Submitted:
13 February 2025
Posted:
14 February 2025
You are already at the latest version
Abstract
Aging is an extensive biological process characterized by morphological and functional alterations in cellular and extracellular components, resulting in a systematic decline in biological functions ultimately leading to death. Although substantial advancements have been made in manipulating lifespan in model organisms like C. elegans and mice through genetic, dietary, and pharmacological means, the fundamental mechanisms driving aging in humans remain elusive and widely debated. In addition, there is no comprehensive computational platform capable of making predictions on aging in multicellular systems and integrating the multiscale competency of lifeforms. We focus on the processes that build and maintain a complex anatomy toward a specific target morphology, and propose the hypothesis that aging arises even in the absence of accumulated cellular or genetic damage, because a homeodynamic system left without any goal in anatomical morphospace will start degrading. This can occur in biological systems because evolution typically prioritizes development over morphostasis, leaving organisms with limited reinforcement of anatomical goals after development. Using an in silico model of homeostatic morphogenesis with a multiscale competency architecture and information dynamics analysis, we find: (1) Absence of Long-Term Morphostasis: Aging emerges naturally after development due to the lack of an evolved regenerative goal, rather than just specific detrimental properties of developmental programs (e.g., antagonistic pleiotropy or hyperfunction); (2) Acceleration Factors vs. Root Cause: Cellular misdifferentiation, reduced competency, communication failures, and genetic damage all accelerate aging but are not its primary cause; (3) Information Dynamics in Aging: Aging correlates with increased active information storage and transfer entropy, while spatial entropy measures distinguish two dynamics—loss of structure and morphological noise accumulation; (4) Dormant Regenerative Potential: Despite organ loss, spatial information persists in the cybernetic tissue, indicating a memory of lost structures, which can be reactivated for organ restoration through targeted regenerative information; and (5) Optimized Regeneration Strategies: Restoration is most efficient when regenerative information includes differential patterns of affected cells and their neighboring tissue, highlighting strategies for rejuvenation. These findings provide a novel perspective on aging dynamics with significant implications for longevity research and regenerative medicine.
Keywords:
aging
; morphogenesis
; cognition
; NCA
; computational modeling
; in silico
1. Introduction
1.1. Theories of Aging
Aging is a system-level, near-ubiquitous biological process characterized by morphological and functional alterations in cellular and extracellular components, resulting in a systematic decline in biological functions [1]. This degenerative process is modulated by genetics, lifestyle, and environmental factors [2]. As cellular functionality diminishes with age, there is a corresponding decrease in physiological capacities and an enhanced vulnerability to a spectrum of diseases, including cardiovascular disorders, cancer, neurodegenerative diseases, type II diabetes, and various infections. These conditions collectively compromise the quality of human life [2,3]. fig:introduction:motivation (A) schematically illustrates the typical life cycle of a complex organism comprising various stages from embryogenesis through maturity, decline, aging, and eventual death.
Several theories have been proposed to explain the aging process, that we can classify into damage-based and programmatic theories [1,4,5], with the former being more extensively studied [1,3,6,7]. Damage-based theories assert that aging results from the accumulation of molecular damage, impacting crucial cellular components like the genome, telomeres, mitochondria, and proteins, driven by imperfect repair mechanisms [6,7,8]. Conversely, programmatic theories suggest that aging is genetically predetermined, or quasi-programmed and encoded within the genome, not merely due to stochastic accumulation of damage [9,10,11]. Here, we formulate and computationally simulate a fundamentally different hypothesis based on the nature of morphogenesis as a homeodynamic system working toward anatomical setpoints: that aging is the result of a proto-cognitive disfunction among the cellular collective, and does not require damage, or evolved antagonistic mechanisms in order to occur. Instead, we propose that, as observed in animal behavior [12], cellular living systems without goals to pursue tend to degrade. Our hypothesis is that aging is the result of intrinsic dynamics of complex cybernetic (goal-directed) systems, not caused by external noise or damage. Nevertheless, it is consistent with other theories of aging [13,14] which propose additional dynamics that can lead to loss of setpoints of anatomical homeostasis, and can be accelerated by various kinds of damage.
Antagonistic pleiotropy posits that certain genes have beneficial effects in early life but detrimental effects in later life. These genes confer advantages such as increased fertility and resilience in youth, which are critical for survival and reproduction. However, as an individual ages, the same genes contribute to aging and decline because their negative effects accumulate. The theory suggests that natural selection favors these genes because of their early-life benefits, despite their harmful subsequent impacts. This results in a trade-off between early benefits and later costs [15]. Another related idea is the hyperfunction theory, which suggests that aging is not primarily a result of accumulating molecular damage or genetic programming, but rather due to a continuous and excessive operation of cellular growth processes that were beneficial during the earlier stages of life [13,14]. Central to this theory is the role of the Target of Rapamycin (TOR) pathway, which is a major regulator of growth and metabolism in cells. During the growth phase of an organism’s life, the TOR pathway promotes cellular and organismal growth by driving processes like protein synthesis, nutrient uptake, and cell division. These functions are vital for development and reproductive success. However, aging arises when these growth-promoting activities are not appropriately down-regulated after the organism has passed its reproductive prime. Instead of adjusting to a maintenance mode that would be more suitable for the non-reproductive phase of life, the TOR pathway continues to stimulate cellular processes actively. This ongoing activity can lead to “hyperfunction”, where cellular components and systems are overstimulated [13,14]. It is not clear, in that scenario, why the efficient processes of morphological homeostasis [16,17] do not orchestrate these active processes toward upkeep of functional anatomy (c.f., fig:introduction:motivation (B,C)).
Our approach is consistent with the antagonistic pleiotropy and hyperfunction models in that we also propose that fundamental morphogenetic processes are involved in aging. However, we do not focus on specific genes with early vs. late roles, but on the role that large-scale morphogenetic plasticity plays in establishing and maintaining bodies and the possible disorders of such cybernetic (goal-driven) processes. We and others have emphasized the indirect link [18,19,20] between genotype and phenotype: not just complexity and pleiotropy, but the fact that the morphogenetic layer has active homeodynamic capabilities that enable collections of cells to be aligned with respect to setpoints in the space of anatomical possibilities [16,21,22] (c.f., Figure 1 (B,C)). Living materials must navigate the space of possible organ and tissue arrangements as needed, starting from diverse initial states – single cells (egg-based development), a different body (in the case of metamorphosis), or a damaged body (in the case of regeneration). In all of these cases, drastic growth and remodeling must occur while failing or no longer needed sub-components must be replaced on-the-fly. This process has been described as a collective intelligence [23,24], because of its problem-solving capacities; like other advanced intelligences, it will be subject to specific failure modes whose origin is not organic disease or damage, but the informational dead end of completing one’s mission and not having a new setpoint toward which to strive.
We formulate, and simulate the consequences of, the hypothesis that the aging phenotype results from morphogenetic processes ceasing their targeted navigation of anatomical morphospace, after development is completed. But adulthood, and the rigors of daily life, are precisely when these processes are needed, in the context of stress, degeneration, and cell replacement, to maintain the Ship of Theseus that is the body. What will bind the cellular components of a body toward common paths in anatomical space after their goals have been reached and the body built? Tissue renewal will continue locally, for a time, but the thing that bound individual cells into a coherent “embryo” – a commitment to a specific navigation path in anatomical space – disappears at the global scale. Damage-based theories of aging are thus hypothesized to be secondary: they induce aging to the extent that they rob cells of clear and consistent anatomical and histological setpoints for which to aim. But the primary new proposal here is that complex goal-driven systems can experience a kind of intrinsic disorganization after they have met their goal. In the absence of strong effort toward targets in morphospace, the cellular collective begins to disband, giving way to systemic disease states such as degeneration and cancer – the inevitable outcome of a body that increasingly becomes a group of individual cells, not a unified higher-order morphogenetic agent [25].
1.2. Knowledge Gaps
Recent research suggests a shift in our understanding of aging, proposing that it results from changes in biological information processing rather than solely from cellular and molecular damage. This concept, linked to a malfunction in the “software of life”, focuses on how aging processes are governed by the epigenetic changes, either bioelectrical or chromatin modifications [5,26,27]. The differing dynamics, capabilities, and failure modes of the molecular layer and the physiological computational layers have long been studied in neuroscience, but are only now beginning to be related to areas such as developmental biology and cancer [28,29,30]. Novel computational frameworks are needed to begin to integrate these ideas in aging and longevity research. Although substantial advancements have been made in extending lifespan in simple model organisms through genetic, dietary, and pharmacological means [2,31,32], the fundamental mechanisms driving aging in humans remain elusive and widely debated. Current life extension strategies show limited effectiveness [33]. Much of the research has adopted a bottom-up approach, focusing on the biological hardware, the cellular components such as genes, molecular pathways, and, more recently, the epigenome. However, managing aging at such a micro-level may not be practical for complex structures like human anatomy [17,34,35]. This suggests that a broader, possibly more integrated approach may be necessary in order to fully understand and effectively intervene in the aging process.
Computational modeling has proven useful in the past to help characterize dynamics guiding complex morphogenesis [36,37,38,39,40,41,42,43,44]. However, to the best of our knowledge, there is currently no computational model in the literature explicating the aging process as a loss of goal-directedness, or directionality within the morphogenetic control system. The field is mostly dominated by experimental approaches and focused on the mathematical and computational modelling of a specific hallmark of aging [6], network approaches to aging or the impact of the environment on aging [45,46,47,48]. One prior study utilizes, similar to our model, a neural cellular automaton (see next section) to perform morphogenesis: with gradient-based deep learning, the self-regulatory interaction dynamics of distributed cellular agents have been optimized to grow a target morphology, fully self-orchestrated. However, without explicitly stabilizing morphogenesis by additionally training the cellular agents on regenerative tasks, anatomies kept growing uncontrollably after development. Thus, the study focused on an adversarial approach to stabilize self-orchestrated morphogenesis rather than studying aging and its causes in such a multi-cellular tissue per se [49].
Existing mathematical models have not yet captured the biological signaling dynamics in aging tissue or revealed the underlying information dynamics that drive aging and the progressive degradation of anatomy over time. Unlike previous approaches, we uniquely address this gap by developing an evolutionary framework that integrates both molecular and systemic aspects of aging within a multi-competency architecture. Our model demonstrates how cells, trained solely through evolution to execute developmental tasks, inherently lack a regenerative goal post-development. By simulating this dynamic, we reveal that aging arises from the absence of targeted navigation in anatomical morphospace after development, offering a novel perspective on the root cause of aging. Moreover, our approach uncovers the potential for dormant regenerative programs to be reactivated, enabling targeted restoration of anatomical structures (c.f., fig:introduction:motivation).
1.3. Biological Architecture of the Model
Morphogenesis is the result of multicellular collectives acting together to achieve a specific endpoint within the space of anatomical possibilities [19] (see fig:introduction:motivation). This process is driven by intercellular communication and information processing, where cells use different communication channels (biochemical, biomechanical or bioelectrical) to exchange information with their neighbors in a tissue [21,50] and implement a process of anatomical homeostasis where deviations from the species-specific target morphology are progressively minimized by active changes in growth and form. This enables cells to adaptively respond in a context-sensitive manner and regulate their behavior through local signaling protocols while collectively following a system-level agenda to grow and maintain an organism’ species-specifis anatomical form in a self-orchestrated way. Thus, the development of embryonic tissues, the growth and remodeling of adult organs, and even the suppression of cancer and aging can be understood as morphological computation [50]. In this sense, morphogenesis can be seen as an intermediate layer of physiological computation between genetic variation and a mature anatomical phenotype (see fig:introduction:motivation (C), significantly effecting the process of evolution [19,41].
Another important aspect of biology, so far neglected in the models of aging is multiscale competency [19,23,51] (see fig:introduction:motivation (B, C)). Biological systems implement an architecture in which context-sensitive problem-solving operates across various levels, from molecular networks to individual cells, tissues, organs, organisms, and even collective groups such as swarms. This structure allows biological entities to solve problems within specific domains and problem spaces relevant to each scale (metabolic, physiological, anatomical, and behavioral state spaces), contributing to the overall adaptability and resilience of life [21,50,52]. At the molecular and cellular level, biological units navigate metabolic spaces to remain within adaptive regions despite changing circumstances. At higher organizational levels, these units form tissues and organs, which tackle more complex and larger-scale physiological and morphological problems. For instance, cells can make autonomous decisions to maintain local balance while contributing to larger-scale biological processes like tissue repair or development and how higher levels of organization constrain and facilitate the behavior of their parts [19,53,54,55,56,57,58]. This enables organisms to adapt to changing conditions and maintain functionality even when faced with disruptions, as observed in regulative development and regeneration where cellular activity is managed toward specific anatomical outcomes despite injury, changes of cell number and size, etc. [22]. This is critical because resistance to both cancer and aging must involve upkeep of complex structures and a resistance to their progressive decline within the Ship of Theseus of the body [59].
A key advantage of this multiscale architecture is that each level is capable of addressing its own problem-spaces without needing constant guidance from higher organizational structures (local homeodynamic competency, see fig:introduction:motivation (C)). This decentralized problem-solving approach ensures that different biological processes, from cell division to behavioral adaptation, occur simultaneously and efficiently. Furthermore, this structure allows biological systems to be both robust and flexible, as failures at one level can often be compensated for by others. Additionally, the interplay between levels within this architecture enables the emergence of complex properties that cannot be explained by examining components in isolation. For example, the collective behavior of cells during processes like regeneration or morphogenesis illustrates how local interactions scale up to contribute to the organism’s overall form and function [23]. Similarly, defects of collective behavior can lead to developmental defects or aging. These interactions are not merely a sum of their parts; rather, they represent emergent capabilities that arise from the coordinated effort of biological subunits. The relative autonomy of layers in biology must surely impact on evolutionary and physiological constraints and drivers of aging, making it essential to study their properties with respect to the lifespan of individuals and evolutionary lineages.
This hierarchical but integrated organization is essential for the evolution of complex life forms. It promotes adaptability, allowing organisms to respond to environmental changes, evolve new features, and develop intricate systems that support survival and reproduction. Moreover, this multiscale competency architecture provides a framework for innovation in fields such as synthetic biology and regenerative medicine [24,34,60], where understanding and harnessing these principles could lead to breakthroughs in tissue engineering, organismal repair and aging [24,26]. This aspect of living forms – the critical layer of computational, problem-solving competency that lies between the genotype and the phenotype is an essential component of any theory of aging that integrates evolutionary and physiological dynamics to understand this complex phenomenon. Thus, we focused our computational model on the role of multiscale morphogenetic competency in the aging process.
1.4. Our Computational Model
Recently [41], we quantitatively investigated in silico the influence of varying levels of cellular competency of a multi-scale competency architecture [19,50] on the evolutionary process. We utilized tools from Artificial Life [61], a field of research focused on developing computational and cybernetic models that exhibit life-like behavior based on ideas derived from biology; prominent examples are Cellular Automata [62,63,64,65] such as Conway’s Game of Life [66]. Specifically, we deployed Neural Cellular Automata [67] (NCAs) as a model for evolving morphogenesis (see Figure 2 (A-C)): NCAs comprise a regular spatial grid of locally interacting cellular agents which maintain a numerical state vector that is regulated by an internal, cell-specific Artificial Neural Network (ANN). In that way, the cells of an NCA can be trained to perform a collective self-orchestrated system-level task, such as morphogenesis [39,40,41] or even collective navigation behavior [51,68], solely via successive perceptions of their local environment, i.e.,, reading neighboring cell state information, and computing corresponding actions to regulate their own states in accordance with a system-level agenda.
Thus, NCAs exhibit striking similarities with the genome- and collective intelligence-based multi-scale competency architectures of biological life [19,20,51,69]: the instructions of how to grow an organism from the bottom up is encoded in its genome, corresponding to the NCA’s (functional) ANN parameters. Moreover, through subsequent localized perception-action cycles of their ANN-based unicellular agents, NCAs model the intercellular communication- and intracellular information processing pathways of the adaptive and flexible internal decision-making mechanisms of biological cells: while biological cells regulate their gene expressions via gene regulatory networks, the cells of an NCA regulate their cell types via their internal ANN, and while biological cells utilize bio-electrical and molecular cues for intercellular signaling, cells in an NCA exchange numerical state information alike. In both cases, such communication pathways are localized between neighboring cells while the collective of cells may still follow a global agenda to self-orchestrate the process of morphogenesis and thereby give rise to the anatomical form of a system-level organism. Here, we use ANNs to model the memory, decision-making, and integrated information processes operating inside of cells [70,71,72,73,74].
Following our previous work [41], we deploy a swarm of virtual unicellular agents on the spatial grid of an NCA. Each agent hosts a decision-making machinery modeled by an ANN (notably, identical ANNs are deployed for each cell throughout a particular NCA instance), enabling the cellular agents to perceive neighboring cell states and independently propose cell-state regulatory actions, as illustrated in fig:introduction:simulation (A, B) and detailed below. Furthermore, we utilize evolutionary algorithms (EAs) [75] to optimize the parameters of NCAs, comprising structural and functional parts [41], such that the cellular agents evolve to self-assemble a particular spatial target pattern of predefined cell states in a fixed number of developmental steps, solely based on local communication rules, see fig:introduction:simulation (C).
Similar to ref. [41], we evolve NCAs to perform morphogenesis and morphostasis tasks, here specifically of a smiley-face-pattern reminiscent of the bioelectric craniofacial prepattern that establishes the rough outlines of the amphibian face during early development7 [76]. We deploy the evolved NCA solution for much longer times compared to the developmental stage at which the evolutionary selection occurs (c.f., fig:introduction:simulation (C, D)) and study the NCA’s long-term behavior with information-dynamic measures such as Active Information Storage (AIS)[77], Transfer Entropy (TE) [78], and Spatial Entropy (SE) . This parallels a biological an organism’s ability to maintain its physiological integrity via multiscale homeodynamic competencies for much longer than necessary for genetic reproduction. In turn, this may lead to an accumulation of a variety of entropically and self-regulatory favored morphological changes that are not directly fatal but potentially increasingly dysfunctional, changing the setpoint of the unicellular ’agents’ self-regulatory scheme and leading to misdifferentation (c.f., fig:introduction:simulation (D)).
Moreover, based on such virtual organisms, we here investigate in silico the information-dynamic implications of temporally affected competencies or information loss at the cellular level of a multi-cellular collective whose parts pursue a system-level agenda. To systematically interfere with the cellular competencies in our NCA model over time, we deploy four different strategies: We (a) increasing the misdifferentiation rate of cell-state expressions, (b) reduce the cellular agent’s ability to self-regulate their respective states by corrupting the reliability at which cellular actions are actually considered in the NCA’s cell state updates, (c) reduce the connectivity between neighboring cells reminiscent of closing an increasing number of gap junctions (GJs) to neighbors, and (d) accumulate genetic damage to the NCA’s functional parameters by randomly modifying the corresponding ANN parameters with Gaussian noise successively throughout the lifetime of our virtual organisms.
Thus, we relate degrading competencies and temporal information loss at the cellular level to aging at the system level. Finally, based on the hypothesis of the information-dynamical implications of aging caused by decaying cellular competencies, we demonstrate - in an silico minimal model - a reliable way of rejuvenation based on targeted interventions at the cellular- or organ level - of facial features.
We suggest that even in the absence of accumulated cellular defect of information processing and communication or genetic damage, a biological system left without any new goal in the morphospace will start degrading anatomically (and that this situation is allowed by evolution because it typically prioritizes development over long-term morphostasis, leaving organisms without any goal in the morphospace after development). Using an in silico model of homeostatic morphogenesis with a multiscale competency architecture and information dynamics analysis, we find: : (1) Absence of Long-Term Morphostasis: Aging emerges naturally after development due to the lack of an evolved regenerative goal, rather than being caused by specific detrimental properties of developmental programs (e.g., antagonistic pleiotropy or hyperfunction); (2) Acceleration Factors vs. Root Cause: Cellular misdifferenciation, reduced competency, communication failures, and genetic damage all accelerate aging but are not its primary cause; (3) Information Dynamics in Aging: Aging correlates with increased active information storage and transfer entropy, while spatial entropy measures distinguish two dynamics, (i() the loss of structure and (ii) morphological noise accumulation; (4) Dormant Regenerative Potential: Despite organ loss, spatial information persists in our cybernetic tissue. This indicates a memory of lost structures, which can be reactivated for organ restoration through targeted regenerative information; and (5) Optimized Regeneration Strategies: Restoration is most efficient when regenerative information includes differential patterns of affected cells and their neighboring tissue, highlighting strategies and protocols for rejuvenation. These findings provide a novel perspective on aging dynamics with significant implications for longevity research and regenerative medicine.
2. Material And Methods
2.1. Neural Cellular Automaton: A Multi-Agent Model for Morphogenesis, and Aging?
Cellular Automata (CAs) were initially introduced by von Neumann to study self-replicating machines [62]. Since then, they have become widely used as simple models for Artificial Life [61]. The core concept behind CAs revolves around maintaining a discrete spatial grid of cells, each individual cell i being assigned a binary, numerical, or even vector-valued state at every time step .
The evolution of these cell states over time follows local update rules. Specifically, we have that , where is a function of the current state of cell i, i.e., , and of the states of its neighboring cells, which we collect in the matrix .
Despite their typically simple and predefined update rules, CAs often exhibit complex dynamics (c.f., Conway’s Game of Life [66]) and have even been employed for universal computation tasks, as in Wolfram’s rule 110[63,64].
Neural Cellular Automata (NCAs) [67] can be seen as an extension to “traditional” CAs. In this context, the local update rule is replaced with a more flexible ANN, , where denotes the set of trainable parameters of the ANN, (see, e.g., Appendix A in Ref. [41]). By employing Machine Learning (ML) techniques, NCAs have been utilized to perform tasks such as self-orchestrated pattern formation [39], the simultaneous co-evolution of a rigid robot’s morphology and controller [51], and have been proposed as a promising candidate for robust, decentralized controllers of autonomous drug delivery systems [68].
An NCA is essentially a grid of interconnected cells, each equipped with an identical ANN that is capable of perceiving the numerical states of its immediate cellular neighbors, , and proposing actions, , to regulate its own cell state, following the equation
where we also account for possible imperfections in the cell-specific state updates via a Gaussian noise term of amplitude .
Thus, the cellular agents of an NCA perceive the numerical states of their immediate, respective neighborhoods, , at any given time step, , to update their own states (c.f., fig:introduction:simulation (A, B)). Importantly, we do not include any explicit signature of aging in our model, i.e.,, no cell-internal marker or explicit input to the cellular agents other than their self-regulated cellular states that would contain temporal information.
However, being immersed in a dynamical multi-cellular environment where every part has its own agenda, the cellular agents’ state updates can also be utilized for active communicating with their neighbors, following a policy . From the perspective of Reinforcement Learning [80], an NCA can thus be considered a trainable multi-agent system that needs to utilize local communication rules to achieve a target system-level outcome.
So far, our approach is agnostic to the particular ANN architecture of the update function, . Closely following Ref. [41], we here briefly describe the genuine ANN architecture deployed in our cellular agents, which is schematically depicted in fig:introduction:simulation (B): Inspired by Ref. [81], we partition an NCA’s cell’s ANN into (i) a sensory part, , preprocessing each neighborhood cell state separately into a respective sensor embedding, which we collect in the matrix . These sensor embeddings are then (ii) averaged across all neighbor embeddings into a single context vector . A subsequent (iii) controller ANN, , potentially with recurrent feedback connections, eventually outputs the cell’s action, based on the cell-specific context vector . Notably, for the controller module we here utilize so-termed RGRN architectures [41] that are inspired by a combination of Recurrent ANNs (RNNs) [82] and Gene Regulatory Networks [83] (see Appendix A in Ref. [41] for details).
Analogous to Ref. [41], we model morphogenesis by employing NCAs on a two-dimensional square grid with the objective that all cells of the grid assume a cell-specific predefined target cell type, , after a fixed number of developmental time steps, starting from an initial cell state configuration . We assign the first elements, , of an NCA’s -dimensional cell state as indicators for expressing one of discrete cell types, while the remaining elements of the cell state represent its hidden states, , that can be utilized by the NCA for intercellular communication. A cell’s “type”, , is now defined as the index, , of the maximum element of its indicator vector :
For an NCA to assemble a predefined target pattern of target cell types , we thus need to find a suitable set of NCA parameters that minimizes the deviation of all actual cell types and the desired ones after developmental time steps. Below, we introduce an evolutionary algorithms to evolve suitable sets of NCA parameters that maximize a fitness score based on comparing the “final” cell types of the NCA, , after the developmental stage to the predefined target cell types . In this context, the NCA parameters thus correspond to the virtual organism’s genotype, while the grid of final cell types represent the system-level phenotype that is seen by the selection mechanism of the evolutionary process (c.f., fig:introduction:simulation (C)).
2.2. Neuroevolution of NCAs: An Evolutionary Algorithm Approach to Morphogenesis
Evolutionary Algorithms (EAs) are heuristic optimization techniques designed to maintain and optimize a set, or population, , comprising parameters or individuals, , over successive generations in order to maximize an objective function or fitness score, . Drawing inspiration from the principles of natural selection and biological life’s DNA-based reproduction mechanisms, EAs (i) predominantly select high-fitness individuals for reproduction, and utilize (ii) crossover and (iii) mutation operations to generate new offspring by (ii) merging the genomic material of two high-quality individuals from the current population, , and (iii) occasionally mutating offspring genomes, by adding typically Gaussian noise to the parameters; the ⨁ symbol signifies a genuine merging operation of two genomes, which may vary according to the specific EA implementation. Thus, populations of individuals are directed towards high-fitness regions within the parameter space , typically across numerous generations of successive selection and reproduction cycles (i)-(iii) [41].
Closely following our previous work [41], and in contrast to many “traditional” EA use-cases, we here explicitly distinguish between the genotypic parameters, , and the corresponding phenotypic realizations, . More specifically, we utilize NCAs (c.f., sec:methods:nca) to model the biologically inspired developmental layer [19] in between genotypes and phenotypes, , akin to the process of morphogenesis: Here, a genotype, , corresponds to a particular realization j of an NCA’s parameters, comprising the set of initial cell states and the corresponding ANN parameters , such that
where we explicitly distinguish between structural (S) and functional (F) genes. We then employ eq:nca-update for developmental steps to successively transform the NCA’s cell states from an initial state into a “final” set of cell types on the NCA’s grid. This mature phenotype,
represents a two-dimensional tissue of cells, and is the input to the fitness score based on which the EA selects: As illustrated in fig:introduction:simulation (C), we here optimize for a fitness score in the phenotype- rather than genotype space, , while still performing genetic recombination and mutation in the genotype space.
Let’s recall: our goal is to achieve morphogenesis of a two-dimensional spatial tissue of cell types, , that optimally resembles a predefined target pattern, , of a total of cells on an square grid of an NCA. Moreover, we aim to derive a virtual organism with this capability by modeling a biologically inspired evolutionary process with EAs to evolve a suitable set of NCA parameters (c.f., fig:introduction:simulation (C)). Thus, we introduce our phenotype-based fitness score, , as [41]
where counts the number of correctly assumed cell types after developmental steps (where ), counts the number of developmental steps at which the target pattern is realized entirely (whenever for all i), and counts pairs of successive time steps, , without cell type updates in the entire tissue (where for all i).
Thus, we design an evolutionary process that primarily selects for NCAs that assume the correct cell type pattern after (precisely) developmental steps and reinforces NCAs (by a factor of in the fitness score) that prematurely achieve and maintain the target pattern. To avoid developmental stagnation and increase the EA’s performance, we specifically discount the fitness score of NCAs with static behavior by . A fitness score of indicates that the problem is solved.
2.3. Information-Theoretic Analysis: Active Information Storage, Transfer Entropy and Spatial Entropy on the NCA
Information theory [84] offers valuable insights into the dynamics of complex systems. In order to understand the NCA’s information dynamics during aging, we used three key information-theoretic metrics to examine the information dynamics: active information storage [77], transfer entropy [78], and the spatial entropy.
Active Information Storage (AIS) quantifies the amount of information from an agent’s past that is pertinent to predicting its future state. Specifically, AIS refers to the portion of stored information currently utilized for determining the agent’s next state [77]. Mathematically, the AIS of an agent Q is expressed as the local mutual information between its semi-infinite past as and its next state at time step :
Here, is an approximation with history length k. The time-averaged value, weighted by the distribution of , is denoted as . In our study, we calculated the local AIS across the states of the cells.
Transfer Entropy (TE) measures the information transferred from a source agent to a destination agent that is not contained in the past of the destination agent. We employed the local TE concept introduced by Lizier [77]. The local TE from a source agent Z to a destination agent Q is defined as the local mutual information between the previous state of the source and the next state of the destination agent , conditioned on the semi-infinite past of the destination (as ):
The transfer entropy is the local TE averaged over time, denoted as , where is an approximation with history length k. Unlike mutual information, which measures static correlation, transfer entropy captures the dynamic, directional flow of information within the agent network.
Spatial Entropy (SE) measures the randomness or disorder within a spatial distribution of states. It provides insight into the complexity of spatial patterns in a system, such as an NCA with multiple states. In our study, we compute SE by evaluating the entropy of the distribution of cell states across the grid at each time step. Formally, the spatial entropy H at time step n is defined as:
where S is the set of possible states, and is the probability of state s occurring in the grid at time step n. This measure provides an indication of the level of unpredictability or complexity in the spatial arrangement of cell states. In this article, we calculate the spatial entropy for the states of the NCA across different time steps.
3. System
Following our previous study [41], we utilize EAs to evolve NCAs to perform morphogenesis tasks. Each cell in our NCA implementation is equipped with a gene regulatory network-inspired recurrent ANN, conceptually substituting the intricate internal decision-making competencies of biological cells (see fig:introduction:simulation (B) for a visualization and Appendix A in [41] for numerical details). In other words, each cell in the NCA is equipped with an information processing pipeline that transforms sensory input of its local environment in the NCA’s grid into control instructions to regularize its own cell type. If done in a coordinated way, these local individual cell-state updates guide the formation of a mature system-level phenotype - in our case of a target tissue - over successive developmental steps.
More specifically, we utilize cells on an NCA with fixed boundary conditions (see sec:methods:nca and Appendix C in Ref. [41]) and train it analogously to [41] via evolutionary algorithms (see sec:methods:ea) to perform morphogenesis - i.e.,, self-orchestrated tissue growth - of a smiley-face pattern in a predefined number of developmental steps (see fig:introduction:simulation (C) for an illustration of the evolutionary process).
The target tissue consists of distinct cell types: one cell type for the background cells, one for the face cells, and a single cell type for the facial organs, i.e.,., for the eyes and the mouth (c.f., blue-, magenta-, and white-colored cells in fig:introduction:simulation (A)). Moreover, we allow the NCA to utilize hidden channel of its cell state channels for unconstrained intercellular communication. In cell-state updates, given by eq:nca-update, we utilize a noise level of , limit the numerical value of the proposed action to , and clip the cell-states post-update to (see Appendix C and Ref. [41] for more details on our NCA implementation).
Our NCAs transform an embryonic genotype, , into a mature phenotype, , and we here quantify an NCA’s performance by evaluating the corresponding fitness score, , defined by eq:fitness with respect to the smiley-face pattern over the NCA’s entire life-span of developmental steps. As illustrated in fig:system:morphogenesis, a successfully evolved NCA maximizes the fitness score to , exhibiting correctly expressed cell types after time steps, then maintaining the target pattern until (see sec:methods:ea for details on the evolutionary process).
Notably, the cellular agents do not perceive any information about the current nor the final fitness score of their corresponding NCA. The fitness score is only utilized by the EA to optimize the NCA’s parameters over evolutionary time-scales. Moreover, there is no contribution neither to the fitness score nor to the NCA’s ANN sensory inputs that would provide the system with explicit temporal information indicating the “age” of any part of the virtual organism, or even a notion of “time” in general. Thus, the self-orchestrated pattern formation of the here - and previously [41] - investigated NCAs is fully driven via emergent, i.e.,, evolved, intercellular communication rules and intracellular decision-making.
4. Computational Results
4.1. Aging as a Loss of Goal-Directedness: Organism Learned Development During Evolution, Not to Maintain Anatomical Homeostasis After Development
We aimed to assess whether the self-regulatory behavior of biological systems that drive morphogenesis induces aging-like phenomena at long time-frames. To this end, we use NCAs evolved in silico to model the process of morphogenesis and then deploy their self-regulatory dynamics for much longer times compared to the reproduction stage during evolution (i.e., when selected for reproduction). We monitor and analyze the NCAs morphological integrity throughout their lifetimes and compare such long-term morphological trajectories with biological aging. The evolutionary process described in sec:methods:ea,sec:system explicitly selects for optimal fitness scores after developmental steps, which corresponds to the fertility age of real-world organisms. However, this implies that the behavior of an NCA with a potentially much longer lifetime of , and particularly , is not subject to selection and may fundamentally be considered ill-defined or ambiguous.
To demonstrate this, we deploy the NCA solution that has been evolved to self-assemble a smiley-face pattern during a morphogenesis phase of developmental steps (c.f., fig:system:morphogenesis) for a much longer lifetime of up to time steps, notably without any further optimization considering the extended lifetime. From the results presented in fig:result:pleiotropy, we learn that after an initial rapid rise in the fitness score during the developmental phase, the NCA’s long-term fitness score gradually declines, primarily due to the loss of different internal organs such as a single or both eyes and, occasionally, the mouth. Moreover, this is not only a feature of the particular NCA demonstrated here, but is a systematic phenomenon of the presented framework.
Thus, we argue that the biologically-ubiquitous self-regulatory behavior of a substrate’s agential parts may induce system-level dynamics that resemble the process of biological aging if applied on timescales that exceed development. The organisms learned during evolution with development as the primary goal; once they reached that goal, we observed that they don’t regenerate by themselves and slowly show signs of morphological deterioration, e.g., aging. In this sense, aging can be seen primarily as a loss of goal-directedness, the goal of development being different from the goal of maintenance of the anatomy over timescales exceeding development. This view on aging is related to programmatic theories of aging like antagonistic pleiotropy or hyperfunction theory [10,14,15] that state that what is good for development could ultimately cause aging. In our framework, the lack of goal-directedness after development may cause some informational antagonistic pleiotropy (or hyperfunction) as cells lose their developmental policy after reaching the target morphology but it is broader in scope as giving a new goal to the cellular collective like regeneration and therefore by reactivating developmental pathways may cure aging (see regenerative experiments below).
4.2. Impact of Defects of Cellular Information Processing at Different Levels on the Rate of Aging in a Multi-Scale Competency Architecture
To determine how the perception-action cycles of the unicellular agents of a collective multi-cellular organism influence, at least in part, the rate of morphological decline, we here investigate the effects of an NCA’s system parameters on its long-term morphological integrity. Based on the NCA system defined in section III, we identify four key mechanisms involved in the corresponding uni-cellular perception-action cycles affecting morphological integrity, namely cell-specific differentiation reliablility (a), reliable state updates (b), reliable communication between neighbors (c), and persistent genetic encoding (d). Our framework allows us to selectively control these corresponding processes (a-d) and explicitly corrupt them independently over the lifetime of the virtual organism. Thereby we demonstrate the severe effects of these processes on the rate of morphological aging.
In the following sections, we therefore study the factors accelerating aging, not the cause of aging per se that we see primarily as a loss of goal-directedness. We introduce these known aging mechanisms (i-iv), respectively, based on and deployed to the biologically motivated unicellular competencies in our NCA system. We then demonstrate and discuss the corresponding implications on enhanced rates of morphological aging in long-term simulations of such virtual organisms. We illustrate the respective results in a compact form in fig:results:schedules(A-D), and continue below with an information-theoretic analysis of these processes.
4.2.1. Cellular Differentiation
One critical parameter in our system is the noise level utilized to corrupt the NCA’s update step given by eq:nca-update. While a moderate noise level can help an evolutionary process to identify robust NCA parameters that solve a specific morphogenesis task, chosen too low, the evolutionary process will in our case most likely identify a direct encoding of the target pattern, largely neglecting cellular competencies in the pattern formation task [41]. In turn, a too-large noise level will not only prevent the evolutionary process from finding a good solution to the morphogenesis task but, in general, drastically limit the cell specification capacities of the NCA’s cell states. Therefore, noisy cell state updates in our system could be seen as the accumulation of misdifferentiated cells, ultimately corrupting the state perceptions of neighboring cells on the NCA creating a default of cellular communication (a hallmark of aging) similarly to [6,85]. This mechanism accelerates aging but is not a cause per se.
We argue that an increasing noise level over the lifetime of such an NCA-akin organism successively affects its morphological integrity. To demonstrate this, we thus utilize the pre-evolved NCA from above (c.f., fig:system:morphogenesis,fig:result:pleiotropy) and deploy it for an extended lifetime of timesteps while successively increasing the noise level linearly from to . The results depicted in fig:results:schedules (A) show that while the correspondingly affected virtual organisms initially follow their unaffected cousins (c.f., fig:system:morphogenesis,fig:result:pleiotropy and black-dashed line in fig:results:schedules (A)), after a particular duration (specific to the particular NCA solution and noise schedule ) the fitness score rapidly drops at , emphasizing an enhanced rate of degradation of the target tissue compared to the baseline with constant noise .
Noise influences the dynamical cell states in our NCA experiments. These states are the inputs and outputs of the virtual tissues’ ANN augmented cells, and represent dynamic variables of the self-regulatory NCA dynamics, with attractor states or instabilities. We thus interpret the effect discussed here as cellular misdifferentation-enhanced morphological aging. Understanding such “run-away” arguments in the “biological software” of cell state expressions, or instabilities in the context of competent tissues seems central to understanding aging.
4.2.2. Cellular Competency
Next, we directly manipulate the cellular decision-making competency - the reliability with which uni-cellular agents can regulate their own cell states within a tissue (e.g., regulative development and regenerative tissue allostasis). Here, we investigate whether degrading unicellular competencies throughout the lifetime of an organism enhances morphological aging: while young, healthy cells are highly reliable in their (re)actions to external perturbations, especially during development, the reliability of such cell decisions might be strongly affected over the lifetime of an organism, inducing morphological decline. To this end, and following 6, we define the decision-making as a probability at which proposed actions of unicellular agents are actually forwarded to the environment at a given time step. More specifically, at every instance in time we draw a uniform random number for every cell in the tissue. Only if we consider the cell’s action in the NCA’s update step, described by eq:nca-update, and omit it otherwise by setting . In that way, we can scale the average reliability of decision-making of all cells in the tissue, i.e.,, while corresponds to fully reliable decision-making of every cell at every time step, parameterizes fully passive cells which can’t make decisions at all.
Here, we utilize as decreasing cellular competency by transforming to in time steps following the functional form (see fig:results:schedules (B)). This increasingly corrupts cellular competencies by successively decreasing the reliability at which every unicellular agent in the grid can regularize, and thus “correct” their states against noise or faulty cell state configurations.
The results of fig:results:schedules (B) reveal that the fitness score of NCAs whose cellular competencies are gradually affected remains similar to the baseline fitness scores of the unperturbed case (c.f., fig:system:morphogenesis,fig:result:pleiotropy). But eventually, at , the small but finite noise of used in our simulations leads to accumulated damage in the NCA’s states that is catastrophic for its morphological integrity, thus destroying the target morphology via diffusive processes.
Thus, decreasing cellular decision-making competencies in a noisy collective organism necessarily lead to morphological aging effects, caused by unreliability in the “biological cell-state reconfiguration software”. However, the rate at which these decreasing cellular competencies lead to significant aging effects is an intricate interplay between the actual noise in the system and the robustness of the unicellular agents to persist against accumulating noise in their respective state configurations.
4.2.3. Cell-Cell Communication
Another necessary feature of systems with decentralized collective decision-making is reliable communication between agents. Similar to biological tissue, the cells of an NCA are integrated into a lattice of functionally homogeneous cells that only diverge in their respective cell state history. However, their neighbor coordination typically remains fixed [39,41,67]. Here, we specifically manipulate the connectivity between the unicellular agents in the NCA’s grid by blocking increasingly many communication channels between neighboring cells as time proceeds. This approach is biologically motivated by cells closing their gap junctions (GJs) to their neighbors, which is known in vivo to shift cells from cooperatively working on large morphogenetic setpoints (organ maintenance) toward individual goals appropriate to unicellular organisms such as proliferation and migration [50,86,87,88]. It is also related to altered cell-cell communication found during aging [6]. Indeed, aging involves deficiencies in neural, neuroendocrine, and hormonal signaling pathways [89] as well as the alterations in the bidirectional communication between human genome and microbiome. Chronic inflammation is also part of this altered cellular communication [6].
We achieve this technically by omitting the corresponding input state of a blocked connection, or GJ from cell j to cell i (see fig:introduction:simulation (B)). More specifically, we utilize a probability which defines the fraction of GJs that should remain open for each cell in the tissue at a given time . In turn, if for a particular cell i, the fraction of connections to its neighbors exceeds at time , we henceforth randomly block another one of its still connected GJs (once closed, GJs remain closed in our case); see sec:methods:nca and [41] for more details on our definition of cellular neighborhoods.
By linearly transforming to , we successively isolate the unicellular agents from their host tissue by blocking an increasing number of their GJs until, eventually at time , all cells are effectively isolated from their surroundings. In fig:results:schedules (C) we demonstrate the effects of this approach: Again, the self-orchestrated morphogenetic program can withstand the effects of increasingly corrupted competencies for a surprisingly long time. However, at , corresponding to and thus half the GJs of every cell being closed, the effects of the removed communication channels can be observed by a resulting rapid drop in the phenotypical fitness scores caused by a rapid degradation of the target pattern.
Thus, decreasing the ability of cells to communicate with their neighbors leads to enhanced effects of morphological aging if a threshold of GJs is closed.
4.2.4. Accumulation of Genetic Damage
Another potential source for self-induced aging is accumulating damage to the cells’ genetic material via successive mutations of the functional genome over the lifetime of the virtual organism: To this end, we utilize the pre-evolved NCA solution discussed above (c.f., fig:system:morphogenesis,fig:result:pleiotropy), but at every time step , we successively manipulate the cells’ ANN parameters throughout their lifetime. This directly introduces accumulating genetic mutations to the self-regulatory mechanisms of the NCA, causes potential dysfunctional decision-making behavior at the uni-cellular level, and correspondingly leads to enhanced morphological decline. More specifically, we add zero-centered normal distributed noise of standard deviation to the functional genes of the NCA, , where . The mutated functional genes are then mapped to the ANN parameters of the unicellular agents’ controllers at every time step , thus directly affecting the intracellular decision-making machinery of the NCA.
From the results depicted in fig:results:schedules (D) we can learn several things: First, the evolved NCA solutions seem incredibly brittle against accumulating genetic damage in the ANN parameters, which is reflected by the fact that most of the deployed simulations (with statistically independent mutations) retain high-fitness target patterns for the entire simulated lifetime of the NCA of timesteps. Second, and in contrast to sec:results:induced-degeneracy:noise,sec:results:induced-degeneracy:competency,sec:results:induced-degeneracy:cell-communication this leads to large qualitative deviations in individual behavior of an ensemble of statistical independently mutated NCA solutions: some solutions die off early and obtain vanishing fitness scores, others degrade rather continuously with a spread-out fitness score of , and many solutions still maintain their integrity although accumulating significant genetic damage of in total.
Thus third, while many mutations appear neutral in our framework, critical mutations to specific genes might induce detrimental modifications to the behavior of the unicellular agents (if they occur at the “correct” moment), typically causing a rapid degradation of the target pattern of the affected NCA as reflected by significant drops in the fitness scores of the corresponding virtual tissue. Notably, due to technical reasons, we mutate the functional genome of every agent homogeneously, thus retaining a functionally homogeneous distributed decision-making in the NCA, which nevertheless deviates functionally from the baseline at . Potentially intriguing effects based on heterogeneous mutations, and thus heterogeneous agents in the NCA are out of the scope of this work, and we leave a more thorough investigation to a further contribution.
4.3. The Acceleration of Aging Is Linked to Increase in AIS and TE, While Spatial Entropy Revealed Two Different Kind of Aging: Loss of Structure and Proliferation, and Accumulation of Morphological Noise
In order to understand the information dynamics of aging and the cellular information processing in a competent tissue, we applied information-theoretic measures such as (i) Active Information Storage (AIS), and Transfer Entropy (TE) and the Spatial Entropy (SE), e.g., the entropy applied to the states of the organism. (see Figure 6).
We observed in the default behavior case, without any add of the noise or induced loss of competency that both the mean AIS and TE are decreasing during development and aging. For some organisms, we can see that TE can increase during aging, indicating episodes of regeneration in the sense that the morphology is impaired and the organisms try to recover. As we can observe it, the spatial entropy is slowly decreasing during aging, and the anatomy is slowly impaired, with the loss of organs like eyes (see Figure 6).
Interestingly, we observe that for the cases where we increase noise, see sec:results:induced-degeneracy:noise, there is a threshold value for increasing the noise level after which a rapid decline in fitness occurs - which we interpret as rapid aging-/dying. Indeed, at the same moment, the SE is decreasing, showing a complete disappearance of the anatomy towards a quasi-random distribution of the states. In this case, the AIS and TE increase around 600 steps, a bit more of half-life. A high TE can be interpreted as a high entropy of actions, the cells trying to act after the decline and disappearance of the anatomy.
However, the loss of competency of the cells does not impact the AIS and TE immediately as in the noise case. It is delayed and is starting to have a lot of random states in the anatomy with an increase of the AIS and TE at the end of the lifespan of the organism . But when noise increases and loss of competency are associated the increase in AIS and TE during aging is higher than in the case when only noise is added. Maybe, the loss of competency alters the capacity of the cells to understand the decline of the anatomy and their potentials to counteract it. Noise in the other hand clearly creates a lot of cellular actions during lifespan.
When the connectivity of the cells is altered, from 500 steps, we can observe an increase in AIS and more variability on TE (decrease and increase during aging). By closing the gap junctions, the cells are not receiving information from their neighbors gradually, increasing the uncertainty about their actions. We can see with the SE that the cells can counteract the loss of connectivity until 500 steps until an important decrease in spatial entropy and a complete inversion of the states of the anatomy.
The case that is creating the highest variability in anatomical outcome is the one where we altered the ANN parameters. AIS decreases and then increases over time slightly, TE is close to 0 after development and SE shows different types of aging, a loss of structure (organs) and proliferation of a specific states in the tissue (cancer), and a random activation of cell states.
We can conclude that if the states of the organisms do not change over time, if the anatomy reached a steady state, the AIS and TE are low. AIS is low because there is no new information to be gained from observing past states—future states are always the same and known, independent of the past. Essentially, the predictability is absolute, but since the system does not utilize past information to determine this (the state is intrinsically static), the AIS has low values. This indicates that in a non-changing system there is no information flow necessary between past and future states. The future state is always known (it’s the same as the current state), and thus, the storage of information from past states is irrelevant for future predictions. However, when AIS increases, it is associated to an increase in TE too. It happens in the cases of catastrophic changes in the morphology, as related to changes in spatial entropy. This important change implies an important information flow between cells, possibly representing the cellular attempts to recover from the high morphological changes. The absence of information flow in the default behavior can indicate that the systems considered that it reached its goal, e.g., development and morphostasis is not one of its goal, leading to a steady decrease of the anatomy, or in other words, aging. SE measures the entropy of the grid states. When SE is very high it corresponds to a random distribution of the states over the tissue. This random distribution can be found mainly in the cases of the loss of competency, noise and when both perturbations of associated. SE reveals also a different kind of aging corresponding to a loss of structure, with the loss of specific organs or the proliferation of a specific states in the tissue, like in the loss of connectivity and ANN parameters cases, meaning that the different perturbations lead to different kind of morphological degradation.
4.4. Regeneration as the Cure for Aging?
4.4.1. Loss of Organs Does Not Imply the Loss of Information About the Organ
In biological systems, certain cells or tissues retain “memories” of past states, influencing future behaviors. For instance, epigenetic memory in cells allows them to “remember” previous environmental exposures or developmental cues. Even if the external trigger is removed, cells can maintain certain gene expression patterns or epigenetic marks [90]. Similarly, tissues may retain information about past structural organization. For example, some amphibians can regenerate limbs or eyes after they are entirely removed [91,92,93]. Planarian flatworms exhibit a kind of holographic memory where, upon cutting into many pieces, each fragment of the animal regenerates everything that is missing to create a perfect and complete copy [94,95]. This persistence of information about the system’s target morphology, which serves as the setpoint for the process of anatomical homeostasis, can be implemented in a variety of biochemical, bioelectrical, and biomechanical information, thus it is important to study this important question.
In order to analyze the information dynamics during loss of organs, we computed the spatial AIS and TE (see Figure 7). We observed that the loss of the eyes during aging does not imply the loss of the AIS at the location of the eye as we can see at step 800 of Figure 7. The AIS is high and located at the position of the eyes in 100% of the cases. This suggests a memory retention in the system. For example, high AIS values at the “eye” locations in our system parallels the prepattern of the developing amphibian face consisting of regions of specialized cellular resting potential that determine the organs [96]. Even after visible features are lost, underlying patterns (like bioelectric signals or AIS) can persist, reflecting the system’s inherent memory of structure. AIS measures how much information about the past states is retained in a system over time. This retention can create a lingering effect, where regions that previously had distinct structures (like the eyes) continue to show high AIS values. Even if the explicit structure is no longer present (e.g., the “eyes” disappear in the smiley face), the underlying system dynamics may still carry information related to those past configurations. This residual AIS indicates that the system “remembers” these regions as important parts of its past structure, maintaining high information storage due to the prior role they played.
We conclude that the loss of organs does not imply the loss of information about the organ (see example Figure 7). Following this result we decided to explore the implications for rejuvenation by inducing the regeneration of the organs in the system in the next section.
4.4.2. Implication for Rejuvenation: A Simulated Experiment of Organ Restoration
The main reason for the long-term fitness degradation in our experiments - as discussed in sec:results:antagonist-pleiotropy, i.e.,, even without artificially affected cellular competencies - is the loss of specific internal organs, such as the eyes or the mouth. Thus, we here aim to correspondingly counteract the loss of detrimentally affected organs by replacing either the entire organ or specifically reconfiguring the affected cells of the NCA with “regenerative” state information (also erasing their internal working memory by resetting their RNN state information, see appendices A and B for details).
To achieve this, we define an organ-specific score function, , that quantifies for every time-step the fraction of correctly expressed target cells out of the total number of target cells of a specific organ, (c.f., blue, green, and red cells in the snapshot of the smiley-face pattern NCA in fig:results:restoration:organ, respectively). Whenever an organ’s score falls below a predefined threshold value of , we perform an intervention to the affected organ, aiming at restoring its morphology locally. To interfere as little as possible with the NCA’s inherent developmental program, we propose utilizing the initial cell states as the regenerative state information of affected cells, (c.f., sec:methods:nca). This corresponds to reconfiguring the affected cells to their initial (primordial) state at the onset of the developmental program of the NCA .
4.4.3. Less Is More: Organ Restoration Induction by Injecting the Regenerative Information Only to Incorrectly-Positioned Cells
We thus deploy the NCA solution discussed in sec:results:antagonist-pleiotropy (c.f., fig:result:pleiotropy) for an extended life-span of time steps and apply the here discussed rejuvenation interventions whenever an organ’s score falls below a threshold value of . In fig:results:restoration:organ (A) and (B) we respectively present the results of this procedure by either (A) resetting all cells of an affected organ with their initial configuration or (B) only resetting the particular cells of an affected organ that express different cell states than their target types. We present for both (A) and (B) the long-term fitness scores of 10 statistically independent simulations, contrast their mean behavior with the baseline case without interventions (see fig:result:pleiotropy), and visualize the organ-specific scores and the corresponding intervention events for selected individuals.
We see that method (A) is not very sustainable: Although we generally measure significant short-term improvements in a respective organs’ scores after such intervention events, the intervened organs dissolve again quickly so that after time steps (which we set as the lower limit of two consecutive interventions, allowing for unperturbed development during this period) the organ needs to be reset again, and again. This periodic need for interventions - also reflected in correspondingly oscillating fitness scores of the entire organism - seems fundamental in our system and is not significantly affected by the delay between two interventions. The resulting fitness scores of the intervened individuals are only slightly improved compared to the baseline case.
Strikingly, if only the wrongly expressed cells of an affected organ are reconfigured to their initial state, as in case (B), very few of such targeted interventions (on the order of , c.f., bar plots in fig:results:restoration:organ (B)) suffice to sustainably restore degrading organs and allow the corresponding virtual organism to maintain its morphological integrity over an extended lifespan. The corresponding long-term fitness scores of the rejuvenated individuals show significant improvement compared to the baseline case and can even be stabilized into a steady state of a constant value of, here, ; notably, the difference to the maximum fitness score of right after the developmental stage at arises due to the accumulative stagnation loss term in eq:fitness.
4.4.4. Boundaries Matter: Organ Restoration Is More Efficient with the Injection of a Differential Pattern Including the Organ Cell States and Neighboring Cell States
Although the above approach is already promising, we further investigate how including the cells at the boundary to adjacent tissue surrounding the respective organs affects regeneration in our procedure. Thus, we additionally consider the respective boundary cells for the left- and right eyes in the evaluation of the respective organs’ scores but intentionally omit the boundaries of the mouth (c.f., emphasized color-coded cells in fig:results:restoration:socket, similar to fig:results:restoration:organ); notably, the mouth is much more stable than the eyes, and its cells already occupy a substantial fraction of the face.
Deploying the same experiments as above in sub:results:restoration:organ, we see in fig:results:restoration:socket that (A) interventions reconfiguring all the cells of affected organs (now also including their respective surrounding tissue) show similar performance to the corresponding case shown in fig:results:restoration:socket (A). However, the more targeted cell-specific interventions depicted in fig:results:restoration:socket (B) of now manipulating not only the wrongly expressed cells of an affected organ but also the wrongly expressed cells of their respective boundaries greatly improve the performance of restoration interventions compared to the cases fig:results:restoration:organ,fig:results:restoration:socket (A) and fig:results:restoration:socket (B). Here, an average number of as little as interventions (and in some cases even one single or two) suffice to stabilize the morphology of the here investigated smiley-face pattern organism in long lifespan experiments.
We conclude that boundaries (and more generally relations) between organs matter in our regeneration experiments: including surrounding tissue in detecting affected organs and in subsequent regeneration experiments that specifically target the wrongly expressed cells of the affected organs can be an efficient approach for regeneration and rejuvenation applications in our system.
A different but likewise promising approach is replacing the affected organs or cells directly with their target states (rather than their respective initial states as here). However, while “transplanting” entire organs can be effective, there is little quantitative difference in rejuvenation performance between replacing cells with their target states and reconfiguring cells into their initial states. From a biological perspective, the latter approach of resetting cells to their primordial states strikes us as conceptually and technologically more feasible, as the number of specialized target states might greatly exceed the number of necessary primordial initial states: resetting cells to their initial cell state, or at least to a particular primordial cell state suitable for starting a specific morphogenetic task, could serve as a general fallback mechanism to induce regeneration of the respective (here virtual) organism by restarting the developmental program locally.
At this point, we would like to stress that the developmental program of our NCA has neither been optimized for such long lifespans nor for the here proposed intervention experiments. The latter can thus be seen as minimally invasive ad-hoc cellular reconfiguration interventions, i.e., a dynamic and targeted “hacking” of the organism’s developmental plan by an external agent [19] (i.e.,, us, in this case).
Overall, we conclude that such a procedure of targeted cellular interventions is a reliable approach for rejuvenation in our system.
5. Discussion and Conclusion
In this paper, we aim to address a significant gap in the aging literature which so-far lacks - to the best of our knowledge – computational models of aging considering cellular information processing and multi-scale competency. We seek to address this knowledge gap by developing an evolutionary framework integrating the multi-scale competency architecture of multicellular tissue.
Specifically, we here utilize numerical tools from Artificial Life [61], so-called Neural Celular Automata (NCAs) [39,41,67] which we trained with an evolutionary algorithm to perform self-orchestrated targeted pattern formation, i.e.,, morphogenesis of a smiley-face pattern. The localized unicellular decision-making centers in our NCA implementation are modeled by Artificial Neural Networks (ANNs) which perceive the numerical states of their direct cellular neighbors on the grid of the NCA to regulate their own cell state individually. In that way, NCAs substitute homeostatic and morphogenetic pathways of “real” multicellular biological tissue, the NCA-specific ANNs and numerical states respectively corresponding to cell-internal decision-making machinery (such as gene regulatory networks) and cell-type expressions.
Analogously to their biological counterparts, the NCA’s cells form their decisions following a global agenda, allowing the collective of cells to assume a predefined target pattern over a predefined developmental time, thus simulating the process of embryogenesis. The evolutionary algorithm selects mature phenotypes directly after the predefined developmental stage (i.e.,, fully grown tissue), specifically quantifying how well the cells of the corresponding NCAs can grow the facial features and organs of the smiley-face pattern in a fully self-orchestrated and decentralized way. Notably, the NCA’s cells do not in any way explicitly perceive time or their age (in terms of simulation time steps), but their dynamics is fully governed by regulating and exchanging noisy state information on the NCA’s grid. Yet, when deploying a successfully trained NCA for much longer times than necessary for development (at which stage evolutionary selection would have occurred), the tissue of the NCA expresses signs of aging, such as successively blurring boundaries between- or entirely losing a single or several organs. We emphasize that this behavior is emergent and happens without any further intervention.
We propose, that even in the absence of accumulated cellular or genetic damage, a biological system left without any goal in the morphospace will start degrading anatomically. Using an in silico model of homeostatic morphogenesis with a multi-scale competency architecture and information dynamics analysis, we find the following, which provides a novel perspective on aging dynamics with significant implications for longevity research and regenerative medicine: (1) Absence of Long-Term Morphostasis goal: Aging emerges naturally after development due to the lack of an evolved regenerative goal, rather than being caused by specific detrimental properties of developmental programs (e.g., antagonistic pleiotropy or hyperfunction); (2) Acceleration Factors vs. Root Cause: Cellular misdifferentiation, reduced competency, communication failures, and genetic damage all accelerate aging but are not its primary cause; (3) Information Dynamics in Aging: Aging correlates with increased active information storage and transfer entropy, while spatial entropy measures distinguish two dynamics, (i) the loss of structure and (ii) morphological noise accumulation; (4) Dormant Regenerative Potential: Despite organ loss, spatial information persists in our cybernetic tissue, indicating a memory of lost structures, which can be reactivated for organ restoration through targeted regenerative information; and (5) Optimized Regeneration Strategies: Restoration is most efficient when regenerative information includes differential patterns of affected cells and their neighboring tissue, highlighting strategies and protocols for rejuvenation.
While the cells in our system learn to perform morphogenesis, i.e, how to grow an adult morphology successfully, we still observe aging effects in long-term simulations even without artificially perturbing the NCA’s parameters. These results suggest that development and morphostasis, while close in terms of morphological trajectories and tasks still significantly diverge. To counteract aging, it is therefore necessary to (re)activate a developmental/regenerative goal. We explicitly demonstrated this by providing our cybernetic tissue with the necessary information for regeneration, thereby inducing rejuvenation through self-organization at the anatomical level. Our results suggest a roadmap for rejuvenation therapies, particularly that curing aging may imply to provide a new goal to the tissue, i.e., regeneration. In other words, this suggests that solving regeneration solves aging. As currently debated, regenerative species, such as salamanders or planaria, might not age at all. While, already known for planaria, this is the subject of current research for salamander. The latter not only lack senescent phenotypes but it is indeed impossible to define an epigenetic clock for specimen older than four years [97]. One extension to the present work thus will be comparing a system that has learned to regenerate and actively maintain morphostasis after development to an aging system that has only been trained on morphogenesis tasks. Understanding their differences in terms of information dynamics will allow us to form a thorough understanding of the homeostatic loops that allow differently expressed cells in a self-assembling tissue to maintain their integrity against adversarial attacks or thermodynamic noise.
This work also emphasized how to optimize regenerative strategies. Our results show the importance of the information at the level of the borders of integrated tissue or organs for regeneration. This importance can be found in the field of bioelectricity. Indeed, research indicates that mutations in critical neurogenesis genes, such as Notch, can lead to developmental defects [98]. These organ defects can be cured through the overexpression or activation of native HCN2 (hyperpolarization-activated cyclic nucleotide-gated) channels [99,100,101]. As voltage-sensitive ion channels, HCN2 channels respond to their cellular environment by enhancing hyperpolarization in cells that are slightly polarized, while remaining inactive in depolarized cells. This mechanism functions analogously to “contrast enhancers,” which sharpen weak differences in membrane potential () across compartment boundaries, much like a sharpening filter brings clarity to a blurred image. In other words, HCN2 enhances and sharpens the borders in the bioelectrical pattern.
Moreover, our results suggest that a multicellular collective is capable of maintaining the memory about fully degraded features of an organism, as explicitly demonstrated here via organ regeneration experiment and information dynamics analysis. This suggests that this information is dormant and can be “retrieved” by injecting the appropriate signals that trigger regeneration in our system. This finding opens up exciting possibilities for future research, particularly in the realm of regenerative medicine and bioengineering. By understanding how cells store and retrieve hierarchical anatomical information, we could potentially develop new methods to promote regeneration in damaged tissues or organs. Additionally, exploring the mechanisms by which cells communicate and coordinate to maintain this memory could lead to breakthroughs in preventing age-related degeneration. For example, targeting specific bioelectrical patterns or enhancing the function of key ion channels like HCN2 could help maintain tissue integrity and function over time. Furthermore, the here discussed principles targeting the “software” of life could be applied to synthetic biology and rejuvenation, where designing artificial systems with similar regenerative capabilities [102,103] as well as new aging treatments might become feasible [26,34].
While our current framework is capable of self-assembling target patterns of heterogeneously distributed cell types, the ANN-based decision-making centers of our NCA’s cells are implemented homogeneously: all cells of an NCA typically share the same ANN structure and parameters, and thus the same basic functionality. Yet, the cells might be configured differently and thus effectively behave heterogeneously due to cell-specific state expressions and input relations, in short, due to their morphogenetic history. In that way, these artificial cells’ functionality mirrors gene regulatory networks. Thus, further in silico studies could investigate minimally complex high-level interventions necessary to induce robust regenerative effects in affected tissue simply by reconfiguring the cellular collective locally. Furthermore, allowing cell-specific functional mutations during the lifetime of such virtual organisms might be an intriguing way to study morphogenetic and aging-related implications of heterogeneity.
Our results validate aspects of both damage-based and programmatic theories of aging [1,4,5] but in very different ways. On the one hand, accelerated morphological decline (or an increased rate of aging) in our system mainly originates from: increasing cellular misdiffentiation, decreasing cellular competency, defective cell-cell communication, and accumulation of genetic damage. While these four characteristics align with damage-based theories of aging [6], they are only sufficient not necessary, i.e., not really causal, as our system undergoes aging also in their absence. Therefore, our model tends towards programmatic theories of aging, antagonistic pleiotropy or hyperfunction [14,15], or the theories of the software design flaw [5,26], in which the causes of aging have to be found in development. Similarly, but not analogously, we here relate aging to the absence of a developmental/regenerative or morphostatic goal, which nonetheless may imply an abnormal continuation of specific developmental pathways that are not beneficial for anatomical homeostasis. However, our results suggest that developmental programs (that are learned during evolution) can also qualify as regenerative pathways when “reactivated” in aging individuals, thus representing a potential protocol for a cure for aging ref. In a sense, regeneration is learned for free and is just dormant after development. This is an important result and indicates that we should focus our research efforts for curing aging on the reactivation of the regenerative capabilities. And our results suggest an experimental research program for regenerative medicine where we create an embryo-like environment allowing the tissue to regenerate. In turn, when interpreting the NCAs’ numerical states as bioelectric degrees of freedom, instead of transcriptomic cell-state expressions, a promising approach toward innovative no-aging or rejuvenation treatments might be to focus on high-level control layers such as bioelectricity that encode collective anatomical goals [26,102].
Our study showed fundamental computational principles in multicellular aging systems, bridging developmental biology with aging research. We explicitly demonstrate that developmental pathways, initially used for morphogenesis, may be dormant and can be reactivated for regeneration and tissue maintenance later in life. Moreover, we observed that cells collectively retain and manipulate morphological information, indicating that targeted interventions could potentially rejuvenate or maintain tissue integrity as organisms age.
Overall, we focused on the loss of goal-directedness in morphospace as the cause of the degradation of the system in this article. However, it may be possible that this cause extends towards nother levels in biological systems but also to artificial ones. For example, it may be possible that to counteract psychological decline during aging by giving new life goals to organism, which is in line with resistance to cognitive decline that has been found in elderly having high social engagement and new activities [104,105,106]. This can take place by providing new patterns (as occurs in metamorphosis [107,108]) within a body, or via communication of signals between bodies, as recently observed in cross-embryo morphogenetic error correction effects [109]. In terms of learning theory, we could ask: what’s the objective of a goal-directed system once it reached its goal? Our results may point towards the idea that a goal-directed system that achieved its goal becomes an aging system in a broad sense: a system deviating from its goal slowly. For a morphogenetic system, this might be reflected by a slowly degrading anatomy, a slight shift in morphospace [21] ultimately leading to death: an anatomical configuration incompatible with life. For AI systems, a decline in performance, similarly to what we found with ChatGPT or other LLMs [110], could be interpreted as aging or developmental defects after the target morphology has been reached [39]. Overall, our results suggest that reaching a goal by generative means is completely different from maintaining it. Thus, to engineer robust and autonomous AI and intelligent robotic systems or synthetic morphology we should integrate the learning of these two different tasks.
Exploring these principles further could lead to significant advancements in delaying or reversing age-related decline. Integrating computational models with experimental biology might pave the way to uncover new strategies to enhance regeneration through manipulating cellular behavior, potentially extending the healthy lifespan of tissues.
Future research will focus on applying these computational insights to real-world biological systems, aiming to develop therapies that modulate aging processes and improve recovery from injuries, thus optimizing the health span of multicellular organisms. These findings not only deepen our understanding of the aging process but also highlight the potential of using computational approaches to predict and control the dynamics of cellular systems in aging. The insights gained from this study open new avenues for research in aging and regenerative medicine, paving the way for novel interventions that could one day transform our approach to aging and chronic disease treatment.
Data Availability Statement
Computational protocols and numerical data that support the findings of this study are shown in this article, in the appendix.
Acknowledgments
We thank the members of the Levin lab for helpful discussions. and Axel de Baat for helpful comments on an earlier version of the paper. BH acknowledges an APART-MINT stipend by the Austrian Academy of Sciences. M.L. gratefully acknowledges We gratefully acknowledge the support of Grant 62212 from the John Templeton Foundation (the opinions expressed in this publication are those of the author(s) and do not necessarily reflect the views of the John Templeton Foundation) and of Astonishing Labs via a sponsored research agreement. We thank Julia Poirier for assistance with the manuscript.
Conflicts of Interest
The authors have no conflicts to disclose. The Levin lab has a sponsored research agreement with Astonishing Labs, a company seeking to impact the longevity and biomedicine of aging field.
References
- de Magalhães, J.P. The biology of ageing: A primer. In Proceedings of the An Introduction to Gerontology; Stuart-Hamilton, I., Ed. Cambridge University Press, 2011, p. 21–47. [CrossRef]
- Partridge, L.; Deelen, J.; Slagboom, P.E. Facing up to the global challenges of ageing. Nature 2018, 561, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L. Biological aging is no longer an unsolved problem. Annals of the New York academy of Sciences 2007, 1100, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, T.B.; Melov, S. On the programmed/non-programmed nature of ageing within the life history. Current Biology 2011, 21, R701–R707. [Google Scholar] [CrossRef] [PubMed]
- de Magalhães, J.P. Ageing as a software design flaw. Genome Biology 2023, 24, 1–20. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023. [Google Scholar] [CrossRef]
- Gladyshev, V.N.; Kritchevsky, S.B.; Clarke, S.G.; Cuervo, A.M.; Fiehn, O.; de Magalhães, J.P.; Mau, T.; Maes, M.; Moritz, R.L.; Niedernhofer, L.J.; et al. Molecular damage in aging. Nature Aging 2021, 1, 1096–1106. [Google Scholar] [CrossRef]
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E.; et al. Geroscience: linking aging to chronic disease. Cell 2014, 159, 709–713. [Google Scholar] [CrossRef]
- de Magalhães, J.P.; Church, G.M. Genomes optimize reproduction: aging as a consequence of the developmental program. Physiology 2005, 20, 252–259. [Google Scholar] [CrossRef]
- Gems, D. The hyperfunction theory: an emerging paradigm for the biology of aging. Ageing research reviews 2022, p. 101557.
- Skulachev, M.; Skulachev, V. New data on programmed aging—slow phenoptosis. Biochemistry (Moscow) 2014, 79, 977–993. [Google Scholar] [CrossRef]
- Raffaelli, Q.; Mills, C.; Christoff, K. The knowns and unknowns of boredom: A review of the literature. Experimental brain research 2018, 236, 2451–2462. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Answering the ultimate question “what is the proximal cause of aging?”. Aging (Albany NY) 2012, 4, 861. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V. The hyperfunction theory of aging: three common misconceptions. Oncoscience 2021, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Austad, S.N.; Hoffman, J.M. Is antagonistic pleiotropy ubiquitous in aging biology? Evolution, medicine, and public health 2018, 2018, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.K. The need for a concept of shape homeostasis. Biosystems 2018, 173, 65–72. [Google Scholar] [CrossRef]
- Pezzulo, G.; Levin, M. Top-down models in biology: explanation and control of complex living systems above the molecular level. Journal of The Royal Society Interface 2016, 13, 20160555. [Google Scholar] [CrossRef]
- Lobo, D.; Solano, M.; Bubenik, G.A.; Levin, M. A linear-encoding model explains the variability of the target morphology in regeneration. Journal of The Royal Society Interface 2014, 11, 20130918. [Google Scholar] [CrossRef]
- Levin, M. Darwin’s agential materials: evolutionary implications of multiscale competency in developmental biology. Cellular and Molecular Life Sciences 2023, 80, 142. [Google Scholar] [CrossRef]
- Mitchell, K.J.; Cheney, N. The genomic code: The genome instantiates a generative model of the organism. arXiv preprint arXiv:2407.15908 2024.
- Fields, C.; Levin, M. Competency in navigating arbitrary spaces as an invariant for analyzing cognition in diverse embodiments. Entropy 2022, 24, 819. [Google Scholar] [CrossRef]
- Levin, M. Collective Intelligence of Morphogenesis as a Teleonomic Process. In Evolution "On Purpose": Teleonomy in Living Systems; The MIT Press, 2023; [https://direct.mit.edu/book/chapter-pdf/2155229/c011300_9780262376013.pdf]. [CrossRef]
- McMillen, P.; Levin, M. Collective intelligence: A unifying concept for integrating biology across scales and substrates. Communications Biology 2024, 7, 378. [Google Scholar] [CrossRef]
- Lagasse, E.; Levin, M. Future medicine: from molecular pathways to the collective intelligence of the body. Trends in Molecular Medicine 2023. [Google Scholar] [CrossRef]
- Levin, M. The computational boundary of a “self”: developmental bioelectricity drives multicellularity and scale-free cognition. Frontiers in psychology 2019, 10, 2688. [Google Scholar] [CrossRef] [PubMed]
- Pio-Lopez, L.; Levin, M. Aging as a loss of morphostatic information: a developmental bioelectricity perspective. Ageing Research Reviews 2024, p. 102310.
- Yang, J.H.; Hayano, M.; Griffin, P.T.; Amorim, J.A.; Bonkowski, M.S.; Apostolides, J.K.; Salfati, E.L.; Blanchette, M.; Munding, E.M.; Bhakta, M.; et al. Loss of epigenetic information as a cause of mammalian aging. Cell 2023, 186, 305–326.e27. [Google Scholar] [CrossRef] [PubMed]
- Pio-Lopez, L.; Kuchling, F.; Tung, A.; Pezzulo, G.; Levin, M. Active inference, morphogenesis, and computational psychiatry. Frontiers in computational neuroscience 2022, 16, 988977. [Google Scholar] [CrossRef]
- Montague, P.R.; Dolan, R.J.; Friston, K.J.; Dayan, P. Computational psychiatry. Trends in cognitive sciences 2012, 16, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Pezzulo, G.; Levin, M. Re-membering the body: applications of computational neuroscience to the top-down control of regeneration of limbs and other complex organs. Integrative Biology 2015, 7, 1487–1517. [Google Scholar] [CrossRef]
- Moskalev, A.; Guvatova, Z.; Lopes, I.D.A.; Beckett, C.W.; Kennedy, B.K.; De Magalhaes, J.P.; Makarov, A.A. Targeting aging mechanisms: pharmacological perspectives. Trends in Endocrinology and Metabolism 2022, 33, 266–280. [Google Scholar] [CrossRef]
- Kenyon, C.J. The genetics of ageing. Nature 2010, 464, 504–512. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Xu, L.; Brunet, A. Turning back time with emerging rejuvenation strategies. Nature cell biology 2019, 21, 32–43. [Google Scholar] [CrossRef]
- Pio-Lopez, L.; Levin, M. Morphoceuticals: Perspectives for discovery of drugs targeting anatomical control mechanisms in regenerative medicine, cancer and aging. Drug Discovery Today 2023, 28, 103585. [Google Scholar] [CrossRef]
- Levin, M. The wisdom of the body: future techniques and approaches to morphogenetic fields in regenerative medicine, developmental biology and cancer. Regenerative medicine 2011, 6, 667–673. [Google Scholar] [CrossRef]
- Sharpe, J. Computer modeling in developmental biology: growing today, essential tomorrow. Development 2017, 144, 4214–4225. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Escuredo, D.; del Álamo, J.C. How computation is helping unravel the dynamics of morphogenesis. Frontiers in Physics 2020, 8, 31. [Google Scholar] [CrossRef]
- Stillman, N.R.; Mayor, R. Generative models of morphogenesis in developmental biology. In Proceedings of the Seminars in Cell & Developmental Biology. Elsevier, 2023, Vol. 147, pp. 83–90.
- Mordvintsev, A.; Randazzo, E.; Niklasson, E.; Levin, M. Growing Neural Cellular Automata. Distill 2020, 5. [Google Scholar] [CrossRef]
- Pio-Lopez, L.; Bischof, J.; LaPalme, J.V.; Levin, M. The scaling of goals from cellular to anatomical homeostasis: an evolutionary simulation, experiment and analysis. Interface Focus 2023, 13, 20220072. [Google Scholar] [CrossRef]
- Hartl, B.; Risi, S.; Levin, M. Evolutionary Implications of Self-Assembling Cybernetic Materials with Collective Problem-Solving Intelligence at Multiple Scales. Entropy 2024, 26. [Google Scholar] [CrossRef]
- Grasso, C.; Bongard, J. Empowered neural cellular automata. In Proceedings of the Proceedings of the Genetic and Evolutionary Computation Conference Companion, 2022, pp. 108–111.
- Najarro, E.; Sudhakaran, S.; Glanois, C.; Risi, S. Hypernca: Growing developmental networks with neural cellular automata. arXiv preprint arXiv:2204.11674 2022. arXiv:2204.11674 2022.
- Mordvintsev, A.; Randazzo, E.; Fouts, C. Growing isotropic neural cellular automata. In Proceedings of the Artificial Life Conference Proceedings 34. MIT Press One Rogers Street, Cambridge, MA 02142-1209, USA journals-info …, 2022, Vol. 2022, p. 65.
- McAuley, M.T.; Kenny, R.A.; Kirkwood, T.B.; Wilkinson, D.J.; Jones, J.J.; Miller, V.M. A mathematical model of aging-related and cortisol induced hippocampal dysfunction. BMC neuroscience 2009, 10, 1–14. [Google Scholar] [CrossRef]
- Viktorov, A.; Kholodnov, V.; Gladkikh, V.; Alekhnovich, A. Influence of environment on aging of living systems: a mathematical model. Advances in Gerontology 2013, 3, 255–260. [Google Scholar] [CrossRef]
- Xue, H.; Xian, B.; Dong, D.; Xia, K.; Zhu, S.; Zhang, Z.; Hou, L.; Zhang, Q.; Zhang, Y.; Han, J.D.J. A modular network model of aging. Molecular systems biology 2007, 3, 147. [Google Scholar] [CrossRef]
- Mc Auley, M.T.; Mooney, K.M. Computational systems biology for aging research. Interdiscip Top Gerontol 2015, 40, 35–48. [Google Scholar]
- Cavuoti, L.; Sacco, F.; Randazzo, E.; Levin, M. Adversarial takeover of neural cellular automata. In Proceedings of the ALIFE 2022: The 2022 Conference on Artificial Life. MIT Press, 2022.
- Levin, M. Technological Approach to Mind Everywhere: An Experimentally-Grounded Framework for Understanding Diverse Bodies and Minds. Frontiers in Systems Neuroscience 2022, 16. [Google Scholar] [CrossRef]
- Pontes-Filho, S.; Walker, K.; Najarro, E.; Nichele, S.; Risi, S. A Unified Substrate for Body-Brain Co-evolution. In Proceedings of the From Cells to Societies: Collective Learning across Scales, 2022.
- Gawne, R.; McKenna, K.Z.; Levin, M. Competitive and coordinative interactions between body parts produce adaptive developmental outcomes. BioEssays 2020, 42, 1900245. [Google Scholar] [CrossRef] [PubMed]
- Noble, D. Genes and causation. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 2008, 366, 3001–3015. [Google Scholar] [CrossRef] [PubMed]
- Ellis, G.F. On the nature of causation in complex systems. Transactions of the Royal Society of South Africa 2008, 63, 69–84. [Google Scholar] [CrossRef]
- Auletta, G.; Ellis, G.F.; Jaeger, L. Top-down causation by information control: from a philosophical problem to a scientific research programme. Journal of the Royal Society Interface 2008, 5, 1159–1172. [Google Scholar] [CrossRef]
- Walker, S.I.; Cisneros, L.; Davies, P.C. Evolutionary transitions and top-down causation. arXiv preprint arXiv:1207.4808 2012.
- Scerri, E.R. Top-down causation regarding the chemistry–physics interface: a sceptical view. Interface Focus 2012, 2, 20–25. [Google Scholar] [CrossRef]
- Okasha, S. Emergence, hierarchy and top-down causation in evolutionary biology. Interface focus 2012, 2, 49–54. [Google Scholar] [CrossRef]
- Hall, D.G. Continuity and the persistence of objects: When the whole is greater than the sum of the parts. Cognitive Psychology 1998, 37, 28–59. [Google Scholar] [CrossRef]
- Mathews, J.; Chang, A.J.; Devlin, L.; Levin, M. Cellular signaling pathways as plastic, proto-cognitive systems: Implications for biomedicine. Patterns 2023, 4. [Google Scholar] [CrossRef]
- Langton, C.G. Artificial life: An overview; Mit Press, 1997.
- Von Neumann, J.; Burks, A.W.; et al. Theory of self-reproducing automata. IEEE Transactions on Neural Networks 1966, 5, 3–14. [Google Scholar]
- Wolfram, S. A New Kind of Science; Wolfram Media, 2002.
- Cook, M. Universality in Elementary Cellular Automata. Complex Syst. 2004, 15. [Google Scholar] [CrossRef]
- Chan, B.W.C. Lenia: Biology of Artificial Life. Complex Systems 2019, 28, 251–286. [Google Scholar] [CrossRef]
- Games, M. The fantastic combinations of John Conway’s new solitaire game “life” by Martin Gardner. Sci. Am. 1970, 223, 120–123. [Google Scholar]
- Li, X.; Yeh, A.G.O. Neural-network-based cellular automata for simulating multiple land use changes using GIS. Int. J. Geogr. Inf. Sci. 2002, 16, 323–343. [Google Scholar] [CrossRef]
- Hartl, B.; Levin, M.; Zöttl, A. Neuroevolution of Decentralized Decision-Making in N-Bead Swimmers Leads to Scalable and Robust Collective Locomotion. ArXiv [in preparation] 2024.
- McMillen, P.; Levin, M. Collective intelligence: A unifying concept for integrating biology across scales and substrates. Communications Biology 2024, 7. [Google Scholar] [CrossRef]
- Katz, Y.; Fontana, W. Probabilistic inference with polymerizing biochemical circuits. Entropy 2022, 24, 629. [Google Scholar] [CrossRef]
- Gyurkó, D.M.; Veres, D.V.; Módos, D.; Lenti, K.; Korcsmáros, T.; Csermely, P. Adaptation and learning of molecular networks as a description of cancer development at the systems-level: potential use in anti-cancer therapies. In Proceedings of the Seminars in Cancer Biology. Elsevier, 2013, Vol. 23, pp. 262–269.
- Csermely, P.; Kunsic, N.; Mendik, P.; Kerestély, M.; Faragó, T.; Veres, D.V.; Tompa, P. Learning of signaling networks: molecular mechanisms. Trends in biochemical sciences 2020, 45, 284–294. [Google Scholar] [CrossRef]
- Herrera-Delgado, E.; Perez-Carrasco, R.; Briscoe, J.; Sollich, P. Memory functions reveal structural properties of gene regulatory networks. PLoS computational biology 2018, 14, e1006003. [Google Scholar] [CrossRef]
- Gabalda-Sagarra, M.; Carey, L.B.; Garcia-Ojalvo, J. Recurrence-based information processing in gene regulatory networks. Chaos: An Interdisciplinary Journal of Nonlinear Science 2018, 28. [Google Scholar] [CrossRef]
- Hansen, N.; Ostermeier, A. Completely Derandomized Self-Adaptation in Evolution Strategies. Evolutionary Computation 2001, 9, 159–195. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Morrie, R.D.; Adams, D.S. V-ATPase-dependent ectodermal voltage and ph regionalization are required for craniofacial morphogenesis. Developmental Dynamics 2011, 240, 1889–1904. [Google Scholar] [CrossRef]
- Lizier, J.T.; Prokopenko, M.; Zomaya, A.Y. Local measures of information storage in complex distributed computation. Information Sciences 2012, 208, 39–54. [Google Scholar] [CrossRef]
- Schreiber, T. Measuring information transfer. Physical review letters 2000, 85, 461. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Morrie, R.D.; Adams, D.S. V-ATPase-dependent ectodermal voltage and pH regionalization are required for craniofacial morphogenesis. Developmental Dynamics 2011, 240, 1889–1904. [Google Scholar] [CrossRef] [PubMed]
- Sutton, R.S.; Barto, A.G. Reinforcement Learning: An Introduction; The MIT Press, 2018.
- Tang, Y.; Ha, D. The Sensory Neuron as a Transformer: Permutation-Invariant Neural Networks for Reinforcement Learning. In Proceedings of the Advances in Neural Information Processing Systems; Beygelzimer, A.; Dauphin, Y.; Liang, P.; Vaughan, J.W., Eds., 2021.
- Rumelhart, D.E.; Hinton, G.E.; Williams, R.J., Learning internal representations by error propagation. In Parallel Distributed Processing: Explorations in the Microstructure of Cognition, Vol. 1: Foundations; MIT Press: Cambridge, MA, USA, 1986; p. 318–362.
- Hiscock, T.W. Adapting machine-learning algorithms to design gene circuits. BMC Bioinformatics 2019, 20, 214. [Google Scholar] [CrossRef]
- MacKay, D.J.; Mac Kay, D.J.; et al. Information theory, inference and learning algorithms; Cambridge university press, 2003.
- Mitchell, A.; Lim, W. Cellular perception and misperception: Internal models for decision-making shaped by evolutionary experience. BioEssays 2016, 38, 845–849. [Google Scholar] [CrossRef]
- Aasen, T.; Hodgins, M.B.; Edward, M.; Graham, S.V. The relationship between connexins, gap junctions, tissue architecture and tumour invasion, as studied in a novel in vitro model of HPV-16-associated cervical cancer progression. Oncogene 2003, 22, 7969–7980. [Google Scholar] [CrossRef]
- Leithe, E.; Sirnes, S.; Omori, Y.; Rivedal, E. Downregulation of gap junctions in cancer cells. Critical Reviews™ in Oncogenesis 2006, 12.
- Levin, M. Bioelectrical approaches to cancer as a problem of the scaling of the cellular self. Progress in biophysics and molecular biology 2021, 165, 102–113. [Google Scholar] [CrossRef]
- Miller, H.A.; Dean, E.S.; Pletcher, S.D.; Leiser, S.F. Cell non-autonomous regulation of health and longevity. Elife 2020, 9, e62659. [Google Scholar] [CrossRef]
- Turner, B.M. Epigenetic responses to environmental change and their evolutionary implications. Philosophical Transactions of the Royal Society B: Biological Sciences 2009, 364, 3403–3418. [Google Scholar] [CrossRef]
- Brockes, J.P. Amphibian limb regeneration: rebuilding a complex structure. Science 1997, 276, 81–87. [Google Scholar] [CrossRef]
- Henry, J.J.; Tsonis, P.A. Molecular and cellular aspects of amphibian lens regeneration. Progress in retinal and eye research 2010, 29, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Maden, M. Axolotl/newt. Molecular Embryology: Methods and Protocols 1999, pp. 415–428.
- Reddien, P.W.; Alvarado, A.S. Fundamentals of planarian regeneration. Annu. Rev. Cell Dev. Biol. 2004, 20, 725–757. [Google Scholar] [CrossRef] [PubMed]
- Owlarn, S.; Bartscherer, K. Go ahead, grow a head! A planarian’s guide to anterior regeneration. Regeneration 2016, 3, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. Reprogramming cells and tissue patterning via bioelectrical pathways: molecular mechanisms and biomedical opportunities. Wiley Interdisciplinary Reviews: Systems Biology and Medicine 2013, 5, 657–676. [Google Scholar] [CrossRef]
- Haluza, Y.; Zoller, J.A.; Lu, A.T.; Walters, H.E.; Lachnit, M.; Lowe, R.; Haghani, A.; Brooke, R.T.; Park, N.; Yun, M.H.; et al. Axolotl epigenetic clocks offer insights into the nature of negligible senescence. BioRxiv 2024, pp. 2024–09.
- Mašek, J.; Andersson, E.R. The developmental biology of genetic Notch disorders. Development 2017, 144, 1743–1763. [Google Scholar] [CrossRef]
- Pai, V.P.; Lemire, J.M.; Paré, J.F.; Lin, G.; Chen, Y.; Levin, M. Endogenous gradients of resting potential instructively pattern embryonic neural tissue via notch signaling and regulation of proliferation. Journal of Neuroscience 2015, 35, 4366–4385. [Google Scholar] [CrossRef]
- Pai, V.P.; Cervera, J.; Mafe, S.; Willocq, V.; Lederer, E.K.; Levin, M. HCN2 channel-induced rescue of brain teratogenesis via local and long-range bioelectric repair. Frontiers in Cellular Neuroscience 2020, 14, 136. [Google Scholar] [CrossRef]
- Pai, V.P.; Levin, M. HCN2 channel-induced rescue of brain, eye, heart and gut teratogenesis caused by nicotine, ethanol and aberrant notch signalling. Wound Repair and Regeneration 2022, 30, 681–706. [Google Scholar] [CrossRef]
- Levin, M.; Selberg, J.; Rolandi, M. Endogenous bioelectrics in development, cancer, and regeneration: drugs and bioelectronic devices as electroceuticals for regenerative medicine. Iscience 2019, 22, 519–533. [Google Scholar] [CrossRef]
- Whited, J.L.; Levin, M. Bioelectrical controls of morphogenesis: from ancient mechanisms of cell coordination to biomedical opportunities. Current opinion in genetics & development 2019, 57, 61–69. [Google Scholar]
- Dause, T.J.; Kirby, E.D. Aging gracefully: social engagement joins exercise and enrichment as a key lifestyle factor in resistance to age-related cognitive decline. Neural regeneration research 2019, 14, 39–42. [Google Scholar] [PubMed]
- Bettio, L.E.; Rajendran, L.; Gil-Mohapel, J. The effects of aging in the hippocampus and cognitive decline. Neuroscience & Biobehavioral Reviews 2017, 79, 66–86. [Google Scholar]
- Fratiglioni, L.; Paillard-Borg, S.; Winblad, B. An active and socially integrated lifestyle in late life might protect against dementia. The Lancet Neurology 2004, 3, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Truman, J.W. The evolution of insect metamorphosis. Current Biology 2019, 29, R1252–R1268. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.D.; Cai, L. Amphibian metamorphosis. Developmental biology 2007, 306, 20–33. [Google Scholar] [CrossRef]
- Tung, A.; Sperry, M.M.; Clawson, W.; Pavuluri, A.; Bulatao, S.; Yue, M.; Flores, R.M.; Pai, V.P.; McMillen, P.; Kuchling, F.; et al. Embryos assist morphogenesis of others through calcium and ATP signaling mechanisms in collective teratogen resistance. Nature Communications 2024, 15, 535. [Google Scholar] [CrossRef]
- Chen, L.; Zaharia, M.; Zou, J. How is ChatGPT’s behavior changing over time? arXiv preprint arXiv:2307.09009 2023.
Figure 1.
(A) A schematic illustration of the stages of life of a complex organism, starting from embryogenesis, development and growth, through maturity, decline, aging, and eventually death. (B) Multicellular organisms are composites of hierarchically interlocked layers of biological self-organization, spanning physical scales from the molecular level, over cells, tissues, organs, to organisms and even groups or collectives of individuals. (C) To develop an organism’s morphology, and sustain its integrity against environmental perturbations, biology maintains a multi-scale closed loop pattern homeostasis mechanism, where the components at every scale of self-organization are capable of self-regulatory and error-correcting behavior. Fundamentally, cells themselves have numerous behavioral and information processing capabilities: To control multicellular morphology during embryogenesis and also in mature and even aging tissue, cells utilize a variety of communication channels, such as bioelectrical, biochemical, and biomechanical processes, enacting local communication protocols via intercellular signaling, intracellular information processing, and cell-state regulation2. This can be seen as a process of physiological computation by which cells interact and respond to their environment to create complex patterns and structures, guiding embryogenesis, maturation, metamorphosis, remodeling, regeneration, and suppression of cancer and aging3. In most organisms, with only very few exceptions, these homeostatic processes eventually break down, leading to irreversible morphological decline, aging, and death.
Figure 1.
(A) A schematic illustration of the stages of life of a complex organism, starting from embryogenesis, development and growth, through maturity, decline, aging, and eventually death. (B) Multicellular organisms are composites of hierarchically interlocked layers of biological self-organization, spanning physical scales from the molecular level, over cells, tissues, organs, to organisms and even groups or collectives of individuals. (C) To develop an organism’s morphology, and sustain its integrity against environmental perturbations, biology maintains a multi-scale closed loop pattern homeostasis mechanism, where the components at every scale of self-organization are capable of self-regulatory and error-correcting behavior. Fundamentally, cells themselves have numerous behavioral and information processing capabilities: To control multicellular morphology during embryogenesis and also in mature and even aging tissue, cells utilize a variety of communication channels, such as bioelectrical, biochemical, and biomechanical processes, enacting local communication protocols via intercellular signaling, intracellular information processing, and cell-state regulation2. This can be seen as a process of physiological computation by which cells interact and respond to their environment to create complex patterns and structures, guiding embryogenesis, maturation, metamorphosis, remodeling, regeneration, and suppression of cancer and aging3. In most organisms, with only very few exceptions, these homeostatic processes eventually break down, leading to irreversible morphological decline, aging, and death.
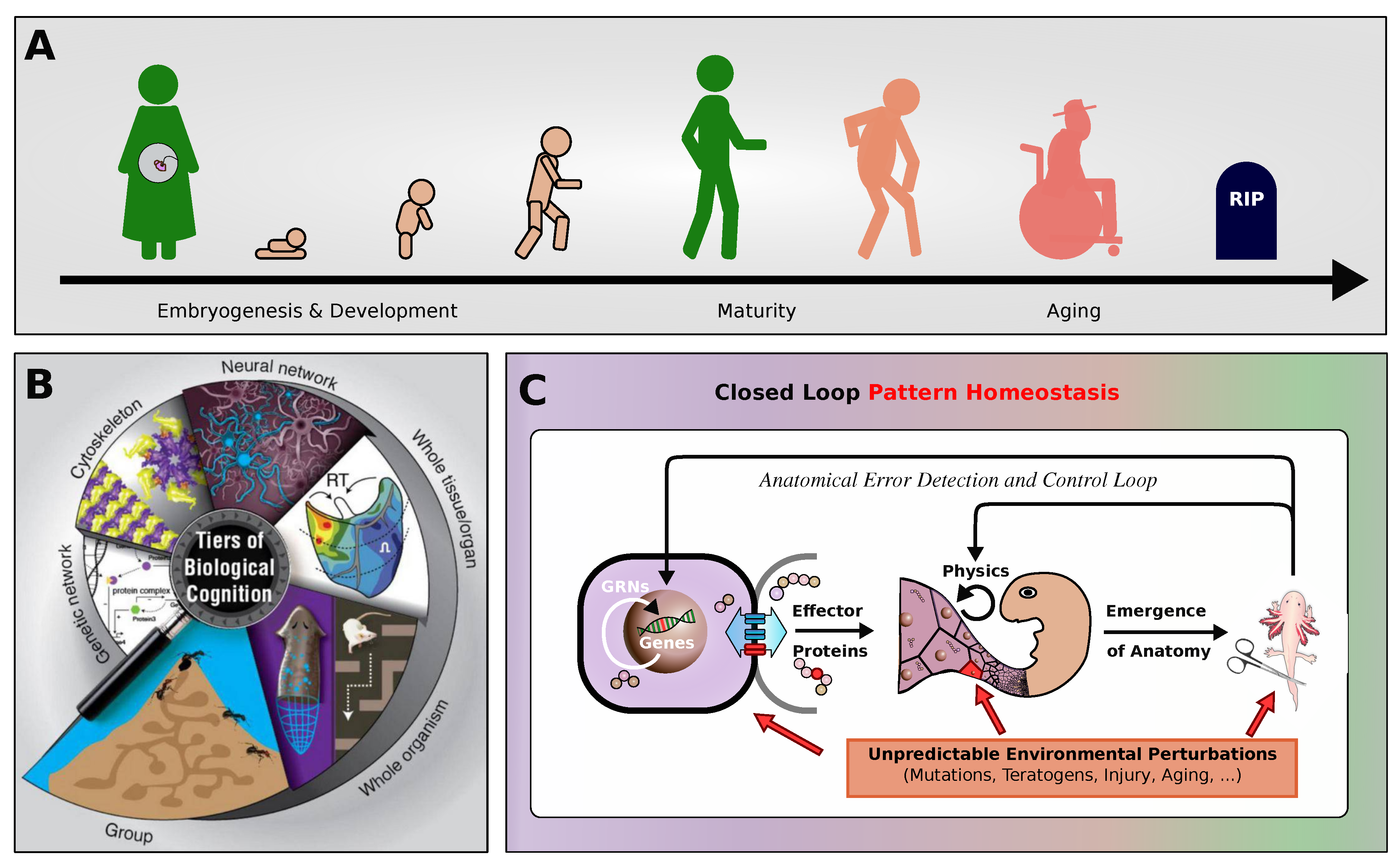
Figure 2.
(A): (A): Schematic computational model of a biologically inspired multi-scale competency architecture [19], relying on Neural Cellular Automata [39,67] (NCAs) in an evolutionary setting [41]:: in an evolutionary NCA, a genetic code, i.e., a cell’s “DNA” (represented by a string of numerical parameters), is compiled into a uni-cellular phenotype containing a proto-cognitive decision-making center (represented by a tiny recurrent neural network parameterized by the “DNA”) by which the cellular agent can actively regulate its own numerical cell state based on local measurements of the states of its immediate cellular neighbors on a square grid of cells. In that way, the collective of cells can be trained, or evolved, to perform morphogenesis of a predefined target pattern of expressed cell types (color-coded in blue, magenta, and white), of, e.g., a smiley-face pattern (reminiscent of the bioelectric craniofacial prepattern defining the amphibian face [79]. (B): : Detailed information flowchart of uni-cellular decision-making of cells on the grid of a NCA, modeling the process of morphogenesis (c.f., (A)): each cell expresses its cell-type based on local communication protocols with its neighboring cells so the collective of cells self-assembles a target pattern. This is implemented via subsequent uni-cellular perception-action cycles that update each cell’s respective state on the NCA’s grid purely based on local measurements of neighboring cell states. (C): Schematics of the evolutionary process of uni-cellular competencies that drive the self-orchestrated morphogenesis of a target pattern (here, of a smiley, face). (D): Schematics of long-term behavior of an NCA evolved in (C) that is not controlled by the evolutionary process. Developmental errors or noise (red wiggly arrows) might lead to a collapse of the target pattern over time, especially long after the selection process occurs (where genetic material is passed on to the next generation. Following pleiotropic considerations, we assume that the long-term “solutions” or the target morphogenetic state might significantly differ from the target state that the evolutionary process “sees” through selection, simply because different states might be more probable (such as, here, a “red” circle without facial features). Moreover, detecting such deviating pleiotropic defects (missing facial features, in our case), and performing targeted interventions of the most affected cells allow us to “reprogram“ the cellular collective and reset it via targeted interventions - without further optimization or adaptation - so the tissue auto-regenerates to the original target pattern.
Figure 2.
(A): (A): Schematic computational model of a biologically inspired multi-scale competency architecture [19], relying on Neural Cellular Automata [39,67] (NCAs) in an evolutionary setting [41]:: in an evolutionary NCA, a genetic code, i.e., a cell’s “DNA” (represented by a string of numerical parameters), is compiled into a uni-cellular phenotype containing a proto-cognitive decision-making center (represented by a tiny recurrent neural network parameterized by the “DNA”) by which the cellular agent can actively regulate its own numerical cell state based on local measurements of the states of its immediate cellular neighbors on a square grid of cells. In that way, the collective of cells can be trained, or evolved, to perform morphogenesis of a predefined target pattern of expressed cell types (color-coded in blue, magenta, and white), of, e.g., a smiley-face pattern (reminiscent of the bioelectric craniofacial prepattern defining the amphibian face [79]. (B): : Detailed information flowchart of uni-cellular decision-making of cells on the grid of a NCA, modeling the process of morphogenesis (c.f., (A)): each cell expresses its cell-type based on local communication protocols with its neighboring cells so the collective of cells self-assembles a target pattern. This is implemented via subsequent uni-cellular perception-action cycles that update each cell’s respective state on the NCA’s grid purely based on local measurements of neighboring cell states. (C): Schematics of the evolutionary process of uni-cellular competencies that drive the self-orchestrated morphogenesis of a target pattern (here, of a smiley, face). (D): Schematics of long-term behavior of an NCA evolved in (C) that is not controlled by the evolutionary process. Developmental errors or noise (red wiggly arrows) might lead to a collapse of the target pattern over time, especially long after the selection process occurs (where genetic material is passed on to the next generation. Following pleiotropic considerations, we assume that the long-term “solutions” or the target morphogenetic state might significantly differ from the target state that the evolutionary process “sees” through selection, simply because different states might be more probable (such as, here, a “red” circle without facial features). Moreover, detecting such deviating pleiotropic defects (missing facial features, in our case), and performing targeted interventions of the most affected cells allow us to “reprogram“ the cellular collective and reset it via targeted interventions - without further optimization or adaptation - so the tissue auto-regenerates to the original target pattern.
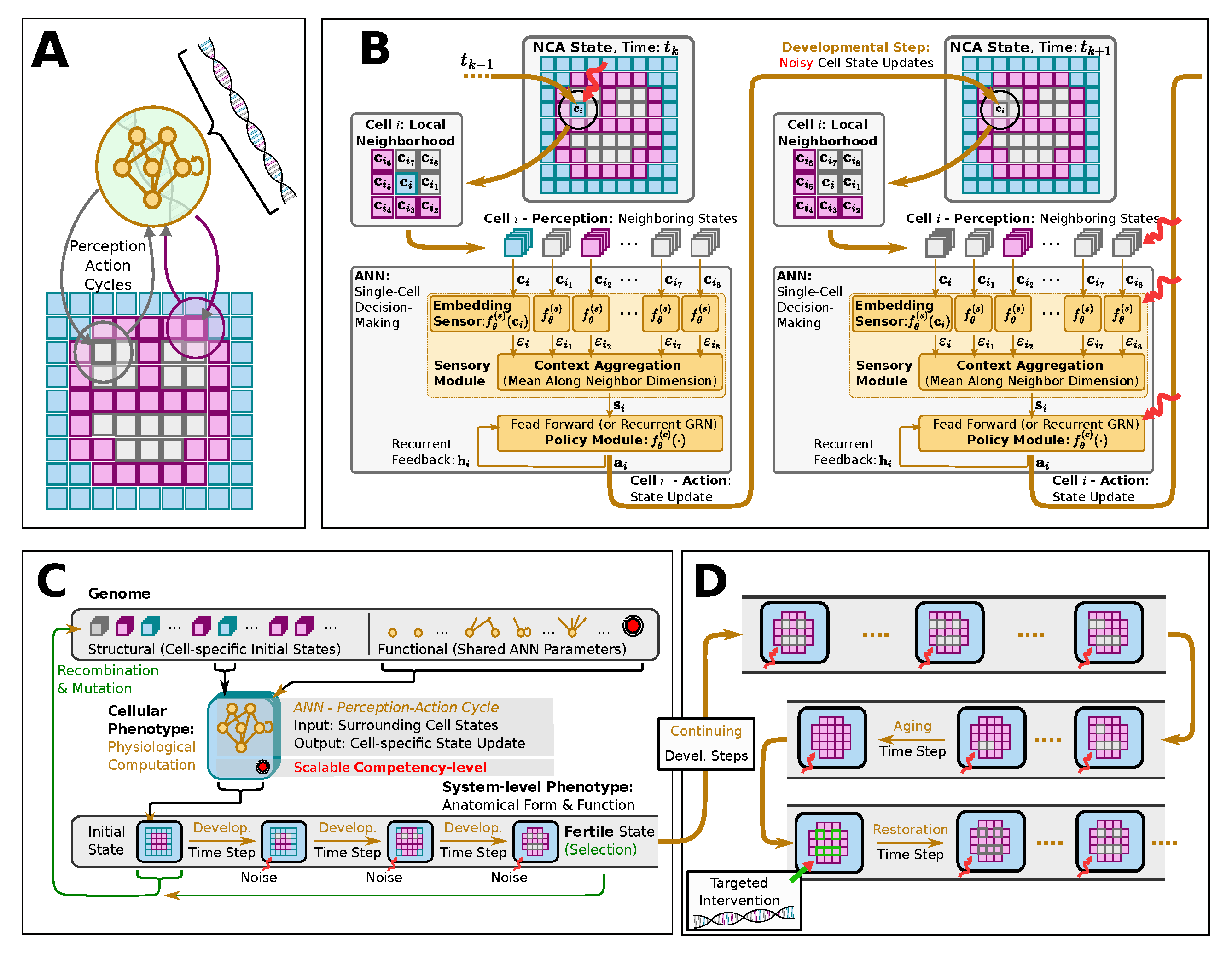
Figure 3.
Typical fitness score evaluations during developmental steps of morphogenesis of 50 statistically independent runs of an NCA that has been evolved to self-assemble a -smiley-face pattern. Example NCA states from the individual with the highest developmental fitness score (c.f., red “elite trajectory” in bottom panel) are presented throughout the developmental phase as temporal snapshots corresponding to the ticks of the horizontal axis; background-, facial-, and internal organ cells (of eyes and mouth) are colored purple, yellow, and red, respectively.
Figure 3.
Typical fitness score evaluations during developmental steps of morphogenesis of 50 statistically independent runs of an NCA that has been evolved to self-assemble a -smiley-face pattern. Example NCA states from the individual with the highest developmental fitness score (c.f., red “elite trajectory” in bottom panel) are presented throughout the developmental phase as temporal snapshots corresponding to the ticks of the horizontal axis; background-, facial-, and internal organ cells (of eyes and mouth) are colored purple, yellow, and red, respectively.
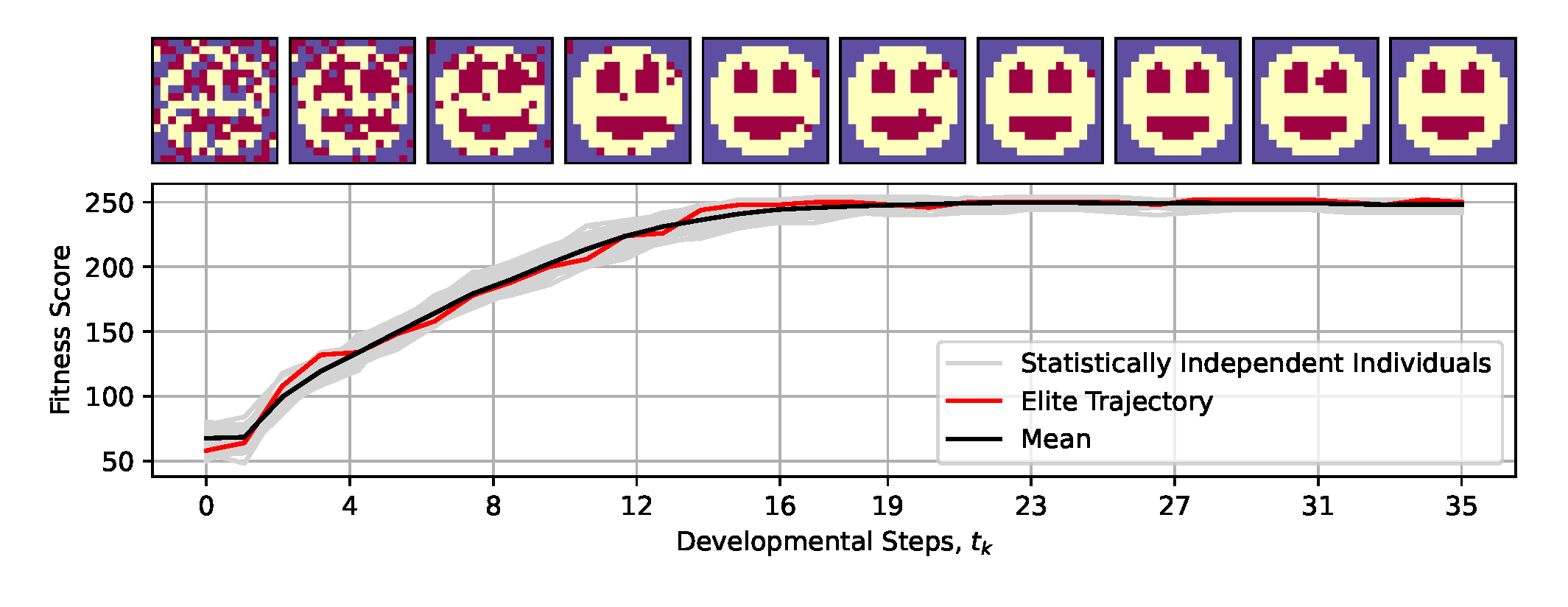
Figure 4.
Same as fig:system:morphogenesis, but evaluated for developmental (or NCA-) time steps, furthermore displaying multiple example trajectories of NCA-states (color-coded): The blue-emphasized individual maintains an approximate smiley-face pattern for exceptionally long times, and thus displays the highest long-term fitness score. The orange- and green-emphasized individuals lose, respectively, either a left or right eye at steps, and both eyes at steps, but maintain their mouth for long times; these individuals display close-to-average long-term fitness scores. The red-emphasized individual eventually loses the mouth, thus displaying the lowest long-term fitness score among the presented trajectories. The spherical face structure seems incredibly robust.
Figure 4.
Same as fig:system:morphogenesis, but evaluated for developmental (or NCA-) time steps, furthermore displaying multiple example trajectories of NCA-states (color-coded): The blue-emphasized individual maintains an approximate smiley-face pattern for exceptionally long times, and thus displays the highest long-term fitness score. The orange- and green-emphasized individuals lose, respectively, either a left or right eye at steps, and both eyes at steps, but maintain their mouth for long times; these individuals display close-to-average long-term fitness scores. The red-emphasized individual eventually loses the mouth, thus displaying the lowest long-term fitness score among the presented trajectories. The spherical face structure seems incredibly robust.
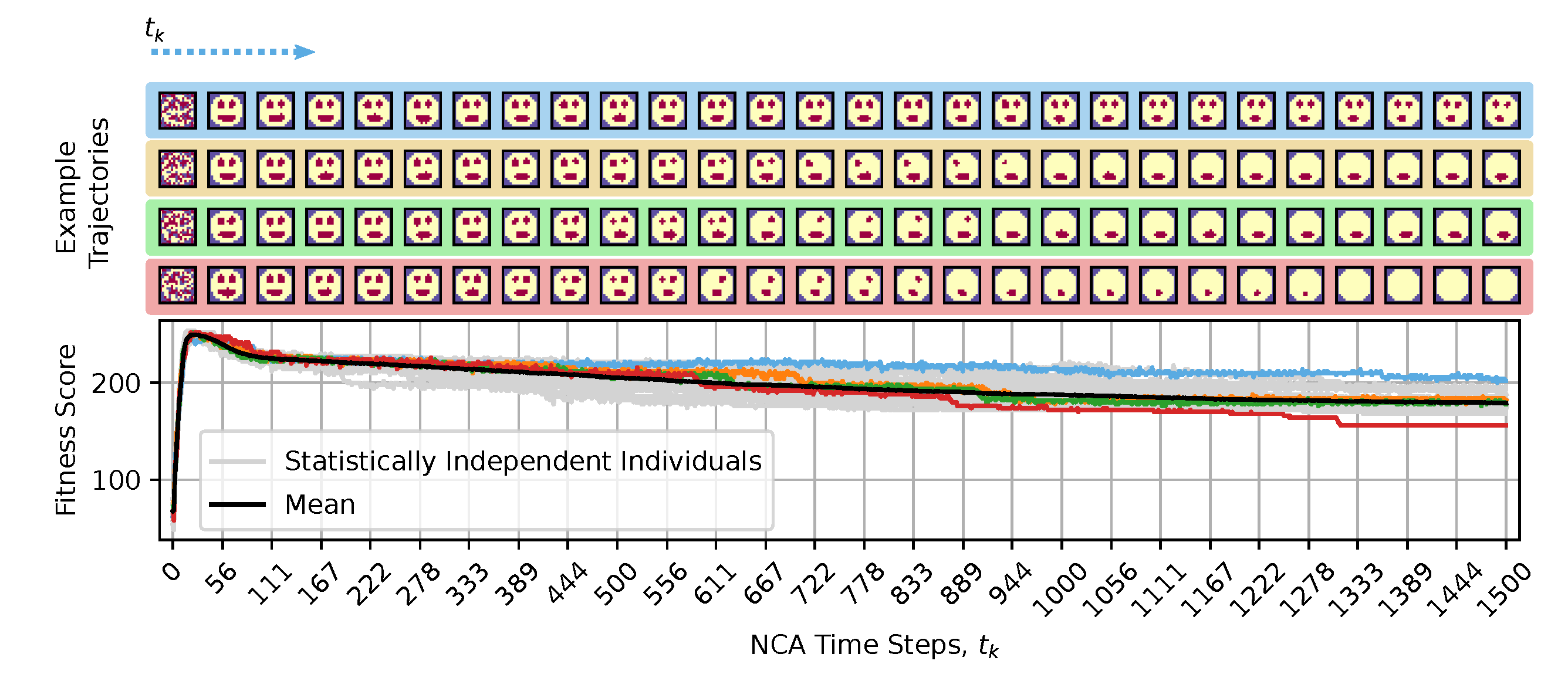
Figure 5.
Panels (A-D): Schematic visualization of the amplified damaging processes to cellular competencies corresponding to sec:results:induced-degeneracy:noise,sec:results:induced-degeneracy:competency,sec:results:induced-degeneracy:cell-communication,sec:results:induced-degeneracy:ann-damage, respectively. (A): The fitness over the lifetime of the smiley-face NCA discussed in sec:system where the noise level for cell-state updates is linearly increased from to (emphasized by red line), for time steps ; gray lines show statistically independent trajectories of individual lifetimes of the NCA, the blue line represents the corresponding ensemble mean, and the black-dashed line is the baseline mean fitness without corrupted competencies (c.f., fig:result:pleiotropy). (B): similar to (A) but corrupting cellular competencies via a successively decreasing decision-making probability to following the functional form (see red line). (C): similar to (A, B) but permanently corrupting the intercellular communication pathways by successively disabling randomly selected communication channels (reminiscent of gap-junctions [50]) between neighboring cells by linearly decreasing the gap-junction prohibiting probability from to (see text and red line). (D): similar to (A-C) but accumulating genetic damage in the unicellular decision-making machinery, i.e.,, corrupting intracellular information processing and decision-making by adding zero-centered Gaussian noise of standard deviation at every time step to the cells’ ANN parameters (which is equivalent to ANN parameters corrupted by at time-step with ). In (A-D), the top panels schematically illustrate the perception-action cycles of every cell over the lifespan of the NCA; the particular processes affecting the cellular competencies, thus enhancing the rate of degradation of the target pattern in each panel (A-D), are emphasized from left to right by red blurry blocks: this involves (A) increasingly misdifferentiation and misperception of cell states by neighbours, (B) increasingly unreliable cell decisions directly affecting the NCA’s action output, (C) successively corrupted intercellular communication channels directly affecting the exchange of information between neighbors, and (D) accumulation of genetic damage by successively diffusing the cells’ ANN parameters directly affecting the unicellular decision-making machinery.
Figure 5.
Panels (A-D): Schematic visualization of the amplified damaging processes to cellular competencies corresponding to sec:results:induced-degeneracy:noise,sec:results:induced-degeneracy:competency,sec:results:induced-degeneracy:cell-communication,sec:results:induced-degeneracy:ann-damage, respectively. (A): The fitness over the lifetime of the smiley-face NCA discussed in sec:system where the noise level for cell-state updates is linearly increased from to (emphasized by red line), for time steps ; gray lines show statistically independent trajectories of individual lifetimes of the NCA, the blue line represents the corresponding ensemble mean, and the black-dashed line is the baseline mean fitness without corrupted competencies (c.f., fig:result:pleiotropy). (B): similar to (A) but corrupting cellular competencies via a successively decreasing decision-making probability to following the functional form (see red line). (C): similar to (A, B) but permanently corrupting the intercellular communication pathways by successively disabling randomly selected communication channels (reminiscent of gap-junctions [50]) between neighboring cells by linearly decreasing the gap-junction prohibiting probability from to (see text and red line). (D): similar to (A-C) but accumulating genetic damage in the unicellular decision-making machinery, i.e.,, corrupting intracellular information processing and decision-making by adding zero-centered Gaussian noise of standard deviation at every time step to the cells’ ANN parameters (which is equivalent to ANN parameters corrupted by at time-step with ). In (A-D), the top panels schematically illustrate the perception-action cycles of every cell over the lifespan of the NCA; the particular processes affecting the cellular competencies, thus enhancing the rate of degradation of the target pattern in each panel (A-D), are emphasized from left to right by red blurry blocks: this involves (A) increasingly misdifferentiation and misperception of cell states by neighbours, (B) increasingly unreliable cell decisions directly affecting the NCA’s action output, (C) successively corrupted intercellular communication channels directly affecting the exchange of information between neighbors, and (D) accumulation of genetic damage by successively diffusing the cells’ ANN parameters directly affecting the unicellular decision-making machinery.
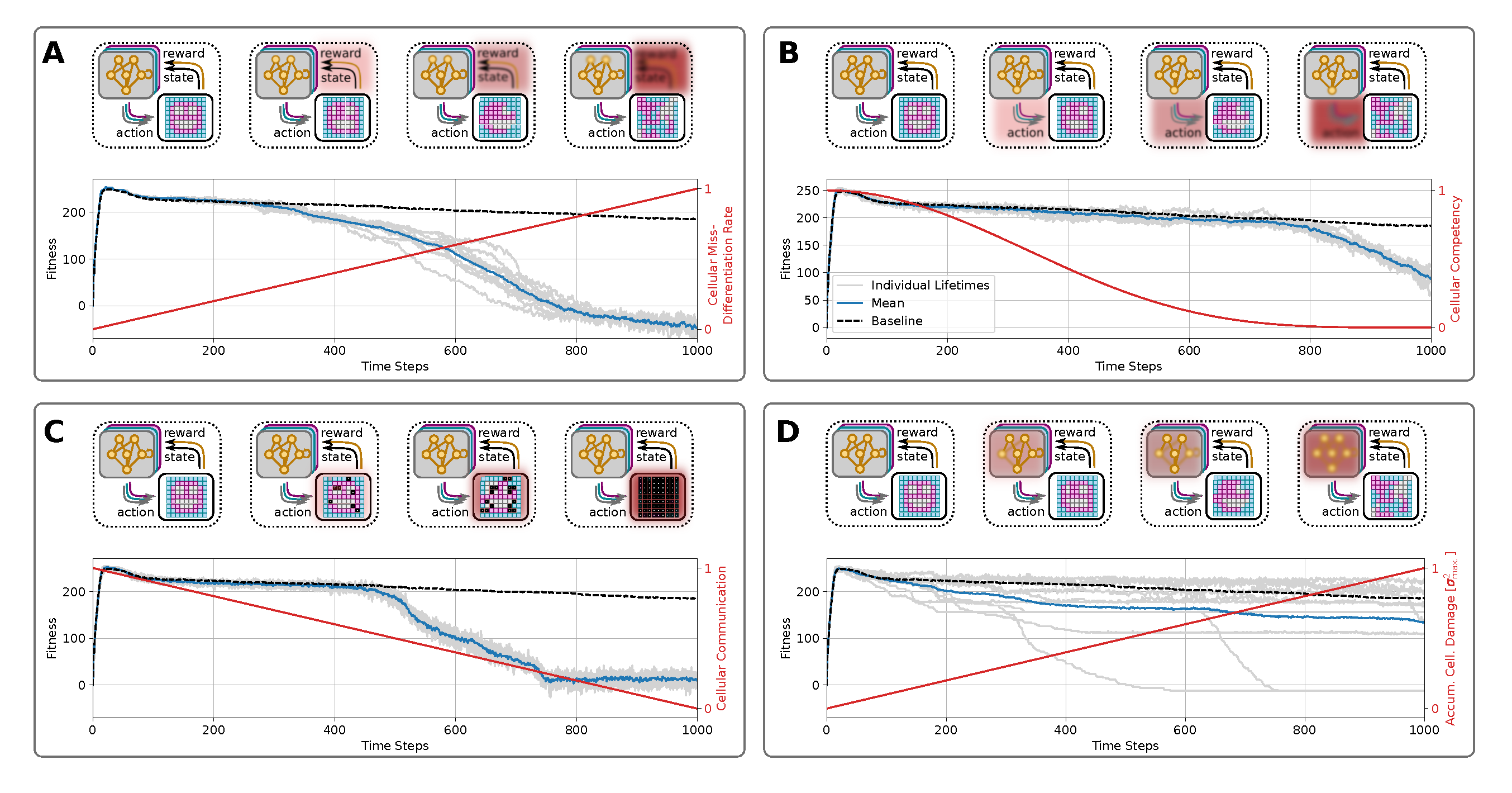
Figure 6.
Active information storage (AIS), transfer entropy (TE) and spatial entropy (SE) during development and aging for 6 conditions: default behavior, noise, competency, connectivity, the alterations of ANN parameters, and noise and competency. Left: in blue we have the mean of the AIS for the 50 organisms, in light grey the individual AIS mean per organism. Middle: in red we have the mean of the TE for the 50 organisms, in light grey the individual TE mean per organism. Right: in green we have the mean of the SE for the 50 organisms, in light grey the individual SE mean per organism. At different stages, we show the states of the organism.
Figure 6.
Active information storage (AIS), transfer entropy (TE) and spatial entropy (SE) during development and aging for 6 conditions: default behavior, noise, competency, connectivity, the alterations of ANN parameters, and noise and competency. Left: in blue we have the mean of the AIS for the 50 organisms, in light grey the individual AIS mean per organism. Middle: in red we have the mean of the TE for the 50 organisms, in light grey the individual TE mean per organism. Right: in green we have the mean of the SE for the 50 organisms, in light grey the individual SE mean per organism. At different stages, we show the states of the organism.
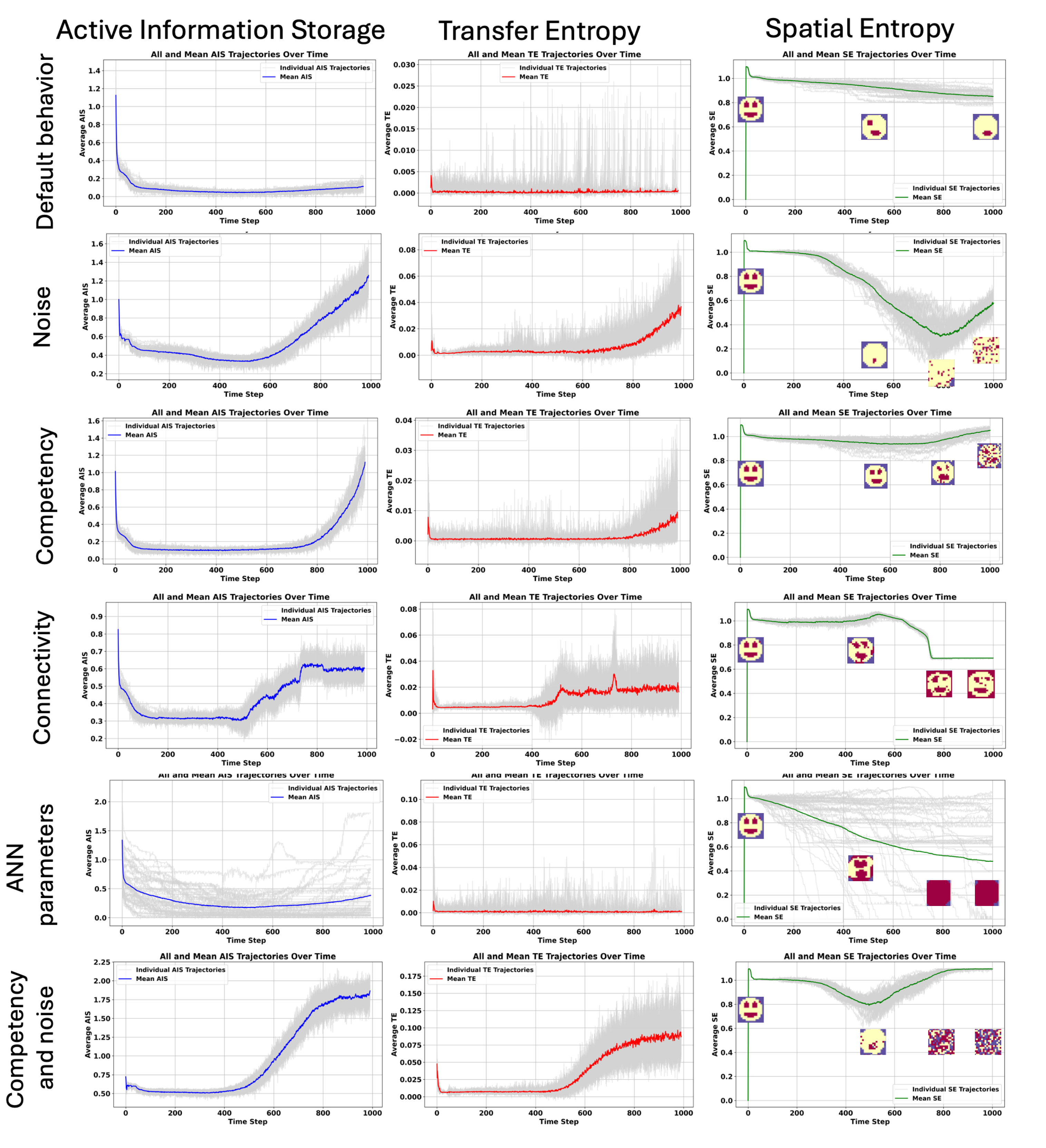
Figure 7.
. Loss of the organs but not of the information about the organ during normal aging. In order to study the information dynamics during aging and morphological decay, we applied AIS and TE during development and aging. Here is an example of a morphogenetic experiment where the organism is losing organs, here the eyes of the smiley face at step 500 in States column. But both AIS and TE are high at the spatial location of these organs (from 500 to 800 steps). This suggests memory retention, similarly to regenerative organisms that can regrow lost organs or limbs, keeping some memory of the lost body parts. This result will lead to the organ regeneration experiment (see Figure 8).
Figure 7.
. Loss of the organs but not of the information about the organ during normal aging. In order to study the information dynamics during aging and morphological decay, we applied AIS and TE during development and aging. Here is an example of a morphogenetic experiment where the organism is losing organs, here the eyes of the smiley face at step 500 in States column. But both AIS and TE are high at the spatial location of these organs (from 500 to 800 steps). This suggests memory retention, similarly to regenerative organisms that can regrow lost organs or limbs, keeping some memory of the lost body parts. This result will lead to the organ regeneration experiment (see Figure 8).
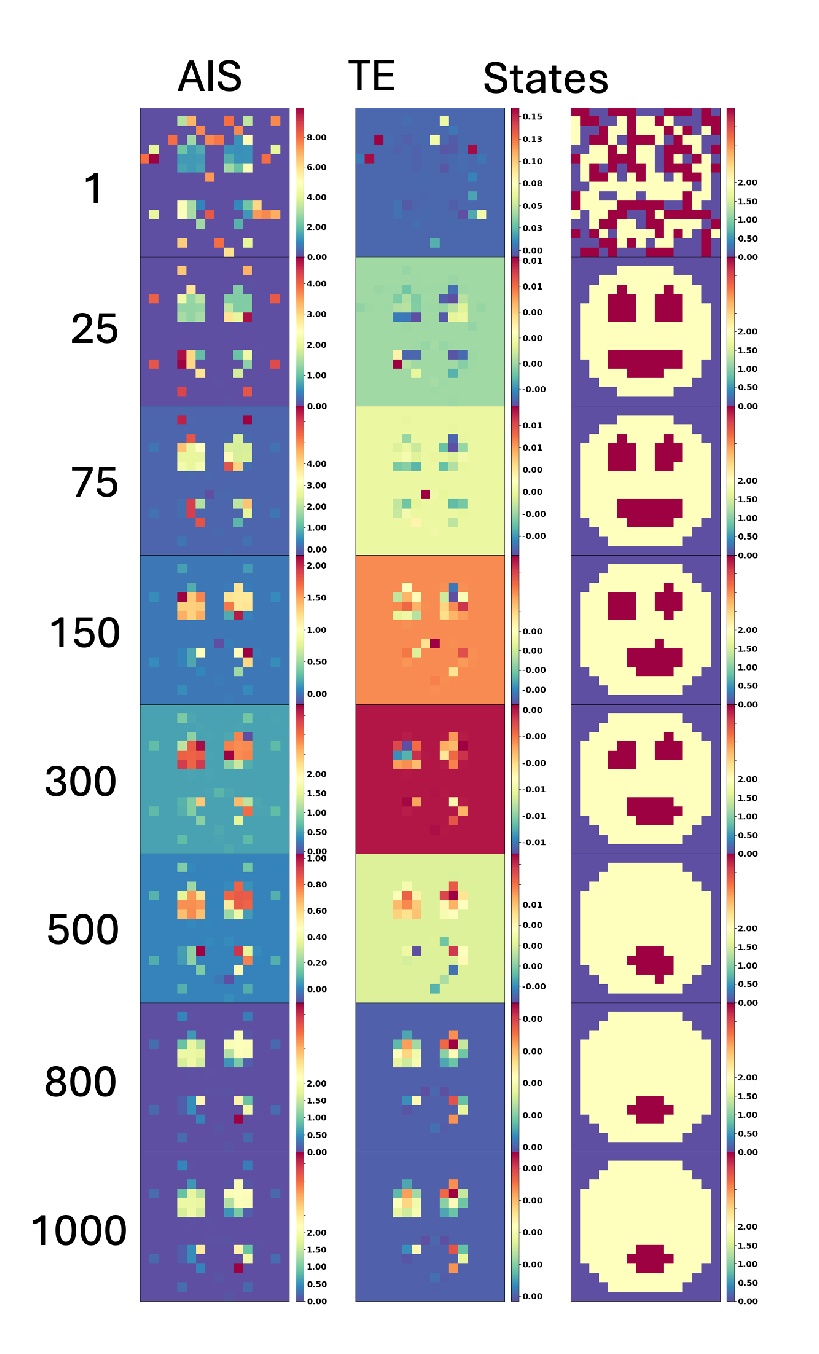
Figure 8.
Organ restoration experiments. (A) by replacing the entire affected organs, i.e.,, the left eye (blue color code), the right eye (green colored), and the mouth (red colored) with their corresponding initial states if the respective organ-specific score is lower than the threshold score of . (B): same as (A) but only reconfiguring the wrongly expressed cells, not the entire organ, of an affected organ. In both, (A, B), the top-left panel presents the mean fitness score (blue) of 10 statistically independent individual simulations of the respective recovery experiments (green), which we contrast with the baseline case without recovery depicted in fig:result:pleiotropy. The four bottom-left panels in (A) and (primordial) respectively present the organ-specific scores (color-coded) of four selected individual lifespan simulations from the top panels; colored stars indicate rejuvenation events. The bar plots indicate the total number of organ-specific rejuvenation events for all trajectories (top right panel) and the selected individual trajectories (bottom right panels) again respectively for (A) and (B). The cartoon inset in the top panel of (B) indicates the respective loci in the NCA used to evaluate the respective organ-specific fitness score (i.e.,, the blue, green, and red organ masks). While the strategy in (A) of replacing detrimentally affected organs entirely with their respective primordial cells does not seem effective (many repetitive interventions are required as a form of symptom control), for the majority of individual simulations in (B), typically only very few intervention events of resetting the affected cells of a corrupted organ to their primordial state suffice to restore sustainable system-level performance.
Figure 8.
Organ restoration experiments. (A) by replacing the entire affected organs, i.e.,, the left eye (blue color code), the right eye (green colored), and the mouth (red colored) with their corresponding initial states if the respective organ-specific score is lower than the threshold score of . (B): same as (A) but only reconfiguring the wrongly expressed cells, not the entire organ, of an affected organ. In both, (A, B), the top-left panel presents the mean fitness score (blue) of 10 statistically independent individual simulations of the respective recovery experiments (green), which we contrast with the baseline case without recovery depicted in fig:result:pleiotropy. The four bottom-left panels in (A) and (primordial) respectively present the organ-specific scores (color-coded) of four selected individual lifespan simulations from the top panels; colored stars indicate rejuvenation events. The bar plots indicate the total number of organ-specific rejuvenation events for all trajectories (top right panel) and the selected individual trajectories (bottom right panels) again respectively for (A) and (B). The cartoon inset in the top panel of (B) indicates the respective loci in the NCA used to evaluate the respective organ-specific fitness score (i.e.,, the blue, green, and red organ masks). While the strategy in (A) of replacing detrimentally affected organs entirely with their respective primordial cells does not seem effective (many repetitive interventions are required as a form of symptom control), for the majority of individual simulations in (B), typically only very few intervention events of resetting the affected cells of a corrupted organ to their primordial state suffice to restore sustainable system-level performance.
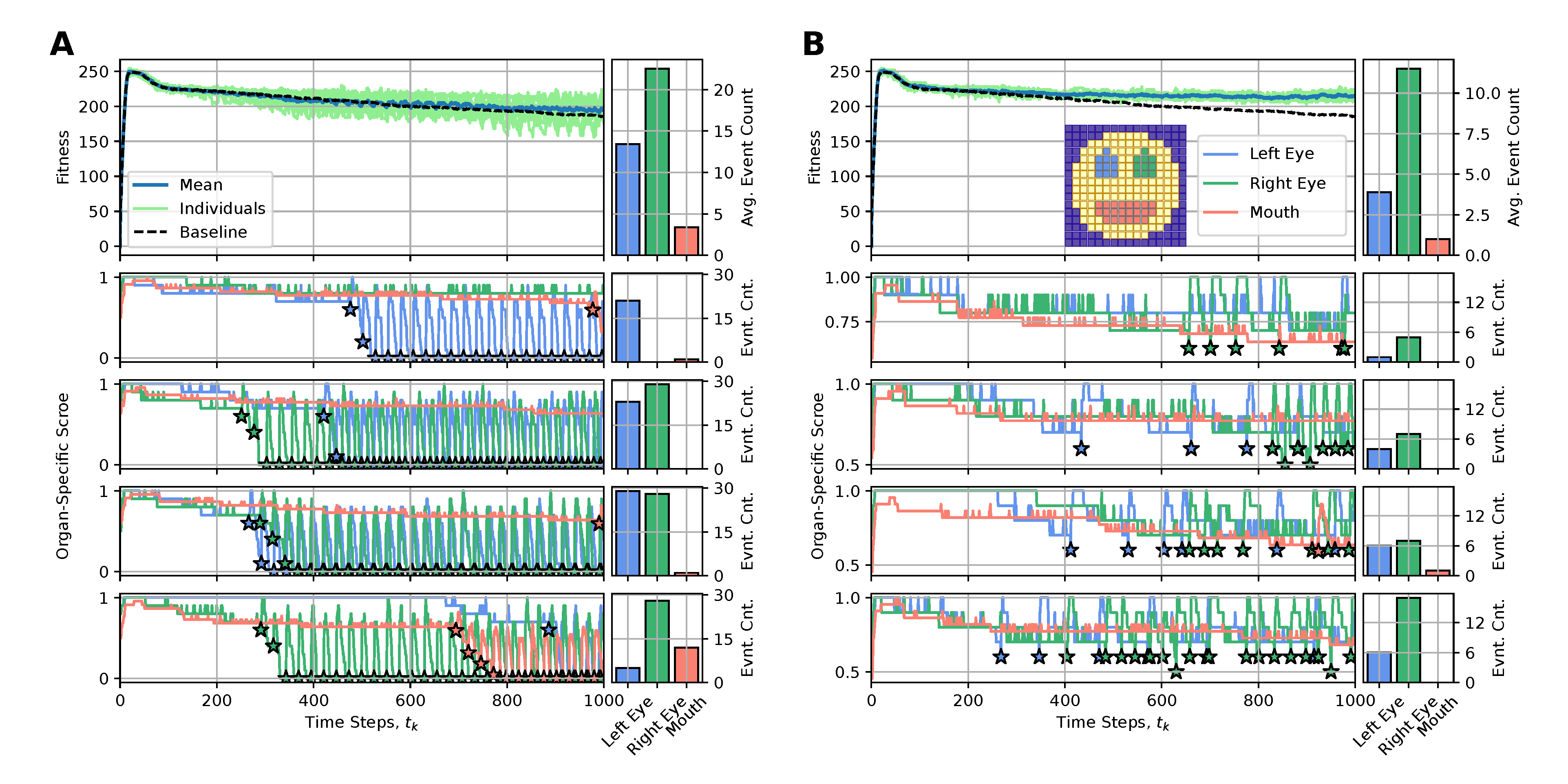
Figure 9.
Organ and surrounding tissue restoration experiments. Similar to fig:results:restoration:organ but not only considering the respective organ cells but also the cells at the boundary to adjacent tissue in detecting affected organs and corresponding interventions (c.f., the eye sockets colored in light-blue and light-green for the left- and right eyes, respectively). Notably, while resetting the entire affected organ and its surrounding tissue (A) shows similar performance to only resetting the organ (see fig:results:restoration:organ (A)), considerably fewer interventions are required in (B) to achieve the same effects as in fig:results:restoration:organ (B) when including the boundaries to reconfiguring wrongly expressed cells of the respective affected organs.
Figure 9.
Organ and surrounding tissue restoration experiments. Similar to fig:results:restoration:organ but not only considering the respective organ cells but also the cells at the boundary to adjacent tissue in detecting affected organs and corresponding interventions (c.f., the eye sockets colored in light-blue and light-green for the left- and right eyes, respectively). Notably, while resetting the entire affected organ and its surrounding tissue (A) shows similar performance to only resetting the organ (see fig:results:restoration:organ (A)), considerably fewer interventions are required in (B) to achieve the same effects as in fig:results:restoration:organ (B) when including the boundaries to reconfiguring wrongly expressed cells of the respective affected organs.
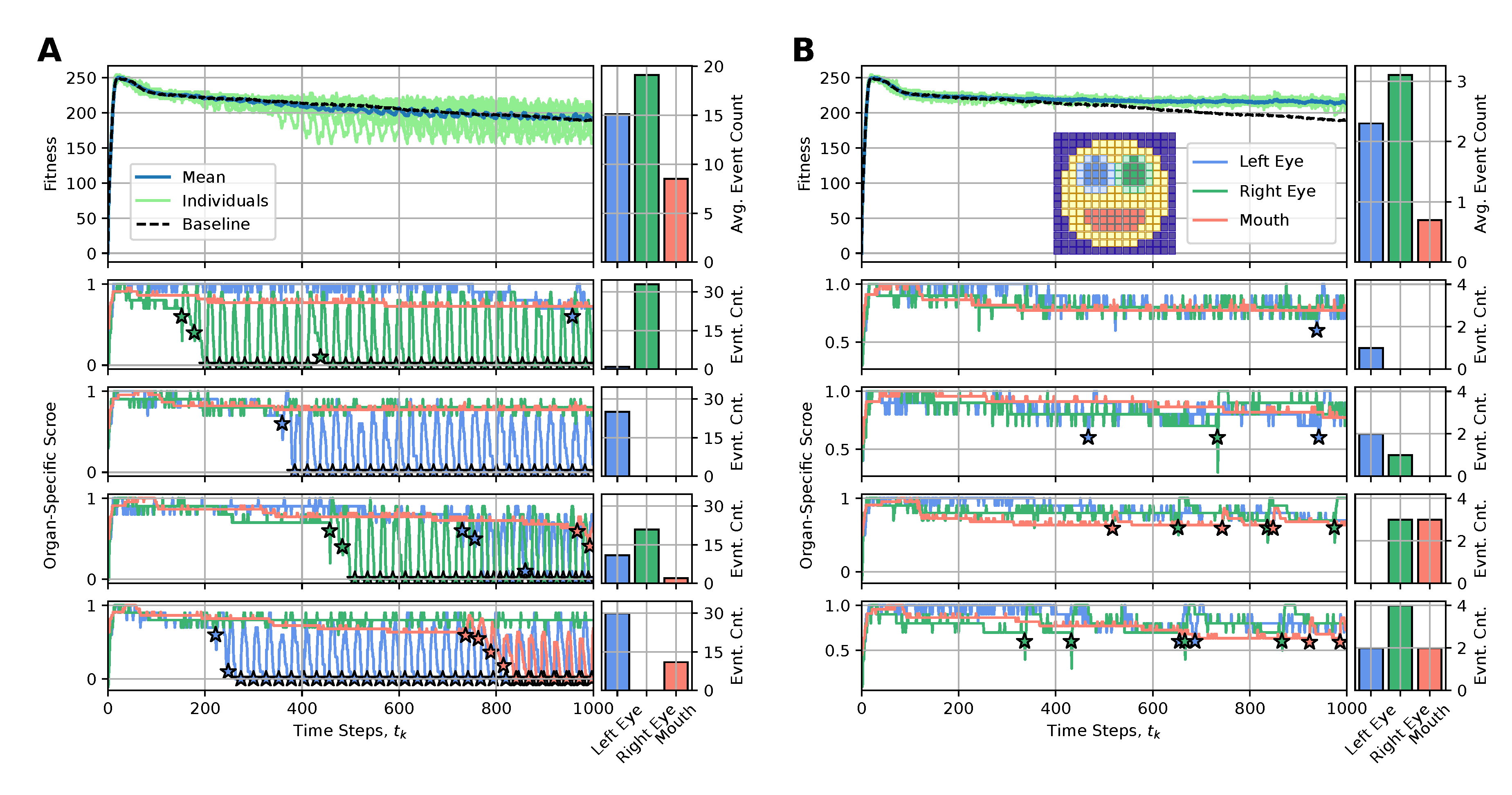
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



