Submitted:
10 November 2024
Posted:
11 November 2024
You are already at the latest version
Abstract
Leaf temperature (Tleaf) is a critical factor influencing plant ecophysiological processes and ecosystem responses to environmental changes. In this study, we investigated the seasonal Tleaf variations among different growth forms (shrubs and trees) of four common tropical woody species in South China and identified the dominant drivers of Tleaf. We hypothesized that Tleaf patterns between shrubs and trees would differ significantly between dry and rainy seasons and that leaf traits and environmental temperature would predominantly affect Tleaf among species. From December 2022 to October 2023, we measured Tleaf, leaf traits, and environmental conditions. The results revealed that Tleaf was consistently greater than air temperature (Tair) across all species, growth forms, and seasons. The regression line slopes between Tair and Tleaf indicated limited homeothermy, with significant differences observed between seasons. Trees had higher Tleaf values than shrubs across dry and rainy seasons, possibly due to their distinct microclimatic adaptations. Leaf chlorophyll content (LCC) and leaf mass per area (LMA) were significant predictors of Tleaf. Furthermore, while soil temperature (Tsoil) positively influenced Tleaf, Tair had a negative effect on Tleaf. Our findings suggest that Tleaf is intricately linked to leaf traits and environmental conditions, with implications for plant performance and ecosystem dynamics.
Keywords:
leaf temperature
; tropical species
; chlorophyll content
; leaf mass per area
; soil and air temperature
1. Introduction
Leaf temperature (Tleaf) plays a pivotal role in the microenvironment of plants [1], significantly influencing their ecophysiological processes, such as photosynthesis and respiration, which determine the carbon assimilation rate [2,3]. It also governs the transpiration rate through stomatal conductance [4]. Beyond the individual plant level, Tleaf is a crucial factor in ecosystems and regional water, carbon, and energy budgets [5,6]. Its role is prominently recognized within the frameworks of dynamic global vegetation models [7,8]. This underscores the importance of understanding and accurately modeling Tleaf in predicting and managing ecosystem responses to environmental changes.
Tleaf can significantly differ from air temperature (Tair), leading to substantial impacts on carbon and water fluxes [9]. Different studies have shown that the difference between Tleaf and Tair can reach 20°C and that this thermal decoupling varies greatly across species and environments [1,10]. In certain environments, such as alpine regions and humid tropics, Tleaf can be as much as 20°C higher than Tair, whereas in warm deserts, it can be 15–20℃ lower [2,9]. These deviations highlight the complex relationships between Tleaf and Tair and their ecological implications.
The thermal response of a leaf to changes in air temperature is crucial for its physiological function, and this response is characterized by the slope of the regression line between Tleaf and Tair [11,12]. A slope less than 1 indicates limited homeothermy, where leaves remain cooler than the air, which is especially useful for carbon uptake at higher Tair [3,13]. A slope equal to 1 represents poikilothermy, where Tleaf closely follows Tair, showing little to no thermal regulation [7]. Conversely, a slope greater than 1 suggests megathermy, where leaves heat up more than the air does, potentially leading to thermal stress [14]. The implications of these responses for plant performance are significant but not fully explored, and the mechanisms behind each type of response require further investigation.
Trees and shrubs may have different physiological and morphological adaptations to their environment, which can affect how their leaves respond to temperature changes. Previous studies have reported that shrubs have exhibited greater sensitivity to climate warming than trees have in recent decades [15,16]. However, Gazol and Camarero [17] reported that shrub species might actually be less affected by temperature fluctuations than trees are, as they are more significantly influenced by microclimatic conditions, topography, and soil temperature. This microclimatic connection allows shrubs to grow at sites that are more sheltered from regional atmospheric temperatures, potentially increasing their survival in areas with more extreme climate conditions [18].
Previous studies have shown that increases in Tleaf are associated with decreased transpiration rates [3,19], which are a consequence of stomatal closure in response to environmental stressors, such as water scarcity [20]. In the rainy season, when soil moisture is plentiful, plants reach their zenith in terms of liquid photosynthesis, stomatal conductance, transpiration, and leaf water potential [21]. This optimal physiological state effectively shields plants from high-temperature damage [22], facilitating normal growth. In contrast, during the dry season, leaf stomata are predominantly impacted by drought conditions [23,24]. A reduction in stomatal density and the closure of stomata lead to a significant decline in photosynthesis and an increase in Tleaf [25].
Tleaf is intricately linked to the leaf’s energy balance, which is influenced by a suite of leaf traits that modulate the absorption and dissipation of energy [7,26]. Specific leaf characteristics, such as the leaf chlorophyll content (LCC) [27], leaf area (LA) [9], leaf thickness (LT) [28], leaf mass per area (LMA) [29], and leaf water content (LWC) [1], play pivotal roles in this context. As the LA increases, the size of the leaf’s boundary layer also correspondingly increases [30]. This increase in boundary layer thickness leads to a decrease in the rate of sensible heat exchange between the leaf and its surrounding air [31]. Consequently, larger leaves typically exhibit higher maximum surface temperatures than smaller leaves do [32,33], assuming that all other factors are equal. When Tleaf increases, the activity of chlorophyll synthesis enzymes increases, leading to increased chlorophyll content [29]. However, once a certain Tleaf (>30°C) is reached, the enzyme activity rapidly decreases, and the chlorophyll content also rapidly decreases [34]. A higher LWC results in greater thermal inertia due to the high specific heat capacity of water. This allows leaves to absorb more heat with less significant temperature increases, thus protecting them from rapid temperature fluctuations and potential damage [35]. Thicker leaves can accumulate more energy and maintain a higher thermal mass [28], which helps stabilize Tleaf with minimal fluctuations. This is facilitated by the positive correlation between LT, LMA, and the content of heat-retaining substances within the leaf, such as water and macromolecules [30]. Consequently, an increase in LT and LMA not only enhances the leaf’s water retention capacity but also slows the rate at which Tleaf increases. Despite the acknowledged influence of leaf traits on Tleaf, the primary determinants that drive Tleaf remain elusive.
In this study, we selected four common tropical woody species in South China on a university campus and examined the seasonal variations (rainy and dry seasons) in Tleaf between different growth forms (shrubs and trees) and their dominant drivers. We hypothesized that (1) Tleaf consistently exceeded Tair, suggesting limited homeothermy; (2) the pattern of Tleaf between shrubs and trees would be significantly different between the dry and rainy seasons; and (3) leaf traits and environmental temperature could strongly affect Tleaf among species.
2. Materials and Methods
2.1. Site Description
The study area was located at Hainan Normal University Longkun South Campus in Haikou, Hainan Province, China (19.59°N, 110.20°E). The area has an annual average sunshine duration of 1752 hours and an average annual temperature of 23.8°C. The climate is characterized by a tropical maritime monsoon with an average annual rainfall of 1724.5 mm, which is highly unevenly distributed. The year is divided into two seasons: the rainy season, which spans from May to October, and the dry season, which spans from November to April of the following year. The rainfall during the rainy season accounts for approximately 80% of the annual total [36]. The average annual evaporation is 1834 mm, with an average relative humidity of 85%. The area is predominantly influenced by northeast and east winds, with an average annual wind speed of 3.4 m s−1.
2.2. Measurement of Leaf Temperature
We selected four common woody plants on the campus for experimentation, including two tree species, Bauhinia blakeana and Fagraea ceilanica, and two shrubs, Ligustrum × vicaryi Rehder and Hibiscum rosa-sinensis Linn. From December 2022 to October 2023, leaf temperature (Tleaf) measurements were conducted on clear days in the middle of each month. Between 12:00 and 14:00 under full sunlight conditions, the leaf canopy temperature was measured remotely via an infrared thermometer gun (Fluke-MT4, Fluke Corporation, Everett, USA) [37]. Canopies emit longwave infrared radiation as a function of their temperature. The infrared thermometer senses this radiation and converts it to an electrical signal, which is displayed as temperature [38]. For each tree species, three individuals were selected for replication, and for each individual, eight random leaves were repeatedly measured, and the average value was taken.
2.3. Leaf Trait Measurements
From each of the three individual plants, twenty fully mature and robust green leaves were collected at random from the sunlit central portion of the canopy. The leaf chlorophyll content (LCC, SPAD) was measured via a chlorophyll meter (SPAD-502, Konica Minolta Holdings, Tokyo, Japan). The fresh leaf mass (LFM, g) was then measured via an analytical balance with a precision of one part in ten thousand. The leaves were subsequently placed into a portable leaf area scanner (LI-3000C, LI-COR, Nebraska, USA) to scan and determine the single leaf area (LA, cm²) [39]. Finally, the leaves were placed in envelopes and dried in a 60°C oven for 48 hours until they reached a constant weight, after which they were weighed again, and the leaf dry mass (LDM, g) was recorded. The leaf mass per area (LMA, g cm−2) and leaf water content (LWC, %) were calculated as follows:
2.4. Environmental Conditions Measurement
Infrared radiation thermometers were used to measure the temperature of the cloudless open sky near each target plant (Tair). Portable soil moisture and temperature meters (TDR 350, Spectrum Technologies Inc., Aurora, USA) were employed to determine the soil temperature (Tsoil) and soil water content (SWC). For each tree species, three individuals were selected for replication, with measurements taken in a circular pattern three times around each plant, and the average value was then calculated [40].
2.5. Data Analysis
We calculated the slope of the regression line between Tair and Tleaf for different growth types (shrub vs. tree), seasons (rainy vs. dry), and species. Analyses of covariance (ANCOVAs) were performed to test whether the slopes among those regression lines were significantly different [41]. To examine the effects of growth type and season on leaf traits, we used linear mixed effect models with sampling date as a random effect to account for temporal autocorrelation.
To estimate the relative importance of the predictor variables, we performed multimodel inference via Akaike’s information criterion (AIC). The relative importance value, regarded as the overall support for each predictor across all models, was estimated as the sum of the Akaike weights for the models in which the predictor appeared. A cutoff relative importance value of 0.8 was set to differentiate between the important and unimportant predictors. Partial regression plots were created for each important predictor to illustrate its relationship with Tleaf while holding all the other variables constant. All the statistical analyses were performed via the R statistical software package (ver. 4.3.0). All the statistical tests were considered significant at P < 0.05.
3. Results
3.1. Leaf Temperature Across Different Groups
Tleaf was consistently greater than Tair with respect to growth type, season, and species (Table 1). In general, Tleaf increased with Tair across the different groups (Figure 1). Across the species, the slope of the regression line between Tair and Tleaf was 0.45 (< 1), indicating limited homeothermy (Table 1 and Figure 1). For different growth types and species, the slopes were lower than one and did not differ among them (Table 1 and Figure 1). During different seasons, the slopes significantly differed (Table 1). The plants in the dry season displayed megathermy (slope >1), whereas the plants in the rainy season presented a slope < 0, suggesting a negative relationship between Tleaf and Tair (Figure 1).
3.2. Leaf Traits According to Growth Type and Season
The LCC was significantly greater in the rainy season than in the dry season (P < 0.05, Figure 2). However, no significant differences between seasons were found for LA, LDM, Tleaf, LMA, or LWC (P > 0.05, Figure 2). Compared with shrubs, trees had greater LCC, LA, LDM, Tleaf, and LMA (P < 0.05), whereas growth form had no significant effect on LWC (P > 0.05, Figure 2). We found no significant interactive effects between season and growth form on any of the traits (P > 0.05, Figure 2).
3.3. The Impact of Environmental Conditions and Leaf Traits
Model selection analysis showed that the best model describing Tleaf included LCC, Tsoil, LMA, and Tair as important predictors (Figure 3). Partial regression analysis indicated that Tleaf increased significantly with increasing LCC and Tsoil but decreased with increasing LMA and Tair (P < 0.05, Figure 4).
4. Discussion
4.1. Thermal Regulation Capacity of Leaves
Consistent with our initial hypothesis, we found that Tleaf consistently surpassed Tair, which aligns with certain prior studies [1,42,43]. In numerous tropical studies, Tleaf can exceed ambient Tair by more than 10°C in sunlit leaves[44,45]. This temperature disparity is attributed to variations in microclimatic conditions [10,28]. For example, wind speeds below 0.5 m s−1 are common in nature [46], and even transient lulls can cause Tleaf to rise by more than 5°C within just a few seconds [47]. Furthermore, the leaf energy balance indicates that the leaf-to-air temperature difference is contingent upon the net energy provided (or lost) through radiation and the energy expended via transpiration [10]. Zhou et al. [1] reported that the effects of leaf physical warming are stronger than those of leaf transpirational cooling in hot environments. Consequently, the Tleaf of all the species was found to be greater than the ambient Tair.
The mechanistic relationship between Tleaf and Tair has received considerable attention in the literature [2,48]. Some studies have argued that leaves exhibit either limited homeothermy [11,49], poikilothermy [50], or megathermy [13,51]. In this study, the regression line slopes between Tair and Tleaf for the overall dataset, each different growth type, and each species were all less than one, indicating limited homeothermy. Energy balance theory suggests that limited homeothermy occurs under conditions of high stomatal conductance and low convective resistance, which implies that transpirational cooling outweighs physical warming [13]. However, our research did not observe a greater transpirational cooling effect over physical warming, as Tleaf consistently exceeded Tair. This unexpected result could be explained by the possibility that, under extremely hot conditions, plants may mitigate physical warming to alleviate heat stress. For example, Zhou et al. [1] reported that savanna woodland species possess the lowest absorptivity and the highest reflectivity, thereby reducing the radiation load and resulting in low levels of physical warming. Our findings indicate that the relationships between Tair and Tleaf, as well as the interactions between leaf physical traits and environmental parameters, are too intricate to be accurately simulated by a simplistic model.
Contrary to our initial hypothesis, plants during the dry season exhibited megathermy, characterized by a slope greater than 1, whereas those in the rainy season presented a slope less than 0, indicating a negative correlation between Tleaf and Tair. The capacity of plants to mitigate heat stress at the leaf level appears to decrease with increasing water stress [52,53]. In light of the anticipated rise in Tair due to global warming, the evaporative demand is projected to increase, resulting in the escalation of the air vapor pressure deficit (VPD) across many regions [54]. Prolonged exposure to elevated temperatures, in conjunction with high leaf-to-air VPD, is anticipated to elicit stomatal closure as a strategy to conserve water in hot-dry environments [43,55]. A reduction in stomatal aperture can lead to a faster increase in Tleaf than in Tair during the dry season, as diminished transpiration results in less cooling of the leaf surface [56]. Conversely, a negative relationship between Tleaf and Tair during the hot–wet season should increase overall carbon uptake by keeping leaves within optimal photosynthetic temperature ranges and below damaging temperatures [9].
4.2. Effects of Growth Form and Seasonal Variation on Tleaf
In this study, we found that the Tleaf of shrubs was significantly lower than that of trees, which may be interpreted in three ways. First, smaller leaves, as typically found on shrubs, have a thinner boundary layer [57], which allows for more efficient cooling through convection rather than evaporation of water [30,58]. Second, shrubs often have a relatively high surface area-to-volume ratio because of their relatively small stature and large number of small leaves [59]. This can lead to increased transpiration rates, which in turn can result in greater cooling of the leaf surface through the evaporation of water [60]. Finally, shrubs, which are closer to the ground, are more susceptible to the influence of soil temperature and moisture [61], which can significantly impact Tleaf. In contrast, trees, which are more exposed to atmospheric conditions, are less affected by ground-level microclimates. The soil temperatures and microclimates beneath shrubs can differ from those under trees, with shrubs often having denser canopies that shade the ground [62], potentially leading to lower soil temperatures and, consequently, reduced Tleaf [63]. As a result, the Tleaf of trees is generally greater than that of shrubs. This differential Tleaf response has implications for our understanding of plant thermal regulation and its ecological consequences for different growth forms.
Our findings revealed that there was no significant seasonal variation in leaf Tleaf across all the species studied. This outcome contradicts the results of a previous study by Rey-Sánchez et al.[45], which indicated that tree canopies in a tropical forest presented higher Tleaf values during the dry season than during the wet season. Additionally, Rey-Sánchez et al. [45] reported a correlation between Tleaf and photosynthetic photon flux density in the wet season, a relationship that was absent in the dry season. This discrepancy was hypothesized to be due to seasonal variations in wind speed, plant physiology, and canopy phenology [64,65]. In contrast, our study demonstrated a consistent Tleaf between the dry and wet seasons, which we attributed to the thermal regulation strategies employed by the plants. These strategies include transpirational cooling, physical warming, and species-specific adaptations that enable them to maintain optimal temperatures for photosynthesis and other physiological processes [1,66,67]. These adaptive mechanisms assist plants in buffering against extreme temperature fluctuations, thereby sustaining relatively stable Tleaf values irrespective of seasonal changes.
In contrast to our second hypothesis, we found that trees presented significantly higher Tleaf values than shrubs did across the rainy and dry seasons. Trees and shrubs may have evolved different adaptive strategies to cope with environmental conditions [68,69]. Trees, as the dominant species in a forest canopy, may have adapted to maintain a relatively high Tleaf as a way to optimize photosynthesis and growth under full sunlight conditions [70]. Shrubs, on the other hand, may have adapted to lower light and temperature conditions characteristic of the understory, leading to lower Tleaf [71]. These factors combined can result in trees maintaining higher Tleaf values than shrubs do, regardless of the seasonal changes in rainfall.
4.3. Tleaf Regulated by LCC and LMA
In agreement with our third hypothesis, we found that Tleaf increased with increasing LCC. This finding aligns with the research of Winter and Koniger [27], who noted significant reductions in LCC and photosynthetic capacity in Gossypium hirsutum L. as Tleaf decreased from approximately 33°C to 27°C under high light conditions. The higher the LCC is, the greater the leaf’s capacity to absorb light energy, leading to increased heat production and, consequently, an increase in Tleaf [72,73]. Conversely, Khan et al. [74] reported that the Tleaf of Triticum aestivum at temperatures ranging from 20°C to 35°C did not affect the LCC or nitrogen content per unit leaf area. Phylogenetic history can significantly influence plant traits, including LCC and Tleaf [34]. Our study indicates that different plant species have evolved distinct adaptive strategies to cope with environmental conditions, affecting their Tleaf and LCC.
In this study, we found a negative correlation between Tleaf and LMA, which is in line with several previous studies [75,76]. Owing to the significant positive correlation between LT and LMA with the substances contained within the leaf, such as water and macromolecules, an increase in LT and LMA not only enhances the water retention of the leaf but also slows the rate of increase in Tleaf [77].
4.4. Tsoil and Tair Mediated Tleaf
In agreement with our third hypothesis, the multivariate analysis revealed that Tleaf was positively regulated by Tsoil and negatively mediated by Tair.
Similar findings were also reported in several previous studies [76,78,79], which reported a positive correlation between Tleaf and Tsoil. The relationship between epidermal cell area and stomatal density follows an inverse pattern, as noted by Rogiers et al. [80]. Consequently, an increase in Tsoil can lead to a reduction in stomatal density, which may be attributed to the increase in epidermal cell size and leaf expansion influenced by Tsoil[81]. This reduction in stomatal density, coupled with stomatal closure, could significantly curtail photosynthesis and consequently increase Tleaf [82]. Furthermore, the stresses induced by elevated Tsoil led to a decrease in the relative water content of the leaves, which in turn resulted in increased Tleaf and cellular rupture [83].
Additionally, Manzi et al. [9] reported a positive correlation between the Tleaf of sun-exposed leaves and Tair across various species. In contrast, our study revealed an inverse relationship, which we attributed to the enhanced transpirational cooling effect under conditions of warming. Several mechanisms have been proposed to explain the stimulation of plant transpiration by elevated temperatures. First, elevated temperatures lead to increased expression of aquaporins in the mesophyll and heightened activity of potassium (K+) channels, which are crucial for stomatal opening, facilitating greater transpiration rates [84]. Second, warming enhances plant transpiration by increasing membrane fluidity, which is essential for water transport, and by reducing water viscosity, thereby lowering resistance to water flow within the plant [85]. Third, our research indicates that under warmer conditions, fine roots adapt by becoming thinner and growing deeper into the soil, which improves their capacity for water absorption [86]. These observations underscore the intricate and heterogeneous responses of diverse vegetation types to climate change, which has profound implications for the field of forest ecology and the formulation of adaptive management strategies.
5. Conclusions
Our study demonstrated that Tleaf consistently surpassed Tair, indicating limited homeothermy in tropical woody species. Trees have higher Tleaf values than shrubs do, potentially due to their distinct microclimatic adaptations. Notably, leaf traits, with a focus on LCC and LMA, substantially influence Tleaf. Tsoil positively modulates Tleaf, whereas Tair exerts a negative regulatory effect. These insights highlight the complex interplay between leaf thermal dynamics and environmental factors. Future studies should broaden the species and climate ranges to obtain more generalizable findings. Examining additional environmental factors and their interactions with leaf traits will deepen our understanding of Tleaf dynamics. Long-term studies will also be crucial for tracking Tleaf responses to climate change, informing ecosystem management strategies.
Author Contributions
Writing-Original Draft, Q.H.; Methodology, Q.H.; Investigation, Q.H., N.T., H.C., K.J., and Y.G.; Conceptualization, D.J.; Formal analysis, D.J.; Funding acquisition, D.J.; Writing-review & editing, D.J.; Supervision, D.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32201332), the Hainan Provincial Natural Science Foundation of China (322QN304, 423RC477), the Innovation Platform for Academicians of Hainan Province (YSPTZX202130), and the Startup Foundation for Advanced Talents of Hainan Normal University.
Data Availability Statement
Data and R code supporting this study will be made available upon request.
Acknowledgments
I am deeply grateful to the editors and reviewers for their valuable feedback, which significantly improved the manuscript. I also appreciate the contributions of undergraduates Yueying Xu, Furong Zhuang, and Fangxia Song to the field sampling. Last, I extend my heartfelt thanks to my wife and family for their unwavering support, which was crucial to the success of this research.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhou, Y.; Kitudom, N.; Fauset, S.; Slot, M.; Fan, Z.; Wang, J.; Liu, W.; Lin, H. Leaf thermal regulation strategies of canopy species across four vegetation types along a temperature and precipitation gradient. Agric. For. Meteorol. 2023, 343, 109766. [Google Scholar] [CrossRef]
- Blonder, B.; Michaletz, S.T. A model for leaf temperature decoupling from air temperature. Agric. For. Meteorol. 2018, 262, 354–360. [Google Scholar] [CrossRef]
- Kibler, C.L.; Trugman, A.T.; Roberts, D.A.; Still, C.J.; Scott, R.L.; Caylor, K.K.; Stella, J.C.; Singer, M.B. Evapotranspiration regulates leaf temperature and respiration in dryland vegetation. Agric. For. Meteorol. 2023, 339, 109560. [Google Scholar] [CrossRef]
- Gauthey, A.; Kahmen, A.; Limousin, J.M.; Vilagrosa, A.; Didion-Gency, M.; Mas, E.; Milano, A.; Tunas, A.; Grossiord, C. High heat tolerance, evaporative cooling, and stomatal decoupling regulate canopy temperature and their safety margins in three European oak species. Global Change Biol. 2024, 30, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yamanaka, T. Application of a two-source model for partitioning evapotranspiration and assessing its controls in temperate grasslands in central Japan. ECOHYDROLOGY 2014, 7, 345–353. [Google Scholar] [CrossRef]
- Gerlein-Safdi, C.; Koohafkan, M.C.; Chung, M.; Rockwell, F.E.; Thompson, S.; Caylor, K.K. Dew deposition suppresses transpiration and carbon uptake in leaves. Agric. For. Meteorol. 2018, 259, 305–316. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Weiser, M.D.; McDowell, N.G.; Zhou, J.; Kaspari, M.; Helliker, B.R.; Enquist, B.J. The energetic and carbon economic origins of leaf thermoregulation. Nat. Plants 2016, 2, 16129. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Madhavji, S.; Way, D.A. Contrasting acclimation responses to elevated CO2 and warming between an evergreen and a deciduous boreal conifer. Global Change Biol. 2020, 26, 3639–3657. [Google Scholar] [CrossRef]
- Manzi, O.J.L.; Wittemann, M.; Dusenge, M.E.; Habimana, J.; Manishimwe, A.; Mujawamariya, M.; Ntirugulirwa, B.; Zibera, E.; Tarvainen, L.; Nsabimana, D. , et al. Canopy temperatures strongly overestimate leaf thermal safety margins of tropical trees. New Phytol. 2024, 243, 2115–2129. [Google Scholar] [CrossRef]
- Fauset, S.; Freitas, H.C.; Galbraith, D.R.; Sullivan, M.J.P.; Aidar, M.P.M.; Joly, C.A.; Phillips, O.L.; Vieira, S.A.; Gloor, M.U. Differences in leaf thermoregulation and water-use strategies between three co-occurring Atlantic forest tree species. Plant, Cell & Environment 2018, 41, 1618–1631. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Weiser, M.D.; Zhou, J.Z.; Kaspari, M.; Helliker, B.R.; Enquist, B.J. Plant Thermoregulation: Energetics, Trait-Environment Interactions, and Carbon Economics. Trends Ecol. Evol. 2015, 30, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, A.T.; Coombs, L.; Ahuanari, R.D.S.; Fortier, R.P.; Feeley, K.J. Leaf thermal safety margins decline at hotter temperatures in a natural warming 'experiment' in the Amazon. New Phytol. 2024, 241, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Still, C.J.; Page, G.; Rastogi, B.; Griffith, D.M.; Aubrecht, D.M.; Kim, Y.; Burns, S.P.; Hanson, C.V.; Kwon, H.; Hawkins, L. , et al. No evidence of canopy-scale leaf thermoregulation to cool leaves below air temperature across a range of forest ecosystems. Proc. Natl. Acad. Sci. U. S. A. 2022, 119, 8. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Still, C.J.; Lee, C.K.F.; Ryu, Y.; Blonder, B.; Wang, J.; Bonebrake, T.C.; Hughes, A.; Li, Y.; Yeung, H.C.H. , et al. Does plant ecosystem thermoregulation occur? An extratropical assessment at different spatial and temporal scales. New Phytol. 2023, 238, 1004–1018. [Google Scholar] [CrossRef]
- García-Cervigón Morales, A.I.; Olano Mendoza, J.M.; Eugenio Gozalbo, M.; Camarero Martínez, J.J. Arboreal and prostrate conifers coexisting in Mediterranean high mountains differ in their climatic responses. DENDROCHRONOLOGIA 2012, 30, 279–286. [Google Scholar] [CrossRef]
- Pellizzari, E.; Camarero, J.J.; Gazol, A.; Granda, E.; Shetti, R.; Wilmking, M.; Moiseev, P.; Pividori, M.; Carrer, M. Diverging shrub and tree growth from the Polar to the Mediterranean biomes across the European continent. Global Change Biol. 2017, 23, 3169–3180. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J. Mediterranean dwarf shrubs and coexisting trees present different radial-growth synchronies and responses to climate. Plant Ecol. 2012, 213, 1687–1698. [Google Scholar] [CrossRef]
- Yang, J.; Cooper, D.J.; Li, Z.; Song, W.; Zhang, Y.; Zhao, B.; Han, S.; Wang, X. Differences in tree and shrub growth responses to climate change in a boreal forest in China. DENDROCHRONOLOGIA 2020, 63, 125744. [Google Scholar] [CrossRef]
- Ranawana, S.R.W.M.C.J.K.; Bramley, H.; Palta, J.A.; Siddique, K.H.M. Role of Transpiration in Regulating Leaf Temperature and its Application in Physiological Breeding. In Translating Physiological Tools to Augment Crop Breeding, Harohalli Masthigowda, M.; Gopalareddy, K.; Khobra, R.; Singh, G.; Pratap Singh, G., Eds. Springer Nature Singapore: Singapore, 2023; pp 91-119.
- Henry, C.; John, G.P.; Pan, R.; Bartlett, M.K.; Fletcher, L.R.; Scoffoni, C.; Sack, L. A stomatal safety-efficiency trade-off constrains responses to leaf dehydration. Nat. Commun. 2019, 10, 3398. [Google Scholar] [CrossRef]
- Joshi, J.; Stocker, B.D.; Hofhansl, F.; Zhou, S.; Dieckmann, U.; Prentice, I.C. Towards a unified theory of plant photosynthesis and hydraulics. Nat. Plants 2022, 8, 1304–1316. [Google Scholar] [CrossRef]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B: Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Barral, A. Stomata feel the pressure. Nat. Plants 2019, 5, 244–244. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sack, L.; Li, Y.; Zhang, J.; Yu, K.; Zhang, Q.; He, N.; Yu, G. Relationships of stomatal morphology to the environment across plant communities. Nat. Commun. 2023, 14, 6629. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Zhu, Y.; Kang, H.; Liu, C. Spatial variations in stomatal traits and their coordination with leaf traits in Quercus variabilis across Eastern Asia. Sci. Total Environ. 2021, 789, 147757. [Google Scholar] [CrossRef]
- Bison, N.N.; Michaletz, S.T. Variation in leaf carbon economics, energy balance, and heat tolerance traits highlights differing timescales of adaptation and acclimation. New Phytol. 2024, 242, 1919–1931. [Google Scholar] [CrossRef]
- Winter, K.; Königer, M. Dry matter production and photosynthetic capacity in Gossypium hirsutum L. under conditions of slightly suboptimum leaf temperatures and high levels of irradiance. Oecologia 1991, 87, 190–197. [Google Scholar] [CrossRef]
- Leigh, A.; Sevanto, S.; Ball, M.C.; Close, J.D.; Ellsworth, D.S.; Knight, C.A.; Nicotra, A.B.; Vogel, S. Do thick leaves avoid thermal damage in critically low wind speeds? New Phytol. 2012, 194, 477–487. [Google Scholar] [CrossRef]
- Li, X.; Zhang, W.; Niu, D.; Liu, X. Effects of abiotic stress on chlorophyll metabolism. Plant Sci. 2024, 342, 112030. [Google Scholar] [CrossRef]
- Tserej, O.; Feeley, K.J. Variation in leaf temperatures of tropical and subtropical trees are related to leaf thermoregulatory traits and not geographic distributions. Biotropica 2021, 53, 868–878. [Google Scholar] [CrossRef]
- Okajima, Y.; Taneda, H.; Noguchi, K.; Terashima, I. Optimum leaf size predicted by a novel leaf energy balance model incorporating dependencies of photosynthesis on light and temperature. Ecol. Res. 2012, 27, 333–346. [Google Scholar] [CrossRef]
- Leigh, A.; Sevanto, S.; Close, J.D.; Nicotra, A.B. The influence of leaf size and shape on leaf thermal dynamics: does theory hold up under natural conditions? Plant, Cell Environ. 2017, 40, 237–248. [Google Scholar] [CrossRef]
- Baird, A.S.; Taylor, S.H.; Pasquet-Kok, J.; Vuong, C.; Zhang, Y.; Watcharamongkol, T.; Scoffoni, C.; Edwards, E.J.; Christin, P.-A.; Osborne, C.P. , et al. Developmental and biophysical determinants of grass leaf size worldwide. Nature 2021, 592, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors Influencing Leaf Chlorophyll Content in Natural Forests at the Biome Scale. FRONT ECOL EVOL 2018, 6. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, Y.; Yue, Z.; Chen, X.; Cao, X.; Ai, X.; Jiang, B.; Xing, Y. The leaf-air temperature difference reflects the variation in water status and photosynthesis of sorghum under waterlogged conditions. PLOS ONE 2019, 14, e0219209. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Zhang, N.; Zhang, Y.; Yin, D.; Hao, J.; Wang, S.; Li, S.; Xu, W.; Yan, W.; Meng, X. , et al. The development of local ambient air quality standards: A case study of Hainan Province, China. EEH 2024, 3, 11–20. [Google Scholar] [CrossRef]
- Muller, J.D.; Rotenberg, E.; Tatarinov, F.; Vishnevetsky, I.; Dingjan, T.; Kribus, A.; Yakir, D. Dual reference method for high precision infrared measurement of leaf surface temperature under field conditions. New Phytol. 2021, 232, 2535–2546. [Google Scholar] [CrossRef]
- Yu, P.; Meng, P.; Tong, X.; Zhang, Y.; Li, J.; Zhang, J.; Liu, P. Temperature sensitivity of leaf flushing in 12 common woody species in eastern China. Sci. Total Environ. 2023, 861, 160337. [Google Scholar] [CrossRef]
- Yang, B.; He, Y.; Chen, W. A simple method for estimation of leaf dry matter content in fresh leaves using leaf scattering albedo. Global Ecol. Conserv. 2020, 23, e01201. [Google Scholar] [CrossRef]
- Urbina-Garcia, O.; Fernandez-Gamiz, U.; Zulueta, E.; Ugarte-Anero, A.; Portal-Porras, K. Indoor Air Quality Measurements in Enclosed Spaces Combining Activities with Different Intensity and Environmental Conditions. In Buildings, 2024; Vol. 14.
- da Silva Duarte, V.; Treu, L.; Campanaro, S.; Fioravante Guerra, A.; Giacomini, A.; Mas, A.; Corich, V.; Lemos Junior, W.J.F. Investigating biological mechanisms of colour changes in sustainable food systems: The role of Starmerella bacillaris in white wine colouration using a combination of genomic and biostatistics strategies. Food Res. Int. 2024, 193, 114862. [Google Scholar] [CrossRef]
- Rey-Sánchez, A.C.; Slot, M.; Posada, J.M.; Kitajima, K. Spatial and seasonal variation in leaf temperature within the canopy of a tropical forest. Clim. Res. 2016, 71, 75–89. [Google Scholar] [CrossRef]
- Crous, K.Y.; Cheesman, A.W.; Middleby, K.; Rogers, E.I.E.; Wujeska-Klause, A.; Bouet, A.Y.M.; Ellsworth, D.S.; Liddell, M.J.; Cernusak, L.A.; Barton, C.V.M. Similar patterns of leaf temperatures and thermal acclimation to warming in temperate and tropical tree canopies. Tree Physiol. 2023, 43, 1383–1399. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.E.; Goulden, M.L. Seasonal patterns of tropical forest leaf area index and CO2 exchange. J GEOPHYS RES-BIOGEO 2008, 113, 12. [Google Scholar] [CrossRef]
- Rey-Sánchez, A.C.; Slot, M.; Posada, J.M.; Kitajima, K. Spatial and seasonal variation in leaf temperature within the canopy of a tropical forest. Clim. Res. 2017, 71, 75–89. [Google Scholar] [CrossRef]
- Vogel, S. Leaves in the lowest and highest winds: temperature, force and shape. New Phytol. 2009, 183, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S. Living in a physical world V. Maintaining temperature. J. Biosci. (Bangalore) 2005, 30, 581–590. [Google Scholar] [CrossRef]
- Cavaleri, M.A. Cold-blooded forests in a warming world. New Phytol. 2020, 228, 1455–1457. [Google Scholar] [CrossRef]
- Cook, A.M.; Berry, N.; Milner, K.V.; Leigh, A. Water availability influences thermal safety margins for leaves. Funct. Ecol. 2021, 35, 2179–2189. [Google Scholar] [CrossRef]
- Drake, J.E.; Harwood, R.; Vårhammar, A.; Barbour, M.M.; Reich, P.B.; Barton, C.V.M.; Tjoelker, M.G. No evidence of homeostatic regulation of leaf temperature in Eucalyptus parramattensis trees: integration of CO2 flux and oxygen isotope methodologies. New Phytol. 2020, 228, 1511–1523. [Google Scholar] [CrossRef]
- Pau, S.; Detto, M.; Kim, Y.; Still, C.J. Tropical forest temperature thresholds for gross primary productivity. Ecosphere 2018, 9, 12. [Google Scholar] [CrossRef]
- Opio, A.; Jones, M.B.; Kansiime, F.; Otiti, T. Influence of climate variables on Cyperus papyrus stomatal conductance in Lubigi wetland, Kampala, Uganda. Afr. J. Aquat. Sci. 2015, 40, 187–192. [Google Scholar] [CrossRef]
- Siebert, S.; Ewert, F.; Eyshi Rezaei, E.; Kage, H.; Graß, R. Impact of heat stress on crop yield—on the importance of considering canopy temperature. Environ. Res. Lett. 2014, 9, 044012. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant responses to rising vapor pressure deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [PubMed]
- Merilo, E.; Yarmolinsky, D.; Jalakas, P.; Parik, H.; Tulva, I.; Rasulov, B.; Kilk, K.; Kollist, H. Stomatal VPD Response: There Is More to the Story Than ABA. Plant Physiol. 2018, 176, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Pankasem, N.; Hsu, P.-K.; Lopez, B.N.K.; Franks, P.J.; Schroeder, J.I. Warming triggers stomatal opening by enhancement of photosynthesis and ensuing guard cell CO2 sensing, whereas higher temperatures induce a photosynthesis-uncoupled response. New Phytol. 2024, n/a. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Qian, H. Drivers of the differentiation between broad-leaved trees and shrubs in the shift from evergreen to deciduous leaf habit in forests of eastern Asian subtropics. Plant Divers. 2023, 45, 535–543. [Google Scholar] [CrossRef]
- Blasini, D.E.; Koepke, D.F.; Bush, S.E.; Allan, G.J.; Gehring, C.A.; Whitham, T.G.; Day, T.A.; Hultine, K.R. Tradeoffs between leaf cooling and hydraulic safety in a dominant arid land riparian tree species. Plant, Cell Environ. 2022, 45, 1664–1681. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Xu, L.; Chen, Z.; He, N. Variation in leaf morphological, stomatal, and anatomical traits and their relationships in temperate and subtropical forests. Sci. Rep. 2019, 9, 5803. [Google Scholar] [CrossRef]
- Wang, R.R.; Zeng, J.X.; Chen, K.H.; Ding, Q.H.; Shen, Q.R.; Wang, M.; Guo, S.W. Nitrogen improves plant cooling capacity under increased environmental temperature. Plant Soil 2022, 472, 329–344. [Google Scholar] [CrossRef]
- Treml, V.; Hejda, T.; Kašpar, J. Differences in growth between shrubs and trees: How does the stature of woody plants influence their ability to thrive in cold regions? Agric. For. Meteorol. 2019, 271, 54–63. [Google Scholar] [CrossRef]
- De Frenne, P.; Zellweger, F.; Rodríguez-Sánchez, F.; Scheffers, B.R.; Hylander, K.; Luoto, M.; Vellend, M.; Verheyen, K.; Lenoir, J. Global buffering of temperatures under forest canopies. Nat. Ecol. Evol. 2019, 3, 744–749. [Google Scholar] [CrossRef]
- Richter, R.; Ballasus, H.; Engelmann, R.A.; Zielhofer, C.; Sanaei, A.; Wirth, C. Tree species matter for forest microclimate regulation during the drought year 2018: disentangling environmental drivers and biotic drivers. Sci. Rep. 2022, 12, 17559. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhu, Z.; Zeng, H.; Myneni, R.B.; Zhang, Y.; Peñuelas, J.; Piao, S. Seasonal peak photosynthesis is hindered by late canopy development in northern ecosystems. Nat. Plants 2022, 8, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Niu, C.; Liu, X.; Hu, T.; Feng, Y.; Zhao, Y.; Liu, S.; Liu, Z.; Dai, G.; Zhang, Y. , et al. Canopy structure regulates autumn phenology by mediating the microclimate in temperate forests. Nat. Clim. Change 2024. [Google Scholar] [CrossRef]
- Huang, J.; Kong, F.; Yin, H.; Middel, A.; Liu, H.; Zheng, X.; Wen, Z.; Wang, D. Transpirational cooling and physiological responses of trees to heat. Agric. For. Meteorol. 2022, 320, 108940. [Google Scholar] [CrossRef]
- Fang, Z.; Zhang, W.; Wang, L.; Schurgers, G.; Ciais, P.; Peñuelas, J.; Brandt, M.; Yang, H.; Huang, K.; Shen, Q. , et al. Global increase in the optimal temperature for the productivity of terrestrial ecosystems. COMMUN EARTH ENVIRON 2024, 5, 466. [Google Scholar] [CrossRef]
- Khan, A.; Yan, L.; Mahadi Hasan, M.; Wang, W.; Xu, K.; Zou, G.; Liu, X.-D.; Fang, X.-W. Leaf traits and leaf nitrogen shift photosynthesis adaptive strategies among functional groups and diverse biomes. Ecol. Indicators 2022, 141, 109098. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, N.; Wei, T.; Liu, B.; Shen, L.; Liu, Y.; Liu, D. Adaptation strategies of leaf traits and leaf economic spectrum of two urban garden plants in China. BMC Plant Biol. 2023, 23, 274. [Google Scholar] [CrossRef]
- Miller, B.D.; Carter, K.R.; Reed, S.C.; Wood, T.E.; Cavaleri, M.A. Only sun-lit leaves of the uppermost canopy exceed both air temperature and photosynthetic thermal optima in a wet tropical forest. Agric. For. Meteorol. 2021, 301-302, 108347. [Google Scholar] [CrossRef]
- Xiong, X.; Wu, H.; Wei, X.; Jiang, M. Contrasting temperature and light sensitivities of spring leaf phenology between understory shrubs and canopy trees: Implications for phenological escape. Agric. For. Meteorol. 2024, 355, 110144. [Google Scholar] [CrossRef]
- Chu, T.; Li, J.; Zhao, J.; Gu, C.; Mumtaz, F.; Dong, Y.; Zhang, H.; Liu, Q. Regional Analysis of Dominant Factors Influencing Leaf Chlorophyll Content in Complex Terrain Regions Using a Geographic Statistical Model. Remote Sens. 2024, 16, 479. [Google Scholar] [CrossRef]
- Xu, M.; Chen, J.M.; Liu, Y.; Wang, R.; Shang, R.; Leng, J.; Shu, L.; Liu, J.; Liu, R.; Liu, Y.; et al. Comparative assessment of leaf photosynthetic capacity datasets for estimating terrestrial gross primary productivity. Sci. Total Environ. 2024, 926, 171400. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Nakamura, Y.; Furbank, R.T.; Evans, J.R. Effect of leaf temperature on the estimation of photosynthetic and other traits of wheat leaves from hyperspectral reflectance. J. Exp. Bot. 2021, 72, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Gratani, L.; Varone, L. Adaptive photosynthetic strategies of the Mediterranean maquis species according to their origin. Photosynthetica 2004, 42, 551–558. [Google Scholar] [CrossRef]
- Barai, K.; Calderwood, L.; Wallhead, M.; Vanhanen, H.; Hall, B.; Drummond, F.; Zhang, Y.J. High Variation in Yield among Wild Blueberry Genotypes: Can Yield Be Predicted by Leaf and Stem Functional Traits? AGRONOMY-BASEL 2022, 12, 18. [Google Scholar] [CrossRef]
- Knight, C.A.; Ackerly, D.D. Evolution and plasticity of photosynthetic thermal tolerance, specific leaf area and leaf size: congeneric species from desert and coastal environments. New Phytol. 2003, 160, 337–347. [Google Scholar] [CrossRef]
- Nóia, R.D.; do Amaral, G.C.; Pezzopane, J.E.M.; Toledo, J.V.; Xavier, T.M.T. Ecophysiology of C3 and C4 plants in terms of responses to extreme soil temperatures. Theor. Exp. Plant Physiol. 2018, 30, 261–274. [Google Scholar] [CrossRef]
- Li, N.N.; Li, J.H.; Shi, X.J.; Shi, F.; Tian, Y.; Wang, J.; Hao, X.Z.; Luo, H.H.; Wang, Z.B. Increasing cotton lint yield and water use efficiency for subsurface drip irrigation without mulching. Front. Plant Sci. 2024, 15, 15. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Hardie, W.J.; Smith, J.P. Stomatal density of grapevine leaves (Vitis vinifera L.) responds to soil temperature and atmospheric carbon dioxide. Aust. J. Grape Wine Res. 2011, 17, 147–152. [Google Scholar] [CrossRef]
- Kilpeläinen, J.; Domisch, T.; Lehto, T.; Piirainen, S.; Silvennoinen, R.; Repo, T. Separating the effects of air and soil temperature on silver birch. Part I. Does soil temperature or resource competition determine the timing of root growth? Tree Physiol. 2022, 42, 2480–2501. [Google Scholar] [CrossRef]
- Bjerring Jensen, N.; Vrobel, O.; Akula Nageshbabu, N.; De Diego, N.; Tarkowski, P.; Ottosen, C.-O.; Zhou, R. Stomatal effects and ABA metabolism mediate differential regulation of leaf and flower cooling in tomato cultivars exposed to heat and drought stress. J. Exp. Bot. 2024, 75, 2156–2175. [Google Scholar] [CrossRef]
- Peer, L.A.; Dar, Z.A.; Lone, A.A.; Bhat, M.Y.; Ahamad, N. High temperature triggered plant responses from whole plant to cellular level. Plant Physiol Rep. 2020, 25, 611–626. [Google Scholar] [CrossRef]
- Sadok, W.; Lopez, J.R.; Smith, K.P. Transpiration increases under high temperature stress: potential mechanisms, trade-offs and prospects for crop resilience in a warming world. Plant, Cell Environ. 2021, 44, 2102–2116. [Google Scholar] [CrossRef] [PubMed]
- Cano-Ramirez, D.L.; Carmona-Salazar, L.; Morales-Cedillo, F.; Ramírez-Salcedo, J.; Cahoon, E.B.; Gavilanes-Ruíz, M. Plasma Membrane Fluidity: An Environment Thermal Detector in Plants. In Cells, 2021; Vol. 10.
- Jiang, Q.; Jia, L.; Wang, X.; Chen, W.; Xiong, D.; Chen, S.; Liu, X.; Yang, Z.; Yao, X.; Chen, T. , et al. Soil warming alters fine root lifespan, phenology, and architecture in a Cunninghamia lanceolata plantation. Agric. For. Meteorol. 2022, 327, 109201. [Google Scholar] [CrossRef]
Figure 1.
Linear regression between leaf temperature (Tleaf) and air temperature (Tair) in plants. (a) All species pooled together, (b) plants classified by growth form, (c) plants classified by season, and (d) plants classified by species. The slope of the dashed line is 1.
Figure 1.
Linear regression between leaf temperature (Tleaf) and air temperature (Tair) in plants. (a) All species pooled together, (b) plants classified by growth form, (c) plants classified by season, and (d) plants classified by species. The slope of the dashed line is 1.
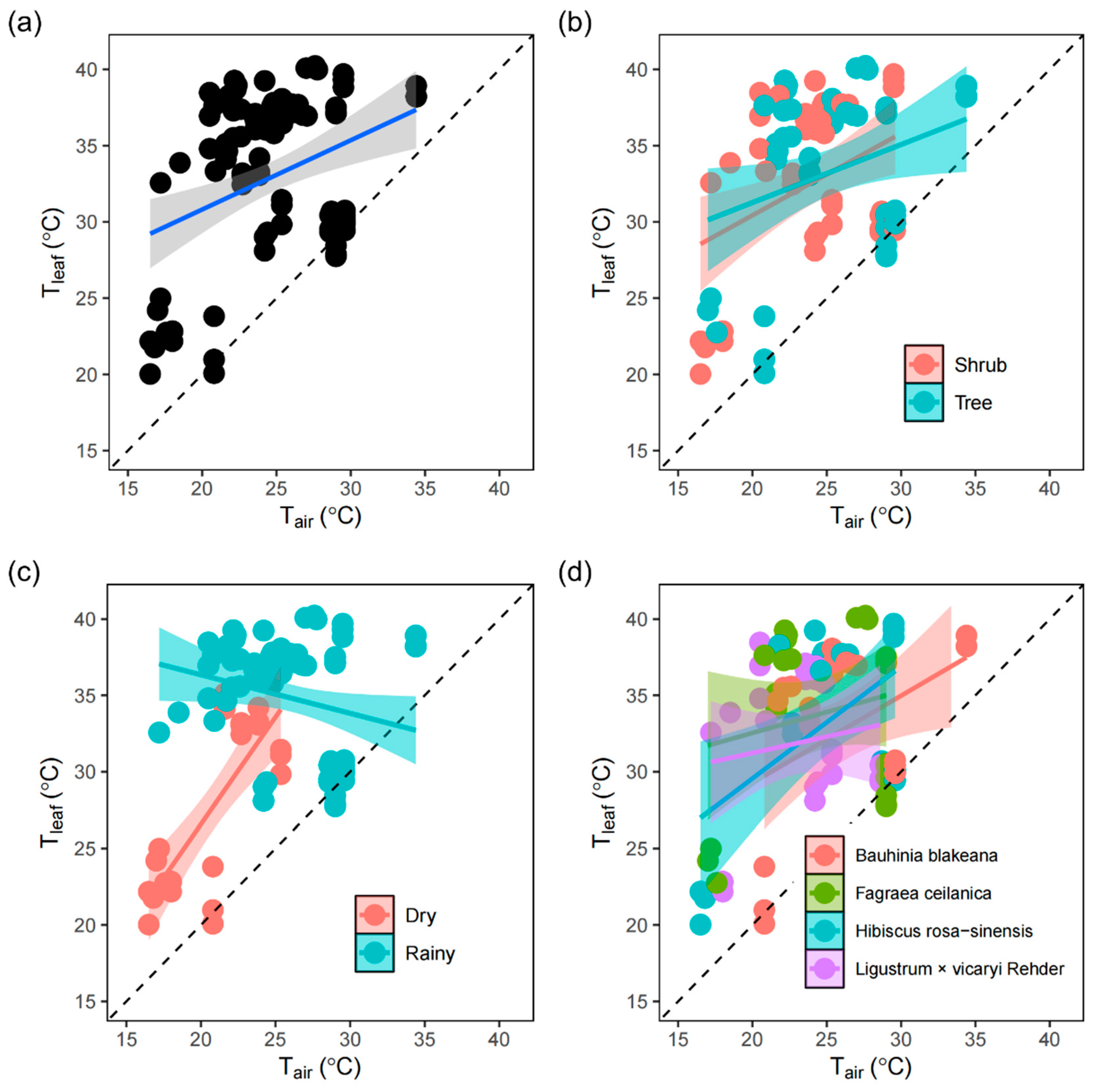
Figure 2.
Leaf traits between growth forms (shrub and tree) within dry and rainy seasons. *: P < 0.05, **: P < 0.01, ***: P < 0.001. Leaf chlorophyll content (SPAD, LCC), leaf area (cm2, LA), leaf dry mass (g, LDM), leaf temperature (°C, Tleaf), leaf mass per area (g m−2, LMA), and leaf water content (%, LWC) were measured.
Figure 2.
Leaf traits between growth forms (shrub and tree) within dry and rainy seasons. *: P < 0.05, **: P < 0.01, ***: P < 0.001. Leaf chlorophyll content (SPAD, LCC), leaf area (cm2, LA), leaf dry mass (g, LDM), leaf temperature (°C, Tleaf), leaf mass per area (g m−2, LMA), and leaf water content (%, LWC) were measured.
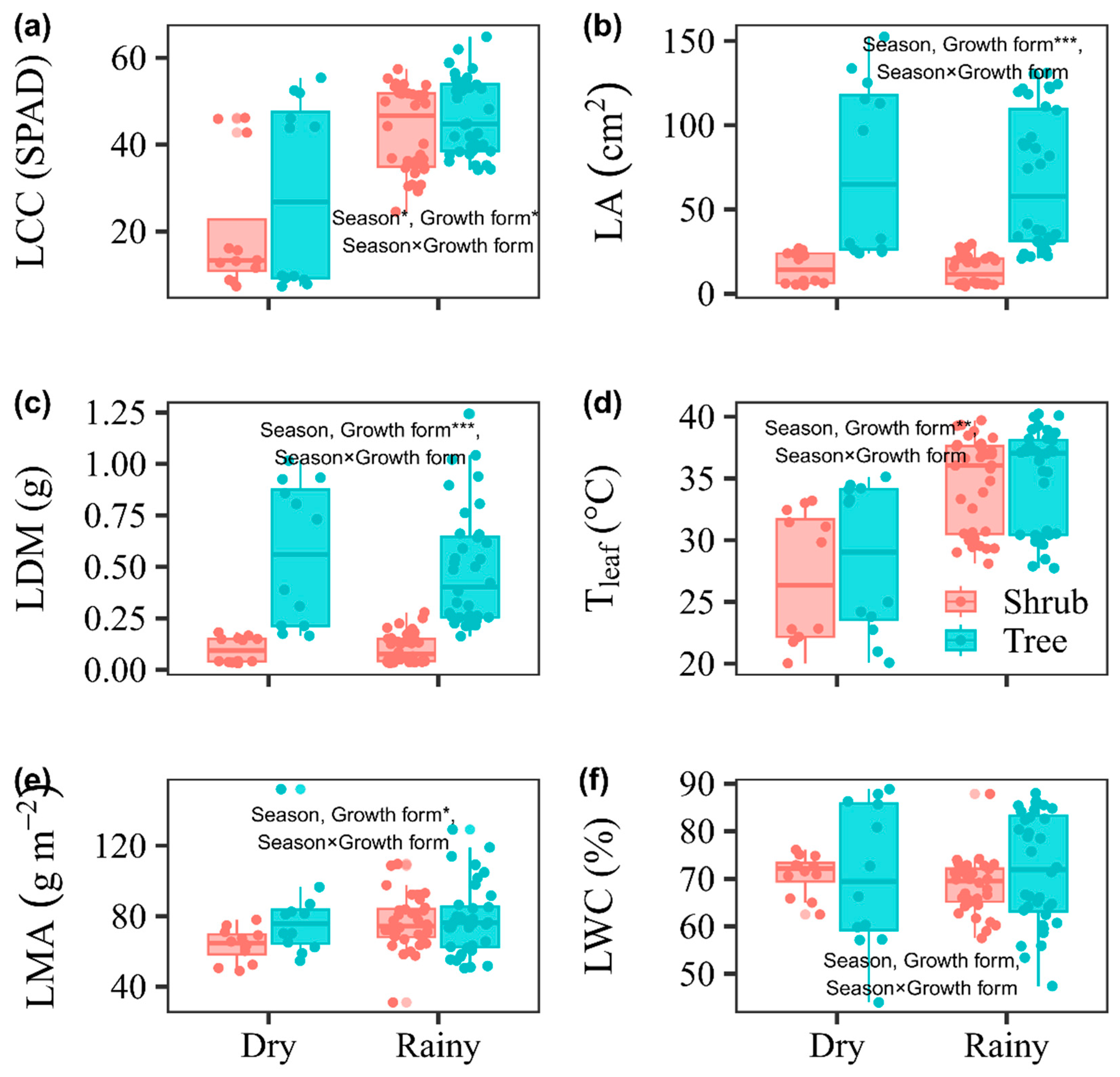
Figure 3.
Model-averaged importance of the predictors of leaf temperature (°C, Tleaf). The importance value is based on the sum of the Akaike weights derived from model selection via the corrected Akaike information criterion. The cutoff is set at 0.8 to differentiate among the most important predictors. Leaf chlorophyll content (SPAD, LCC), soil temperature (°C, Tsoil), leaf mass per area (g m−2, LMA), air temperature (°C, Tair), leaf dry mass (g, LDM), leaf water content (%, LWC), leaf area (cm2, LA), and soil water content (%, SWC) were measured.
Figure 3.
Model-averaged importance of the predictors of leaf temperature (°C, Tleaf). The importance value is based on the sum of the Akaike weights derived from model selection via the corrected Akaike information criterion. The cutoff is set at 0.8 to differentiate among the most important predictors. Leaf chlorophyll content (SPAD, LCC), soil temperature (°C, Tsoil), leaf mass per area (g m−2, LMA), air temperature (°C, Tair), leaf dry mass (g, LDM), leaf water content (%, LWC), leaf area (cm2, LA), and soil water content (%, SWC) were measured.
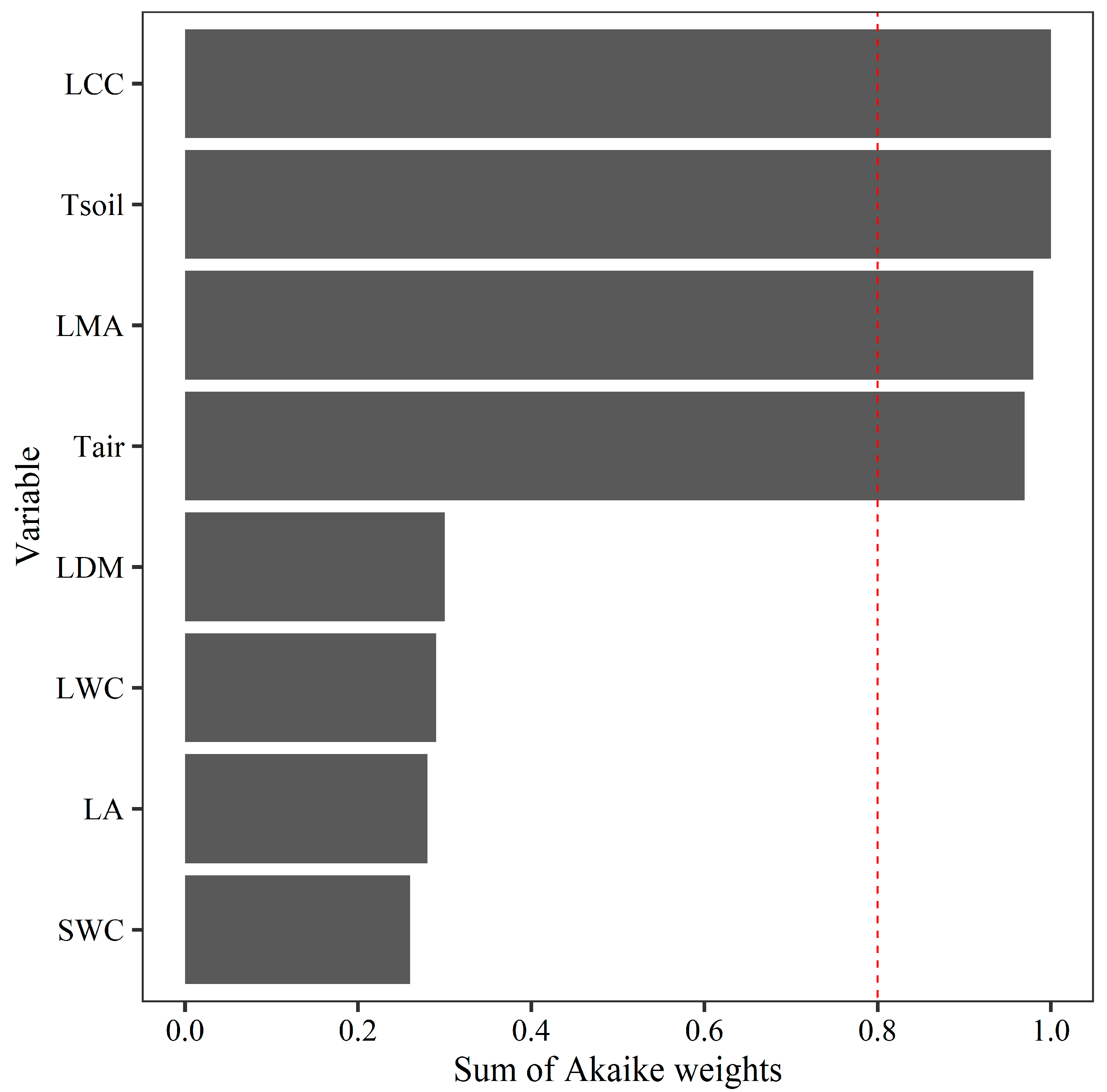
Figure 4.
Partial regression plots of the leaf chlorophyll content (SPAD, LCC), leaf mass per area (g m−2, LMA), soil temperature (°C, Tsoil), and air temperature (°C, Tair) in relation to the leaf temperature (°C, Tleaf) from multiple linear regression models once all other variables in the model were statistically controlled.
Figure 4.
Partial regression plots of the leaf chlorophyll content (SPAD, LCC), leaf mass per area (g m−2, LMA), soil temperature (°C, Tsoil), and air temperature (°C, Tair) in relation to the leaf temperature (°C, Tleaf) from multiple linear regression models once all other variables in the model were statistically controlled.
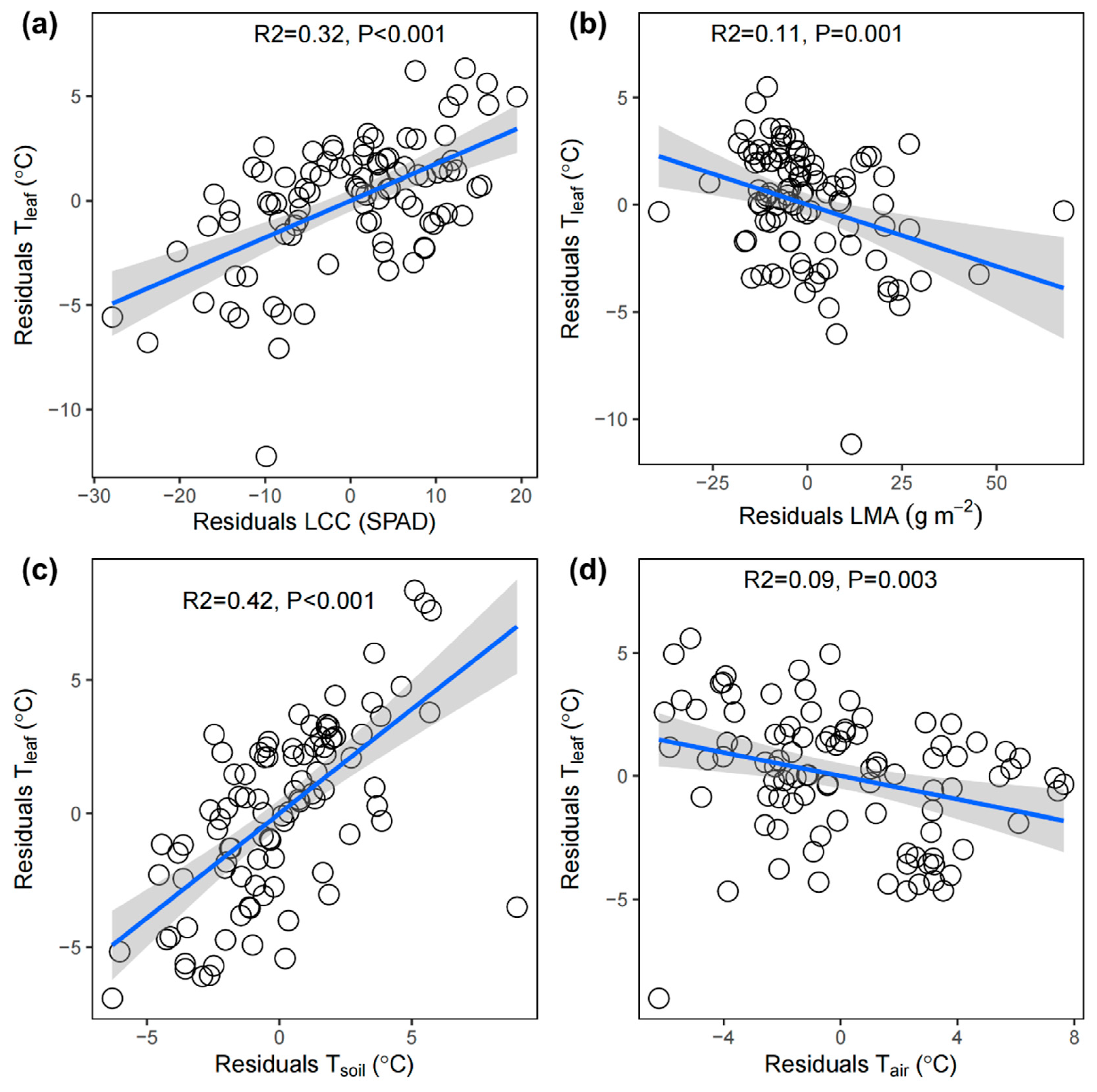

Table 1.
Summary of the linear regressions between leaf temperature (Tleaf) and air temperature (Tair) and the statistics (mean ± standard error) of Tleaf and Tair in plants of different growth types, seasons, and species. Different lowercase letters represent significant differences at P <0.05 for different growth types, seasons, and species.
Table 1.
Summary of the linear regressions between leaf temperature (Tleaf) and air temperature (Tair) and the statistics (mean ± standard error) of Tleaf and Tair in plants of different growth types, seasons, and species. Different lowercase letters represent significant differences at P <0.05 for different growth types, seasons, and species.
| Groups | Slope (95% CI) | R2 | P | Tleaf (°C) | Tair (°C) |
| All | 0.45 (0.21, 0.70) | 0.13 | <0.001 | 33.01±0.55 | 24.83±0.43 |
| Growth type | |||||
| Shrub | 0.54 (0.18. 0.90)a | 0.16 | 0.004 | 32.62±5.31 | 24.09±3.98 |
| Tree | 0.38 (0.03, 0.73)a | 0.09 | 0.035 | 33.39±5.40 | 25.57±4.32 |
| Season | |||||
| Dry | 1.40 (0.88, 1.92)a | 0.59 | <0.001 | 27.66±5.60 | 20.77±3.07 |
| Rainy | −0.25 (−0.50, −0.004)b | 0.06 | 0.047 | 34.79±3.89 | 26.18±3.63 |
| Species | |||||
| Hibiscus rosa-sinensis | 0.73 (0.22, 1.25)a | 0.28 | 0.008 | 33.39±5.82 | 25.22±4.21 |
| Bauhinia blakeana | 0.57 (0.07, 1.07)a | 0.21 | 0.026 | 32.96±5.33 | 26.46±4.23 |
| Ligustrum × vicaryi Rehder | 0.21 (−0.39, 0.81)a | 0.02 | >0.05 | 31.86±4.74 | 22.95±3.46 |
| Fagraea ceilanica | 0.27 (−0.28, 0.83)a | 0.05 | >0.05 | 33.82±5.55 | 24.68±4.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



