
Submitted:
18 November 2024
Posted:
18 November 2024
You are already at the latest version
Abstract
Canine monocytic ehrlichiosis (CME) is a highly infectious disease with zoonotic potential. Ehr-lichia canis, the causative agent, is primarily transmitted by Rhipicephalus sanguineus. Tick infesta-tion and tick-borne diseases (TBD) are serious human and veterinary health problems due to lack of efficient control measures. This review aims to provide information about CME, detailing ep-idemiology, pathogenesis, clinical manifestations and current strategies for diagnosis, treatment and prevention. The review delves into biology of R. sanguineus, exploring lifecycle, habitat and mechanisms of E. canis transmission. Ehrlichia canis infection follows a three-phase transition: acute, subclinical and chronic, with distinct clinical signs, from mild to severe and even life threatening in some cases, with potential for asymptomatic long-term carriers acting as reservoirs. Pathogenesis involves complex interactions between the pathogen and host immune responses, influencing disease severity and progression. Additionally, the review highlights challenges in controlling CME due to the widespread distribution of R. sanguineus. Genetic diversity within R. sanguineus populations and their varying vector competence further complicate control efforts. Role of environmental changes in ticks’ distribution align control of CME within the OneHealth concept, integrating approaches to improve outcomes for humans, animals and environment. Preventive measures on tick control and potential vaccine development are also reviewed.
Keywords:
Canine Monocytic Ehrlichiosis
; Rhipicephalus sanguineus
; Ehrlichia canis
; Zoonosis
; Human ehr-lichiosis
; Rickettsia
; canine haemorrhagic fever
; feline ehrlichiosis
1. Introduction
Over the past decade, tick-borne diseases (TBD) caused by Ehrlichiae have garnered increasing attention from the veterinary, medical and biological fields [1].
The importance of ticks (Arachnida: Ixodida) as vectors of human and animal infectious diseases makes them second only to mosquitoes (Insecta: Diptera) in this regard, and tick-borne ehrlichial diseases are important emerging infections with a global prevalence [2,3]. Canine monocytic ehrlichiosis (CME) is a potentially fatal tick-borne infectious disease transmitted mainly by the brown dog tick Rhipicephalus sanguineus, also known to act as the vector of several other infectious diseases, underlining its efficiency at transmitting disease [4].
The etiological agent of CME is the obligate intracellular rickettsia Ehrlichia canis that invades and develops in canine monocytes, some cases leading to death [2]. Notably, E. canis presents a significant concern for canine health all over the world.
Human ehrlichiosis is a newly recognised disease and human infection with E. canis has been reported with increasing frequency since its discovery in 1987 [5,6,7,8,9]. This suggests that its zoonotic potential is either greatly underappreciated or steadily increasing. Such reports combined with changing environmental conditions and the increasing distribution of several tick species, firmly aligns the control of CME with the aim of the “One Health” concept that emphasises the importance of improving healthcare for humans, animals and the environment.
1.1. Canine Monocytic Ehrlichiosis
1.1.1. Background
CME is a tick-borne infectious disease of dogs, which has the potential to be fatal, but is not contagious. In the past, it has also been referred to as canine rickettsiosis or canine haemorrhagic fever [3,10]. The aetiological agent is the obligate intracellular rickettsia E. canis (family Anaplasmataceae, order Rickettsiales), a gram-negative bacterium [11]. In canine hosts E. canis invades and then replicates in monocytes and macrophages. Infection can range from asymptomatic infection to severe and even life-threatening disease, with animals presenting clinical signs such as depression, fever, leukopenia and thrombocytopaenia (among others). The brown dog tick, R. sanguineus is the principal vector responsible for transmission of the E. canis parasite [1,12,13]. The first recognised incidence of CME occurred in 1935, when veterinarians at the Pasteur Institute (Algeria) described a disease in dogs that caused acute fever and conspicuous anaemia [12]. Numerous dogs used by the United States of America (USA) military during the Vietnam War also succumbed to what was later identified as tropical canine pancytopaenia, another older name given to CME [14]. Reports of the disease also emanated from parts of Africa, the Middle East and the Orient some years later [10]. CME is now a disease prevalent worldwide, responsible for extensive morbidity and mortality in domestic and wild canid populations [13].
1.1.2. Hosts
Ehrlichia canis predominantly infects dogs and other members of the Canidae family. All breeds and ages are susceptible, although the severity and clinical signs may vary. From an outbreak in a Florida zoo, significant mortality amongst different species of wild canids, including wolves and wolf-dog crosses, as well as many dogs, occurred in response to E. canis infection and CME [15]. Coyotes, red foxes and grey foxes were shown to be susceptible to infection after inoculation with blood from infected dogs. Furthermore, R. sanguineus larvae were infected after feeding a blood meal on a grey fox [16,17]. More recently, E. canis was also detected in a small number of red foxes in Portugal [18]. Though less common than in dogs, E. canis can also infect cats, leading to feline ehrlichiosis, which was first reported through a natural infection in France by Charpentier and Groulade (1986), and it has subsequently been recognised worldwide with increasing frequency [19,20]. There are two recognised forms of feline ehrlichiosis, but only one is caused by E. canis, and infects mononuclear cells. Immune responses have also been demonstrated to E. canis, through seroconversion, in jackals, a raccoon and a puma, but not in capybara, deer or lemurs [21]. Though only a low percentage of raccoons were positive for antibodies reactive to E. canis in another study, suggesting they are unlikely to be important in transmission [22].
1.1.3. Ehrlichia canis Development in the Host
E. canis is a small (0.5 – to 1.5 µm), gram-negative, obligate intracellular bacterium. It has selective tropism for peripheral blood monocytes and macrophages in dogs and other mammalian hosts. It is also deposited in target organs, invading the bone marrow, liver, spleen, and lymph nodes [23,24,25]. The development of E. canis in the host follows a sequential three step transition of acute, subclinical, and chronic phases after the initial incubation period. When an infected tick is feeding on the host, mononuclear cells are attracted to the inflamed site where a tick is attached. Salivary secretions from the tick may support the development of E. canis infection in monocytes by modulating host immunity [13]. This is the acute phase of the disease and is characterised by E. canis bacteria infecting and multiplying in granular lymphocytes and monocytes. Bacteria reside inside these cells in membrane-bound cytoplasmic vacuoles or modified parasitophorous endosomes known as morulae [23,26,27] (Figure 1).
The host gets infected through a tick bite and, once in the perivascular area, E. canis adheres to the membrane of mononuclear cells. Through endocytosis (a) it enters the cell and forms pleomorphic elementary bodies (EB) in phagosomes for 2 or 3 days. Elementary bodies grow and divide by binary fission and exit the phagosome as initial bodies (IB), that in turn continue to multiply and mature into morulae (M). Bacteria exit the cells by exocytosis (b), after morulae rupture, and enter in the host circulation to be phagocytised by other granular lymphocytes and monocytes (Image created with BioRender.com).
1.1.4. Vectors
Ticks are arthropods (phylum Arthropoda) that have a significant impact on medical and veterinary fields, despite being generally underappreciated. They can cause direct damage to the species they feed off, as well as existing as vectors of numerous different pathogens. These include bacteria, protozoa, helminths and viruses [28,29]. Currently, all of the known Ehrlichia species are transmitted by different hard-bodied ticks within the Ixodidae family [21]. The principal vector of CME, in both experimental and biological settings, is the brown dog tick R. sanguineus [10,13,15]. It has also been described as the “kennel tick” and was first distinguished taxonomically as Ixodes sanguineus in 1806 by Latreille [1,4,12,13]. Prior to 1998, there were no reports of experimental transmission of E. canis by any ixodid ticks other than R. sanguineus. Eight years before this, an experiment was attempted to transmit the pathogen with the argasid tick Otobius megini, but it was not successful [30]. The transstadial transmission of E. canis was eventually demonstrated in the American dog tick Dermacentor variabilis [31].
During the past years, the taxonomic classification of R. sanguineus sensu stricto (s.s.) has been under ongoing debate and some genetic findings strongly suggest the existence of different identities under the same name [32]. According to multiple biological parameters, phylogenetic analysis and reproductive compatibility, at least two distinct populations were reported in the Americas and named as R. sanguineus sensu lato (s.l.) tropical lineage and R. sanguineus s.l. tropical temperate lineage [33,34,35,36,37,38,39,40,41,42]. Morphologically, the two lineages are very similar [38,43]. Some reports suggest that the absence of established temperate lineage ticks in tropical climates is because adults would become inactive i.e., in diapause, immediately after moulting throughout the year. Likewise, the absence of established tropical lineage ticks in temperate climates is because they would not start diapause to avoid the fatal consequences of adverse winters [41,44].
The vector competence for E. canis was investigated by Moraes-Filho et al. (2015) in larvae and nymphs exposed to infection by feeding on an infected host. After moulting into nymphs and adults, respectively, the E. canis polymerase chain reaction (PCR)-positive ticks were allowed to feed on a naïve dog. For this study four populations of ticks were used: São Paulo state, south-eastern Brazil (BSP), Rio Grande do Sul state, southern Brazil (BRS), Argentina (ARG), and Uruguay (URU). Only dogs infested with BSP adult ticks became clinically ill, presented E. canis seroconversion and were E. canis-PCR positive in the blood. Furthermore, only the population of unfed BSP nymphs and adults were positive for E. canis deoxyribonucleic acid (DNA) 30 days after moulting. Accordingly, the BSP population was shown to be a competent vector for this bacterium, contrary to BRS, ARG and URU tick populations [45]. Likewise, the presence of E. canis infection in the tropical lineage was reported in Argentina [46], tropical areas of Brazil [47,48], but not in ticks collected from dogs in temperate areas of Uruguay [49]. The specific reason why the tropical lineage is a competent vector, whereas the temperate lineage is not, is still unclear. A recent study suggested that Coxiella species, part of the tick prokaryotic microbiome composition, may be different in both lineages, influencing the interaction between the vector and the harboured pathogens [50]. Lastly, the comparison between both the temperate and tropical lineages infected and non-infected tissues has shown a different pattern of protein representation (Sanches et al. 2021).
In domestic dogs, R. sanguineus is a very common ectoparasite [4], resulting in the distribution of CME being directly proportional to the prevalence of the vector [51]. Riphicephalus sanguineus is the most widely distributed tick in the world, being especially prevalent in tropical and subtropical areas [3,24,52,53,54,55]. In regions with a moderate climate, R. sanguineus tick activity is increased from late spring to early autumn, however, they are continually active in tropical and subtropical regions [3]. As R. sanguineus are a monotropic tick, all life cycle stages favour dogs as the principal hosts regardless of geographical area, despite their ability to feed on other domestic or wild animals [3]. Ticks in immature life stages can sometimes be found on various mammals, such as rabbits, cats, rodents, wild canids and humans, as well as pigeons; however, adults prefer larger mammals [19,20,56].
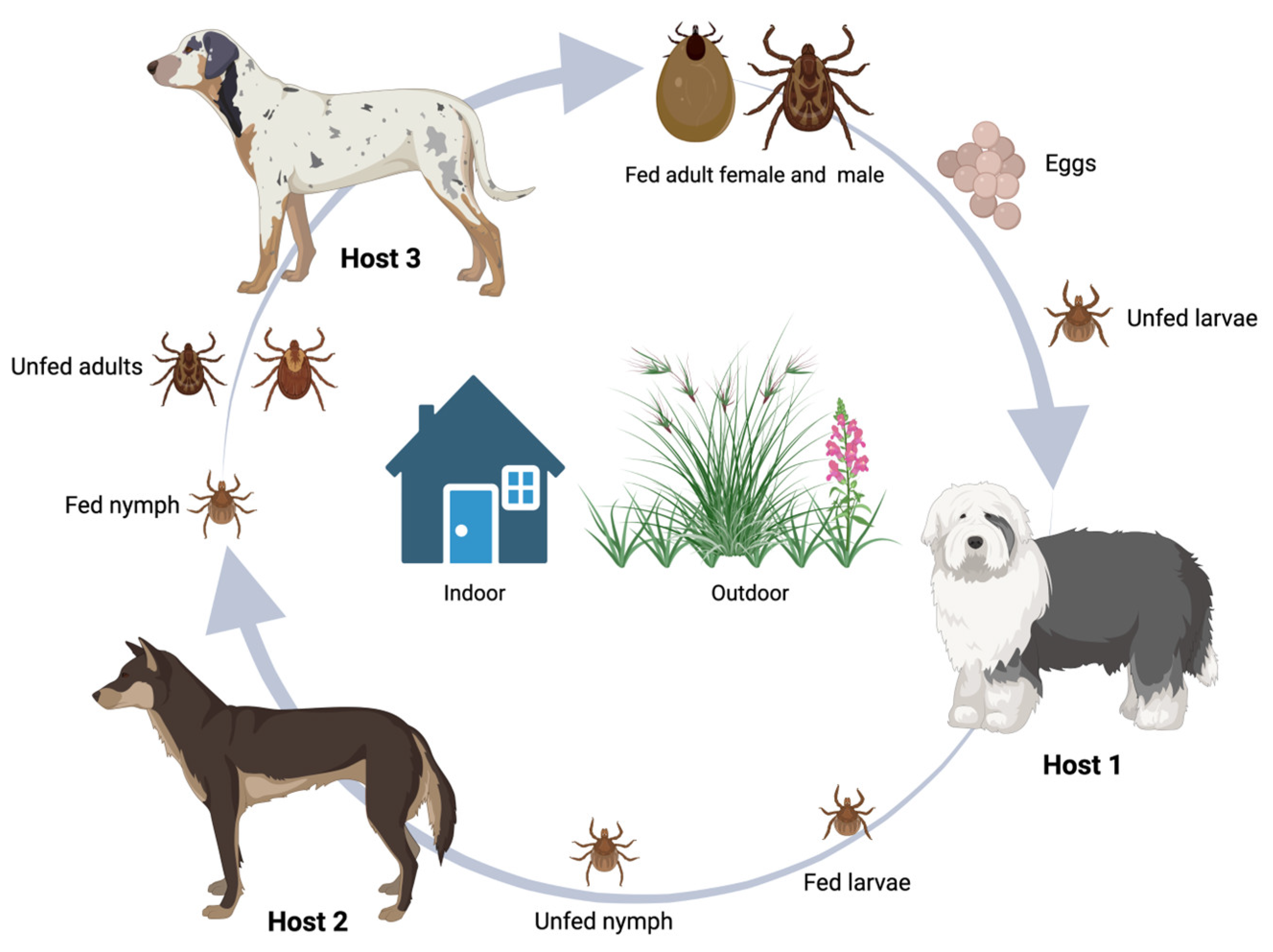
Morphological features of R. sanguineus ticks include a small, elongated body, free of any ornamentation, which is red-brown in colour. They have short palps, which are sensory structures that resemble a pair of legs, and a distinguishing feature known as the basis capituli, which is a flat hexagonal surface, where the mouthparts are attached [4]. All ixodid ticks, including R. sanguineus undergo four developmental stages: egg, larva, nymph and adult. To complete their development, a blood meal is required at each of the three stages after the egg [57]. A different host is required by R. sanguineus at each active developmental phase. After feeding on one host, the tick leaves that host to develop and moult and this happens twice: between larval and nymphal, and nymphal and adult stages [4,58]. R. sanguineus spend most of their life off-host, but they only mate when on a host [58,59]. When seeking a new host, they exhibit hunter behaviour, and, on occasion, can also outstretch their first pair of legs, which is a positioning known as questing [3]. After feeding on a host, which can last for 5 to 21 days, engorged adult females seek out sheltered places like cracks or crevices indoors to lay a large batch of eggs [4,58]. For each female, egg laying represents the end of their life - they lay around 4000 eggs and then die [4,60]. After a gestation period lasting 1 to 3 weeks, the larvae hatch and seek out a host. Larvae take a blood meal over the course of 3 to 10 days, then drop off the host and moult into nymphs. As nymphs, they follow the same cycle again for around 3 days to 2 weeks, before moulting into adults. Male and female adult ticks both feed off hosts. The entire life cycle lasts for just over two months when conditions are favourable. Contrary to many other tick species, R. sanguineus are endophilic, meaning they are frequently found indoors [4]. The life cycle is represented in Figure 2.
In addition to their role in E. canis transmission, R. sanguineus ticks act as vectors for other pathogens including the etiological agents of canine babesiosis, Babesia canis [61] and Babesia gibsoni [62]; the agent of Q fever Coxiella burnetii [63]; the agent of canine hepatozoonosis Hepatozoon canis [64], Rickettsia conorii, which causes Mediterranean spotted fever [65] and the bacterium that causes Rocky Mountain spotted fever, Rickettsia rickettsii [66]. Visceral leishmaniasis, caused by Leishmania infantum, may be yet another pathogen for which R. sanguineus ticks act as a vector [67], although data is not conclusive or proven.
After the blood meal, engorged females drop-off and lay eggs in the environment, normally in sheltered places with easy access to a vertebrate host. Unfed larvae hatch from these eggs, attach to a host to feed for about 4 days and drop-off to mould into nymphs. Nymphs will attach and feed on a dog for another 4 days, drop-off and moult into adults. Unfed adults will also attach and feed on a host, for about a week, mate, drop-off and lay eggs to restart the life cycle. In normal conditions, the entire cycle can be completed in 2 months (Image created with BioRender.com).
1.1.5. Ehrlichia canis Development in the Vector
There are important epidemiological implications associated with pathogen acquisition and transmission at the different tick developmental stages [1]. After feeding on dogs infected with E. canis, R. sanguineus ticks take up the bacterium and become infectious [21]. After multiplying within the digestive tract of engorged larvae, nymphs and adults, E. canis then infects the midgut (MG) epithelium cells, haemocytes and salivary glands (SG) cells [68]. Transstadial transmission, which occurs during “moulting” is where a pathogen needs to survive in the vector from one stage of the life cycle to the next. Transstadial transmission occurs so E. canis can transmit infection between two hosts. Ticks facilitate transstadial transmission between canine hosts through salivary secretions whilst ingesting a blood meal at the attachment site [13]. Male R. sanguineus ticks were shown to be capable of intrastadial i.e., within the same developmental stage, and transstadial acquisition and transmission of E. canis to susceptible dogs under experimental conditions. This was possible without the presence of female ticks [1]. Transovarial transmission, which is the vertical passage of parasites from an adult tick to its offspring by infecting developing eggs, has not been demonstrated with E. canis in R. sanguineus ticks [13].
1.1.6. Pathogenesis
CME pathogenesis begins once a host animal becomes infected with E. canis, either from an infected tick, transfusion of infected blood, or the transfer of infected leukocytes [31], with an incubation period of 8 to 20 days followed by three sequential phases: acute, subclinical, and chronic, though not all dogs advance to the latter [24].
At around day 10 after the initial infection, a collection of mild to severe and non-specific clinical signs characterize the acute phase. These include anorexia, fever, depression, dyspnoea, haemorrhages, lethargy, lymphadenopathy, splenomegaly and weight loss, which can progress to death [15,21,69,70,71]. Furthermore, anterior uveitis and retinal lesions are frequently observed in dogs with CME at each of the three phases [72]. Acute phase typically lasts 1 to 3 weeks [27], after which, E. canis infection perpetuates due to either inadequate treatment or spontaneous (clinical) resolution. Consequently, these animals can appear to be disease-free but may advance to the subclinical phase of CME [21,71,73]. Therefore, clinically healthy dogs in the subclinical phase harbour rickettsia for years without progressing to clinical disease. These dogs act as reservoirs for CME as ticks can continue to acquire and spread E. canis to other hosts [1,73].
It remains uncertain why some dogs progress to the chronic phase of CME [71]. This phase is characterised by a spectrum of symptoms, which may be mild or severe and include emaciation, epistaxis, haemorrhages, peripheral oedema, ocular problems, and hypotensive shock, which can result in death [21,74,75]. Dogs that only experience the milder form of chronic CME can carry an infection devoid of clinical signs for several years, in effect, making them the natural reservoir of E. canis [13].
The severity of CME can vary from an asymptomatic infection to a severe and even life-threatening disease due to several factors, including E. canis strain pathogenicity, host immunocompetence, canine breed, dose of infection and the impact of co-infections. Two known pathogens that can also be transmitted by R. sanguineus and co-infect dogs and influence CME severity are Babesia canis vogeli and H. canis [23,76]. Though all dogs can be infected, German shepherds seem to be more susceptible to E. canis infection. They often develop a chronic haemorrhagic syndrome, coupled with higher rates of morbidity and mortality [77,78]. Haematological alterations associated with the acute phase include anaemia and leukopenia, both mild, as well as thrombocytopaenia [79]. In the subclinical phase, which follows the acute phase, dogs typically lack clinical signs; haematological parameters are generally normal, though platelet (PLT) count can be low [80]. It has been suggested that E. canis organisms are located in the spleen during this phase [73]. The spleen is known to be the major reservoir of monocytes in mammals [81] and splenic macrophages are also well described, which would support this suggestion. In the chronic phase, pancytopaenia is highly prevalent, and other most common clinical signs of the disease in this stage are anorexia, bleeding diathesis, depression, lethargy, lymphadenomegaly, pyrexia, splenomegaly and weight loss [79].
Different aspects of the canine immune response to E. canis infection have been investigated to varying degrees. Numerous studies have been carried out in experimentally infected dogs, though far fewer exist in animals that were naturally infected. Comparing two groups of dogs with either clinical or subclinical infection, a study found the former to present lower numbers of eosinophils, red blood cells (RBC), packed cell volume (PCV), haemoglobin, and albumin, and a reduced albumin/globulins ratio [82]. Dogs with clinical infection also had higher γ-globulin levels. There were no statistically significant differences between the groups following analysis of total CD3+ T cells, CD4+ helper T cells, CD8+ cytotoxic T cells, B cells or major histocompatibility complex (MHC) II+ lymphocytes. Though CD21+ B cell numbers, both relative and absolute, were lower in dogs with clinical infection. This may be due to passage of cells from blood into specific organs [82].
Other studies have sought to directly compare experimentally and naturally infected dogs. For instance, one relatively recent study examined leukocyte abnormalities in both groups of dogs experiencing acute CME. During the first 14 days of experimental infection, a clear decrease in nearly all leukocyte populations, including segmented neutrophils, lymphocytes, monocytes and eosinophils, was observed. By contrast, a statistically significant increase in band neutrophils was apparent. By day 28 after infection, cell counts had returned to at least, and in some cases slightly higher than, pre-infection levels [83]. Neutropenia was the most striking leukocyte anomaly in this group, which may be due to impaired granulopoiesis, a more rapid egress of these cells from bone marrow or destruction caused by anti-neutrophil antibodies. Though neutrophil levels did increase again from day 28 after infection, making this less likely. Data in naturally infected dogs was far more variable with both neutropenia and neutrophilia found with similar frequency. Though this discrepancy may be due to the small and heterogenous population of naturally infected dogs that were sampled. Neutrophils are the first immune cell population to respond to infections and tissue damage [84]. Although several studies have reported neutropenia in dogs infected with CME, neutrophil function during the acute phase of the disease has been largely, though not completely, ignored. One study has quantified oxidative metabolism in neutrophils isolated from the peripheral blood of dogs during the first six weeks of infection, reporting that there were no significant differences in oxidative metabolism between cells from infected and uninfected dogs. However, 2 weeks post-infection, infected neutrophils that were re-stimulated with an inactivated bacterial extract, were much more reactive than those from uninfected dogs receiving the same treatment. This response was sustained until the end of the 6-week experiment. This is a particularly interesting observation and may be an example of trained immunity in these cells. This mechanism was not formally characterised until 2011 [85], six years after the CME study in question. More recently, it has been shown to occur in neutrophils [86].
The acute phase response, which forms part of the innate immune response to pathogens presence, involves the production of several acute phase proteins (APP). In response to stimulation by pro-inflammatory cytokines released by monocytes and macrophages, the liver synthesizes these proteins. In dogs, the assessment of innate inflammatory responses after vaccination against CME are not common. In one study, the production kinetics of a selection of APP was compared in unvaccinated and attenuated E. canis strain-vaccinated dogs that were subsequently challenged with a wild strain. In vaccinated dogs, levels of C-reactive protein (CRP), serum amyloid A (SAA), haptoglobin, albumin, paraoxonase-1 and total antioxidant capacity were significantly lower than in unvaccinated dogs. Post-challenge increases in these markers were also more delayed in the vaccinated dogs than those lacking protection. A strong positive correlation was apparent between rickettsial load and APP such as CRP and SAA, indicative of a direct impact by the pathogen on these inflammatory markers, as would be expected. These findings fit with those from an earlier study examining naturally infected dogs [87].
As previously discussed, E. canis has tropism for and invades canine monocytes and macrophages. Insights into the intercellular spreading process between macrophages were gained through use of a panel of drugs on the E. canis-infected canine monocyte-macrophage cell line, DH82 cells [88]. A reduced infection rate was observed in cytochalasin D-treated cells, identifying the actin cytoskeleton as crucial. This compound inhibits actin polymerisation, which is known to be important for pathogen spreading. Bacterial growth was also decreased in cells exposed to deferoxamine, which chelates iron. Earlier studies had revealed iron influx as crucial for the proliferation and spreading of other bacterial species. This study confirmed that this is also relevant for E. canis spreading. Finally, the most significant impact on bacterial spreading was observed when verapamil, a cell membrane and endoplasmic reticulum calcium channel blocker, was used. It completely inhibited the spread of E. canis to adjacent cells. Analysis of acid phosphatase labelling to observe phagosome-lysosome fusion indicated that E. canis also evades lysosomal fusion to survive and replicate [88]. This could be due to an active type IV secretion system (TFSS), which is known to inhibit the transport of bacteria to lysosomes and is conserved [89] and expressed [90] in E. canis. As is often the case with successful pathogens, E. canis can evade elimination by host cells through more than one strategy. Evidence of its capacity to modulate host immune responses was shown in DH82 cells. Using two different antibodies, MHC class II molecules were expressed on 46.9% of uninfected cells. By contrast, E. canis completely abrogated surface expression, identifying a mechanism used by the bacterium to subvert the host immune response [91].
To better understand the pathogenesis of canine ehrlichiosis as well as the host response to the pathogen, the E. canis genome was sequenced for the first time in 2006 [89]. Of particular interest was a conserved group of twelve cell wall proteins that contain tandem repeats. Prior studies had shown that this family is involved in host-pathogen interactions and pathogenicity. All of these twelve E. canis proteins have identifiable tandem repeat-containing orthologues in other Rickettsiales genomes. Another group of genes engaged in host-pathogen interactions and pathogenicity are the Vir proteins. The E. canis genome build shows they are arranged in two clusters: virB8/virB9/ virB10/virB11/virD4, and virB3/virB4/ virB6 plus three virB6-related genes. An earlier work revealed virB/D operons drive expression of the TFSS, which is used to deliver virulence factors from bacteria to host cell cytoplasm [92]. Of interest, the E. canis VirB9 is expressed in canine and tick hosts, with modelling identifying several antigenic epitopes [90]. Added to the fact that virB/D operons needed to express TFSS components were identified and functionally characterised in E. chaffeensis [93]; it is highly likely E. canis relies on its orthologous proteins for intracellular survival and replication. Further evidence of the ability of E. canis to evade the host immune response can be inferred from absent cell wall components. The genes required for the synthesis of both lipopolysaccharide and peptidoglycan are lacking in the E. canis genome (as well as other Ehrlichia). This makes its recognition by Toll-like receptors 2 and 4 highly unlikely, limiting the innate immune response [89].
The ongoing “success” of E. canis to invade and persist in hosts can be partially attributed to our limited understanding of its functional genes and proteins. In fact, it has been suggested the lack of a vaccine is hindered by its arsenal of protective and immunoreactive proteins that alter antibody responses in infected dogs [94]. The known repertoire of these proteins is also believed to be a fraction of the true total, with many undefined. With that in mind, two recent studies have offered a significant advance in this area through discovery of several novel immunoreactive proteins that may assist future vaccine development [94,95]. In both studies, proteins with conformation-dependent antibody epitopes were identified using novel approaches. Previous efforts had revealed a small selection of proteins with only linear antibody epitopes, since experimental methods were limited. Notably, sixteen out of the thirty-four proteins identified in the two studies were predicted to be secreted as well as immunoreactive [94,95]. As they are probably targeted by the host immune response to neutralize their function, they may be responsible for subverting canonical host processes to promote infection.
1.1.7. Diagnosis
The different phases and multiple clinical manifestations of CME can make its diagnosis challenging [71]. Typically, a combination of anamnesis i.e. residing in or travelling to an endemic region as well as previous exposure to ticks, characteristic clinical signs, as described previously, plus haematological and biochemical alterations leads to a diagnosis [71]. To definitively confirm this, laboratory tests comprising of one or more of microscopy, serology, cell culture or PCR [71,96] are carried out to directly or indirectly detect E. canis. Intracytoplasmic morula-like bodies, either single or multiple, observed in monocytes by microscopy during the acute phase, is conclusive [69,71]. Despite this, it is a method lacking sensitivity, often reporting false negatives and false positives with other ehrlichiae of the Anaplasmatacea family. For example, canine monocytes may be infected with E. chaffeensis, Ehrlichia ruminantium or Ehrlichia risticii [97,98,99]. This method is also less than ideal due to its time-consuming workflow, and low numbers of organisms make diagnosis difficult [21,71,96]. Just 4% of blood smears from dogs with ehrlichiosis are sufficiently unambiguous to observe the presence of E. canis morulae [100].
There are now improved serological methods available to accurately screen and/or diagnose CME [71]. One example is the use of indirect fluorescent antibody tests (IFAT) for anti-E. canis immunoglobulin (Ig) G (IgG), which were long considered to be the “gold standard” [71,101,102] . Enzyme-linked immunosorbent assays (ELISA) are also used to diagnose E. canis infection [101,102,103,104]. In the past, a problematic aspect of serological assays was the potential for cross-reactivity with other ehrlichial organisms. These were reported in endemic areas with other Ehrlichia species, for instance: Ehrlichia ewingii, E. chaffeensis and E. risticii [71,102,105]. Ehrlichia canis antibodies may also cross react with Anaplasma phagocytophilum antigens [102]. Improvements in the sensitivity and specificity of ELISA have made them a more robust method for diagnosing E. canis infection. This is, in part, due to a deeper molecular characterization of E. canis proteins. For example, two conserved immunoreactive antigens, the glycoproteins gp19 and gp36, were identified as able to induce specific antibody responses to E. canis [106,107]. These antigens, as well as the glycoprotein gp200 were expressed as recombinant glycoproteins, and used to develop an ELISA that demonstrated 100% sensitivity and specificity compared to IFAT. This ELISA can also detect antibodies two weeks earlier than IFAT in samples from experimentally infected dogs. Furthermore, it is species-specific, without cross reactivity with antibodies in sera from dogs infected with E. chaffeensis [108]. If strain-specific detection is needed, there are genotype-specific ELISA now available that offer this degree of specificity. Taking advantage of a tandem repeat motif within the TRP36, an ELISA that can distinguish between Brazilian and American E. canis infections derived from these alternative genotypes, was developed [109]. In addition to serological assays, molecular detection methods also provide a definitive diagnosis.
The PCR coupled with amplicon sequencing can sensitively detect and potentially reveal the genotype of E. canis DNA [71,110]. PCR assays targeting a diverse group of target genes, for e.g., 16S ribosomal RNA (16S rRNA), outer membrane multigenes 28 and 30 (p28; p30), disulphide bond formation protein (dsb) and outer membrane protein 1 (omp-1), have been developed. The most frequently performed amplify 16S rRNA and p30 [71,111,112].
By combining IFAT tests with nested-PCR, a laboratory diagnosis could be made during the acute phase of infection, prior to detectable antibody levels in sera, and also in the subclinical phase [113,114]. By combining PCR with DNA hybridisation, E. canis infection can be detected with greater sensitivity than conventional PCR [115,116]. A recent study has highlighted the challenge associated with diagnosing the disease in dogs in the subclinical and chronic phases of CME. In the former, there is a lack of clinical signs, whilst in both disease states, low parasitaemia can be observed because bacteria reside in target organs. Consequently, these dogs are negative for E. canis in the blood when PCR is used to screen. A group of fifty-nine dogs that lacked clinical signs but had ticks were euthanised, after which blood, bone marrow, liver, lymph node and spleen tissue samples were screened for E. canis by PCR. A negative blood PCR was confirmed in 52.5% of dogs, however, 61.3% were positive in tissue samples [117]. In line with other studies these last authors cited, there might be a case for combining detection in blood with splenic aspirates as an improved diagnostic approach and/or determining CME distribution.
Quantitative real-time PCR (qPCR) is more sensitive than conventional PCR, and it rapidly became the principal method of E. canis diagnosis. Some of the earliest examples of this assay being developed immediately underlined its usefulness. Based on amplification of 16S rRNA, a qPCR TaqMan assay was created to discriminate single infections in canine blood with Ehrlichia chaffeensis, E. canis, E. ewingii, A. phagocytophilum and Anaplasma platys. It could also determine co-infection with E. canis and A. platys [118]. This assay was later shown to be effective at detecting some Ehrlichia and Anaplasma in Amblyomma americanum, Dermacentor and Ixodes species, though not E. canis [119]. The first tricolour TaqMan assay to detect and discriminate between different Ehrlichia species in a single reaction was based on amplification of dsb. This genus-specific target allowed singleplex quantification of E. chaffeensis, E. ewingii or E. canis from naturally or experimentally infected dogs without cross hybridisation of the probe [120]. The primers also would not amplify A. platys, A. phagocytophilum, R. conorii or Rickettsia typhi, emphasising its specificity. An alternative variant to these assays has been subsequently developed to amplify the 16S rRNA in E. canis and the heat shock protein (HSP) 70 (hsp70) in B. canis vogeli in a multiplex assay with dual labelled probes [121]. Another multiplex qPCR assay using SYBR green is also available to amplify B. gibsoni, B. vogeli, E. canis and H. canis. The virB9 is the target in E. canis [122].
1.1.8. Treatment, Control and Prevention
The first line treatment for CME consists in administrations of doxycycline, a broad-spectrum tetracycline-class antibiotic, since it exhibits greater intracellular penetration and inhibits bacterial protein synthesis to prevent growth [123,124]. Tetracycline hydrochloride, oxytetracycline, minocycline and chloramphenicol have also been shown to target E. canis with variable level of efficacy [100]. As reviewed by Mylonakis et al. (2019), CME treatment with antimicrobials should be able to cause clinical remission and resolution of clinical and pathological alterations [125]. Timely diagnosis and administration of antibiotics, primarily doxycycline, have been effective in treating acute cases. However, chronic cases can present challenges due to the persistence of the pathogen. Complete parasite elimination is not always possible or proved by diagnostic means [113].
A commercial vaccine for CME has yet to be developed; therefore, measures to control ticks remain the best preventative approach to restrict the transmission of E. canis. Strategies for tick control can be focussed on the on-host phase of the tick life cycle, achieved through targeting dogs as well as indoor spaces. The phase of the tick life cycle when it is off host can also be targeted, relying on knowledge of tick ecology. Often, a combination of both approaches is adopted. Naturally, controlling ticks outdoors is very challenging and since R. sanguineus ticks prefer indoor spaces, limiting infestations in these areas as well as on the host should be the primary focus of control measures. A combination of chemical and non-chemical approaches should be pursued [4]. Chemical control on the host can be established using liquid formulations, sprays, repellents and impregnated collars, containing acaricides and/or insecticides such as fipronil, amitraz or permethrin. A comparison study using novel chewable compounds, afoxolaner (NexGard™) and fluralaner (Bravecto™) has found an impaired efficacy against R. sanguineus infestation with low efficacy times allowing E. canis transmission. Contrary, Advantix® (50 % permethrin:10 % imidacloprid) effectively blocked transmission of E. canis to dogs, providing adequate protection for dogs against CME. Commercially available drugs such as pyrethroids, phenylpyrazoles, isoxazolines, and amitraz show good effectiveness regarding the reduction of ticks engaged in disease spread. However, owners must be educated about the development of resistance against such drugs and should be encouraged to alternatively use of different acaricides [126].
Altering tick habitats’ through sealing cracks and crevices, and/or removing grass and weeds represent outdoor controls that do not require chemicals It is essential that canine blood donors be screened to confirm seronegativity for anti-tick antibodies to avoid E. canis transmission from a blood transfusion [4].
An integrated approach combining environmental changes, host management, chemical interventions, and veterinary care can reduce tick infestations and therefore E. canis transmission.
2. Conclusions
The persistent challenge of CME underscores the critical need for advancing our understanding of E. canis and its primary vector, R. sanguineus. This review highlights the significant medical and veterinary impact of CME, emphasizing the disease's zoonotic potential and the complexities involved in its control and prevention. Despite advancements in diagnostic and therapeutic approaches, the global distribution and adaptability of R. sanguineus, coupled with the multifaceted interactions between pathogen, vector and host continue to hinder effective management strategies.
Addressing CME requires a multifaceted approach integrating advancements in molecular diagnostics, targeted therapeutic regimens and robust preventive measures, including vector control and potential vaccine development, as pointed by Aziz et al. (2023). The One Health framework, which considers the interconnectedness of human, animal and environmental health, is paramount in developing sustainable solutions to mitigate the impacts of CME.
Future research should focus on exploring the genetic diversity of R. sanguineus, understanding the ecological factors influencing tick distribution, and developing novel intervention strategies. Collaborative efforts between veterinary and medical professionals, researchers and policymakers are essential to enhance our ability to manage CME effectively and protect both canine and human health.
By consolidating current knowledge and identifying gaps, this review aims to inform and inspire further research and policy initiatives, ultimately contributing to the global effort to control TBD and improve health outcomes.
Author Contributions
All authors have read and agreed to the published version of the manuscript.
Funding
This review was funded by the Portuguese Foundation for Science and Technology (FCT), projects UIDB/00772/2020 (Doi:10.54499/UIDB/00772/2020).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bremer, W.G.; Schaefer, J.J.; Wagner, E.R.; Ewing, S.A.; Rikihisa, Y.; Needham, G.R.; Jittapalapong, S.; Moore, D.L.; Stich, R.W. Transstadial and intrastadial experimental transmission of Ehrlichia canis by male Rhipicephalus sanguineus. Vet. Parasitol. 2005, 131, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Cao, W.; Pan, H. Ehrlichiae and Ehrlichial Diseases in China. Ann. New York Acad. Sci. 2003, 990, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasites Vectors 2010, 3, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. The brown dog tick, Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae): From taxonomy to control. Vet. Parasitol. 2008, 152, 173–185. [Google Scholar] [CrossRef]
- Maeda, K.; Markowitz, N.; Hawley, R.C.; Ristic, M.; Cox, D.; McDade, J.E. Human Infection withEhrlichia canis, a Leukocytic Rickettsia. N. Engl. J. Med. 1987, 316, 853–856. [Google Scholar] [CrossRef]
- J. E. Dawson et al., "Isolation and characterization of an Ehrlichia sp. from a patient diagnosed with human ehrlichiosis," J Clin Microbiol, vol. 29, no. 12, pp. 2741-5, Dec 1991.
- Brouqui, P.; Le Cam, C.; Kelly, P.J.; Laurens, R.; Tounkara, A.; Sawadogo, S.; Lo-Marcel, V.; Gondao, L.; Faugere, B.; Delmont, J.; et al. Serologic evidence for human ehrlichiosis in Africa. Eur. J. Epidemiology 1994, 10, 695–698. [Google Scholar] [CrossRef]
- Perez, M.; Rikihisa, Y.; Wen, B. Ehrlichia canis-like agent isolated from a man in Venezuela: antigenic and genetic characterization. J. Clin. Microbiol. 1996, 34, 2133–9. [Google Scholar] [CrossRef]
- Perez, M.; Bodor, M.; Zhang, C.; Xiong, Q.; Rikihisa, Y. Human Infection with Ehrlichia Canis Accompanied by Clinical Signs in Venezuela. Ann. N. Y. Acad. Sci. 2006, 1078, 110–117. [Google Scholar] [CrossRef]
- S. A. Ewing, "Canine ehrlichiosis," Adv Vet Sci Comp Med, vol. 13, pp. 331-53, 1969.
- J. S. Dumler et al., "Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and 'HGE agent' as subjective synonyms of Ehrlichia phagocytophila," Int J Syst Evol Microbiol, vol. 51, no. Pt 6, pp. 2145-65, Nov 2001.
- Donatien and, F. Lestoquard, "Existence in Algerie d'une Rickettsia du chien.," Bull. Soc. Pathol. Exot., vol. 28, pp. 418-419, 1935.
- M. G. Groves, G. L. Dennis, H. L. Amyx, and D. L. Huxsoll, "Transmission of Ehrlichia canis to dogs by ticks (Rhipicephalus sanguineus)," Am J Vet Res, vol. 36, no. 7, pp. 937-40, Jul 1975.
- Walker, J.S.; Rundquist, J.D.; Taylor, R.; Wilson, B.L.; Andrews, M.R.; Barck, J.; Hogge, A.L.; Huxsoll, D.L.; Hildebrandt, P.K.; Nims, R.M. Clinical and clinicopathologic findings in tropical canine pancytopenia. J Am Vet Med Assoc 1970, 157, 43–55. [Google Scholar]
- J. W. Harvey, C. F. Simpson, J. M. Gaskin, and J. H. Sameck, "Ehrlichiosis in wolves, dogs, and wolf-dog crosses," J Am Vet Med Assoc, vol. 175, no. 9, pp. 901-5, Nov 1 1979.
- Ewing, S.A.; Buckner, R.G.; Stringer, B.G. The Coyote, a Potential Host for Babesia canis and Ehrlichia sp. J. Parasitol. 1964, 50, 704. [Google Scholar] [CrossRef]
- Amyx, H.L.; Huxsoll, D.L. RED AND GRAY FOXES — POTENTIAL RESERVOIR HOSTS FOR Ehrlichia canis. J. Wildl. Dis. 1973, 9, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.; Gilad, M.; Cortes, H.C.; Nachum-Biala, Y.; Lopes, A.P.; Vila-Viçosa, M.J.; Simões, M.; A Rodrigues, P.; Baneth, G. First report of Anaplasma platys infection in red foxes (Vulpes vulpes) and molecular detection of Ehrlichia canis and Leishmania infantum in foxes from Portugal. Parasites Vectors 2015, 8, 144–144. [Google Scholar] [CrossRef] [PubMed]
- Bouloy, R.P.; Lappin, M.R.; Holland, C.H.; Thrall, M.A.; Baker, D.; O'Neil, S. Clinical ehrlichiosis in a cat. J. Am. Veter- Med Assoc. 1994, 204, 1475–1478. [Google Scholar] [CrossRef]
- Breitschwerdt, E.B.; Abrams-Ogg, A.C.; Lappin, M.R.; Bienzle, D.; Hancock, S.I.; Cowan, S.M.; Clooten, J.K.; Hegarty, B.C.; Hawkins, E.C. Molecular Evidence Supporting Ehrlichia canis–Like Infection in Cats. J. Veter- Intern. Med. 2002, 16, 642–9. [Google Scholar] [CrossRef]
- Stich, R.; Schaefer, J.J.; Bremer, W.G.; Needham, G.R.; Jittapalapong, S. Host surveys, ixodid tick biology and transmission scenarios as related to the tick-borne pathogen, Ehrlichia canis. Veter- Parasitol. 2008, 158, 256–273. [Google Scholar] [CrossRef]
- Yabsley, M.J.; Murphy, S.M.; Luttrell, M.P.; Little, S.E.; Massung, R.F.; Stallknecht, D.E.; Conti, L.A.; Blackmore, C.G.; Durden, L.A. Experimental and Field Studies on the Suitability of Raccoons (Procyon lotor) as Hosts for Tick-Borne Pathogens. Vector-Borne Zoonotic Dis. 2008, 8, 491–504. [Google Scholar] [CrossRef]
- Y. Rikihisa, "The tribe Ehrlichieae and ehrlichial diseases," Clin Microbiol Rev, vol. 4, no. 3, pp. 286-308, Jul 1991.
- Z. Woldehiwet and M. Ristic, Rickettsial and chlamydial diseases of domestic animals, 1st ed. Oxford, England ; New York: Pergamon Press, 1993, pp. xiii, 427 p.
- M. Ristic and C. J. Holland, "Canine ehrlichiosis," Pergamon Press - Oxford, 1993.
- Popov, V.L.; Han, V.C.; Chen, S.-M.; Dumler, J.S.; Feng, H.-M.; Andreadis, T.G.; Tesh, R.B.; Walker, D.H. Ultrastructural differentiation of the genogroups in the genus Ehrlichia. J. Med Microbiol. 1998, 47, 235–251. [Google Scholar] [CrossRef]
- McQuiston, J.H.; McCall, C.L.; Nicholson, W.L. Ehrlichiosis and related infections. J. Am. Veter- Med Assoc. 2003, 223, 1750–1756. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Zweygarth, E.; Blouin, E.F.; Gould, E.A.; Jongejan, F. Tick cell lines: tools for tick and tick-borne disease research. Trends Parasitol. 2007, 23, 450–457. [Google Scholar] [CrossRef]
- Otranto, D.; Dantas-Torres, F.; Brianti, E.; Traversa, D.; Petrić, D.; Genchi, C.; Capelli, G. Vector-borne helminths of dogs and humans in Europe. Parasites Vectors 2013, 6, 1–14. [Google Scholar] [CrossRef]
- Ewing, S.A.; Harkess, J.R.; Kocan, K.M.; Barker, R.W.; Fox, J.C.; Tyler, R.D.; Cowell, R.L.; Morton, R.B. Failure to Transmit Ehrlichia canis (Rickettsiales: Ehrlichieae) with Otobius megnini (Acari: Argasidae). J. Med Èntomol. 1990, 27, 803–806. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.; Ewing, S.; Barker, R.; Fox, J.; Crow, D.; Kocan, K. Experimental transmission of Ehrlichia canis (Rickettsiales: Ehrlichieae) by Dermacentor variabilis (Acari: Ixodidae). Veter- Parasitol. 1998, 74, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Nava, S.; Estrada-Peña, A.; Petney, T.; Beati, L.; Labruna, M.B.; Szabó, M.P.; Venzal, J.M.; Mastropaolo, M.; Mangold, A.J.; Guglielmone, A.A. The taxonomic status of Rhipicephalus sanguineus (Latreille, 1806). Veter- Parasitol. 2015, 208, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Szabó, M.P.; Mangold, A.J.; João, C.F.; Bechara, G.H.; Guglielmone, A.A. Biological and DNA evidence of two dissimilar populations of the Rhipicephalus sanguineus tick group (Acari: Ixodidae) in South America. Veter- Parasitol. 2005, 130, 131–140. [Google Scholar] [CrossRef]
- Burlini, L.; Teixeira, K.R.S.; Szabó, M.P.J.; Famadas, K.M. Molecular dissimilarities of Rhipicephalus sanguineus (Acari: Ixodidae) in Brazil and its relation with samples throughout the world: is there a geographical pattern? Exp. Appl. Acarol. 2009, 50, 361–374. [Google Scholar] [CrossRef]
- Moraes-Filho, J.; Marcili, A.; Nieri-Bastos, F.A.; Richtzenhain, L.J.; Labruna, M.B. Genetic analysis of ticks belonging to the Rhipicephalus sanguineus group in Latin America. Acta Trop. 2011, 117, 51–55. [Google Scholar] [CrossRef]
- Levin, M.L.; Studer, E.; Killmaster, L.; Zemtsova, G.; Mumcuoglu, K.Y. Crossbreeding between different geographical populations of the brown dog tick, Rhipicephalus sanguineus (Acari: Ixodidae). Exp. Appl. Acarol. 2012, 58, 51–68. [Google Scholar] [CrossRef]
- Nava, S.; Mastropaolo, M.; Venzal, J.M.; Mangold, A.J.; Guglielmone, A.A. Mitochondrial DNA analysis of Rhipicephalus sanguineus sensu lato (Acari: Ixodidae) in the Southern Cone of South America. Veter- Parasitol. 2012, 190, 547–555. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Latrofa, M.S.; Annoscia, G.; Giannelli, A.; Parisi, A.; Otranto, D. Morphological and genetic diversity of Rhipicephalus sanguineus sensu lato from the New and Old Worlds. Parasites Vectors 2013, 6, 213–213. [Google Scholar] [CrossRef]
- Liu, G.-H.; Chen, F.; Chen, Y.-Z.; Song, H.-Q.; Lin, R.-Q.; Zhou, D.-H.; Zhu, X.-Q. Complete Mitochondrial Genome Sequence Data Provides Genetic Evidence That the Brown Dog Tick Rhipicephalus sanguineus (Acari: Ixodidae) Represents a Species Complex. Int. J. Biol. Sci. 2013, 9, 361–369. [Google Scholar] [CrossRef]
- Sanches, G.S.; Évora, P.M.; Mangold, A.J.; Jittapalapong, S.; Rodriguez-Mallon, A.; Guzmán, P.E.; Bechara, G.H.; Camargo-Mathias, M.I. Molecular, biological, and morphometric comparisons between different geographical populations of Rhipicephalus sanguineus sensu lato (Acari: Ixodidae). Veter- Parasitol. 2016, 215, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.O.; Gruntmeir, J.M.; Hamer, S.A.; Little, S.E. Temperate and tropical lineages of brown dog ticks in North America. Veter- Parasitol. Reg. Stud. Rep. 2017, 7, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Latrofa, M.S.; Ramos, R.A.N.; Lia, R.P.; Capelli, G.; Parisi, A.; Porretta, D.; Urbanelli, S.; Otranto, D. Biological compatibility between two temperate lineages of brown dog ticks, Rhipicephalus sanguineus (sensu lato). Parasites Vectors 2018, 11, 398. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, P.R.; Bechara, G.H.; Denardi, S.E.; Saito, K.C.; Nunes, E.T.; Szabó, M.P.J.; Mathias, M.I.C. Comparison of the external morphology of Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae) ticks from Brazil and Argentina. Veter- Parasitol. 2005, 129, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Labruna, M.B.; Gerardi, M.; Krawczak, F.S.; Moraes-Filho, J. Comparative biology of the tropical and temperate species of Rhipicephalus sanguineus sensu lato (Acari: Ixodidae) under different laboratory conditions. Ticks Tick-borne Dis. 2017, 8, 146–156. [Google Scholar] [CrossRef]
- Moraes-Filho, J.; Krawczak, F.S.; Costa, F.B.; Soares, J.F.; Labruna, M.B. Comparative Evaluation of the Vector Competence of Four South American Populations of the Rhipicephalus sanguineus Group for the Bacterium Ehrlichia canis, the Agent of Canine Monocytic Ehrlichiosis. PLOS ONE 2015, 10, e0139386. [Google Scholar] [CrossRef]
- Cicuttin, G.L.; Tarragona, E.L.; De Salvo, M.N.; Mangold, A.J.; Nava, S. Infection with Ehrlichia canis and Anaplasma platys (Rickettsiales: Anaplasmataceae) in two lineages of Rhipicephalus sanguineus sensu lato (Acari: Ixodidae) from Argentina. Ticks Tick-borne Dis. 2015, 6, 724–729. [Google Scholar] [CrossRef]
- D. M. Aguiar, G. T. Cavalcante, A. Pinter, S. M. Gennari, L. M. Camargo, and M. B. Labruna, "Prevalence of Ehrlichia canis (Rickettsiales: Anaplasmataceae) in dogs and Rhipicephalus sanguineus (Acari: Ixodidae) ticks from Brazil," J Med Entomol, vol. 44, no. 1, pp. 126-32, Jan 2007. [CrossRef]
- Souza, B.M.P.d.S.; Leal, D.C.; Barboza, D.C.P.M.; Uzêda, R.S.; De Alcântara, A.C.; Ferreira, F.; Labruna, M.B.; Gondim, L.F.P.; Franke, C.R. Prevalence of ehrlichial infection among dogs and ticks in Northeastern Brazil. Rev. Bras. De Parasitol. Veter- 2010, 19, 89–93. [Google Scholar] [CrossRef]
- J. M. Venzal, A. Estrada-Peña, O. Castro, C. G. De Souza, A. Portillo, and O. J.A., "Study on seasonal activity in dogs and ehrlichial infection in Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae) from southern Uruguay," Parasitol. Latinoam., vol. 62, pp. 23-26, 2007.
- Luzzi, M.d.C.; de Carvalho, L.A.L.; Pinheiro, D.G.; Lima-Duarte, L.; Camargo, J.V.; Kishi, L.T.; Fernandes, C.C.; Machado, R.Z.; Soares, J.F.; André, M.R.; et al. Analysis on the prokaryotic microbiome in females and embryonic cell cultures of Rhipicephalus sanguineus tropical and temperate lineages from two specific localities in Brazil. Rev. Bras. De Parasitol. Veter- 2021, 30, e005721. [Google Scholar] [CrossRef]
- T. M. Neer, E. B. Breitschwerdt, R. T. Greene, and M. R. Lappin, "Consensus statement on ehrlichial disease of small animals from the infectious disease study group of the ACVIM. American College of Veterinary Internal Medicine," J Vet Intern Med, vol. 16, no. 3, pp. 309-15, May-Jun 2002.
- Keefe, T.J.; Holland, C.J.; E Salyer, P.; Ristic, M. Distribution of Ehrlichia canis among military working dogs in the world and selected civilian dogs in the United States. . 1982, 181, 236–8. [Google Scholar]
- Estrada-Peña, A.; Jongejan, F. Ticks Feeding on Humans: A Review of Records on Human-Biting Ixodoidea with Special Reference to Pathogen Transmission. Exp. Appl. Acarol. 1999, 23, 685–715. [Google Scholar] [CrossRef] [PubMed]
- Hua, P.; Yuhai, M.; Shide, T.; Yang, S.; Bohai, W.; Xiangrui, C. Canine Ehrlichiosis Caused Simultaneously by Ehrlichia canis and Ehrlichia platys. Microbiol. Immunol. 2000, 44, 737–739. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, E.; Sainz, A.; Dunner, S.; Amusategui, I.; López, L.; Rodríguez-Franco, F.; Luaces, I.; Cortés, O.; Tesouro, M.A. First isolation and molecular characterization of Ehrlichia canis in Spain. Veter- Parasitol. 2004, 125, 365–372. [Google Scholar] [CrossRef]
- Maxwell, I. C. O. C. G. O. "Environmental and multi-host infestation of the brown dog tick, Rhipicephalus sanguineus in Owerri, South-east Nigeria- a case report," Veterinarski arhiv, vol. 76, no. 1, 2006.
- D. E. Sonenshine, Biology of ticks. New York: Oxford University Press, 1991.
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Otranto, D. Rhipicephalus sanguineus on dogs: relationships between attachment sites and tick developmental stages. Exp. Appl. Acarol. 2010, 53, 389–397. [Google Scholar] [CrossRef]
- H. G. Koch, "Oviposition of the Brown Dog Tick (Acari, Ixodidae) in the Laboratory," (in English), Annals of the Entomological Society of America, vol. 75, no. 5, pp. 583-586, 1982.
- Regendanz, P.; Muniz, J. O Rhipicephalus sanguineus como transmissor da Piroplasmose canina no Brasil. Mem. Do Inst. Oswaldo Cruz 1936, 31, 81–84. [Google Scholar] [CrossRef]
- S. K. Sen, "The vector of canine piroplasmosis due to Piroplasma gibsoni," Ind. J. Vet. Sci. Anim. Husband, vol. 3, pp. 356-363, 1933.
- Mantovani, A.; Benazzi, P. THE ISOLATION OF COXIELLA BURNETII FROM RHIPICEPHALUS-SANGUINEUS ON NATURALLY INFECTED DOGS. 1953, 122, 117–118.
- Nordgren, R.; Craig, T. Experimental transmission of the texas strain of Hepatozoon canis. Veter- Parasitol. 1984, 16, 207–214. [Google Scholar] [CrossRef]
- E. Brumpt, "Longevite du virus de la fievre boutonneuse chez la tique, Rhipicephalus sanguineus.," Compt Rend Soc Biol, vol. 8, pp. 1199-1202, 1932.
- R. R. Parker, C. B. Philip, and W. L. Jellison, "Rocky Mountain spotted fever: potentialities of tick transmission in relation to geographical occurence in the United States," Am. J. Trop. Med. Hyg., vol. 13, pp. 341-379, 1933.
- Coutinho, M.T.Z.; Bueno, L.L.; Sterzik, A.; Fujiwara, R.T.; Botelho, J.R.; De Maria, M.; Genaro, O.; Linardi, P.M. Participation of Rhipicephalus sanguineus (Acari: Ixodidae) in the epidemiology of canine visceral leishmaniasis. Veter- Parasitol. 2005, 128, 149–155. [Google Scholar] [CrossRef]
- R. D. Smith, D. M. Sells, E. H. Stephenson, M. R. Ristic, and D. L. Huxsoll, "Development of Ehrlichia canis, causative agent of canine ehrlichiosis, in the tick Rhipicephalus sanguineus and its differentiation from a symbiotic Rickettsia," Am J Vet Res, vol. 37, no. 2, pp. 119-26, Feb 1976.
- Huxsoll, D.L.; Hildebrandt, P.K.; Nims, R.M.; Amyx, H.L.; Ferguson, J.A. Epizootiology of Tropical Canine Pancytopenia. J. Wildl. Dis. 1970, 6, 220–225. [Google Scholar] [CrossRef]
- Harrus, S.; Kass, P.H.; Klement, E.; Waner, T. Canine monocytic ehrlichiosis: a retrospective study of 100 cases, and an epidemiological investigation of prognostic indicators for the disease. Veter- Rec. 1997, 141, 360–363. [Google Scholar] [CrossRef]
- Harrus, S.; Waner, T. Diagnosis of canine monocytotropic ehrlichiosis (Ehrlichia canis): An overview. Veter- J. 2011, 187, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Leiva, M.; Naranjo, C.; Peña, M.T. Ocular signs of canine monocytic ehrlichiosis: a retrospective study in dogs from Barcelona, Spain. Veter- Ophthalmol. 2005, 8, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Harrus, S.; Waner, T.; Aizenberg, I.; Foley, J.E.; Poland, A.M.; Bark, H. Amplification of Ehrlichial DNA from Dogs 34 Months after Infection with Ehrlichia canis. J. Clin. Microbiol. 1998, 36, 73–76. [Google Scholar] [CrossRef] [PubMed]
- W. C. Buhles, Jr., D. L. Huxsoll, and M. Ristic, "Tropical canine pancytopenia: Clinical, hematologic, and serologic response of dogs to Ehrlichia canis infection, tetracycline therapy, and challenge inoculation," J Infect Dis, vol. 130, no. 4, pp. 357-67, Oct 1974.
- C. E. Greene, Clinical microbiology and infectious diseases of the dog and cat. Philadelphia: Saunders, 1984, pp. xviii, 967 p.
- Gal, A.; Harrus, S.; Arcoh, I.; Lavy, E.; Aizenberg, I.; Mekuzas-Yisaschar, Y.; Baneth, G. Coinfection with multiple tick-borne and intestinal parasites in a 6-week-old dog. Can Vet J 2007, 48, 619–22. [Google Scholar]
- Hildebrandt, P.K.; Huxsoll, D.L.; Walker, J.S.; Nims, R.M.; Taylor, R.; Andrews, M. Pathology of canine ehrlichiosis (tropical canine pancytopenia). Am J Vet Res 1973, 34, 1309–20. [Google Scholar]
- Nyindo, M.; Huxsoll, D.; Ristic, M.; Kakoma, I.; Brown, J.; Carson, C.; Stephenson, E. CELL-MEDIATED AND HUMORAL IMMUNE-RESPONSES OF GERMAN SHEPHERD DOGS AND BEAGLES TO EXPERIMENTAL-INFECTION WITH EHRLICHIA-CANIS. 1980, 41, 250–254.
- Harrus, S.; Waner, T.; Bark, H.; Jongejan, F.; Cornelissen, A.W.C.A. Recent Advances in Determining the Pathogenesis of Canine Monocytic Ehrlichiosis. J. Clin. Microbiol. 1999, 37, 2745–2749. [Google Scholar] [CrossRef]
- Waner, T.; Harrus, S.; Bark, H.; Bogin, E.; Avidar, Y.; Keysary, A. Characterization of the subclinical phase of canine ehrlichiosis in experimentally infected beagle dogs. Veter- Parasitol. 1997, 69, 307–317. [Google Scholar] [CrossRef]
- Swirski, F.K.; Nahrendorf, M.; Etzrodt, M.; Wildgruber, M.; Cortez-Retamozo, V.; Panizzi, P.; Figueiredo, J.-L.; Kohler, R.H.; Chudnovskiy, A.; Waterman, P.; Aikawa, E.; Mempel, T.R.; Libby, P.; Weissleder, R.; Pittet, M.J. Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science 2009, 325, 612–616. [Google Scholar] [CrossRef]
- Villaescusa, A.; Tesouro, M.A.; García-Sancho, M.; Ayllón, T.; Rodríguez-Franco, F.; Sainz, A. Evaluation of lymphocyte populations in dogs naturally infected by Ehrlichia canis with and without clinical signs. Ticks Tick-borne Dis. 2012, 3, 279–282. [Google Scholar] [CrossRef]
- Gianopoulos, A.; Mylonakis, M.E.; Theodorou, K.; Christopher, M.M. Quantitative and qualitative leukocyte abnormalities in dogs with experimental and naturally occurring acute canine monocytic ehrlichiosis. Veter- Clin. Pathol. 2016, 45, 281–290. [Google Scholar] [CrossRef]
- Silvestre-Roig, C.; Fridlender, Z.G.; Glogauer, M.; Scapini, P. Neutrophil Diversity in Health and Disease. 2019, 40, 565–583. [CrossRef]
- Netea, M.G.; Quintin, J.; van der Meer, J.W. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Moorlag, S.J.; Rodriguez-Rosales, Y.A.; Gillard, J.; Fanucchi, S.; Theunissen, K.; Novakovic, B.; de Bont, C.M.; Negishi, Y.; Fok, E.T.; Kalafati, L.; et al. BCG Vaccination Induces Long-Term Functional Reprogramming of Human Neutrophils. Cell Rep. 2020, 33, 108387. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, M.; Ceron, J.; Leontides, L.; Siarkou, V.; Martinez, S.; Tvarijonaviciute, A.; Koutinas, A.; Harrus, S. Serum Acute Phase Proteins as Clinical Phase Indicators and Outcome Predictors in Naturally Occurring Canine Monocytic Ehrlichiosis. J. Veter- Intern. Med. 2011, 25, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.N.; Levenhagen, M.; Rieck, S.; Labruna, M.; Beletti, M. The spreading process of Ehrlichia canis in macrophages is dependent on actin cytoskeleton, calcium and iron influx and lysosomal evasion. Veter- Microbiol. 2013, 168, 442–446. [Google Scholar] [CrossRef]
- Mavromatis, K.; Doyle, C.K.; Lykidis, A.; Ivanova, N.; Francino, M.P.; Chain, P.; Shin, M.; Malfatti, S.; Larimer, F.; Copeland, A.; et al. The Genome of the Obligately Intracellular Bacterium Ehrlichia canis Reveals Themes of Complex Membrane Structure and Immune Evasion Strategies. J. Bacteriol. 2006, 188, 4015–4023. [Google Scholar] [CrossRef]
- Felek, S.; Huang, H.; Rikihisa, Y. Sequence and Expression Analysis of virB9 of the Type IV Secretion System of Ehrlichia canis Strains in Ticks, Dogs, and Cultured Cells. Infect. Immun. 2003, 71, 6063–6067. [Google Scholar] [CrossRef]
- Harrus, S.; Waner, T.; Friedmann-Morvinski, D.; Fishman, Z.; Bark, H.; Harmelin, A. Down-regulation of MHC class II receptors of DH82 cells, following infection with Ehrlichia canis. Veter- Immunol. Immunopathol. 2003, 96, 239–243. [Google Scholar] [CrossRef]
- Boschiroli, M.L.; Ouahrani-Bettache, S.; Foulongne, V.; Michaux-Charachon, S.; Bourg, G.; Allardet-Servent, A.; Cazevieille, C.; Lavigne, J.-P.; Liautard, J.P.; Ramuz, M.; et al. Type IV secretion and Brucella virulence. Veter- Microbiol. 2002, 90, 341–348. [Google Scholar] [CrossRef]
- Ohashi, N.; Zhi, N.; Lin, Q.; Rikihisa, Y. Characterization and Transcriptional Analysis of Gene Clusters for a Type IV Secretion Machinery in Human Granulocytic and Monocytic Ehrlichiosis Agents. Infect. Immun. 2002, 70, 2128–2138. [Google Scholar] [CrossRef]
- Luo, T.; Patel, J.G.; Zhang, X.; Walker, D.H.; McBride, J.W. Immunoreactive Protein Repertoires of Ehrlichia chaffeensis and E. canis Reveal the Dominance of Hypothetical Proteins and Conformation-Dependent Antibody Epitopes. Infect. Immun. 2021, 89, e0022421. [Google Scholar] [CrossRef]
- Luo, T.; Patel, J.G.; Zhang, X.; Walker, D.H.; McBride, J.W. Ehrlichia chaffeensis and E. canis hypothetical protein immunoanalysis reveals small secreted immunodominant proteins and conformation-dependent antibody epitopes. npj Vaccines 2020, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Chaichanasiriwithaya, W.; Rikihisa, Y. Comparison of PCR with other tests for early diagnosis of canine ehrlichiosis. J. Clin. Microbiol. 1994, 32, 1658–62. [Google Scholar] [CrossRef] [PubMed]
- Kakoma, I.; Hansen, R.D.; E Anderson, B.; A Hanley, T.; Sims, K.G.; Liu, L.; Bellamy, C.; Long, M.T.; Baek, B.K. Cultural, molecular, and immunological characterization of the etiologic agent for atypical canine ehrlichiosis. J. Clin. Microbiol. 1994, 32, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.; Matthewman, L.; Mahan, S.; Semu, S.; Peter, T.; Mason, P.; Brouqui, P.; Raoult, D. Serological evidence for antigenic relationships between Ehrlichia canis and Cowdria ruminantium. Res. Veter- Sci. 1994, 56, 170–174. [Google Scholar] [CrossRef]
- E. B. Breitschwerdt, B. C. Hegarty, and S. I. Hancock, "Sequential evaluation of dogs naturally infected with Ehrlichia canis, Ehrlichia chaffeensis, Ehrlichia equi, Ehrlichia ewingii, or Bartonella vinsonii," J Clin Microbiol, vol. 36, no. 9, pp. 2645-51, Sep 1998.
- B. J. Woody and J. D. Hoskins, "Ehrlichial diseases of dogs," Vet Clin North Am Small Anim Pract, vol. 21, no. 1, pp. 75-98, Jan 1991.
- Cadman, H.; Kelly, P.; Matthewman, L.; Zhou, R.; Mason, P. Comparison of the dot-blot enzyme linked immunoassay with immunofluorescence for detecting antibodies to Ehrlichia canis. Vet Rec 1994, 135, 362–362. [Google Scholar] [CrossRef]
- Waner, T.; Strenger, C.; Keysary, A.; Harrus, S. Kinetics of serologic cross-reactions between Ehrlichia canis and the Ehrlichia phagocytophila genogroups in experimental E. canis infection in dogs. Veter- Immunol. Immunopathol. 1998, 66, 237–243. [Google Scholar] [CrossRef]
- Rikihisa, Y.; A Ewing, S.; Fox, J.C.; Siregar, A.G.; Pasaribu, F.H.; Malole, M.B. Analyses of Ehrlichia canis and a canine granulocytic Ehrlichia infection. J. Clin. Microbiol. 1992, 30, 143–148. [Google Scholar] [CrossRef]
- Harrus, S.; Alleman, A.; Bark, H.; Mahan, S.M.; Waner, T. Comparison of three enzyme-linked immunosorbant assays with the indirect immunofluorescent antibody test for the diagnosis of canine infection with Ehrlichia canis. Veter- Microbiol. 2002, 86, 361–368. [Google Scholar] [CrossRef]
- G. L. Murphy, S. A. Ewing, L. C. Whitworth, J. C. Fox, and A. A. Kocan, "A molecular and serologic survey of Ehrlichia canis, E. chaffeensis, and E. ewingii in dogs and ticks from Oklahoma," Vet Parasitol, vol. 79, no. 4, pp. 325-39, Nov 27 1998.
- Doyle, C.K.; Nethery, K.A.; Popov, V.L.; McBride, J.W. Differentially Expressed and Secreted Major Immunoreactive Protein Orthologs of Ehrlichia canis and E. chaffeensis Elicit Early Antibody Responses to Epitopes on Glycosylated Tandem Repeats. Infect. Immun. 2006, 74, 711–720. [Google Scholar] [CrossRef]
- McBride, J.W.; Doyle, C.K.; Zhang, X.; Cardenas, A.M.; Popov, V.L.; Nethery, K.A.; Woods, M.E. Identification of a Glycosylated Ehrlichia canis 19-Kilodalton Major Immunoreactive Protein with a Species-Specific Serine-Rich Glycopeptide Epitope. Infect. Immun. 2007, 75, 74–82. [Google Scholar] [CrossRef]
- Cárdenas, A.M.; Doyle, C.K.; Zhang, X.; Nethery, K.; Corstvet, R.E.; Walker, D.H.; McBride, J.W. Enzyme-Linked Immunosorbent Assay with Conserved Immunoreactive Glycoproteins gp36 and gp19 Has Enhanced Sensitivity and Provides Species-Specific Immunodiagnosis ofEhrlichia canisInfection. Clin. Vaccine Immunol. 2007, 14, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, D.M.; Zhang, X.; Braga, I.A.; Taques, I.I.G.G.; McBride, J.W. Detection of genotype-specific Ehrlichia canis exposure in Brazilian dogs by TRP36 peptide ELISA. Ticks Tick-borne Dis. 2016, 7, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Lim, S.Y.; Sharma, R.S.K.; Cheng, N.A.B.Y.; Watanabe, M. Molecular Detection of Ehrlichia canis in Dogs in Malaysia. PLOS Neglected Trop. Dis. 2013, 7, e1982. [Google Scholar] [CrossRef] [PubMed]
- Stich, R.W.; Rikihisa, Y.; Ewing, S.A.; Needham, G.R.; Grover, D.L.; Jittapalapong, S. Detection of Ehrlichia canis in Canine Carrier Blood and in Individual Experimentally Infected Ticks with a p30 -Based PCR Assay. J. Clin. Microbiol. 2002, 40, 540–546. [Google Scholar] [CrossRef]
- McClure, J.C.; Crothers, M.L.; Schaefer, J.J.; Stanley, P.D.; Needham, G.R.; Ewing, S.A.; Stich, R.W. Efficacy of a Doxycycline Treatment Regimen Initiated during Three Different Phases of Experimental Ehrlichiosis. Antimicrob. Agents Chemother. 2010, 54, 5012–5020. [Google Scholar] [CrossRef]
- Wen, B.; Rikihisa, Y.; Mott, J.M.; Greene, R.; Kim, H.Y.; Zhi, N.; Couto, G.C.; Unver, A.; Bartsch, R. Comparison of nested PCR with immunofluorescent-antibody assay for detection of Ehrlichia canis infection in dogs treated with doxycycline. J. Clin. Microbiol. 1997, 35, 1852–5. [Google Scholar] [CrossRef]
- Alexandre, N.; Santos, A.S.; Núncio, M.S.; de Sousa, R.; Boinas, F.; Bacellar, F. Detection of Ehrlichia canis by polymerase chain reaction in dogs from Portugal. Veter- J. 2009, 181, 343–344. [Google Scholar] [CrossRef]
- McBride, J.W.; Corstvet, R.E.; Gaunt, S.D.; Chinsangaram, J.; Akita, G.Y.; Osburn, B.I. PCR Detection of Acute Ehrlichia Canis Infection in Dogs. J. Veter- Diagn. Investig. 1996, 8, 441–447. [Google Scholar] [CrossRef]
- M. J. Homer, I. Aguilar-Delfin, S. R. Telford, 3rd, P. J. Krause, and D. H. Persing, "Babesiosis," Clin Microbiol Rev, vol. 13, no. 3, pp. 451-69, Jul 2000.
- Rodríguez-Alarcón, C.A.; Beristain-Ruiz, D.M.; Olivares-Muñoz, A.; Quezada-Casasola, A.; Pérez-Casio, F.; Álvarez-Martínez, J.A.; Tapia-Alanís, J.; Lira-Amaya, J.J.; Rivera-Barreno, R.; Cera-Hurtado, O.S.; et al. Demonstrating the presence of Ehrlichia canis DNA from different tissues of dogs with suspected subclinical ehrlichiosis. Parasites Vectors 2020, 13, 1–7. [Google Scholar] [CrossRef]
- Sirigireddy, K.R.; Ganta, R.R. Multiplex Detection of Ehrlichia and Anaplasma Species Pathogens in Peripheral Blood by Real-Time Reverse Transcriptase-Polymerase Chain Reaction. J. Mol. Diagn. 2005, 7, 308–316. [Google Scholar] [CrossRef]
- Sirigireddy, K.R.; Mock, D.C.; Ganta, R.R. Multiplex Detection of Ehrlichia and Anaplasma Pathogens in Vertebrate and Tick Hosts by Real-Time RT-PCR. Ann. New York Acad. Sci. 2006, 1078, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Doyle, C.K.; Labruna, M.B.; Breitschwerdt, E.B.; Tang, Y.-W.; Corstvet, R.E.; Hegarty, B.C.; Bloch, K.C.; Li, P.; Walker, D.H.; McBride, J.W. Detection of Medically Important Ehrlichia by Quantitative Multicolor TaqMan Real-Time Polymerase Chain Reaction of the dsb Gene. J. Mol. Diagn. 2005, 7, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Peleg, O.; Baneth, G.; Eyal, O.; Inbar, J.; Harrus, S. Multiplex real-time qPCR for the detection of Ehrlichia canis and Babesia canis vogeli. Veter- Parasitol. 2010, 173, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Padmaja, M.; Singh, H.; Panwar, H.; Jyoti, *!!! REPLACE !!!*; Singh, N.K. Development and validation of multiplex SYBR Green real-time PCR assays for detection and molecular surveillance of four tick-borne canine haemoparasites. Ticks Tick-borne Dis. 2022, 13, 101937. [Google Scholar] [CrossRef]
- Iqbal, Z.; Rikihisa, Y. Reisolation of Ehrlichia canis from blood and tissues of dogs after doxycycline treatment. J. Clin. Microbiol. 1994, 32, 1644–1649. [Google Scholar] [CrossRef]
- Breitschwerdt, E.B.; Hegarty, B.C.; Hancock, S.I. Doxycycline Hyclate Treatment of Experimental Canine Ehrlichiosis Followed by Challenge Inoculation with TwoEhrlichia canisStrains. Antimicrob. Agents Chemother. 1998, 42, 362–368. [Google Scholar] [CrossRef]
- Mylonakis, M.E.; Harrus, S.; Breitschwerdt, E.B. An update on the treatment of canine monocytic ehrlichiosis (Ehrlichia canis). Veter- J. 2019, 246, 45–53. [Google Scholar] [CrossRef]
- Jongejan, F.; Crafford, D.; Erasmus, H.; Fourie, J.J.; Schunack, B. Comparative efficacy of oral administrated afoxolaner (NexGard™) and fluralaner (Bravecto™) with topically applied permethrin/imidacloprid (Advantix®) against transmission of Ehrlichia canis by infected Rhipicephalus sanguineus ticks to dogs. Parasites Vectors 2016, 9, 348. [Google Scholar] [CrossRef]
Figure 1.
Ehrlichia canis life cycle in mononuclear cells.
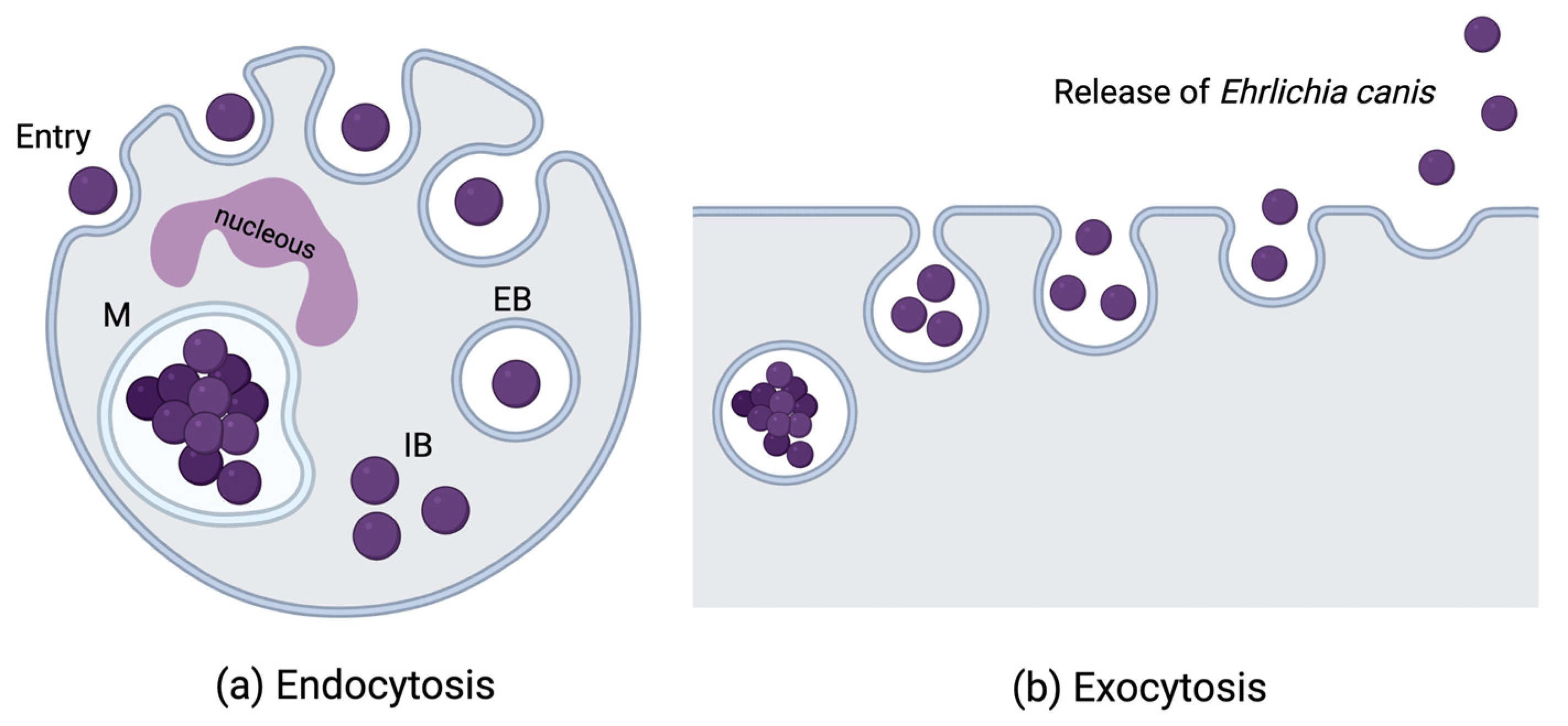
Figure 2.
Rhipicephalus sanguineus life cycle.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



