaining in the area [43]. The behaviors and unique life history of O. chierchiae aligns with Wodinsky’s original hypothesis that the inhibition of feeding and maternal death in other octopus species serves to prevent cannibalism of the young by the mother [29].
The O. chierchiae optic glands likely rely on alternate signaling systems or compensatory mechanisms to mediate iteroparity. Transcripts that are upregulated in mated O. bimaculoides, such as the steroidogenic enzymes, may be more dynamically up- and downregulated through each cycle of mating and brooding in O. chierchiae. Additionally, the steady increase in ImpL2 production during O. bimaculoides senescence may be entirely absent in O. chierchiae. Future comparison of brooding O. chierchiae neuroendocrinology to senescent O. bimaculoides profiles will be instrumental in clarifying the functional roles of transcriptional changes in senescent octopuses.
Bumblebees
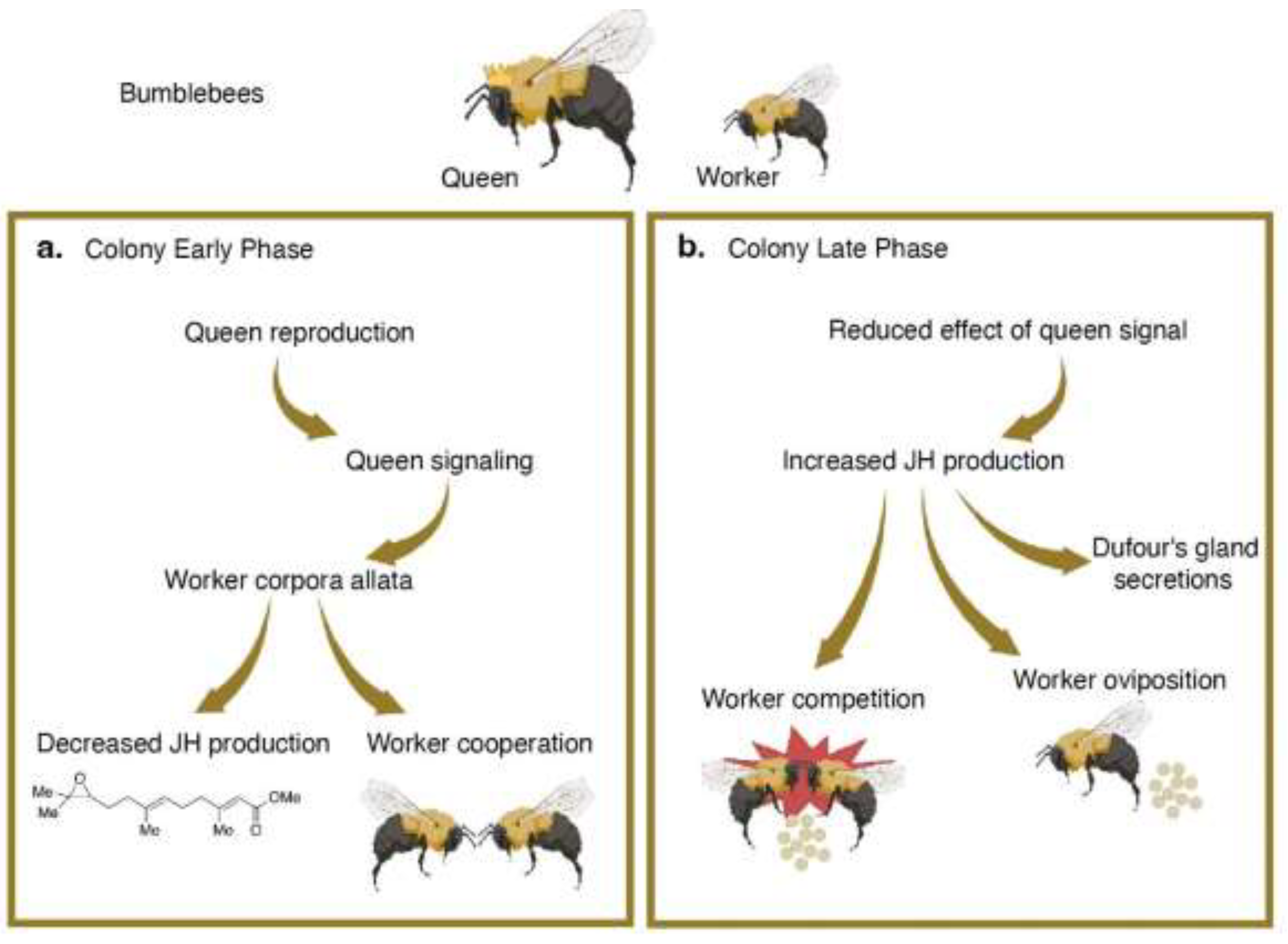
Bumblebees offer the opportunity to understand post-reproductive senescence and death on the scale of the whole colony as well as on an individual basis. In both Bombus impatiens and Bombus terrestris, colonies are founded in the spring and die by the fall [44]. In the beginning of the colony cycle, the queen only produces diploid female workers, who cooperatively perform nursing and foraging tasks to keep the colony running (Figure 3a). As the colony ages, the queen begins to produce future queens, called gynes, and haploid males. At this stage, previously cooperative workers abruptly become aggressive, competing with each other and the queen to produce males (Figure 3b) [45]. Food provisioning and other nest tasks are neglected in favor of reproduction, and few broods are successfully reared. Instead, workers remove or eat each other’s eggs, including the eggs of the queen, and attempt to oviposit. The fierce competition phase of the colony may result in the premature death of the queen [44,46,47]. At the end of the colony cycle, female workers, males, and the queen all die. Gynes are the lone survivors of the colony, undergoing diapause before initiating their own colonies the next year. Ultimately, reproductive competition destabilizes the maintenance and productivity of the colony, leading to its demise. Examining the neuroendocrinology of reproductive competition can elucidate mechanisms by which the behaviors of individuals contribute to the death of the entire social group.
In bumblebees, signaling systems in both the queen and the workers act simultaneously to modulate reproduction, aggression, and ultimately, colony longevity. Exocrine organs, including the Dufour’s gland and the mandibular glands, produce compounds that are essential for chemical communication and social identification [48,49,50]. The queen’s chemical secretions reduce reproductive output of the workers. In Bombus terrestris, this occurs through the suppression of the workers’ corpora allata, an endocrine gland that produces juvenile hormone (Figure 3a) [51,52]. Juvenile hormone controls development and metamorphosis in arthropods and serves as a major gonadotropic hormone in social insects [53,54]. High levels of juvenile hormone induce oogenesis and impacts social dominance in bumblebees. By suppressing juvenile hormone synthesis, the queen can monopolize reproduction: workers that lack juvenile hormone have underdeveloped ovaries, do not lay eggs, are less aggressive, and hold a less dominant position in worker groups (Figure 3a) [48,53].
As the colony develops, the queen to worker ratio diminishes. Faced with a high worker population, the effectiveness of the queen’s chemical secretions may also decline [46,55]. This enables individual workers to overcome reproductive suppression and take advantage of the opportunity to oviposit [56]. Reproductive workers exhibit elevated juvenile hormone levels and increased social dominance, effectively inhibiting the reproduction of other workers (Figure 3b) [57,58]. Additionally, Dufour’s gland secretions of reproductively active workers become similar to that of bumblebee queens; high juvenile hormone levels cause the removal of esters that signal reproductive sterility in nonreproductive workers [48,59,60]. This difference in chemical identity may function to protect sterile workers during the competition phase, signaling to reproductive workers that they are not a threat, or act as an ancillary chemical cue for inhibiting reproduction of other workers.
The death of the bumblebee colony as a result of reproductive competition arises from dynamic actions of hormone and pheromone signaling. The queen’s chemical secretions and reproductive behaviors are key drivers in controlling competition amongst her workers, but modulation of chemical cues in the workers themselves may also contribute to the transition from cooperativity to antagonism. For example, juvenile hormone exerts gonadotropic effects which have downstream consequences on the social milieu [48,53]. In bumblebees, rapid and socially-dynamic chemical signaling causes a stark shift from cooperativity to reproductive aggression that ultimately destroys the colony.
Conclusions
Research conducted in these three systems emphasize the highly intertwined roles that developmental and metabolism-related signaling pathways play in the control of reproduction and lifespan. In C. elegans, reproduction triggers IIS and dafachronic acid based senescent phenotypes in hermaphrodites [13]. Octopuses brood their young until dying, undergoing a dramatic decline that culminates in starvation, self-cannibalism, and self-injury [29,30]. Bumblebee colonies undergo a JH-mediated rapid decline during the production of fertile offspring [44,48].
Although the phenotypes, behaviors, or life histories discussed here may seem extreme, they only begin to represent the full spectrum of reproductive death found in invertebrates. For example, aphagous mayflies lack a fully formed gut or mouth as reproductive adults. Because mayfly adults have no opportunity to replenish energy reserves, regulation of metabolism likely plays a key role in the regulation of their reproduction and death. They mate and die rapidly, living only days to weeks as adults [5,61]. Future experimental studies might test whether administration of nutrients during the adult phase could extend lifespan, or if adults that do not reproduce tend to live longer than those who do. Reproductive cannibalism is prevalent in several species of spiders and mantids: females kill and eat their male mates [62,63,64,65,66]. In a particularly noteworthy case, the male dies without female intervention immediately upon insertion of the second pedipalp [64]. It is possible this behavior promotes the fitness of the female during reproduction, and thus increases the chances of reproductive success for both the male and female. Future research into the mechanistic basis of this phenomenon should not only examine the offspring fitness of mothers who consumed their mates, but also how the death of the male changes the reproductive success of other males. Deepening our understanding of the three animals we discuss in depth here and broadening the neuroendocrinology of reproductive death to other non-model animals will greatly increase our appreciation for the evolutionary richness and diversity of life history strategies present amongst all animals.
References
- Northcutt RG: Evolution of centralized nervous systems: two schools of evolutionary thought. Proc Natl Acad Sci U S A 2012, 109 Suppl 1:10626–10633. [CrossRef]
- Hartenstein V: The Central Nervous System of Invertebrates. In The Wiley Handbook of Evolutionary Neuroscience. Shepherd, S V. John Wiley & Sons, Ltd; 2016:173–235.
- Picard MAL, Vicoso B, Bertrand S, Escriva H: Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict. Genes (Basel) 2021, 12:1136A sweeping examination of invertebrate reproduction, focusing on sexual vs asexual reproduction and sex-determination systems. This broader analysis is a valuable tool in analyzing the implications of these systems.
- Hughes PW: Between semelparity and iteroparity: Empirical evidence for a continuum of modes of parity. Ecology and Evolution 2017, 7:8232–8261. [CrossRef]
- Finch CE: Longevity, senescence, and the genome. University of Chicago Press; 1990.
- Mack HID, Heimbucher T, Murphy CT: The nematode Caenorhabditis elegans as a model for aging research. Drug Discovery Today: Disease Models 2018, 27:3–13. [CrossRef]
- Cassada RC, Russell RL: The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Developmental Biology 1975, 46:326–342. [CrossRef]
- Albert PS, Riddle DL: Mutants of Caenorhabditis elegans that form dauer-like larvae. Developmental Biology 1988, 126:270–293. [CrossRef]
- Riddle DL, Swanson MM, Albert PS: Interacting genes in nematode dauer larva formation. Nature 1981, 290:668–671. [CrossRef]
- Mahanti P, Bose N, Bethke A, Judkins JC, Wollam J, Dumas KJ, Zimmerman AM, Campbell SL, Hu PJ, Antebi A, et al.: Comparative metabolomics reveals endogenous ligands of DAF-12, a nuclear hormone receptor, regulating C. elegans development and lifespan. Cell Metab 2014, 19:73–83.
- Chasnov JR, Chow KL: Why are there males in the hermaphroditic species Caenorhabditis elegans? Genetics 2002, 160:983–994. [CrossRef]
- Maures TJ, Booth LN, Benayoun BA, Izrayelit Y, Schroeder FC, Brunet A: Males Shorten the Life Span of C. elegans Hermaphrodites via Secreted Compounds. Science 2013, 343:541–544. [CrossRef]
- Ezcurra M, Benedetto A, Sornda T, Gilliat AF, Au C, Zhang Q, van Schelt S, Petrache AL, Wang H, de la Guardia Y, et al.: C. elegans Eats Its Own Intestine to Make Yolk Leading to Multiple Senescent Pathologies. Current Biology 2018, 28:2544-2556.e5.
- Kern CC, Srivastava S, Ezcurra M, Hsiung KC, Hui N, Townsend S, Maczik D, Zhang B, Tse V, Konstantellos V, et al.: C. elegans ageing is accelerated by a self-destructive reproductive programme. Nat Commun 2023, 14:4381. A comparison of Caenorhabditis and Pristionchus reproductive programs, focusing on presence or absence of programmed death phenotypes in other species. The findings support C. elegans reproductive death, as yolk venting and other phenotypes only occur in hermaphrodites.
- Shi C, Murphy CT: Mating Induces Shrinking and Death in Caenorhabditis Mothers. Science 2014, 343:536–540.
- DePina AS, Iser WB, Park S-S, Maudsley S, Wilson MA, Wolkow CA: Regulation of Caenorhabditis elegans vitellogenesis by DAF-2/IIS through separable transcriptional and posttranscriptional mechanisms. BMC Physiology 2011, 11:11. [CrossRef]
- Venz R, Pekec T, Katic I, Ciosk R, Ewald CY: End-of-life targeted degradation of DAF-2 insulin/IGF-1 receptor promotes longevity free from growth-related pathologies. eLife 2021, 10:e71335. Degradation of DAF-2 receptors during aging extends C. elegans lifespan beyond that of daf-2 mutants. This is notable because it delivers relief from aging phenotypes without the detrimental effects of reduced IIS signaling in early life.
- Kim S-S, Lee C-K: Growth signaling and longevity in mouse models. BMB Rep 2019, 52:70–85. [CrossRef]
- Kenyon C: The Plasticity of Aging: Insights from Long-Lived Mutants. Cell 2005, 120:449–460.
- Kimura KD, Tissenbaum HA, Liu Y, Ruvkun G: daf-2, an Insulin Receptor-Like Gene That Regulates Longevity and Diapause in Caenorhabditis elegans. Science 1997, 277:942–946.
- Kenyon C, Chang J, Gensch E, Rudner A, Tabtiang R: A C. elegans mutant that lives twice as long as wild type. Nature 1993, 366:461–464. [CrossRef]
- Kern CC, Townsend SJ, Salzmann A, Rendell NB, Taylor GW, Comisel RM, Foukas LC, Bähler J, Gems D: C. elegans feed yolk to their young in a form of primitive lactation. Nature Communications 2021, 12:5801. Demonstrates that yolk venting in C. elegans benefits the offpsring and delves further into the mechanics of IIS control of yolk production and venting.
- Zhao Y, Gilliat AF, Ziehm M, Turmaine M, Wang H, Ezcurra M, Yang C, Phillips G, McBay D, Zhang WB, et al.: Two forms of death in ageing Caenorhabditis elegans. Nat Commun 2017, 8:15458. [CrossRef]
- Wang H, Zhao Y, Ezcurra M, Benedetto A, Gilliat AF, Hellberg J, Ren Z, Galimov ER, Athigapanich T, Girstmair J, et al.: A parthenogenetic quasi-program causes teratoma-like tumors during aging in wild-type C. elegans. NPJ Aging Mech Dis 2018, 4:6. [CrossRef]
- Sharma KK, Wang Z, Motola DL, Cummins CL, Mangelsdorf DJ, Auchus RJ: Synthesis and activity of dafachronic acid ligands for the C. elegans DAF-12 nuclear hormone receptor. Molecular Endocrinology 2009, 23:640–648. [CrossRef]
- Hsin H, Kenyon C: Signals from the reproductive system regulate the lifespan of C. elegans. Nature 1999, 399:362–366. [CrossRef]
- Dumas KJ, Guo C, Wang X, Burkhart KB, Adams EJ, Alam H, Hu PJ: Functional divergence of dafachronic acid pathways in the control of C. elegans development and lifespan. Dev Biol 2010, 340:605–612. [CrossRef]
- Packard A: Cephalopods and Fish: The Limits of Convergence. Biological Reviews 1972, 47:241–307. [CrossRef]
- Wodinsky J: Hormonal Inhibition of Feeding and Death in Octopus: Control by Optic Gland Secretion. Science 1977, 198:948–951.
- Wang ZY, Ragsdale CW: Multiple optic gland signaling pathways implicated in octopus maternal behaviors and death. Journal of Experimental Biology 2018, 221:jeb185751. [CrossRef]
- Wells MJ, Wells J: Pituitary Analogue in the Octopus. Nature 1969, 222:293–294. [CrossRef]
- Rocha F, Guerra A, González AF: A review of reproductive strategies in cephalopods. Biol Rev Camb Philos Soc 2001, 76:291–304. [CrossRef]
- Wang ZY, Pergande MR, Ragsdale CW, Cologna SM: Steroid hormones of the octopus self-destruct system. Current Biology 2022, 32:2572-2579.e4. Characterizes the steroid secretions from the optic gland, identifying cholesterol metabolites that are upregulated at the end of life. The findings provide evidence for daf-36 enzyme activity leading to the increased production of 7-DHC in mated octopuses.
- Gerisch B, Rottiers V, Li D, Motola DL, Cummins CL, Lehrach H, Mangelsdorf DJ, Antebi A: A bile acid-like steroid modulates Caenorhabditis elegans lifespan through nuclear receptor signaling. Proceedings of the National Academy of Sciences 2007, 104:5014–5019. [CrossRef]
- Yoshiyama-Yanagawa T, Enya S, Shimada-Niwa Y, Yaguchi S, Haramoto Y, Matsuya T, Shiomi K, Sasakura Y, Takahashi S, Asashima M, et al.: The Conserved Rieske Oxygenase DAF-36/Neverland Is a Novel Cholesterol-metabolizing Enzyme*. Journal of Biological Chemistry 2011, 286:25756–25762.
- Porter FD: Smith–Lemli–Opitz syndrome: pathogenesis, diagnosis and management. Eur J Hum Genet 2008, 16:535–541.
- Ryan AK, Bartlett K, Clayton P, Eaton S, Mills L, Donnai D, Winter RM, Burn J: Smith-Lemli-Opitz syndrome: a variable clinical and biochemical phenotype. Journal of Medical Genetics 1998, 35:558–565. [CrossRef]
- Honegger B, Galic M, Köhler K, Wittwer F, Brogiolo W, Hafen E, Stocker H: Imp-L2, a putative homolog of vertebrate IGF-binding protein 7, counteracts insulin signaling in Drosophila and is essential for starvation resistance. Journal of Biology 2008, 7:10. [CrossRef]
- Bian L, Li F, Chang Q, Liu C, Tan J, Zhang S, Li X, Li M, Sun Y, Xu R, et al.: Comparison of Behavior, Histology and ImpL2 Gene Expression of Octopus sinensis Under Starvation and Senescence Conditions. Frontiers in Marine Science 2022, 9. [CrossRef]
- Figueroa-Clarevega A, Bilder D: Malignant Drosophila tumors interrupt insulin signaling to induce cachexia-like wasting. Developmental Cell 2015, 33:47–55.
- Kwon Y, Song W, Droujinine IA, Hu Y, Asara JM, Perrimon N: Systemic Organ Wasting Induced by Localized Expression of the Secreted Insulin/IGF Antagonist ImpL2. Developmental Cell 2015, 33:36–46.
- Grearson AG, Dugan A, Sakmar T, Sivitilli DM, Gire DH, Caldwell RL, Niell CM, Dölen G, Wang ZY, Grasse B: The Lesser Pacific Striped Octopus, Octopus chierchiae: An Emerging Laboratory Model. Frontiers in Marine Science 2021, 8. Examines O. chierchiae through its life cycle under laboratory mariculture conditions, providing first detailed descriptions of iteroparous brooding behaviors.
- Rodaniche AF: Iteroparity in the Lesser Pacific Striped Octopus Octopus Chierchiae (Jatta, 1889). Bulletin of Marine Science 1984, 35:99–104.
- Goulson D: Bumblebees: Behaviour, Ecology, and Conservation. Oxford University Press; 2010.
- Duchateau MJ, Velthuis HHW: Development and Reproductive Strategies in Bombus Terrestris Colonies. Behaviour 1988, 107:186–207.
- Amsalem E, Grozinger CM, Padilla M, Hefetz A: Chapter Two - The Physiological and Genomic Bases of Bumble Bee Social Behaviour. In Advances in Insect Physiology. Edited by Zayed A, Kent CF. Academic Press; 2015:37–93.
- Princen SA, Van Oystaeyen A, van Zweden JS, Wenseleers T: Worker dominance and reproduction in the bumblebee Bombus terrestris: when does it pay to bare one’s mandibles? Animal Behaviour 2020, 166:41–50.An analysis of the colony-wide effect of the queen and reproductive workers on overall worker reproduction. The authors show that reproduction is controlled largely through dominance of the queen or of dominant workers.
- Shpigler H, Amsalem E, Huang ZY, Cohen M, Siegel AJ, Hefetz A, Bloch G: Gonadotropic and Physiological Functions of Juvenile Hormone in Bumblebee (Bombus terrestris) Workers. PLOS ONE 2014, 9:e100650. [CrossRef]
- Krieger GM, Duchateau M-J, Van Doorn A, Ibarra F, Francke W, Ayasse M: Identification of Queen Sex Pheromone Components of the Bumblebee Bombus terrestris. J Chem Ecol 2006, 32:453–471. [CrossRef]
- Amsalem E, Orlova M, Grozinger CM: A conserved class of queen pheromones? Re-evaluating the evidence in bumblebees (Bombus impatiens). Proceedings of the Royal Society B: Biological Sciences 2015, 282:20151800. [CrossRef]
- Röseler P-F, Röseler I, van Honk CGJ: Evidence for inhibition of corpora allata activity in workers ofBombus terrestris by a pheromone from the queen’s mandibular glands. Experientia 1981, 37:348–351. [CrossRef]
- Van Honk CGJ, Velthuis HHW, Röseler P-F, Malotaux ME: THE MANDIBULAR GLANDS OF BOMBUS TERRESTRIS QUEENS AS A SOURCE OF QUEEN PHEROMONES. Entomologia Experimentalis et Applicata 1980, 28:191–198. [CrossRef]
- Pandey A, Motro U, Bloch G: Juvenile hormone interacts with multiple factors to modulate aggression and dominance in groups of orphan bumble bee (Bombus terrestris) workers. Hormones and Behavior 2020, 117:104602. Examines the effect of decreased juvenile hormone levels on egg laying, dominance, and aggression in bumblebee workers. In groups of worker bees outside the colony, reduction in juvenile hormone leads to lower aggressiveness, less egg laying, and less dominance. Juvenile hormone reduction does not, however, change existing hierarchies.
- Hefetz A, Grozinger CM: Hormonal Regulation of Behavioral and Phenotypic Plasticity in Bumblebees. In Hormones, Brain and Behavior. . Elsevier; 2017:453–464.
- Bloch G, Borst DW, Huang Z-Y, Robinson GE, Hefetz A: Effects of social conditions on Juvenile Hormone mediated reproductive development in Bombus terrestris workers. Physiological Entomology 1996, 21:257–267. [CrossRef]
- Amsalem E, Hefetz A: The appeasement effect of sterility signaling in dominance contests among Bombus terrestris workers. Behav Ecol Sociobiol 2010, 64:1685–1694. [CrossRef]
- Amsalem E, Shamia D, Hefetz A: Aggression or ovarian development as determinants of reproductive dominance in Bombus terrestris: interpretation using a simulation model. Insect Soc 2013, 60:213–222. [CrossRef]
- Bloch G, Hefetz A: Regulation of reproduction by dominant workers in bumblebee (Bombus terrestris) queenright colonies. Behav Ecol Sociobiol 1999, 45:125–135. [CrossRef]
- Derstine NT, Villar G, Orlova M, Hefetz A, Millar J, Amsalem E: Dufour’s gland analysis reveals caste and physiology specific signals in Bombus impatiens. Sci Rep 2021, 11:2821. Dufour's gland is responsible for pheromone secretions in bumblebees. The authors characterize Dufour's gland secretions, identifying compounds specific to reproductive status.
- Amsalem E, Twele R, Francke W, Hefetz A: Reproductive competition in the bumble-bee Bombus terrestris: do workers advertise sterility? Proc Biol Sci 2009, 276:1295–1304. [CrossRef]
- Carey JR: Longevity minimalists: life table studies of two species of northern Michigan adult mayflies. Experimental Gerontology 2002, 37:567–570. [CrossRef]
- Arnqvist G, Henriksson S: Sexual cannibalism in the fishing spider and a model for the evolution of sexual cannibalism based on genetic constraints. Evolutionary Ecology 1997, 11:255–273. [CrossRef]
- Roggenbuck H, Pekár S, Schneider JM: Sexual cannibalism in the European garden spider Araneus diadematus: the roles of female hunger and mate size dimorphism. Animal Behaviour 2011, 81:749–755. [CrossRef]
- Foellmer MW, Fairbairn DJ: Spontaneous male death during copulation in an orb-weaving spider. Proc R Soc Lond 2003, 270:S183-185. [CrossRef]
- Barry KL, Holwell GI, Herberstein ME: Female praying mantids use sexual cannibalism as a foraging strategy to increase fecundity. Behavioral Ecology 2008, 19:710–715. [CrossRef]
- Burke NW, Holwell GI: Male coercion and female injury in a sexually cannibalistic mantis. Biology Letters 2021, 17:20200811. [CrossRef]
Figure 1.
Overview of senescent phenotypes induced by reproduction in C. elegans. a. Dauers are an alternate developmental stage that promotes survival under stressful conditions. Most C. elegans worms are self-fertilizing hermaphrodites. b. Reproduction leads to intestinal atrophy, increased vitellogenesis, and shrinking.
Figure 1.
Overview of senescent phenotypes induced by reproduction in C. elegans. a. Dauers are an alternate developmental stage that promotes survival under stressful conditions. Most C. elegans worms are self-fertilizing hermaphrodites. b. Reproduction leads to intestinal atrophy, increased vitellogenesis, and shrinking.
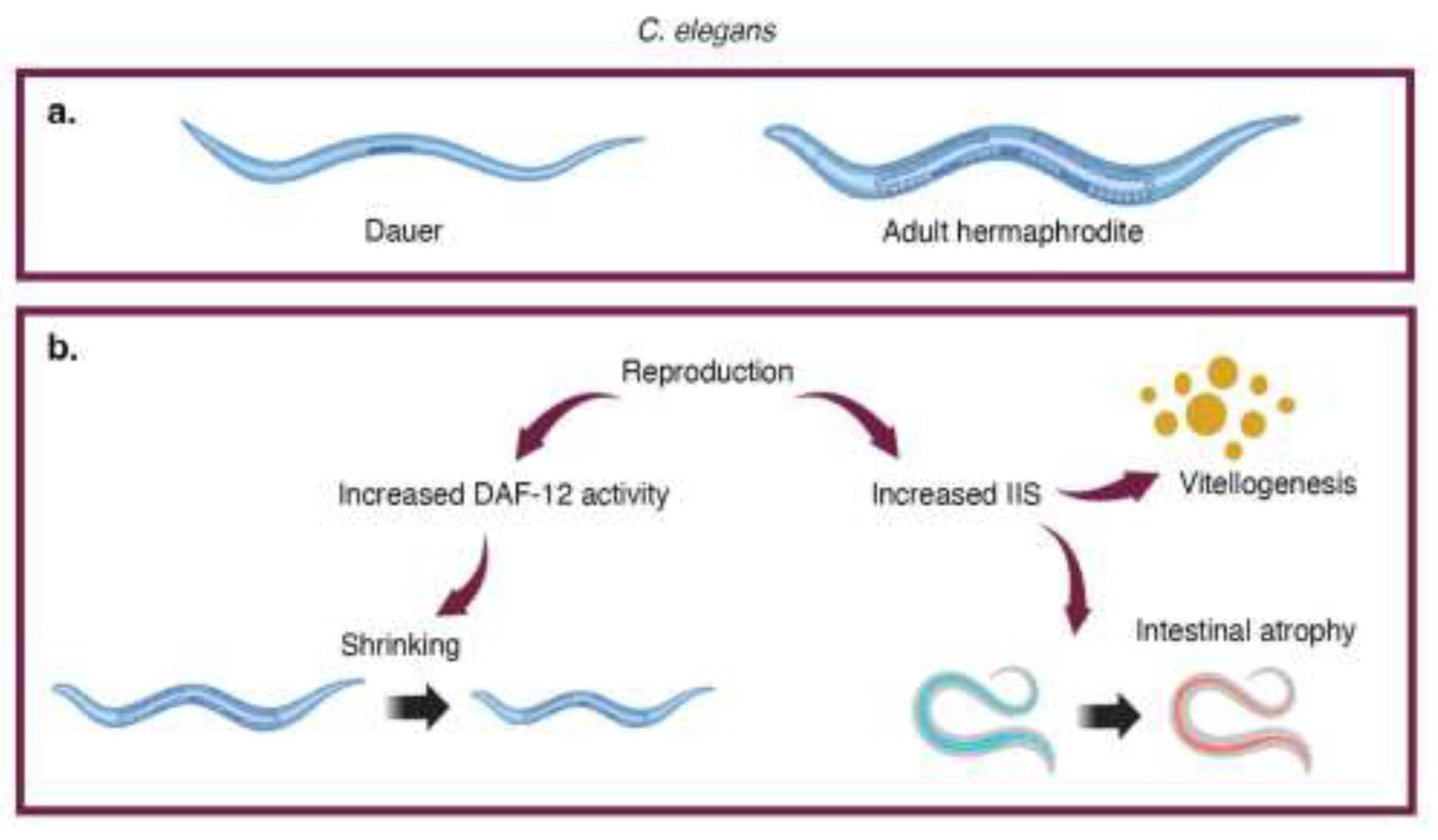
Figure 2.
Effects of reproductive senescence in octopuses. The daf-36 and IIS pathways are involved in reproduction-triggered death in octopuses.
Figure 2.
Effects of reproductive senescence in octopuses. The daf-36 and IIS pathways are involved in reproduction-triggered death in octopuses.
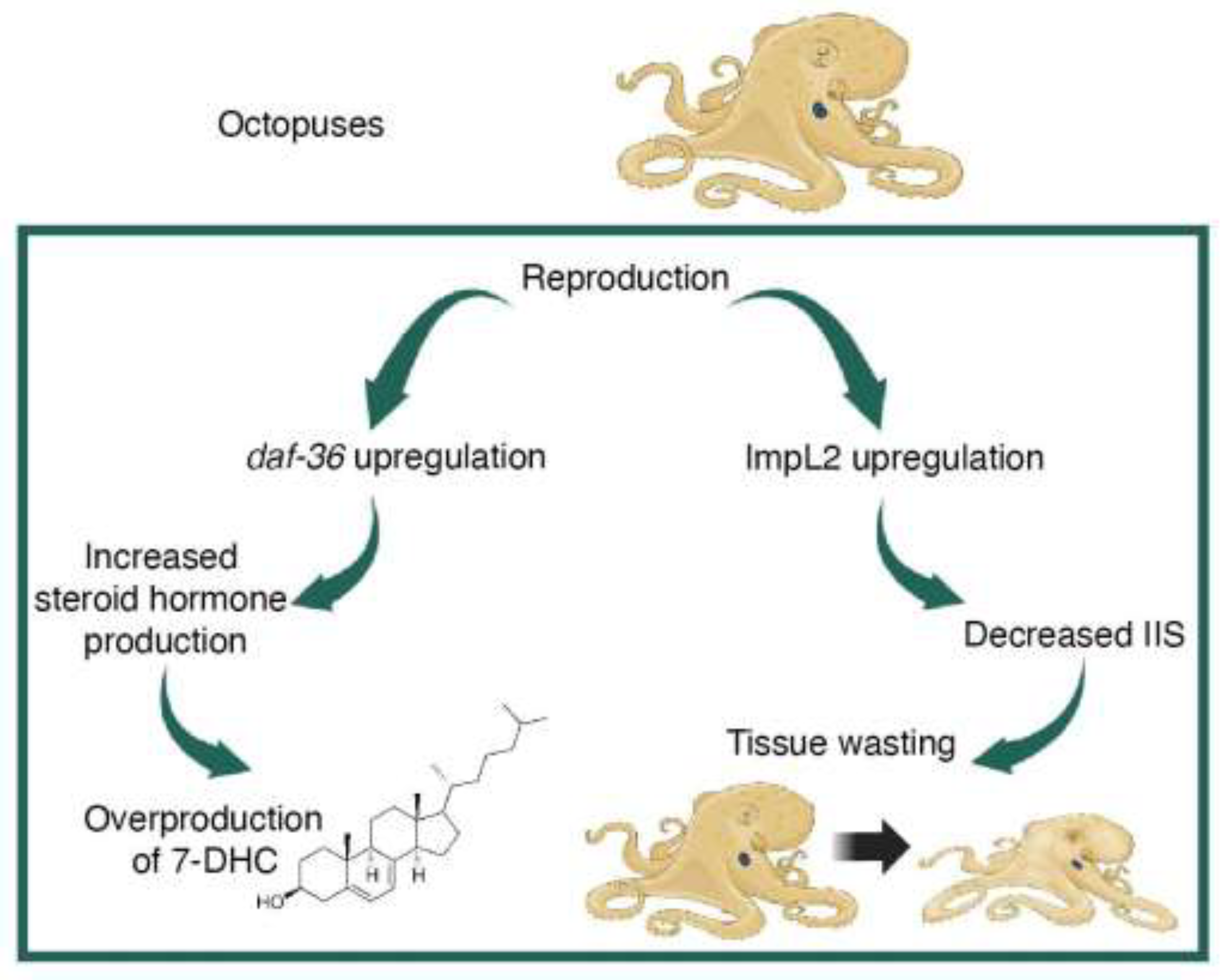
Figure 3.
Colony death in bumblebees. a. In the early phase of the colony, workers are cooperative and non-reproductive. b. In the late phase of the colony, changes in queen signaling and worker JH production cause workers to exhibit overt antagonism with each other and the queen as they compete to lay eggs.
Figure 3.
Colony death in bumblebees. a. In the early phase of the colony, workers are cooperative and non-reproductive. b. In the late phase of the colony, changes in queen signaling and worker JH production cause workers to exhibit overt antagonism with each other and the queen as they compete to lay eggs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



