
Submitted:
08 November 2024
Posted:
08 November 2024
You are already at the latest version
Abstract
Fungi and soil bacteria are vital for organic matter decomposition and biogeochemical cycles, but excessive synthetic fertilizer use contributes to soil degradation and loss of biodiversity. Despite this, about 97% of soil microorganisms are unculturable, making them difficult to study. Metagenomics offers a solution, enabling the direct extraction of DNA from soil to uncover microbial diversity and functions. This study utilized metagenomics to analyze the rhizosphere of 2-year-old Tonda di Giffoni hazelnut saplings treated with synthetic NPK, composted olive pomace, and an innovative fertilizer derived from sulfur-based agro-industrial waste stabilized with bentonite clay. Using 16S rDNA for bacteria and ITS2 for fungi, Illumina sequencing provided insights into microbial responses to different fertilizer treatments. Results highlighted a significant increase in the abundance of beneficial microorganisms such as Thiobacillus, Pseudoxanthomonas, and Thermomyces, especially when organic material was included. Additionally, microbial biodiversity improved with organic inputs, as shown by increased species richness (Chao1) and diversity (Bray-Curtis). These findings emphasize the importance of organic fertilization in enhancing soil microbial health, offering a sustainable approach to improving soil quality and hazelnut productivity.
Keywords:
Organic Waste
; Sulfur
; Olive Pomace
; Metagenomics
; Biodiversity
; Microbiome
; Corylus Avellana
1. Introduction
Hazelnut (Corylus avellana L.), a member of the Betulaceae family, ranks as the second most widely grown nut in the world, following only almonds [1]. The main hub of hazelnut production is located along the Black Sea coast of Turkey, with other significant cultivation in Italy, United States and Spain [2]. To meet the growing consumer demand for hazelnuts and increase productivity, it is imperative to adopt strategies that ensure robust yields while preserving soil and the environment. Several studies have been conducted to improve the productivity and fertility of hazelnut cultivation [3,4,5] but as reported by Vincze et al [6], a better understanding of the role of microbes in agroecosystem functioning in the context of plant growth and soil fertility is critical for sustainable agricultural production. Plant nutrition is closely influenced by microbial community in the rhizosphere [7]. The soil rhizosphere is a micro-ecosystem in which the composition and structure of microorganisms can influence nutrient transformations in the soil, nutrient uptake by plants, and thus plant growth and development [8]. According to Roeland et al [9], the rhizosphere functions as a unique region where the interaction between plant, root and soil microbiome takes place; in fact, as stated by Philippot et al [10], the rhizosphere is one of the most dynamic interfaces on Earth.
In recent years, in response to climate change and the widespread use of synthetic/mineral fertilizers, the effects of different fertilization techniques on the rhizosphere microbial community have begun to be studied [7,11,12].
A study by Chavez-Romero et al [13] states that the application of organic fertilizers has beneficial effects compared to inorganic fertilizers in promoting the diversity of soil microbial communities and enriching the soil with organic carbon, nitrogen and other nutrients. Dai X.B et al [14], observed that the increased application of synthetic nitrogen and phosphate fertilizers changed the composition of the soil microbial community by reducing the abundance of gram-positive bacteria such as actinomycetes. Guo et al [15], found that synthetic fertilization negatively influenced microbial communities in the rhizosphere, and that positive effects on soil microbial communities were observed with addition of organic fertilizer compared to chemical fertilization [16]. Similarly, Legrand et al [17], found greater bacterial richness and uniformity with manure application compared to chemical fertilization. In contrast, Orr et al [18], observed no discernible influence on the microbial community when comparing conventional and organic farming systems. The composition and biodiversity of the observed microbiota were attributed exclusively to environmental and soil chemical variables.
Although several previous studies have focused mainly on long-term field experiments, as stated by, organic fertilizers can have an action even in the short term, within a single growing season, especially in crop productivity [19,20]. Nevertheless, no study has yet been conducted on the evaluation of mineral and organic synthetic fertilizers on hazelnut saplings in the short term and specifically on the influence of these fertilizations on rhizosphere microbial processes. Studies that have employed culture-based methods [21,22] capture only a limited fraction of the overall microbial diversity. In contrast, next-generation sequencing (NGS) techniques offer a powerful approach for achieving a more comprehensive understanding of microbial community composition and diversity [23].
The aim of this study is to evaluate the short-term changes in microbial diversity and community in the rhizosphere of hazelnut saplings. The rhizosphere microbial community of unfertilized hazelnut saplings was compared with the rhizosphere microbial community of hazelnut saplings fertilized with synthetic commercial NPK fertilizer (NPK), composted olive pomace (OP), sulfur bentonite (SB) and sulfur bentonite + composted olive pomace (SBOP).
SB and SBOP are new fertilizers obtained from agro-industrial wastes composed as follows: SB is comprised of elemental sulfur (S) residue from the hydrocarbon refining process stabilized with bentonite clay, without composted olive pomace; SBOP is comprised of elemental S residue from the hydrocarbon refining process stabilized with bentonite clay, with the addition of composted olive pomace. The effectiveness of Sulfur-based fertilizers, both with and without an organic component, has been demonstrated in different studies [25] but no studies up to now have been carried on tree species. This study aims to verify if fertilization yields different outcomes compared to the previously studied horticultural plants. Advanced techniques are needed to study the effects of different fertilizations on soil microbiome, given that about 97 percent of bacterial species cannot be cultivated due to the lack of knowledge of different microorganism cultivation requirements as nutritional needs, natural environment physiochemical conditions, and the symbiotic or parasitic relationships that are maintained in a microbial community [26].
2. Materials and Methods
2.1. Fertilizers Manufacturing
The olive pomace compost used in this study was produced at the farm composting plant of Nuovo Cilento agricultural company, in San Mauro Cilento (SA), Italy.
The chemical characteristics of olive pomace compost are reported in Table 1 and analyzed according to Muscolo et al [24]. The synthetic fertilizers were produced by Steel Belt System s.r.l and the process of manufacturing is reported in Panuccio et al [27]. The composition of different fertilizers is reported in Table 2.
2.2. Experimental Design
In February 2023, 2 years old hazelnut saplings var. Tonda di Giffoni were placed in 30 litre capacity pots filled with a sandy loam soil (64% sand, 28% loam, 8% clay) according to Agricultural Organization of the United Nations (FAO) [28]. The soil contained an organic matter content of 20 g/kg, a total nitrogen concentration of 1 g/kg, and a carbon-to-nitrogen ratio of 12.
Four replications were made per treatment, for a total of 20 pots following here doses of each treatment per pot:
- 45 g of composted olive pomace (OP)
- 5 g of Sulfur Bentonite + Olive Pomace (SBOP)
- 5 g of Sulfur Bentonite (SB)
- 4.2 g of synthetic fertilizer (NPK).
- Unfertilized soil (CTR)
For each treatment fertilizer was applied twice, before flowering in February and before vegetative growth in June. To limit weed development and avoid excessive soil overheating, the pots were isolated by placing them in larger-diameter containers (50 cm), and the surface soil and defined chamber was filled with expanded clay (Figure 1). Soil water content, through irrigation with potable water, was always maintained at 70% of field capacity.
2.3. Samples Collection and DNA Extraction
In July rhizosphere sampling was performed using the protocol of Simmon et al [29], with some modifications: 2 mm of soil attached to the roots were left and a representative section were cut and placed in 50ml tube with 25 ml of phosphoric buffer, centrifuged for 10 minutes at 4,000 g at 4 °C and root were removed to obtained rhizosphere fraction. Rhizosphere DNA was extracted using a commercial kit (QIAGEN DNeasy PowerSoil Pro kit) following the protocol with one modification, 10 mg of skim milk has been added to all the samples as suggested by Hoshino et al [30] to increase DNA extraction yield from volcanic soil. The final DNA concentrations were determined by NanoDrop 2000 UV-vis spectrophotometer (Thermo Scientific, Wilmington, USA), and the DNA quality was checked by 1% agarose gel electrophoresis on a Luminescence Image Analyzer System (LAS 4000; ImageQuant, Life sciences) (Figure 2).
DNA samples were screened for the V3-V4 hypervariable regions by PCR using primers 341F (5’-GCGGTAATTCCAGCTCCAA – 3’) and 806R (5’ -GGACTACNNGGGTATCTAAT) for 16S rDNA. For ITS2 using the primers ITS3-2024F (5’-GCATCGATGAAGAACGCAGC-3’) and ITS4 -2409R (5’-TCCTCCGCTTATTGATATGC-3’) [31]. All PCR reactions were carried out with 15 μL of Phusion® High -Fidelity PCR Master Mix (New England Biolabs); 2 μM of forward and reverse primers, and about 10 ng template DNA. Thermal cycling consisted of initial denaturation at 98°C for 1 min, followed by 30 cycles of denaturation at 98°C for 10 s, annealing at 50°C for 30 s, and elongation at 72°C for 30 s. Sequencing libraries were generated using TruSeq® DNA PCR- Free Sample Preparation Kit (Illumina, San Diego, CA, USA) following manufacturer's recommendations and index codes were added. The library quality was assessed on the Qubit@ 2.0 Fluorometer (Thermo Scientific) and Agilent Bioanalyzer 2100 system. At last, the library was sequenced on an Illumina NovaSeq 6000 platform (Illumina, San Diego, CA, USA) and 250 bp paired-end reads were generated. The DNA library preparation and sequencing were performed by Novogene Co, Ltd (Beijing, China, http://www.novogene.com/). The raw sequence files generated (fastq files) underwent quality control analysis with FastQC.
2.4. Data Analysis
The NGS datasets were analysed using the EBI Metagenomics service pipeline, which provides quality control, taxonomic analysis based on SSU rDNA sequences, and sequence assembly. (MGnify. 2023. “Analysis Pipeline V5.0.” November 9, 2023. https://docs.mgnify.org/src/docs/analysis.html)
2.5. Biodiversity Assessment and Statistical Analysis
Sequence data from the rhizosphere soil community was analyzed using Microbiome Analyst (https://www.microbiomeanalyst.ca/) [32]. Microbial biodiversity was assessed for quantify differences between groups at two levels: within-samples (alpha-diversity) and between samples (beta-diversity) [33]. From the different six measures supported in Microbiome Analyst described by Chong et al [34], Chao1 index [35] and Shannon index [36,37] were selected. From the five different beta-diversity indexes supported by Microbiome Analyst, “Bray-Curtis dissimilarity” [38] was utilized. The index measures the compositional dissimilarly between the microbial communities based on counts of each samples. Microbiome Analyst can measure Beta-diversity using PCoA or nonmetric multidimensional scaling (NMDS); for this study PCoA was selected because it maximizes the linear correlation between samples [34]. For identifying microbial taxa that were significantly different between groups, LEfSe (Linear discriminant analysis Effect Size), a non-parametric statistical method, was selected. A significance level of P < 0.05 and LogLDA score of ± 2 were applied. The jveen tool [39] was used to compare the genera of fungi and bacteria that were found to be statistically significant using LeFSE. ‘Class’ level was selected for bacteria and fungi for generating the Heatmap outputs. The distance between data points in the clustering input is measured with the standard Euclidean (as-the-crow-files) distances. All sequences were deposited at European Nucleotide Archive (ENA, http://www.ebi.ac.uk/ena) under project number PRJEB70816 for bacteria and PRJEB68325 for fungi.
3. Results
Sequencing generated a total of 1656509 (16S rDNA) and 1749133 (ITS2) sequences, respectively, for 20 samples, classified into 1202 OTUs for the 16S rDNA and 1642 OTUs for the ITS2.
3.1 Bacteria Taxa Abundance
Results showed that amongst the 10 most abundant bacterial phyla, Actinobacteria were the most dominant, constituting 46% in rhizosphere soil treated with OP, 45% in control soil, 43% in SBOP-treated soil, 41% in soil with NPK, and 39% in SB-treated soil. (Figure 3a).
The second most abundant phylum were the Proteobacteria: representing 23% in SBOP treated soil, 22% in soil with OP, 21% with NPK, 20% in CTR soil, and 18% in SB treated soil. Acidobacteria increased in all treated soils, compared to CTR (7%), especially with SB (10%).
About 76% of the genus classification was not identified. The genus Bacillus was most prevalent in CTR control plants (3%) but decreased with SB (2%) and SBOP (2%) and accounted for only 1 % in NPK and OP treatments. Contrastingly, the genus Streptomyces, was present at an equal percentage of 3% in all treatments except for SB-treated plants where it decreased to 2%. (Figure 3 b).
3.2. Fungi Taxa Abundance
Taxonomic classification showed that the most abundant phylum among the samples was Ascomycota. The unfertilized CTR soils had the highest percentage of Ascomycota fungi (73%), followed by SB treatment (69%), SBOP (59%), NPK (59%), OP (55%). Fungi belonging to the phylum Streptophyta showed an increase compared to the control (19%) across all treatments: SB (25%), NPK (29%), OP (29%), and SBOP (30%). Fungi from the phylum Basidiomycota were predominantly found in the OP treatment (9%). (Fig 4a)
About 76% of the genus classification was not identified. The most abundant genus for all samples was Talaromycetes, especially in those treated with SB (13%). This relative abundance decreased in soils fertilized with NPK (7%), SBOP (6%), OP (5%) and in control soil (4%). (Figure 4.b)
3.3. Bacteria Alpha and Beta Diversity
Species richness calculated using the Chao1 index for bacteria OTUs showed that, all treatments resulted in an increase compared to control plants (without fertilization) (Fig. 5a). Specifically, the largest increase was observed in plants treated with sulfur bentonite + olive pomace fertilizer (SBOP), followed by those treated with olive pomace (OP). Treatment with the synthetic fertilizer (NPK) also resulted in a statistically significant increase, while a smaller increase was noted for sulfur bentonite (SB) treatment. (Fig 5a). The results of the post-hoc pairwise comparison (multiple-group only) revealed statistically significant differences (p < 0.05) between several treatments. Specifically, there was a significant difference between OP and NPK (p = 0.034), OP and SB (p = 0.039), and CTR and OP (p = 0.039). The differences in community composition between samples calculated with Bray-Curtis index showed a dissimilarity of 35.6% for axis 1 and a dissimilarity of 16.2 % for axis 2. (Fig 5c). Although no clear clustering pattern was shown by using unsupervised PCoA on Bray-Curtis dissimilarity, the Principal coordinate 2 highlights a trend along the sample distribution, mainly driven by the use of compost on one side and mineral fertilization and sulfur use on the other side. From the result of pairwise PERMANOVA analysis and the multi-testing adjustment, based on Benjamini-Hochberg procedure procedure (FDR), a statistically significant difference for p value < 0.05 can be seen between the CTR vs NPK treatments with a p value of 0.037 and OP vs NPK with a p value of 0.046 (Table 1 Supplementary Material).
3.4 Fungi Alpha and Beta Diversity
Species richness, calculated using the Chao1 index for fungal OTUs, demonstrated an increase in all treatments compared to control plants (without fertilization). Specifically, the Olive pomace (OP) treatment exhibited the greatest increase in this index, followed by sulfur bentonite + olive pomace (SBOP) and then the synthetic fertilizer (NPK) (Fig. 5b) (Figure 5b).
From the results of post-hoc pairwise comparison (multiple-group only), statistically significant differences (p < 0.05) were observed between several treatments. These included OP vs CTR (p = 0.0033), SBOP vs CTR (p = 0.0073), OP vs SB (p = 0.0079), and SBOP vs SB (p = 0.018).
The dissimilarity in community composition between samples calculated with Bray-Curtis dissimilarly showed a dissimilarity of 29.5% for axis 1 and a dissimilarly of 19.4 for axis 2. (Fig 5c). Although no clear clustering pattern is shown by using unsupervised PCoA on Bray-Curtis index, the second principal coordinate 2 highlights a trend along the samples distribution mainly driven by the use of compost on one side and mineral fertilization and sulfur use on the other side. From the result of pairwise PERMANOVA analysis and the multi-testing adjustment based on Benjamini-Hochberg procedure procedure (FDR), a statistically significant difference for p value < 0.05 can be seen between treatments of OP vs NPK with a p value of 0.032 and OP vs CTR with a p value of 0.047 (Table2 Supplementary Material).
3.5. Comparison Analysis with LEfSe, Heatmap of Relative Abundance and Veen Diagram
For Bacteria, the linear discriminant analysis Effect size (LEfSe) to feature level with p-value cutoff 0.05 and LogLDA score 2.0 identified a total of 44 significant features. (Table. 3 Supplementary materials). There were 10 significant genera observed (Figure 6a).
From the Venn diagram analysis, bacteria belonging to the Pseudonocardia, Phaselicystis, Hirschia, Paludibaculum, Nitrosospira, Anaeromyxobacter, Adhaeribacter, and Reyranella were found to be common across all treatments and the unfertilized soil (CTR) (Figure 7). The genus Thiobacillus was common to treatments with OP, SB, and SBOP. In contrast, the genus Pseudoxanthomonas was found to be common between treatments with OP and SBOP, as well as in the unfertilized soil (CTR) (Figure 7).
Histogram of the LDA scores computed for features differentially abundant on different treatment. LEfSe scores can be interpreted as the degree of consistent difference in relative abundance between features of analyzed bacterial and fungal communities (Figure 6). The histogram thus identifies which clades among all those detected as statistically and biologically differential explain the greatest differences between genus communities.
The genus Pseudonocardia appeared more abundant in the OP treatment, with no significant differences observed compared to other treatments. Meanwhile, the genus Thiobacillus was notably absent in both control soil and NPK-fertilized plants but showed an increase with all other treatments, particularly with SBOP. Hirschia, Paludibaculum, and Reyranella genera increased in abundance when organic components were added to fertilizers, notably in SBOP and OP treatments. Conversely, the genus Pseudoxanthomonas showed an increase compared to the control in OP and SBOP treatments, while decreased in plants treated with NPK and SB. Nitrosospira bacteria were more abundant in NPK-treated soil. However, bacteria belonging to the genus Anaeromyxobacter decreased in OP and SBOP treatments. Adhaeribacter bacteria decreased across all treatments, with a significant reduction observed in mineral treatments with SB and NPK (Figure 7).
For Fungi, the linear discriminant analysis Effect size (LEfSe) was employed up to the taxonomy level feature level, with a p-value cutoff of 0.05 and a LogLDA score of 2.0, resulting in a total of 28 significant features (Table 4, Supplementary Materials). Among these, 12 significant genera were identified (Figure 6b).
From the Venn diagram analysis (Figure 8), it was observed that the fungi genera Podospora, Chaetomium, and Berkleasmium are common across all treatments and the unfertilized soil (CTR). Genera Thermomyces, Pseudoallescheria, and Peziza were common across all soils treated with fertilizers. The genus Malbranchea was common to the unfertilized soil (CTR), OP, SB, and SBOP treatments but not in NPK treatment. On the other hand, the genus Veronae was common to the unfertilized soil (CTR) and all other treatments except SB. Soil treatments with NPK, OP, and SBOP shared the genera Monascus and Pleuroteciella. Only treatments with OP and SBOP shared the genera Tritirachium and Dactylella. Fungi of the genus Pseudoallescheria, Thermomyces, and Podospora were predominantly in soil with organic components, such as OP and SBOP fertilizers.
Conversely, fungi of the genus Chaetomium and Berkleasmium behaved consistently across OP, SB and SBOP fertilizer inputs, decreasing compared to soil without fertilizer (CTR) and NPK synthetic fertilizer.
Meanwhile, fungi belonging to the genus Malbranchea and Peziza showed a significant increase in soils fertilized with SB compared to all other treatments. In contrast, fungi of the genus Veronae decreased in treatments with OP and SB compared to unfertilized soil (CTR) and NPK-fertilized soil but increased in soil fertilized with SBOP. (Figure 8).
Heatmaps representing bacterial diversity at a class level (Figure 9) indicated that SBOP treatment showed high abundance for Gammaproteobacteria and Bacilli, suggesting that this treatment created a favourable environment for the proliferation of these classes. In contrast, SB treatment showed a low abundance for Alphaproteobacterial and Actinobacteria. Treatment with SB varied between replicates, showing a less uniform response than the other treatments. OP treatment appears less favourable for the proliferation of Acidobacteria and Anaerolineae, indicating a possible negative impact of OP treatment on this class of bacteria. The NPK treatment showed a variable abundance especially on the classes Betaproteobacteria and Clostridia. Soils without treatment (CTR) showed that the abundance of the bacterial classes is generally moderate to low, especially for the classes Planctomycetacia and Verrucomicrobiae. The dendrogram shows how the bacterial classes are grouped according to their abundance. Classes such as Gemmaproteobacteria and Bacilli tend to cluster together, indicating a similar response to treatments. In contrast, the upper dendrogram shows how samples with similar abundance patterns cluster together. This clustering confirms that SBOP-treated samples tend to have more similar abundance profiles than other treatments.
Heatmap of fungi at class level (Figure 10) shows that the SBOP treatment exhibits high abundance for several taxa, in particular this treatment could promote the growth of fungi belonging to these groups, suggesting a favourable environment for the proliferation of Dothideomycetes and Eurotiomycetes. The SB treatment shows a variable abundance pattern, with moderate Saccharomycetes and higher abundance of Sordariomycetes. The effect of SB treatment varied between replicates, suggesting a less uniform response than other treatments. OP treatment generally shows low abundance for many taxa, especially in Geoglossomycetes and Leotiomycetes. The NPK treatment shows considerable variation in the abundance of taxa: High abundance of Orbiliomycetes and low abundance of Sordariomycetes. The CTR treatment shows generally lower abundance for several taxa: Agaricomycetes and Trebouxiophyceae, comparated with OP and SBOP. The dendrogram shows how taxa are grouped according to their abundance. Taxa Dothideomycetes Eurotiomycetes tend to cluster together, indicating a similar response to treatments.
4. Discussion
The combined application of chemical and organic fertilizers has previously been documented to offer several advantages over synthetic fertilizers, including a balanced nutrient supply and a reduction in the environmental risks associated with excessive usage [40,41]. While some studies [42,43,44] suggest that short-term effects from applying organic fertilizers may not significantly impact microbial richness due to competition between microbes in bio-organic fertilizers and the existing local bacterial community, others present a contrasting view. For instance, Tian et al [44] reported a decrease in soil bacterial diversity following organic fertilizer application, whereas others study [45,46] highlighted the potential of organic fertilizers to enhance the soil microbiome, promoting bacterial growth and increasing population diversity.
Our study demonstrated that the combined fertilizer treatment, composed of organic, composted olive pomace (including a manure component) and sulfur and bentonite minerals, enhanced the microbial abundance.in a similar manner to organic fertilizer (OP), rather than to sulfur-bentonite fertilizer (SB) alone
Our research indicates that adding just 5 % of organic material to sulfur-based mineral fertilizer can leverage beneficial increases in various diverse bacteria and fungi, offering a potential improvement in soil quality.
This observation agrees with the study conducted by Marra et al [47] who demonstrated that bacterial communities were influenced by a sulfur-bentonite-based fertilizer combined with organic components (orange waste) in alkaline soils, at the same application rates used in our study, whilst increases in fungal communities were correlated with an organic fertilizer (horse manure), and actinomycetes were linked to NPK treatments. A recent study by Maffia et al [48] further showed that compost derived from olive pomace, similar in composition to the one used in this study, significantly increased the number of actinomycete, fungal, and other bacterial taxa compared to the control and other treatments. However, these studies relied on traditional microbial counting methods, which have inherent limitations, such as selective culturing and under-representation of non-culturable microbes. Despite the positive increase in microbial abundance/diversity with 5 % organic matter, the optimal fertilization rate at which to achieve maximum increases still requires further experimental verification. For example Han et al [49] found that 10-30 % organic fertilizer significantly increased bacterial diversity in maize, while other argue for a higher percentage [50].
4.1. Effect of Fertilization on Microbial Composition and Biodiversity
At the phylum level, we observed that Actinobacteria and Proteobacteria were the dominant bacterial groups in the rhizosphere of Corylus avellana across all treatments. The increased abundance of Actinobacteria in the olive pomace compost (OP) treatment is noteworthy, as this bacterial phylum plays a key role in soil organic matter turnover and the breakdown of complex molecules such as cellulose and polycyclic aromatic hydrocarbons [51,52]. Similarly, the increased presence of Proteobacteria in the SBOP and OP treatments may be explained by the rise in soil carbon content following the application of organic amendments, as observed in previous studies [53]. As α-, β-, and γ-Proteobacteria are classified as "copiotrophs" that utilize labile carbon for growth, they tend to thrive in nutrient-rich environments [54,55], such as those in the OP and SBOP treatments.
Interestingly, an increase in Acidobacteria was also observed across all treatments, particularly in the SB treatment. According to Kalam et al [56], these bacteria possess genes that enable them to survive in and competitively colonize the rhizosphere, fostering beneficial relationships with plants. Additionally, Acidobacteria are equipped with genes that enable them to metabolize both inorganic and organic nitrogen sources, effectively reducing nitrates, nitrites, and nitric oxide [57].
While Firmicutes were identified as the fifth most abundant phylum, the genus Bacillus—a member of Firmicutes—was the most dominant genus across treatments. Bacillus is a highly adaptable bacterium capable of surviving adverse environmental conditions by forming spores and degrading organic materials such as cellulose. Though Firmicutes as a whole may be less abundant relative to other phyla, the dominance of Bacillus is consistent with its ability to thrive in diverse soil environments [58]. However, while several studies have shown that Bacillus abundance increases with the application of organic fertilizers [59,60], our results align with Wu J et al [61], who observed a sharp decline in Bacillus relative abundance in fertilized soils compared to unfertilized soils.
Regarding fungi, Ascomycota and Streptophyta were the dominant phyla in the rhizosphere of Corylus avellana across all treatments. Our study showed a lower relative abundance of Ascomycota in most treatments compared to the control (CTR). This finding is in line with a recent study by Sivojienė et al [61], which also noted a reduction in the abundance of Ascomycota following organic fertilizer application, such as poultry manure. This could have been attributed to the more efficient competition for resources by other microorganisms or their better adaptation to new soil chemical conditions, such as fungi from the genus Talaromyces. This genus was particularly abundant in the OP treatment. A similar study reported a negative correlation between Basidiomycota abundance and high levels of total nitrogen (TN), soil organic carbon (SOC), and soil moisture (SM), but a positive correlation with aromatic substance availability (SA) [63]. This suggests that Basidiomycota may thrive in soils with high carbon and nitrogen content but where aromatic substances are more prevalent, as is often the case with organic fertilizers.
The second most abundant fungal phylum was Streptophyta, a group of land plants and algae that may compete with crops for nitrogen [64,65,66], potentially coming from contamination of the water used for this field experiment. Considering the competitive role of Streptophyta in nutrient uptake, particularly nitrogen, the SB treatment was observed to be associated with the smallest increase in Streptophyta abundance (25%) so could therefore be considered the most favorable treatment for minimizing competition between crops and this phylum.
Fungi belonging to the genus Talaromyces were notable for their role as primary decomposers of plant residues and as antagonists towards other fungi [67]. Some species of Talaromyces secrete organic acids and phosphatase, aiding in the dissolution of inorganic calcium phosphate and phosphate ester, which promotes phosphorus uptake by plants [68,69]. In our study, the SB treatment demonstrated the highest response in Talaromyces abundance compared to the control. This may have been due to the soil acidification induced by sulfur, which creates a favorable environment for Talaromyces, however additional assessments of soil pH would be required to verify this. Additionally, bentonite present in SB treatments could have contributed to improvements in soil structure, moisture retention, and increases in phosphorus availability, thereby enhancing the growth-promoting effects of Talaromyces on plants.
The increase in alpha-diversity, particularly bacterial species richness as measured by the Chao1 index, in soils treated with sulfur bentonite (SB) and organic sulfur bentonite (SBOP), is consistent with the findings of Damo et al [79] who reported that sulfur applications significantly increased both microbial abundance and diversity compared to sulfur-free treatments. Notably, when comparing SB and SBOP, the organic component of compost in the SBOP treatment plays a critical role in further enhancing species richness for both bacteria and fungi, even in short-term treatments. This observation aligns with other studies [71,72] which emphasize the pivotal role of organic matter in fostering microbial biodiversity.
Alpha diversity, as quantified through the Chao1 index, reflects the richness or the number of distinct species in the soil microbial community. Our results show that the inclusion of even a small percentage (5%) of organic material in the SBOP treatment resulted in a significant increase in fungal richness compared to the control (CTR), underscoring the importance of organic components in soil biodiversity. These results corroborate the findings of Hu et al [43] who demonstrated that the use of organic fertilizers can significantly enhance microbial composition even after short-term application. This suggests that the composition and quality of organic matter are key factors in promoting soil alpha-diversity, a conclusion also supported by Guo et al [73]
In terms of beta-diversity, which refers to the differences in community composition between treatments, the Bray-Curtis dissimilarity index highlighted the strong influence of fertilizer type on microbial community structure. Organic amendments, such as compost (OP) or sulfur bentonite, supported the establishment of distinct microbial communities compared to mineral fertilizers like NPK. This differentiation suggests that the organic matter, in combination with other components like sulfur, significantly reshapes soil microbial populations, which is a phenomenon widely observed in soil ecology [43]. However, the high variability within treatment groups, as if often seen in complex soil environments, may obscure clear clustering patterns, which can be influenced by several factors, including nutrient availability, microbial interactions, and soil structure.
This variability in microbial response suggests a nuanced interaction between the fertilizer types and the inherent soil microbial community, where the specific components of the organic matter (e.g. compost) play a critical role in driving both alpha and beta-diversity changes.
4.2. Sulfur with Organic Matter Enhances Beneficial Microbial Component
The results obtained with LEfSe (Linear discriminant analysis Effect Size) analysis showed significant differences in bacterial and fungal communities between different fertilisation treatments, allowing us to identify key taxa that influence soil health and plant-soil interactions. In particular, fertilisers containing organic components, such as the OP and SBOP, favored the proliferation of bacteria and fungi closely linked to the nutrient cycle, improving nutritional efficiency and soil quality.
Among bacteria, Thiobacillus was found to have a significantly higher relative abundance in SBOP-treated soils, emphasizing the critical role of this genus in the sulfur cycle. Thiobacillus oxidises elemental sulfur and reduced compounds such as H₂S and thiosulphates, converting them to sulphate (SO₄²-), a process that not only enriches the soil with essential sulphates, but also facilitates the solubilisation of other nutrients, including phosphates and micronutrient metals [74]. This microbial activity promoted by the organic component of sulfur fertiliser highlights how the incorporation of organic materials can enhance the bioavailability of key nutrients.
Pseudoxanthomonas, another bacterial genus found with significant abundance in OP and SBOP treatments, contributes to the decomposition of complex organic compounds, such as lignin and cellulose, improving nutrient availability to plants. The presence of this genus has also been associated with an increase in soil biodiversity, with potential positive effects on rhizosphere health [75,76]. Along with bacteria such as Pseudoxanthomonas, fungi such as Thermomyces were also found to be more abundant in SBOP treatments. Thermomyces is crucial for the degradation of lignocellulose and facilitates the mineralisation of organic matter, supporting the carbon and nitrogen cycle in the soil [77,78]. This synergy between bacteria and fungi creates a nutrient-rich environment that supports plant growth and health.
Bacteria belonging to the genus Reyranella, present in SBOP and OP treatments, play a significant role in carbon transformation and stabilisation of soil microbial communities, contributing to soil resilience and health. This genus is known for its ability to adapt to variable soil conditions and interact positively with other microorganisms, improving the stability of microbial communities and promoting effective biogeochemical cycles. In parallel, the thermophilic fungus Pseudallescheria has been found in abundance with the addition of organic fertilisers, especially with SBOP, playing an important role in mineralizing organic matter and improving soil fertility [79,80]. This ability to decompose organic matter makes Pseudallescheria a key player in the nutrient cycle, particularly in agricultural systems using organic fertilisers.
The genus Phaselicystis, was also present in abundance in the OP and SBOP treatments is a crucial component linking soil nutrient cycling and plant defence. This genus, and in particular the only known species Phaselicystis flava, contributes to the degradation of complex organic compounds, improving the availability of nutrients such as organic carbon and nitrogen, which are essential for plant growth support [81]. However, Phaselicystis also stands out for its role in plant defense due to its ability to produce arachidonic acid, an allelopathic metabolite that mediates interactions between microorganisms in the soil, particularly between arbuscular mycorrhizal fungi and bacteria[82]. Arachidonic acid not only modulates microbial interactions, but also recruits beneficial microorganisms into the rhizosphere, promoting nutrient turnover and protecting plants from abiotic and biotic stresses [82,83].
Next to bacteria, many fungi also play a crucial role in plant defense. Tritirachium, a fungus abundant in OP and SBOP treatments, is known for its entomopathogenic properties and protease production, which contribute to plant protection from pest attacks [84]. (This highlights how the organic component of fertilisers, in addition to improving nutrition, promote the growth of microorganisms with defensive properties.
Dactylella, another fungus present exclusively in OP and SBOP treatments, is known for its ability to trap nematodes and for its use in the biocontrol of fungal pathogens, contributing to a synergistic approach in natural plant protection. Its presence suggests a reduction in the need for external chemical inputs, improving soil resilience and promoting more sustainable agriculture [85]. Finally, the fungus Chaetomium, known to produce numerous metabolites with antifungal and photoprotective activity, showed a decrease in all treatments compared to the control, especially with OP. This could indicate that enriching the soil with treatments that promote beneficial microorganisms reduces the need for the protective action Chaetomium provides in untreated soil, highlighting the complexity of microbial interactions in an organic fertilization context [86,87]
5. Conclusions
The present study demonstrates that the combined application of chemical and organic fertilizers—specifically, an innovative blend of composted olive pomace and mineral constituents such as sulfur and bentonite—represents a promising strategy for enhancing soil biodiversity. Our findings indicate that even a modest addition of organic matter (5%) to sulfur-based fertilizers can significantly enhance both bacterial and fungal diversity, yielding effects comparable to those observed with purely organic fertilizers. These results underscore the potential of mixed fertilization strategies to sustain soil health and promote plant growth by optimizing microbial diversity and functionality. Notably, we observed substantial changes in the rhizosphere microbiome of Corylus avellana following the application of various fertilizers, which influenced microbiome composition. The application of organic fertilizers, including composted olive pomace and a sulfur-bentonite-olive pomace combination, notably improved the Chao1 index for both bacterial and fungal communities. It is crucial to emphasize that our assessments focus on the short-term effects of these fertilization strategies. Future research will continue to explore the long-term implications of these practices on soil health and sustainability. Nonetheless, the current findings are promising, indicating positive outcomes for agricultural sustainability and the effective use of fertilizers, thereby contributing to a more balanced and sustainable approach to soil management.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Bacteria Beta diversity Bray-Curtis. The table below summarizes the result of pairwise PERMANOVA analysis. The multi-testing adjustment is based on Benjamini-Hochberg procedure procedure (FDR); Table S2: Fungi Beta diversity Bray-Curtis. The table below summarizes the result of pairwise PERMANOVA analysis. The multi-testing adjustment is based on Benjamini-Hochberg procedure procedure (FDR); Table S3: Bacteria LEfSe results with P value cut of 0.05 and Log LDA score 2.0. The table below shows the most 44 features ranked by their p values that are statistically significant; Table S4: Fungi LEfSe results with P value cut of 0.05 and Log LDA score 2.0. The table below shows the most 44 features ranked by their p values that are statistically significant;.
Author Contributions
Conceptualization, A.M (Angela Maffia) and GC; methodology, A.M (Angela Maffia), RS; software RS, AM (Angela Maffia); validation TW, AM (Adele Muscolo) and G.C.; formal analysis, A.M( Angela Maffia), AM (Adele Muscolo), RS.; investigation, T.W.; resources, A.M (Angela Maffia), RS; data curation AL, EA.; writing—original draft preparation, A.M (Angela Maffia), RS; writing—review and editing, A.M (Adele Muscolo), TW; visualization, E.A, A.L.; supervision, GC.; project administration, G.C.; funding acquisition, G.C. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation.
Funding
This work was supported by the project “Evaluation of the rhizosphere environment of hazelnut plants,” funded by the University of Salerno and the doctoral research funds of the Mediterranean University of Reggio Calabria.
Acknowledgments
Thanks to hospitality of National Institute of Agricultural Botany (NIAB, Cambridge, UK).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ciemniewska-Żytkiewicz, H.; Verardo, V.; Pasini, F.; Bryś, J.; Koczoń, P.; Caboni, M. F. Determination of lipid and phenolic fraction in two hazelnut (Corylus avellana L.) cultivars grown in Poland. Food Chemistry 2014, 168, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Özmen, S. Responses of hazelnut trees to organic and conventional managements in the dryland. Erwerbs-Obstbau 2017, 60, 21–30. [Google Scholar] [CrossRef]
- Wei, L. Zhai, Q. The dynamics and correlation between nitrogen, phosphorus, potassium and calcium in a hazelnut fruit during its development. Frontiers of Agriculture in China 2010, 4, 352–357. [Google Scholar] [CrossRef]
- Silvestri, C.; Bacchetta, L.; Bellincontro, A.; Cristofori, V. Advances in cultivar choice, hazelnut orchard management, and nut storage to enhance product quality and safety: an overview. Journal of the Science of Food and Agriculture 2020, 101, 27–43. [Google Scholar] [CrossRef]
- Pannico, A.; Modarelli, G. C.; Stazi, S. R.; Giaccone, M.; Romano, R.; Rouphael, Y.; Cirillo, C. Foliar Nutrition Influences Yield, Nut Quality and Kernel Composition in Hazelnut cv Mortarella. Plants 2023, 12, 2219. [Google Scholar] [CrossRef]
- Vincze, É.-B.; Becze, A.; Laslo, É.; Mara, G. Beneficial soil microbiomes and their potential role in plant growth and soil fertility. Agriculture 2024, 14, 152. [Google Scholar] [CrossRef]
- Lambers, H.; Mougel, C.; Jaillard, B.; Hinsinger, P. Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective. Plant and Soil 2009, 321, 83–115. [Google Scholar]
- Das, P. P.; Singh, K. R.; Nagpure, G.; Mansoori, A.; Singh, R. P.; Ghazi, I. A.; Kumar, A.; Singh, J. Plant-soil-microbes: A tripartite interaction for nutrient acquisition and better plant growth for sustainable agricultural practices. Environmental Research 2022, 214, 113821. [Google Scholar] [CrossRef]
- Berendsen, R. L.; Pieterse, C. M. J.; Bakker, P. A. H. M. The rhizosphere microbiome and plant health. Trends in Plant Science 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J. M.; Lemanceau, P.; Van Der Putten, W. H. Going back to the roots: the microbial ecology of the rhizosphere. Nature Reviews Microbiology 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Cheng, W. , Parton, W. J., Gonzalez-Meler, M. A., Phillips, R., Asao, S., McNickle, G. G., Brzostek, E., & Jastrow, J. D. Synthesis and modeling perspectives of rhizosphere priming. New Phytologist 2014, 201, 31–44. [Google Scholar] [PubMed]
- Lau, J. A.; Lennon, J. T. Evolutionary ecology of plant–microbe interactions: soil microbial structure alters selection on plant traits. New Phytologist 2011, 192, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Romero, Y.; Navarro-Noya, Y. E.; Reynoso-Martínez, S. C.; Sarria-Guzmán, Y.; Govaerts, B.; Verhulst, N.; Dendooven, L.; Luna-Guido, M. 16S metagenomics reveals changes in the soil bacterial community driven by soil organic C, N-fertilizer and tillage-crop residue management. Soil and Tillage Research 2016, 159, 1–8. [Google Scholar] [CrossRef]
- X.B Dai et al. Soil microbial community composition and its role in carbon mineralization in long term fertilization paddy soils. Sci total Environ 2015, 580, 556–53.
- Guo, Z.; Wan, S.; Hua, K.; Yin, Y.; Chu, H.; Wang, D.; Guo, X. Fertilization regime has a greater effect on soil microbial community structure than crop rotation and growth stage in an agroecosystem. Applied Soil Ecology 2020, 149, 103510. [Google Scholar] [CrossRef]
- Lori, M.; Hartmann, M.; Kundel, D.; Mayer, J.; Mueller, R. C.; Mäder, P.; Krause, H.-M. Soil microbial communities are sensitive to differences in fertilization intensity in organic and conventional farming systems. FEMS Microbiology Ecology 2023, 99. [Google Scholar] [CrossRef]
- Legrand, T.P.R.A. , Catalano, S.R., Wos-Oxley, M.L., Stephens, F., Landos, M., Bansemer, M.S., Stone, D.A.J., Qin, J.G., Oxley, A.P.A., 2018. The Inner Workings of the Outer Surface: Skin and Gill Microbiota as Indicators of Changing Gut Health in Yellowtail Kingfish. Front. Microbiol 2664, 8, 2664. [Google Scholar] [CrossRef]
- Orr, C. H.; Leifert, C.; Cummings, S. P.; Cooper, J. M. Impacts of organic and conventional crop management on diversity and activity of Free-Living nitrogen fixing bacteria and total bacteria are subsidiary to temporal effects. PLoS ONE 2012, 7, e52891. [Google Scholar] [CrossRef]
- Arancon, N.Q. , Edwards, C.A., Bierman, P., Metzger, J.D., Lucht, C., . Effects of vermicomposts produced from cattle manure, food waste and paper waste on the growth and yield of peppers in the field. Pedobiologia 2005, 49, 297–306. [Google Scholar] [CrossRef]
- Lazcano, C.; Gómez-Brandón, M.; Revilla, P.; Domínguez, J. Short-term effects of organic and inorganic fertilizers on soil microbial community structure and function. Biology and Fertility of Soils 2012, 49, 723–733. [Google Scholar] [CrossRef]
- Morales-Rodríguez, C.; Palo, C.; Palo, E.; Rodríguez-Molina, M.C. Control of Phytophthora nicotianae with Mefenoxam, Fresh Brassica Tissues, and Brassica Pellets. Plant Dis. 2014, 98, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Blaya, J.; Lacasa, C.; Lacasa, A.; Martínez, V.; Santísima-Trinidad, A.B.; Pascual, J.A.; Ros, M. Characterization of Phytophthora nicotianae isolates in southeast Spain and their detection and quantification through a real-time TaqMan PCR. J. Sci. Food Agric. 2015, 95, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Scotti, R.; Mitchell, A. L.; Pane, C.; Finn, R. D.; Zaccardelli, M. Microbiota characterization of agricultural green Waste-Based suppressive composts using omics and classic approaches. Agriculture 2020, 10, 61. [Google Scholar] [CrossRef]
- Muscolo, A.; Mallamaci, C.; Settineri, G.; Calamarà, G. Increasing Soil and Crop Productivity by Using Agricultural Wastes Pelletized with Elemental Sulfur and Bentonite. Agronomy Journal 2017, 109, 1900–1910. [Google Scholar] [CrossRef]
- Maffia, A.; Marra, F.; Canino, F.; Oliva, M.; Mallamaci, C.; Celano, G.; Muscolo, A. Comparative Study of Fertilizers in Tomato-Grown Soils: Soil quality, Sustainability, and Carbon/Water Footprints. Soil Systems 2023, 7, 109. [Google Scholar] [CrossRef]
- Liu, S. , Moon, C.D., Zheng, N. et al. Opportunities and challenges of using metagenomic data to bring uncultured microbes into cultivation. Microbiome 10, 76. [CrossRef]
- Mr, P.; F, M.; A, M.; C, M.; A, M. Recycling of agricultural (orange and olive) bio-wastes into ecofriendly fertilizers for improving soil and garlic quality. Resources Conservation & Recycling Advances 2022, 15, 200083. [Google Scholar] [CrossRef]
- FAO. Methods of Analysis for Soils of Arid and Semi-Arid Regions; Food and Agricultural Organization: Rome, Italy, 2007; p. 57. [Google Scholar]
- Simmons, T.; Caddell, D. F.; Deng, S.; Coleman-Derr, D. Exploring the Root Microbiome: Extracting Bacterial Community Data from the Soil, Rhizosphere, and Root Endosphere. Journal of Visualized Experiments No. 135. 2018. [Google Scholar] [CrossRef]
- Hoshino, T.Y. , and Matsumoto, M. () An improved DNA extraction method using skim milk from soils that strongly adsorb DNA. Microbes Environ 2004, 19, 13–19. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef]
- Dhariwal, A. Microbiome Analyst: a web based tool for comprehensive statistical, visual and meta-analysis of microbioma data. Nucleic Acids Res. 2017, 45. [Google Scholar] [CrossRef] [PubMed]
- Kers, J. G.; Saccenti, E. The Power of Microbiome Studies: Some considerations on which alpha and beta metrics to use and how to report results. Frontiers in Microbiology 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Chong, J. , Liu P Zhou G, Xia J. Using microbiome analyst for comprehensive statistical, functional and meta-analysis of microbiome data. Nature Protocols 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Chao, A. Nonparametric-estimation of the number of classes in a Population. Scandinavian Journal of Statistics 1984, 11, 265e270. [Google Scholar]
- Lemos, L. N.; Fulthorpe, R. R.; Triplett, E. W.; Roesch, L. F. W. Rethinking microbial diversity analysis in the high throughput sequencing era. Journal of Microbiological Methods 2011, 86, 42–51. [Google Scholar] [CrossRef]
- Magurran, A. E. (2013). Measuring Biological Diversity. Hoboken NJ: John Wiley & Sons.
- Bray RJ, Curtis JT. An ordination of the upland forestcommunities of southern Wisconsin. Ecol Monogr 1957, 27, 325–349. [CrossRef]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: an interactive Venn diagram viewer. BMC Bioinformatics 2014, 15. [Google Scholar] [CrossRef]
- Liu, W.; Cheng, Y.; Guo, J.; Duan, Y.; Wang, S.; Xu, Q.; Liu, M.; Xue, C.; Guo, S.; Shen, Q.; Ling, N. Long-term manure inputs induce a deep selection on agroecosystem soil antibiotic resistome. Journal of Hazardous Materials 2022, 436, 129163. [Google Scholar] [CrossRef]
- Wu, X.; Hu, H.; Li, S.; Zhao, J.; Li, J.; Zhang, G.; Li, G.; Xiu, W. Chemical fertilizer reduction with organic material amendments alters co-occurrence network patterns of bacterium-fungus-nematode communities under the wheat–maize rotation regime. Plant and Soil 2022, 473, (1–2). [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, Y.; Xiong, W.; Ran, W.; Shen, B.; Shen, Q.; Zhang, R. Responses of bacterial communities in arable soils in a Rice-Wheat cropping system to different fertilizer regimes and sampling times. PLoS ONE 2014, 9, e85301. [Google Scholar] [CrossRef]
- Hu, Z.; Ji, L.; Wan, Q.; Li, H.; Li, R.; Yang, Y. Short-Term Effects of Bio-Organic Fertilizer on Soil Fertility and Bacterial Community Composition in Tea Plantation Soils. Agronomy 2022, 12, 2168. [Google Scholar] [CrossRef]
- Tian, W.; Wang, L.; Li, Y.; Zhuang, K.; Li, G.; Zhang, J.; Xiao, X.; Xi, Y. Responses of microbial activity, abundance, and community in wheat soil after three years of heavy fertilization with manure-based compost and inorganic nitrogen. Agriculture Ecosystems & Environment 2015, 213, 219–227. [Google Scholar] [CrossRef]
- Ji, L.; Wu, Z.; You, Z.; Yi, X.; Ni, K.; Guo, S.; Ruan, J. Effects of organic substitution for synthetic N fertilizer on soil bacterial diversity and community composition: A 10-year field trial in a tea plantation. Agriculture Ecosystems & Environment 2018, 268, 124–132. [Google Scholar] [CrossRef]
- Feng, H.; Fu, R.; Hou, X.; Lv, Y.; Zhang, N.; Liu, Y.; Xu, Z.; Miao, Y.; Krell, T.; Shen, Q.; Zhang, R. Chemotaxis of Beneficial Rhizobacteria to Root Exudates: The First Step towards Root–Microbe Rhizosphere Interactions. International Journal of Molecular Sciences 2021, 22, 6655. [Google Scholar] [CrossRef]
- Marra, F.; Maffia, A.; Canino, F.; Greco, C.; Mallamaci, C.; Adele, M. Effects of fertilizer produced from agro-industrial wastes on the quality of two different soils. Archives of Agronomy and Soil Science 2023, 69, 3600–3618. [Google Scholar] [CrossRef]
- Maffia, A.; Marra, F.; Celano, G.; Oliva, M.; Mallamaci, C.; Hussain, M.I.; Muscolo, A. Exploring the Potential and Obstacles of Agro- Industrial Waste-Based Fertilizers. Land 2024, 13, 1166. [Google Scholar] [CrossRef]
- Han, J.; Dong, Y.; Zhang, M. Chemical fertilizer reduction with organic fertilizer effectively improve soil fertility and microbial community from newly cultivated land in the Loess Plateau of China. Applied Soil Ecology 2021, 165, 103966. [Google Scholar] [CrossRef]
- Ren, J.; Liu, X.; Yang, W.; Yang, X.; Li, W.; Xia, Q.; Li, J.; Gao, Z.; Yang, Z. Rhizosphere soil properties, microbial community, and enzyme activities: Short-term responses to partial substitution of chemical fertilizer with organic manure. Journal of Environmental Management 2021, 299, 113650. [Google Scholar] [CrossRef]
- De Menezes, A. B.; et al. C/N ratio drives soil actinobacterial cellobiohydrolase gene diversity. Appl. Environ. Microbiol. 2015, 81, 3016–3028. [Google Scholar] [CrossRef]
- Yang, Z. , Singh, B. & Sitaula, B. Soil organic carbon fractions under diferent land uses in Mardi watershed of Nepal. Commun. Soil Sci. Plan. 35, 615–629.
- Liu, Z. , Guo, Q., Feng, Z., Liu, Z., Li, H., Sun, Y., Liu, C., & Lai, H. (2019). Long-term organic fertilization improves the productivity of kiwifruit (Actinidia chinensis Planch.) through increasing rhizosphere microbial diversity and network complexity. Applied Soil Ecology 2019, 147, 103426. [Google Scholar] [CrossRef]
- Noah, F. , Bradford, M.A., Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar]
- Trivedi, P. , Anderson, I. C., & Singh, B. K. Microbial modulators of soil carbon storage: integrating genomic and metabolic knowledge for global prediction. Trends in Microbiology 2013, 21, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R. Z.; El-Enshasy, H. A.; Dailin, D. J.; Suriani, N. L. Recent understanding of soil acidobacteria and their ecological significance: A Critical review. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef]
- Eichorst, S. , Basu, A., Ahmad, I., Sayyed, R. Z., El-Enshasy, H. A., Dailin, D. J., & Suriani, N. L. Recent understanding of soil acidobacteria and their ecological significance: A Critical review. Frontiers in Microbiology 2020, 11. [Google Scholar]
- Dobrzyński, J.; Wróbel, B.; Górska, E. B. Taxonomy, ecology, and cellulolytic properties of the genus bacillus and related genera. Agriculture 2023, 13, 1979. [Google Scholar] [CrossRef]
- Wu, L.; Jiang, Y.; Zhao, F.; He, X.; Liu, H.; Yu, K. Increased organic fertilizer application and reduced chemical fertilizer application affect the soil properties and bacterial communities of grape rhizosphere soil. Scientific Reports 2020, 10. [Google Scholar] [CrossRef]
- Liu, W.; Cui, S.; Wu, L.; Qi, W.; Chen, J.; Ye, Z.; Ma, J.; Liu, D. Effects of bio-organic fertilizer on soil fertility, yield, and quality of tea. Journal of Soil Science and Plant Nutrition 2023, 23, 5109–5121. [Google Scholar] [CrossRef]
- Wu, J.; Sha, C.; Wang, M.; Ye, C.; Li, P.; Huang, S. Effect of organic fertilizer on soil bacteria in maize fields. Land 2021, 10, 328. [Google Scholar] [CrossRef]
- Sivojienė, D.; Masevičienė, A.; Žičkienė, L.; Ražukas, A.; Kačergius, A. Soil Microbial Community Structure and Carbon Stocks Following Fertilization with Organic Fertilizers and Biological Inputs. Biology 2024, 13, 534. [Google Scholar] [CrossRef]
- Tang, H.; Li, C.; Xiao, X.; Shi, L.; Cheng, K.; Wen, L.; Li, W. Effects of short-term manure nitrogen input on soil microbial community structure and diversity in a double-cropping paddy field of southern China. Scientific Reports 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Chang H-X, Haudenshield JS, Bowen CR and Hartman GL.Metagenome-Wide Association Study and Machine Learning Prediction of Bulk Soil Microbiome and Crop Productivity. Front. Microbiol 2017, 8, 519. [CrossRef]
- Leliaert, F.; Smith, D. R.; Moreau, H.; Herron, M. D.; Verbruggen, H.; Delwiche, C. F.; De Clerck, O. Phylogeny and molecular evolution of the green algae. Critical Reviews in Plant Sciences 2012, 31, 1–46. [Google Scholar] [CrossRef]
- Becker, B. Snow ball earth and the split of Streptophyta and Chlorophyta. Trends in Plant Science 2012, 18, 180–183. [Google Scholar] [CrossRef]
- Zhai, F.H. , Li T.L., Qin X.R., Zhao X.D., Jiang L.W., Xie Y.H. Effect of fertilisation on fungal community in topsoil of winter wheat field. Plant Soil Environ 2022, 68, 317–327. [Google Scholar] [CrossRef]
- Yadav, B.K. , Tarafdar J.C. Penicillium purpurogenum, unique P mobilizers in arid agro-ecosystems. Arid Land Research and Management 2011, 25, 87–99. [Google Scholar] [CrossRef]
- Maity, A. , Pal R.K., Chandra R., Singh N.V. Penicillium pinophilum – a novel microorganism for nutrient management in pomegranate (Punica granatum L.). Scientia Horticulturae 2014, 169, 111–117. [Google Scholar] [CrossRef]
- Damo, J.L.C.; Shimizu, T.; Sugiura, H.; Yamamoto, S.; Agake, S.-i.; Anarna, J.; Tanaka, H.; Sugihara, S.; Okazaki, S.; Yokoyama, T.; et al. The Application of Sulfur Influences Microbiome of Soybean Rhizosphere and Nutrient-Mobilizing Bacteria in Andosol. Microorganisms 2023, 11, 1193. [Google Scholar] [CrossRef]
- Semenov, M. V.; Krasnov, G. S.; Semenov, V. M.; Van Bruggen, A. Mineral and organic fertilizers distinctly affect fungal communities in the crop rhizosphere. Journal of Fungi 2022, 8, 251. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, X.; Hu, Y.; Zhao, Y. Effects of Different Proportions of Organic Fertilizer in Place of Chemical Fertilizer on Microbial Diversity and Community Structure of Pineapple Rhizosphere Soil. Agronomy 2024, 14, 59. [Google Scholar] [CrossRef]
- Guo, X.; Liu, J.; Xu, L. ; Sun,F.; Ma, Y.; Yin, D.; Gao, Q.; Zheng, G.;Lv, Y. Combined Organic and Inorganic Fertilization Can EnhanceDry Direct-Seeded Rice Yield byImproving Soil Fungal Communityand Structure. Agronomy 2022, 12, 1213. [Google Scholar] [CrossRef]
- Agha, A. B. A.; Kahrizi, D.; Ahmadvand, A.; Bashiri, H.; Fakhri, R. Identification of Thiobacillus bacteria in agricultural soil in Iran using the 16S rRNA gene. Molecular Biology Reports 2018, 45, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Li, J.; Lyu, J.; Feng, Z.; Zhang, G.; Yang, H.; Gao, C.; Jin, L.; Yu, J. Chemical fertilizer reduction combined with bio-organic fertilizers increases cauliflower yield via regulation of soil biochemical properties and bacterial communities in Northwest China. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, X.; Li, R.; Wang, G.; Jin, Y.; Xu, W.; Wang, H.; Qu, J. Organic amendment improves rhizosphere environment and shapes soil bacterial community in black and red soil under lead stress. Journal of Hazardous Materials 2021, 416, 125805. [Google Scholar] [CrossRef]
- Tan, Y.; Wang, J.; He, Y.; Yu, X.; Chen, S.; Penttinen, P.; Liu, S.; Yang, Y.; Zhao, K.; Zou, L. Organic fertilizers shape soil microbial communities and increase soil amino acid metabolites content in a blueberry orchard. Microbial Ecology 2022, 85, 232–246. [Google Scholar] [CrossRef]
- Krell, T. Microcalorimetry: a response to challenges in modern biotechnology. Microbial Biotechnology 2007, 1, 126–136. [Google Scholar] [CrossRef]
- Wen, Y.-C.; Li, H.-Y.; Lin, Z.-A.; Zhao, B.-Q.; Sun, Z.-B.; Yuan, L.; Xu, J.-K.; Li, Y.-Q. Long-term fertilization alters soil properties and fungal community composition in fluvo-aquic soil of the North China Plain. Scientific Reports 2020, 10. [Google Scholar] [CrossRef]
- Stromberger, M. E.; Shah, Z.; Westfall, D. G. High specific activity in low microbial biomass soils across a no-till evapotranspiration gradient in Colorado. Soil Biology and Biochemistry 2010, 43, 97–105. [Google Scholar] [CrossRef]
- Rodríguez-Berbel, N.; Ortega, R.; Lucas-Borja, M. E.; Solé-Benet, A.; Miralles, I. Long-term effects of two organic amendments on bacterial communities of calcareous mediterranean soils degraded by mining. Journal of Environmental Management 2020, 271, 110920. [Google Scholar] [CrossRef]
- Lu, P.; Shi, H.; Tao, J.; Jin, J.; Wang, S.; Zheng, Q.; Liu, P.; Xiang, B.; Chen, Q.; Xu, Y.; Li, Z.; Tan, J.; Cao, P. Metagenomic insights into the changes in the rhizosphere microbial community caused by the root-knot nematode Meloidogyne incognita in tobacco. Environmental Research 2022, 216, 114848. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.; Ma, D.; Wang, L.; Zhang, X.; Ding, Y.; Fan, K.; Xu, Z.; Yuan, C.; Jia, H.; Ren, Y.; Ding, Z. Differential responses of the rhizosphere microbiome structure and soil metabolites in tea (Camellia sinensis) upon application of cow manure. BMC Microbiology 2022, 22. [Google Scholar] [CrossRef] [PubMed]
- Betzel, C. , et al. . “Structure of a serine protease proteinase K from Tritirachium album Limber at 0.98 Å resolution. Biochemistry 2001, 40, 3080–3088. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, T. , Priya, R., & Sunitha, T. (2020). Dactylella. In Elsevier eBooks (pp. 809–816). [CrossRef]
- Fatima N, Muhammad SA, Khan I, Qazi MA et al Chaetomium endophytes: a repository of pharmacologically active metabolites. Acta Physiol Plant 2016, 38, 136. [CrossRef]
- Zhao, S.-S.; Zhang, Y.-Y.; Yan, W.; Cao, L.-L.; Xiao, Y.; Ye, Y.-H. Chaetomium globosum CDW7, a potential biological control strain and its antifungal metabolites. FEMS Microbiology Letters 2016, fnw287. [Google Scholar] [CrossRef]
Figure 1.
Experimental site and pots with Corylus avellana plants and different treatments.

Figure 2.
DNA quality control of DNA extraction samples obtained by Luminescence Image Analyzer System (LAS).
Figure 2.
DNA quality control of DNA extraction samples obtained by Luminescence Image Analyzer System (LAS).
Figure 3.
Top 10 Bacteria taxa relative abundance for phylum (a) and genus (b) of soils treated with synthetic fertilizer (NPK), Olive Pomace (OP), Sulfur bentonite (SB), Sulfur bentonite + Olive Pomace (SBOP), CTR is unfertilized soils. .
Figure 3.
Top 10 Bacteria taxa relative abundance for phylum (a) and genus (b) of soils treated with synthetic fertilizer (NPK), Olive Pomace (OP), Sulfur bentonite (SB), Sulfur bentonite + Olive Pomace (SBOP), CTR is unfertilized soils. .
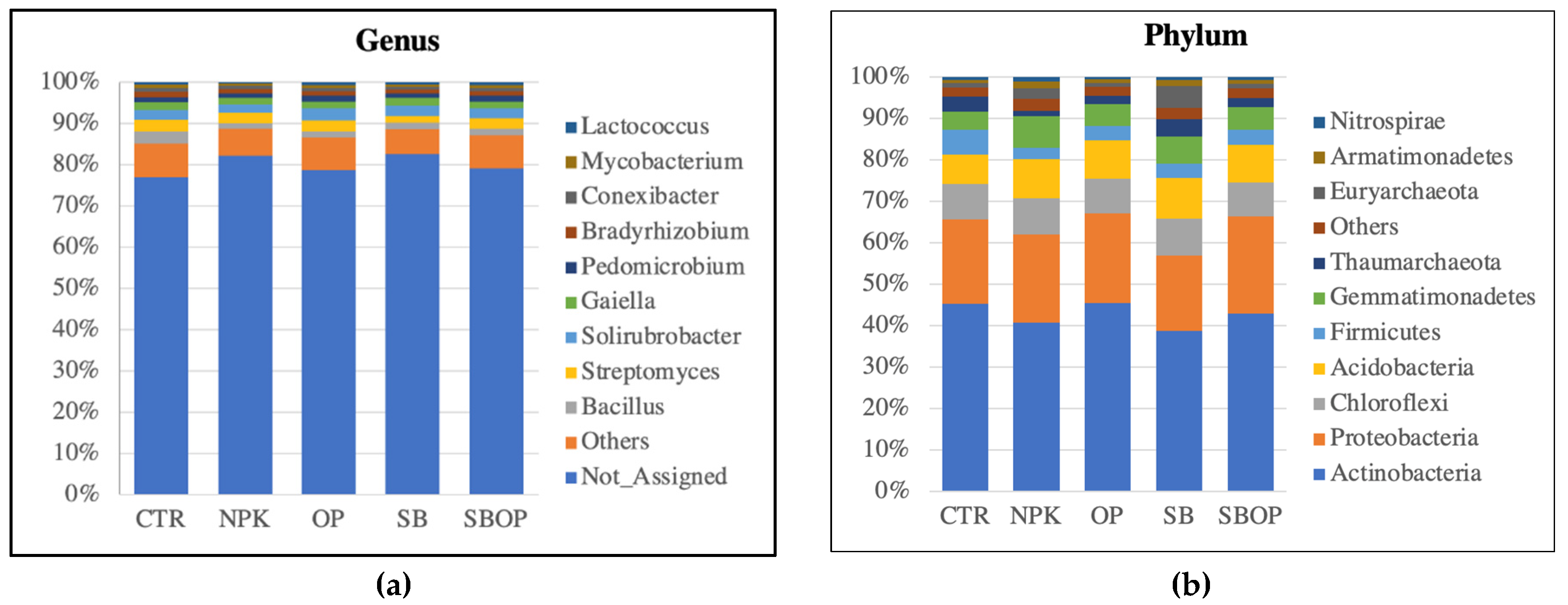
Figure 4.
Top 10 Fungi taxa abundance for phylum (a) and genus (b).
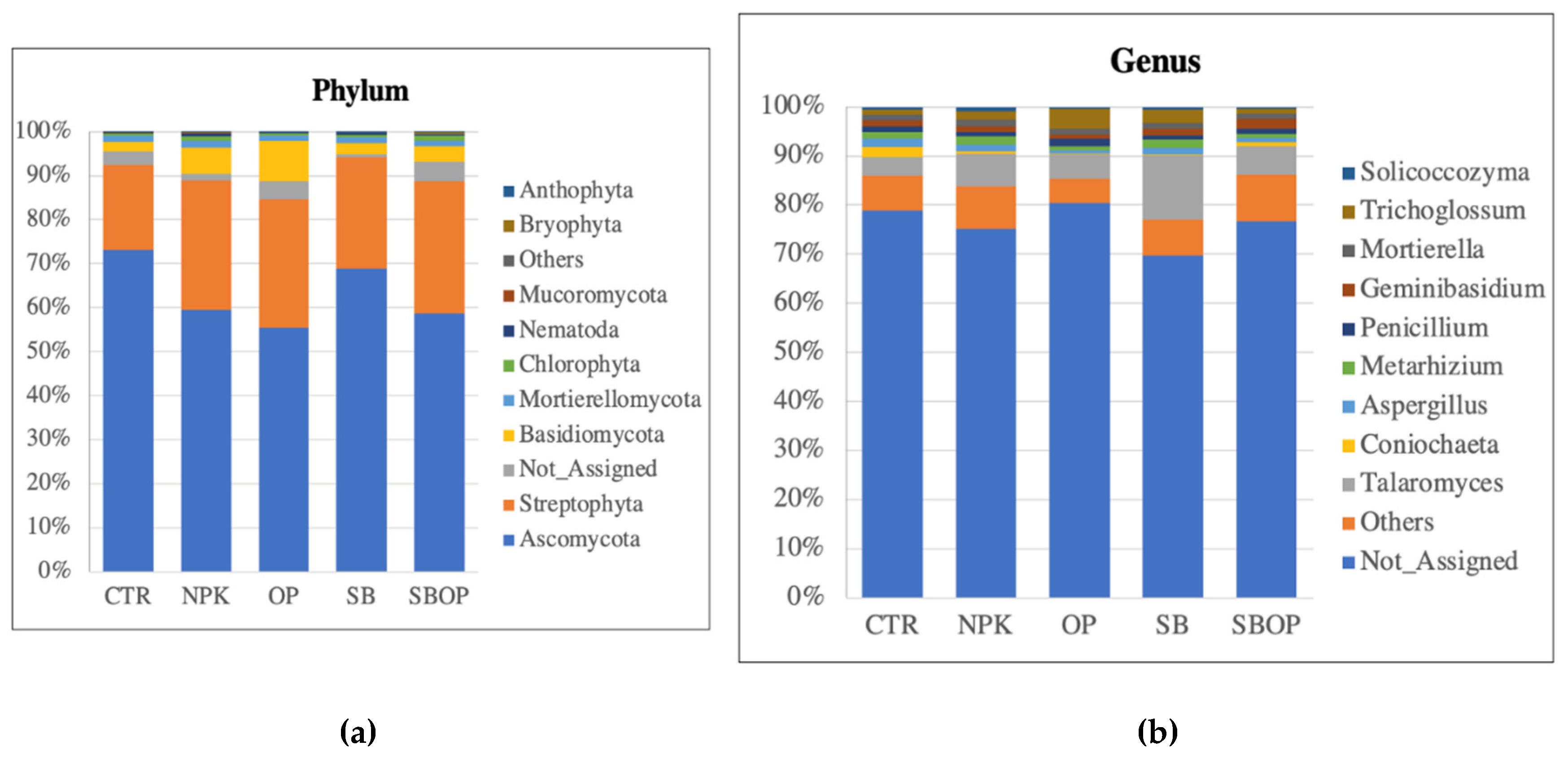
Figure 5.
Chao1 index of Bacteria (a) and Fungi (b). Statistically significant differences are expressed with (*) for p value < 0.05 through post-hoc pairwise comparison (multiple-groups only). Principal Coordinate analysis (PCoA) of bacteria (c) and fungi (d) calculated with Bray-Curtis dissimilarity of soils treated with synthetic fertilizer (NPK), Olive Pomace (OP), Sulfur bentonite (SB), Sulfur bentonite + Olive Pomace (SBOP), CTR is unfertilized soils. .
Figure 5.
Chao1 index of Bacteria (a) and Fungi (b). Statistically significant differences are expressed with (*) for p value < 0.05 through post-hoc pairwise comparison (multiple-groups only). Principal Coordinate analysis (PCoA) of bacteria (c) and fungi (d) calculated with Bray-Curtis dissimilarity of soils treated with synthetic fertilizer (NPK), Olive Pomace (OP), Sulfur bentonite (SB), Sulfur bentonite + Olive Pomace (SBOP), CTR is unfertilized soils. .
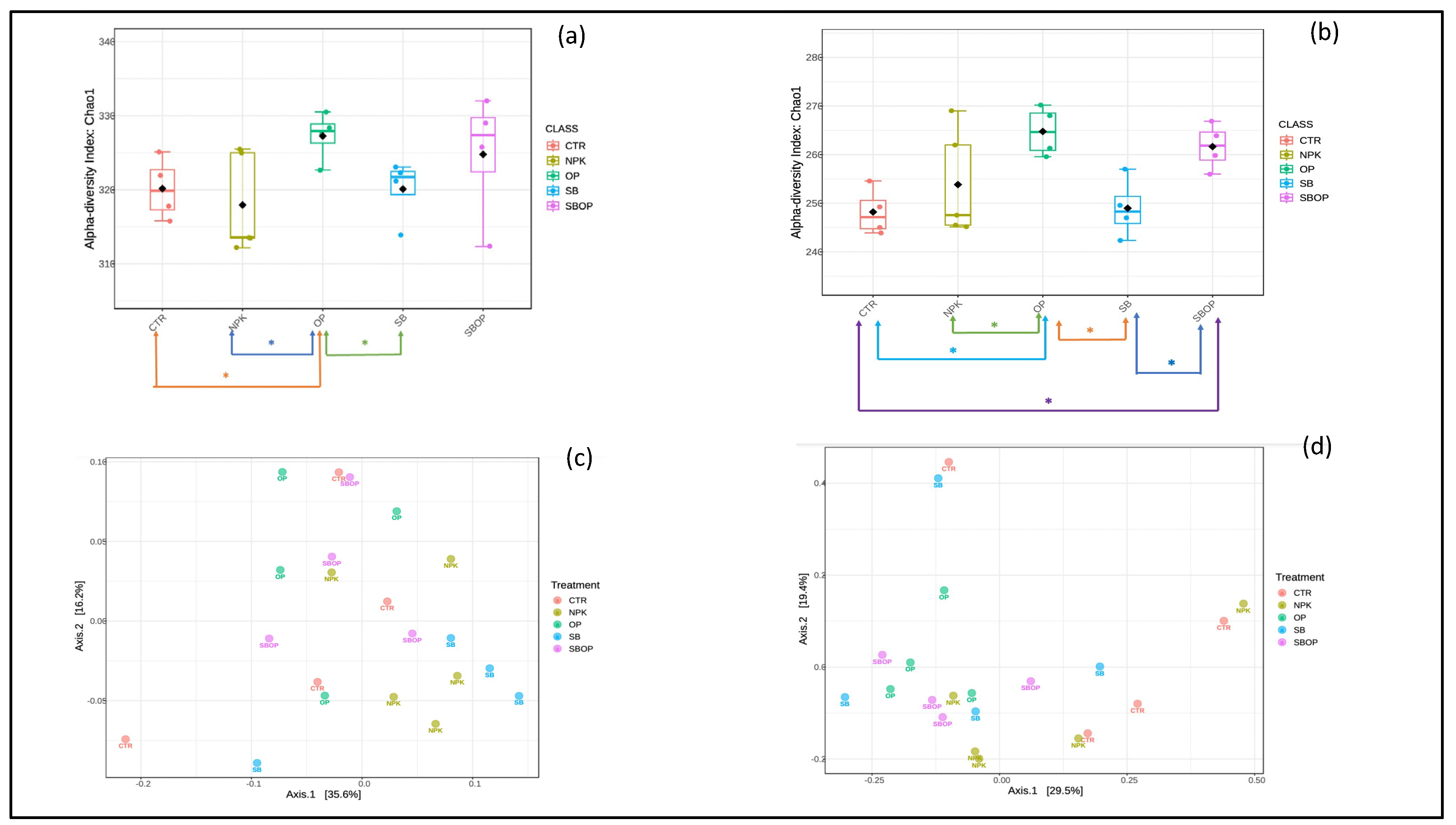
Figure 6.
LEfSe results on soil bacterial (a) and fungal (g) communities. Histogram of the LDA scores computed for features differentially abundant on different treatment. LEfSe scores can be interpreted as the degree of consistent difference in relative abundance between features of analyzed bacterial and fungal communities. The histogram thus identifies which clades among all those detected as statistically and biologically differential explain the greatest differences between genus communities.
Figure 6.
LEfSe results on soil bacterial (a) and fungal (g) communities. Histogram of the LDA scores computed for features differentially abundant on different treatment. LEfSe scores can be interpreted as the degree of consistent difference in relative abundance between features of analyzed bacterial and fungal communities. The histogram thus identifies which clades among all those detected as statistically and biologically differential explain the greatest differences between genus communities.
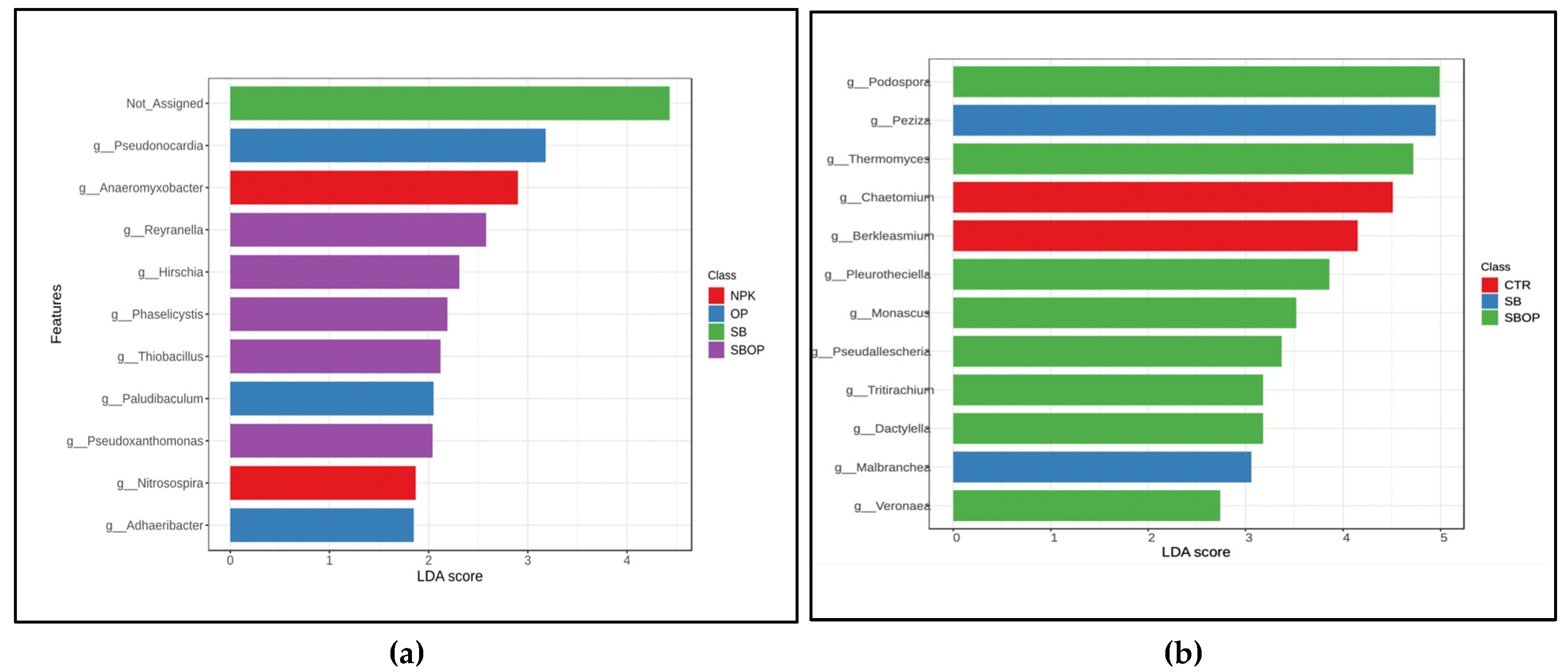
Figure 7.
Veen diagram of genus of bacteria which were found to be statistically significant from linear discriminant analysis effect size (LEfSE) and and related box plots.
Figure 7.
Veen diagram of genus of bacteria which were found to be statistically significant from linear discriminant analysis effect size (LEfSE) and and related box plots.
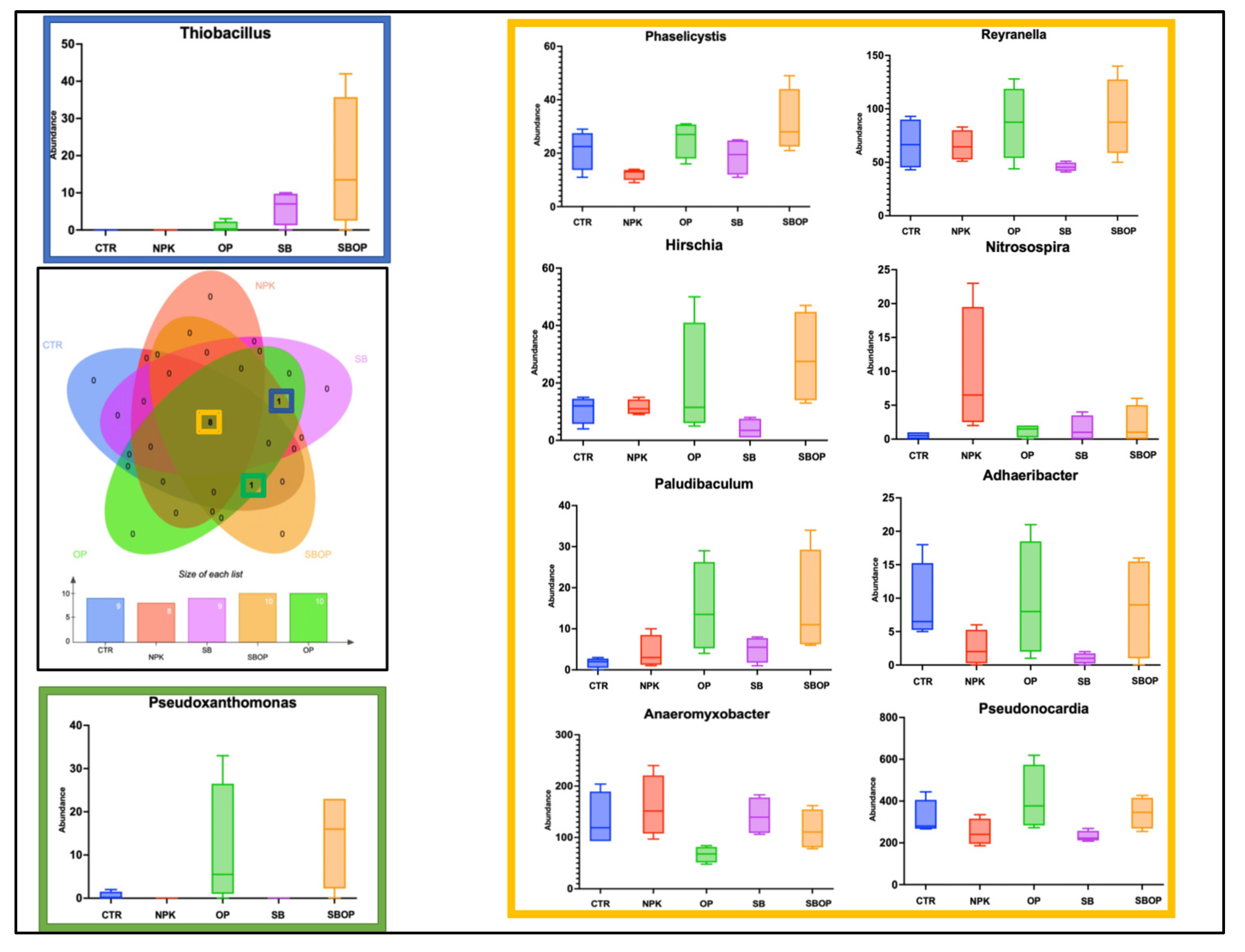
Figure 8.
Veen diagram of genus of fungi which were found to be statistically significant from linear discriminant analysis effect size (LEfSE) and and related box plots.
Figure 8.
Veen diagram of genus of fungi which were found to be statistically significant from linear discriminant analysis effect size (LEfSE) and and related box plots.
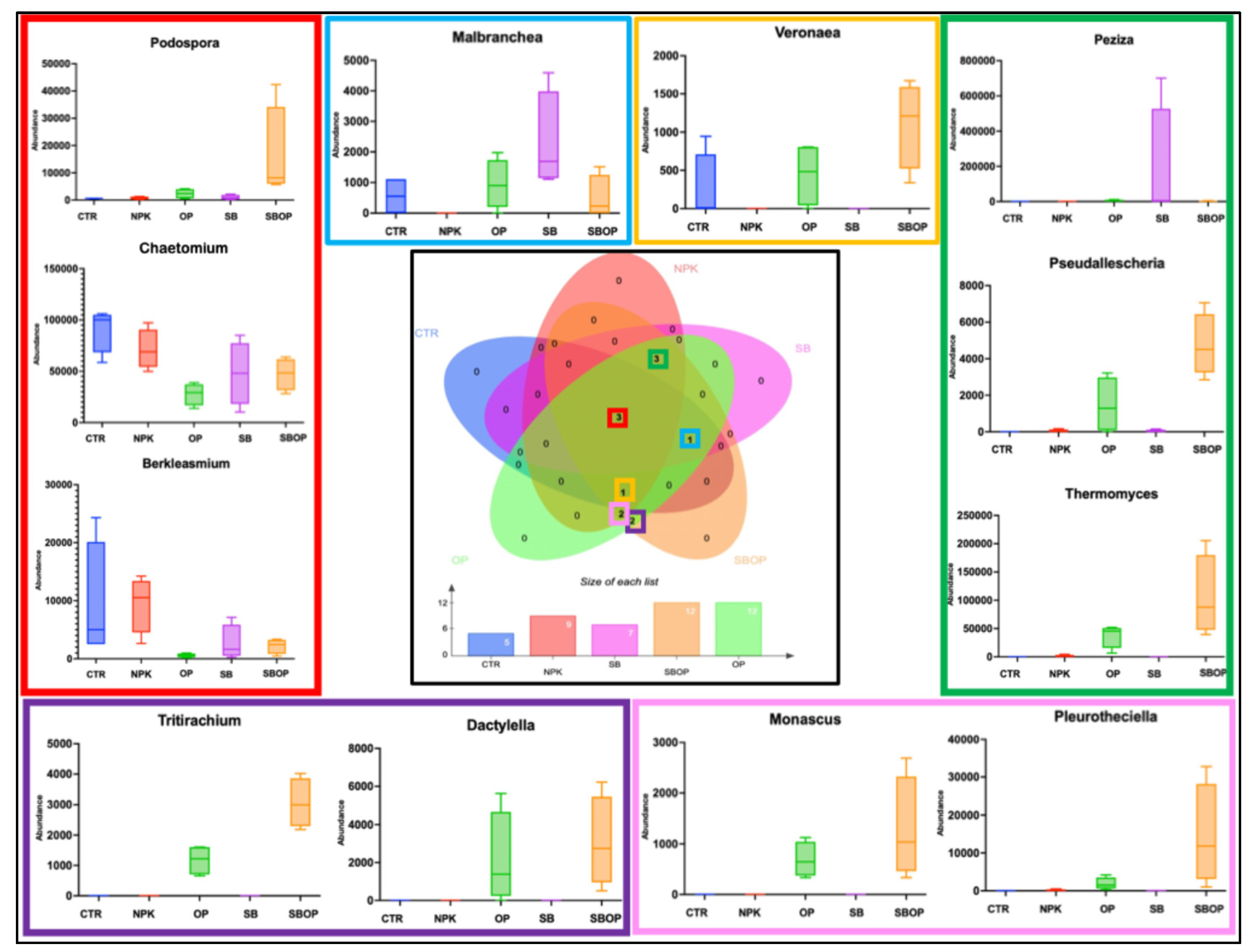
Figure 9.
Bacteria Heatmap of relative abundance at the class level.
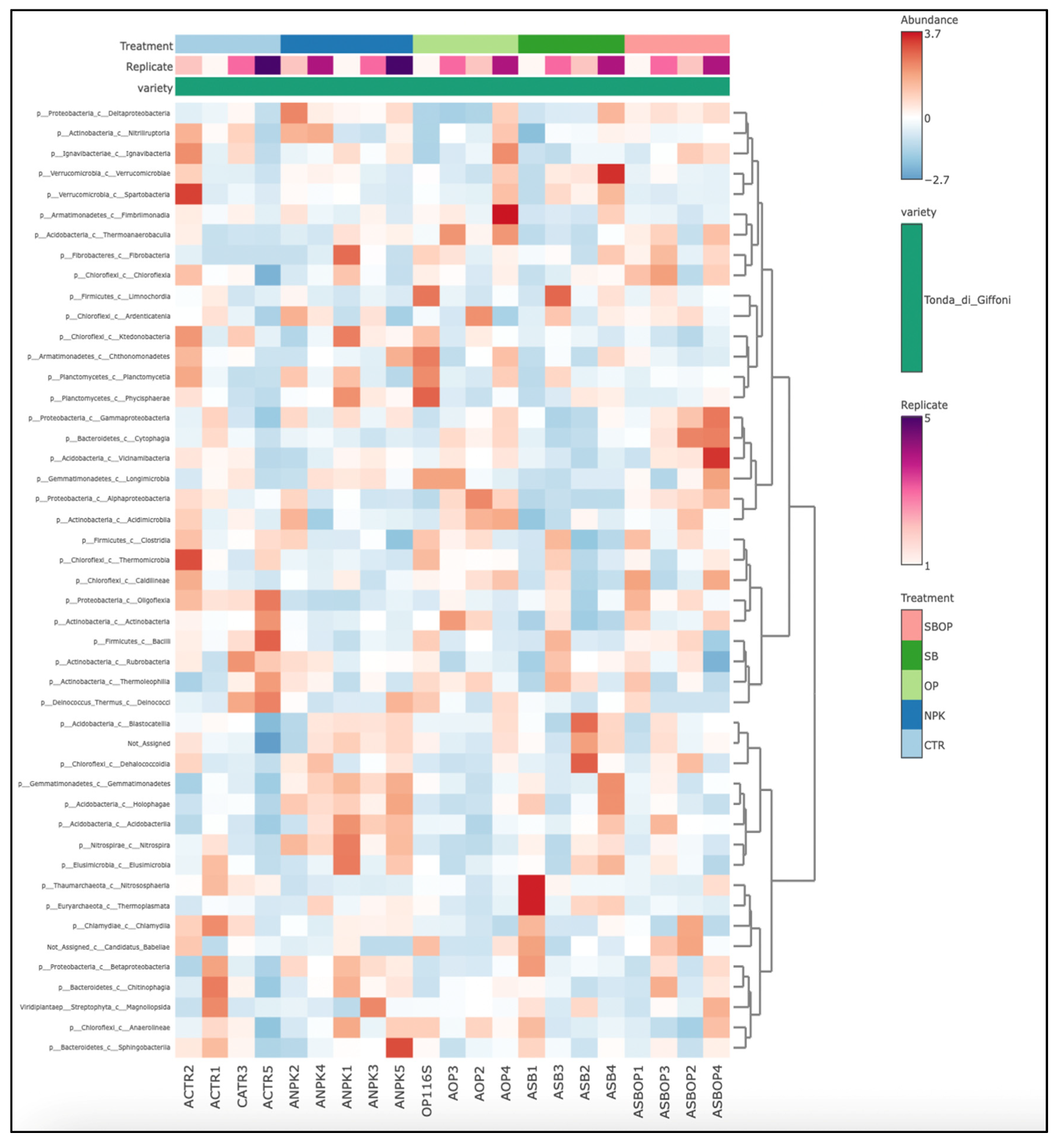
Figure 10.
Fungi Heatmap of relative abundance at the class.
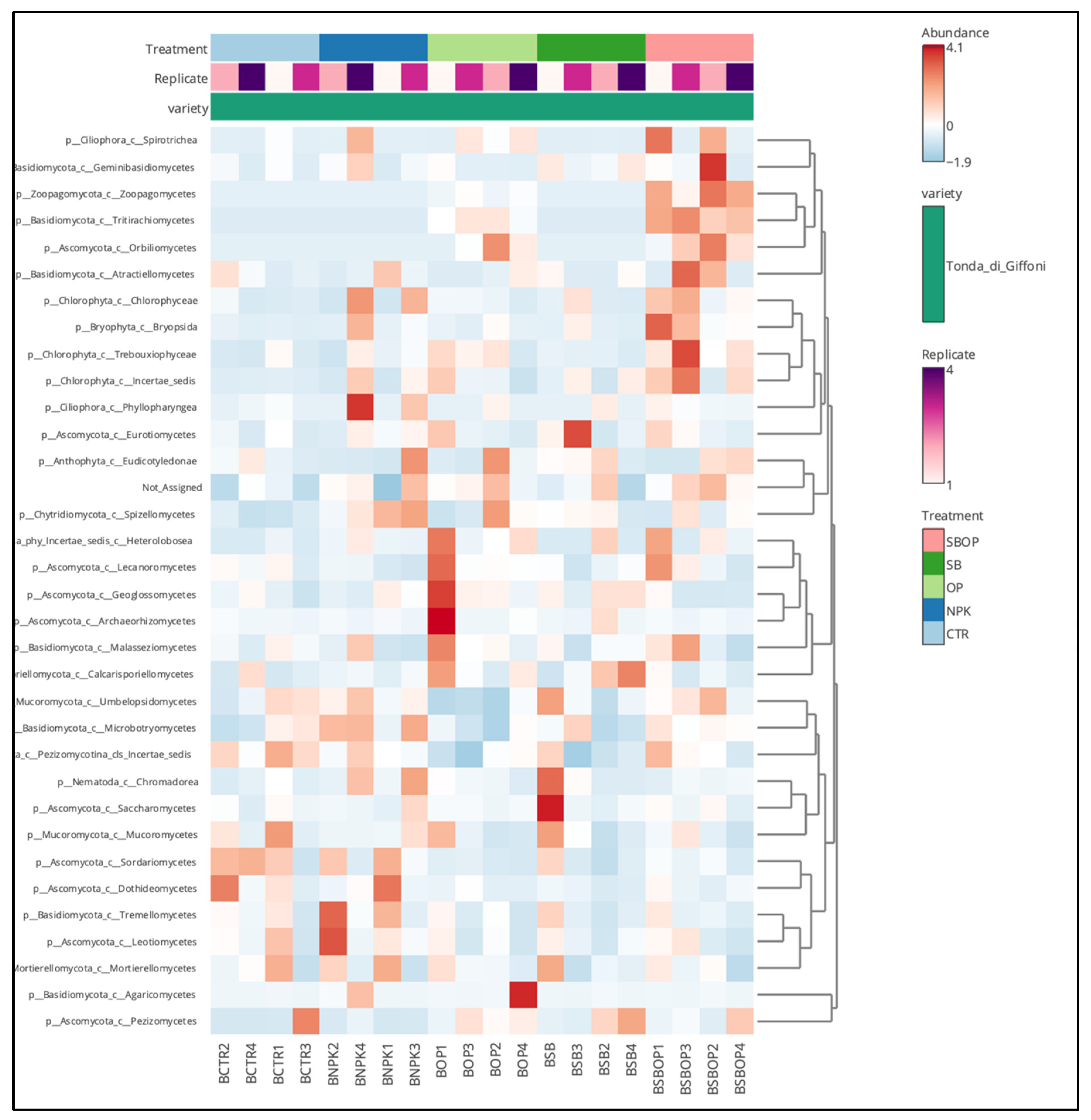

Table 1.
Chemical properties of composted olive pomace. The data are the mean of three replicates ± standard deviation.
Table 1.
Chemical properties of composted olive pomace. The data are the mean of three replicates ± standard deviation.
| Chemical properties | Value |
| pH (H2O) | 7.8 ± 0.04 |
| EC (mS cm-1) | 2.6 ± 0.27 |
| Moisture (g kg-1 fw) | 383 ± 0.3 |
| C % (g kg-1 dw) | 426 ± 0.4 |
| Total N (g kg-1 dw) | 19.6 ± 0.1 |
| C/N | 21.7 ± 0.3 |
| Na + (mg g-1 dw) | 4.35 ± 0.02 |
| NH4+ (mg g-1 dw) | 1.50 ± 0.01 |
| K+ (mg g-1 dw) | 9.13 ± 0.06 |
| Mg2+ (mg g-1 dw) | 1.45 ± 0.43 |
| Ca2+ (mg g-1 dw) | 14.06 ± 1.4 |
| Cl- (mg g-1 dw) | 0.02 ± 0.5 |
| PO43- (mg g-1 dw) | 0.40 ± 0.4 |
| SO42- (mg g-1 dw) | 0.18 ± 2.9 |
| WSB (mg TAE g-1 dw) | 2.5 ± 0.05 |
Table 2.
Composition of different fertilizers.
| Fertilizers | Composition |
|---|---|
| Sulfur Bentonite + Olive Pomace (SBOP) | 5 % of composted olive pomace recovered by a two-phase oil mill 10 % of bentonite clay 85% of elementar Sulfur. |
| Sulfur Bentonite (SB): | 90% of elementar Sulfur 10 % of Bentonite clay |
| Composted Olive Pomace (OP) | 34% of composted olive pomace recovered by a two-phase oil mill 33% of buffalo manure 33% of a mixture consisting of wood defibrate and olive leaves. |
| Synthetic fertilizer (NPK) | 20% of N 10% of P2O5 10% of K2O |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



