
Submitted:
11 September 2024
Posted:
13 September 2024
You are already at the latest version
Abstract
Almost fifty years after the introduction of antibiotics as an antimicrobial agent, we are facing an increasing number of multi-resistant microbials. The search for new antimicrobial compounds emerged as a promising alternative to treatments for diseases caused by microorganisms such as fungi and bacteria. Thus, endophytic fungi are gaining recognition for their varied metabolites that may possess antimicrobial properties. In our research, we isolated, from Calea pinnatifida, six endophytic fungi identified as Colletotrichum siamense, Colletotrichum karstii, Hypomontagnella barbarensis, Neopestalotiopsis clavispora, Nigrospora sacchari-officinarum and Annulohypoxylon moriforme. Analyses of microbial secondary metabolites by utilizing UHPLC-HR-ESI-MS/MS in conjunction with molecular networking, allowed the annotation of 9 compounds, 5 of them from GNPS spectral library match, and 4 from prediction-based on the molecular network, and allowed the isolation of two pairs of Acyl-tetronic acid derivatives distereoisomeric identified by NMR spectrum. In this sense, the present study underlines the value of endophytic fungi as a reservoir of potentially new antimicrobial agents.
Keywords:
Calea pinnatifida
; Endophytic fungi
; Hypomontagnella barbarensis
; antimicrobial activity
; dereplication
1. Introduction
The search for bioactive compounds as an alternative treatment for diseases caused by microorganisms such as fungi and bacteria has proved to be of great interest to researchers, since some treatments has already demonstrated a certain specific resistance to some common strains, thus triggering a threat to a global scale [1]. Modern medicine, which relies heavily on effective antimicrobial drugs, faces significant challenges in the face of these new demands [2,3]. Some actions can be linked to resistance to existing antimicrobials, such as their excessive and inappropriate use, in human treatment, food production, and the environment [4,5]. In response to this threat, over the last almost one hundred years, research has been dedicated to discovering and applying bioactive compounds from various organisms as a strategy for alternative solutions to the problem [6]. The use of natural products from fungi, including terpenes, peptides, and mainly polyketides has demonstrated properties that can be effective against bacteria and fungi and has emerged as an alternative to treatments [7,8,9,10,11,12,13]. Thus, from Ampelomyces sp., isolated from Urospermum picroides (Asteraceae), are described the compounds 6-O-Methylalaternin and Altersolanol A, which displayed antimicrobial activity against the gram-positive pathogens, Staphylococcus aureus, S. epidermidis and Enterococcus faecalis [14]. Antimicrobial metabolites are also described from extract of the endophyte Nodulisporium sp., isolated from the plant Erica arborea (Ericaceae), including Nodulisporins D–F with antibacterial and fungicidal properties [15]. Asteraceae, a family noticeable for its medicinal uses, comprises approximately 33,000 species grouped into 1,911 genera [16], some of which harbor endophytic fungi [9]. Although there is a great diversity and abundance of Asteraceae, the associated species of fungi are small, and a low percent had their endophytic fungi studied. According to Caruso et al. [9], studies carried out on the endophytes of some Asteraceae species have shown interesting compounds, such as Isofusidiene I, A, B, C and D from endophytes of the genus Chalara associated with Artemisia vulgaris, showing antibacterial and antifungal activity [17] and Phomosin K with antimicrobial activity, from Phomopsis fungi inhabiting the internal tissues of Notobasis syriaca [18]. In this context, Calea pinnatifida, has emerged as a plant with remarkable pharmacological potential, due to its traditional medicinal properties, including use in folk medicine as an antimalarial, analgesic, anti-inflammatory, and for the treatment of gastrointestinal diseases [19]. However, despite their relevance in traditional medicine, the endophytic fungi associated with C. pinnatifida remain largely unknown and little explored. So, this research gap offers a unique opportunity to investigate and identify bioactive compounds with pharmaceutical potential [9,17,20,21]. Therefore, this study investigates endophytic fungi associated with C. pinnatifida to identify new antimicrobial compounds. To achieve that, the analysis of antimicrobial extracts was carried out by UHPLC-HRMSMS combined with molecular networking. This strategy led to selecting one endophytic fungi whose extract allowed the isolation of two pairs of diastereoisomeric Tetronic acid derivatives that includes the novel compound Noduslifuranol.
2. Materials and Methods
2.1. General Experimental Procedures
The 1H and 13C NMR spectra were recorded at 300 and 75 MHz, respectively, on Ultrashield 300 Avance III spectrometer (Bruker−BioSpin, Ettlingen, Germany). It was equipped with a 5 mm trinuclear inverse detection probe with z-gradient (TXI). The temperature was controlled by a BCU I accessory at 25°C. Deuterated methanol (MeOD-d4) (Sigma-Aldrich, St. Louis, USA) was used as the solvent and internal standard. High Performance Liquid Chromatography with Diode Array Detection (HPLC-DAD) analysis was carried out on the Ultimate 3000 liquid chromatograph with UV-DAD detector (Dionex, Sunnyvale, USA), using a Kinetex C18 analytical column (Core-Shell − 4.6 μm − 150.0 mm × 4.6 mm) for method standardization and chemical profile analysis and a Kinetex column C18 semi-preparative column (Core-Shell − 4.6 μm − 150.0 mm × 10.0 mm) for compound isolation. The analyses by UHPLC-HR-MS/MS were performed in a mass spectrometer Q-ToF maxis 3G (Bruker−Daltonics, Billerica, USA), equipped with an electrospray ionization source (ESI) operating with a resolution of 60,000 FWHM in positive mode with detection in a range of masses from m/z 50 to 1500 Da. The capillary voltage was fixed at 4500 V and the nozzle at 500 V. The conditions applied for the drying gas (N2) were 8.0 mL/min, pressure 4.0 Bar, and temperature 200°C for the flow of 0.3 mL/min. UV-Vis detection (PDA) was performed at a rate of 4.6 Hz in a spectral window from 200 to 700 nm [22]. Calibration was set at less than 2 ppm using sodium formiate.
2.2. Plant Material, Isolation, and Cultivation of Endophytic Fungi
Branches and leaves of C. pinnatifida, were collected at the Botanical Garden of São Paulo − SP (coordinates 23 38'29.47" S, 46 37'15.83" W). The specimen (Ferreira, M. J. P. 27) has been deposited in the SPF Herbarium of the Botany Department from the Biosciences Institute of the University of São Paulo.
The methodology used for the isolation of endophytic fungi was based on the procedure described by Santos [23]. Briefly, from the fungi isolated on agar plates, fragments of cultivated mycelium were inoculated in 150 mL Erlenmeyer flask containing 70 mL of potato dextrose broth (PDB) (Becton Dickinson, São Paulo, Brazil), kept under stirring at 28°C for 7 days in an incubator (28°C) in order to obtain a seed culture for the solid medium inoculation. An aliquot of 1 mL of seed culture was inoculated in rice medium, in a 250 mL Erlenmeyer flask containing 50 g of autoclaved rice grains and 90 mL of water. This culture remained in a static medium for 28 days in an incubator at 28°C [24]. After cultivation, the fungi was inactivated and extracted with ethyl acetate (50 mL) under stirring for 24 h, then vacuum filtered (total volume of 150 mL). The filtrate was dried in a rotatory evaporator under low pressure at 40°C and the crude extracts were obtained [25].
The isolated fungi were preserved using the Castellani’s method [26], which consists of storing fungi in sterilized water or saline solution [27] under refrigeration (4oC). This technique is preferably applied to young cultures to reduce the metabolism and consequently the latency of the cells in the face of restricted nutritional sources [28], guaranteeing the preservation of the original characteristics of the culture for long periods and promoting the absence of contamination [29,30].
2.3. Identification of Endophytic Fungi
Total DNA was obtained from a 2-day-old at 37°C monosporic colonies on yeast extract sucrose agar (YES) (10 g yeast extract, 75 g sucrose, 10 g agar, and 500 mL distilled water) by PrepMan Ultra kit (Applied Biosystems, Carlsbad, CA, USA), according to the manufacturer’s recommendations. For accurate species identification was sequenced: i) fragments of internal transcribed spacer (ITS) of ribosomal DNA region using the primer pairs ITS1 and ITS4 and fragments of tubulin gene (TUB2) using the primer primers Bt-2a and Bt-2b. The amplification reactions were performed in a final volume of 25 µL containing 1 µL DNA (40 ng), 12.5 µL 2 PCR Master Mix (Promega Corporation, Madison, WI, USA) containing 3 mM MgCl2, 400 mM each dNTP, and 50 U/mL of Taq Polymerase, 9.5 µL Milli-Q water, and 1 µL of each primer. The amplification protocol included an initial denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing for 30 s (at 52°C and 55°C for ITS and TUB2, respectively), extension final for 10 min at 72°C. Amplicons were purified using Gel and PCR Clean-Up Kit (Promega Corporation, Madison, WI, USA) as manufacturer’s recommendations. All cycling reactions were performed in Eppendorf Mastercycler X50 (Eppendorf Norge AS, Oslo, Norway). The amplified fragments were sequenced in both directions using the BigDye™ Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA). The samples were run on a SeqStudio Genetic Analyzer (Applied Biosystems, Foster City, CA, USA) at the Laboratory of Emerging Fungal Pathogens (Federal University of São Paulo, São Paulo, Brazil) under previously described conditions [31].
The obtained sequences were assembled and edited using the DNA sequence assembly software Sequencher 4.1.4 (Gene Codes Corporation, Ann Arbor, MI, USA). Successful assembly of the contigs required a minimum match percentage of ≥85 and a minimum overlap of 20. The consensus sequences were compared with the sequences deposited in public genomic databases GenBank (http://www.ncbi.nlm.nih.gov/genbank/) and Mycobank (http:// https://www.mycobank.org/Pairwise_alignment) using BLASTn tool. We considered the parameters of E-value <10−5 and identity ≥99% for accurate species identification.
To determine the phylogenetic distance of strains in the same family, phylogenetic trees were elaborated. Thus, ITS sequences from the fungi were aligned with MAFFT [32], and visualized with AliViewer. Poorly aligned regions were removed using trimal v1.2 (parameter - automated1) [33]. A maximum likelihood tree was built with the ITS sequences from the samples (CPFF12, CPFF14, CPFF16, CPFF41, CPFF42, and CPFF52), and ITS sequences from similar related genera. Amanita muscaria ITS sequence was used as an outlier (AB015700.1). The tree was built with IQ-TREE v.2 [34] with model finder (parameters –bb 1000 – bnni -nt AUTO). The resulting trees were visualized using iTOL [35].
For morphological characterization, the isolates were incubated on PDA at 25°C for 7 to 15 days to evaluate their viability and purity. For genus identification, each isolate was cultured on PDA, Oatmeal agar (OA), and Synthetic nutrient agar (SNA) [36] at 25°C for 14 days. The macromorphology features such as colony color, diffused pigment, and texture were observed. The vegetative and reproductive mycelium characteristics, including hyphae, conidia, and conidiogenous cells, were examined. Micromorphological observations were carried out using a light microscope (Leica® DM500) on mounts prepared in lactophenol [37,38].
After cultivation, the fungi were inactivated and extracted with ethyl acetate (50 mL) under stirring for 24 h, then vacuum filtered (total volume of 150 mL). The filtrate was dried in a rotatory evaporator under low pressure at 40°C and the crude extracts were obtained [25].
2.4. Data Acquisition by UHPLC-HRMS/MS and Molecular Networking
Each extract was solubilized in methanol (1 mg/mL), filtered in a 0.45 µm syringe filter (Mili-pore PTFE, 13 mm, 22 µm – Allcrom, São Paulo, Brazil) and analyzed by UHPLC-HRMS/MS. Analysis conditions are in mobile phase gradient mode (A) H2O + 0.01% formic acid and (B) methanol, flow rate 300 μL/min: 0-3 min: 2% B; 3-18 min from 2% to 100%; 18-21 min: 100% B constant; 21-25 min returns to the initial condition of 2% B; 25-28 min. with 2% B for column reconditioning, using a reverse phase column (Kinetex EVO C18, 100 mm x 2.1 mm, 2.6 μm, 100 Å) at a constant temperature of 50°C.
The files generated in the UHPLC-HRMS/MS analyses were converted from the original “.d” format to the open ".mzXML" extension for the construction of molecular networking. The newly converted files were checked by SeeMS software (Proteowizard®). The ".mzXML" files were uploaded to the Global Natural Products Social Molecular Networking (GNPS) platform server using WinSCP software [39]. A molecular network was created using the online workflow on the GNPS platform (http://gnps.ucsd.edu) based on MSCluster algorithm [40]. Data were filtered by removing all ions from MS/MS fragments within +/- 17 Da of the m/z precursor. MS/MS spectra acquired on a quadrupole hyphenated time-of-flight (QTof) type mass analyzer were filtered by picking only the top 6 fragment ions in the +/- 50Da window across the spectrum. The precursor ion mass tolerance was set to 0.02 Da and the MS/MS fragment ion tolerance to 0.02 Da. A network was created where edges were filtered to have a cosine score above 0.7 and more than 6 corresponding peaks. In addition, edges between two nodes were kept in the network if and only if each of the nodes appeared in the top 5 most similar nodes to each other. Finally, the maximum size of a molecular family was set to 100, and the lowest-scoring edges were removed from molecular families until the molecular family size was below this threshold. The network spectra were then searched against the GNPS spectral libraries and for dereplication, the library spectra were filtered in the same way as the input data. All matches maintained between the network spectra and library spectra were required to have a score above 0.7 and at least 6 matched peaks [39]. Clusters detected in blank were removed from the networking, which includes spectral data from solvents used in the extraction and chromatographic procedures (dried hexane, chloroform, ethyl acetate, methanol). The molecular networking view and edition were performed in Cytoscape version 3.10.2 [41].
2.5. Chromatographic Fractionation and ISOLATION of compounds
The extract obtained from Hypomontagnella barbarensis - CPFF41 was selected from profile in molecular networking for further purification. So, part of the extract (150 mg) was subjected to HPLC-DAD in semi-preparative RP-18 in H2O with 0.01% formic acid (A) and methanol (MeOH) (B) in gradient mode: 0 min.: 40% B; 7 min.: 15% B; 9 min.: 85% B and gradually returns to the initial condition up to 10 min and oven temperature at 45°C (flow rates 1.8 mL/min, UV 255 nm) to afford compound 1 (25 mg) and compound 2 (8 mg). These compounds were then analyzed by NMR and HRMS-MS for structural identification.
2.6. Antimicrobial Activity Assay - Strains and Growth of Microorganisms in Solid Culture
The antimicrobial activity assays of the extracts were evaluated against both yeast strains (Table 1) and bacterial strains (Table 2).
The cultures were performed from the inoculum of the colonies isolated from the strains of interest in standard medium for YPD yeasts (1% yeast extract, 2% peptone, 2% dextrose and 2% agar) and Luria−Bertani medium (Bio basic inc. S516), containing 25 mL of culture medium in each Petri dish. Temperature and growth time ranged from 24 to 48 h and from 30 to 37 ° C, depending on experimental planning. Eugenol (Sigma Aldrich, ST Louis, USA) was used as positive control for yeasts and kanamycin 50 μg/mL and the same concentration for ampicillin (AppliChem) as positive control for bacteria.
2.7. Disc Diffusion Method
The disc diffusion method to assess the antimicrobial susceptibility of the microorganisms was performed following the CLSI protocol (M2-A8) [42]. The microorganisms were cultured in LB (bacteria) or YPD (yeast) 24 to 48 h before. The inoculum was made by picking 3 colonies and transferred to a test tube containing 5 mL of 0.9% saline solution. The suspension was adjusted cell density similar to the Mcfarland scale of 0.5 (corresponds to 1.5x108 cells per mL). Subsequently, with a sterile swab, the solution was spread evenly under the plate containing Mueller Hinton for bacteria or Mueller Hinton (BD 212322) supplemented with 2% D-glucose + 0.5 μL methylene blue for yeasts. Then, sterile filter paper discs of 5 mm diameter that received 5 to 10 μL of the compounds of interest with a final concentration ranging from 200 μg to 400 μg, a positive control was also placed (disc containing 200 μg/disc of eugenol for yeasts or kanamycin 50 μg/disc for bacteria) and a negative control (disc containing water). The plates were incubated in a bacteriological oven at 30°C for 24 to 48 h depending on the microorganism cultivated, after which the halos were measured and the data duly recorded. The results are presented as mean of three biological replicates and statical analysis (mean comparation) were performed using the PrismGraph version 8.0.
2.8. Minimum Inhibitory Concentration (MIC)
Broth microdilution assays were performed in a 96-well plate for bacteria using the EUCAST (European Committee on Antimicrobial Susceptibility Testing) method [43]. Briefly, the microorganisms were grown on a solid LB medium and incubated for 24 h at 37°C. Three fresh colonies were used and transferred to 5 mL of saline solution (0.9% NaCl), this cell suspension was adjusted to a cell density of approximately 1-2 x108 x CFU/mL (0.5 McFarland scale). The extracts were diluted in water and adjusted to an initial concentration of 0.0007 μg/μL, then a serial dilution was made, and the extracts were tested, the growth control contained only 100 μL of control contained only 100μL of the liquid culture, and the negative control was made with 100 μL of the sterile RPMI medium the 96−well plates were incubated at 30°C for 24 h, after this period the optical density (0.0 μL) was measured. During this period, the optical density (0.23-0.27) was determined in a microplate reader (Epoche 2 from Biotek). After reading the OD530, 50 μL of a 0.005% resazurin solution was added to check the microorganisms’ viability, and the plate was again incubated on a shaker at a temperature of 30°C for 2 h. Experiments were conducted with two technical replicates and in biological triplicates.
3. Results
3.1. Endophytic Fungi Obtained from Leaves
Seven endophytic fungi (designated as CPFF12, CPFF14, CPFF16, CPFF41, CPFF42, CPFF52 and CPFC2) were isolated from Calea pinnatifida leaves (Figure S1- Supplementary Material). Five different genera were identified through macro and microscopic visualization of conidiophores (reproductive structures), hyphae, and growth aspects (Figure S2 A-E - Supplementary Material). Furthermore, molecular analyses using sequences obtained from ITS region and fragment of the TUB2 gene allowed the identification of the genus Colletotrichum for three isolates, CPFF16 being Colletotrichum siamense (Figure S2C -- Supplementary Material) and CPFF12 / CPFF14 as Colletotrichum karstii (Figure S2A and S2B - Supplementary Material), having been confirmed as identical strains only after molecular analysis. Another species found for CPFF41 was Hypomontagnella barbarensis (Figure S2D - Supplementary Material), segregated from the Hypoxylon group by a polyphasic taxonomic approach [44], CPFF42 was identified as Neopestalotiopsis clavispora (Figure S2E - Supplementary Material), a species inserted in the recently known genus within the Pestalotiopsis group. CPFF52 was found to be Nigrospora sacchari-officinarum (Figure S2F - Supplementary Material), and finally, CPFC2 was identified as Annulohypoxylon moriforme (Figure S2G - Supplementary Material). The database used to identify endophytic fungi was the NCBI platform, using the BLAST tool, and the sequences obtained were deposited in GenBank (Table 3). The blast of the nucleotide sequence of endophytic fungi are in Table S1 - Supplementary Material.
3.2. Dereplication Based on Molecular Networking Organization from HRMS-MS Data
The crude EtOAc extracts obtained from the seven endophytic fungi isolated from C. pinnatifida were analyzed by UHPLC-HRMS/MS and, subsequently, dereplicated using the GNPS platform, to generate molecular networking and to make annotations of secondary metabolites (Figure S3A and B - Supplementary Material). The dereplication process showed, for the analyzed extracts, a total of 5 library matches in positive ionization mode and annotations made by GNPS suggest 3 compounds from terpenoid class, Fusaproliferin (cembrane diterpene), Alismol (guaiane sesquiterpene) annotated for the C. karstii and N. sacchari-officinarum and 1-Acetoxy-7-isopropylidene-1,4a-dimethyl-6-oxodecahydro-2-naphthalenyl-2,3-dimethyl-2-oxiranecarboxylate (eudesmane sesquiterpene), annotated for the Colletotrichum genus (Table 4).
Furthermore, considering that no known compounds were annotated by GNPS from the extract from Hypomontagnella barbarensis (CPFF41), besides fatty acid derivative (Table 4), the molecular networking of this extract was analyzed in detail. Thus, molecular networking for this extract showed a cluster in which the major compound at m/z 309.134 was observed, in agreement with the node size in the molecular network, indicating a more significant number of detected spectra (Figure 1). This compound (1) was then isolated in a more substantial proportion from the H. barbarensis extract and identified as E/Z Nodulisporacid A (see item 3.3) [45]. In addition to that, other nodes referring to analogous compounds were observed in the cluster, which includes the methyl ester of Nodulisporacid A at m/z 323.147 ([M+H]+) (compound 3), already described in the literature [45]; a derivative of Nodulisporacid A with one less methyl at m/z 295.116 ([M+H]+), reported here for the first time, for which the name nor-Nodulisporacid A is proposed (compound 4); and the compound at m/z 291.123 ([M+H]+) whose proposed structure includes an additional furane ring, Nodulisfuranol. The additional furan ring is proposed based on a biogenesis hypothesis involving the nucleophilic attachment of a hydroxyl group at C-3 (formed from the enolic tautomer of Tetronic acid A) to the C-6 position of the carboxylic acid, followed by dehydration. This last derivative, Nodulisfuranol - compound 2, was isolated from the extract, and its structure was determined by NMR (see section 3.3) and HR-ESI-MS-MS² (Figure 1 and Figure 3).
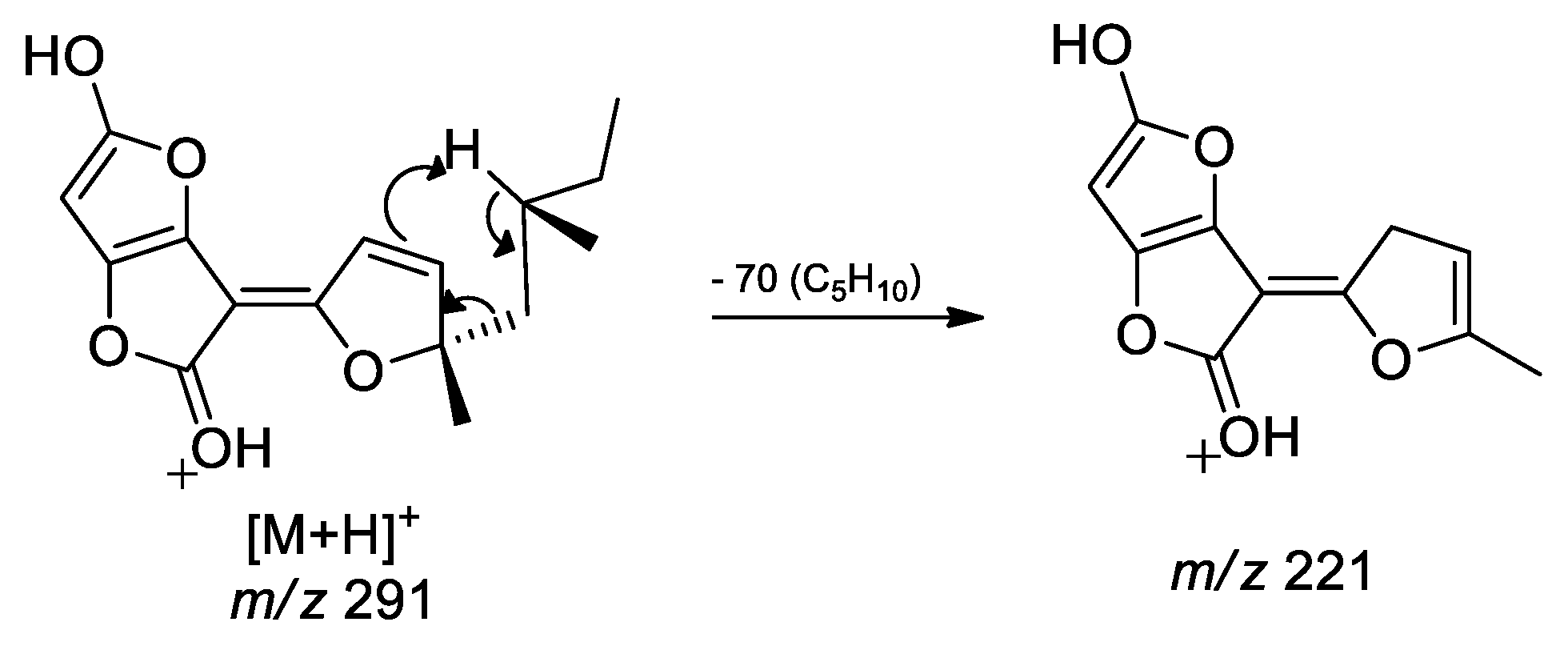
The structures 1 to 4 share similar MS² spectra, as observed in the molecular networking (Figure 1). Based on this observation, fragmentation proposals were put forth to corroborate and complement their structural similarity and annotation, considering characteristic and complementary fragmentation mechanisms. In the fragmentation spectra of structures 1, 3, and 4, dehydration of the carboxylic acid (1 and 4) or elimination of MeOH from the methyl ester (3) was observed, resulting in the loss of 18 Da (H2O) or 32 Da (MeOH), respectively, corresponding to mechanism (a) in the Figure 2. Another pathway, which combines mechanisms (a) and (b), leads to concurrent loss of H₂O or MeOH along with the McLafferty rearrangement (loss of 70 Da, C₅H₁₀) in structures of 1, 3 and 4. This rearrangement occurs due to the deprotonation of the methyne carbon C-6’ in the side chain by the nucleophilic attach of π electrons of the dihydrofuran ring, resulting in an expected loss of 70 Da in structures 1, 3, and 4. Additionally, based on the proposed alkene loss mechanism (70 Da), it was suggested that structure 4 lacks a methyl group at position C-10´ to maintain similarity with structures 1 and 3. Consequently, structure 4 has no a stereogenic center at position C-4´. Furthermore, via pathway (a), can occur a loss of C₂H₂O (42 Da), leading to fragments with m/z ratios of 249 (compounds 1 and 3) and m/z 235 (compound 4). Subsequently, the intermediate ion can lose 28 Da in the form of CO, supporting an alternative pathway to form ions with ratios of m/z 221 (compounds 1 and 3) and m/z 207 (compound 4) (Figure 2). Finally, the compound 2 exhibits a base peak with an m/z ratio of 221, corresponding to the previously proposed McLafferty rearrangement mechanism for structures 1, 3, and 4. The additional furane ring also corroborate to the less fragmentation pattern in comparison to the structures 1, 3 and 4 (Figure 1 and 3).
Figure 1.
Consensus MS² mass spectra and Molecular network created by GNPS. Colors of the nodes: Green: CPFF41; Orange: CPFF42; Yellow: CPFF52. Edge labels between nodes represent m/z differences. Node labels represent parent masses (m/z). Edge labels between nodes represent cosine scores. The size of the nodes is proportional to the spectral detection rate.
Figure 1.
Consensus MS² mass spectra and Molecular network created by GNPS. Colors of the nodes: Green: CPFF41; Orange: CPFF42; Yellow: CPFF52. Edge labels between nodes represent m/z differences. Node labels represent parent masses (m/z). Edge labels between nodes represent cosine scores. The size of the nodes is proportional to the spectral detection rate.
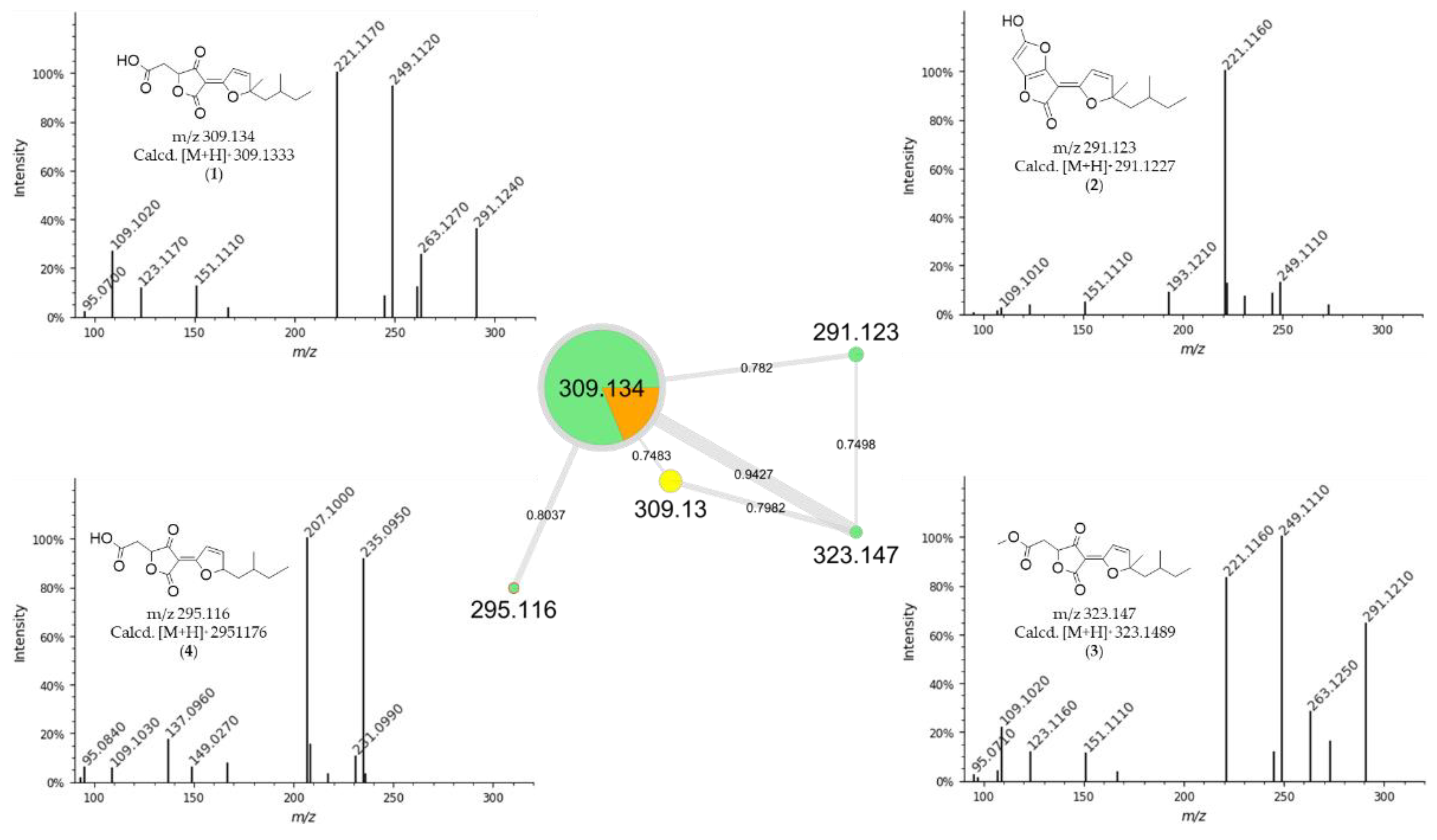
Figure 2.
Fragmentation proposes for compounds 1 (m/z 309), 3 (m/z 323) and 4 (m/z 295) in ESI-(+)-QTof.
Figure 2.
Fragmentation proposes for compounds 1 (m/z 309), 3 (m/z 323) and 4 (m/z 295) in ESI-(+)-QTof.
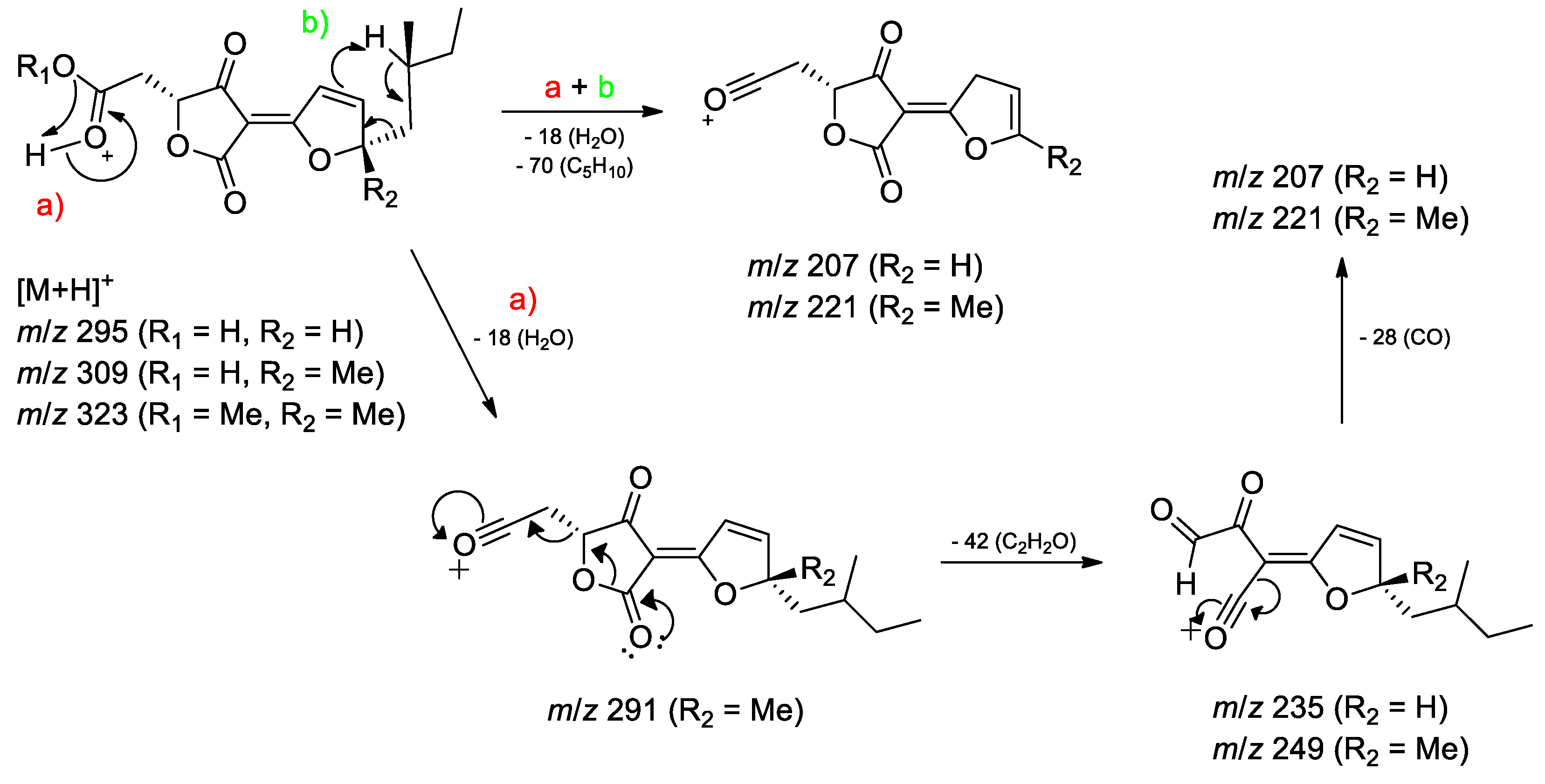
Figure 3.
Fragmentation proposes for compound 2 (m/z 291) in ESI-(+)-QTof.
3.3. Identification of Compounds from Fungi Hypomontagnella barbarensis
Considering a molecular family of Tetronic acid derivatives and the annotations corresponding to Nodulisporacid A and three other derivatives (Figure 1), associated with antimicrobial activity evaluation with lower values of MIC against S. aureus, the extract from H. barbarensis was then selected for chromatographic fractionation. Thus, purification by HPLC allowed the isolation of compound 1 as a pair of isomers (E and Z). The compound was subjected to 1H and 13C NMR and UHPLC-HR-MS/MS analyses for structural elucidation. The 13C NMR data indicated the presence of 32 carbon signals, including six methyl groups, six methylenes, four methines, and twelve fully substituted carbons. The carbon resonance at δ 170.7/172.6 was attributed to the carboxylic function. Signals were also assigned to an ester (δ 163.9/164.0) and an α,β-unsaturated ketone (δ 196.9/198.3). Other signals included a 1,2-disubstituted olefin unit (δ 123.0/123.1 and 164.0/164.1) and fully substituted carbons (δ 180.9/181.7, 103.5/104.1, and 94.5/94.9). Thus, the framework of this compound contains a furandione and a furylidene fragments connected by a double bond characteristic of Tetronic acid derivatives. Additionally, both E and Z isomers were observed for the double bond, and the E- and Z-isomers were interconverted spontaneously and reached an equilibrium state at a ca. ratio of 1:1. Therefore, the data were consistent with the structures diastereoisomers Z and E of Nodulisporacid A (Figure 4) and confirmed by comparison of NMR data with those reported in the literature [46]. For compound 2, the 13C NMR showed the same skeleton containing sixteen carbon signals duplicated, also revealing spontaneous inter-conversion between E- and Z-isomers, with the ratio of 1:1. The difference to compound 1 is the absence of a ketone carbonyl signal, indicating that the tetronic ring could be in the enolic form, which may have favored the attack of the enol hydroxyl on the carboxyl leading to the formation of a new furan ring after dehydration in this second ring, evidenced by signals at δ 5.67/5.68 (s, H-5) referring to the two E/Z stereoisomers in 1H-NMR. These signals, in the HSQC spectrum presents a cross peak with shifts at 98.3/98.4 (C-5). Furthermore, it is possible to observe the cross peaks in the HMBC spectrum between H-5 with signals at δ 153.2/153.6 (C-3), in addition to the signal at δ 182.3/182.5 (C-4). The structure with the additional furan ring was also suggested based on the analysis of the mass spectrum that showed [M+H]+ at m/z 291.1232 corresponding to the molecular formula of C16H18O5 indicating the presence of 8 unsaturations. Thus, the proposed structure is a derivative of compound 1 and, being the first time reported in the literature, the name Nodulisfuranol is proposed (Figure 4).
Compound 1: Nodulisporacid A (E:Z mixture); yellow amorphous solid; Z-isomer: 1H NMR (MeOH-d4, 300 MHz) δ 4.84 (m, H-4), 2.87 (m, H-5a), 2.73 (m, H-5b), 7.32 (d, J = 5.7 Hz, H-2'), 7.81 (d, J = 5.7 Hz, H-3'), 2.06 (m, H-5' a), 1.65 (m, H-5' b), 1.25 (m, H-6'), 1.25 (m, H-7'a), 1.14 (m, H-7'b), 0.78 (t, J = 7.0 Hz, H-8'); 0.85 (d, J = 6.1 Hz, H-9'); 1.54 (s, H-10'). 13C NMR (MeOH-d4, 75 MHz) δ 172.5 (C, C-1), 94.9 (C, C-2), 198.3 (C, C-3), 80.7 (CH, C-4), 36.8 (CH2, C-5), 173.1 (C, C-6), 181.6 (C, C-1’), 123.1 (CH, C-2'), 164.1 (CH, C-3'), 103.4 (C, C-4'), 45.2 (CH2, C-5'), 31.3 (CH3, C-6'), 31.5 (CH2, C-7'), 11.4 (CH3, C-8'); 21.3 (CH3, C-9'), 23.8 CH3, C-10'). E-isomer: 1H NMR (MeOH-d4, 300 MHz) δ 4.87 (m, H-4), 2.93 (m, H-5a), 2.79 (m, H-5b), 7.47 (d, J = 5.8 Hz, H-2'), 7.83 (d, J = 5.7 Hz, H-3'), 2.12 (m, H-5'a), 1.72 (m, H-5'b), 1.25 (m, H-6'), 1.25 (m, H-7'a), 1.14 (m, H-7'b), 0.77 (t, J = 7.0 Hz, H-8'); 0.80 (d, J = 6.1 Hz, H-9'); 1.54 (s, H-10'). 13C NMR (MeOH-d4, 75 MHz) δ 170.7 (C, C-1), 94.5 (C, C-2), 196.8 (C, C-3), 80.3 (CH, C-4), 36.7 (CH2, C-5), 172.9 (C, C-6), 180.9 (C, C-1'), 123.0 (CH, C-2'), 163.9 (CH, C-3'), 103.4 (C, C-4'), 45.1 (CH2, C-5'), 31.3 (CH, C-6'), 31.4 (CH2, C-7'), 11.4 (CH3, C8'), 21.2 (CH3, C-9'), 23.7 (CH3, C-10').
Compound 2: Nodulisfuranol (E:Z mixture); yellow amorphous solid; Z-isomer: 1H NMR (MeOH-d4, 300 MHz) δ 5.67 (s, H-5), 7.25 (d, J = 5.8 Hz, H-2'), 7.86 (d, J = 5.8 Hz, H-3'), 2.01 (m, H-5'a), 1.60 (m, H-5'b), 1.15-1.19 (m, H-6'), 1.15-1.19 (m, H-7'a), 1.04 (m, H-7'b), 0.74 (t, J = 7.0 Hz, H-8'), 0.78 (d, J = 6.0 Hz, H-9'), 1.47 (s, H-10'), 7.87 (s, OH). 13C NMR (MeOH-d4, 75 MHz) δ 166.1 (C, C-1), 92.5 (C, C-2), 153.2 (C, C-3), 182.3 (C, C-4), 98.3 (C, C-5), 165.6 (C, C-6), 182.4 (C, C-1'), 123.1 (CH, C-2'), 163.6 (CH, C-3'), 104.7 (C, C-4'), 44.9 (CH2, C-5'), 31.3 (CH, C-6'), 31.5 (CH2, C-7'), 11.4 (CH3, C-8'), 21.1 (CH3, C-9'), 23.4 (CH3, C-10'). E-isomer: 1H NMR (MeOH-d4, 300 MHz) δ 5.68 (s, H-5), 7.40 (d, J = 5.7 Hz, H-2'), 7.89 (d, J = 5.7 Hz, H-3'), 2.03 (m, H-5'a), 1.68 (m, H-5'b), 1.15-1.19 (m, H-6'), 1.15-1.19 (m, H-7'a), 1.04 (m, H-7'b), 0.74 (t, J = 7.0 Hz, H-8'), 0.79 (d, J = 6.0 Hz, H-9'), 1.50 (s, H-10'), 7.90 (s, OH). 13C NMR (MeOH-d4, 75 MHz) δ 167.9 (C, C-1), 93.0 (C, C-2), 153.6 (C, C-3), 182.5 (CH, C-4), 98.4 (C, C-5), 165.9 (C, C-6),182.5 (C, C-1’), 123.2 (CH, C-2'), 163.9 (CH, C-3'), 105.4 (C, C-4'), 45.0 (CH2, C-5'), 31.4 (CH3, C-6'), 31.5 (CH2, C-7'), 11.4 (CH3, C-8'), 21.2 (CH3, C-9'), 23.5 (CH3, C-10').
3.4. Antimicrobial Activity
Initially, disk diffusion assays were conducted with the yeasts and bacterial strains listed in Table 1 and 2 in order to screen fungal extracts for antimicrobial activity. The CPFF14 and CPFF41 extracts inhibited the S. aureus strain and CPFF12 extract proved promising against two yeasts strains, S. cerevisiae and C. albicans in disk diffusion assay. The other extracts evaluated did not show significant inhibition values (Figure 5 and Table 5).
In sequence, the species listed in Table 1 and 2 were tested to determine the minimum inhibitory concentration (MIC). The results showed that yeasts were more sensitive to the extract CPFF12 (C. karstii), while the bacteria S. aureus was more sensitive to the extracts CPFF14 and CPFF41. The H. barbarensis (CPFF41) extract showed 90% growth inhibition at 0.05 mg/mL (Table 6). Other extracts also had no significant activity against microbial strains.
4. Discussion
Asteraceae family, to which C. pinnatifida belongs, has common or cosmopolitan endophytic species [47]. The genera found, Colletotrichum, Nigrospora, and Pestalotiopsis, according to Caruso [9] and Nicoletti [48], were isolated from most plant species in this family, while the genus Xylaria, was observed in a lower percentage. Also, these fungi were described as phytopathogens, a diverse group of endophytic fungi that can affect a wide range of economically important crops, including fruit, vegetables, ornamental plants, and grasses, and some species in this group have also become the object of study for biotechnological applications, seeking new bioactive compounds that can be used in new environments [49].
From C. pinnatifida were isolated 3 species of endophytic fungi from the Colletotrichum genus including C. siamense and C. karstii, besides species of Hypomontagnella barbarensis, Neopestalotiopsis clavispora, Nigrospora sacchari-officinarum and Annulohypoxylon moriforme. These genera of endophytes have already been described as endophytes of other species and produce metabolites of interest, such as Colletotrichum gloesporioides, which has stood out in studies in terms of the production of toxic metabolites and promising biological activity [50]. Wang [51] reported that Pestalachlorides E and F, isolated from fungi of the genus Pestalotiopsis, showed potent agronomic activity, which was also reported by Yang [52], regarding the isolation of the anticancer Taxol present in the species Pestalotiopsis microspora. Other compounds with interesting activities come from the Nigrospora genus, ranging from herbicides to insecticides and antimicrobials [53,54,55,56]. These secondary metabolites have demonstrated a wide range of biological activities, such as antifungal, antibacterial, antiviral, antitumor, and cytotoxic properties [57,58,59]. Hypoxylaceae family was recently elevated from Xylariaceae, representing one of the predominant groups of fungal endophytes. Furthermore, from this family, it was identified the species Hypomontagnella barbarensis, a new genus of the Hypoxylaceae family, segregated from Hypoxylon by a polyphasic taxonomic approach [44]. Regarding to chemical composition of extracts, some metabolites, already described in the literature, isolated from other organisms were annotated by the platform, such as Fusaproliferin, a compound with anti-inflammatory action, isolated from Fusarium proliferatum [60] (Figure S3A Supplementary Material and Table 4). Most of these bioactive compounds are derived from the secondary metabolism of these organisms, through which they generate compounds belonging to the most diverse chemical classes, such as terpenes, alkaloids, polyketides, and peptides, among others [61]. The molecular networking analysis also allowed the dereplication of a molecular family of Tetronic acid derivatives. It made possible the annotation of derivatives from H. barbariensis, which drives chromatographic fractionation. Furthermore, compound 1, isolated from H. barbariensis, identified as two isomers (E and Z) of Nodulisporacid A, has been characterized by Kasettrathat et al [62], and it has already been reported by Sumiya et al. [63] being isolated from the marine fungi Nodulisporium sp. and by Huang et al., [45] isolated from an endophytic fungus Hypomontagnella monticulosa, belonging to the same genus as the fungus found. This compound has been tested for its bioactivity and cytotoxicity, showing moderate activity against cancer cell lines developed by Kasettrathat et al. [63] and moderate antimalarial activity by Huang et al [45]. However, compound 2, which was isolated from the same extract, was identified as a derivative of Nodulisporacid A named Nodulisfuranol, and has not yet been described in the literature, suggesting that it is a new substance, as well as compound 4 (nor-Nodulisporacid A) of m/z 295 annotated in the Nodulisporacid A cluster.
The antimicrobial activity of the endophytic fungal extracts was evaluated by disk diffusion and MIC assays against 8 bacterial strains including S. aureus, and 10 yeast strains. However, only the extracts of C. Karstii and H. barbarensis were active against, at least, one of the strains, with the extract of H. barbarensis being active against S. aureus. It was also demonstrated that this extract is composed of Noduliporacid A and the triple-bonded derivative called Nodulisfuranol. Considering that Nodulisporacid A has already been evaluated against S. aureus in the work of Huang et al [45], and that it showed no activity, it can be suggested that the activity observed for the CPFF41 extract may be due to the presence of the derivative, or even due to the synergistic action between the two compounds.
Furthermore, combining the data from the GNPS annotations and the microbiological assays, we can suggest that the presence of the annotated compounds, such as Fusaproliferin and alismol, is related to the antimicrobial activity of the extracts. For the CPFF12 extract, was observed the annotation of these two compounds, already described as active and with cytotoxic/antimicrobial action [64,65]. They may justify the inhibition, especially of C. albicans, with a low extract concentration. This makes it clear that synergy between compounds can occur, and thus, the activity can be associated with more than one compound.
Due to these promising activities, knowing the metabolic profile of these individuals was of great interest. The presence of compounds already described as active, and the possibility of isolating new ones leaves open the opportunity for new discoveries. Additionally, considering the large number of secondary metabolites that fungi might contain, dereplication tools such as LC-MSMS and molecular networking, have become important in prospecting metabolites, saving time and work on isolation and characterization of natural products. However, whereas genome analyses have revealed a great diversity of biosynthetic gene clusters (BGCs) that far exceeds the number of known substances, most of its secondary metabolites remain still unknown [66]. Thus, dereplication of the extracts from fungi, with the aid of GNPS to annotate metabolites, can be complemented with genomic analysis integrated with metabolomics, to identify gene clusters related to the biosynthesis of the annotated molecules, and thus can allow access full biosynthetic potential of the fungi.
5. Conclusions
Investigating endophytic fungi offers a promising avenue for discovering new bioactive compounds with potential pharmaceutical uses. Through the methods outlined in this study, we isolated 6 endophytic fungi from C. pinnatifida leaves, belonging to 4 distinct genera: Colletotrichum, Neopestalotiopsis, Nigrospora, Hypomontagnella and Annulohypoxylon. Although these genera are recognized for their phytopathogenic roles, they have also demonstrated biotechnological promise in producing bioactive metabolites. Additionally, our findings regarding chemical composition of extracts from fungi indicate that the molecular network approach based on GNPS showed to be a rapid and efficient tool to organize into clusters and annotate bioactive metabolites from a crude extract, not only based on the annotation of known molecules, but also for new ones, as Acyl tetronic acid derivatives suggested for the first time in the present study. Moreover, it was isolated two Tetronic acid derivatives identified by NMR, from the most active extract against S. aureus. Thus, this approach can be used for drug discovery. Remarkably, the array of bioactive compounds synthesized by these endophytic fungi underscores their potential as valuable reservoirs for new pharmaceuticals, agrochemicals, and industrial compounds. Overall, this study underscores the significance of investigating endophytic fungi associated with medicinal plants like C. pinnatifida. It provides a fertile ground for discovering new bioactive compounds with a wide range of pharmacological properties.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Blast of the nucleotide sequence of endophytic fungi from Calea pinnatifida. Figure S2: Micromorphology slides of endophytic fungi. Figure S3: Molecular networking of the crude extracts of endophytic fungi isolated from Calea pinnatifida. Figure S4: NMR spectra of the isolated compounds.
Author Contributions
Conceptualization, B.B., S.P.V. and P.S.; methodology, B.B., L.A.C., J.M., L.F.L., M.J.P.F., R.C.P., M.A.V., S.S.G., G.Q.S., A.M.R. and J.A.C.; software, B.B., L.A.C. and J.A.C.; formal analysis, B.B., L.A.C., A.L.S., P.S. and S.P.V.; resources, P.S., S.P.V., M.J.P.F.; data curation, P.S. and S.P.V.; preparation of original draft, P.S., L.A.C., B.B.; review and editing, B.B., P.S. and L.A.C..; supervision, P.S. and S.P.V.; project administration, P.S.; acquisition of funding, P.S. and S.P.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo - FAPESP, grant number 2022/09202-4 and 2020/16554-9. Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - CAPES fellowships to BB and JM (Finance Code 001, Brazil) and Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq (scientific research awards of PS and MJPF). SSG is a FAPES Capixaba Research Fellow (FAPES 692/2024). This study is an activity within the CEPID ARIES – Antimicrobial Resistance Institute of São Paulo (FAPESP 2021/10599-3) (https://site.unifesp.br/cepidaries/). Finnally, this research was registered in the Sistema Nacional de Gestão do Patrimônio Genético e do Conhecimento Tradicional Associado [SisGen # A90708B].
Data Availability Statement
All Mass Spectrometry data used in this work is available at MassIVE repository MSV000095564 according to the DOI-code 10.25345/C5BC3T821. The repository is accessible online (ftp://massive.ucsd.edu/v08/MSV000095564/ ).
Conflicts of Interest
The authors declare that they have no known competing financial interest or personal relationships that could have appeared to influence the work reported in this paper.
References
- Alkatheri AH, Yap PSX, Abushelaibi A, et al (2023) Microbial Genomics: Innovative Targets and Mechanisms. Antibiotics 12:. [CrossRef]
- World Health Organization (2024) Ten threats to global health in 2019. In: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019.
- Murray CJ, Ikuta KS, Sharara F, et al (2022) Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. The Lancet 399:629–655. [CrossRef]
- Lewis K (2013) Platforms for antibiotic discovery. Nat Rev Drug Discov 12:371–387. [CrossRef]
- World Health Organization (2024) Antimicrobial resistance. In: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance.
- Loureiro RJ, Roque F, Teixeira Rodrigues A, et al (2016) Use of antibiotics and bacterial resistances: Brief notes on its evolution. Revista Portuguesa de Saude Publica 34:77–84. [CrossRef]
- Bessada SMF, Barreira JCM, Oliveira MBPP (2015) Asteraceae species with most prominent bioactivity and their potential applications: A review. Ind Crops Prod 76:604–615. [CrossRef]
- Caldas LA, Rodrigues MT, Batista ANL, et al (2020) Sesquiterpene lactones from Calea pinnatifida: Absolute configuration and structural requirements for antitumor activity. Molecules 25:1–11. [CrossRef]
- Caruso G, Abdelhamid MT, Kalisz A, Sekara A (2020) Linking endophytic fungi to medicinal plants therapeutic activity. A case study on Asteraceae. Agriculture (Switzerland) 10:1–23. [CrossRef]
- Deshmukh SK, Gupta MK, Prakash V, Saxena S (2018) Endophytic fungi: A source of potential antifungal compounds. Journal of Fungi 4:. [CrossRef]
- Golinska P, Wypij M, Agarkar G, et al (2015) Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. International Journal of General and Molecular Microbiology 108:267–289. [CrossRef]
- Kharwar RN, Mishra A, Gond SK, et al (2011) Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat Prod Rep 28:1208–1228. [CrossRef]
- Mousa WK, Raizada MN (2013) The Diversity of Anti-Microbial Secondary Metabolites Produced by Fungal Endophytes: An Interdisciplinary Perspective. Front Microbiol 4:. [CrossRef]
- Aly AH, Edrada-Ebel RA, Indriani ID, et al (2008) Cytotoxic metabolites from the fungal endophyte Alternaria sp. and their subsequent detection in its host plant Polygonum senegalense. J Nat Prod 71:972–980. [CrossRef]
- Dai J, Krohn K, Draeger S, Schulz B (2009) New naphthalene-chroman coupling products from the endophytic fungus, Nodulisporium sp. From erica arboreal. European J Org Chem 1564–1569. [CrossRef]
- WFO Plant list (2024) Familia Asteraceae. In: https://wfoplantlist.org/taxon/wfo-7000000146-2023-12?page=1.
- Lösgen S, Magull J, Schulz B, et al (2008) Isofusidienols: Novel chromone-3-oxepines produced by the endophytic fungus Chalara sp. European J Org Chem 698–703. [CrossRef]
- Hussain H, Tchimene MK, Ahmed I, et al (2011) Antimicrobial chemical constituents from the endophytic fungus Phomopsis sp. from Notobasis syriaca. Nat Prod Commun 6:1905–1906. [CrossRef]
- Amaral CR do, Mazarotto EJ, Gregório PC, Favretto G (2023) Identification of endophytic fungi in Hamelia patens Jacq. and evaluation of the antimicrobial potential of fungal extracts. Research, Society and Development 12:e23012541767. [CrossRef]
- Hatamzadeh S, Rahnama K, Nasrollahnejad S, et al (2020) Isolation and identification of L-asparaginase-producing endophytic fungi from the Asteraceae family plant species of Iran. PeerJ 8:e8309. [CrossRef]
- Lu H, Zou WX, Meng JC, et al (2000) New bioactive metabolites produced by Colletotrichum sp., an endophytic fungus in Artemisia annua. Plant Science 151:67–73.
- Aron AT, Gentry EC, McPhail KL, et al (2020) Reproducible molecular networking of untargeted mass spectrometry data using GNPS. Nat Protoc 15:1954–1991. [CrossRef]
- Santos AL, Ionta M, Horvath R, et al (2021) Molecular network for accessing polyketide derivatives from Phomopsis sp., an endophytic fungus of Casearia arborea (Salicaceae). Phytochem Lett 42:1–7. [CrossRef]
- Xiao J, Zhang Q, Gao YQ, et al (2014) Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J Agric Food Chem 62:3584–3590. [CrossRef]
- Oliveira CM, Regasini LO, Silva GH, et al (2011) Dihydroisocoumarins produced by Xylaria sp. and Penicillium sp., endophytic fungi associated with Piper aduncum and Alibertia macrophylla. Phytochem Lett 4:93–96. [CrossRef]
- Catellani A (1967) A Maintenance and Cultivation of the common pathogenic fungi of man in sterile distilled water. American Journal of Tropical Medicine and Hygiene 70:181–184.
- Neufeld PM, Oliveira PC de (2008) Preservation of dermatophyte fungi by sterile distilled water technique. Revista Brasileira de Análises Clínicas 40:167–169.
- Abreu MMV, Tutunji VL (2008) Implantation and maintenance of microbial culture collection in UniCEUB. Universitas: Ciências da Saúde 2:236–251.
- Aparecido CC, Rosa EC, Costa IAM, Jorge CM (2018) Avaliação de diferentes métodos para preservação de fungos fitopatogênicos. Biológico 80:1–7.
- Pires GCC, Aparecido CC, Finatti D (2012) Divulgação técnica de preservação em laboratório de fungos filamentosos por longos períodos de tempo. Biológico 74:9–16.
- Negri CE, Gonçalves SS, Xafranski H, et al (2014) Cryptic and Rare Aspergillus species in Brazil: Prevalence in clinical samples and in Vitro susceptibility to Triazoles. J Clin Microbiol 52:3633–3640. [CrossRef]
- Katoh K, Rozewicki J, Yamada KD (2018) MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform 20:1160–1166. [CrossRef]
- Capella-Gutiérrez S, Silla-Martínez JM, Gabaldón T (2009) trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 25:1972–1973. [CrossRef]
- Minh BQ, Schmidt HA, Chernomor O, et al (2020) IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol Biol Evol 37:1530–1534. [CrossRef]
- Letunic I, Bork P (2021) Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res 49:W293–W296. [CrossRef]
- Hoog GS, Guarro J, Gené J (2020) The Ultimate Benchtool for Diagnostics. In: Atlas of Clinical Fungi:, 4th ed. Foundation Atlas of Clinical Fungi, p 776.
- Gonçalves SS, Cano JF, Stchigel AM, et al (2012) Molecular phylogeny and phenotypic variability of clinical and environmental strains of Aspergillus flavus. Fungal Biol 116:1146–1155. [CrossRef]
- Silva WC, Gonçalves SS, Santos DWCL, et al (2017) Species diversity, antifungal susceptibility and phenotypic and genotypic characterization of Exophiala spp. infecting patients in different medical centers in Brazil. Mycoses 60:328–337. [CrossRef]
- Wang M, Carver JJ, Phelan V V., et al (2016) Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat Biotechnol 34:828–837. [CrossRef]
- Frank AM, Bandeira N, Shen Z, et al (2008) Clustering millions of tandem mass spectra. J Proteome Res 7:113–122. [CrossRef]
- Shannon P, Markiel A, Ozier O, et al (2003) Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res 13:2498–2504. [CrossRef]
- 2003; 42. Costa SF, Gales A, Machado AM de O (2003) Padronização dos testes de sensibilidade a antimicrobianos por disco-difusão.
- Santos NO, Mariane B, Lago JHG, et al (2015) Assessing the chemical composition and antimicrobial activity of essential oils from Brazilian plants - Eremanthus erythropappus (Asteraceae), Plectrantuns barbatus, and P. amboinicus (Lamiaceae). Molecules 20:8440–8452. [CrossRef]
- Lambert C, Wendt L, Hladki AI, et al (2019) Hypomontagnella (Hypoxylaceae): a new genus segregated from Hypoxylon by a polyphasic taxonomic approach. Mycol Prog 18:187–201. [CrossRef]
- Huang Y, Chen Z, Huang M, et al (2023) Cytotoxic tetronic acid derivatives from the mangrove endophytic fungus Hypomontagnella monticulosa YX702. Fitoterapia 170:. [CrossRef]
- Sumiya T, Ishigami K, Watanabe H (2010) Determination of the absolute configuration of nodulisporacid A by the concise synthesis of four stereoisomers via three-component reaction and one-pot construction of the framework. Tetrahedron Lett 51:2765–2767. [CrossRef]
- Vieira MLA, Johann S, Hughes FM, et al (2014) The diversity and antimicrobial activity of endophytic fungi associated with medicinal 1 plant Baccharis trimera (Asteraceae) from the Brazilian savannah 2 3. Canadian Jornal of Microbiology 60:847–856.
- Nicoletti R, Fiorentino A (2015) Plant bioactive metabolites and drugs produced by endophytic fungi of Spermatophyta. Agriculture (Switzerland) 5:918–970.
- Pamphile JA, Costa AT, Rosseto P, et al (2017) Biotchnological applications of secondary metabolites extracted from endophytic fungi: The case of Colletotrichum sp. Revista UNINGÁ 53:113–119.
- García-Pajón CM, Collado IG (2003) Secondary metabolites isolated from Colletotrichum species. Nat Prod Rep 20:426–431. [CrossRef]
- Wang KL, Wu ZH, Wang Y, et al (2017) Mini-review: Antifouling natural products from marine microorganisms and their synthetic analogs. Mar Drugs 15:266. [CrossRef]
- Yang XL, Zhang JZ, Luo DQ (2012) The taxonomy, biology and chemistry of the fungal Pestalotiopsis genus. Nat Prod Rep 29:622–641. [CrossRef]
- Chen Z, Dong Z, Wen J, et al (2016) A New Sesquiterpene from the Endophytic Fungus Nigrospora sphaerica #003. Natural Product 10:307–310.
- Fukushima T, Tanaka M, Gohbara M, Fuworzl T (1998) Phytotoxicity of three lactones sacchari from Nigrospora. Phytochemistry 48:625–630.
- Meepagala KM, Becnel JJ, Estep AS (2015) Phomalactone as the Active Constituent against Mosquitoes from Nigrospora spherica. Agricultural Sciences 06:1195–1201. [CrossRef]
- Ibrahim D, Chai Lee C, Yenn TW, et al (2015) Effect of the extract of endophytic fungus, Nigrospora sphaerica cl-op 30, against the growth of ethicillin-resistant Staphylococcus aureus (Mrsa) and Klebsiella pneumonia cells. Tropical Journal of Pharmaceutical Research 14:2091–2097. [CrossRef]
- Aly AH, Debbab A, Proksch P (2011) Fungal endophytes: Unique plant inhabitants with great promises. Appl Microbiol Biotechnol 90:1829–1845. [CrossRef]
- Jalgaonwala RE, Vishwas Mohite B, Mahajan RT (2011) A review: Natural products from plant associated endophytic fungi. J Microbiol Biotechnol 1:21–32.
- Strobel G, Daisy B (2003) Bioprospecting for Microbial Endophytes and Their Natural Products. Microbiology and Molecular Biology Reviews 67:491–502. [CrossRef]
- Ćeranić A, Svoboda T, Berthiller F, et al (2021) Identification and functional characterization of the gene cluster responsible for fusaproliferin biosynthesis in fusarium proliferatum. Toxins (Basel) 13:. [CrossRef]
- 2012; 61. Canuto KM, Rodrigues THS, Oliveira FSA, Gonçalves FJT (2012) Fungos endofíticos.
- Sumiya T, Ishigami K, Watanabe H (2010) Determination of the absolute configuration of nodulisporacid A by the concise synthesis of four stereoisomers via three-component reaction and one-pot construction of the framework. Tetrahedron Lett 51:2765–2767. [CrossRef]
- Kasettrathat C, Ngamrojanavanich N, Wiyakrutta S, et al (2008) Cytotoxic and antiplasmodial substances from marine-derived fungi, Nodulisporium sp. and CRI247-01. Phytochemistry 69:2621–2626. [CrossRef]
- 1996; 64. Logrieco A, Moretti A, Fornelli F, et al (1996) Fusaproliferin Production by Fusarium subglutinans and Its Toxicity to Artemia salina, SF-9 Insect Cells, and IARC/LCL 171 Human B Lymphocytes.
- Pereira RG, Garcia VL, Nova Rodrigues MV, Martínez J (2016) Extraction of lignans from Phyllanthus amarus Schum. & Thonn using pressurized liquids and low-pressure methods. Sep Purif Technol 158:204–211. [CrossRef]
- Hemmerling F, Piel J (2022) Strategies to access biosynthetic novelty in bacterial genomes for drug discovery. Nat Rev Drug Discov 21:359–378.
Figure 4.
Structures of the Z and E isomers of Nodulisporacid A (1) and Nodulisfuranol (2).
Figure 5.
Evaluation of antimicrobial activity of crude extracts (CPFF12 and CPFF14) by disc diffusion test. Inhibition halo formed by the crude extract CPPF12 against A) C. albicans; B) S. cerevisiae, crude extract CPPF12 against C) S. aureus and crude extract CPPF41 against D) S. aureus.
Figure 5.
Evaluation of antimicrobial activity of crude extracts (CPFF12 and CPFF14) by disc diffusion test. Inhibition halo formed by the crude extract CPPF12 against A) C. albicans; B) S. cerevisiae, crude extract CPPF12 against C) S. aureus and crude extract CPPF41 against D) S. aureus.
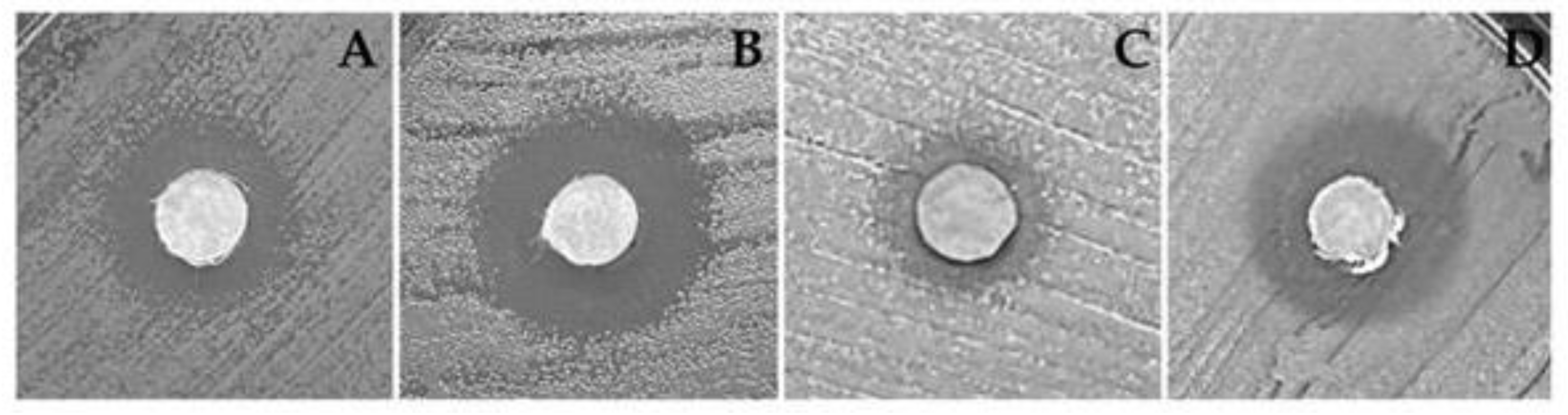

Table 1.
Yeast strains used in yeast inhibitory activity tests.
| Species | Cell line |
|---|---|
| Cryptococcus neoformans | KN99α (serotype A) |
| Cryptococcus neoformans | JEC21 (serotype D) |
| Cryptococcus gattii | NIH312 (serotype C) |
| Cryptococcus gattii | R265 (serotype B) |
| Saccharomyces cerevisiae | BY4647 |
| Candida krusei | Clinical isolate 9602 |
| Candida parapsilosis | Clinical isolate 68 |
| Candida albicans | CBmay 560 |
| Candida tropicalis | ATCC 1303 |
| Candida dubliniensis | ATCC 7876 |
Table 2.
Bacterial strain used for disc diffusion and minimum inhibitory concentration assays.
| Species | Cell line |
|---|---|
| Enterococcus faecium | ATCC CCB076 |
| Pseudomonas aeruginosa | ATCC 27853 |
| Klebsiella pneumoniae | ATCC 700603 |
| Escherichia coli | ATCC 25922 |
| Shigella flexneri | ATCC 12022 |
| Salmonella enterica | ATCC 14028 |
| Staphylococcus aureus | ATCC25923 |
| Acinetobacter baumanii | ATCC 19606 |
Table 3.
Species identification of isolates based on sequences obtained from ITS region and fragment of the TUB2 gene.
Table 3.
Species identification of isolates based on sequences obtained from ITS region and fragment of the TUB2 gene.
| Strain Number | Species | % Identity* | |||
| ITS | Cod. | TUB2 | Cod. | ||
| CPFF12 | Colletotrichum karstii | 100 | PP831945 | 100 | PP840075 |
| CPFF14 | Colletotrichum karstii | 99.39 | PP829193 | 99.72 | PP840076 |
| CPFF16 | Colletotrichum siamense | 100 | PP829196 | 99.60 | PP840077 |
| CPFF41 | Hypomontagnella barbarensis | 99.23 | PP829195 | 93.24 | PP840078 |
| CPFF42 | Neopestalotiopsis clavispora | 100 | PP829194 | 99.09 | PP942538 |
| CPFF52 | Nigrospora sacchari-officinarum | 99.08 | PP832015 | 100.00 | PP840079 |
| CPFC2 | Annulohypoxylon moriforme | 98.50 | PP834406 | 98.09 | PP840080 |
* Blast NCBI.
Table 4.
Annotation of compounds on GNPS spectral library match.
| Compound name |
Molecular formula |
Calculated mass |
Precursor | Error (ppm) | Sample | Structural formula |
|
|---|---|---|---|---|---|---|---|
| 1 | 1-Acetoxy-7-isopropylidene-1,4a-dimethyl-6-oxodecahydro-2-naphthalenyl 2,3-dimethyl-2-oxiranecarboxylate | C22H32O6 | 415.2097 [M+Na]+ |
415.2097 | 0.1 | CPFF12 CPFF16 |
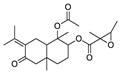 |
| 2 | 13-keto-9Z,11E-octadecadienoic acid | C18H30O3 | 295.2273 [M+H]+ |
295.2275 | 0.7 | CPFF41 CPFF42 |
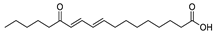 |
| 3 | Alismol | C15H24O | 203.1799 [M+H]+ |
203.1797 | 1.0 | CPFF14 CPFF52 |
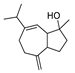 |
| 4 | Phytosphingosine | C18H39NO3 | 318.3008 [M+H]+ |
318.3000 | 2.6 | CPFF12 CPFF16 CPFF52 |
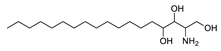 |
| 5 | Fusaproliferin | C27H40O5 | 445.2954 [M+H]+ |
445.2949 | 1,1 | CPFF12 CPFF14 |
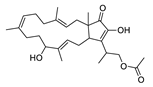 |
Table 5.
Disk diffusion method (DDM) − Zone of inhibition (mm).
|
Extract (400 μg) |
Species (lineage) | ||
|
S. cerevisiae (BY4647) |
C. albicans (CBmay 560) |
S. aureus (ATCC 25923) |
|
| CPFF12 | 1.4 ± 0.26 | 1.23 ± 0.25 | n/a |
| CPFF14 | n/a | n/a | 0.73 ± 0.23 |
| CPFF41 | n/a | n/a | 1.37 ± 0.32 |
| Control* | 2.07 ± 0.12a | 2.07 ± 0.12b | 2.5 ± 0.0b |
* aEEugenol (200 μg); bKanamicin (50 μg). n/a – not active.
Table 6.
Minimum inhibitory concentration (MIC).
| Specie | Extract - MIC90 (mg/mL) | Control* | ||
|---|---|---|---|---|
| CPFF12 | CPFF14 | CPFF41 | ||
| S. aureus (ATCC25923) | n/a | 0.20 (91.6±1.5) | 0.05 (92.0±1.0) | 0.004a |
| C. albicans (CBmay 560) | 0.01 (92.0±1.9) | n/a | n/a | 0.025b |
| S. cerevisiae (BY4647) | 0.02 (90.3±1.5) | n/a | n/a | 0.013c |
*a Vancomycin; bChloramphenicol; cFluconazole. n/a – not active.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



