
Submitted:
02 April 2024
Posted:
03 April 2024
You are already at the latest version
Abstract
Piscine lactococcosis is a disease caused by Lactococcus garvieae, L. petauri and L. formosensis. It is considered to be an emerging disease and poses a risk to global aquaculture production. Studies of the genetic diversity of Lactococcus spp. strains have epidemiological importance since they provide information on the genetic relationships between isolates and indicate geographic and temporal distribution. The aim of this study was to evaluate the genetic diversity of Brazilian Lactococcus spp. isolates obtained from native fish species using PCR-based DNA fingerprinting techniques. For this, 36 isolates were selected and submitted to gyrB sequencing, molecular serotyping, REP-, BOX-, and RAPD-PCR approaches. The discriminatory power and the congruence between the typing methods were calculated. After the correct taxonomic discrimination of the isolate, a total of 14, 5 and 17 isolates were reclassified as L. garvieae, L. formosensis and L. petauri, respectively. L. garvieae and L. petauri are related to serotype I, while L. formosensis strains were classified as serotype II. The typing methods tested are useful for evaluating the genetic diversity of isolates and a comprehensive indication of the diversity was found. Heterogeneous and homogeneous populations were observed for L. garvieae and L. petauri, respectively. RAPD-PCR demonstrated a greater discriminatory power for L. formosensis and L. petauri, while REP-PCR showed better results for L. garvieae strains, thus these methodologies are recommended for genotyping these bacterial species. In conclusion, Brazilian isolates of Lactococcus spp. obtained from native fish species constitute a genetically diverse population and the techniques used in this study allowed a better genetic characterization of the isolates.
Keywords:
fish
; lactococcosis
; identification
; serotyping
; genotyping
1. Introduction
Piscine lactococcosis is a disease that affects both marine and freshwater fish species, whether on farms or in free-living environments [1,2]. Disease outbreaks have occurred in the Americas [2,3,4,5,6,7,8], Africa [9,10], Europe [11,12,13], Asia [14,15,16,17] and Oceania [18]. One of the most impactful was the outbreak that occurred in 2020, in the state of California (USA), which resulted in the culling of more than 3.2 million fish, with L. petauri being identified as causing the disease [8].
Morphologically, lactococci are 0.5–1.5 μm Gram-positive, chain-forming, normally mesophilic cocci that ferment lactose-producing lactic acid and have complex growth requirements [19]. For a long time, Lactococcus garvieae was considered the main causative agent of piscine lactococcosis; however, in recent studies, it has been revealed that piscine lactococcosis can also be caused by closely related species such as L. petauri and L. formosensis [5,20]. These three species have great similarity in relation to their clinical manifestations and genetics, and are almost identical in the polymerase chain reaction (PCR), based on 16s rRNA gene sequencing, biochemical and MALDI-TOF identification, which generates an unreliable result at the species level [1,20,21]. Currently, the gyrB gene sequencing is the best technique for discriminating isolates from the genus Lactococcus [5].
Several molecular genetic diversity methods have already been applied to discriminate strains previously classified as Lactococcus garvieae, such as pulse-field gel electrophoresis (PFGE), repetitive sequence-based PCR (rep-PCR), random amplification of polymorphic DNA-PCR (RAPD), multilocus sequence analysis (MLSA) and multilocus sequence typing (MLST) [8,22,23]. Rapid and inexpensive typing using PCR-based techniques such as rep-PCR can be used to trace gene parentage, while potentially more discriminatory techniques such as MLST help to confirm the results [24]. The combination of these methods has already been used to genotype Lactococcus spp. strains and demonstrates the existence of genetic heterogeneity within this species [22,25]. Molecular genotyping studies of this pathogen permits the acquisition of epidemiological data and provides temporal and geographic records of the pathogen, as well as the ability to track the genetic relationship of bacterial populations [26].
In recent years, piscine lactococcosis has been detected in Brazil in different species of fish, such as Pseudoplatystoma sp. [3,4,27], Lophiosilurus alexandri [28], Colossoma macropomum [29], Arapaima gigas [30], Oreochromis niloticus [3,5,27] and ornamental fishes [31], and an increasing number of outbreaks have been occurring in the country. However, little is known about the genetic diversity of Lactococcus spp. strains obtained from native fish species. Therefore, the aims of this study were to evaluate the genetic diversity of Lactococcus spp. strains isolated from native Brazilian fish species using PCR-based DNA fingerprinting techniques and evaluate the discriminatory power and congruence of typing methods.
2. Materials and Methods
2.1. Isolates
A total of 36 Lactococcus spp. strains obtained from nine species of native fish (Arapaima gigas, Brycon amazonicus, Colossoma macropomum, Hoplias macrophtalmus, Lophiosilurus alexandri, Phractocephalus hemioliopterus, Pseudoplatystoma corruscans, Pseudoplatystoma fasciatum and a hybrid of Pseudoplatystoma) reared between 2012 and 2023 on commercial farms in six Brazilian states (Amazonas, Bahia, Mato Grosso do Sul, Minas Gerais, Pará and São Paulo) were selected (Table 1, Figure 1). These isolates were previously identified as Lactococcus garvieae using matrix-assisted laser desorption ionization time-of-flight (MALDI-ToF) mass spectrometry (Bruker Daltonics) and then stored until use at -70 °C in BHI broth with 15% glycerol.2.2. Lactococcus spp. identification
2.2.1. DNA Extraction
The selected Lactococcus spp. isolates were thawed, inoculated on MRS (Man, Rogosa, Sharpe) agar, and incubated at 28 ºC for 3 days. Subsequently, colonies were collected and diluted in 180 µL of lysis solution (20 mg mL-1 lysozyme; 20 mM Tris-HCl, pH 8; 2 mM EDTA; and 1.2% Triton) and incubated overnight at 37 ºC. Bacterial DNA was extracted using the Maxwell 16 Tissue DNA purification kit, according to the manufacturer’s recommendations. The amount of DNA extracted was quantified via spectrophotometry (Nanodrop, Thermo Scientific). The DNA samples were stored at -20 ºC until use.
2.2.2. gyrB Sequencing
Molecular identification of the Lactococcus spp. strains at the species level was performed using gyrB gene sequencing, as previously described by Egger et al. [5] with some modifications. The PCR reaction was carried out using the GoTaq PCR Core System kit (Promega), consisting of a mixture with 150 ng of DNA template (3 µL) plus 22 µL of PCR mix (1X PCR buffer, 0.5 µM of each primer [gyrB-F: CATGCTGGTGGTAAATTTGG, gyrB-R: GTCATCCATTTCTCCTAAACC], 0.2 mM dNTP, 2.5 mM MgCl2, 0.05 UµL-1 Taq DNA polymerase, and sterile water). PCR amplification was carried out in a Veriti 96-well thermal cycler (Applied Biosystems), using the following conditions: 1 cycle of 94 ºC for 5 minutes for denaturation, followed by 35 cycles of 94 ºC for 45 seconds, 58 ºC for 45 seconds and 72 ºC for 70 seconds; and the final extension at 72 ºC for 5 minutes. The amplicons were separated via capillary electrophoresis (QIAxcel Advanced, Qiagen), according to the manufacturer’s recommendations.
After visualizing the size of the amplicons, the PCR products were purified using the Agencourt AMPure® XP kit (Beckman Coulter), according to the manufacturer’s instructions. The sequencing reaction was performed using the BigDye Terminator Cycle sequencing kit (Applied Biosystems) and the run was performed on the ABI 3500 Genetic Analyzer (Applied Biosystems). The contigs were assembled and curated using BioEdit software (Ibis BioSciences). Available gyrB gene sequences of isolates of L. garvieae (strains Lg2 and ATCC 49156), L. petauri (strains B1726 and 159469) and L. formosensis (strain I4/6O and NBRC 109475) that had been previously confirmed at species level were downloaded from NCBI. All the gyrB gene sequences were aligned using the MEGA v.11 (Molecular Evolutionary Genetics Analysis Version 11) program [32], using the maximum composite likelihood method [33] to calculate the evolutionary distances. The phylogenetic representation was assigned based on the evolutionary history inferred by the neighbor-joining method [34] with a bootstrap analysis with 1,000 replicates to check the stability of the tree generated. The iTOL software (http://itol.embl.de/) was used to edit the phylogenetic tree.
2.3. Molecular Serotyping
The selected isolates were subjected to serotyping using a multiplex reaction, as previously described by Ohbayashi et al. [15] with some modifications. In summary, the reaction was performed with the GoTaq PCR Core System kit in a total volume of 25 μL. The reaction mixture consisted of 100 ng of template DNA (2 μL) plus 23 μL of PCR mix (1X PCR buffer, 0.5 μM of each primer [LGD-F: GGATTGAACTTCCTGCCACA, LGD-R: ATCCTTGAGGACAACGAAGG], 0.2 mM dNTP, 2.5 mM MgCl2, 0.06 U μL-1 Taq DNA polymerase, and sterile water). PCR amplification was performed in a Veriti 96-well thermal cycler, using the following conditions: 1 cycle of 95 ºC for 5 minutes for denaturation, followed by 30 cycles at 95 ºC for 30 seconds, 55 ºC for 30 seconds and 72 ºC for 90 seconds; and the final extension at 72 ºC for 7 minutes. The amplicons were separated via capillary electrophoresis (QIAxcel Advanced), according to the manufacturer’s recommendations.
2.4. Genetic Typing Using PCR-Based DNA Fingerprinting Techniques
2.4.1. REP-PCR
The genetic diversity of the Lactococcus spp. strains was evaluated by amplifying palindromic extragenic sequences (REP-PCR), as previously described by Costa et al. [26] with some modifications. The reaction was carried out using the GoTaq PCR Core System kit in a total reaction volume of 25 µL. The reaction mixture consisted of 300 ng of template DNA (6 µL) plus 19 µL of PCR mix (1X PCR buffer, 0.5 µM of GTG5 primer [GTGGTGGTGGTGGTG], 0.2 mM dNTP, 3.0 mM MgCl2, 0.1 U µL-1 Taq DNA polymerase, and sterile water). PCR amplification was carried out in a Veriti 96-well thermal cycler, using the following conditions: 1 cycle at 95 ºC for 5 minutes for denaturation, followed by 30 cycles at 95 ºC for 30 seconds, 45 ºC for 1 minute and 72 ºC for 4 minutes; and the final extension at 72 ºC for 16 minutes.
2.4.2. BOX-PCR
Genotyping was also carried out using the BOXA1R primer (CTACGGCAAGGCGACGCTGACG), as previously described by Ferrario et al. [22], with some modifications. The reaction was carried out using the GoTaq PCR Core System kit in a total reaction volume of 25 μL. The reaction mixture consisted of 100 ng of template DNA (2 μL) plus 23 μL of PCR mix (1X PCR buffer, 0.5 μM of primer, 0.2 mM dNTP, 3.0 mM MgCl2, 0.06 U µL-1 Taq DNA polymerase, and sterile water). PCR amplification was carried out in a Veriti 96-well thermal cycler, using the following conditions: 1 cycle at 95 ºC for 5 minutes for denaturation, followed by 40 cycles at 95 ºC for 1 minute, 48 ºC for 20 seconds and 72 ºC for 2 minutes; and the final extension was carried out at 72 ºC for 10 minutes.
2.4.3. RAPD-PCR
PCR amplification of genomic DNA using M13 arbitrary sequence primers (GAGGGTGGCGGTTCT) was performed as previously described by Ferrario et al. [22], with some modifications. The reaction was carried out using the GoTaq PCR Core System kit in a total reaction volume of 25 μL. The reaction mixture consisted of 100 ng of template DNA (2 μL) plus 23 μL of PCR mix (1X PCR buffer, 0.5 μM primer, 0.2 mM dNTP, 3.0 mM MgCl2, 0.06 U µL-1 Taq DNA polymerase, and sterile water). PCR amplification was carried out in a Veriti 96-well thermal cycler, using the following conditions: 1 cycle of 95 ºC for 5 minutes for denaturation, followed by 40 cycles of 95 ºC for 1 minute, 46 ºC for 20 seconds and 72 ºC for 2 minutes; and the final extension at 72 ºC for 10 minutes.
2.4.4. Analysis of Agarose Gels
The PCR products were separated via electrophoresis on a 1.5% agarose gel, stained with ethidium bromide (0.5 μg mL-1 for 30 minutes), then visualized in a UV transilluminator, and the images were captured using the photo-documentation system L-Pix EX (Loccus Biotechnology). A 1 kb marker (Invitrogen) was used as a molecular weight standard.
The REP-PCR, BOX-PCR and RAPD-PCR images were analyzed using BioNumerics software version 6.6 (Applied Maths). The Dice coefficient was used to assess the similarity between the identified band patterns [35]. The following parameters were used: optimization = 2%; band matching tolerance = 1%. Dendrograms were created using the unweighted pair group method with the arithmetic mean (UPGMA) approach [36]. To assess clonality between strains, a cutoff of ≥ 80% was proposed based on the DNA fingerprint pattern that considers this value to determine the genetic similarity between strains [37,38]. The discriminatory power of the REP-, BOX-, and RAPD-PCR techniques was determined using Simpson’s index of diversity (SDI) [39]. The adjusted Rand index (ARI) was used to measure the congruence among the different typing methods [40]. Wallace coefficients (WC) were calculated to estimate the probability that two isolates assigned together by one typing method are also classified in the same type using another method (bidirectional values) [41]. SDI, ARI and WC analyses were performed using the online software Comparing Partitions (available at http://www.comparingpartitions.info/) [42].
3. Results
3.1. Bacterial Identification
From the sequencing of the gyrB gene and the phylogenetic analysis, the new taxonomic identifications of the isolates previously identified as L. garvieae by MALDI-TOF MS were obtained. Thus, fourteen isolates were identified as L. garvieae, seventeen were identified as L. petauri and five as L. formosensis (Figure 2).
3.2. Serotyping of the Lactococcus spp. Strains
The presence of two serotypes for the Lactococcus spp. isolates was observed. Serotype I was related to the strains classified as L. garvieae and L. petauri, while serotype II was related to strains identified as L. formosensis (Figure 2).
3.3. Genotyping of the Lactococcus spp. Isolates
The REP-PCR resulted in the amplification of 12, 13 and 10 bands ranging in size from 400 to 6,000 bp, with similarities of 73.9%, 51.4% and 70.4% for L. formosensis, L. garvieae and L. petauri, respectively. Two, seven and four different REP patterns were detected for L. formosensis, L. garvieae and L. petauri, respectively (Figure 3).The BOX-PCR resulted in the amplification of 9, 14 and 7 bands ranging in size from 500 to 6,000 bp, with similarities of 56.2%, 52.6% and 57.2% for L. formosensis, L. garvieae and L. petauri, respectively. Two, six and three different BOX patterns were detected for L. formosensis, L. garvieae and L. petauri, respectively (Figure 4).The RAPD-PCR resulted in the amplification of 9, 7 and 13 bands ranging in size from 550 to 2,000 bp, with similarities of 37.6%, 29.3% and 45.8% for L. formosensis, L. garvieae and L. petauri, respectively. Four, six and seven different RAPD patterns were detected for L. formosensis, L. garvieae and L. petauri, respectively (Figure 5).
The SDI values are presented in Table 2. There was no equivalence of discriminatory powers among the genotyping methods for L. formosensis and L. petauri. However, for the L. garvieae strains, the discriminatory power of each method was high (SDI ≥ 0.824), suggesting an equivalent level of discrimination between them. A greater discriminatory power was observed in the RAPD-PCR assay (SDI ≥ 0.765). In relation to the bacterial species, L. petauri showed the lowest diversity values, demonstrating less intraspecific variation, regardless of the typing methods.
Furthermore, the ARI and WC were used to compare the congruency between type assignments of the different typing methods. These values are presented in Table 2. In general, there was a weak correlation between the information provided by the methods tested. Using ARI, a low level of agreement was observed between REP-, BOX-, and RAPD-PCR, regardless of the bacterial species evaluated. The maximum value of ARI of 0.429 was found at a threshold level of 80% similarity. The WC values between BOX- and REP-PCR for the L. formosensis strains, or when comparing these two typing methods with RAPD-PCR, evidenced a low bidirectional correspondence among the techniques (Table 2). On the other hand, WC for RAPD-PCR and REP- or BOX-PCR approach the estimated coverage (WCrapd→rep = 1.000 and WCrapd→box = 1.000). Therefore, RAPD-PCR was better predictor for the genotype of this bacterial species.
For the L. garvieae strains, in the WC values, irrespective of the typing methods compared, low bidirectional correspondence values were observed (Table 2). As such, the additional information provided by a second typing method is useful for assessing the diversity of this species.
In relation to the L. petauri isolates, the WC values also showed that RAPD-PCR was a better predictor of REP- (WCrapd→rep = 0.969) or BOX-PCR (WCrapd→box = 0.750) types than the contrary. In addition, BOX-PCR was also better predictor of the REP-PCR type (WCbox→rep = 0.838) than REP-PCR was of the BOX-PCR type; however, these methodologies had a strong bidirectional correspondence (Table 2).
4. Discussion
Piscine lactococcosis has been detected in a wide range of fish species [10], and has generated significant economic losses in the aquaculture industry [43] due to the ability of the pathogen to adapt, survive and spread under different environmental conditions [44,45]. Here, we present a set of Lactococcus spp. strains obtained from native fish species in Brazil. These isolates came from both clinical cases of fish diseases and those intended for the isolation of a potential probiotic candidate (intestinal samples). A high rate of isolates was obtained from Pseudoplatystoma (n = 16, 44.4%) and Arapaima gigas (n = 10, 27.7%) and Brycon amazonicus, Hoplias macrophtalmus and Phractocephalus hemioliopterus were identified as new hosts of this pathogen.
The diagnosis of piscine lactococcosis is a crucial step in improving the management of an outbreak. The identification methods most used by diagnostic laboratories are based on bacterial isolation, followed by phenotypic and biochemical characterization [25]. However, due to the high genetic heterogeneity of Lactococcus spp. isolates, mainly for the L. garvieae/petauri/formosensis group, definitive identification of isolates based solely on the aforementioned methodologies may be inaccurate, thus requiring the application of molecular techniques to confirm the taxonomic position of the isolates [5,46]. As such, the correct discrimination between Lactococcus spp. is vital. In the present study, the sequencing of the gyrB gene was used to confirm the identification of all the isolates selected. Our results showed that five strains were L. formosensis, 14 were L. garvieae and 17 were L. petauri, which permits the first description of L. petauri and L. formosensis as a pathogen in native Brazilian fish species.
Lactococcus spp. strains can be classified into two serotypes using molecular methods, since they differ in the size of the fragments obtained. Fragments measuring 285 bp correspond to serotype I, while fragments measuring 1,285 bp correspond to serotype II [15]. Using this methodology, serotype I was detected in diseased Anguilla anguilla, Oncorhynchus mykiss, Paralichthys olivaceus, Pseudocaranx dentex, Rachycentron canadum, Seriola quinqueradiata, Seriola dumerili, and Trachinotus blochii [15,16,25,47,48,49]. On the other hand, serotype II was detected in Pseudocaranx dentex, Seriola quinqueradiata, Seriola dumerili and Seriola lalandi [15,50]. Our study showed that L. garvieae and L. petauri strains were classified as serotype I, while the isolates of L. formosensis were characterized as serotype II (Figure 2). In addition, one isolate that was previously classified as serotype II [50] was recently described as belonging to L. formosensis [47]. Furthermore, we performed an in silico PCR, using java web tools [51], for all the complete genomes (I4/60, GenBank accession number: GCA_023822125.1; 122061, GCA_002355575.1; and MS200408A, GCA_030295725.1) and the reference genome (NBRC 109475, GCA_018403745.1) of L. formosensis strains, with taxonomy checked on the NCBI database, and also found an expected PCR product size of 1,285 bp. This leads us to suspect that all L. formosensis strains are serotype II and that the molecular serotyping can be a good tool for diagnosing this bacterial species; however, future studies with more L. formosensis strains need to be conducted to prove this hypothesis.
PCR-based typing is an effective approach and has been used to identify different genotypes of a specific pathogen, which is fundamental for understanding the spread of diseases and their evolution [7,52]. DNA fingerprinting techniques can be applied to access the genetic diversity of different microorganisms and the use of primers specific to certain regions of the genome provides amplification patterns that make it possible to differentiate genetic groups [53]. RAPD-PCR is a methodology that is based on the amplification of various fragments using small and arbitrary primers at low annealing temperatures. This enables the hybridization of many mismatched sequences in a bacterial genome and generates an amplicon coverage of less than 3 kb. On the other hand, REP- and BOX-PCR techniques use primers that hybridize the non-coding repeated sequences, with the banding pattern depending on the number and distribution of these repeating elements throughout the genome [54]. Although these methodologies have less discriminatory power than other genotyping methods, such as PFGE and MLST, they are relatively simple, cheap, easy to perform and provide fast results [55], and are, therefore, advantageous in studies of genetic diversity. Here, we evaluated the genetic diversity of Lactococcus spp. strains isolated from different native Brazilian fish species using REP-, BOX-, and RAPD-PCR.
Previous studies using genotyping methods have demonstrated genetic variability between L. garvieae strains from different fishes, terrestrial animals and other sources, as well as from different geographic origins. Using RAPD-, REP- and ERIC-PCR, with oligonucleotides different to those used in our study, some authors have observed a wide diversity of genetic groups for the isolates studied [7,23,25,46]. It is known that clustering is strictly dependent on the chosen oligonucleotides and the efficiency of the technique [56]. In this way, when we examined studies that used BOX1R [22], GTG5 [8,22] and M13 [22,23,56,57,58] primers, the same ones used in the current study, we also observed a high heterogeneity within L. garvieae strains; however, without taxonomically distinguishing the isolates within L. garvieae/petauri/formosensis group. This is due to the fact that the suggestion to reclassify the isolates is recent [12]. Therefore, our study is the first to evaluate the genetic profiles in Lactococcus spp. for each bacterial species after their correct taxonomic classification.
We demonstrated that the molecular typing methods tested are useful for evaluating the genetic diversity of L. garvieae, L. petauri and L. formosensis strains. For the L. garvieae isolates, all the typing methods showed a considerably similar discriminatory power (SDI ≥ 0.824). The high values of SDI suggest a considerable genetic divergence among the strains evaluated, which is corroborated by the identification of different profiles in each methodology. REP-, BOX- and RAPD-PCR tends to cluster isolates obtained from the same outbreak or sample collection (candidate probiotic isolation) with 100% similarity, for example, CRPB53 and CRBP54 or LG88-23 and LG89-23 strains, which in all the techniques were grouped together. Therefore, non-epidemiologically related L. garvieae strains are heterogeneous. Based on the ARI and WC values, REP-PCR seems to be the most appropriate L. garvieae typing strategy when compared to the other methods evaluated.
In relation to the Lactococcus formosensis isolates, the discriminatory power of the typing methods was not equivalent and the congruence between them was low. There was no clear relationship between the typing methods and geographical origin, fish species or tissue collected. Only the BOX-PCR separated isolates from clinical diseases and the intestinal samples (Figure 4). RAPD-PCR resulted in more diverse band profiles when compared to the other two techniques, demonstrating that the isolates are epidemiologically unrelated, including those from the same mortality outbreak (167/23-02 and 167/23-06), which clustered into different genetic groups (Figure 5). Therefore, we suggest that RAPD-PCR type could be considered a better predictor of genetic groups for this bacterial species.
The L. petauri strains are more homogeneous when compared to those of L. garvieae. This homogeneity is demonstrated in the dendrogram cluster analysis of all the genotyping methods, especially in REP-PCR (SDI = 0.331, 14 isolates clustered in the same group) and BOX-PCR (SDI = 0.412, 13 isolates clustered in the same group). The similar banding patterns of REP- and RAPD-PCR, with 100% similarity, grouped the isolates of the same fish species more satisfactorily, and suggests that these isolates are closely related. A low congruence between the typing methods was observed and RAPD-PCR has the greatest potential to be used in epidemiological studies of L. petauri strains based on WC values.
Although PCR-based DNA fingerprinting techniques are widely used for the molecular genotyping of Lactococcus spp. strains, providing the identification of genetic relatedness of different isolates and track the origin, distribution, and evolution of pathogen [48], the different typing methods, which are each based on different attributes of genetic variation, would hardly present complete agreement among them [24], and may have relative low intra-laboratory reproducibility [54]. In that regard, PFGE and MLST are a well-established methodologies for the study of the genetic diversity of L. garvieae [23,59] and L. petauri [5]. Consequently, future studies using these methods should be performed to provided new insights into the genetic and evolutionary characteristics of the L. garvieae/petauri/formosensis group using the isolates obtained from the native Brazilian fish species.
5. Conclusion
The current study demonstrated a comprehensive indication of the diversity found within the Lactococcus spp. strains isolated from the native Brazilian fish species. L. petauri and L. formosensis were identified among the isolates selected after the correct discrimination of bacterial species. Serotype II was detected only in L. formosensis. Using PCR-based DNA fingerprinting techniques, we observed that the L. garvieae strains had a greater genetic heterogeneity, while L. petauri behaved like a more homogeneous population. A low agreement among typing methods was observed, regardless of bacterial species evaluated. However, based on SDI and WC values, REP-PCR is the most suitable method for evaluating the genetic diversity of L. garvieae strains, while RAPD-PCR is a better predictor of L. formosensis and L. petauri diversity among the techniques evaluated.
Author Contributions
Angelo Carlo Chaparro Barbanti: Conceptualization, Investigation, Methodology, Formal analyses, Writing – original draft, Writing – review & editing; Angélica Emanuely Costa do Rosário: Conceptualization, Investigation, Methodology, Formal analyses, Writing – review & editing; Cynthia Rafaela Monteiro da Silva: Investigation, Methodology, Writing – review & editing; Victória Pontes Rocha: Methodology, Writing – review & editing; Henrique Lopes Costa: Investigation, Formal analyses, Visualization, Writing – review & editing; Júlia Miranda Trindade: Methodology, Writing – review & editing; Luiz Fagner Ferreira Nogueira: Investigation, Formal analyses, Visualization, Data curation, Supervision, Writing – review & editing; Júlio César Câmara Rosa: Methodology, Writing – review & editing; Maria José Tavares Ranzani-Paiva: Resources, Writing – review & editing; Fabiana Pilarski: Investigation, Resources, Writing – review & editing; Silvia Umeda Gallani: Investigation, Resources, Supervision, Writing – review & editing; Carlos Augusto Gomes Leal: Investigation, Resources, Writing – review & editing; Henrique César Pereira Figueiredo: Conceptualization, Methodology, Supervision, Resources, writing – review & editing; Guilherme Campos Tavares: Conceptualization, Investigation, Methodology, Funding acquisition, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing.
Acknowledgments
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES), through the PROCAD/Amazônia (grant number 88881.200614/2018-01), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, grant numbers APQ-01227-22 and APQ-04309-22), and Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM).
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Approval
This research did not involve the use of live animals.
References
- Altinok, I.; Ozturk, R.C.; Ture, M. NGS Analysis Revealed That Lactococcus garvieae Lg-Per Was Lactococcus petauri in Türkiye. J. Fish Dis. 2022, 45, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Abraham, T.; Yazdi, Z.; Littman, E.; Shahin, K.; Heckman, T.I.; Quijano Cardé, E.M.; Nguyen, D.T.; Hu, R.; Adkison, M.; Veek, T.; et al. Detection and Virulence of Lactococcus garvieae and L. petauri from Four Lakes in Southern California. J. Aquat. Anim. Health 2023, 35, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.J.; Klesius, P.H.; Shoemaker, C.A. First Isolation and Characterization of Lactococcus garvieae from Brazilian Nile Tilapia, Oreochromis niloticus (L.), and Pintado, Pseudoplathystoma corruscans (Spix & Agassiz). J. Fish Dis. 2009, 32, 943–951. [Google Scholar] [CrossRef]
- Fukushima, H.C.S.; Leal, C.A.G.; Cavalcante, R.B.; Figueiredo, H.C.P.; Arijo, S.; Moriñigo, M.A.; Ishikawa, M.; Borra, R.C.; Ranzani-Paiva, M.J.T. Lactococcus garvieae Outbreaks in Brazilian Farms Lactococcosis in Pseudoplatystoma Sp. – Development of an Autogenous Vaccine as a Control Strategy. J. Fish Dis. 2017, 40, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Egger, R.C.; Rosa, J.C.C.; Resende, L.F.L.; de Pádua, S.B.; de Oliveira Barbosa, F.; Zerbini, M.T.; Tavares, G.C.; Figueiredo, H.C.P. Emerging Fish Pathogens Lactococcus petauri and L. garvieae in Nile Tilapia (Oreochromis niloticus) Farmed in Brazil. Aquaculture 2023, 565, 739093. [Google Scholar] [CrossRef]
- Quintana, C.I.F.; Guidoli, M.G.; Domitrovic, H.A.; Blanco, T.K. Lactococosis En Pseudoplatystoma reticulatum. Rev. Vet. 2017, 28, 132–137. [Google Scholar] [CrossRef]
- Ortega, C.; Irgang, R.; Valladares-Carranza, B.; Collarte, C.; Avendaño-Herrera, R. First Identification and Characterization of Lactococcus garvieae Isolated from Rainbow Trout (Oncorhynchus mykiss) Cultured in Mexico. Animals 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Shahin, K.; Veek, T.; Heckman, T.I.; Littman, E.; Mukkatira, K.; Adkison, M.; Welch, T.J.; Imai, D.M.; Pastenkos, G.; Camus, A.; et al. Isolation and Characterization of Lactococcus garvieae from Rainbow Trout, Onchorhyncus mykiss, from California, USA. Transbound. Emerg. Dis. 2022, 69, 2326–2343. [Google Scholar] [CrossRef] [PubMed]
- Bwalya, P.; Hang’ombe, B.M.; Evensen, Ø.; Mutoloki, S. Lactococcus garvieae Isolated from Lake Kariba (Zambia) Has Low Invasive Potential in Nile Tilapia (Oreochromis niloticus). J. Fish Dis. 2021, 44, 721–727. [Google Scholar] [CrossRef]
- Meyburgh, C.M.; Bragg, R.R.; Boucher, C.E. Lactococcus garvieae: An Emerging Bacterial Pathogen of Fish. Dis. Aquat. Organ. 2017, 123, 67–79. [Google Scholar] [CrossRef]
- Salogni, C.; Bertasio, C.; Accini, A.; Gibelli, L.R.; Pigoli, C.; Susini, F.; Podavini, E.; Scali, F.; Varisco, G.; Alborali, G.L. The Characterisation of Lactococcus garvieae Isolated in an Outbreak of Septicaemic Disease in Farmed Sea Bass (Dicentrarchus labrax, Linnaues 1758) in Italy. Pathogens 2024, 13. [Google Scholar] [CrossRef]
- Vela, A.I.; del Mar Blanco, M.; Colussi, S.; Kotzamanidis, C.; Prearo, M.; Altinok, I.; Acutis, P.L.; Volpatti, D.; Alba, P.; Feltrin, F.; et al. The Association of Lactococcus petauri with Lactococcosis Is Older than Expected. Aquaculture 2024, 578, 740057. [Google Scholar] [CrossRef]
- Eyngor, M.; Zlotkin, A.; Ghittino, C.; Prearo, M.; Douet, D.-G.; Chilmonczyk, S.; Eldar, A. Clonality and Diversity of the Fish Pathogen Lactococcus garvieae in Mediterranean Countries. Appl. Environ. Microbiol. 2004, 70, 5132–5137. [Google Scholar] [CrossRef]
- Choi, H.J.; Hur, J.W.; Cho, J.B.; Park, K.H.; Jung, H.J.; Kang, Y.J. Introduction of Bacterial and Viral Pathogens from Imported Ornamental Finfish in South Korea. Fish. Aquat. Sci. 2019, 22, 5. [Google Scholar] [CrossRef]
- Ohbayashi, K.; Oinaka, D.; Hoai, T.D.; Yoshida, T.; Nishiki, I. PCR-Mediated Identification of the Newly Emerging Pathogen Lactococcus garvieae Serotype II from Seriola quinqueradiata and S. dumerili. Fish Pathol. 2017, 52, 46–49. [Google Scholar] [CrossRef]
- Rao, S.; Pham, T.H.; Poudyal, S.; Cheng, L.-W.; Nazareth, S.C.; Wang, P.-C.; Chen, S.-C. First Report on Genetic Characterization, Cell-Surface Properties and Pathogenicity of Lactococcus garvieae, Emerging Pathogen Isolated from Cage-Cultured Cobia (Rachycentron canadum). Transbound. Emerg. Dis. 2022, 69, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Shahi, N.; Mallik, S.K.; Sahoo, M.; Chandra, S.; Singh, A.K. First Report on Characterization and Pathogenicity Study of Emerging Lactococcus garvieae Infection in Farmed Rainbow Trout, Oncorhynchus mykiss (Walbaum), from India. Transbound. Emerg. Dis. 2018, 65, 1039–1048. [Google Scholar] [CrossRef]
- Eldar, A.; Goria, M.; Ghittino, C.; Zlotkin, A. Herve Bercovier Biodiversity of Lactococcus garvieae Strains Isolated from Fish in Europe, Asia, and Australia. Appl. Environ. Microbiol. 1999, 65, 1005–1008. [Google Scholar] [CrossRef]
- von Wright, A. Genus Lactococcus. In Lactic Acid Bacteria: Microbiological and Functional Aspects; Vinderola, G., Ouwehand, A., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, 2019; pp. 33–46. [Google Scholar]
- de Ruyter, T.; Littman, E.; Yazdi, Z.; Adkison, M.; Camus, A.; Yun, S.; Welch, T.J.; Keleher, W.R.; Soto, E. Comparative Evaluation of Booster Vaccine Efficacy by Intracoelomic Injection and Immersion with a Whole-Cell Killed Vaccine against Lactococcus petauri Infection in Rainbow Trout (Oncorhynchus mykiss). Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Church, D.L.; Cerutti, L.; Gürtler, A.; Griener, T.; Zelazny, A. Stefan Emler Performance and Application of 16S RRNA Gene Cycle Sequencing for Routine Identification of Bacteria in the Clinical Microbiology Laboratory. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef]
- Ferrario, C.; Ricci, G.; Borgo, F.; Rollando, A.; Fortina, M.G. Genetic Investigation within Lactococcus garvieae Revealed Two Genomic Lineages. FEMS Microbiol. Lett. 2012, 332, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Chen, M.-Y.; Sudpraseart, C.; Lin, P.; Yoshida, T.; Wang, P.-C.; Chen, S.-C. Genotyping and Phenotyping of Lactococcus garvieae Isolates from Fish by Pulse-Field Gel Electrophoresis (PFGE) and Electron Microscopy Indicate Geographical and Capsular Variations. J. Fish Dis. 2022, 45, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Kidd, T.J.; Grimwood, K.; Ramsay, K.A.; Rainey, P.B.; Bell, S.C. Comparison of Three Molecular Techniques for Typing Pseudomonas aeruginosa Isolates in Sputum Samples from Patients with Cystic Fibrosis. J. Clin. Microbiol. 2011, 49, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Torres-Corral, Y.; Santos, Y. Clonality of Lactococcus garvieae Isolated from Rainbow Trout Cultured in Spain: A Molecular, Immunological, and Proteomic Approach. Aquaculture 2021, 545, 737190. [Google Scholar] [CrossRef]
- Costa, F.A.A.; Leal, C.A.G.; Leite, R.C.; Figueiredo, H.C.P. Genotyping of Streptococcus dysgalactiae Strains Isolated from Nile Tilapia, Oreochromis niloticus (L.). J. Fish Dis. 2014, 37, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Sebastião, F.A.; Furlan, L.R.; Hashimoto, D.T.; Pilarski, F. Identification of Bacterial Fish Pathogens in Brazil by Direct Colony PCR and 16S RRNA Gene Sequencing. Adv. Microbiol. 2015, 05, 409–424. [Google Scholar] [CrossRef]
- Assis, G.B.N.; Pereira, F.L.; Zegarra, A.U.; Tavares, G.C.; Leal, C.A.; Figueiredo, H.C.P. Use of MALDI-TOF Mass Spectrometry for the Fast Identification of Gram-Positive Fish Pathogens. Front. Microbiol. 2017, 8, 1492. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.L.A.; Chagas, E.C.; Oliveira, P.M.; Benavides, M.V.; Majolo, C.; Boijink, C. de L.; Tavares-Dias, M.; Ishikawa, M.M.; Fujimoto, R.Y.; Rodrigues, F.B.; et al. Agentes Patogênicos de Tambaquis Cultivados, Com Destaque Para Registros Em Rio Preto Da Eva, AM 2016, 80.
- Rosário, A.E.C. do; Henrique, M.C.; Costa, É.J.C. da; Barbanti, A.C.C.; Tavares, G.C. Surtos de Bacteriose Em Juvenis de Pirarucu (Arapaima gigas) Provevientes de Pisciculturas Amazônicas e Avaliação Da Sensibilidade de Aeromonas jandaei a Antimicrobianos. In Enfermidades parasitárias e bacterianas na piscicultura brasileira: insights e perspectivas; Cruz, M.G. da, Castro, J. da S., Jerônimo, G.T., Eds.; i-EDUCAM, 2023; pp. 33–46.
- Cardoso, P.H.M.; Moreno, L.Z.; de Oliveira, C.H.; Gomes, V.T.M.; Silva, A.P.S.; Barbosa, M.R.F.; Sato, M.I.Z.; Balian, S.C.; Moreno, A.M. Main Bacterial Species Causing Clinical Disease in Ornamental Freshwater Fish in Brazil. Folia Microbiol. (Praha). 2021, 66, 231–239. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for Inferring Very Large Phylogenies by Using the Neighbor-Joining Method. Proc. Natl. Acad. Sci. 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Dice, L.R. Measures of the Amount of Ecologic Association between Species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Sokal, R.; Michener, C. A Statistical Method for Evaluating Systematic Relationship. Univ. Kansas Sci. Bull. 1958, 38, 1409–1438. [Google Scholar]
- Armitage, P.; Berry, G.; Matthews, J.N.S. Statistical Methods in Medical Research; 4th ed.; Blackwell Science Ltd: Bodmin, 2002; ISBN 9780470773666.
- Hunter, P.R. Reproducibility and Indices of Discriminatory Power of Microbial Typing Methods. J. Clin. Microbiol. 1990, 28, 1903–1905. [Google Scholar] [CrossRef] [PubMed]
- Hunter, P.R.; Gaston, M.A. Numerical Index of the Discriminatory Ability of Typing Systems: An Application of Simpson’s Index of Diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [PubMed]
- Hubert, L.; Arabie, P. Comparing Partitions. J. Classif. 1985, 2, 193–218. [Google Scholar] [CrossRef]
- Wallace, D.L. A Method for Comparing Two Hierarchical Clusterings: Comment. J. Am. Stat. Assoc. 1983, 78, 569–576. [Google Scholar] [CrossRef]
- Pinto, F.R.; Melo-Cristino, J.; Ramirez, M. A Confidence Interval for the Wallace Coefficient of Concordance and Its Application to Microbial Typing Methods. PLoS One 2008, 3, e3696. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.-A.; Wang, P.-C.; Yoshida, T.; Liaw, L.-L.; Chen, S.-C. Development of a Sensitive and Specific LAMP PCR Assay for Detection of Fish Pathogen Lactococcus garvieae. Dis. Aquat. Organ. 2013, 102, 225–235. [Google Scholar] [CrossRef]
- Gibello, A.; Galán-Sánchez, F.; Blanco, M.M.; Rodríguez-Iglesias, M.; Domínguez, L.; Fernández-Garayzábal, J.F. The Zoonotic Potential of Lactococcus garvieae: An Overview on Microbiology, Epidemiology, Virulence Factors and Relationship with Its Presence in Foods. Res. Vet. Sci. 2016, 109, 59–70. [Google Scholar] [CrossRef]
- Vendrell, D.; Balcázar, J.L.; Ruiz-Zarzuela, I.; de Blas, I.; Gironés, O.; Múzquiz, J.L. Lactococcus garvieae in Fish: A Review. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 177–198. [Google Scholar] [CrossRef] [PubMed]
- Ravelo, C.; Magariños, B.; López-Romalde, S.; Alicia, E.T.; Jesús, L.R. Molecular Fingerprinting of Fish-Pathogenic Lactococcus garvieae Strains by Random Amplified Polymorphic DNA Analysis. J. Clin. Microbiol. 2003, 41, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.M.; Abdelsalam, M.; Kawato, S.; Harakawa, S.; Kawakami, H.; Hirono, I.; Kondo, H. Comparative Genome Analyses of Three Serotypes of Lactococcus Bacteria Isolated from Diseased Cultured Striped Jack (Pseudocaranx dentex). J. Fish Dis. 2023, 46, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Neupane, S.; Rao, S.; Yan, W.-X.; Wang, P.-C.; Chen, S.-C. First Identification, Molecular Characterization, and Pathogenicity Assessment of Lactococcus garvieae Isolated from Cultured Pompano in Taiwan. J. Fish Dis. 2023, 46, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Salighehzadeh, R.; Sharifiyazdi, H.; Akhlaghi, M.; Soltanian, S. Serotypes, Virulence Genes and Polymorphism of Capsule Gene Cluster in Lactococcus garvieae Isolated from Diseased Rainbow Trout (Oncorhynchus mykiss) and Mugger Crocodile (Crocodylus palustris) in Iran. Iran. J. Vet. Res. 2020, 21, 26–32. [Google Scholar] [PubMed]
- Shi, Y.Z.; Nishiki, I.; Yanagi, S.; Yoshida, T. Epidemiological Study on Newly Emerging Lactococcus garvieae Serotype II Isolated from Marine Fish Species in Japan. Fish Pathol. 2019, 54, 51–57. [Google Scholar] [CrossRef]
- Kalendar, R.; Lee, D.; Schulman, A.H. Java Web Tools for PCR, in Silico PCR, and Oligonucleotide Assembly and Analysis. Genomics 2011, 98, 137–144. [Google Scholar] [CrossRef]
- Neoh, H.; Tan, X.-E.; Sapri, H.F.; Tan, T.L. Pulsed-Field Gel Electrophoresis (PFGE): A Review of the “Gold Standard” for Bacteria Typing and Current Alternatives. Infect. Genet. Evol. 2019, 74, 103935. [Google Scholar] [CrossRef] [PubMed]
- Borba, M.P.; Ballarini, A.E.; Witusk, J.P.D.; Lavin, P.; Van Der Sand, S. Evaluation of BOX-PCR and REP-PCR as Molecular Typing Tools for Antarctic Streptomyces. Curr. Microbiol. 2020, 77, 3573–3581. [Google Scholar] [CrossRef]
- Ramadan, A.A. Bacterial Typing Methods from Past to Present: A Comprehensive Overview. Gene Reports 2022, 29, 101675. [Google Scholar] [CrossRef]
- Stumpf, A.N.; Roggenkamp, A.; Hoffmann, H. Specificity of Enterobacterial Repetitive Intergenic Consensus and Repetitive Extragenic Palindromic Polymerase Chain Reaction for the Detection of Clonality within the Enterobacter cloacae Complex. Diagn. Microbiol. Infect. Dis. 2005, 53, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Foschino, R.; Nucera, D.; Volponi, G.; Picozzi, C.; Ortoffi, M.; Bottero, M.T. Comparison of Lactococcus garvieae Strains Isolated in Northern Italy from Dairy Products and Fishes through Molecular Typing. J. Appl. Microbiol. 2008, 105, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Duman, M.; Buyukekiz, A.G.; Saticioglu, I.B.; Cengiz, M.; Sahinturk, P.; Altun, S. Epidemiology, Genotypic Diversity, and Antimicrobial Resistance of Lactococcus garvieae in Farmed Rainbow Trout (Oncorhynchus mykiss) TT -. IFRO 2020, 19, 1–18. [Google Scholar]
- Fadaeifard, F.; Rabiei, M.; Sharifpour, M.F. Genetic Characterization of Lactococcus garvieae Isolated from Farmed Rainbow Trout by Random Amplified Polymorphic DNA-PCR in Iran. J. Hell. Vet. Med. Soc. 2020, 71, 2023–2030. [Google Scholar] [CrossRef]
- Ferrario, C.; Ricci, G.; Milani, C.; Lugli, G.A.; Ventura, M.; Eraclio, G.; Borgo, F.; Fortina, M.G. Lactococcus garvieae: Where Is It From? A First Approach to Explore the Evolutionary History of This Emerging Pathogen. PLoS One 2013, 8, e84796. [Google Scholar] [CrossRef]
- Tavares, G.C.; de Queiroz, G.A.; Assis, G.B.N.; Leibowitz, M.P.; Teixeira, J.P.; Figueiredo, H.C.P.; Leal, C.A.G. Disease Outbreaks in Farmed Amazon Catfish (Leiarius marmoratus x Pseudoplatystoma corruscans) Caused by Streptococcus agalactiae, S. iniae, and S. dysgalactiae. Aquaculture 2018, 495, 384–392. [Google Scholar] [CrossRef]
Figure 1.
Map of the distribution of Lactococcus spp. according to the geographic location of the farms. Colors represent bacterial species and sizes represent the number of isolates per location.
Figure 1.
Map of the distribution of Lactococcus spp. according to the geographic location of the farms. Colors represent bacterial species and sizes represent the number of isolates per location.
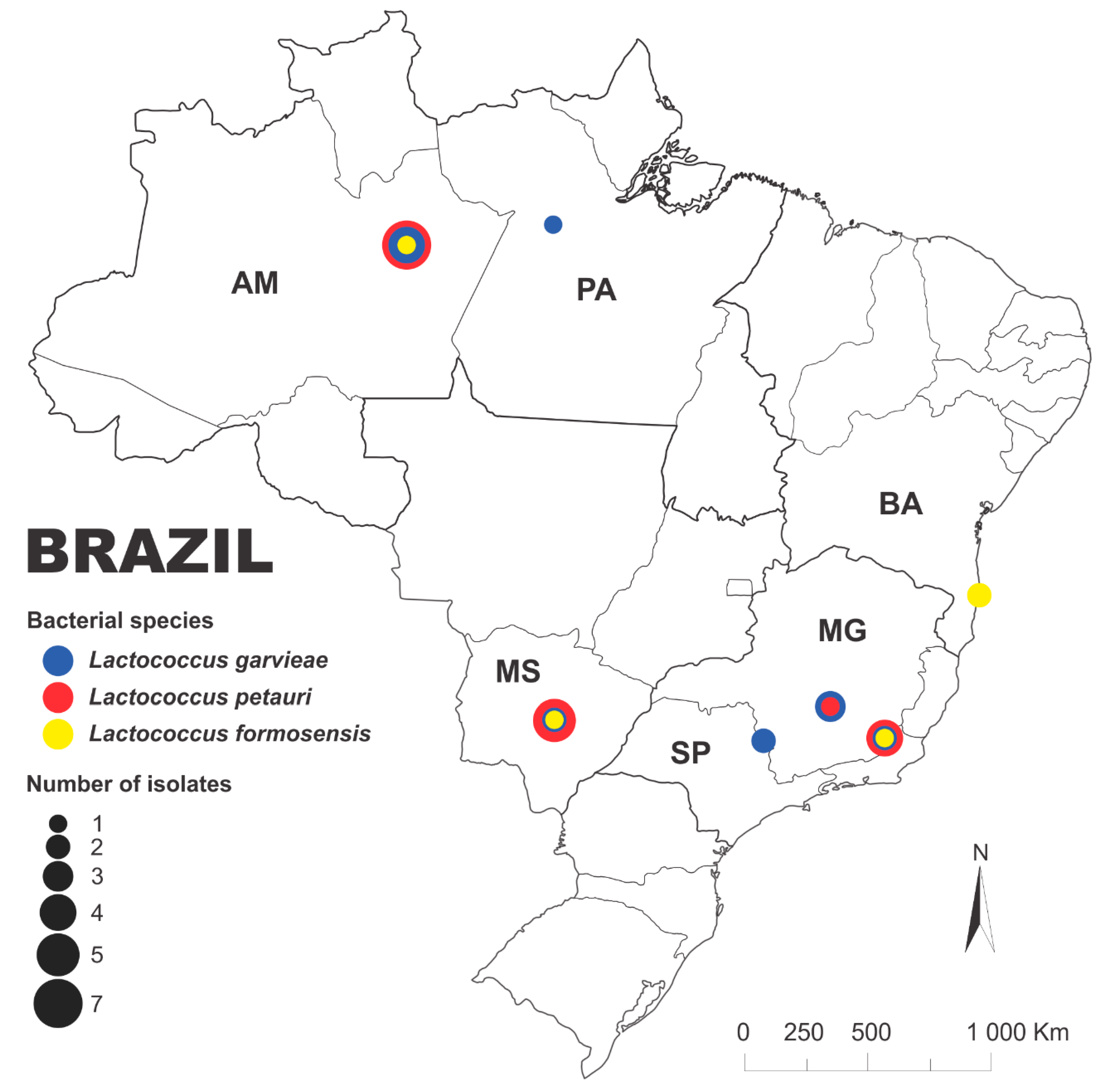
Figure 2.
Phylogenetic tree based on gyrB sequences of the isolates from this study and the representative strains of L. formosensis, L. garvieae and L. petauri obtained from NCBI were constructed using the neighbor-joining method. Bootstrap percentages (based on 1,000 replications) are shown at branch points. L. raffinolactis WiKim0068 was used as an outgroup. Molecular serotyping for the ATCC 49156 (GenBank accession number: GCA_000269925.1), Lg2 (GCA_000269945.1), B1726 (GCA_023499275.1), 159469 (GCA_002154895.1), I4/6O (GCA_023822125.1) and NBRC 109475 (GCA_018403745.1) strains was performed using java web tools for in silico PCR [51].
Figure 2.
Phylogenetic tree based on gyrB sequences of the isolates from this study and the representative strains of L. formosensis, L. garvieae and L. petauri obtained from NCBI were constructed using the neighbor-joining method. Bootstrap percentages (based on 1,000 replications) are shown at branch points. L. raffinolactis WiKim0068 was used as an outgroup. Molecular serotyping for the ATCC 49156 (GenBank accession number: GCA_000269925.1), Lg2 (GCA_000269945.1), B1726 (GCA_023499275.1), 159469 (GCA_002154895.1), I4/6O (GCA_023822125.1) and NBRC 109475 (GCA_018403745.1) strains was performed using java web tools for in silico PCR [51].
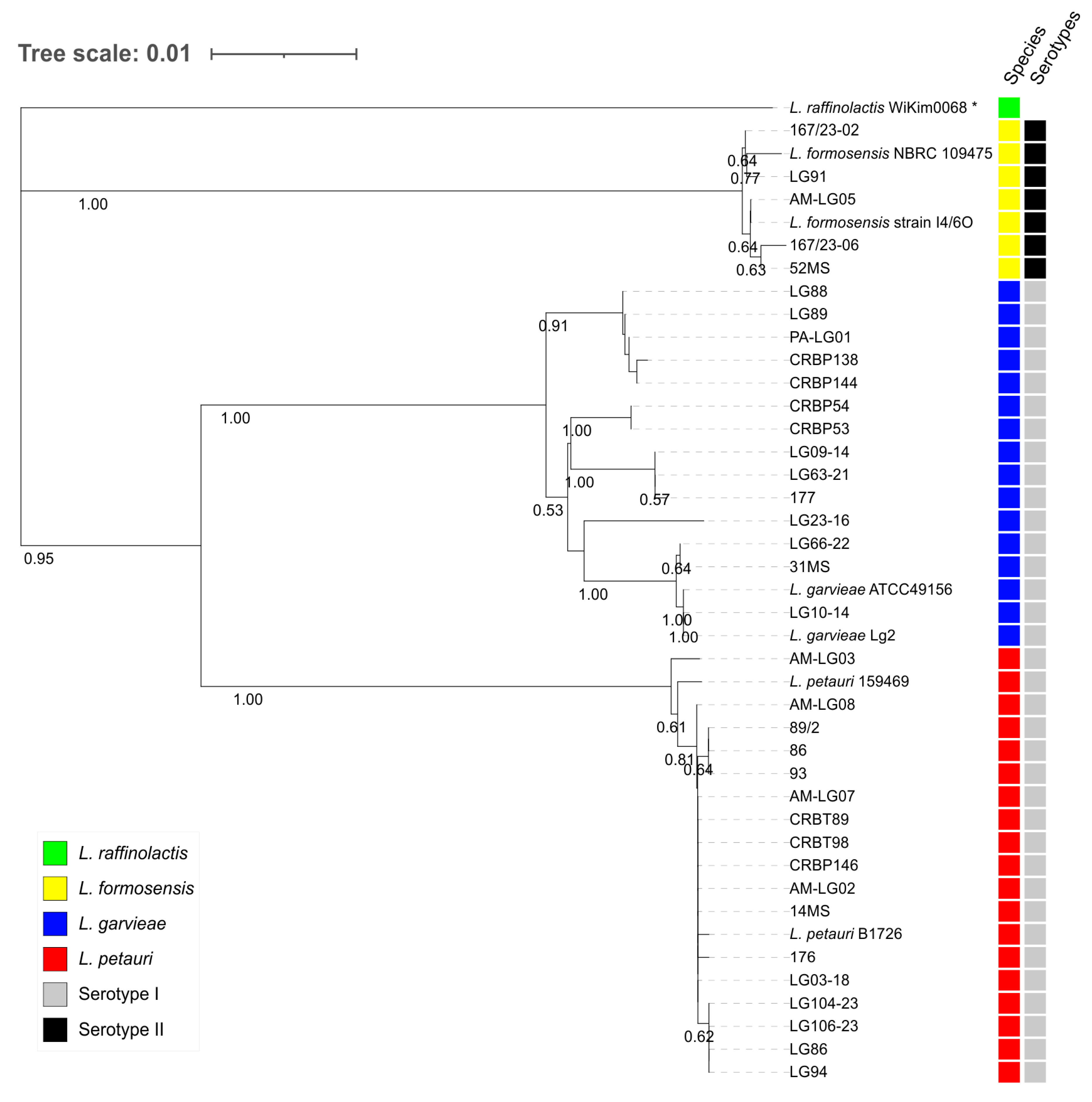
Figure 3.
Dendrograms obtained using REP-PCR for L. formosensis (A), L. garvieae (B) and L. petauri (C). The dendrogram was constructed using Dice’s coefficient and the UPGMA approach. Colors represent the host from which the bacteria were isolated (color code: forest green = Pseudoplatystoma sp., gray = Colossoma macropomum, red = Arapaima gigas, lime = Pseudoplatystoma fasciatum, green = Pseudoplatystoma corruscans, purple = Lophiosilurus alexandri, yellow = Brycon amazonicus, blue = Hoplias macrophtalmus, and pink = Phractocephalus hemioliopterus).
Figure 3.
Dendrograms obtained using REP-PCR for L. formosensis (A), L. garvieae (B) and L. petauri (C). The dendrogram was constructed using Dice’s coefficient and the UPGMA approach. Colors represent the host from which the bacteria were isolated (color code: forest green = Pseudoplatystoma sp., gray = Colossoma macropomum, red = Arapaima gigas, lime = Pseudoplatystoma fasciatum, green = Pseudoplatystoma corruscans, purple = Lophiosilurus alexandri, yellow = Brycon amazonicus, blue = Hoplias macrophtalmus, and pink = Phractocephalus hemioliopterus).

Figure 4.
Dendrograms obtained using BOX-PCR for L. formosensis (A), L. garvieae (B) and L. petauri (C). The dendrogram was constructed using Dice’s coefficient and the UPGMA approach. Colors represent the host from which the bacteria were isolated (color code: forest green = Pseudoplatystoma sp., gray = Colossoma macropomum, red = Arapaima gigas, lime = Pseudoplatystoma fasciatum, green = Pseudoplatystoma corruscans, purple = Lophiosilurus alexandri, yellow = Brycon amazonicus, blue = Hoplias macrophtalmus, and pink = Phractocephalus hemioliopterus).
Figure 4.
Dendrograms obtained using BOX-PCR for L. formosensis (A), L. garvieae (B) and L. petauri (C). The dendrogram was constructed using Dice’s coefficient and the UPGMA approach. Colors represent the host from which the bacteria were isolated (color code: forest green = Pseudoplatystoma sp., gray = Colossoma macropomum, red = Arapaima gigas, lime = Pseudoplatystoma fasciatum, green = Pseudoplatystoma corruscans, purple = Lophiosilurus alexandri, yellow = Brycon amazonicus, blue = Hoplias macrophtalmus, and pink = Phractocephalus hemioliopterus).

Figure 5.
Dendrograms obtained using RAPD-PCR for L. formosensis (A), L. garvieae (B) and L. petauri (C). The dendrogram was constructed using Dice’s coefficient and the UPGMA approach. Colors represent the host from which the bacteria were isolated (color code: forest green = Pseudoplatystoma sp., gray = Colossoma macropomum, red = Arapaima gigas, lime = Pseudoplatystoma fasciatum, green = Pseudoplatystoma corruscans, purple = Lophiosilurus alexandri, yellow = Brycon amazonicus, blue = Hoplias macrophtalmus, and pink = Phractocephalus hemioliopterus).
Figure 5.
Dendrograms obtained using RAPD-PCR for L. formosensis (A), L. garvieae (B) and L. petauri (C). The dendrogram was constructed using Dice’s coefficient and the UPGMA approach. Colors represent the host from which the bacteria were isolated (color code: forest green = Pseudoplatystoma sp., gray = Colossoma macropomum, red = Arapaima gigas, lime = Pseudoplatystoma fasciatum, green = Pseudoplatystoma corruscans, purple = Lophiosilurus alexandri, yellow = Brycon amazonicus, blue = Hoplias macrophtalmus, and pink = Phractocephalus hemioliopterus).


Table 1.
Geographical origin of the 36 strains of Lactococcus spp. isolated from the native Brazilian fish species.
Table 1.
Geographical origin of the 36 strains of Lactococcus spp. isolated from the native Brazilian fish species.
| Isolate | Species | Host | Tissue | Year | Brazilian state | Reference |
|---|---|---|---|---|---|---|
| 14MS | L. petauri | Pseudoplatystoma fasciatum | Kidney | 2012 | MS | [27] |
| 167/23-02 | L. formosensis | Arapaima gigas | Brain | 2023 | BA | This study |
| 167/23-06 | L. formosensis | Arapaima gigas | Brain | 2023 | BA | This study |
| 176 | L. petauri | Pseudoplatystoma fasciatum | Brain | 2012 | MS | [4] |
| 177 | L. garvieae | Pseudoplatystoma fasciatum | Brain | 2012 | MS | [4] |
| 31MS | L. garvieae | Pseudoplatystoma fasciatum | Kidney | 2012 | MS | [27] |
| 52MS | L. formosensis | Pseudoplatystoma fasciatum | Brain | 2012 | MS | [27] |
| 86 | L. petauri | Pseudoplatystoma sp. | Brain | 2012 | MS | [4] |
| 89/2 | L. petauri | Pseudoplatystoma sp. | Brain | 2012 | MS | [4] |
| 93 | L. petauri | Pseudoplatystoma sp. | Brain | 2012 | MS | [4] |
| AM-LG02 | L. petauri | Colossoma macropomum | Intestine | 2020 | AM | This study |
| AM-LG03 | L. petauri | Colossoma macropomum | Intestine | 2022 | AM | This study |
| AM-LG05 | L. formosensis | Colossoma macropomum | Intestine | 2022 | AM | This study |
| AM-LG07 | L. petauri | Brycon amazonicus | Brain | 2022 | AM | This study |
| AM-LG08 | L. petauri | Brycon amazonicus | Brain | 2022 | AM | This study |
| CRBP138 | L. garvieae | Arapaima gigas | Intestine | 2023 | AM | This study |
| CRBP144 | L. garvieae | Arapaima gigas | Intestine | 2023 | AM | This study |
| CRBP146 | L. petauri | Arapaima gigas | Intestine | 2023 | AM | This study |
| CRBP53 | L. garvieae | Arapaima gigas | Intestine | 2023 | AM | This study |
| CRBP54 | L. garvieae | Arapaima gigas | Intestine | 2023 | AM | This study |
| CRBT89 | L. petauri | Arapaima gigas | Intestine | 2023 | AM | This study |
| CRBT98 | L. petauri | Arapaima gigas | Intestine | 2023 | AM | This study |
| LG03-18 | L. petauri | Pseudoplatystoma corruscans | Brain | 2018 | MG | This study |
| LG09-14 | L. garvieae | Pseudoplatystoma corruscans | Kidney | 2014 | SP | [28] |
| LG10-14 | L. garvieae | Lophiosilurus alexandri | Brain | 2014 | MG | [28] |
| LG104-23 | L. petauri | Pseudoplatystoma sp. | Brain | 2023 | MG | This study |
| LG106-23 | L. petauri | Pseudoplatystoma sp. | Kidney | 2023 | MG | This study |
| LG23-16 | L. garvieae | Pseudoplatystoma corruscans | Brain | 2016 | SP | [60] |
| LG63-21 | L. garvieae | Hoplias macrophtalmus | Kidney | 2021 | MG | This study |
| LG66-22 | L. garvieae | Phractocephalus hemioliopterus | Kidney | 2022 | MG | This study |
| LG86-23 | L. petauri | Pseudoplatystoma sp. | Kidney | 2023 | MG | This study |
| LG88-23 | L. garvieae | Brycon amazonicus | Brain | 2023 | MG | This study |
| LG89-23 | L. garvieae | Brycon amazonicus | Kidney | 2023 | MG | This study |
| LG91-23 | L. formosensis | Pseudoplatystoma sp. | Brain | 2023 | MG | This study |
| LG94-23 | L. petauri | Pseudoplatystoma sp. | Brain | 2023 | MG | This study |
| PA-LG01 | L. garvieae | Arapaima gigas | Brain | 2018 | PA | [30] |
AM: Amazonas; BA: Bahia; MS: Mato Grosso do Sul; MG: Minas Gerais; PA: Pará; SP: São Paulo.
Table 2.
Number of types, Simpson’s index of diversity (SDI), adjusted Rand indices (ARI), and Wallace coefficients (WC) for REP-PCR, BOX-PCR and RAPD-PCR of L. formosensis (LF), L. garvieae (LG) and L. petauri (LP) strains.
Table 2.
Number of types, Simpson’s index of diversity (SDI), adjusted Rand indices (ARI), and Wallace coefficients (WC) for REP-PCR, BOX-PCR and RAPD-PCR of L. formosensis (LF), L. garvieae (LG) and L. petauri (LP) strains.
| Bacterial species | Evaluation Method | Number of types | SDI | ARI | WC | ||||
|---|---|---|---|---|---|---|---|---|---|
| REP | BOX | RAPD | REP | BOX | RAPD | ||||
| LF | REP | 2 | 0.600 | 1.000 | 0.000 | 0.286 | 1.000 | 0.500 | 0.250 |
| BOX | 2 | 0.400 | 1.000 | 0.138 | 0.333 | 1.000 | 0.167 | ||
| RAPD | 4 | 0.900 | 1.000 | 1.000 | 1.000 | 1.000 | |||
| LG | REP | 7 | 0.901 | 1.000 | 0.429 | 0.221 | 1.000 | 0.667 | 0.444 |
| BOX | 6 | 0.835 | 1.000 | 0.261 | 0.400 | 1.000 | 0.400 | ||
| RAPD | 6 | 0.824 | 1.000 | 0.250 | 0.375 | 1.000 | |||
| LP | REP | 4 | 0.331 | 1.000 | 0.421 | 0.239 | 1.000 | 0.736 | 0.341 |
| BOX | 3 | 0.412 | 1.000 | 0.139 | 0.838 | 1.000 | 0.300 | ||
| RAPD | 7 | 0.765 | 1.000 | 0.969 | 0.750 | 1.000 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



