
Submitted:
13 September 2024
Posted:
14 September 2024
You are already at the latest version
Abstract
Cotton (Gossypium spp.) is the most important fibre crop worldwide. Black root rot and Fusarium wilt are two major diseases of cotton caused by soil-borne Berkeleyomyces rouxiae and Fusarium oxysporum f. sp. vasinfectum (Fov), respectively. Phenotyping plant symptoms caused by soil-borne pathogens has always been a challenge. To increase the uniformity of infection, we adapted a seedling screening method that directly uses liquid cultures to inoculate the plant roots and the soil. Four isolates each of B. rouxiae and Fov were collected from cotton fields in Australia and were characterised for virulence on cotton under controlled plant growth conditions. While the identity of all four B. rouxiae isolates were confirmed by multi-loci sequencing, only two of them were found to be pathogenic on cotton, suggesting variability in the ability of isolates of this species to cause disease. The four Fov isolates were phylogenetically clustered together with the other Australian Fov isolates and displayed both external and internal symptoms characteristic of Fusarium wilt on cotton plants. Furthermore, the isolates appeared to induce varied levels of plant disease severity indicating differences in their virulence on cotton. To contrast the virulence of the Fov isolates, four putatively non-pathogenic Fusarium oxysporum (Fo) isolates collected from cotton seedlings exhibiting atypical wilt symptoms were assessed for their ability to colonise cotton host. Despite the absence of Secreted In Xylem genes (SIX6, SIX11, SIX13 and SIX14) characteristic of Fov, all four Fo isolates retained the ability to colonise cotton and induce wilt symptoms. This suggests that slightly virulent strains of Fo may contribute to the overall occurrence of Fusarium wilt in cotton fields. Findings from this study will allow better distinction to be made between plant pathogens and endophytes and allow fungal effectors underpinning pathogenicity to be explored.
Keywords:
Upland cotton
; pathogenicity
; fungal effectors
; Fusarium wilt
; black root rot
; Secreted In Xylem effector genes
; endophyte
; Translation elongation factor-1 alpha subunit
1. Introduction
Cotton (Gossypium hirsutum) is the most important fibre crop in the world. Approximately 25 million tons of cotton are produced worldwide per annum with an estimated value of 12 billion USD [1]. In Australia, the cotton industry is primarily based in the inland regions of New South Wales and southern Queensland generating an annual revenue of 1.9 billion AUD [2].
Two important diseases of cotton in Australia are black root rot (BRR) and Fusarium wilt caused by Berkeleyomyces rouxiae and Fusarium oxysporum f. sp. vasinfectum (Fov) respectively [2,3]. Both fungal pathogens are considered hemibiotrophic plant pathogens. B. rouxiae was formerly known as Thielaviopsis basicola within the family Ceratocystidaceae [4]. B. rouxiae first colonises living root tissues and then consumes the root cells causing black discolouration, which results in brittle root tissues and reduced water and nutrient uptake by the plants [5]. Severe black root rot infection typically results in stunted seedlings which can delay flowering of the plants [6] but it does not often cause plant death in the cotton fields [3]. Yield losses of up to 10% caused by BRR have been reported in Australia [3].
Fov is a soil-borne pathogen that colonises the vascular system of the plants. Its proliferation inside the water-conducting vessels causing blockage of water supply to the upper parts of the plant, leads to leaf wilting, chlorosis, necrosis, vascular discolouration, plant stunting, defoliation and plant death [7]. Globally there are multiple races of Fov, but isolates originating from Australia belong to their own distinct lineage [8,9]. Vegetative compatibility groupings (VCGs) are also used to classify Fov isolates. Initially, in a study by Fernandez et al. (1994), ten VCGs were identified in Fov pathogenic to cotton, these being 0111-01110, with the first 3 digits representing Fov and the last number (1-10) identifying the VCG subgroup. They found that each of the races 1, 2, 3, 4, and 6 corresponded to a distinct VCG [10]. The Australian Fov isolates were then identified with distinct VCGs, 01111 and 01112 [11]. Studies showed that most isolates within the same VCG had the same rDNA and mtDNA haplotypes although multiple haplotypes were observed in some cases. Overall, three ancestral lineages of F. oxysporum pathogenic towards cotton were observed which includes: race 3 which is common in Egypt, Sudan, and Israel and considered a single clone; the Australian biotypes; and the all others including Californian races and race 1 from the Americas, races 7 and 8 from China, and race 4 from India [7].
In recent years, many isolates of F. oxysporum from cotton seedlings exhibited atypical that wilt symptoms and that do not phylogenetically conform to the standard Australian biotypes have been identified [12]. This highlights a need to investigate further the potential of F. oxysporum to cause disease on cotton plants. There are currently a number of undetermined VCGs within Fov [13] as well as the presence of Australian isolates that are not compatible with VCG01111 and VCG0112 but never-the-less showed the ability to infect cotton plants [2,12]. This study aims to address whether minor or mildly virulent races of Fov may be contributing to the overall pathogenicity of the F. oxysporum populations in cotton growing regions of Australia.
Pathogenicity of black root rot pathogen relied on using naturally or artificially infested soil or commercial potting mix, respectively [14]. On the other hand, there is an array of methods such as root dip [15], soil drench [16], stem puncture, and agar plugs amended potting mix [17] to assess pathogenicity of cotton-Fusarium spp. Different inoculation methods also resulted in different implications. Therefore, in this study we sought to assess virulence of both black root rot pathogen and cotton-Fusarium spp. using a rapid and robust seedling and soil inoculation assay using a spore suspension obtained from unfiltered liquid cultures. Assessing pathogenicity of these important cotton pathogens is critical in understanding not just virulence but also detecting new biotypes that may contribute to the evolution of pathogen populations across the wide cotton growing regions.
2. Materials and Methods
2.1. Fungal Isolates
A total of 12 fungal isolates were used in this study. Four confirmed Fusarium oxysporum f. sp. vasinfectum isolates, namely Fov SG1, Fov SG26, Fov SG55 and Fov TH1 that were recovered from diseased stems with vascular browning, a typical characteristic of Fusarium wilt, and were collected during a cotton disease survey in regional Queensland (Table 1). Fusarium oxysporum isolates Fo BRF1, Fo BRF2, Fo SHF6 and Fo WRF2 were collected from hypocotyls of cotton seedlings in regional New South Wales (NSW). These seedlings displayed collar rot in the form of red-brown lesions around the seedling collars [12] (Table 1). The Berkeleyomyces rouxiae isolates were collected from the roots (StrB22, BRR4, RVB4.1) and the crown vascular tissue (22BRR77) of cotton plants showing black root rot symptoms in regional NSW (Table 1). Using the single spore isolation method [3], monoconidial cultures for all 12 isolates were obtained from spores grown on cultures of half-strength potato dextrose agar (PDA) (F. oxysporum) or 10% carrot extract agar (carrot juice extracted from 100 g of carrots per litre of water) carrot extract agar (B. berkeleyomyces).
Diameter of the colonies was recorded and then used to normalise conidia production per unit area of the colony. Slides for microscopic examination were prepared using a tape touch method [18]. Fungal structures were examined under a Leica DM1000 compound microscope with ICC50 W Camera using the 40× objective (Leica Camera, Wetzlar, Germany). F. oxysporum isolates were stained with lactophenol cotton blue before visualisation under the microscope.
To prepare a spore suspension, culture plates were flooded with 10mL of sterile water. Mycelia and conidia were gently scraped off the plates using a sterile disposable spreader. The suspension was collected, and the conidia concentration was determined using a hemocytometer and an appropriate dilution factor.
2.2. DNA Extraction, PCR and Phylogenetic Analysis
DNA extraction was performed using mycelial scraped off a plate and then subjected to the microwave oven method [12]. PCR was performed using a 1 in 10 diluted DNA template and Dreamtaq (Thermo Fisher Scientific, Waltham, MA, USA). PCR amplicons of the correct size were confirmed on a 1% agarose gel, purified using GeneJET PCR purification kit (Thermo Fisher Scientific, Waltham, MA, USA), and Sanger-sequenced (Australian Genome Research Facility, Melbourne, Australia).
Universal primers for Secreted In Xylem (SIX) genes 1 to 14 that specifically detects the presence or the absence of these effector genes in Fusarium oxysporum species complex were obtained from a previous study [19] (Table S1). Fov-specific primers for detecting SIX6 in the Australian Fov biotypes were obtained from a previous study [20] (Table S1). PCR was performed to retrieve gene fragments of the translation elongation factor 1-alpha (TEF-1α), mitochondrial small subunit rDNA (mtSSU rDNA), nitrate reductase (NIR), and phosphate permease (PHO) using previously specified primers and conditions [9,21,22,23]. Minichromosome Maintenance Complex Component 7 (MCM7), the RNA polymerase II gene encoding the second largest protein subunit (RPB2) was PCR-amplified using primers derived from previous studies [24,25] and are listed along with their respective PCR conditions and PCR amplicon sizes in Table S1. The collection of sequences forming the backbone of the phylogenetic analysis were retrieved from the respective studies for F. oxysproum [2,9,21] and B. rouxiae [4].
Geneious Prime v 2024.0.7 (Biomatter Pty. Ltd., Auckland, New Zealand) was used to perform the phylogenetic analysis. DNA sequences were aligned using MAFFT v 7.490 [26] and edited over two iterations to remove gaps and derive a consensus alignment sequence. Bayesian inference was used to construct the phylogenetic trees using MrBayes v 3.2.6 [27]. The settings included the GTR-G-I model of substitution and two independent analyses on four Markov chain Monte Carlo (MCMC) chains for 2,000,000 generations, with a burn-in rate of 25% for every 1000 generations sampled. Orthologous gene sequences from other Fusarium spp. were used to anchor the whole phylogeny of the same gene. The tree branches were transformed into a cladogram and visualised in Geneious Prime.
2.3. Growth Chamber Experiments
Seeds of the cotton cv. Sicot746 B3F were directly sown into 105 mm (width) x 105 mm (length) x 120 mm (depth) square pots. The potting mix (UQ23) contained 70% composted pine bark of 0-5 mm in size and 30% Coco Peat and has a pH range of 5.5-6.5. UQ23 potting mix was then pasteurised using steam. For potting, each pot had three seeds which produced one to three seedlings per pot. Pots were lightly fertilised with a teaspoon of a balanced fertiliser (Osmocote). For each treatment, six pots containing seeds were put in a rectangular plastic tray with dimensions of 35 cm (length) x 29 cm (Width) x 5.5 cm (depth). The trays had a layer of soil approximately 2-3 cm deep underneath the pots.
A double-tier Conviron growth chamber of model GEN1000 SH (Conviron, Winnipeg, Canada) was used for plant growth. Plants for both Fov and B. rouxiae pathogenicity testing were grown concurrently in the same chamber. A single tier can hold 5 trays of plants, and each tray represents a single treatment. The top tier contained the four Fov treatments and the bottom tier contained the four B. rouxiae treatments (Figure S1, S2). Each tier also had their own uninoculated controls (sterile water). The growth chamber conditions were set to 16 h of photoperiod with a day/night temperature of 26°C / 22°C, respectively. Relative humidity was set to default at 60%. Plants were lightly watered every two to three days. The F. oxysporum experiment was set up later independent of the first experiment, but in the same chamber with the same running conditions.
2.4. Plant Inoculation
For establishing the liquid cultures, four to five mycelial plugs were used to inoculate sterile liquid media. Half-strength potato dextrose broth (PBD, 500mL) was used to culture each of the eight F. oxysporum isolates whereas 10% carrot juice (200mL) was used to culture the B. rouxiae isolates. Cultures were continuously shaken on a rotating platform at a speed of 120 rpm and a temperature of 26°C for 7 days. The spore concentration of each culture was determined using a hemacytometer on the day of inoculation (Table S2). The Fov and Fo cultures produced spores in the range of 2.7×105 to 3.8×107 conidia per mL, whereas the B. rouxiae culture produced spores in the range of 1.8×105 to 1.4×106 condia per mL for all but StrB22, which produced a low number of spores but abundant mycelia (Table S2).
Unfiltered culture (500mL) with the above-mentioned concentrations was used for root dipping. Firstly, 2-week-old cotton seedlings were uprooted from the pots (Figures S1, S2). Their roots were cleaned by rinsing with distilled water and then blotted dry on paper towels. Plants were then root-dipped in the unfiltered culture for 5 minutes (Figures S1D, S2D). Soil from the pots of each treatment was tipped into a disposable bag and mixed evenly with 250mL of the unfiltered culture before seedlings were re-potted. After inoculation, each tray was then bagged with a disposable (700 x 1000mm) waste bag (Richlands, Australia) to avoid cross-contamination (Figures S1B, S2B).
2.5. Symptom Assessment and Pathogen Reisolation from Plant Tissues
Plants inoculated with B. rouxiae were examined at 15 days post inoculation (dpi). Plants inoculated with Fov were examined at 27 dpi. Due to the observation that some of these Fov inoculated plants were hitting the ceiling of the growth chamber at the time of harvest, in the subsequent growth chamber trial with F. oxysporum, plants were examined at an earlier stage, at 17 dpi.
Plants were examined for external discolouration on the roots and stems. Plant weight was determined using both the above and below-ground portions. Plant height was measured from the basal node to the terminal bud of the main stem. Root discolouration was measured as the length of discoloured roots and was expressed as a percentage of the total length of the roots. External stem discolouration was scored at a 5-10 cm region above the basal node of the main stem. Internal discolouration was assessed by longitudinally cutting the stems. Leaf wilting was scored as the number of leaves that were either dropped, wilting or showing yellowing symptoms over the number of leaf nodes along the main stem, with or without leaves attached.
Koch's postulate was performed to assess the extent of the colonisation by the pathogen in the host using a previously described method [28]. The plant tissues were surface sterilised in 70% ethanol for 60 s and were blotted dry under aseptic conditions. Roots (main root node and fine roots), stem (basal node of the stem and 7-10 cm above ground), and petiole (leaf nodes joining stem) tissues of approximately 5 mm x 2 mm pieces were embedded into half-strength PDA (Fov and F. oxysporum) or 10% carrot agar (B. rouxiae) containing 100 ppm streptomycin sulfate. Four tissues per tissue type per plant were plated. Plates were incubated in the dark at 26°C for 4 days and then scored for the presence of F. oxysporum and B. rouxiae colonies under a microscope. A positive was determined firstly by the presence of microconidia and mycelia (F. oxysporum) or endoconidia chains and aleuriospores (B. rouxiae). A visual identification was also made by comparing the morphology and colour of the colonies with those of the pure culture plates.
2.6. Statistical Analysis
Statistical analysis was performed in SPSS statistics for Macintosh v29 (IBM Corp, Armonk, NY, USA). Shapiro–Wilk normality test was first performed to assess whether the dataset fits a normal distribution. One-way analysis of variance (ANOVA) was then performed to derive the descriptive statistics including the means and the 95% confidence interval of each treatment group. Homogeneity of variance was also assessed using Levene’s statistics. A post-hoc Tukey's Honestly Significant Difference test was performed using sample size harmonic means for unequal sample sizes to separate means for groups in homogeneous subsets.
3. Results
3.1. Morphology of the Isolates
On carrot agar, the B. rouxiae isolates were olive green initially and darkened over time (Figure 1A). The isolates had uniform colony margins. RVB4.1, 22BRR77 and BRR4 were flat in elevation. StrB22 appeared white in the centre, with raised elevation from aerial hyphae, and circular. BRR4 and StrB22 produced few spores in liquid media containing 10-20% (w/v) carrot juice (Table S2). All four B. rouxiae isolates grew well on half-strength PDA. On both half-strength PDA and 10% carrot agar, all four isolates showed the presence of endoconidia and aleuriospores under a microscope (Figure 2A-H). Endoconidia were unicellular, mostly hyaline, sometimes brown, cylindrical and produced in single or chains. No endoconidia chains were observed in StrB22. Endoconidial size varied from 10.2–16.0 μm in length and 4.7–5.4 μm in width (Figure S3A). Aleuriospores were dark brown, formed laterally or terminally on hyphal branches in chains of 2–6 cylindrical segments (Figure 2A-H). Size of a single aleuriospore varied from 32.7–40.6 μm in length and 11.4–12.3 μm in width (Figure S3B). Conidia production on half-strength PDA ranged from 16.7–26,800 endoconidia per mm2 colony (Figure S3C).
The Fov colonies appeared pale to light pink on half-strength PDA plates (Figure 1B) whereas the F. oxysporum colonies appeared to vary in colour which includes white (Fo BRF1), pale yellow (Fo BRF2), orange (Fo WRF2) and dark pink (Fo SHF6) (Figure 1C). For Fov SG1, Fov SG26, Fov SG55 and Fo SH6 white aerial hyphae were observed whereas the mycelia of Fov TH1, Fo BRF1, Fo BRF2 and Fo WRF2 were flat and closely adhered to the media. Microconidia were on average 6.0–12.4 μm in length and 2.4–2.7 μm in width (Figure S4A), and hyaline, single-celled, oval to reniform, and produced abundantly in false heads on short monophialides (Figure 1I-P). Macroconidia were on average 20–30 μm long, generally three-septate, hyaline, sometimes slightly curved, and had an apical hook (Figure 2I-P). Conidia production on half-strength PDA ranged from 300–65,460 endoconidia per unit area of the colony (Figure S4B).
3.2. Phylogenetic Analysis Confirms the Australian Isolates as Belonging to Berkeleyomyces Rouxiae
Bayesian inference analysis based on the concatenated sequences of MCM7 and RPB2 genes showed that all four B. rouxiae isolates clustered within the B. rouxiae subclade of the genus Berkeleyomyces (Figure 3). The subclade of B. rouxiae was phylogenetically separated from that of the closely related species, B. basicola. The phylogenetic positions of our strains confirmed their B. rouxiae identity.
3.3. Phylogenetic Analysis and SIX Gene Profiles Places the New Fov Isolates with Other Australian Fov, but Suggests the Fo Isolates Are Unique
To resolve the phylogenetic positions of the F. oxysporum isolates within the pathogen forms that infect cotton, Bayersian analysis was performed using concatenated partial sequences of TEF-1α, mtSSU rDNA, PHO, and NIR gene sequences. The Fov representative isolates belonging to distinct lineages I to V were retrieved from previous studies [9,21] and were used to anchor this phylogey (Figure 4). The topology of the phylogeny can be correlated with race and geographic origin. Lineage I contains mostly race 3 strains from Egypt, and race 5 from Sudan. Lineage II contains race 1 and 2, primarily from the United States, along with race 6 from Brazil. Lineage III contains race 8 strains from China. Lineage IV has strains of race 4 and race 7 from India and China, respectively. Lineage IV is represented by the Australian biotypes, namely VCG01111 and VCG01112 (Figure 4). An additional lineage, independent of lineage I to V, was identified in another study [21] and it contained F. oxysporum isolates that were endophytic or slightly pathogenic on cotton plants.
With the lineages of Fov correctly defined by the topology of phylogenetic tree, the four Fov isolates, namely Fov SG1, Fov SG26, Fov SG55 and Fov TH1, were phylogenetically positioned within Lineage V (Figure 4). They grouped amongst the VCG01111 isolates while further distinctions allowed VCG01112 isolates to form a subclade within lineage V. Of the 4 Fusarium oxysporum isolates, Fo BRF1 descended from an ancestral split from Lineage V, suggesting that it shares the same evolutionary descent as lineage V isolates (Figure 4). The divergence event separated the lineage that gave rise to Fo BRF1 from the lineage that gave rise to lineage V. On the other hand, Fo BRF2, Fo SHF6 and Fo WRF2 were phylogenetically clustered within a lineage that is independent of lineages I to V (Figure 4). Within this phylogroup, there are other F. oxysporum strains isolated from the rhizosphere of wild relatives of cotton in Australia and they appeared to cause little to no Fusarium wilt symptoms on cotton plants [21].
To ascertain whether any of the F. oxysporum isolates were Fov, we used the marker FovSIX6 that specifically detects the presence of SIX6 in the Australian Fov biotypes [20]. FovSIX6 was detected in Fov SG1, Fov SG26, Fov SG55, Fov TH1 but the primers did not amplify a product in Fo BRF1, Fo BRF2, Fo SHF6 and Fo WRF2 (Table 2). To further validate these results, the FovSIX6 gene products were sequenced and then used to construct a Bayesian inference phylogeny comprised of SIX6 gene sequences of other pathogen forms of F. oxysporum (Figure 5). Fov-specific SIX6 genes showed that the Fov isolates presented here and other Fov representatives formed a distinct lineage that is separated from the rest of the F. oxysporum strains (Figure 5). This confirmed that our Fov isolates included in this study belongs to the Australian biotypes.
To confirm the absence of SIX6 in the F. oxysporum isolates and to generate an effector gene profile for each SIX gene, we performed a diagnostic PCR screen on these isolates using a set of universal primers for SIX1-SIX14. These primers were developed in a previous study [19]. The results showed that Fov SG1, Fov SG26, Fov SG55 and Fov TH1 possessed SIX11, SIX13 and SIX14 in addition to SIX6 (Table 2). The effector profiles of these Fov isolates are consistent in comparison to those of the other Fov accessions (BRIP) characterised in the previous study [19]. None of the SIX genes were detected in Fo BRF1, Fo BRF2, Fo SHF6 and Fo WRF2 suggesting that they are most likely absent in these isolates (Table 2).
3.4. Berkeleyomyces Rouxiae Isolates Showed Variability in Disease Severity on Cotton
Sicot746 B3F plants inoculated with B. rouxiae were assessed for both internal and external symptoms at 15 dpi (Figure 6). The isolates 22BRR77 and BRR4 significantly reduced plant height and weight of Sicot746 B3F when compared with the uninoculated controls (Figure 6A-C, Figure 7A). These plants also developed black lesions on the epidermis of stems at 1-6 cm above the basal node (Figure 7A). Internal discolouration was observed in the vascular tissues of some of the stems. Furthermore, the roots of 22BRR77 and BRR4 inoculated plants appeared blackened and stunted, a characteristic symptom of black root rot (Figure 7A). The leaves of these plants also displayed significant wilt symptoms (Figure 6A, 6F). Reductions in weight and height were negatively associated with black discolouration in the lower stems of the plants and the roots of the symptomatic plants (Figure 6D-E, Figure 7A). These symptoms appeared to develop uniformly amongst infected plants (Figure S5-S7). Plants inoculated with StrB22 and RVB4.1 did not have any visible external symptoms (Figure 6, Figure S5-S6) and their plant height and weight were comparable to that of the uninoculated controls (Figure 6B-C). The roots of these plants were still discoloured but to a lesser extent when compared to 22BRR77 and BRR4 inoculated plants (Figure 6D, Figure 7A, Figure S5-S6).
Reisolation of B. rouxiae from B. rouxiae-inoculated plants on 10% carrot agar plants produced characteristic dark colonies, although not all of them were identified as positives (Figure S8). A root reisolation frequency of 67% and 68.3% were observed for plants inoculated with 22BRR77 and BRR4, respectively (Figure 6G). The reisolation frequency was higher in the stem than in the roots, with 96.9% and 100% of the basal stem issues being identified as positives for 22BRR77 and BRR4 inoculated plants, respectively (Figure 6G). Above-ground stems and petiole tissues showed less than 10% reisolation frequency for 22BRR77 and BRR4. Reisolation frequency for RVB4.1 was less than 10% for all tissue types assayed. No positives were identified from StrB22 inoculated plants or uninoculated control plants.
3.5. Isolates of Fusarium Oxysporum f. sp. Vasinfectum Varied in Their Virulence towards Cotton
Fov-inoculated plants did not show any significant differences in plant weight at harvest (Figure 8B). However, Fov SG1 and Fov SG55 inoculated plants were significantly shorter than uninoculated plants, Fov TH1 and Fov SG26 inoculated plants (Figure 8C). A brown discolouration was observed at the basal stem and in the roots of plants inoculated with the Fov isolates (Figure 7B, 8D-E) but these symptoms were only sporadically detected (Figure S9-S11). However, these effects were significant when compared to the uninoculated controls (Figure 8D-E). Fov SG1 inoculated plants had 82% roots discoloured which were significantly higher than the plants inoculated with the other isolates (Figure 8D). Fov SG1, Fov SG55 and Fov TH1 inoculated plants also showed significantly higher levels of leaf wilting when compared to that of the uninoculated controls or plants inoculated with Fov SG26 (Figure 8F). Internally, vascular discolouration, which is a characteristic symptom of Fusarium wilt, was consistently identified in the main root nodes and the cut stems of all Fov inoculated plants (Figure S9-S11). Disease incidence, expressed as the number of plants carrying internal stem discolouration over total number of plants assessed, was estimated at 100% for Fov SG1 and Fov SG26, 92.8% for Fov SG55, and 43.7% for Fov TH1.
Tissue reisolations produced white to light pink colonies which were further confirmed to be F. oxysporum-like under a microscope (Figure S8). Reisolation frequencies were high in both the roots (53.1 - 96.9%) and lower stems (71.9 - 92.9%) for all Fov inoculated plants (Figure 8G). The upper stem reisolation frequency was the highest in Fov SG1 inoculated plants (85.7%) and lowest in Fov SG26 inoculated plants (15.6%). The presence of Fov at the base of the petiole was detected in all but Fov SG26, amid at a relatively low reisolation frequency of 7.8-14.3% (Figure 8G).
3.6. Virulence of Fusarium Oxysporum on Cotton
Plants inoculated with the four F. oxysporum isolates were harvested at 17 dpi which means that the plants were a lot younger than the Fov inoculated plants at harvest (27 dpi) (Figure 9A). No significant differences in plant height and weight were detected between plants inoculated with the F. oxysporum isolates and the uninoculated controls (Figure 9 B-C). Light brown discolouration in the roots and the stems were evident (Figure 9 D-E). Internally, black discolouration in the vascular regions of the stems was also observed (Figure 9C). These effects are significant when compared to uninoculated controls (Figure 9D-E), although the symptoms were not consistently identified amongst the plants of each treatment group (Figure S12-S14). Based on the internal discolouration of the stem, F. oxysporum inoculated plants showed disease incidences at 71.4% for Fo WRF2, 60% for Fo SHF6, 58.3% for Fo BRF2, and 30.8% for Fo BRF1. The inoculated plants also showed significantly elevated leaf wilting when compared to the uninoculated controls (Figure 9F).
Reisolation from plants inoculated with the F. oxysporum isolates showed that the colour of recovered colonies was comparable to that of the pure culture (Figure S8). Reisolation frequencies were 83.3 - 90.4% from the roots, 57.1 - 80.8% from the lower stem regions, and 7.1 - 25% from the upper stem regions (Figure 9G). F. oxysporum-like colonies were only recovered at a low frequency (2.5-11.5%) from the petioles of Fo BRF1, Fo SHF6 and Fo BRF2 inoculated plants.
4. Discussion
In this study, we successfully employed a simple and robust inoculation method to assess and differentiate virulence of both B. rouxiae and Fusarium spp. on cotton. Using unfiltered suspension for root dipping and soil inculcation, virulence of B. rouxiae was visually detected at 15 dpi in comparison to 19 to 30 dpi using artificially or naturally infested soil [14]. Similarly, screening for Fusarium wilt typically takes 1 - 3 months for disease phenotypes to be expressed and assessed in different plant species [29,30,31]. In our study, we were able to terminate and assess the aggressiveness of cotton-Fusarium spp. as soon as 15 dpi. A seedling screening system such as this can be scaled up to provide preliminary assays for pathogen virulence at a high throughput. A reliable screening system is important in determining the host range of specific plant pathogens and genetics of plant and pathogen interactions.
Spores and mycelia were not filtered before being applied to the plants and the soil. Given that spore production varies significantly amongst the different Fov and Berkeleyomyces isolates (Figure S3-S4), a pre-determined number of spores per pot can still lead to variations in the build-up of inoculum for infection. Instead, sufficient spores and mycelia were applied to the soil at a high density. Fine roots and the new root growth in soil saturated with the fungus can achieve good infection rates, thus providing uniformity in infection levels. Furthermore, plant growth under a controlled environment with a set of constant parameters can reduce variability in growth and varied disease expression due to environmental effects. Although varied amount of spores and mycelia in the inoculum applied may also contribute to varied disease severity.
The result shows that B. rouxiae BRR4 and 22BRR77 were clearly pathogenic towards cotton. The discolourations in the stems were distinct and appeared more severe when compared to the discolouration levels in the roots (Figure 7). This contrasts the pot trial of another study in which root discolouration appeared to be the most dominant trait associated with the black root rot of cotton [3]. In that study, Berkeleyomyces spore culture was mixed with the potting mix and seeds are directly sown into the Berkeleyomyces-infested soil. This inoculation method mimics the field conditions in which residual Berkeleyomyces spores in the soil provides a natural inoculum for the germinating cotton seeds.
In plants severely affected by 22BRR77, some discolouration was observed in the vascular regions of the stems. However, black root rot infection is commonly observed on the epidermis of below ground hypocotyls and roots. Vascular infection associated with Berkeleyomyces is rarely detected in Australia. Although it has been shown that in the presence of root-knot nematodes, B. basicola can induce necrosis within the vascular tissues of cotton plants [32]. Therefore, open lesions and wounds caused by nematodes or wounds such as those artificially induced during root dipping and washing in our study can potentially lead to the movement of B. rouxiae through the vasculature in cotton plants. Never-the-less, both BRR2 and 22BRR77 were only recovered at a low level in the arial parts of the plants suggesting that their ability to infect plants is primarily through the roots and lower parts of the plants. The strong discolouration on the stem surface could be used as a trait marker to identify and confirm Berkeleyomyces infections in the cotton field. Despite being identified as B. rouxiae, RVB4.1 and StrB22 did not have any effects on plant growth, suggesting that there is variability in the ability of B. rouxiae strains to infect specific plant species. This may potentially be explained by differences in the effector profile of these isolates. Pac-bio sequencing of both RVB4.1 and 22BRR77 are underway. Differences between the genomes of these two isolates will be investigated.
DNA sequences have been used to investigate the evolutionary relationships among the Fov races. Based on multiple conserved gene sequences, different races of Fov were separated into five distinct lineages [7,9,16,33]. In our study, the four Fov isolates conformed to the other Australian Fov biotypes in lineage V and the SIX gene analysis further supported their phylogenetic positions within this lineage. Recently, it was shown that non-Australian Fov isolates are a part of lineage V and can be distinguished phylogenetically from the Australian isolates [33]. Taken together, these results support the proposal that the different lineages of Fov are independently evolving [10], highlighting a need to further investigate the diversity of Fov isolates beyond the current classification system.
All 4 F. oxysporum isolates were isolated from the hypocotyls of cotton seedlings displaying the collar rot disease [12]. They were not initially identified as Fov due to the fact that the plants the isolates were obtained from did not display Fusarium wilt symptoms. Effector gene profiling based on the presence or absence of SIX genes has been used to differentiate the different pathogen forms of F. oxysporum as well as F. oxysporum endophytes in a previous study [19]. By using this diagnostic system, it was determined that none of the 4 F. oxysporum isolates used in this study possessed any SIX genes. Despite not having any SIX genes, these isolates were able to colonise the vascular tissues within the stems and cause disease on cotton plants. These results suggest that they behave in a manner characteristic of a vascular wilt pathogen.
Fo BRF1 share the same evolutionary descent as lineage V but appeared to form a lineage that is independent of lineage V. Similarly, Fo BRF2, Fo SHF6 and Fo WRF2 were clustered within a lineage that is independent of the Fov lineages. This lineage was previously defined by F. oxysporum isolates that were endophytic or slightly pathogenic on cotton cultivars [21]. Despite having reduced virulence on cotton cultivars, the authors observed that these isolates were highly pathogenic on a wild relative of cotton, therefore suggesting that these cotton hosts could serve as inoculum reservoir for the co-occurrence of these 'wild' Fov populations in the field [21]. Moreover, selection pressures exerted by the changing environment and Fov resistance in cotton cultivars may have favoured the selection of the 'wild' Fov populations with enhanced pathogenicity. The 4 F. oxysporum isolates used in this study were part of a collection consisting of 186 F. oxysporum isolates recovered from cotton seedlings showing the collar rot disease [12]. Fusarium wilt symptoms were not observed on these plants. Out of this collection, only 21.5% of the isolates grouped with the Australian biotypes lineage V while 78.5% of the isolates clustered with the other lineages [12]. Our results are consistent with these findings. Taken together, these results collectively suggests that under disease inducive conditions, F. oxysporum strains can act opportunistically on cotton plants and their ability to colonise and exert virulence on hosts are dependent on host genotypes. The specificity of the Fusarium-cotton interactions may have been encoded by the cotton hosts. Thus, the F. oxysporum populations may have been enriched for mildly virulent strains that are capable of causing Fov-like symptoms on cotton plants in the field. They apparently lack the SIX gene effectors and are part of lineages that are independent of the Fov phylogroups.
5. Conclusions
Black root rot and Fusarium wilt are significant diseases constraining cotton production world-wide. Here we adopted a plant screening system where the virulence of these pathogens can be assessed quickly and effectively on cotton plants. Our results contribute towards understanding the pathogenicity of these pathogen populations in the cotton fields and allow strategies in surveillance and detection to be improved.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: plant growth before and after inoculation with Berkeleyomyces rouxiae isolates; Figure S2: plant growth before and after inoculation with Fusarium oxysporum f. sp. vasinfectum isolates; Figure S3: the spore size and growth rates of Berkeleyomyces rouxiae isolates after 3 weeks of growth on half-strength PDA plates; Figure S4: the spore size and growth rates of Fusarium oxysporum isolates after seven days of growth on half-strength PDA plates; Figure S5-S7: symptomatology of Sicot746 B3F seedlings inoculated with Berkeleyomyces rouxiae at harvest (15 — 20 days post inoculation); Figure S8: reisolation of the Berkeleyomyces rouxiae and Fusarium oxysporum isolates from cotton plants on half-strength PDA plates (reverse view); Figure S9-S11: symptomatology of Sicot746 B3F seedlings inoculated with Fusarium oxysporum f. sp. vasinfectum at harvest (27 — 34 days post inoculation). Figure S12-S14: symptomatology of Sicot746 B3F seedlings inoculated with Fusarium oxysporum spp. at harvest (17 — 22 days post inoculation). Table S1: primers used for PCR in this study; Table S2: spore concentrations of the liquid cultures determined on the day of inoculation.
Author Contributions
Conceptualization, A.C., E.A.B.A. and D.M.G.; methodology, A.C.; software, A.C.; validation, A.C., E.A.B.A. and D.M.G.; formal analysis, A.C.; investigation, A.C.; resources, D.P.L., L.J.S., D.K.; data curation, A.C.; writing—original draft preparation, A.C.; writing—review and editing, A.C., D.P.L., L.J.S., D.K., E.A.B.A. and D.M.G.; supervision, A.C., E.A.B.A. and D.M.G.; project administration, E.A.B.A and D.M.G.; funding acquisition, E.A.B.A. and D.M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Australian Research Council Research Hub for Sustainable Crop Protection (project number IH190100022) and funded by the Australian Government.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s. Gene sequences generated in this study have been deposited in Genbank and can be accessed under project 9788723 (submitted).
Acknowledgments
We would like to kindly thank Leela Rizal for the preliminary assessment made on the four Fov isolates. We would also like to thank Andrea Matthews for the technical support she provided during this project.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Khan, M.A.; Wahid, A.; Ahmad, M.; Tahir, M.T.; Ahmed, M.; Ahmad, S.; Hasanuzzaman, M. World Cotton Production and Consumption: An Overview. In: Ahmad, S., Hasanuzzaman, M. (eds) Cotton Production and Uses. Springer, Singapore. [CrossRef]
- Le, D.P.; Nguyen, C.P.T.; Kafle, D.; Scheikowski, L.; Montgomery, J.; Lambeth, E.; Thomas, A.; O’Keeffe, K.; Shakeshaft, B.; Young, A.; et al. Surveillance, Diversity and Vegetative Compatibility Groups of Fusarium oxysporum f. sp. vasinfectum Collected in Cotton Fields in Australia (2017 to 2022). Pathogens 2022, 11, 1537. [Google Scholar] [CrossRef] [PubMed]
- Le, D.P.; Gregson, A.; Jackson, R. Identification of Berkeleyomyces rouxiae causing black root rot disease on cotton seedlings in New South Wales, Australia. J Gen Plant Pathol 2022, 88, 155–159. [Google Scholar] [CrossRef]
- Nel, W.J.; Duong, T.A.; Wingfield, B.D.; Wingfield, M.J.; de Beer, Z.W. A new genus and species for the globally important, multihost root pathogen Thielaviopsis basicola. Plant Pathol 2018, 67, 871–882. [Google Scholar] [CrossRef]
- Nel, W.J.; Duong, T.A.; de Beer, Z.W.; Wingfield, M.J. Black root rot: a long known but little understood disease. Plant Pathol 2019, 68, 834–842. [Google Scholar] [CrossRef]
- Nehl, D.B.; Allen, S.J.; Mondal, A.H.; Lonergan, P.A. Black root rot: a pandemic in Australian cotton. Australas Plant Pathol 2004, 33, 87–95. [Google Scholar] [CrossRef]
- Davis, R.M.; Colyer, P.D.; Rothrock, C.S.; Kochman, J.K. Fusarium Wilt of Cotton: Population Diversity and Implications for Management. Plant Dis 2006, 90, 692–703. [Google Scholar] [CrossRef]
- Davis, R.; Moore, N.; Kochman, J. Characterisation of a population of Fusarium oxysporum f. sp. vasinfectum causing wilt of cotton in Australia. Aust J Agric Res 1996, 47, 1143–1156. [Google Scholar] [CrossRef]
- Skovgaard, K.; Nirenberg, H.I.; O'Donnell, K.; Rosendahl, S. Evolution of Fusarium oxysporum f. sp. vasinfectum races inferred from multigene genealogies. Phytopathology 2001, 91, 1231–1237. [Google Scholar] [CrossRef]
- Fernandez, D.; Assigbese, K.; Dubois, M.P.; Geiger, J.P. Molecular characterization of races and vegetative compatibility groups in Fusarium oxysporum f. sp. vasinfectum. Appl Environ Microbiol 1994, 60, 4039–4046. [Google Scholar] [CrossRef]
- Wang, B.; Brubaker, C.L.; Tate, W.; Woods, M.J.; Matheson, B.A.; Burdon, J.J. Genetic variation and population structure of Fusarium oxysporum f.sp. vasinfectum in Australia. Plant Pathol 2006, 55, 746–755. [Google Scholar] [CrossRef]
- Le, D.P.; Tran, T.T.; Gregson, A.; Jackson, R. TEF1 sequence-based diversity of Fusarium species recovered from collar rot diseased cotton seedlings in New South Wales, Australia. Australas Plant Pathol 2020, 49, 277–284. [Google Scholar] [CrossRef]
- Bell, A.A.; Gu, A.; Olvey, J.; Wagner, T.A.; Tashpulatov, J.J.; Prom, S.; Quintana, J.; Nichols, R.L.; Liu, J. Detection and characterization of Fusarium oxysporum f. sp. vasinfectum VCG0114 (Race 4) isolates of diverse geographic origins. Plant Dis 2019, 103, 1998–2009. [Google Scholar] [CrossRef] [PubMed]
- Le, D.P.; Nguyen, C.P.T.; Manjarrez, M.; Giblot-Ducray, D. A diagnostic guide for Berkeleyomyces rouxiae causing black root rot of cotton. Plant Health Prog 2024. [Google Scholar] [CrossRef]
- Kim, Y.; Hutmacher, R.; Davis, R. Characterization of California isolates of Fusarium oxysporum f. sp. vasinfectum. Plant Dis 2005, 89, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Abdelraheem, A.; Lujan, P.; Idowu, J.; Sullivan, P.; Nichols, R.; Wedegaertner, T.; Zhang, J. Detection and characterization of Fusarium wilt (Fusarium oxysporum f. sp. vasinfectum) race 4 causing Fusarium wilt of cotton seedlings in New Mexico. Plant Dis 2021, 105, 3353–3367. [Google Scholar] [CrossRef]
- Ogle, H.; Stirling, A.; Dart, P. Pathogenicity of fungi associated with seedling disease of cotton. Aust J Exp Agric 1993, 33, 923–929. [Google Scholar] [CrossRef]
- Harris, J.L. Safe, low-distortion tape touch method for fungal slide mounts. J Clin Microbiol 2000, 38, 4683–4684. [Google Scholar] [CrossRef]
- Czislowski, E.; Zeil-Rolfe, I.; Aitken, E.A.B. Effector Profiles of Endophytic Fusarium Associated with Asymptomatic Banana (Musa sp.) Hosts. Int J Mol Sci 2021, 22, 2508. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Rep, M.; Wang, B.; Ashton, A.; Dodds, P.; Ellis, J. Variation in potential effector genes distinguishing Australian and non-Australian isolates of the cotton wilt pathogen Fusarium oxysporum f.sp. vasinfectum. Plant Pathol 2011, 60, 232–243. [Google Scholar] [CrossRef]
- Wang, B.; Brubaker, C.L.; Summerell, B.A.; Thrall, P.H.; Burdon, J.J. Local origin of two vegetative compatibility groups of Fusarium oxysporum f. sp. vasinfectum in Australia. Evol Appl 2010, 3, 505–524. [Google Scholar] [CrossRef]
- O'Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. 1990, 31, 315–322. [Google Scholar]
- de Beer, Z.W.; Duong, T.A.; Barnes, I.; Wingfield, B.D.; Wingfield, M.J. Redefining Ceratocystis and allied genera. Stud Mycol 2014, 79, 187–219. [Google Scholar] [CrossRef] [PubMed]
- Fourie, A.; Wingfield, M.J.; Wingfield, B.D.; Barnes, I. Molecular markers delimit cryptic species in Ceratocystis sensu stricto. Mycological Progress 2015, 14, 1–18. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol biol evol 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Morrison, S.; Gregson, A.; Le, D.P.; Urquhart, A.S.; Smith, L.J.; Aitken, E.A.B.; Gardiner, D.M. Fluorescently Tagged Verticillium dahliae to Understand the Infection Process on Cotton (Gossypium hirsutum) and Weed Plant Species. Pathogens 2024, 13, 442. [Google Scholar] [CrossRef]
- Chen, A.; Sun, J.; Matthews, A.; Armas-Egas, L.; Chen, N.; Hamill, S.; Mintoff, S.; Tran-Nguyen, L.T.T.; Batley, J.; Aitken, E.A.B. Assessing Variations in host resistance to Fusarium oxysporum f sp. cubense race 4 in Musa species, with a focus on the subtropical race 4. Front Microbiol 2019, 10, 1062. [Google Scholar] [CrossRef]
- Cong, L.L.; Sun, Y.; Wang, Z.; Kang, J.M.; Zhang, T.J.; Biligetu, B.; Yang, Q.C. A rapid screening method for evaluating resistance of alfalfa (Medicago sativa L.) to Fusarium root rot. Can J Plant Pathol 2018, 40, 61–69. [Google Scholar] [CrossRef]
- Matthews, A.; Muthukumar, S.P.; Hamill, S.; Aitken, E.A.; Chen, A. Impact of inoculum density of Fusarium oxysporum f. sp. zingiberi on symptomatic appearances and yield of ginger (Zingiber officinale Roscoe). Access Microbiol 2023, 5, 000605–v000603. [Google Scholar] [CrossRef]
- Walker, N.R.; Kirkpatrick, T.L.; Rothrock, C.S. Effect of temperature on and histopathology of the interaction between Meloidogyne incognita and Thielaviopsis basicola on cotton. Phytopathology 1999, 89, 613–617. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Li, S.; Lu, X.; Gao, H.; Wang, X.; Ma, Y.; Zhang, X.; Wang, P.; Ma, P. Identification of a new genotype of Fusarium oxysporum f. sp. vasinfectum on cotton in China. Plant Dis 2015, 99, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Colony morphology of Berkeleyomyces rouxiae and Fusarium oxysporum isolates used in this study. A) B. rouxiae isolates RVB4.1, BRR4, 22BRR77, and StrB22 grown for 2 weeks on 10% carrot agar. B) Fusarium oxysporum f. sp. vasinfectum isolates Fov SG1, Fov SG26, Fov SG55, and Fov TH1 grown for 2 weeks on half-strength potato dextrose agar (PDA). (C) Fusarium oxsporum isolates Fo BRF1, Fo BRF2, Fo WRF2, and Fo SHF6 grown for 2 weeks on half-strength PDA.
Figure 1.
Colony morphology of Berkeleyomyces rouxiae and Fusarium oxysporum isolates used in this study. A) B. rouxiae isolates RVB4.1, BRR4, 22BRR77, and StrB22 grown for 2 weeks on 10% carrot agar. B) Fusarium oxysporum f. sp. vasinfectum isolates Fov SG1, Fov SG26, Fov SG55, and Fov TH1 grown for 2 weeks on half-strength potato dextrose agar (PDA). (C) Fusarium oxsporum isolates Fo BRF1, Fo BRF2, Fo WRF2, and Fo SHF6 grown for 2 weeks on half-strength PDA.
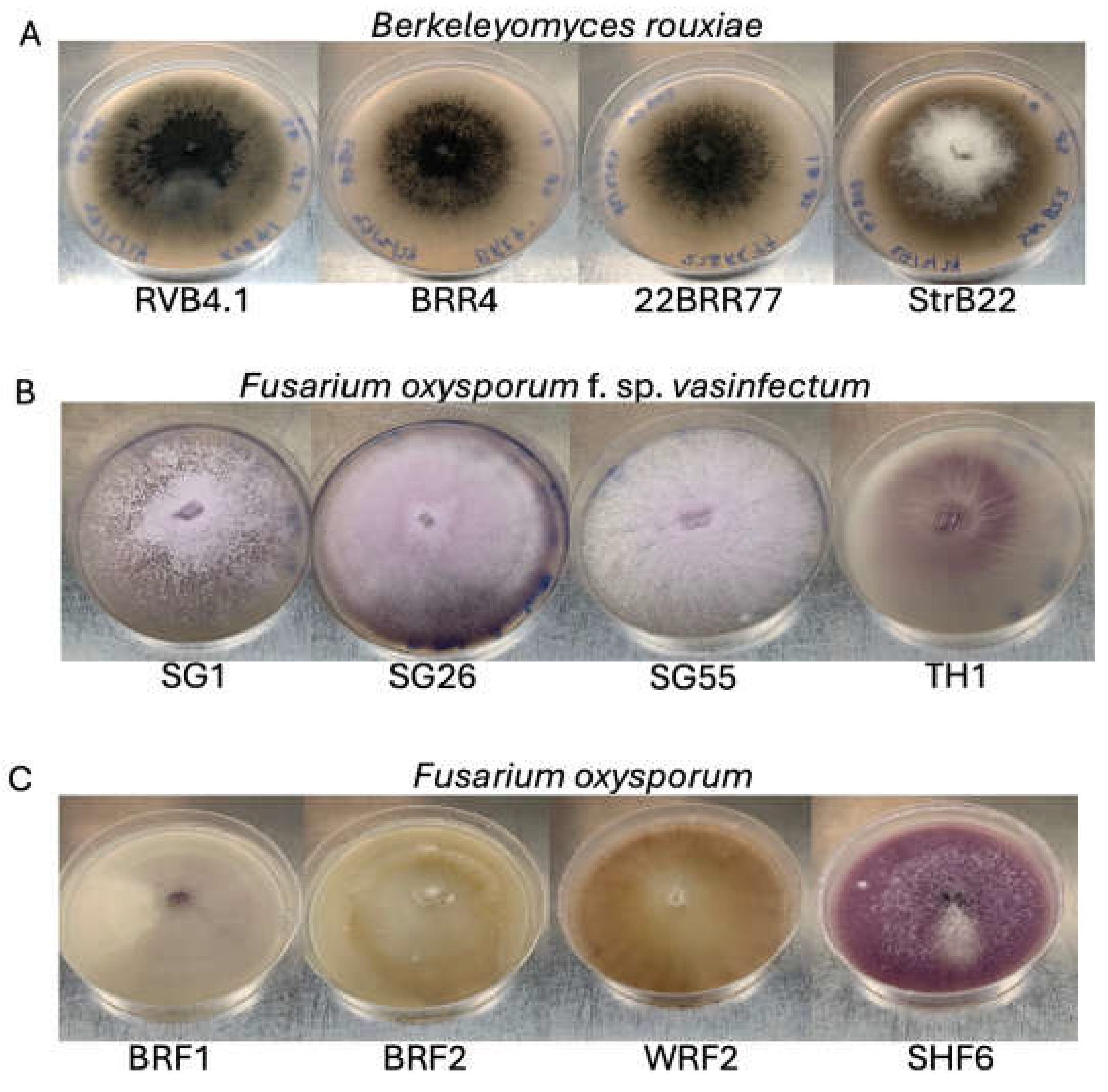
Figure 2.
Compound microscopic images of Berkeleyomyces rouxiae and Fusarium oxysporum isolates used in this study. (A-B) B. rouxiae isolate RVB4.1. (C-D) B. rouxiae isolate 22BRR77. (E-F) B. rouxiae isolate BRR4 (G-H) B. rouxiae isolate StrB22. (I) Fusarium oxysporum f. sp. vasinfectum isolate (Fov) SG1. (J) Fov isolate SG26. (K) Fov isolate SG55. (L) Fov isolate TH1. (M) Fusarium oxsporum (Fo) isolate BRF1. Inset: a cluster of conidia. (N) Fo isolate BRF2. Inset: microconidia. (O) Fo isolate SHF6. Inset: a cluster of macroconidia. (P) Fo isolate WRF2. Inset: microconidia. e = endoconidia; ec = endoconidia chains; a = aleuriospores; c = microconidia; ma = macroconidia. Bars indicate a scale of 15 µm.
Figure 2.
Compound microscopic images of Berkeleyomyces rouxiae and Fusarium oxysporum isolates used in this study. (A-B) B. rouxiae isolate RVB4.1. (C-D) B. rouxiae isolate 22BRR77. (E-F) B. rouxiae isolate BRR4 (G-H) B. rouxiae isolate StrB22. (I) Fusarium oxysporum f. sp. vasinfectum isolate (Fov) SG1. (J) Fov isolate SG26. (K) Fov isolate SG55. (L) Fov isolate TH1. (M) Fusarium oxsporum (Fo) isolate BRF1. Inset: a cluster of conidia. (N) Fo isolate BRF2. Inset: microconidia. (O) Fo isolate SHF6. Inset: a cluster of macroconidia. (P) Fo isolate WRF2. Inset: microconidia. e = endoconidia; ec = endoconidia chains; a = aleuriospores; c = microconidia; ma = macroconidia. Bars indicate a scale of 15 µm.
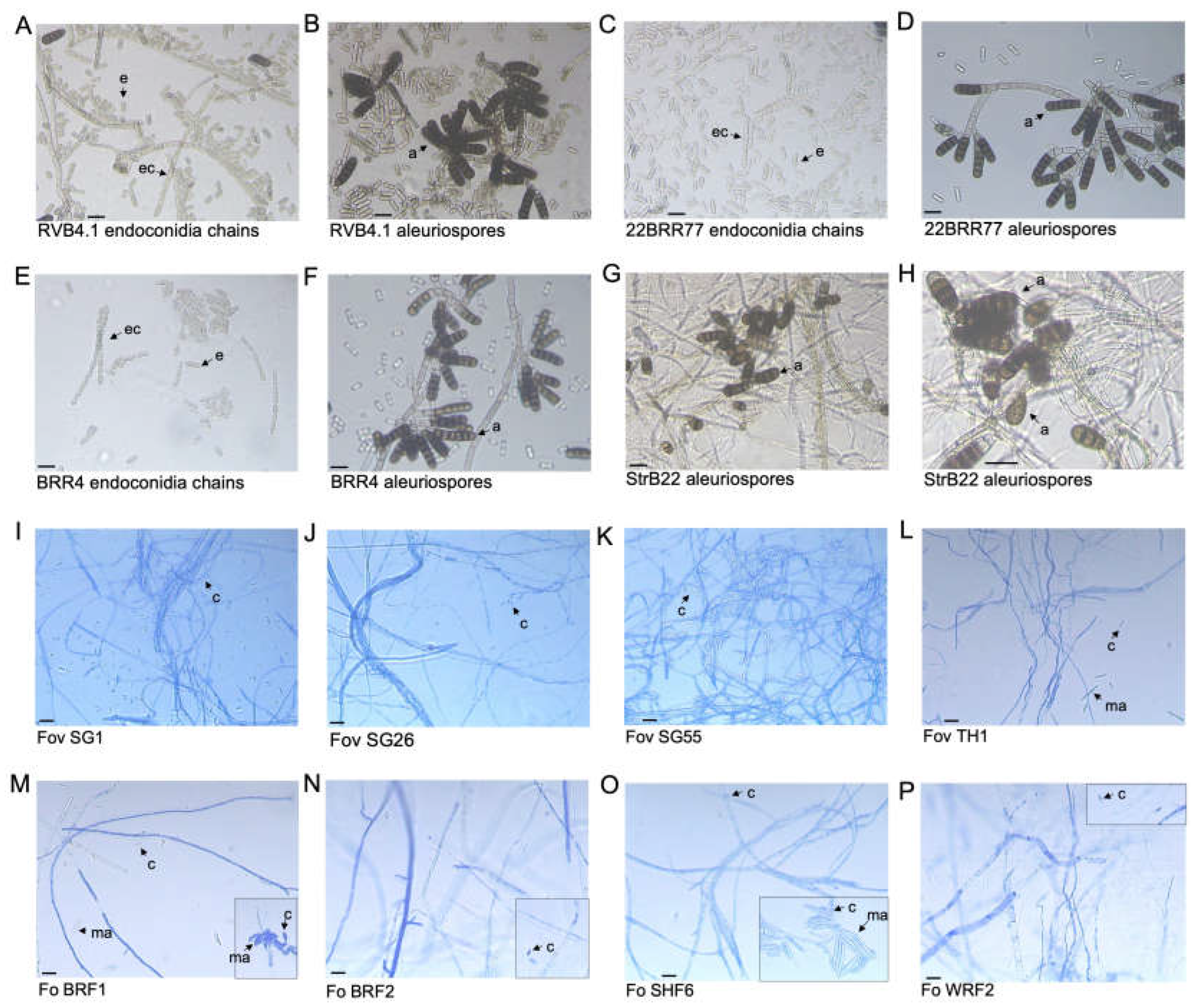
Figure 3.
Phylogenetic positions of Berkeleyomyces rouxiae isolates within the genus Ceratocystidaceae determined using Bayesian inference. Phylogenetic relationships reconstructed using concatenated sequences of MCM7 and RPB2 genes. Bars indicate a scale range of 0.05. Node values show the posterior probability. Genus is abbreviated as Ch. for Chalaropsis, C. for Ceratocystis, B. for Berkeleyomyces, L. for Lignincola.
Figure 3.
Phylogenetic positions of Berkeleyomyces rouxiae isolates within the genus Ceratocystidaceae determined using Bayesian inference. Phylogenetic relationships reconstructed using concatenated sequences of MCM7 and RPB2 genes. Bars indicate a scale range of 0.05. Node values show the posterior probability. Genus is abbreviated as Ch. for Chalaropsis, C. for Ceratocystis, B. for Berkeleyomyces, L. for Lignincola.
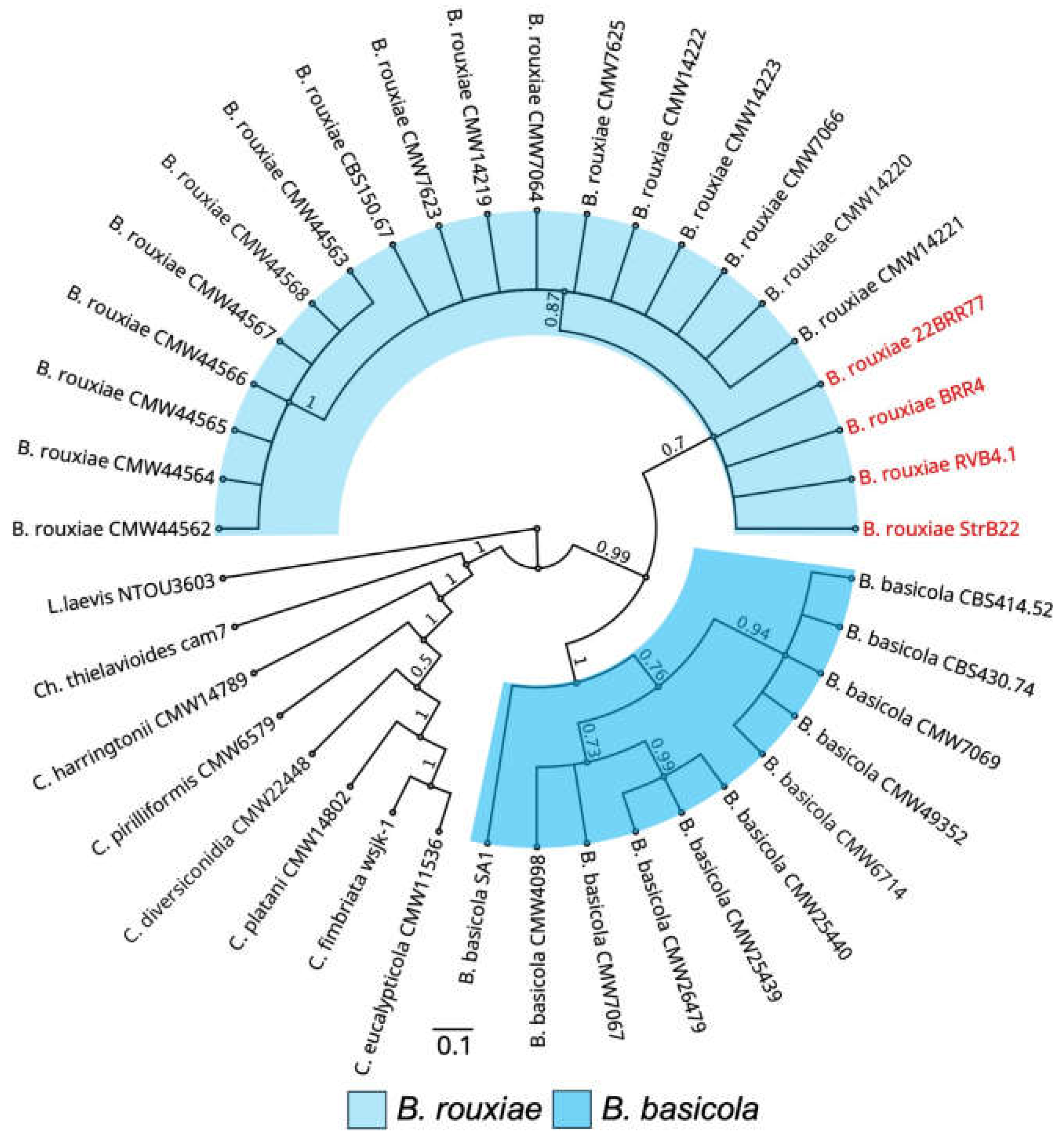
Figure 4.
Bayesian phylogeny of the Fusarium oxysporum isolates inferred from combined analysis of translation elongation factor, mitochondrial small subunit rDNA, nitrate reductase, and phosphate permease gene sequences. Fusarium oxysporum f. sp. vasinfectum isolates are classified into lineages I-V [7,9]. The endophytic and slightly pathogenic F. oxysporum isolates from cotton were classified as a distinct lineage [21] and these isolates were characterised to be either non-pathogenic (green circle) or slightly pathogenic (orange circles) on cotton plants. VCG01111 and VCG01112 are abbreviated as VCG11 and VCG12, respectively. Bars indicate a scale range of 0.05. Node values show the posterior probability.
Figure 4.
Bayesian phylogeny of the Fusarium oxysporum isolates inferred from combined analysis of translation elongation factor, mitochondrial small subunit rDNA, nitrate reductase, and phosphate permease gene sequences. Fusarium oxysporum f. sp. vasinfectum isolates are classified into lineages I-V [7,9]. The endophytic and slightly pathogenic F. oxysporum isolates from cotton were classified as a distinct lineage [21] and these isolates were characterised to be either non-pathogenic (green circle) or slightly pathogenic (orange circles) on cotton plants. VCG01111 and VCG01112 are abbreviated as VCG11 and VCG12, respectively. Bars indicate a scale range of 0.05. Node values show the posterior probability.
Figure 5.
Bayesian inference phylogenetic reconstruction of Secreted In Xylem 6 gene (SIX6) in Fusarium oxysporum f. sp. vasinfectum isolates and isolates from other formae speciales of Fusarium oxysporum. Focuc = Fusarium oxysporum f. sp. cucumerinum; Fol = Fusarium oxysporum f. sp. lycopersici; Fopas = Fusarium oxysporum f. sp. passiflora; Fon = Fusarium oxysporum f. sp. niveum; Fop = Fusarium oxysporum f. sp. pisi; Foradc = Fusarium oxysporum f. sp. radicis-cucumerinum; Foses = Fusarium oxysporum f. sp. sesami. Bars indicate a scale range of 0.05. Node values show the posterior probability.
Figure 5.
Bayesian inference phylogenetic reconstruction of Secreted In Xylem 6 gene (SIX6) in Fusarium oxysporum f. sp. vasinfectum isolates and isolates from other formae speciales of Fusarium oxysporum. Focuc = Fusarium oxysporum f. sp. cucumerinum; Fol = Fusarium oxysporum f. sp. lycopersici; Fopas = Fusarium oxysporum f. sp. passiflora; Fon = Fusarium oxysporum f. sp. niveum; Fop = Fusarium oxysporum f. sp. pisi; Foradc = Fusarium oxysporum f. sp. radicis-cucumerinum; Foses = Fusarium oxysporum f. sp. sesami. Bars indicate a scale range of 0.05. Node values show the posterior probability.
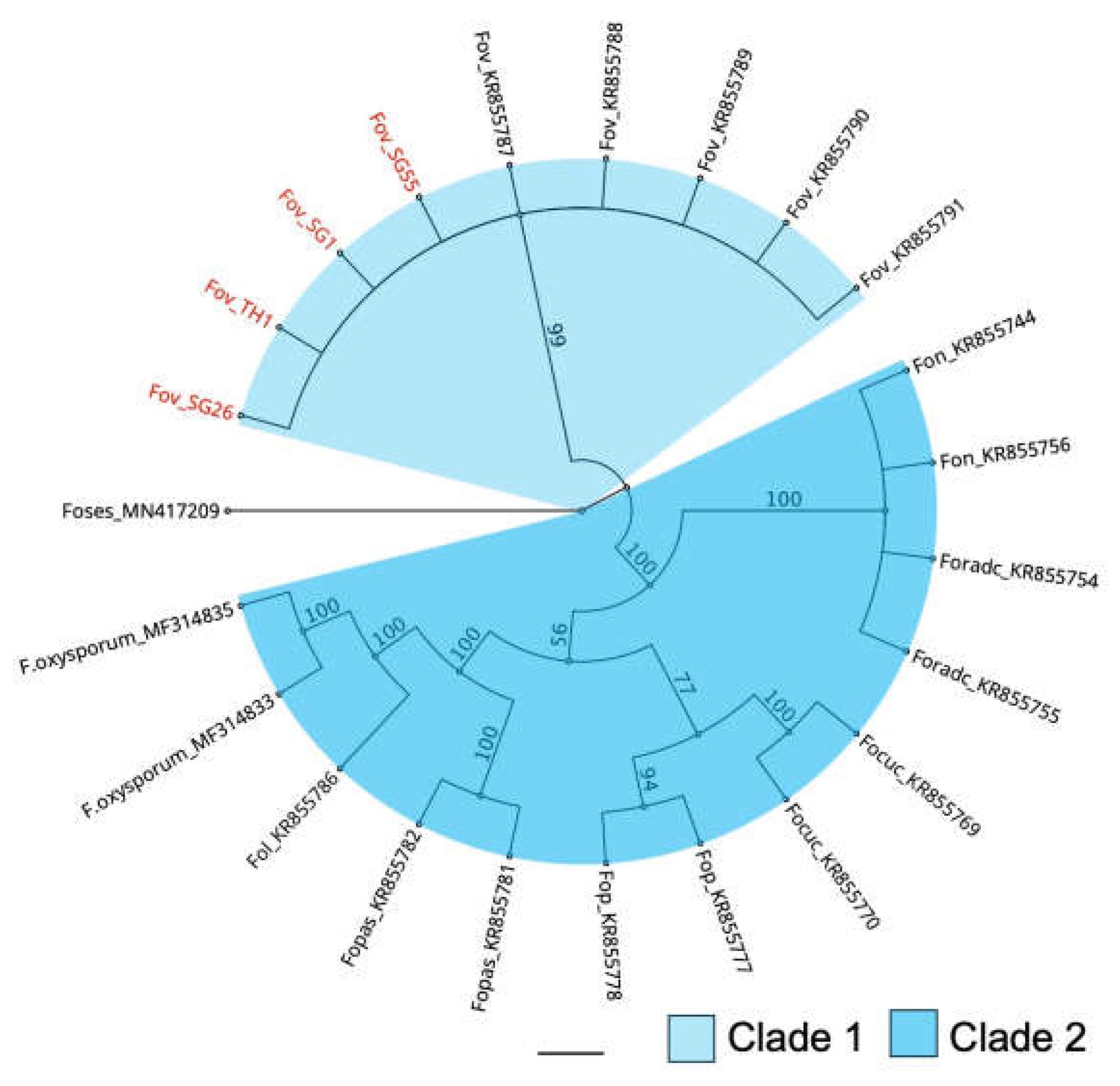
Figure 6.
Pathogenicity testing of Berkeleyomyces rouxiae isolates on cotton cv. Sicot746 B3F. (A) Plants at harvest (15 — 20 days post inoculation). (B) Total (above and below ground) plant weight. (C) Plant height. (D) Percentage of roots discoloured. (E) Total stem discoloured. (F) Percentage of leaves wilted or dropped. (G) Reisolations of B. rouxiae-like colonies on half-strength potato dextrose agar from the roots, stems and petioles of uninoculated plants and plants inoculated with the B. rouxiae isolates. Numbers indicate the number of samples assayed per tissue type. Letters indicate a significant separation of means by Tukey's range test at p = 0.05. Error bars indicate a 95% confidence interval.
Figure 6.
Pathogenicity testing of Berkeleyomyces rouxiae isolates on cotton cv. Sicot746 B3F. (A) Plants at harvest (15 — 20 days post inoculation). (B) Total (above and below ground) plant weight. (C) Plant height. (D) Percentage of roots discoloured. (E) Total stem discoloured. (F) Percentage of leaves wilted or dropped. (G) Reisolations of B. rouxiae-like colonies on half-strength potato dextrose agar from the roots, stems and petioles of uninoculated plants and plants inoculated with the B. rouxiae isolates. Numbers indicate the number of samples assayed per tissue type. Letters indicate a significant separation of means by Tukey's range test at p = 0.05. Error bars indicate a 95% confidence interval.
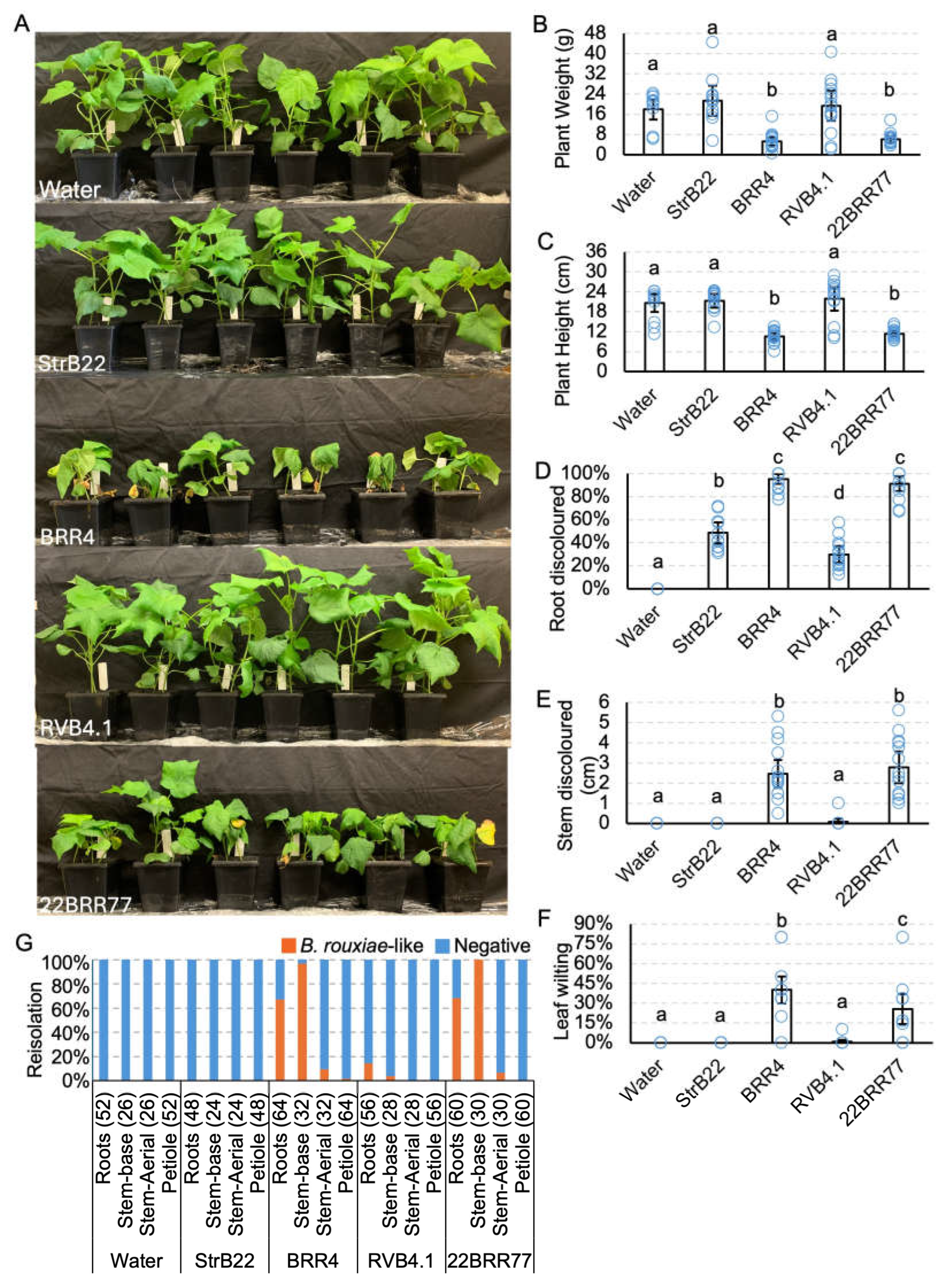
Figure 7.
Symptomatology of Berkeleyomyces rouxiae and Fusarium oxysporum isolates on cotton cv. Sicot746 B3F at harvest. Red vertical bars indicate a scale of 10 cm.
Figure 7.
Symptomatology of Berkeleyomyces rouxiae and Fusarium oxysporum isolates on cotton cv. Sicot746 B3F at harvest. Red vertical bars indicate a scale of 10 cm.
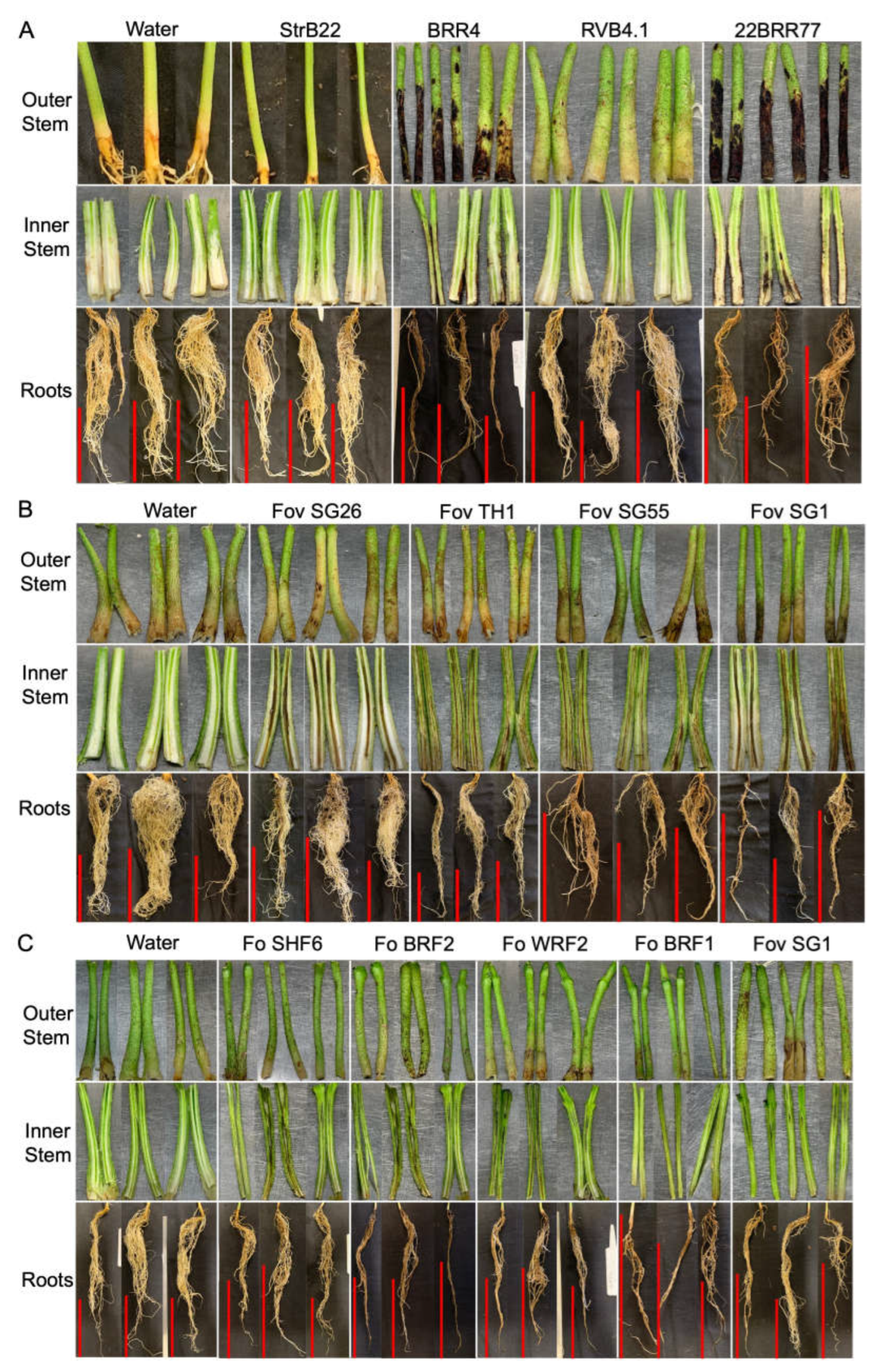
Figure 8.
Pathogenicity testing of Fusarium oxysporum f. sp. vasinfectum (Fov) isolates on cotton cv. Sicot746 B3F. (A) Plants at harvest (27 — 34 days post inoculation). (B) Total (above and below ground) plant weight. (C) Plant height. (D) Percentage of roots discoloured. (E) Total stem discoloured. (F) Percentage of leaves wilted or dropped. (G) Reisolations of F. oxysporum-like colonies on half-strength potato dextrose agar from the roots, stems and petioles of uninoculated plants and plants inoculated with the Fov isolates. Numbers indicate the number of samples assayed per tissue type. Letters indicate a significant separation of means by Tukey's range test at p = 0.05. Error bars indicate a 95% confidence interval.
Figure 8.
Pathogenicity testing of Fusarium oxysporum f. sp. vasinfectum (Fov) isolates on cotton cv. Sicot746 B3F. (A) Plants at harvest (27 — 34 days post inoculation). (B) Total (above and below ground) plant weight. (C) Plant height. (D) Percentage of roots discoloured. (E) Total stem discoloured. (F) Percentage of leaves wilted or dropped. (G) Reisolations of F. oxysporum-like colonies on half-strength potato dextrose agar from the roots, stems and petioles of uninoculated plants and plants inoculated with the Fov isolates. Numbers indicate the number of samples assayed per tissue type. Letters indicate a significant separation of means by Tukey's range test at p = 0.05. Error bars indicate a 95% confidence interval.
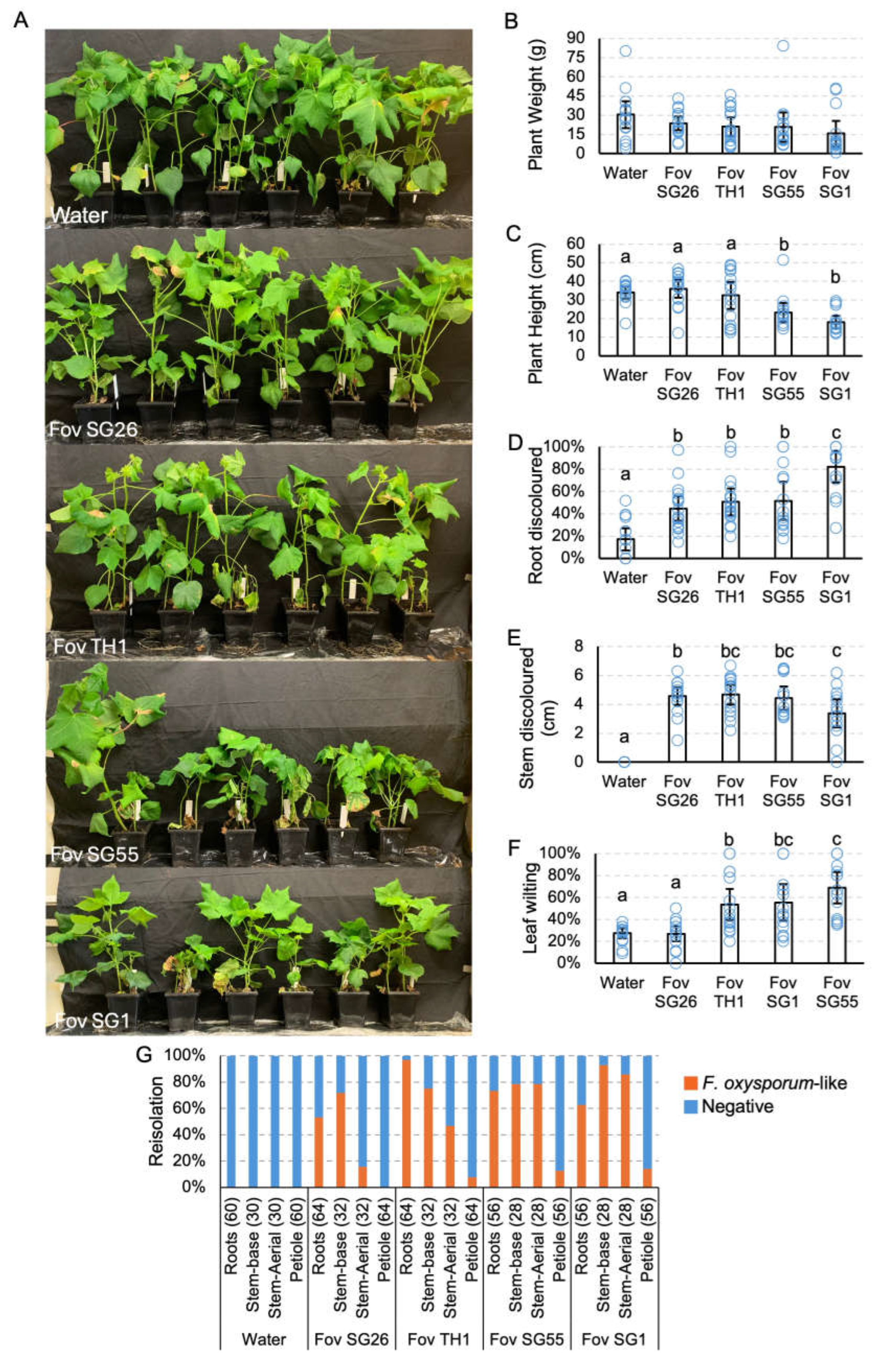
Figure 9.
Pathogenicity testing of Fusarium oxysporum (Fo) isolates on cotton cv. Sicot746 B3F. (A) Plants at harvest (17 — 22 days post inoculation). (B) Total (above and below ground) plant weight. (C) Plant height. (D) Percentage of roots discoloured. (E) Total stem discoloured. (F) Percentage of leaves wilted or dropped. (G) Reisolations of F. oxysporum-like colonies on half-strength potato dextrose agar from the roots, stems and petioles of uninoculated plants and plants inoculated with the Fov isolates. Numbers indicate the number of samples assayed per tissue type. Letters indicate significant a separation of means by Tukey's range test at p = 0.05. Error bars indicate a 95% confidence interval.
Figure 9.
Pathogenicity testing of Fusarium oxysporum (Fo) isolates on cotton cv. Sicot746 B3F. (A) Plants at harvest (17 — 22 days post inoculation). (B) Total (above and below ground) plant weight. (C) Plant height. (D) Percentage of roots discoloured. (E) Total stem discoloured. (F) Percentage of leaves wilted or dropped. (G) Reisolations of F. oxysporum-like colonies on half-strength potato dextrose agar from the roots, stems and petioles of uninoculated plants and plants inoculated with the Fov isolates. Numbers indicate the number of samples assayed per tissue type. Letters indicate significant a separation of means by Tukey's range test at p = 0.05. Error bars indicate a 95% confidence interval.
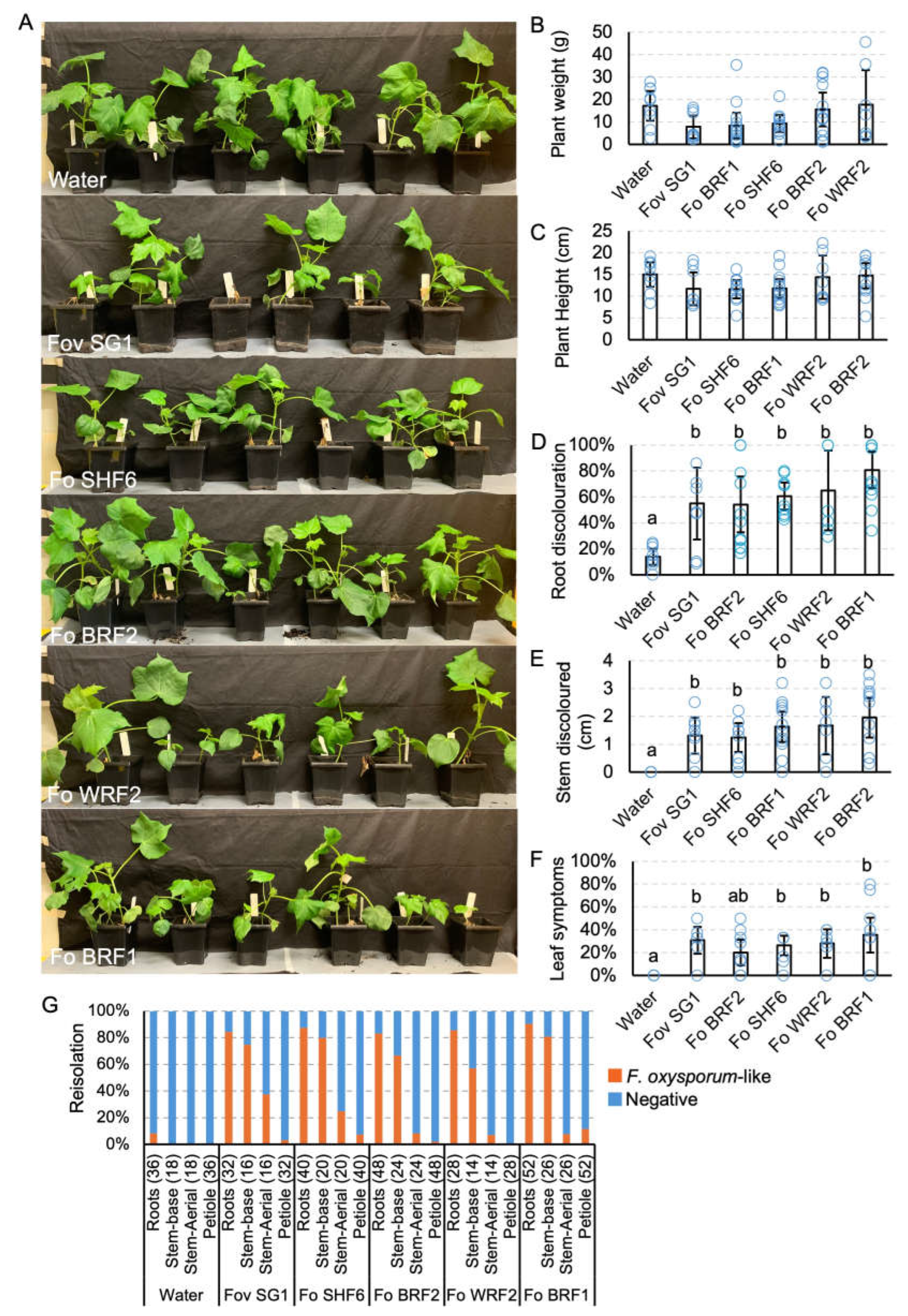

Table 1.
Fungal isolates used in this study indicating their origins and date of collection.
| Isolates1 | Species name | Location | Cotton tissue sampled2 | Collection date (d/m/y) |
|---|---|---|---|---|
| Fov SG1 | Fusarium oxysporum f. sp. vasinfectum | Saint George, Queensland | Stem | 20/04/2022 |
| Fov SG26 | Fusarium oxysporum f. sp. vasinfectum | Saint George, Queensland | Stem | 20/04/2022 |
| Fov SG55 | Fusarium oxysporum f. sp. vasinfectum | Saint George, Queensland | Stem | 21/04/2022 |
| Fov TH1 | Fusarium oxysporum f. sp. vasinfectum | Theodore, Queensland | Stem | 31/12/2022 |
| Fo BRF1 | Fusarium oxysporum | Walgett, New South Wales | Hypocotyls | 21/12/2017 |
| Fo BRF2 | Fusarium oxysporum | Walgett, New South Wales | Hypocotyls | 21/12/2017 |
| Fo SHF6 | Fusarium oxysporum | Wee Waa, New South Wales | Hypocotyls | 21/12/2017 |
| Fo WRF2 | Fusarium oxysporum | Mungindi, New South Wales | Hypocotyls | 21/12/2017 |
| RVB4.1 | Berkeleyomyces rouxiae | Hillston, New South Wales | Roots | 06/12/2017 |
| StrB22 | Berkeleyomyces rouxiae | Mungindi, New South Wales | Roots | 06/12/2017 |
| BRR4 (DAR85827) | Berkeleyomyces rouxiae | Condobolin, New South Wales | Roots | 06/12/2017 |
| 22BRR77 | Berkeleyomyces rouxiae | Wee Waa, New South Wales | Crown | 02/06/2022 |
1 All isolates were archived and maintained locally, except for BRR4 isolate which was deposited in the NSW Plant Pathology and Mycology Herbarium with the accession number DAR85827. 2 Diseased cotton plant parts where the pathogens were recovered.
Table 2.
SIX gene profiles of the isolates used in this study.
| Isolate | SIX1 | SIX2 | SIX3 | SIX4 | SIX5 | SIX6 | SIX6(Fov) | SIX7 | SIX8 | SIX9-1 | SIX10 | SIX11 | SIX12 | SIX13 | SIX14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fov SG11 | — | — | — | — | — | + | + | — | — | — | — | + | — | + | + |
| Fov SG261 | — | — | — | — | — | + | + | — | — | — | — | + | — | + | + |
| Fov SG551 | — | — | — | — | — | + | + | — | — | — | — | + | — | + | + |
| Fov TH11 | — | — | — | — | — | + | + | — | — | — | — | + | — | + | + |
| Fo BRF11 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Fo BRF21 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Fo SHF61 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| Fo WRF21 | — | — | — | — | — | — | — | — | — | — | — | — | — | — | — |
| NRRL254332 | — | — | — | — | — | — | n/t | — | — | — | — | — | — | + | — |
| BRIP636072 | — | — | — | — | — | + | n/t | — | — | — | — | + | — | + | + |
| BRIP433512 | — | — | — | — | — | + | n/t | — | — | — | — | + | — | + | + |
| BRIP253742 | — | — | — | — | — | + | n/t | — | — | — | — | + | — | + | + |
| BRIP433442 | — | — | — | — | — | + | n/t | — | — | — | — | + | — | + | + |
| BRIP433362 | — | — | — | — | — | + | n/t | — | — | — | — | + | — | + | + |
| BRIP433392 | — | — | — | — | — | + | n/t | — | — | — | — | + | — | + | + |
| BRIP433562 | — | — | — | — | — | + | n/t | — | — | — | — | + | — | + | + |
1 SIX gene products for these isolates were amplified using universal primers based on conserved coding regions of the SIX genes [19], except for SIX6 (Fov) which used primers that can specifically detect SIX6 in Australian Fov isolates [20]. 2 The SIX gene profiles of these Fov isolates were obtained from a previous study [19]. — indicates absence of the SIX gene amplicon. + indicates the presence of the SIX gene amplicon. n/t abbreviates 'not tested'.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



