
Submitted:
01 November 2024
Posted:
04 November 2024
You are already at the latest version
Abstract
Streptococcus agalactiae is an important pathogen responsible for cases of high mortality in farmed and wild fish worldwide. In Brazil, this bacterium has been commonly associated with outbreaks in Nile tilapia farms, but other native fish species are also susceptible. Since floating cages are one of the most common culture systems used in the country, the close contact between farmed tilapia and native fish species presents a risk concerning the transmission of this pathogen. In this study, we characterized a mortality outbreak in free-living trahira and in farmed arapaima, as well as the genetic and antimicrobial susceptibility patterns of the isolates obtained. During the outbreaks, moribund fish were sampled and subjected to bacterial examination, after which the isolates were identified via MALDI-ToF analysis. Genotyping was evaluated using repetitive sequence-based PCR (REP-PCR) and multilocus sequence typing (MLST). Antimicrobial susceptibility was evaluated using disc diffusion assays. In addition, whole genome analysis also was performed. S. agalactiae was identified in all diseased fish, all of which belonged to serotype Ib, however, trahira strains were classified as non-typeable lineages in the MLST assay, while arapaima strains were classified as ST260. These isolates were shown to be similar to the main genotype found in Nile tilapia in Brazil, using REP-PCR, MLST and phylogenomic analysis. The pathogenicity of bacterium was confirmed by Kock’s postulates for both fish species. The antimicrobial susceptibility assay showed variable results to the same antibiotics among the isolates, prompting four of the isolates to be classified as multidrug-resistant. This study represents the first report of a natural outbreak of Streptococcus agalactiae infection in wild trahira and farmed arapaima inhabiting the same aquatic environment with Nile tilapia.
Keywords:
antimicrobial resistance
; bacteria
; fish
; genetic diversity
; genomic analysis
1. Introduction
Streptococcus agalactiae infections have been described in a large number of hosts [1], especially humans [2], cattle [3], and fish [4]. This Gram-positive bacterium is considered an important pathogen for aquaculture due to the high mortality on farms worldwide and the septicemia and meningoencephalitis caused in a wide range of fish species, including captive and wild fish, with tilapia and its hybrids the most affected [5,6]. In Brazil, this pathogen has been isolated from the Nile tilapia (Oreochromis niloticus), Amazon catfish (Leiarius marmoratus × Pseudoplatystoma corruscans), barred sorubim (Pseudoplatystoma reticulatum), prochilod (Prochilodus lineatus), and pearl cichlid (Geophagus brasiliensis) in monoculture or polyculture with tilapia [4,7,8,9]. There are no descriptions concerning the natural outbreak of S. agalactiae infections in free-living fish in Brazil.
Trahira (Hoplias malabaricus, Bloch) is a native freshwater fish species, widely distributed in the hydrographic basins of Central and South America, and found in different aquatic environments, such as rivers, lakes, floodplains, and reservoirs [10,11]. This predatory fish, which is essentially piscivorous (Rodrigues et al., 2017), has also been used as an ornamental fish [13], and its meat is much appreciated in Brazilian cuisine [11]. Thereby, there was an increase in the trahira production in Brazilian aquaculture [13], especially in the southern region, where the total fish production reached 793,224 tons in 2022 [14]. Currently, the causes of diseases outbreaks in trahira are unknown. The identification of parasite communities, such as protozoans, metacercariae, Digenea, nematodes, and parasitic crustaceans [15,16,17], as well as the isolation of Aeromonas spp. from fillets sold in a commercial establishment [18] have been reported in trahira. Nevertheless, there is still no evidence supporting that such organisms may be involved in the disease etiology of this aquatic host. Beyond this, there are no other reports concerning S. agalactiae infections in trahira.
Arapaima (Arapaima gigas, Schinz) is considered one of the largest species of freshwater scale fish in the world, reaching more than 2 meters in length and weighing about 125 kg [19,20]. Popularly known as “pirarucu”, “paiche” or “arapaima”, this fish species is commonly found in lowland lentic waters of the Amazon, Araguaia-Tocantins, and Orinoco River basins, as lakes, floodplains and flooded forest [21,22,23]. Moreover, the presence of arapaima has already been confirmed in the upper Paraná River basin [24]. The species is currently present in 12 countries, being a native species in Brazil, Colombia, Ecuador, Guyana, and Peru, and introduced in Bolivia, China, Cuba, Mexico, Philippines, Singapore, Thailand and USA [25,26,27,28]. This fish species is carnivorous, obligate air-breathing and its commercial production have been increasing in different Brazilian states due to its high growth rate, rusticity, adaptability to commercial feed, high carcass yield, absence of thorns in the meat, resistance to the environment with high stocking densities and low concentration of oxygen dissolved, and acceptance of its meat in the consumer market [29]. These characteristics made it possible for the arapaima production in Brazilian aquaculture to reach 2028,247 tons in 2022 [14]. However, the production of this fish species has some obstacles, especially those related to artificial reproduction, nutrition, high mortality rates in early life stages, and the occurrence of outbreaks of parasitic and infectious diseases [30,31,32]. Among the parasitic agents that cause diseases in arapaima farms stand out: Ichthyophthirius multifiliis [33], Dawestrema sp. [30], Hysterothylacium sp. [34], and Polyacanthorhynchus macrorhynchus [33]. On the other hand, bacterial infections caused by Aeromonas hydrophila [31], Aeromonas jandaei [35], Edwardsiella tarda [36], Salmonella enterica subsp. arizonae [37], Serratia fonticola [38] and lactococcosis-causing bacteria [39] have already been reported in diseased arapaima. A previous study detected a higher prevalence of Streptococcus spp. (16.7%) in arapaima without clinical signs from Peruvian farms that reported 1 a 2% of mortality for undiagnosed causes [40]. However, the authors did not determine the isolates obtained at the species level.
Therefore, this study aimed to identify the etiological agent responsible for the mortalities in free-living trahiras and farmed arapaima in Brazil. The genotype and antibiotic resistance patterns of the isolates were evaluated. In addition, the whole genome sequencing was performed for a representative strain of each fish species to elucidate interspecies transmission and detected antimicrobial resistance genes.
2. Materials and Methods
2.1. Outbreaks
2.1.1. Free-Living Trahiras
In March 2017, cases of mortality were observed in free-living trahiras inhabiting a lake on a farm in the city of Mateus Leme/MG (Geographic coordinates: 19º59’23” S, 44º25’21” W). In this lake, there are other wild native fish species and Nile tilapia reared in floating cages. During the evaluation period, the concentration of dissolved oxygen remained above 6.0 mg/L, the water temperature varied between 28 and 30.5 ºC, and eight diseased adult trahiras were observed. Of these, three moribund trahiras (average length of 50 ± 4.24 cm) were sampled and transported on ice to the laboratory for analysis. The remaining fish were found dead and in an advanced stage of autolysis. Previous cases of streptococcosis in farmed tilapia were reported by the farmer in the previous fish production cycle, approximately 8 months earlier. However, during the studied outbreak, only wild trahira presented clinical signs of diseases.
2.1.2. Farmed Arapaima
Between April and July 2017, a disease outbreak occurred in a commercial fish farm located in the city of Montes Claros/MG (Geographic coordinates: 16º43’41” S, 43º51’54” W). On this farm, Nile tilapia and arapaima are reared in excavated tanks, however, mortalities were verified in arapaima tanks. Six moribund fish (average weight of 40 ± 12 g) were sampled and transported on ice to the laboratory for analysis.
2.2. Bacteriological Examinations
Brain and kidney tissues were aseptically sampled, streaked onto tryptone soybean agar (Kasvi, Italy) supplemented with 5% defibrinated sheep blood (TSAB), and incubated at 28 ℃ for 48 h for bacterial isolation. Next, pure colonies were subjected to Gram staining, catalase and oxidase tests, and Lancefield’s serotyping using the Slidex Strepto Plus kit (BioMerieux, France). The bacterial species were confirmed via MALDI-ToF mass spectrometry analysis using Bruker Microflex MALDI Biotyper 2.0 (Bruker Daltonics, Germany), as previously described [41]. Then, the isolates were stored in 15% glycerol BHI (“Brain Heart Infusion”, Himedia, India) broth at – 70 ℃ until further use.
2.3. DNA Extraction
The Streptococcus agalactiae isolates obtained from this study were thawed, streaked onto TSAB, and incubated at 28 ℃ for 48 h. Then, the bacterial DNA was extracted with the Wizard Genomic DNA Purification kit (Promega, USA), according to the manufacturer’s instructions. The extracted DNA was quantified with the Nanodrop 2000 spectrophotometer (Thermo Scientific, USA) and stored at – 20 ℃ until use as a template in the PCR assays.
2.4. Capsular Typing
The capsular polysaccharide typing was performed via a multiplex PCR assay, as described by Poyart et al. [42] with some modifications. The multiplex PCR was performed with a GoTaq DNA Polymerase kit (Promega, USA) and a final volume of 25 µL. The mix was composed of 5 µL of 1x PCR buffer, 0.25 µL of each previously designed PCR primer [42], 0.5 µL of dNTPs, 2 µL of MgCl2, 2 U (0.4 µL) of Taq DNA polymerase, and 100 ng of template DNA. The PCR conditions consisted of an initial step of 95 ℃ for 15 min, followed by 35 cycles of 95 ℃ for 1 min, 63 ℃ for 1 min, and 72 ℃ for 1 min, with a final elongation step of 72 ℃ for 10 min. The primers used were acquired from Integrated DNA Technologies (IDT, USA). The multiplex PCR was performed using the Veriti 96-well Thermal Cycler equipment (Life Technologies, USA). The PCR products were separated via 1.5% agarose gel electrophoresis (70 V for 180 min), with the 100 bp marker (Invitrogen, USA) as the molecular size standard. The gel was stained with ethidium bromide (0.5 μg mL-1 for 30 min) and subsequently visualized via UV transillumination. Images were captured using the L-Pix EX equipment (Loccus Biotechnology, Brazil). The SA07 (serotype Ia), SA16 (serotype Ib), and NEM316 (serotype III) strains were included as positive controls for the multiplex PCR. To confirm the serotype, the isolates were subjected to serotyping using the Strep B Latex Ib (batch LBIb-P1; Statens Serum Institute, Denmark) according to the manufacturer's recommendations.
2.5. Repetitive Extragenic Palindromic-PCR (REP-PCR)
The genetic diversity of the isolates was evaluated by repetitive extragenic palindromic-PCR (REP-PCR), described by Costa et al. [43] with some modifications. In addition, Streptococcus agalactiae strains isolated from diseased Nile tilapia (SA52 and SA95) and Amazon catfish (SA103) were included in this analysis. These strains had the DNA extracted as described above.
PCR was performed with a HotStart Taq Polymerase kit and a final volume of 25 µL. The mix was composed of 5 µL of 1x PCR buffer, 1.25 µL of GTG5 (5'-GTGGTGGTGGTGGTG-3') (Integrated DNA Technologies, EUA), 0.5 µL of dNTPs, 3 µL of MgCl2, 0.5 µL of Taq DNA polymerase, and 100 ng of template DNA. The PCR conditions consisted of an initial step of 95 ℃ for 15 min, followed by 30 cycles of 95 ℃ for 30 s, 45 ℃ for 1 min, and 72 ℃ for 8 min, with a final elongation step of 72 ℃ for 16 min. The Veriti 96-Well Thermal Cycler also was used. The amplicons were separated by electrophoresis on a 2% agarose gel, stained with ethidium bromide (0.5 mg mL-1 for 30 min), visualized by UV transillumination, and the images were captured using an L-Pix EX. A 1 kb DNA ladder (Invitrogen, USA) was used as the molecular size standard. The REP-PCR images were analyzed using BioNumerics version 6.6 (Applied Maths, Belgium). The Dice coefficient was used to analyse the similarities of the banding patterns [44]. Dendrograms were created using the unweighted pair group method with an average (UPGMA) approach. Isolates that showed similarity ≥ 92% were considered clonally related [45]. The discriminatory power of REP-PCR was calculated using Simpson’s index of diversity (D) [46]. This analysis was performed using the online software Comparing Partitions (available at http://www.comparingpartitions.info/) [47].
2.6. Multilocus Sequence Typing (MLST)
The MLST analysis was performed for the sequencing of the seven housekeeping genes (adhP, pheS, atr, glnA, sdhA, tkt, and glcK), following the methodology described by Jones et al. [48] with some modifications. The PCR was performed with a HotStart Taq Polymerase kit (Qiagen, USA) and a final volume of 25 µL. The reaction mixture and PCR conditions for each gene are shown in Table 1. All reactions were performed using the Veriti 96-well Thermal Cycler equipment. The PCR products were separated via 1.5% agarose gel electrophoresis, and stained images were captured as described above. Then, the PCR products were purified with the Purelink PCR purification kit (Invitrogen, USA), according to the manufacturer’s instructions. The gene sequencing was performed using a BigDyeTM Termination v3.1 Cycle sequencing kit (Applied Biosystems, USA) and ABI 3500 Genetic Analyzer (Life Technologies). Contigs were generated for each gene using the BioEdit software (Ibis Biosciences, USA). The ST of the isolates was determined using the Streptococcus agalactiae MLST database (https://pubmlst.org/organisms/streptococcus-agalactiae) [49].
2.7. Whole-Genome Sequencing and Phylogenomic Analysis
The whole-genome sequencing was performed on the Illumina HiSeq 2000 platform with 125 bp paired-end reads. Sequencing libraries were constructed according to the Illumina protocol using 100 ng of genomic DNA. Annotated assemblies were produced using a pipeline described by Page et al. [50]. To determine the genomic relationship between S. agalactiae isolates from trahira and arapaima with other isolates obtained of fish in Brazil, we conducted whole-genome sequencing of SA32-17 and SA45-17 strains and compared phylogenomically with genomes deposited on GenBank (Supplementary Table S1). Reads were assembled using VelvetOptimiser v2.2.5 (https://github.com/tseemann/VelvetOptimiser) and Velvet v1.2 [51]. SSPACE [52] and GapFiller [53] were used to scaffold the contigs and sequence gaps filling, respectively. Assembly quality and statistics were evaluated with QUAST v.5.2 [54]. Automated annotation was performed using PROKKA v1.11 [55] and a Streptococcus-specific database from RefSeq [56]. Assembled contigs were also evaluated for identification and characterization of antimicrobial resistance gene using the PanViTa tool [57].
To infer a phylogenomic tree on the selected genomes via the codon tree method, which uses RAxML to align single-copy cross-genus protein families (PGFams). In parallel, we also inferred the degree of genomic relatedness among SA32-17, SA45-17 and other Brazilian Streptococcus agalactiae fish strains (Supplementary Table S1) with Average Nucleotide Identity (ANI) analysis through the MUMmer alignment method using pyANI (v3.0) [58]. Further, we used Orthofinder for tree plotting and visualization [59].
2.8. Antimicrobial Susceptibility
Streptococcus agalactiae strains were submitted to antimicrobial susceptibility tests via the disc diffusion methodology, according to the Methods for Antimicrobial Disk Susceptibility Testing of Bacteria Isolated from Aquatic Animals (VET03) guidelines, with adaptations recommended for streptococci bacteria (Group 4) [60]. The antimicrobial discs (Oxoid, UK) tested were oxytetracycline (OXY, 30 μg), florfenicol (FLO, 30 μg), sulfamethoxazole/trimethoprim (SXT, 25 μg), amoxicillin (AMO, 10 μg), norfloxacin (NOR, 10 μg) and erythromycin (ERY, 15 μg). The Streptococcus agalactiae isolates were thawed, streaked onto TSAB, and incubated at 28 ℃ for 48 h. Colonies of each strain were transferred to a sterile saline solution to achieve an absorbance value of 0.08-0.13 (OD625), as assessed using a spectrophotometer, and the bacteria were streaked onto a Müller Hinton standard agar medium (Kasvi, Italy) supplemented with 5% defibrinated sheep blood. Subsequently, three discs of antibiotics were applied per plate. After incubation at 28 ℃ for 48 h, the diameters of the inhibition halos were measured with a digital caliper (Vernier Caliper). This step was performed in triplicate. The isolates were classified as wild- (susceptible) or non-wild-types (resistant), according to Streptococcus agalactiae epidemiological cutoff values (COWT) previously calculated for isolates from fish [9,61] using normalized resistance interpretation (NRI) [62]. In addition, the criteria and standardized international terminology for defining multidrug-resistance (MDR) created by the European Centre for Disease Prevention and Control (ECDC) and the Centers for Disease Control and Prevention (CDC) were followed [63]. MDR was defined as an isolate that was non-susceptible to at least one agent in three or more antimicrobial categories.
2.9. Fish and Challenge Assay
To fulfill Koch’s postulate, trahira (average length of 13.8 ± 1.1 cm) and arapaima (19.09 ± 1.49 cm) juveniles that had previously been conditioned to the acceptance of dried food, were donated from a commercial hatchery. Fishes were acclimated in two 57 L aquaria (covered with black plastic to keep the animals under low luminosity) supplied with flow-through dechlorinated tap water (0.5 L h-1) at 28 ℃ for 15 d. The fish were subject to a 12:12 h light/dark photoperiod and were fed to satiation with Nutripiscis 42% PB (Presence, Brazil) twice a day. One Streptococcus agalactiae strain obtained of each host was randomly selected for the challenge assay.
For trahira infection, SA32-17 strain was inoculated in BHI broth and incubated at 28 ℃ for approximately 20 h under low agitation until a bacterial concentration of 1.19 × 106 CFU mL-1 was reached. Before the challenge assay, fish were anesthetized by immersion in a benzocaine solution (100 mg L-1) (Sigma-Aldrich, USA). Four fish were treated with intraperitoneal injections (infected group, G1) with 0.1 mL of SA32-17 inoculum (bacteria in BHI broth) corresponding to a dose of 105 CFU fish-1, whereas, two fish (control group, G2) were injected with 0.1 mL of sterile BHI.
For arapaima infection, SA45-17 strain was selected. This isolate was inoculated in BHI broth and incubated at 28 ℃ for approximately 19 h under low agitation until a bacterial concentration of 5.2 × 107 CFU mL-1 was reached. Before the challenge assay, fish also were anesthetized by immersion in a benzocaine solution (100 mg L-1). Eight fish were treated with intraperitoneal injections (infected group, G3) with 0.1 mL of SA45-17 inoculum corresponding to a dose of 106 CFU fish-1, whereas, eight (control group, G4) were injected with 0.1 mL of sterile BHI.
The infected fish (G1 and G3) and control groups (G2 and G4) were monitored and fed twice a day for 14 d. Dead fish were removed and subjected to bacterial isolation from the sampling of brain and kidney tissues. These tissues were aseptically collected, streaked onto TSAB, and incubated at 28 ℃ for 72 h. Then, the isolates were identified via MALDI-ToF, as previously described. The same procedure was performed in all fish that remained alive at the end of the experimental period after euthanasia with a benzocaine overdose (300 mg L-1) for detection of carrier fish. This in vivo experiment was approved by the Ethics Committee for Animal Use of the Federal University of Minas Gerais (CEUA-UFMG – 316/2019 and 143/2021).
2.10. Histopathological Analysis
Fragments of the brain, kidney, liver, spleen, and heart were collected for histopathological analysis. The tissues collected from each fish of infected and control groups were fixed in neutral buffered 10% formalin for 24h. Next, the tissues were dehydrated through immersion in ethanol solutions of increasing concentrations (70%-100%), cleared with xylene, and embedded in paraffin using a vacuum infiltration processor (TP 1020, Leica Biosystems, Germany). Tissues were sectioned in paraffin blocks at 4 µm using the semi-automated rotary microtome (RM2245, Leica Biosystems) and stained with hematoxylin-eosin (HE) [64]. Stained sections were examined using light microscopy (Olympus BX41, Japan) and photographed using an automated upright microscope system (Leica DM4000 B).
3. Results
3.1. Clinical Signs at Field Conditions and Identification of Streptococcus Agalactiae Isolates
The main clinical signs verified in the diseased trahira were lethargy, exophthalmia (Figure 1A), and corneal opacity. While in diseased arapaima were observed anorexia, lethargy, exophthalmia, corneal opacity (Figure 1B), erratic swimming and ascites.
In the bacteriological analysis, a pure culture with small, translucent, and nonhemolytic colonies was obtained from all tissues sampled from the analyzed fish. The isolates were characterized as Gram-positive cocci arranged in chains, negative in catalase and oxidase tests, and classified as Lancefield Group B. Bacterial identification using MALDI-ToF confirmed that Streptococcus agalactiae was the etiological agent associated with the disease outbreak in the wild trahira and farmed arapaima.
3.2. Genetic Diversity and Relationship Among the Isolates
All Streptococcus agalactiae isolates (SA31, SA32, SA33, SA45, SA100, SA108, SA110, SA112 and SA174 – strain codes from culture collection; one strain from each fish analyzed) obtained belonged to serotype Ib by multiplex PCR and confirmed conventional serotyping with Strep B Latex Ib.
REP-PCR of the isolates resulted in amplification of eight bands, ranging in size from 500 to 5000 bp. A single REP profile was detected, with a similarity of 94% (D = 1.0), indicating that the isolates obtained from trahira, arapaima, Amazon catfish and Nile tilapia are clonally related (Figure 2).
The MLST analysis of Streptococcus agalactiae strains obtained from trahira showed positive PCR results for six of the seven genes tested, corresponding to allelic profiles adhP: 52; atr: 31; glnA: 4; pheS: 17; sdhA: 26; and tkt: 19. Negative results were obtained for the glcK gene. The occurrence of a partially deleted glcK gene was previously characterized in Streptococcus agalactiae isolates from fish in Brazil [65]. In this manner, these isolates cannot be categorized into an ST group, instead of being determined as a non-typeable (NT) lineage. On the other hand, the isolates obtained from arapaima showed positive PCR results for all seven genes tested, corresponding to allelic profiles adhP: 52; atr: 31; glnA: 28; pheS: 17; sdhA: 26; glcK: 26 and tkt: 19. These isolates were categorized into ST-260.
3.3. Genomic Similarity and Phylogenomic Analysis
A summary of the genomic features of SA32-17 and SA45-17 is provided in Table 2. The genome project has been deposited at GenBank database under BioProject accession number PRJNA1179504.
The ANI analysis showed that SA32-17 and SA45-17 genomes was > 99% identical to other piscine Streptococcus agalactiae strains. Although the isolates showed a high similarity, in the phylogenomic analysis it is possible to observe the formation of two slightly distinct groups, one constituted with NT strains e other with CC260 (ST260 and ST927) strains. SA32-17 strain grouped with NT and SA45-17 with CC260 strains in accordance with the MLST results (Figure 3).
3.4. Antimicrobial Susceptibility and In Silico Identification of AMR Genes
In the disc diffusion assay, Streptococcus agalactiae strains exhibited zones ranging between 6 and 32 mm (Table 3). SA32, SA100, SA108, SA112 and SA174 strains were classified as MDR, nevertheless they showed non-wild-type phenotypes to three or four different categories of antibiotics. All isolates showed a wild-type phenotype to OXY and FLO, and non-wild-type to SXT. A total of 33.3%, 33.3% and 66.7% of isolates showed wild-type phenotype to AMO, NOR and ERY, respectively (Table 3).
A search on the genome sequences of SA32-17 and SA45-17 strains for AMR genes detected genes that suggest the fluoroquinolone (arlR, norB, pmrA), macrolide (RlmA(II), efrA, mreA), lincosamide (lmrD and lmrP), rifamycin (rpoB2) antibiotics resistance, and peptide antibiotic (Saga_mprF). QacJ, related to disinfecting agents and antiseptics resistance, was exclusively detected in SA32 strain, while efrB, related to macrolide resistance, was detected just in SA45 strain (Table 2).
3.5. Challenge Assay and Histopathological Analysis
Clinical diseases caused by Streptococcus agalactiae were successfully reproduced under experimental conditions (Groups 1 and 3). Anorexia, pallor of the integument, buccal hyperemia, and corneal opacity were the main clinical signs observed in infected trahira. Only one instance of mortality was observed in the infected group (72 h post-infection) (Figure 4). However, positive bacterial recovery was verified in all remaining infected fish. During the necropsy, two of the three fish that remained alive showed signs of renal congestion. Neither clinical signs nor mortalities were observed in the control group (Group 2). In histopathological analysis, trahira of control group showed normal tissue architecture. However, the infected fish exhibited meningitis (Figure 5a), inflammatory cell infiltration in brain and heart tissues, congestion and thrombosis of blood vessels in brain and liver tissues, vacuolar degeneration of hepatocytes (Figure 5b) and hemorrhagic areas in myocardium and pericardium (Figure 5c).
On the other hand, mortality was observed in arapaima infected with Streptococcus agalactiae (Group 3) on the 2nd day post-infection (dpi). In this group, the mortality rate of 100% was achieved after five days dpi (Figure 4), with positive bacterial recovery from all dead fish. Clinical signs such as anorexia, lethargy and fish at the bottom of aquarium were verified in fish approximately 48 h dpi. After 96h dpi, anorexia, corneal opacity, loss of equilibrium, despigmentation of the skin and hyperemia at the bases of the pectoral fins also observed in this group. The post mortem examination revealed liver pallor. None clinical signs were observed in the control group, however, at 5th dpi, one fish dead. Negative bacteriological result was obtained from this animal. No other mortality was observed in the control group until the end of the experimental period. In histopathology, cellular necrosis associated with the presence of Gram-positive bacteria was observed in spleen of challenged arapaima (Figure 5d).
4. Discussion
This study represents the first report concerning a natural outbreak of Streptococcus agalactiae infection in wild trahira and farmed arapaima that shared the same aquatic environment with Nile tilapia. The clinical manifestation of disease in the field and laboratory conditions were compatible with Streptococcus agalactiae infection. Anorexia, lethargy, exophthalmia, corneal opacity, and hyperemia are clinical signs commonly visualized due to this disease [4,6] and were observed in moribund fishes. Pathological changes in brain, liver, heart and spleen tissues of infected trahira and arapaima were compatible with histopathological findings in other fish infected with Streptococcus agalactiae [66,67,68].
Streptococcus agalactiae has regularly been isolated from farmed Nile tilapia during outbreaks, mainly during the summer season [4], that correspond to December to March in Brazil. Recently, this pathogen has expanded its host spectrum to other native species reared in Brazil. Tavares et al. [9] described streptococcosis outbreaks caused by Streptococcus agalactiae and other streptococci in farmed Amazon catfish (Leiarius marmoratus × Pseudoplatystoma reticulatum). Similar to this study, in Tavares et al. [9] the fish were in close contact (reared in the same culture) with Nile tilapia. Based on the previous and present results, the main risk for the interspecies transmission of Streptococcus agalactiae in Brazil seems to be the close contact, through the aquatic environment, or co-cultivation of Nile tilapia (the main Streptococcus agalactiae host in Brazil) and native species. This raises two important concerns. First, farms that adopt polyculture systems between Nile tilapia and native species could create an opportunity for the interspecies adaptation and transmission of Streptococcus agalactiae, which is an important health issue. Second, since aquaculture expansion in Brazil has been based on the use of floating cages in lakes or dam reservoirs, this could allow the transmission of Streptococcus agalactiae to wild fish. As arapaima and trahira are carnivorous—particularly in the case of arapaima, where tilapia has been used as a forage species in captive feeding [69,70], a common practice mainly among breeders, and in the case of trahiras, where sharing the same aquatic environment facilitates the capture of tilapia that may escape from the tanks—the oral transmission hypothesis is suggested. Some studies have indicated that the main route of Streptococcus agalactiae entry in fish is through the gastrointestinal tract via ingestion of contaminated water or infected fish [5,71], which corroborates our hypothesis.
Streptococcus agalactiae isolates from trahira were classified as serotype Ib and non-typeable. This genetic pattern has commonly been identified in diseased Nile tilapia reared in the Central-South region of Brazil, including the Minas Gerais state [65], where a disease outbreak in free-living trahira occurred. On the other hand, the isolates from arapaima were also classified as serotype Ib, but ST260. This genetic pattern has been identified in Northeast region of Brazil and Minas Gerais state [65], also in Nile tilapia. In this manner, the identification of the genotypes obtained in this study, regardless of the molecular typing method (MLST and REP-PCR), is similar to the strains obtained from streptococcal infections in Brazilian tilapia farms, which suggests transmission between tilapia and trahira or tilapia and arapaima. This hypothesis was corroborated by the genomic analyses of our sequenced strains when compared with other Streptococcus agalactiae strains isolates from Nile tilapia in Brazil. The high sequence similarity (> 99%) of Streptococcus agalactiae strains obtained from trahira, arapaima and tilapia suggest that transmission between these host is likely.
Streptococcus agalactiae strains presented wild-type phenotypes to the tested antibiotics, with four isolates considered as MDR (SA32, SA100, SA108 and SA174). Moreover, the identification of fluoroquinolone and macrolide resistance genes, in genomic analysis, not corroborate with disk diffusion result of SA32 and SA45 strains for norfloxacin and erythromycin, respectively. The phenomenon of multi-resistance has previously been described in Streptococcus agalactiae isolated from fish [9,72]. All isolates were classified as wild-types to florfenicol and oxytetracycline, and both antibiotics were approved for use in Brazilian aquaculture establishments for the treatment of Streptococcus agalactiae but had demonstrated therapeutic failures concerning disease control in Nile tilapia in field and laboratory conditions [73,74]. The susceptibility to the two antibiotics suggests a possible way of treating the disease for these aquatic hosts, but a future study should be carried out to prove this efficiency.
5. Conclusions
In conclusion, this is the first report of mortalities caused by Streptococcus agalactiae in free-living trahira and in arapaima farmed, of which the isolates belonged to the same genetic pattern commonly identified in Nile tilapia in Brazil. This may suggest that the commercial production of tilapia in the same aquatic environment with other native fish species may enable the adaptation of Streptococcus agalactiae to other host fish.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Strains, host, origin, and ST of Streptococcus agalactiae strains used in phylogenomic analysis.
Author Contributions
Conceptualization, RGCX, GCT and CAGL; methodology, RGCX, GAQ, TMFS, JPT, GCT and CAGL; formal analysis, GAQ and FFA; investigation, RGCX, GAQ, TMFS, JPT, and CAGL; resources, CAGL; data curation, FFA and GCT; writing—original draft preparation, RGCX, GCT and CAGL; writing—review and editing, RGCX, GAQ, TMFS, JPT, FFA, GCT and CAGL; visualization, FFA and GCT; supervision, GCT and CAGL; project administration, CAGL; funding acquisition, GCT and CAGL. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financed in part by the CNPq (Grant nº 301915/2018-0), FAPEMIG (APQ-01227-22), and CAPES (Finance Code 001).
Data Availability Statement
The genome sequences of the Streptococcus agalactiae isolated from trahira and arapaima were included in the NCBI database under BioProject accession number PRJNA1179504.
Conflicts of Interest
The authors declare no conflicts of interest.
Ethical Approval
All in vivo protocols were approved by the Ethics Committee for Animal Use at the Federal University of Minas Gerais (protocols No. 316/2019 and 143/2021).
References
- Lusiastuti, A.M.; Textor, M.; Seeger, H.; Akineden, Ö.; Zschöck, M. The Occurrence of Streptococcus agalactiae Sequence Type 261 from Fish Disease Outbreaks of Tilapia Oreochromis niloticus in Indonesia. Aquac. Res. 2014, 45, 1260–1263. [Google Scholar] [CrossRef]
- Johri, A.K.; Paoletti, L.C.; Glaser, P.; Dua, M.; Sharma, P.K.; Grandi, G.; Rappuoli, R. Group B Streptococcus: Global Incidence and Vaccine Development. Nat. Rev. Microbiol. 2006, 4, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Keefe, G.P. Streptococcus agalactiae Mastitis: A Review. Can. Vet. J. = La Rev. Vet. Can. 1997, 38, 429–437. [Google Scholar]
- Mian, G.F.; Godoy, D.T.; Leal, C.A.G.; Yuhara, T.Y.; Costa, G.M.; Figueiredo, H.C.P. Aspects of the Natural History and Virulence of S. agalactiae Infection in Nile Tilapia. Vet. Microbiol. 2009, 136, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Bowater, R.O.; Forbes-Faulkner, J.; Anderson, I.G.; Condon, K.; Robinson, B.; Kong, F.; Gilbert, G.L.; Reynolds, a.; Hyland, S.; McPherson, G.; et al. Natural Outbreak of Streptococcus agalactiae (GBS) Infection in Wild Giant Queensland Grouper, Epinephelus lanceolatus (Bloch), and Other Wild Fish in Northern Queensland, Australia. J. Fish Dis. 2012, 35, 173–186. [Google Scholar] [CrossRef]
- Evans, J.J.; Klesius, P.H.; Gilbert, P.M.; Shoemaker, C.A.; Sarawi, M.A. Al; Landsberg, J. Characterization of B-Haemolytic Group B Streptococcus agalactiae in Cultured Seabream, Sparus auratus L., and Wild Mullet, Liza klunzingeri ( Day ), in Kuwait. J. Fish Biol. 2002, 25, 505–513. [Google Scholar] [CrossRef]
- Leira, M.H.; Botelho, H.A.; Lago, A.A.; De Freitas, R.T.F.; Garcia, A.M.; De Azevedo Soares Dos Santos, H.C. Identification of Pathogens in Fish Polyculture Systems in Southern Minas Gerais, Brazil. Acta Vet. Bras. 2019, 13, 13–17. [Google Scholar] [CrossRef]
- Sebastião, F.A.; Furlan, L.R.; Hashimoto, D.T.; Pilarski, F. Identification of Bacterial Fish Pathogens in Brazil by Direct Colony PCR and 16S rRNA Gene Sequencing. Adv. Microbiol. 2015, 05, 409–424. [Google Scholar] [CrossRef]
- Tavares, G.C.; de Queiroz, G.A.; Assis, G.B.N.; Leibowitz, M.P.; Teixeira, J.P.; Figueiredo, H.C.P.; Leal, C.A.G. Disease Outbreaks in Farmed Amazon Catfish (Leiarius marmoratus x Pseudoplatystoma corruscans) Caused by Streptococcus agalactiae, S. iniae, and S. dysgalactiae. Aquaculture 2018, 495, 384–392. [Google Scholar] [CrossRef]
- Bialetzki, A.; Nakatani, K.; Sanches, P. V.; Baumgartner, G. Spatial and Temporal Distribution of Larvae and Juveniles of Hoplias malabaricus (Characiformes, Erythrinidae) in the Upper Paraná River Floodplain, Brazil. Brazilian J. Biol. 2002, 62, 211–222. [Google Scholar] [CrossRef]
- da Silva, C.A.; Oba, E.T.; Ramsdorf, W.A.; Magalhães, V.F.; Cestari, M.M.; Oliveira Ribeiro, C.A.; Silva de Assis, H.C. First Report about Saxitoxins in Freshwater Fish Hoplias malabaricus through Trophic Exposure. Toxicon 2011, 57, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.C.; Santos, A.C.G.; Ferreira, E.M.; Teófilo, T.S.; Pereira, D.M.; Costa, F.N. Aspectos Parasitológicos Da Traíra (Hoplias malabaricus) Proveniente Da Cidade de São Bento, MA. Arq. Bras. Med. Vet. e Zootec. 2017, 69, 264–268. [Google Scholar] [CrossRef]
- Benigno, R.N.M.; Knoff, M.; Matos, E.R.; Gomes, D.C.; Pinto, R.M.; Clemente, S.C.S. Morphological Aspects of Clinostomidae Metacercariae (Trematoda: Digenea) in Hoplerytrinus unitaeniatus and Hoplias malabaricus (Pisces: Erythrinidae) of the Neotropical Region, Brazil. An. Acad. Bras. Cienc. 2014, 86, 733–744. [Google Scholar] [CrossRef] [PubMed]
- IBGE Produção Da Pecuária Municipal, 50th ed.; Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, Brasil, 2023.
- Baia, R.R.J.; Florentino, A.C.; Silva, L.M.A.; Tavares-Dias, M. Patterns of the Parasite Communities in a Fish Assemblage of a River in the Brazilian Amazon Region. Acta Parasitol. 2018, 63, 304–316. [Google Scholar] [CrossRef]
- Meneguetti, D.U. de O.; Laray, M.P. de O.; Camargo, L.M.A. Primeiro Relato de Larvas de Eustrongylides Sp. (Nematoda: Dioctophymatidae) Em Hoplias malabaricus (Characiformes: Erythrinidae) No Estado de Rondônia, Amazônia Ocidental, Brasil. Rev. Pan-Amazônica Saúde 2013, 4, 55–58. [Google Scholar] [CrossRef]
- Oliveira, M.S.B.; Corrêa, L.L.; Oliveira Ferreira, D.; Neves, L.R.; Tavares-Dias, M. Records of New Localities and Hosts for Crustacean Parasites in Fish from the Eastern Amazon in Northern Brazil. J. Parasit. Dis. 2017, 41, 565–570. [Google Scholar] [CrossRef]
- Guimarães, L.; Santos, A.C. dos; Ferreira, E.; Pereira, D.; Costa, F. Microbiological Quality of Trahira Fish (Hoplias malabaricus) from Baixada Maranhense, Municipality of São Bento, MA. Arq. Inst. Biol. (Sao. Paulo). 2018, 84, 1–7. [Google Scholar] [CrossRef]
- Honczaryk, A.; Inoue, L.A.K.A. Anesthesia in Pirarucu by Benzocaine Sprays in the Gills. Ciência Rural 2010, 40, 204–207. [Google Scholar] [CrossRef]
- Imbiriba, E.P. Potencial de Criação de Pirarucu, Arapaima gigas,Em Cativeiro. Acta Amaz. 2001, 31, 299. [Google Scholar] [CrossRef]
- Fazzi-Gomes, P.F.; Melo, N.; Palheta, G.; Guerreiro, S.; Amador, M.; Ribeiro-Dos-santos, A.K.; Santos, S.; Hamoy, I. Genetic Diversity and Differentiation in Natural Populations of Arapaima gigas from Lower Amazon Revealed by Microsatellites. Genet. Mol. Res. 2017, 16, 1–6. [Google Scholar] [CrossRef]
- Ferraris Jr., C.J. Family Arapaimatidae (Bonytongues). In Check list of the freshwater fishes of South and Central America; Reis, R.E., Kullander, S.O., Ferraris, C.J., Jr., Eds.; EDIPUCRS: Porto Alegre, Brazil, 2003; pp. 32–33. [Google Scholar]
- Pereira-Filho, M.; Roubach, R. Pirarucu (Arapaima gigas). In Espécies nativas para piscicultura no Brasil; Baldisserotto, B., de C. Gomes, L., Eds.; Editora UFSM: Santa Maria, Brazil, 2010; pp. 27–56. [Google Scholar]
- Carvalho, F.R.; Casatti, L.; Manzotti, A.R.; Ravazzi, D.C.W. First Record of Arapaima gigas (Schinz, 1822) (Teleostei: Osteoglossomorpha), the “Pirarucu”, in the Upper Paraná River Basin, Southeast Brazil. Check List 2015, 11, 1–4. [Google Scholar] [CrossRef]
- Lawson, L.L.; Tuckett, Q.M.; Lawson, K.M.; Watson, C.A.; Hill, J.E. Lower Lethal Temperature for Arapaima Arapaima gigas: Potential Implications for Culture and Establishment in Florida. N. Am. J. Aquac. 2015, 77, 497–502. [Google Scholar] [CrossRef]
- Miranda-Chumacero, G.; Wallace, R.; Calderón, H.; Calderón, G.; Willink, P.; Guerrero, M.; Siles, T.M.; Lara, K.; Chuqui, D. Distribution of Arapaima (Arapaima gigas) (Pisces: Arapaimatidae) in Bolivia: Implications in the Control and Management of a Non-Native Population. BioInvasions Rec. 2012, 1, 129–138. [Google Scholar] [CrossRef]
- Ohs, C.; Hill, J.; Wright, S.; Giddings, H.M.; Durland Donahou, A. Candidate Species for Florida Aquaculture: Arapaima Arapaima gigas. Edis 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Wyman-Grothem, K.; Castello, L.; DTBS, C.; CRC, D.; ALB, M.; Patoka, J.; Stewart, D.; Watson, C. Invasion Risk to the United States from Arapaima spp. Hinges on Climate Suitability. Aquac. Environ. Interact. 2024, 16, 175–188. [Google Scholar] [CrossRef]
- Drumond, G.V.F.; Caixeiro, A.P. de A.; Tavares-Dias, M.; Marcon, J.L.; Affonso, E.G. Características Bioquímicas e Hematológicas Do Pirarucu Arapaima gigas Schinz, 1822 (Arapaimidae) de Cultivo Semi-Intensivo Na Amazônia. Acta Amaz. 2010, 40, 591–595. [Google Scholar] [CrossRef]
- Cruz, M.G. da; Jerônimo, G.T.; Bentes, S.P.C.; Gonçalves, L.U. Trichlorfon Is Effective against Dawestrema cycloancistrium and Does Not Alter the Physiological Parameters of Arapaima (Arapaima gigas): A Large Neotropical Fish from the Amazon. J. Fish Dis. 2022, 45, 203–212. [Google Scholar] [CrossRef]
- Dias, M.K.R.; Sampaio, L.S.; Proietti-Junior, A.A.; Yoshioka, E.T.O.; Rodrigues, D.P.; Rodriguez, A.F.R.; Ribeiro, R.A.; Faria, F.S.E.D.V.; Ozório, R.O.A.; Tavares-Dias, M. Lethal Dose and Clinical Signs of Aeromonas hydrophila in Arapaima gigas (Arapaimidae), the Giant Fish from Amazon. Vet. Microbiol. 2016, 188, 12–15. [Google Scholar] [CrossRef]
- Ribeiro, M. da S.; da Fonseca, F.A.L.; de Queiroz, M.N.; Affonso, E.G.; da Conceição, L.E.C.; Gonçalves, L.U. Fish Protein Hydrolysate as an Ingredient in Diets for Arapaima gigas Juveniles. Bol. do Inst. Pesca 2017, 43, 85–92. [Google Scholar] [CrossRef]
- Marinho, R.G.B.; Tavares-Dias, M.; Dias-Grigório, M.K.R.; Neves, L.R.; Yoshioka, E.T.O.; Boijink, C.L.; Takemoto, R.M. Helminthes and Protozoan of Farmed Pirarucu (Arapaima gigas) in Eastern Amazon and Host-Parasite Relationship. Arq. Bras. Med. Vet. e Zootec. 2013, 65, 1192–1202. [Google Scholar] [CrossRef]
- Andrade-Porto, S.M.; Cárdenas, M.Q.; Martins, M.L.; Oliveira, J.K.Q.; Pereira, J.N.; Araújo, C.S.O.; Malta, J.C.O. First Record of Larvae of Hysterothylacium (Nematoda: Anisakidae) with Zoonotic Potential in the Pirarucu Arapaima gigas (Osteichthyes: Arapaimidae) from South America. Brazilian J. Biol. 2015, 75, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Proietti-Junior, A.A.; Lima, L.S.; Roges, E.M.; Rodrigues, Y.C.; Lima, K.V.B.; Rodrigues, D.P.; Tavares-Dias, M. Experimental Co-Infection by Aeromonas hydrophila and Aeromonas jandaei in Pirarucu Arapaima gigas (Pisces: Arapaimidae). Aquac. Res. 2021, 52, 1688–1696. [Google Scholar] [CrossRef]
- Choresca Jr., C.H.; Gomez, D.K.; Shin, S.P.; Kim, J.H.; Han, J.E.; Jun, J.W.; Park, S.C. Molecular Detection of Edwardsiella tarda with gyrB Gene Isolated from Pirarucu, Arapaima gigas Which Is Exhibited in an Indoor Private Commercial Aquarium. African J. Biotechnol. 2011, 10, 848–850. [Google Scholar]
- Kodama, H.; Nakanishi, Y.; Yamamoto, F.; Mikami, T.; Izawa, H.; Imagawa, T.; Hashimoto, Y.; Kudo, N. Salmonella arizonae Isolated from a Pirarucu, Arapaima gigas Cuvier, with Septicaemia. J. Fish Dis. 1987, 10, 509–512. [Google Scholar] [CrossRef]
- Choresca Jr., C.H.; Kim, J.H.; Gomez, D.K.; Jang, H.; Joh, S.J.; Park, S.C. Isolation of Serratia fonticola from Pirarucu Arapaima gigas. Korean J. Vet. Res. 2008, 48, 89–92. [Google Scholar]
- Barbanti, A.C.C.; do Rosário, A.E.C.; da Silva Maia, C.R.M.; Rocha, V.P.; Costa, H.L.; Trindade, J.M.; Nogueira, L.F.F.; Rosa, J.C.C.; Ranzani-Paiva, M.J.T.; Pilarski, F.; et al. Genetic Characterization of Lactococcosis-Causing Bacteria Isolated from Brazilian Native Fish Species. Aquaculture 2024, 593, 741305. [Google Scholar] [CrossRef]
- Serrano-Martínez, E.; Verónica, C.P.; Marco, Q.H.; Gina, C. V.; Jorge, L.Q. Isolation of Bacteria and Fungi in Tissues of Paiche (Arapaima gigas) Reared in Captivity. Rev. Investig. Vet. del Peru 2014, 25, 117–122. [Google Scholar]
- Assis, G.B.N.; Pereira, F.L.; Zegarra, A.U.; Tavares, G.C.; Leal, C.A.; Figueiredo, H.C.P. Use of MALDI-TOF Mass Spectrometry for the Fast Identification of Gram-Positive Fish Pathogens. Front. Microbiol. 2017, 8, 1492. [Google Scholar] [CrossRef]
- Poyart, C.; Tazi, A.; Réglier-Poupet, H.; Billoët, A.; Tavares, N.; Raymond, J.; Trieu-Cuot, P. Multiplex PCR Assay for Rapid and Accurate Capsular Typing of Group B Streptococci. J. Clin. Microbiol. 2007, 45, 1985–1988. [Google Scholar] [CrossRef]
- Costa, F.A.A.; Leal, C.A.G.; Leite, R.C.; Figueiredo, H.C.P. Genotyping of Streptococcus dysgalactiae Strains Isolated from Nile Tilapia, Oreochromis niloticus (L.). J. Fish Dis. 2014, 37, 463–469. [Google Scholar] [CrossRef]
- Dice, L.R. Measures of the Amount of Ecologic Association between Species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Ishii, S.; Sadowsky, M.J. Applications of the Rep-PCR DNA Fingerprinting Technique to Study Microbial Diversity, Ecology and Evolution. Environ. Microbiol. 2009, 11, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R.; Gaston, M.A. Numerical Index of the Discriminatory Ability of Typing Systems: An Application of Simpson’s Index of Diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [PubMed]
- Pinto, F.R.; Melo-Cristino, J.; Ramirez, M. A Confidence Interval for the Wallace Coefficient of Concordance and Its Application to Microbial Typing Methods. PLoS One 2008, 3, e3696. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.; Bohnsack, J.F.; Takahashi, S.; Oliver, K.A.; Chan, M.-S.; Kunst, F.; Glaser, P.; Rusniok, C.; Crook, D.W.M.; Harding, R.M.; et al. Multilocus Sequence Typing System for Group B Streptococcus. J. Clin. Microbiol. 2003, 41, 2530–2536. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software, the PubMLST.Org Website and Their Applications. Wellcome Open Res. 2018, 3, 1–20. [Google Scholar] [CrossRef]
- Page, A.J.; De Silva, N.; Hunt, M.; Quail, M.A.; Parkhill, J.; Harris, S.R.; Otto, T.D.; Keane, J.A. Robust High-Throughput Prokaryote de Novo Assembly and Improvement Pipeline for Illumina Data. Microb. genomics 2016, 2, e000083. [Google Scholar] [CrossRef]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de Novo Short Read Assembly Using de Bruijn Graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Boetzer, M.; Henkel, C. V; Jansen, H.J.; Butler, D.; Pirovano, W. Scaffolding Pre-Assembled Contigs Using SSPACE. Bioinformatics 2011, 27, 578–579. [Google Scholar] [CrossRef]
- Boetzer, M.; Pirovano, W. Toward Almost Closed Genomes with GapFiller. Genome Biol. 2012, 13, R56. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, K.D.; Tatusova, T.; Brown, G.R.; Maglott, D.R. NCBI Reference Sequences (RefSeq): Current Status, New Features and Genome Annotation Policy. Nucleic Acids Res. 2012, 40, D130–5. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.L.N.; Ariute, J.C.; Rodrigues da Costa, F.M.; Benko-Iseppon, A.M.; Barh, D.; Azevedo, V.; Aburjaile, F. PanViTa: Pan Virulence and ResisTance Analysis. Front. Bioinforma. 2023, 3. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and Taxonomy in Diagnostics for Food Security: Soft-Rotting Enterobacterial Plant Pathogens. Anal. Methods 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic Orthology Inference for Comparative Genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef]
- CLSI VET03: Methods for Antimicrobial Broth Dilution and Disk Diffusion Susceptibility Testing of Bacteria Isolated from Aquatic Animals; Clinical and Laboratory Standards Institute: Wayne, USA, 2020.
- Leal, C.A.G.; Silva, B.A.; Colombo, S.A. Susceptibility Profile and Epidemiological Cut-Off Values Are Influenced by Serotype in Fish Pathogenic Streptococcus agalactiae. Antibiotics 2023, 12. [Google Scholar] [CrossRef]
- Kronvall, G.; Smith, P. Normalized Resistance Interpretation, the NRI Method. APMIS 2016, 124, 1023–1030. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Luna, L.G. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology, 3rd ed. Blakiston Division: New York, 1968.
- Barony, G.M.; Tavares, G.C.; Pereira, F.L.; Carvalho, A.F.; Dorella, F.A.; Leal, C.A.G.; Figueiredo, H.C.P. Large-Scale Genomic Analyses Reveal the Population Structure and Evolutionary Trends of Streptococcus agalactiae Strains in Brazilian Fish Farms. Sci. Rep. 2017, 7, 13538. [Google Scholar] [CrossRef]
- Laith, A.A.; Ambak, M.A.; Hassan, M.; Sheriff, S.M.; Nadirah, M.; Draman, A.S.; Wahab, W.; Ibrahim, W.N.W.; Aznan, A.S.; Jabar, A.; et al. Molecular Identification and Histopathological Study of Natural Streptococcus agalactiae Infection in Hybrid Tilapia (Oreochromis niloticus). Vet. world 2017, 10, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Delamare-Deboutteville, J.; Bowater, R.; Condon, K.; Reynolds, A.; Fisk, A.; Aviles, F.; Barnes, A.C. Infection and Pathology in Queensland Grouper, Epinephelus lanceolatus, (Bloch), Caused by Exposure to Streptococcus agalactiae via Different Routes. J. Fish Dis. 2015, 38, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Ortega Asencios, Y.; Barreiro Sánchez, F.; Bueno Mendizábal, H.; Huancaré Pusari, K.; Ostos Alfonso, H.; Manchego Sayán, A.; Pereira Figueiredo, M.A.; Gómez Manrique, W.; de Andrade Belo, M.A.; Sandoval Chaupe, N. First Report of Streptococcus agalactiae Isolated from Oreochromis niloticus in Piura, Peru: Molecular Identification and Histopathological Lesions. Aquac. Reports 2016, 4, 74–79. [Google Scholar] [CrossRef]
- Gonzalez-Callirgos, L.; da Costa, J.I.; Yunis-Aguinaga, J. Economic Evaluation of Arapaima gigas Production in Earth Ponds: Case Study of a Small Fish Farm at San Martin-Peru. Bol. do Inst. Pesca 2023, 49, 1–8. [Google Scholar] [CrossRef]
- Venturieri, R.; Bernardino, G. Pirarucu: Espécie Ameaçada Pode Ser Salva Através Do Cultivo. Panor. da Aquicultura 1999, 9. [Google Scholar]
- Iregui, C.A.; Comas, J.; Vásquez, G.M.; Verján, N. Experimental Early Pathogenesis of Streptococcus agalactiae Infection in Red Tilapia Oreochromis spp. J. Fish Dis. 2016, 39, 205–215. [Google Scholar] [CrossRef]
- Chideroli, R.T.; Amoroso, N.; Mainardi, R.M.; Suphoronski, S.A.; de Padua, S.B.; Alfieri, A.F.; Alfieri, A.A.; Mosela, M.; Moralez, A.T.P.; de Oliveira, A.G.; et al. Emergence of a New Multidrug-Resistant and Highly Virulent Serotype of Streptococcus agalactiae in Fish Farms from Brazil. Aquaculture 2017, 479, 45–51. [Google Scholar] [CrossRef]
- de Oliveira, T.F.; Queiroz, G.A.; Teixeira, J.P.; Figueiredo, H.C.P.; Leal, C.A.G. Recurrent Streptoccoccus agalactiae Infection in Nile Tilapia (Oreochromis niloticus) Treated with Florfenicol. Aquaculture 2018, 493, 51–60. [Google Scholar] [CrossRef]
- Faria, F.C.; Leal, C. a G.; Carvalho-Castro, G. a; Leite, R.C.; Figueiredo, H.C.P. Carrier State Induced by Oxytetracycline Therapy against Streptococcosis in Nile Tilapia, Oreochromis niloticus (L.). J. Fish Dis. 2014, 37, 853–857. [Google Scholar] [CrossRef]
Figure 1.
Natural Streptococcus agalactiae infection in native fish species. A. Diseased wild trahira with exophthalmia (arrow) found dead during the outbreak; B. Diseased arapaima with exophtalmia and corneal opacity, alive in the tank.
Figure 1.
Natural Streptococcus agalactiae infection in native fish species. A. Diseased wild trahira with exophthalmia (arrow) found dead during the outbreak; B. Diseased arapaima with exophtalmia and corneal opacity, alive in the tank.

Figure 2.
REP-PCR results for five Streptococcus agalactiae strains isolated from trahira, arapaima, Amazon catfish and Nile tilapia. The dendrogram was constructed using Dice’s coefficient and the UPGMA method.
Figure 2.
REP-PCR results for five Streptococcus agalactiae strains isolated from trahira, arapaima, Amazon catfish and Nile tilapia. The dendrogram was constructed using Dice’s coefficient and the UPGMA method.

Figure 3.
Whole-genome sequence RAxML phylogenetic tree constructed using SA32-17 and SA45-17 strains together other Brazilian Streptococcus agalactiae strains deposited in the GenBank database (bootstrap test, 1000 replicates). Two distantly related groups of isolates are visualized one on the right side (CC260, marked with *) and one on the left side (NT) of the figure. Bootstrap values are presented as color gradients at the branches for better visualization.
Figure 3.
Whole-genome sequence RAxML phylogenetic tree constructed using SA32-17 and SA45-17 strains together other Brazilian Streptococcus agalactiae strains deposited in the GenBank database (bootstrap test, 1000 replicates). Two distantly related groups of isolates are visualized one on the right side (CC260, marked with *) and one on the left side (NT) of the figure. Bootstrap values are presented as color gradients at the branches for better visualization.
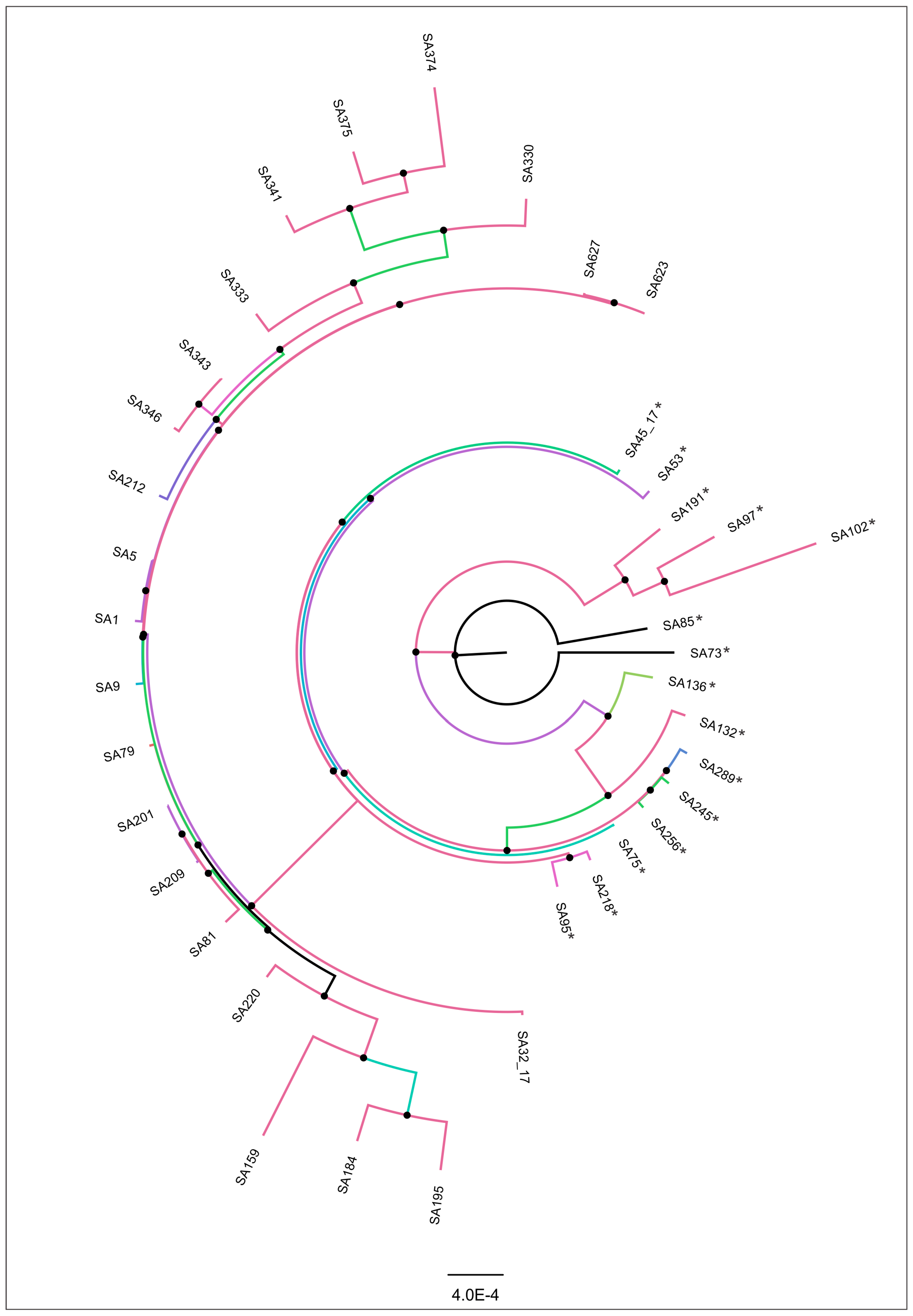
Figure 4.
Mortality rate of the trahira and arapaima challenged with Streptococcus agalactiae during the experimental period.
Figure 4.
Mortality rate of the trahira and arapaima challenged with Streptococcus agalactiae during the experimental period.
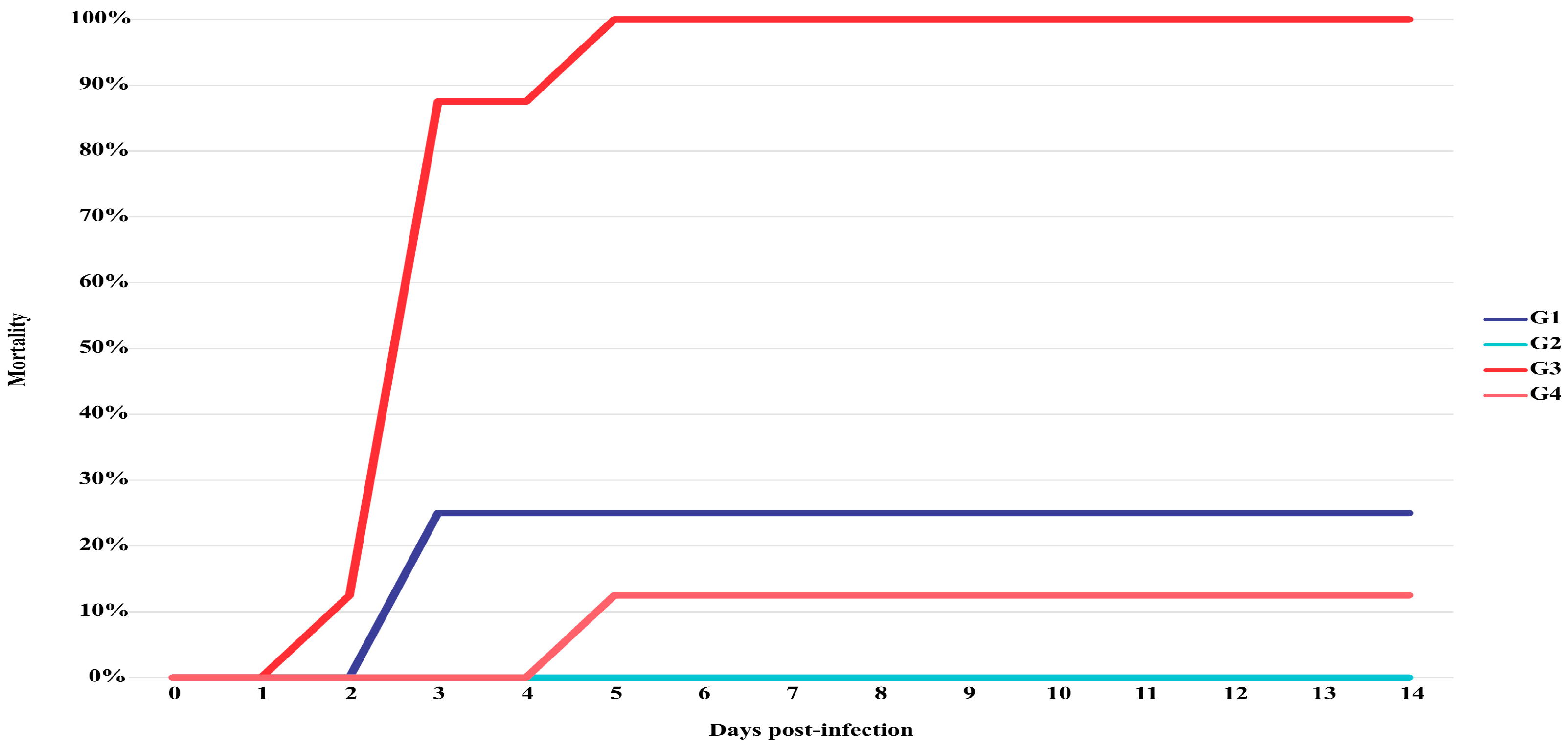
Figure 5.
Histology of challenged trahira (A-C) and arapaima (D). A. Brain – meningitis was and obvious sign characterized by thickening with congested blood vessels and inflammatory cells infiltration; B. Liver – congestion and thrombosis of portal blood vessel accompanied with hepatic sinusoids with marked vacuolar degeneration change of hepatocytes; C. Heart – Accumulation of inflammatory cells (arrows) and hemorrhagic areas in myocardium and pericardium; D. Spleen – cellular necrosis (picnosis = arrow) in white pulp associated with the presence of Gram-positive bacteria (arrow head).
Figure 5.
Histology of challenged trahira (A-C) and arapaima (D). A. Brain – meningitis was and obvious sign characterized by thickening with congested blood vessels and inflammatory cells infiltration; B. Liver – congestion and thrombosis of portal blood vessel accompanied with hepatic sinusoids with marked vacuolar degeneration change of hepatocytes; C. Heart – Accumulation of inflammatory cells (arrows) and hemorrhagic areas in myocardium and pericardium; D. Spleen – cellular necrosis (picnosis = arrow) in white pulp associated with the presence of Gram-positive bacteria (arrow head).
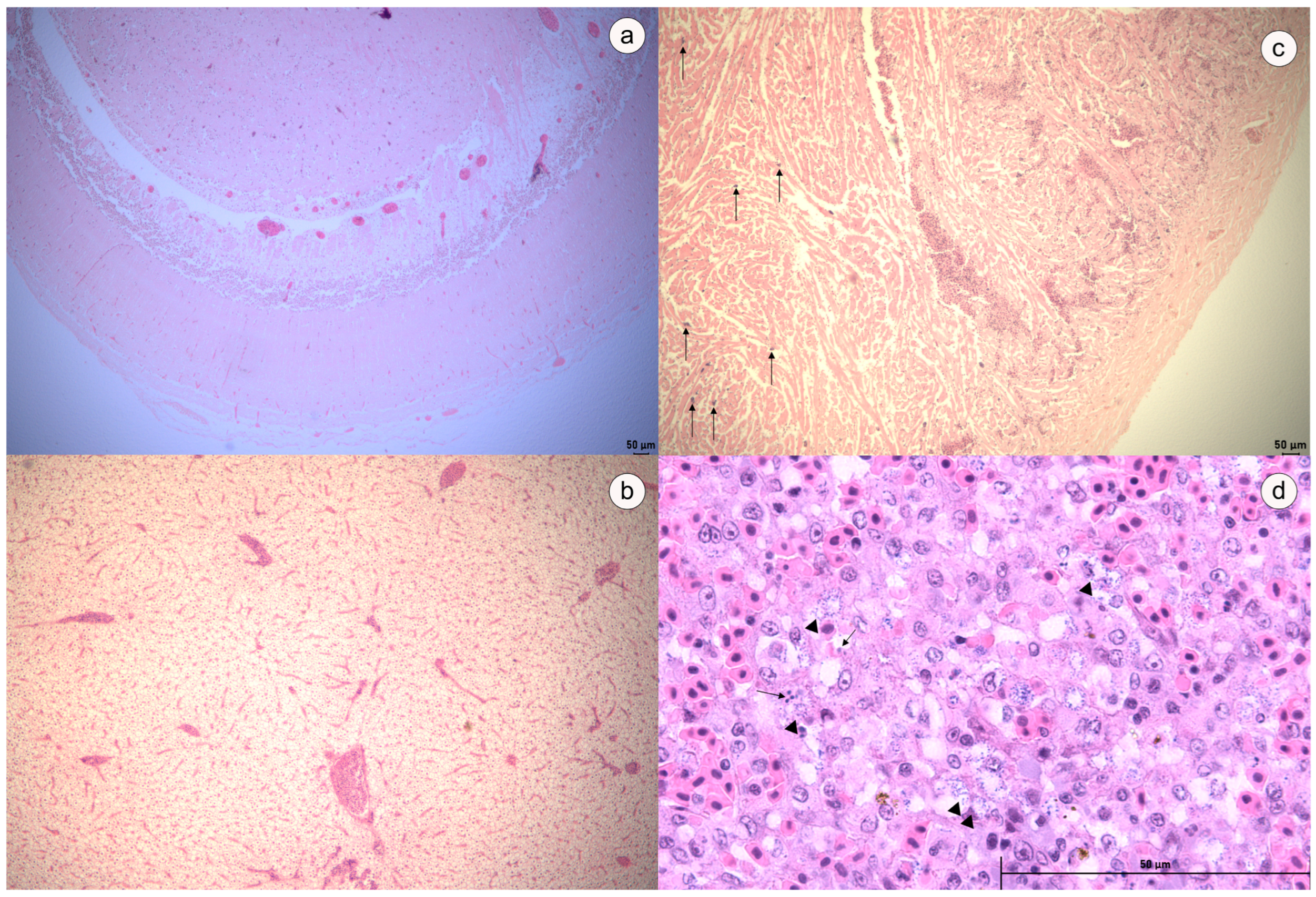

Table 1.
PCR mix composition (final reaction volume of 25 μL) and thermal cycling conditions used in the MLST analysis of Streptococcus agalactiae.
Table 1.
PCR mix composition (final reaction volume of 25 μL) and thermal cycling conditions used in the MLST analysis of Streptococcus agalactiae.
| Genes | Reaction mixture | PCR conditions |
|---|---|---|
|
atr glnA tkt |
Buffer 10X: 2.5 μL 25 mM MgCl2: 1.5 μL 10 mM dNTP: 0.5 μL 10 mM each primer: 0.5 μL Taq DNA Polymerase: 0.25 μL |
Initial step: 95 ℃ for 5 min Cycling: 35 cycles of 95 ℃ for 1 min, 56 ℃ for 45 s, and 72 ℃ for 1 min; Final elongation: 72 ℃ for 15 min |
|
glcK pheS |
Buffer 10X: 2.5 μL 25 mM MgCl2: 1.5 μL 10 mM dNTP: 0.5 μL 10 mM each primer: 0.5 μL Taq DNA Polymerase: 0.25 μL |
Initial step: 95 ℃ for 15 min Cycling: 35 cycles of 95 ℃ for 1 min, 56 ℃ for 1 min, and 72 ℃ for 1 min; Final elongation: 72 ℃ for 10 min |
|
adhP sdhA |
Buffer 10X: 2.5 μL 25 mM MgCl2: 1.0 μL 10 mM dNTP: 0.25 μL 10 mM each primer: 0.2 μL Taq DNA Polymerase: 0.05 μL |
Initial step: 95 ℃ for 5 min Cycling: 35 cycles of 95 ℃ for 1 min, 56 ℃ for 40 s, and 72 ℃ for 1 min; Final elongation: 72 ℃ for 10 min |
Table 2.
Main genomic features of SA32-17 and SA45-17 strains.
| SA32-17 | SA45-17 | |
|---|---|---|
| Genome statistics by QUAST tool | ||
| #contigs (>= 0 bp) | 29 | 23 |
| #contigs (>= 1000 bp) | 28 | 23 |
| Total length (>= 0 bp) | 1,824,566 | 1,817,806 |
| Total length (>= 1000 bp) | 1,823,865 | 1,817,806 |
| #contigs | 29 | 23 |
| Largest contig | 368,081 | 283,920 |
| Total length | 1,824,566 | 1,817,806 |
| G+C (%) | 35.34 | 35.32 |
| N50 | 136,190 | 124,698 |
| N90 | 42,369 | 56,228 |
| auN | 192,272.4 | 144,941.0 |
| L50 | 4 | 5 |
| L90 | 13 | 15 |
| # N’s per 100 kbp | 1.32 | 0.00 |
| Attributes | ||
| Genes (total) | 1,957 | 1,922 |
| Coding sequences (total) | 1,852 | 1,834 |
| Pseudogenes | 29 | 26 |
| rRNA | 11 | 10 |
| tRNA | 64 | 51 |
| tmRNA | 1 | 1 |
| AMR gene detection using PanViTa tool | ||
| arlR (%) | 50.4 | 52.2 |
| RlmA(II) (%) | 53.2 | 53.2 |
| lmrD (%) | 61.2 | 61.2 |
| efrA (%) | 52.8 | 52.6 |
| norB (%) | 59.7 | 59.5 |
| pmrA (%) | 57.5 | 57.3 |
| mreA (%) | 99.7 | 99.4 |
| efrB (%) | 0.0 | 53.3 |
| lmrP (%) | 54.4 | 54.4 |
| Saga_mprF (%) | 99.4 | 99.1 |
| rpoB2 (%) | 52.2 | 52.2 |
| qacJ (%) | 58.9 | 0.0 |
Table 3.
Disc diffusion results of GBS strains from diseased trahira. The inhibition zone diameters (mm) of each strain were measured after growth in Muller-Hinton agar enriched with 5% defibrinated sheep blood at 28 ℃ for 48 h. WT (wild type) and NWT (non-wild type), considered susceptible and resistant, respectively. Gray boxes represent NWT isolates.
Table 3.
Disc diffusion results of GBS strains from diseased trahira. The inhibition zone diameters (mm) of each strain were measured after growth in Muller-Hinton agar enriched with 5% defibrinated sheep blood at 28 ℃ for 48 h. WT (wild type) and NWT (non-wild type), considered susceptible and resistant, respectively. Gray boxes represent NWT isolates.
| Antimicrobial agent* | ||||||
|---|---|---|---|---|---|---|
| Strain ID | SXTa | OXYb | FLOb | AMOb | NORa | ERYa |
| SA31 | 20 (NWT) | 28 (WT) | 30 (WT) | 28 (NWT) | 30 (WT) | 25 (WT) |
| SA32 | 19 (NWT) | 28 (WT) | 30 (WT) | 28 (NWT) | 20 (WT) | 17 (NWT) |
| SA33 | 18 (NWT) | 30 (WT) | 32 (WT) | 32 (WT) | 10 (NWT) | 32 (WT) |
| SA45 | 6 (NWT) | 25 (WT) | 24 (WT) | 27 (NWT) | 16 (WT) | 27 (WT) |
| SA100 | 6 (NWT) | 27 (WT) | 25 (WT) | 27 (NWT) | 6 (NWT) | 27 (WT) |
| SA108 | 6 (NWT) | 25 (WT) | 26 (WT) | 31 (WT) | 6 (NWT) | 24 (NWT) |
| SA110 | 6 (NWT) | 23 (WT) | 26 (WT) | 34 (WT) | 6 (NWT) | 27 (WT) |
| SA112 | 6 (NWT) | 25 (WT) | 25 (WT) | 27 (NWT) | 6 (NWT) | 28 (WT) |
| SA174 | 6 (NWT) | 25 (WT) | 24 (WT) | 28 (NWT) | 6 (NWT) | 22 (NWT) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



