
Submitted:
06 April 2023
Posted:
07 April 2023
You are already at the latest version
Abstract
The rhizosphere is a rich source of actinomycetes which can produce several potential biologically-active secondary metabolites. The principal goals for this research are to extract, purify and characterize the bioactive secondary metabolites produced by three different strains of actinomycetes isolated from the rhizosphere of rosemary, black locust and olive. The plant growth-promoting effect (PGPE) of the studied strains of actinomycetes on Ocimum basilicum L. (basil) and disease-control effect on necrotic stem lesions of “black leg” caused by Fusarium tabacinum on basil, were in silico evaluated. The cell-free culture filtrates from the studied actinomycetes isolates were evaluated in vitro for their antimicrobial activity against some common phytopathogens. The secondary metabolites obtained from the cell-free culture filtrates have been chemically characterized using high‐resolution electrospray ionization of liquid-chromatography/mass-spectrometric detection (ESI-(HR)Orbitrap-MS). Results of in silico trial showed that all studied isolates demonstrated PGPE on basil seedlings improving some Eco-physiological characteristics and reduced the disease incidence of F. tabacinum. The extracted metabolites from the studied actinomycetes demonstrated antimicrobial activity in Petri-plates assay. The chemical analysis revealed the presence of totally 20 different components. This research emphasizes how valuable the examined isolates are for producing bioactive compounds, indicating their putative antimicrobial activity and their potential employment as fungal-biocontrol agents. In particular, the obtained results revealed the possibility of green-synthesis of some important secondary metabolites such as N-Acetyl-L-histidinol, Rhizocticin A and Eponemycin from actinomycetes. The bioactive metabolites may be successively used to develop novel bio-formulations for both crop protection and/or PGPE.
Keywords:
antimicrobial activity
; natural products
; microbial metabolites
; plant diseases
; biological control
1. Introduction
Bioactive substances are abundantly produced by soil microorganisms [1]. Due to bacteria's capacity to produce a variety of useful products, such as antibiotics, fungicides, herbicides, hydrolytic enzymes, antitumor, antivirals and immune-suppressants, there is increased interest in employing them for medical and agricultural applications [2,3,4]. Recently, the pathogen resistance demands to discover novel antimicrobial substances that are effective against serious phytopathogens; so, a high interest to screen new microbes from various environments, with antimicrobial activity, was arised to explore novel potential medications against infections that are resistant to drugs. Among these new isolates are actinomycetes, unicellular filamentous gram-positive bacteria, found throughout nature in a wide range of environments. Actinomycetes are prominent and significant producers of numerous biological by-products such as antibiotics and plant growth promoting substances [5]. Actinomycetes are very similar to fungi due to their ability to form a mycelium (hyphae), but their hyphae are much smaller than fungal ones [5,6].
Actinobacteria is regarded as one of the major groups of actinomycetes, which includes the genus Streptomyces which produces several know antibiotics [4]. According to Berdy [7], the majority of the discovered bioactive substances are produced by genus Streptomyces. Girão et al. [8] reported the presence of several bioactive compounds, which account for around 45% of all known microbial bioactive metabolites, have been isolated from actinobacteria from terrestrial sources. Girão et al. [8] investigated the ability of several actinobacteria isolated from Laminaria ochroleucahe to control Candida albicans and Staphylococcus aureus. Therefore, new actinomycetes can be isolated for discovering novel bioactive compounds for agriculture or medicine purposes.
Ocimum basilicum L. (basil) is a culinary herb of Lamiaceae family. It is native of tropical zones of central Africa and southeast Asia and can be also found in Mediterranean region [9]. Basil can be infected by several phytopathogens such as Fusarium sp. (wilt disease), Pythium sp (damping off), Botrytis sp. (gray mold), Colletotrichum sp. (black-spot) and Peronospora sp. (downy mildew) [10]. In particular, F. tabacinum W. Gams (Beyma) causes the necrotic stem lesions “black leg” disease on basil [11].
The primary goals of the present research were: i) extraction and the purification of the extracts produced from three studied actinomycetes isolates; ii) chemical characterization of the metabolites of the three isolates using HR-LC-ESI-Orbitrap-MS; iii) in vitro evaluation of their antimicrobial activity against some phytopathogenic strains; iv) in silico evaluation of the promoting effect of the three actinomycetes strains on O. basilicum and controlling “black leg disease” on basil caused by F. tabacinum.
2. Results
2.1. Actinomycetes isolation and indentification
The morphological examination using the optical microscope of the three pure-cultures showed the typical like-mycelia structure of actinomycetes characterized by branching and filamentous growth. In particular, the colony of isolate Act1 produced well-developed vegetative radial-hyphae ranged between(1.5 – 2.0 µm Ø, consisting of long-semi-straight filaments bearing colonies arranged vertically in whorls with irregular shape (Figure 1A,B). The isolate Act2 showed a dense radial-hyphae ranged between 1.5 – 2.0 µm Ø and consists of long-semi-straight filaments bearing colonies arranged vertically in whorls with irregular shape (Figure 1C,D). The isolate Act3 showed a radial-hyphae consist of long straight filaments bearing colonies with regular coccoid-shape (Figure 1E,F).
The sequences of the amplified DNA have been carried out in BMR Genomics (Padova, Italy) as previously reported by Elshafie and Camele [12]. BLAST software has been used for comparing the obtained sequences with those available in GenBank. The analysis demonsrated strong similarities percentages with Streptomyces sp., Streptomyces atratus and Arthrobacter humicola present in GenBank. One sequence for each studied isolate was deposited in the NCBI GenBank with the following accession numbers ON241810, ON241816 and ON241806 for Streptomyces sp. (Act1), Streptomyces atratus (Act2) and Arthrobacter humicola (Act3), respectively.
2.2. Growth-promoting and disease-control effects
2.2.1. Eco-physiological characteristics
All bacterized plants with the actinomycetes isolates resulted able to promote the growth of basil seedlings and showed higher values of Eco-physiological characteristics compared to the negative control (non-bacterized plants) (Table 1). In particular, seedlings inoculated with Act1 and Act3 showed the higher significant values of NL, SL and TDwS (Table 1). In addition, Act1 and Act2 showed the higher significant values regarding TFwS (Table 1).
On the other hand, the Table 2 reports the Eco-physiological characteristics of bacterized basil infected with F. tabacinum. In particular, seedlings inoculated with Act1 and Act3 demonstrated higher significant values of NT, SL and TFwS (Table 2). In addition, Act1 showed the higher significant values regarding TDwS (Table 1). Furthermore, Act2 showed a moderate PGPE on basil seedlings especially NT, SL and TDwS (Table 2).
2.2.2. Disease-control effect
The bacterized-plants with Streptomyces sp. and A. humicola exhibited no foliar or radical symptoms due to S. sclerotiorum infection. In fact, the disease indexes of Streptomyces sp. and A. humicola were 0.3 and 0.2 %, respectively whereas the control effects were 99.2 and 99.5 %, respectively (Table 3). On the other hand, S. atratus-bacterized seedlings had a moderate disease index higher than 7 % and control effect higher than 79 % (Figure 2 and Figure 3).
Results showed also that the C+ve (plants only inoculated with F. tabacinum) developed leaf chlorosis 20 DPI and later turns necrotic. In addition, a high percentage of leaf wilt and root necrosis were also observed 35 DPI. In particular, a significant high percentage of symptomatic leaves was observed in the case of seedlings infected with F. tabacinum, where the disease-index was higher than 36 % compared to C-ve (plants not inoculated with F. tabacinum or actinomycetes isolates) and plants bacterized with actinomycetes isolates (Figure 2, Table 3). F. tabacinum was always re-isolated from the infected-plants.
2.3. Antimicrobial assay of metabolites
2.3.1. Antifungal activity
The extracts have been evaluated for their antifungal activity against some food and phytopathogenic fungi following the incorporation method (Figure 4). A. humicola showed the highest antifungal effect against all tested fungi. Streptomyces sp. showed a moderate antifungal effect higher than S. atratus particularly against M. fructigena, R. solani, F. oxysporum, P. expansum, P. digitatum. On the other hand, S. atratus showed a promising effect against S. sclerotiorum and P. commune.
2.3.2. Antibacterial activity
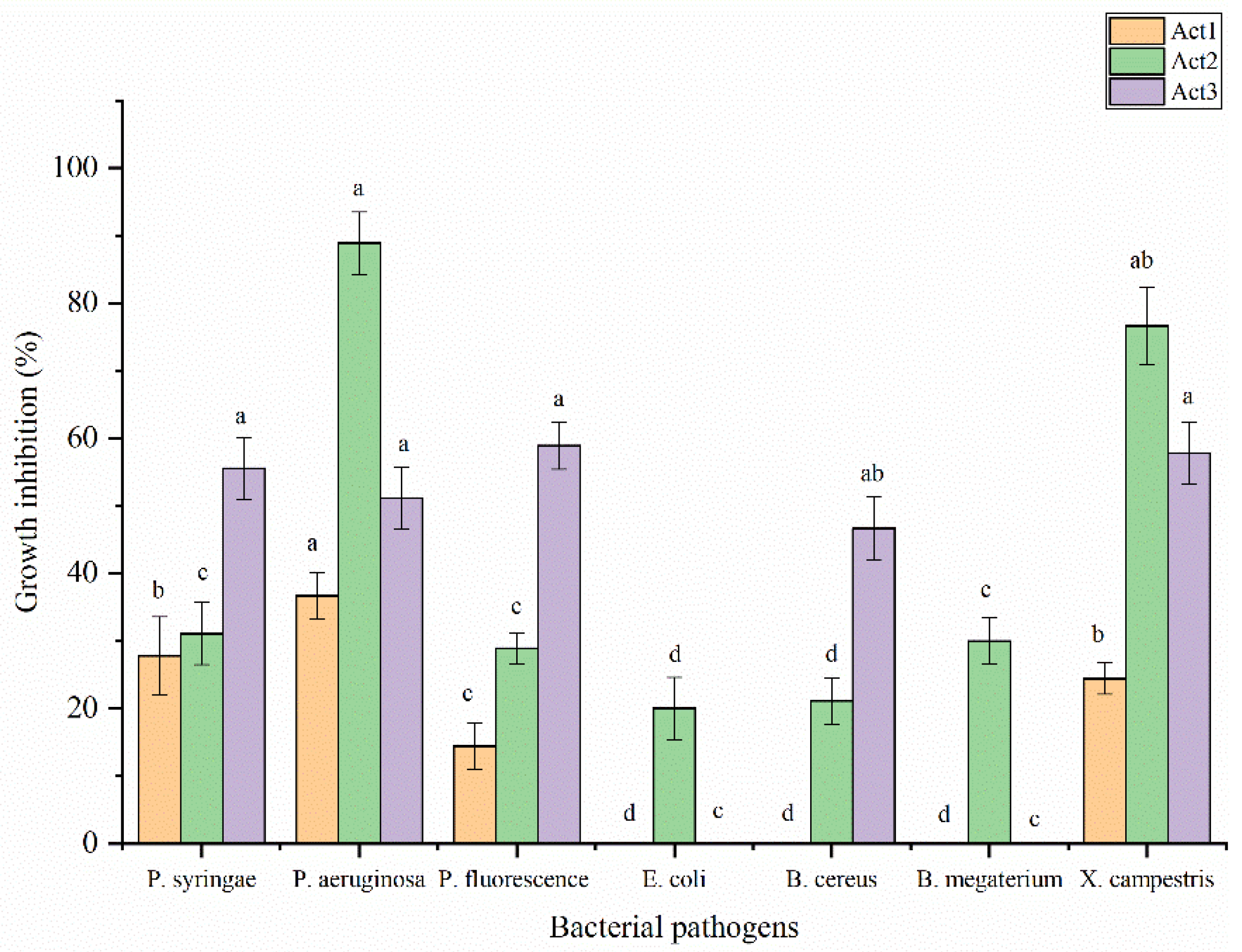
Regarding the antibacterial activity, the extracts have been evaluated against some food and plant pathogenic bacteria following the disc diffusion method (Figure 5). The highest significant activity of S. atratus has been observed against P. aeruginosa and X. campestris showing BGI% at 88.9 and at 76.7%, respectively. A. humicola showed the highest activity against P. fluorescens, X. campestris, P. syringae and P. aeruginosa showing BGI% at 58.9, 57.8, 55.6 and 51.1%, respectively. Streptomyces sp. showed moderate activity against P. aeruginosa, P. syringae and X. campestris with BGI% at 36.7, 27.8 and 24. %, respectively. In addition, the lowest activity has been observed in case of S. atratus against B. cereus and E. coli with BGI% at 21.1 and 20.0%, respectively. On the other hand, Streptomyces sp. resulted not active against E. coli, B. cereus and B. megaterium. In case of A. humicola no activity was observed against E. coli and B. megaterium.
Figure 4.
Antifungal activity of extracted metabolites. Bars with different letters between different phytopathogenic fungi for each treatment are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
Figure 4.
Antifungal activity of extracted metabolites. Bars with different letters between different phytopathogenic fungi for each treatment are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
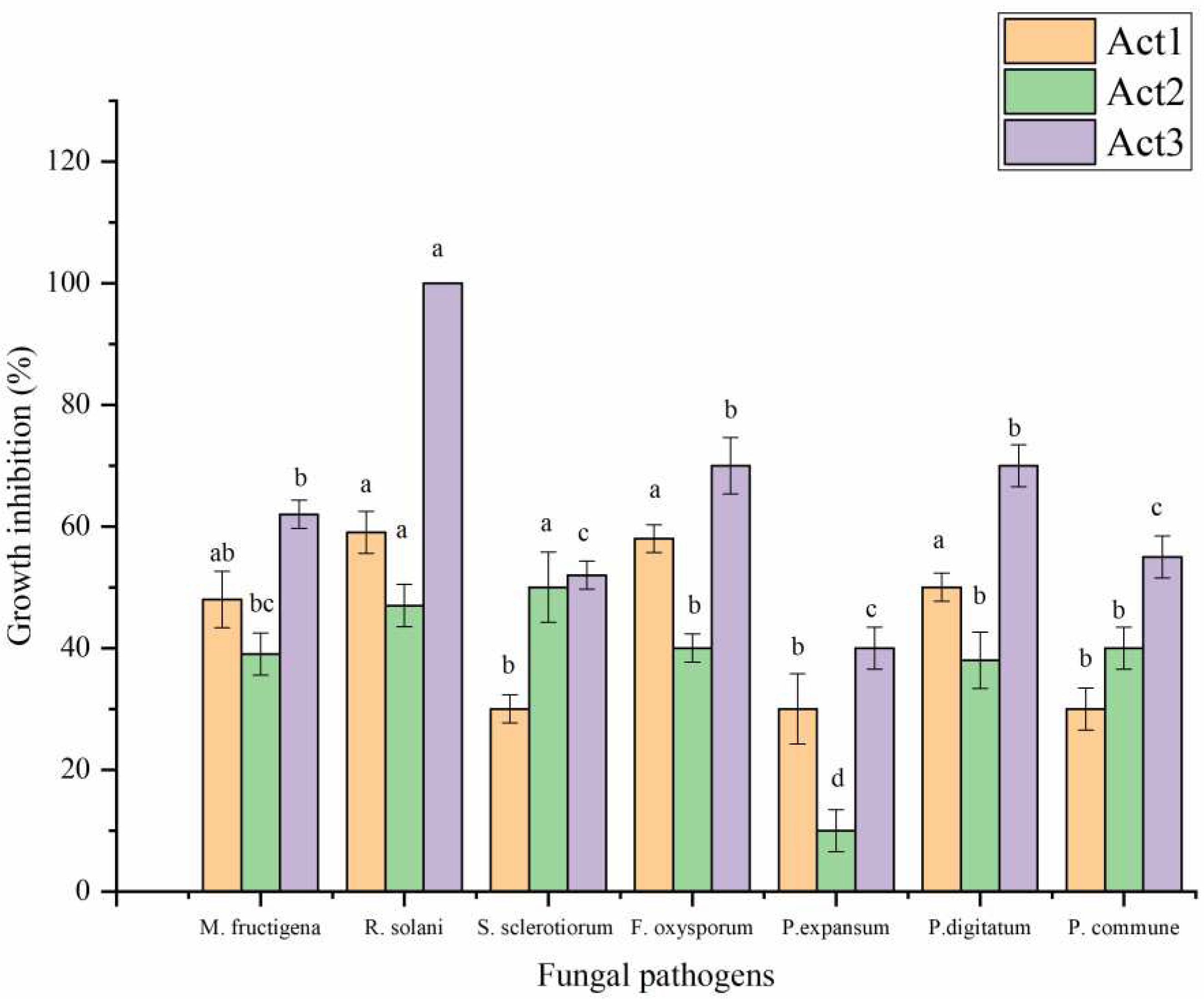
Figure 5.
Antibacterial activity of extracted metabolites. Bars with different letters between different phytopathogenic bacteria for each treatment are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
Figure 5.
Antibacterial activity of extracted metabolites. Bars with different letters between different phytopathogenic bacteria for each treatment are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
2.4. ESI/(HR)orbitrap/MS metabolic profiles
The chemical analysis of three studied actinomycetes isolates revealed the presence of 20 components, as illustrated in Table 4, (Figures S1–S3). Particularly, compound 1: N-Acetyl-L-histidinol, identified both in Streptomyces sp. (Act1) and S. atratus (Act2) extracts, which eluted at a retention time of 0.87 min, showed a peak [M+H]+ ion at m/z 184.1076, corresponding to a protonated molecular ion. From the spectrum, a produced ion at m/z 166 corresponding to a loss of oxydrile group, was also evident. We noted that the compound generating the protonated ion at 184.1076, is a prominent peak in all the extracts and this compound was previously isolated from Streptomyces coelicolor [13].
Compound 2: N,N′-Diacetyl-2-deoxystreptamine, was identified only in A. humicola (Act3) extract, has molecular ion [M+H]+ at m/z 247.1283. 2-Deoxystreptamine antibiotics constitute a large group of aminoglycoside antibiotics to which over 20 members have been assigned to date, including the paromomycins, the kanamycins, the gentamicins, the nebramycins, ribostamycin, the lividomycins, validomycin and ambutyrosin [14].
Compound 3: Indolacatm V, displaying sodiated ion of 324.1659 which is consistent with Indolacatm V, only identified in Streptomyces sp. (Act1) extract, with retention time 1.18 min. This compound was previously isolated from Streptomyces blastmyceticus showing antagonistic activity toward protein kinase C [15].
Compound 4: Hexahydro-2H-pyrido[1,2-a]pyrazin-3(4H)-one, plue protonated ion [M+H]+ (m/z 155.117), detected only in S. atratus (Act2) extract. This compound was previously found in a Streptomyces extract, as reported by Nithya et al [16].
Compounds 5,6: Compound 5 corresponds to valyldetoxinine with molecular ion [M+H]+ at m/z 275.159, whereas 6 corresponds to paromomycin, with potassium molecular ion [M+K]+ at m/z 654.265. These two compounds were found only in A. humicola (Act3) extract. Valyldetoxinine was a member of detoxin complex, a group of depsipeptide metabolites produced by Streptomyces caesputosus var. detoxus, present in the soil [17]. The detoxins exhibit antimicrobial activity against some microorganisms. Paromomycin is a naturally occurring aminoglycoside antibiotic, produced by Streptomyces rimosus sp. paromomycinus, that affects both prokaryotic and eukaryotic through the chelation with A-site of ribosome [18,19]. Valyldetoxinine and paromomycin hadn’t ever been previously reported in A. humicola.
Compound 7: N-[3-[5-(2-methylpropyl)-3,6-dioxo-2-piperazinyl]propyl]-N′-nitro-,(2S-cis)-Guanidine, present in both extracts of Streptomyces sp. (Act1) and S. atratus (Act2), was eluted at 7.28 min, with a hydrate molecular ion [M+H2O]+ at m/z 332.1812 [20]. This compound was a cyclic dipeptide (CDP), widely biosynthesized during the cyclo-dipeptide synthsis cycle either within prokaryotic and/or eukaryotic cells [21]. This compound was also found in Streptomyces strains [22].
Compound 8: n-Hexyl-β-D-glucoside, detected in Streptomyces sp. (Act1) and S. atratus (Act2) extracts, with a [M+H]+ ion at m/z 265.1654; it is a substrate of β-glucosidases, hydrolytic enzymes, normally present in Streptomyces strains, that cleave β-glycosidic bonds of carbohydrates [23].
Compounds 9-11: Nonactic acid (9) and homononactinic acid (11) identified only in S. atratus (R3) extract, with [M+H]+ ion at m/z 203.1274 and m/z 217.1430 corresponding to protonated molecular ions, respectively. These two compounds have been isolated from Streptomyces and assayed against a panel of cancer cell lines as reported by Lu et al. [24].
Compound 10: (11aS)-1,2,3,11a-Tetrahydro-8-hydroxy-7-methoxy-5H-pyrrolo[2,1-c][1,4]benzodiazepin-5-one, also known as antibiotic DC81, present in Streptomyces sp. (Act1) and S. atratus (Act2), with m/z 247.1073, previously reported as antitumor and antibiotic compound originated from some actinomycetes [25].
Compounds 12-15: these compounds were identified only in Streptomyces sp (Act1) extract. Compound 12 gave a [M+H]+ ion at m/z 307.1646 attributed to phthoxazolins B, C or D. These compounds were isolated from the culture broth of Streptomyces sp., as reported by Shiomi et al. [26], but the limitation of LC-MS is that metabolites are structural isomers which cannot be distinguished. Compound 13 was identified as Chandrananimycin D, based on accurate mass at m/z 301.0811, attributing to a protonated molecular ion [M+H]+: the phenoxazinone chandrananimycin D was firstly characterized and isolated from Streptomyces griseus; it also reported for its antiproliferative activity [27]. The compound 14 gave a [M+H]+ ion at m/z 299.0659, attributed to a protonated molecular ion of carboxyexfoliazone. This compound was firstly report from a wild-type of Streptomyces strain, as reported by Abdelfattah et al. [28]. Compound 15 eluted at 10.52 min, had a molecular ion [M+H]+ at m/z 283.0705 corresponding to a protonated molecular ion of phencomycin and previously reported by Chatterjee et al. [29].
Compounds 16-18: these compounds were detected only in A. humicola (Act3) extract, giving [M+H]+ ion at m/z 288.2889 and [M+H] + ion at 346.2220, respectively. Particularly, compound 16 was attributed to a protonated molecular ion of 2-amino-3-hydroxyhexadecanoic acid, and has been already reported previously in Arthrobacter genus [30]. Compound 18 was attributed to a protonated molecular ion of Maoxianamides A or B. These two compounds were isolated from Streptomyces maoxianensis, as reported by Li et al., [31]. Compound 17: 1,1-Dimethylethyl 2-[2-(ethoxycarbonyl)-1-cyclopenten-1-yl]diazenecarboxylate, found only in S. atratus (Act2) extract, gave a [M+H]+ ion at m/z 269.14923.
Compound 19: this compound was attributed to a protonated molecular ion Rhizocticin A, was identified both in Streptomyces sp. (Act1) and S. atratus (Act2) extracts, gave a [M+H]+ ion at m/z 352.30490. This compound is a natural phosphonate antibiotic produced by the bacterial strain Bacillus subtilis [32] and it is very similar to plumbemycin, isolated from Streptomyces plumbeus, for amino acid (Z)-2-amino-5-phosphono-3-pentenoic acid, present in both antibiotics [33].
Compound 20: this compound was attributed to a protonated molecular ion of Eponemycin, was identified in all three studied isolates, gave a [M+H]+ ion at m/z 399.24997. This compound was reported to be produced by Streptomyces hygroscopicus and showed potent growth inhibition against various tumor cells [34]. Recently, Fitri et al. [35] reported that this compound is a potential candidate for a new antimalarial drug due to its efficacy against Plasmodium berghei.

Table 4.
Chromatographic analysis of extracted secondary metabolites from studied actinomycetes isolates.
Table 4.
Chromatographic analysis of extracted secondary metabolites from studied actinomycetes isolates.
| No. | Retention time (min) | Measured m/z |
Molecular formula | Identification | Act1 | Act2 | Act3 |
|---|---|---|---|---|---|---|---|
| 1 | 0.87 | 184.1075 | C8H13N3O2 | N-Acetyl-L-histidinol | X | X | |
| 2 | 1.17 | 247.12834 | C10H18N2O5 | N,N′-Diacetyl-2-deoxystreptamine | X | ||
| 3 | 1.18 | 324.16592 | C17H23N3O | Indolactam V | X | ||
| 4 | 1.19 | 155.11748 | C8H14N2O | Hexahydro-2H-pyrido[1,2-a]pyrazin-3(4H)-one | X | ||
| 5 | 2.30 | 275.15955 | C12H22N2O5 | Valyldetoxinine | X | ||
| 6 | 7.11 | 654.2653 | C23H45N5O14 | Paromomycin | X | ||
| 7 | 7.28 | 332.18118 | C12H22N6O4 | Guanidine, N-[3-[5-(2-methylpropyl)-3,6-dioxo-2-piperazinyl]propyl]-N′-nitro-, (2S-cis) | X | X | |
| 8 | 7.73 | 265.16544 | C12H24O6 | n-Hexyl-β-D-glucoside | X | X | |
| 9 | 7.88 | 203.12737 | C10H18O4 | Nonactinic acid | X | ||
| 10 | 8.47 | 247.10730 | C13H14N2O3 | (11aS)-1,2,3,11a-Tetrahydro-8-hydroxy-7-methoxy-5H-pyrrolo[2,1-c][1,4]benzodiazepin-5-one | X | X | |
| 11 | 8.61 | 217.14296 | C11H20O4 | Homononactinic acid | X | ||
| 12 | 8.81 | 307.16461 | C16H22N2O4 | phthoxazolins B, C and D | X | ||
| 13 | 9.43 | 301.08112 | C15H12 N2O5 | Chandrananimycin D | X | ||
| 14 | 9.75 | 299.06592 | C15H10N2O5 | Carboxyexfoliazone | X | ||
| 15 | 10.52 | 283.07053 | C15H10O4N2 | Phencomycin | X | ||
| 16 | 11.32 | 288.2889 | C16H33NO3 | 2-Amino-3-hydroxyhexadecanoic acid | X | ||
| 17 | 11.45 | 269.14923 | C13H20N2O4 | 1,1-Dimethylethyl 2-[2-(ethoxycarbonyl)-1-cyclopenten-1-yl]diazenecarboxylate | X | ||
| 18 | 12.36 | 346.2220 | C17H31O6N | Maoxianamide A or B | X | ||
| 19 | 12.20 | 352.30490 | C11H22N5O6P | Rhizocticin A | X | X | |
| 20 | 14.09 | 399.24997 | C20H34N2O6 | Eponemycin | X | X | X |
3. Discussion
The obtained results are promising for biocontrol of F. tabacinum where the treatments with actinomycetes isolates showed a high reduction of disease symptoms on O. basilicum seedlings, demonstrating the control effect exerted by Streptomyces sp. and Arthrobacter humicola against F. tabacinum. Furthermore, the treatment with the three studied isolates of actinomycetes may induce also the resistance effect against F. tabacinum. In addition, the obtained results of the current study are in agree with the results reported by Elshafie and Camele [12], where the same studied strains showed the capacity to stimulate the development of S. lycopersicum and reduced the disease incidence of S. sclerotiorum. The results agree with several research reporting that many actinomycetes from the soil can inhibit some harmful phytopathogenic fungi [36,37,38].
The PGPE of the studied isolates showed also significant influences on the Eco-physiological characteristics of basil plants which may be due to synthesis of phytohormones such as gibberellic acid, Indole 3-acetic acid and zeatine (Z) produced by symbiotic and saprophytic actinomycetes [39].
Chaudhary et al. [1] investigated the antagonistic behavior of some actinomycetes isolates obtained from various niche environments in India, and observed the bioactivity of some of studied isolates against Bacillus cereus, Shigella dysenteriae and Methicillin-resistant Staphylococcus. The same authors also noted that, none of the studied isolates were able to stop mycelium from growing inside cells, however were all able to stop the extracellular growth of filaments from the examined bacteria [1].
Our results are in agreement with Odumosu et al. [40], who reported that various Streptomyces species demonstrated efficient antibacterial activity against a variety of human and food-borne diseases including Staphylococcus, Escherichia coli and Salmonella. The same authors used GC-MS to examine the secondary metabolites generated by the species under study. The analysis which revealed the presence of some bioactive substances with antibiotic properties, therefore a potential source of novel antibiotics. On the other hand, the bioactive secondary metabolites produced by Streptomyces strains with antifungal activity, such as isoikarugamycin, produced by Streptomyces zhaozhouensis against C. albicans, may be in charge of the antibacterial activity of these microorganisms [41].
The chemical analysis of the bioactive extracts revealed the presence of several components, including phenoxazinones and detoxin complex, to which the bioactivities could have In particular, chandrananimycin D, was reported in literature [42] for its antimicrobial activity. Valyldetoxinine, a member of detoxin complex, was distinguished by its remarkable detoxifying effect against the antibiotic blasticidin S, both in animal and plant cells [43]. Paromomycin, an antibiotic depsipeptide, can be used as biocontrol agent to suppress the soilborne diseases and as plant protection agent, in particular against Pectobacterium carotovorum, the responsible for bacterial soft rot producing pectolytic enzymes that hydrolyze pectin between individual plant cells, and against Phytophthora capsici, the responsible for blight and fruit rot of peppers and other important commercial crops [44].
Jizba et al. [45] reported stimulatory effects of nonactic and homonanctinic acids on Cucumis sativus growth: the substances were also considered later as pesticidal metabolites by Jizba and Skibova [46]. Li et al. [31] reported a moderate antifungal activity of maoxianamide A and B against S. sclerotiorum, the fungal phytopathogen responsible for white mold, ubiquitous and highly destructive disease. Also, rhizocticin A was reported for its effect on inhibition of Rhizoctonia solani, the fungal pathogen causing brown patch [47].
4. Materials and Methods
4.1. Actinomycetes isolation and indentification
The three studied actinomycetes used in this study, have been isolated from the rhizosphere of black locust (Robinia pseudoacacia L.) (Act1), rosemary (Rosmarinus officinalis L.) (Act2) and olive (Olea europaea L.) (Act3) at Potenza city (Basilicata region, Sothern Italy), following the Membrane Filter Technique [48]. The morphological identification of the three actinomycetes isolates has been carried out previously based on their microscopic features using light microscope (Axioskop—ZEISS, Oberkochen, Germany) and also by molecular methods-based PCR techniques as reported by Elshafie and Camele [12]. The isolates have been sub-cultured and conserved in peptone yeast calcium agar (PY-CA) nutrient media contains (g/L) peptone 5, yeast extract 3 and calcium chloride 0.7.
4.2. Plant growth-promoting & disease-control effects
A greenhouse-trial was undertaken out to evaluate the PGPE of the three isolates for basil as well as their disease-control effect (DC) against the necrotic stem lesions “black leg” caused by F. tabacinum. Basil seeds were surface sterilized with ethanol (70%), rinsed three times with sterile distilled water and then have been sowed in polystyrene seed-trays. The greenhouse's temperature and relative humidity were adjusted 24±2°C and 60-70 %, respectively, during the entire experiment.
Regarding actinomycetes treatment, a suspension 106 CFU.mL-1 of each isolate obtained from 5 days-fresh PY-Ca culture and inoculated into Minimal Mineral (MM) media [10.5 dipotassium phosphate, 4.5 potassium dihydrogen phosphate, 1.0 Ammonium sulfate, 0.5 trisodium Citrate Dihydrate, 0.2 Magnesium sulfate, 5.0 dextrose (g/L), pH 7] and left in Rotary-Incubator for 8 days at 180 rpm under constant temperature at 28°C. Broth culture (100 ml/pot) were poured into the basil-rhizosphere, 15 Days Post-Seed germination (DPSg).
For artificial fungal-infection, a conidial-suspension (108 spore/mL) of F. tabacinum was inoculated in potato dextrose broth (PDB) flask and incubated in agitation (180 rpm) for 7 days at 22 °C. Fifty mL broth was inoculated into the basil-rhizosphere 10 days after actinomycetes treatment. Twenty seedlings for each experiment i) untreated health; ii) treated only with actinomycetes; and iii) treated only with fungi, have been used.
At the end of the trial, plant growth was examined for the Eco-physiological characteristics 40 DPSg, following the method explained by Elshafie et al [49], going to measure stem length (SL), leaf number (NL), twigs number (NT), total shoot fresh-weight (TFwS) and total shoot dry-weight (TDwS). The disease incidence was monitored daily for 15 days post-infection (DPI), using the following scale: 0= less than 5 % symptomatic leaf; 1= 6 to 20 % of symptomatic leaf; 2= 21 to 50 % of symptomatic leaf; 3= 51 to 80 % of symptomatic leaf; 4 ≥ 80 % of symptomatic leaf [12]. Using Formula 1, the infection proportion (IP%) was calculated, whereas the Formula 2 and 3 were used for evaluating the disease-index (DI%) and control-effect (CE%), respectively [50].
| (1) | |
| (2) | |
| (3) |
Where: NSL= Number of Symptomatic Leaves; TLN= Total Leaf Number; Hi.S = Highest Scale; DI-P = Disease Index of Pathogen treatment; DI-B = Disease Index of actinomycetes-treated.
4.3. Extraction of metabolites
The secondary metabolites of the three studied strains were obtained following the method of Lavermicocca et al. [51] with little changes as follows: an Erlenmeyer-flask filled with 140 mL of MM broth and seeded with 2.0 mL of each actinomycete suspension at 107 CFU/mL and left in Rotary-Incubator for 8 days at 180 rpm under constant temperature at 28°C. The broth culture of each isolate was centrifuged at 20,000 g/10 min, the precipitate was discarded and the upper-phase was filtered using Millipore 0.22 μM. The purified filtrate was extracted in equal volume of suitable organic solvent (ethyl-acetate) and shacked for 5 min using a separator funnel. The combined organic fractions were concentrated by using an Eevaporator (Heidolf 2000, Germany) at 180 rpm/80°C for 20 min. The dried extracts were resuspended in 1 mL of sterile distilled water [52].
4.4. Microbicidal test
4.4.1. Antifungal
The in vitro antifungal activity of the extracted metabolites from the three studied isolates was evaluated against the following phytopathogenic fungi, Monilinia fructigena, Rhizoctonia solani, Fusarium oxysporum, Sclerotinia sclerotiorum, Penicillium expansum, P. digitatum and P. commune, using the incorporation method [53,54]. Ten μL of each extract, at concentration 100 and 50 %, was deposited on a Potato Dextrose Agar (PDA) [55] pre-inoculated with each tested fungal disc. To assess the antifungal activity, the diameter of mycelium grew in Millimetre. The inhibition percentage of fungal growth (FGI %) was calculated following Formula (4) [56]. Cycloheximide was used as positive control (C+ve) at 100 µg/mL.
where: FGI (%), mycelium inhibition percentage: D.MGt, mean diameter of fungal mycelium in treated Petri dish (mm); D.MGc, mean diameter of mycelium control Petri dish (mm).
4.4.2. Antibacterial assay
The in vitro antibacterial activity of the extracts was evaluated against the following pathogenic bacteria, Pseudomonas syringae, P. fluorescence, P. aeruginosa, E. coli, Bacillus cereus, B. megaterium and Xanthomonas campestris, using the disc diffusion method [57,58,59]. King’B (KB) nutrient media was used for reculturing the studied bacterial strains [60]. The bacterial suspensions were prepared in sterile distilled water (SDW) and adjusted at 106 CFU/mL. Four mL of each bacterial suspension diluted in soft agar (0.7 %) at 9:1 v/v, were poured into a KB Petri dish (90 mm). Fifteen μL of each extract, at concentration 100 and 50 %, was deposited on filter discs (6 mm-OXOID) previously placed on plates and left for 30 min under laminar flow. The eventually bactericidal effect was evaluated measuring the diameter of inhibition zone (D.Iz) in Millimetre compared to Tetracycline (1,600 µg/mL), used as C+ve. The bacterial growth inhibition percentage (BGI %) was calculated following Formula (5). All tested treatments were carried out in triplicates ± standard deviations (SDs).
where: BGI (%) represents the bacterial inhibition percentage; D.Iz, mean diameter of inhibition zone in treated Petri dish (mm); D.Cc, mean diameter of bacterial grown in control Petri dish (mm).
BGI (%) = D.Iz / D.Cc x 100
4.5. ESI/(HR)orbitrap/MS metabolic profiles
The qualitative analysis of three studied actinomycetes extracts was performed by High Performance Liquid Chromatography-Mass Spectrometry (HPLC-MS) using LTQ-XL Ion Trap mass spectrometer equipped with an Ultimate 3000 HPLC. Chromatographic separation was obtained using a Kinetex Polar C18 column (100 x 3.0 mm, 100Å, 2.6 µm). The injection volume was 0.5 mL/min and a mobile phase consisting of a combination of A (0.1% formic acid in water, v/v) and B (0.1% formic acid in acetonitrile MeCN); a linear gradient ranged between 5 to 60% B in 25 min, from 60 to 95 % B in 10 min, and held at 95 % B for 5 min was used. The mass spectrometer was set in positive ion mode. ESI source conditions were the following: capillary voltage −48 V; tube lens voltage −176.47; capillary temperature 280°C; sheath 15 and auxiliary gas flow (N2) 5, sweep gas 0, spray voltage 5. MS spectra were obtained, at 30 000 resolutions, by full-range acquisition with scan range between 150− 1500 m/z.
5. Conclusions
This research revealed the biological activity of actinomycetes especially Streptomyces and Arthrobacter. This study highlighted also the value of the new studied strains in terms of their capacity to produce significant bioactive by-products that can be employed as biocontrol agents against several serious fungi, including Fusarium species. In conclusion, some metabolites, detected in the extracted of the studied isolates, were recognized as effective against phytopathogenic fungi and bacteria, which may represent the potential for their uses in future sustainable strategies to control outbreaks in the vast range of crops. In fact, this research demonstrated the possibility of green-synthesis of some important secondary metabolites such as N-Acetyl-L-histidinol, rhizocticin A and Eponemycin from actinomycete. In particular, N-Acetyl-L-histidinol (detected in the current study from Streptomyces sp. and S. atratus), is considered an important derivative of the primary metabolite L-Histidinol, in agree with Ballio et al [13] who reported that N-acetyl-L-histidinol was produced in cultures of Streptomyces coelicolor. Regarding rhizocticin A (detected in the current study from Streptomyces sp. and S. atratus), is considered a natural phosphonate antibiotic and hydrophilic phosphono-oligopeptides extracted from different biocontrol agents such as Bacillus subtilis [47]. Eponemycin (detected in the current study from the three studied isolates) is considered an important antibiotic with specific in vivo antitumor effect against B16 melanoma [34]. Nevertheless, these potential bioactive compounds may be used to develop new commercial formulations either as plant-growth promoters or for crop protection.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Hazem Elshafie and Ippolito Camele; Data curation, Vincenzo De Feo; Formal analysis, Carmen Formisano; Funding acquisition, Ippolito Camele; Investigation, Laura De Martino, Lucia Caputo and Ippolito Camele; Methodology, Hazem Elshafie, Laura De Martino, Carmen Formisano and Lucia Caputo; Supervision, Vincenzo De Feo and Ippolito Camele; Writing – original draft, Hazem Elshafie; Writing – review & editing, Hazem Elshafie, Laura De Martino, Vincenzo De Feo and Ippolito Camele.
Funding
This study was carried out within the Agritech National Research Center and received funding from the European Union Next-GenerationEU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR) – MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4 – D.D. 1032 17/06/2022, CN00000022). This manuscript reflects only the authors’ views and opinions, neither the European Union nor the European Commission can be considered responsible for them.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chaudhary, H.S.; Yadav, J.; Shrivastava, A.R.; Singh, S.; Singh, A.K. , Gopalan, N. Antibacterial activity of actinomycetes isolated from different soil samples of Sheopur (A city of central India). J. Adv. Pharma. Technol. Res. 2013, 4, 118–23. [Google Scholar] [CrossRef]
- Vikineswary, S.; Nadaraj, P.; Wong, W.H.; Balabaskaran, S. . Actinomycetes from a tropical mangrove ecosystem – antifungal activity of selected strains. Asia Pac. J. Mol. Biol. Biotechnol. 1997, 5, 81–86. [Google Scholar]
- Jeffrey, L.S.H.; Norzaimawati, A.N.; Rosnah, H. Prescreening of bioactivities from actinomycetes isolated from forest peat soil in Sarawak. J. Trp. Agric. Fd. Sci. 2011, 39, 245–254. [Google Scholar]
- Manivasagan, P.; Kang, K.-H.; Sivakumar, K.; Li-Chan, E.C.; Oh, H.M.; Kim, S.K. Marine actinobacteria: an important source of bioactive natural products. Environ. Toxicol. Pharmacol. 2014, 38, 172–188. [Google Scholar] [CrossRef]
- Pepper, I.L.; Gentry, T.J. Chapter 4 – “Earth Environments” in: Environmental Microbiology (Third Edition) – 2015 pp. 59-88.
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s manual of determinative bacteriology (9th ed). Williams and Wilkins, Baltimore. 1994.
- Berdy, J. . Bioactive microbial metabolites. J. 2005Antibiotics 58, 1.
- Girão, M.; Ribeiro, I.; Ribeiro, T.; Azevedo, I.C.; Pereira, F.; Urbatzka, R.; Leão, P.N.; Carvalho, M.F. Actinobacteria Isolated from Laminaria ochroleuca: A Source of New Bioactive Compounds. Front. Microbiol. 2019, 10, 683, 1–13. [Google Scholar] [CrossRef]
- Simon, J.E. Basil. Center for New Crops & Plant Products, Department of Horticulture, Purdue University, West Lafayette, IN. 1998.
- Garibaldi, A.; Minuto, A.; Minuto, G.; Gullino, M.L. First Report of Downy Mildew on Basil (Ocimum basilicum) in Italy. Plant Dis. 2004, 88, 312. [Google Scholar] [CrossRef]
- Matta, A. Fusarium tabacinum (Beyma) W. Gams patogeno in natura su basilico e pomodoro. Rivista di Patologia Vegetale, S. 1978, IV, 14, 119-125.
- Elshafie, H.S.; Camele, I. Rhizospheric actinomycetes revealed antifungal and plant-growth-promoting activities under controlled environment. Plants 2022, 11, 1872. [Google Scholar] [CrossRef]
- Ballio, A.; Russi, S.; Vlasić, D. Isolation and identification of N-acetyl-L-histidinol in cultures of mutants of Streptomyces coelicolor requiring histidine. Annali Dell'istituto Superiore di Sanita 1966, 2(4), 523–530. [Google Scholar]
- Seligman, SJ. Frequency of resistance to kanamycin, tobramycin, netilmicin, and amikacin in gentamicinresistant gram-negative bacteria. Antimicrob Agents Chemother. 1978, Jan;13(1):70-3. PMID: 626492; PMCID: PMC352186. [CrossRef]
- He, F.; Takahiro, M.; Iori, M.; Hitomi, N.; Miroslava, A.; Shotaro, H.; Takayoshi, A.; Ikuro, A. Molecular basis for the P450-catalyzed C-N bond formation in indolactam biosynthesis. Nature Chem. Biol. 2019, 15, 1206–1213. [Google Scholar] [CrossRef]
- Nithya, K.; Muthukumar, C.; Biswas, B.; Alharbi, N.S.; Kadaikunn, S.; Khaled, J.M.; Dhanasekaran, D. Desert actinobacteria as a source of bioactive compounds production with a special emphasis on Pyridine-2,5-diacetamide a new pyridine alkaloid produced by Streptomyces sp. DA3-7. Microbiol. Res 2018, 207, 116–133. [Google Scholar] [CrossRef]
- Li, W.R.; Han, S.Y.; Joullie, M.M. Total synthesis of (+)-valyldetoxinine and (-)-detoxin D1. Tetrahedron 1993, 49, 785–802. [Google Scholar] [CrossRef]
- El-Housseiny, G.S.; Ibrahim, A.A.; Yassien, M.A.; Aboshanab, K.M. Production and statistical optimization Paromomycin by Streptomyces rimosusNRRL 2455 in solid state fermentation. BMC Microbiol. 2021, 21, 34. [Google Scholar] [CrossRef]
- Cho, R.M.; Kogan, H.V.; Elikan, A.B.; Snow, J.W. Paromomycin Reduces Vairimorpha (Nosema) ceranae Infection in Honey Bees but Perturbs Microbiome Levels and Midgut Cell Function. Microorganisms 2022, 10, 1107. [Google Scholar] [CrossRef]
- Furukawa, T.; Akutagawa, T.; Funatani, H.; Uchida, T.; Hotta, Y.; Niwa, M.; Takaya, Y. Cyclic dipeptides exhibit potency for scavenging radicals. Bioorganic & Med. Chem. 2012, 20, 2002–2009. [Google Scholar]
- Simon, G.; Berube, C.; Voyer, N.; Grenier, D. Anti-biofilm and anti-adherence properties of novel cyclic dipeptides against oral pathogens. Bioorganic & Medicinal Chem. 2019, 27, 2323–2331. [Google Scholar]
- Tatsuta, K.; Tsuchiya, T.; Umezawa, S.; Naganawa, H.; Umezawa, H. Revised structure for arglecin. J. Antibiotics 1972, 25, 674–676. [Google Scholar] [CrossRef]
- Kumar, P.; Ryan, B.; Henehan, G.T.M. b-Glucosidase from Streptomyces griseus: Nanoparticle immobilization and application to alkyl glucoside synthesis. Protein Exp. Purif. 2017, 132, 164–170. [Google Scholar] [CrossRef]
- Lu, S.; Hu, J.; Xie, X.; Zhou, R.; Li, F.; Huang, R.; He, J. Secondary Metabolites with Cytotoxic Activities from Streptomyces sp. BM-8 Isolated from the Feces of Equusquagga. Molecules 2021, 26, 24–7556. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chen, Y.K.; Wang, J.J.; Hsu, C.C.; Tsai, F.Y.; Sung, P.J.; Lin, H.C.; Chang, L.S.; Hu, W.P. DC-81-enediyne induces apoptosis of human melanoma A375 cells: involvement of the ROS, p38 MAPK, and AP-1 signaling pathways. Cell Biol. Toxicol. 2013, 29, 85–99. [Google Scholar] [CrossRef]
- Shiomi, K.; Arai, N.; Shinose, M.; Takahashi, Y.; Yoshida, H.; Iwabuchi, J.; Tanaka, Y.; Omura, S. New antibiotics phthoxazolins B, C and D produced by Streptomyces sp. KO-7888. J. Antibiot. 1995, 48(7), 714–9. [Google Scholar] [CrossRef]
- Gomes, P.B. : Nett, M.; Dahse, H.M.; Hertweck, C.P. Structural merger of a phenoxazinone with an epoxyquinone antibiotic. J. Nat. Prod. 2010, 73, 1461–1464. [Google Scholar] [CrossRef]
- Abdelfattah, M.S. A new bioactive aminophenoxazinone alkaloid from a marine-derived actinomycete. Nat. Prod. Res. 2013, 27, 2126–2131. [Google Scholar] [CrossRef]
- Chatterjee, S.; Vijayakumar, E.K.; Franco, C.M.; Maurya, R.; Blumbach, J.; Ganguli, B.N. Phencomycin, a new antibiotic from a Streptomyces species HIL Y-9031725. J. Antibiot. 1995, 8(11), 1353-4. [Google Scholar] [CrossRef]
- Hidetsugu, N.; Hitoshi, E.; Koji, K.; Okumura, K. Biological method of producing serine and serinol derivatives. U.S. Patent No 3871958A. Washington, DC: U.S. Patent and Trademark Office. 1975.
- Li, J.M.; Yan, K.; Zhang, H.; Qi, H.; Zhang, J.; Xiang, W.S.; Wang, J.D.; Wang, X.J. New aliphatic acid amides from Streptomyces maoxianensis sp. nov. J. Antibiotics 2017, 70, 187–189. [Google Scholar] [CrossRef]
- Gahungu, M.; Arguelles-Arias, A.; Fickers, P.; Zervosen, A.; Joris, B.; Damblon, C.; Luxen, A. Synthesis and biological evaluation of potential threonine synthase inhibitors: Rhizocticin A and Plumbemycin A. Bioorganic & Med. Chem. 2013, 21, 4958–4967. [Google Scholar]
- Fredenhagen, A.; Angst, C.; Peter, H.H. Digestion of rhizocticins to (Z)-L-2-amino-5-phosphono-3-pentenoic acid: revision of the absolute configuration of plumbemycins A and B. J. Antibiotics 1995, 48, 1043–1045. [Google Scholar] [CrossRef]
- Sugawara; K. ; Hatori M.; Nishiyama, Y.; Tomita, K.; Kamei, H.; Konishi, M., Oki, T. Eponemycin, a new antibiotic active against B16 melanoma. I. Production, isolation, structure and biological activity. J. Antibiot. (Tokyo). 1990, 43(1), 8–18. [Google Scholar]
- Fitri, L.E.; Cahyono, A.W.; Nugraha, R.Y.B.; Alkarimah, A.; Ramadhani, N.N.; Laksmi, D.A.; Trianty, L.; Noviyanti, R. Analysis of eponemycin (α'β' epoxyketone) analog compound from Streptomyces hygroscopicus subsp Hygroscopicus extracts and its antiplasmodial activity during Plasmodium berghei infection. Biomed. Res. 2017, 28, 164–172. [Google Scholar]
- Suwan, N.; Wassamon, B.; Sarunya, N. Antifungal activity of soil actinomycetes to control chilli anthracnose caused by Colletotrichum gloeosporioides. J. Agric. Technol. 2012, 8, 725–737. [Google Scholar]
- Sacramento, D.R.; Coelho, R.R.R.; Wigg, M.D.; Wigg, M.D. , Linhares, L.F.T.; Santos, M.G.M.; Semêdo, L.T.A.; Silva, A.J.R. Antimicrobial and antiviral activities of an actinomycete (Streptomyces sp.) isolated from a Brazilian tropical forest soil. World J. Microbiol. Biotechnol. 2004, 20, 225–229. [Google Scholar] [CrossRef]
- Prapangdee, B.; Kuekulvong, C.; Mongkolsuk, S. Antifungal Potential of Extracellular Metabolites Produced by Streptomyces hygroscopicus against Phytopathogenic Fungi. Int. J. Biol. Sci. 2008, 4, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Solans, M.; Vobis, G.; Cassa´n, F.; Luna, V.; Wall, L.G. Production of phytohormones by root-associated saprophytic actinomycetes isolated from the actinorhizal plant Ochetophila trinervis. World J. Microbiol. Biotechnol. 2011, 27, 2195–2202. [Google Scholar] [CrossRef]
- Odumosu, B.T.; Buraimoh, O.M.; Okeke, C.J.; Ogah, J.O.; Michel, Jr. F.C. ; Antimicrobial activities of the Streptomyces ceolicolor strain AOBKF977550 isolated from a tropical estuary. J. Taibah Univer. Sci. 2017, 11, 836–841. [Google Scholar] [CrossRef]
- Lacret, R. , Oves-Costales, D.; Gómez, C.; Díaz, C.; de la Cruz, M.; Pérez-Victoria, I.; Vicente, F.; Genilloud, O.; Reyes, F. New ikarugamycin derivatives with antifungal and antibacterial properties from Streptomyces zhaozhouensis. Mar. Drugs 2014, 13, 128–140. [Google Scholar] [CrossRef]
- Zhang, X.M.; Liu, X.; Wang, Z.; Tian, Z.H.; Xie, W.D. Viridobrunnines A B, antimicrobial phenoxazinone alkaloids from a soil associated Streptomyces sp. Heterocycles 2015, 91(9), 1809–1814. [Google Scholar]
- Ogita, T.; Seto, H.; Ōtake, N.; Yonehara, H. The Structures of Minor Congeners of the Detoxin Complex. Agricul. Biol. Chem. 1981, 45, 2605–2611. [Google Scholar] [CrossRef]
- Park, J.W.; Lee, S.W. , Balaraju, K.; Kim, C.J.; Park, K. Chili - Pepper Protection from Phytophthora capsici and Pectobacterium carotovorum SCC1 by Encapsulated Paromomycin Derived from Streptomyces sp. AMG-P1. J. Pure & Appl. Microbiol. 2012, 6, 1517–1522. [Google Scholar]
- Jizba, J.; Pfukrylova, V.; Ujhelyova, L.; Varkonda, S. Insecticidal properties of nonactic acid and homononactic acid, the precursors of macrotetrolide antibiotics. Folia Microbiol. 1992, 37, 299–303. [Google Scholar] [CrossRef]
- Jizba, J.; Skibova, I. Regulation of biosynthesis of pesticidal metabolic complexes in Streptomyces griseus. Folia Microbiologica 1994, 39, 119–28. [Google Scholar] [CrossRef]
- Kugler, M.; Loeffler, W.; Rapp, C.; Kern, A.; Jung, G. Rhizocticin A, an antifungal phosphono-oligopeptide of Bacillus subtilis ATCC 6633: biological properties. Arch Microbiol. 1990, 153(3), 276–81. [Google Scholar] [CrossRef]
- Njenga, W.P.; Mwaura, F.B.; Wagacha, J.M.; Gathuru, E.M. Methods of Isolating Actinomycetes from the Soils of Menengai Crater in Kenya. Arch Clin Microbiol. 2017, 8, 3:45. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Sakr, S.; Bufo, S.A.; Camele, I. An attempt of biocontrol the tomato-wilt disease caused by Verticillium dahliae using Burkholderia gladioli pv. agaricicola and its bioactive secondary metabolites. Int. J. Plant Biol. 2017, 8, 57–60. [Google Scholar]
- Lee, K.J.; Kamala-Kannan, S.; Sub, H.S.; Seong, C.K.; Lee, G.W. World J. Microbiol. Biotecnol. 2008, 24, 1139–1145. [CrossRef]
- Lavermicocca, P.; Iacobellis, N.S.; Simmaco, M.; Graniti, A. Biological properties and spectrum of activity of Pseudomonas syringae pv. syringae toxins. Physiol. Mol. Plant Pathol. 1997, 50, 129–140. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Viggiani, L.; Mostafa, M.S.; El-Hashash, M.A.; Bufo, S.A.; Camele, I. Biological activity and chemical identification of ornithine lipid produced by Burkholderia gladioli pv. agaricicola ICMP 11096 using LC-MS and NMR analyses. J. Biol. Res. 2017, 90, 96–103. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Amato, M.; De Feo, V.; Camele, I. Chemical composition and antimicrobial activity of Chia (Salvia hispanica l.) Essential oil. Europ. Food Res. Technol. 2018, 244, 1675–1682. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Gruľová, D.; Baranová, B.; Caputo, L.; De Martino, L.; Sedlák, V.; Camele, I.; De Feo, V. Antimicrobial activity and chemical composition of essential oil extracted from Solidago canadensis L. growing wild in Slovakia. Molecules 2019, 24, 1206. [Google Scholar] [CrossRef]
- Aryal, S. Potato Dextrose Agar (PDA)- Principle, Uses, Composition, Procedure and Colony Characteristics. Microbiol. Info. Com. 2018.
- Zygadlo, J.A.; Guzman, C.A.; Grosso, N.R. Antifungal properties of the leaf oils of Tagetes minuta L. and Tagetes filifolia Lag. J. Essent. Oil Res. 1994, 6, 617–621. [Google Scholar] [CrossRef]
- Bhunia, A.; Johnson, M.C. , Ray, B. Purification, characterization and antimicrobial spectrum of a bacteriocin produced by Pediococcus acidilactici. J. Appl. Bacteriol. 1988, 65, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Camele, I.; Elshafie, H.S.; Caputo, L.; Sakr, S.H.; De Feo, V. Bacillus mojavensis: Biofilm formation and biochemical investigation of its bioactive metabolites. J. Biol. Res. 2019, 92, 39–45. [Google Scholar] [CrossRef]
- Gruľová, D.; Caputo, L.; Elshafie, H.S.; Baranová, B.; De Martino, L.; Sedlák, V.; Camele, I.; De Feo, V. Thymol chemotype Origanum vulgare L. essential oil as a potential selective bio-based herbicide on monocot plant species. Molecules 2020, 25(3), 595. [Google Scholar] [CrossRef]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
Figure 1.
Microscopic-morphological features of the three studied actinomycetes isolates. A,C,E are cultures of Streptomyces sp. (Act1), Streptomyces atratus (Act2) and Arthrobacter humicola (Act3) grown in PY-CA media, respectively. B,D,F are the colonies of Streptomyces sp. (Act1), Streptomyces atratus (Act2) and Arthrobacter humicola (Act3), respectively examined under stereo-Microscope (1000x).
Figure 1.
Microscopic-morphological features of the three studied actinomycetes isolates. A,C,E are cultures of Streptomyces sp. (Act1), Streptomyces atratus (Act2) and Arthrobacter humicola (Act3) grown in PY-CA media, respectively. B,D,F are the colonies of Streptomyces sp. (Act1), Streptomyces atratus (Act2) and Arthrobacter humicola (Act3), respectively examined under stereo-Microscope (1000x).
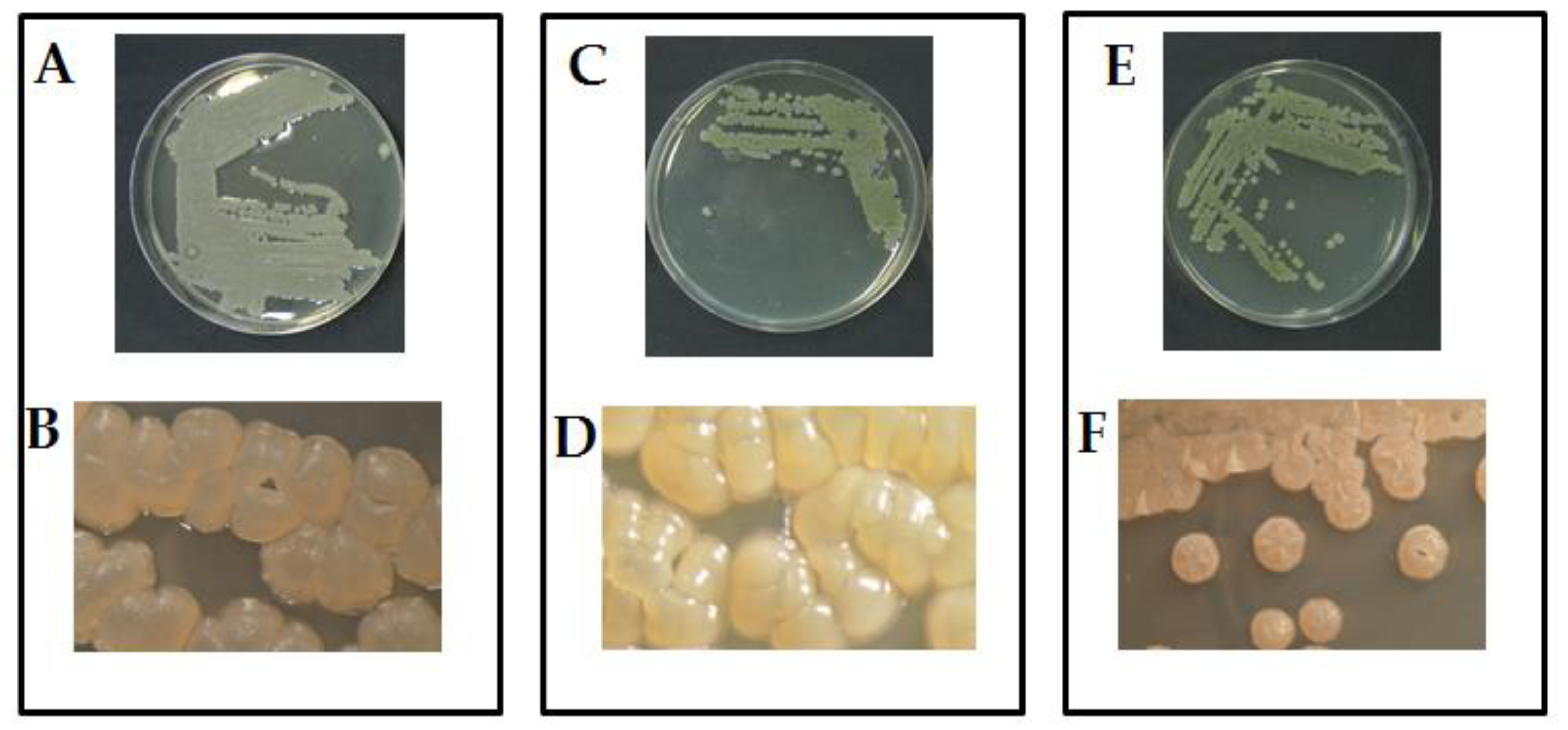
Figure 2.
Disease-index of basil inoculated with F. tabacinum. Where: Act1+F; Act2+F; Act3+F are inoculated plants with Streptomyces sp., A S. atratus and humicola, respectively. Bars with different letters between different treatments are significantly different at p < 0.05 according to SPSS software. values are mean of 3 replicates ± SDs.
Figure 2.
Disease-index of basil inoculated with F. tabacinum. Where: Act1+F; Act2+F; Act3+F are inoculated plants with Streptomyces sp., A S. atratus and humicola, respectively. Bars with different letters between different treatments are significantly different at p < 0.05 according to SPSS software. values are mean of 3 replicates ± SDs.
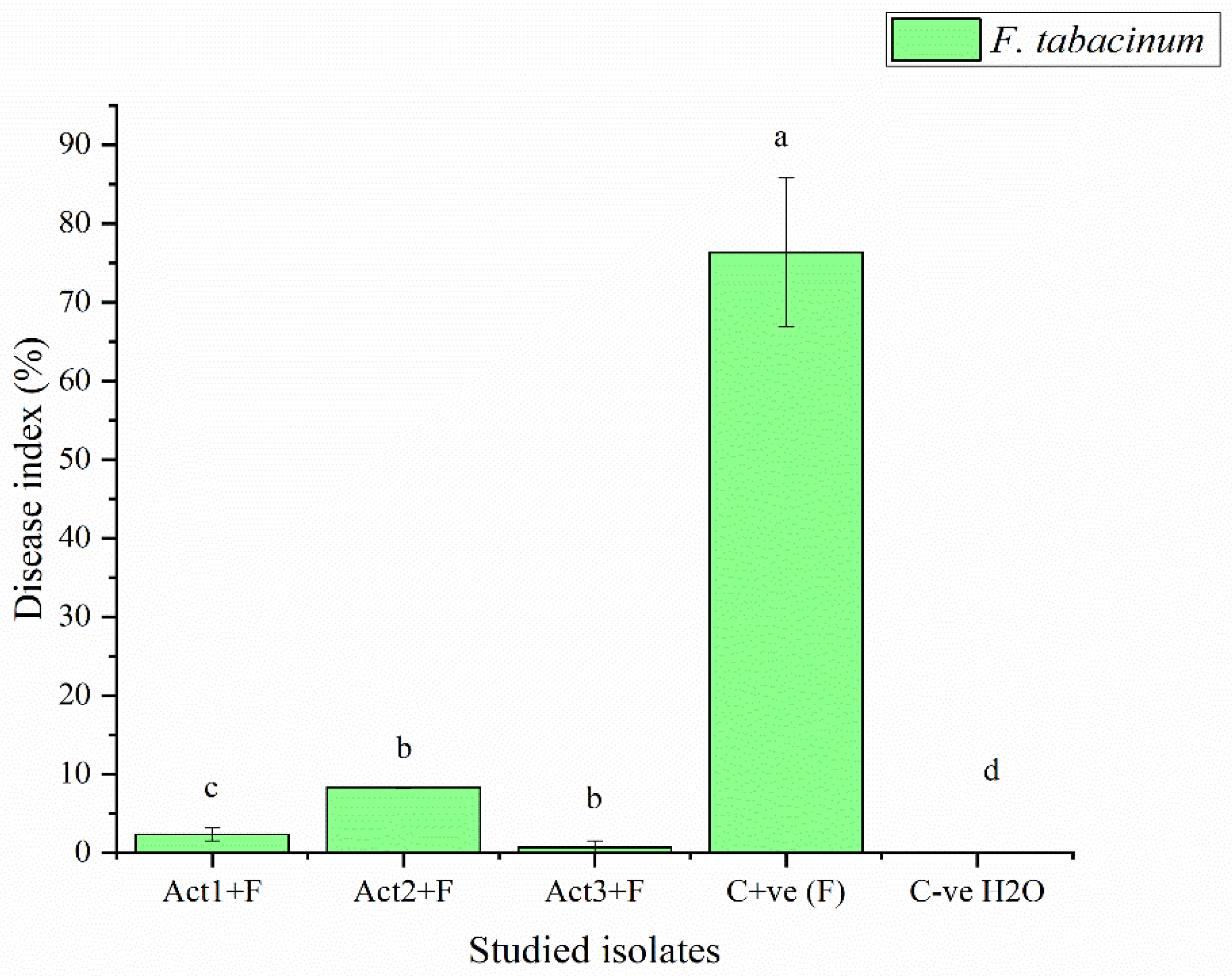
Figure 3.
Control effect of basil after infection with pathogen. Bars with different letters between different treatments are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
Figure 3.
Control effect of basil after infection with pathogen. Bars with different letters between different treatments are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
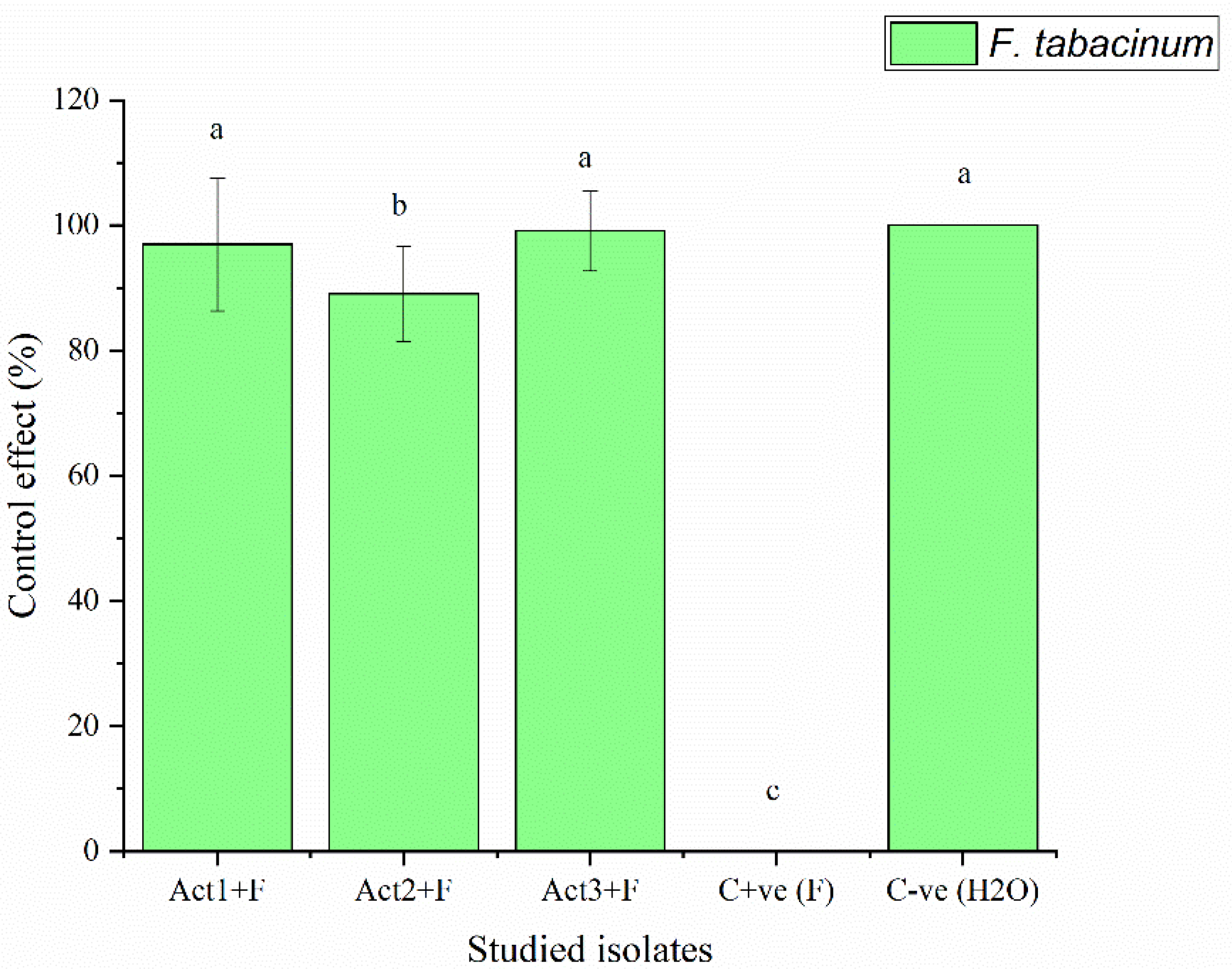
Table 1.
Eco-physiological characteristics of basil post-actinomycetes inoculation (healthy plant).
| Actinomycetes isolates |
Eco-Physiological Characteristics | ||||
|---|---|---|---|---|---|
| NL (n) | NT (n) | SL (cm) | TFwS (g) | TDwS (g) | |
| Control | 89±4b | 8±2a | 27±2b | 85±12b | 5±2c |
| Act1: Streptomyces sp. | 113±12a | 4±2b | 39±4a | 114±15a | 23±2a |
| Act2: A. humicola | 105±5a | 7±1a | 41±6a | 99±9b | 18±6a |
| Act3: S. atratus | 85±7b | 3±1b | 27±2b | 111±27a | 14±2b |
NL: number of leaves; NT: number of twigs; SL: shoot length; TFwS: total fresh weight of shoot; TDwS: total dry
weight of shoot. Values followed by different letters in each vertical column are significantly different at p < 0.05
according to SPSS software. Data are expressed as mean of 3 replicates ± SDs.
Table 2.
Eco-physiological characteristics of basil post-actinomycetes inoculation and (infected plants).
Table 2.
Eco-physiological characteristics of basil post-actinomycetes inoculation and (infected plants).
| Actinomycetes isolates |
Eco-Physiological Characteristics | ||||
|---|---|---|---|---|---|
| NL (n) | NT (n) | SL (cm) | TFwS (g) | TDwS (g) | |
| Cont -ve | 85±9c | 2±1ab | 34±4b | 75±4c | 15±2b |
| Act1: Streptomyces sp. | 146±7a | 7±1a | 45±5a | 241±13a | 40±4a |
| Act2: A. humicola | 111±10b | 5±1a | 47±3a | 246±7a | 31±4ab |
| Act3: S. atratus | 109±9b | 3±1ab | 34±4b | 191±5b | 20±3b |
NL: number of leaves; NT: twigs count; SL: stem length; TFwS: total fresh weight of shoot; TDwS: total dry weight of shoot. Values followed by different letters in each column are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
Table 3.
Determination of disease-index and control-effect on basil after different applications.
| Treatments | Disease-index | Control-effect |
|---|---|---|
| DI % | CE % | |
| Act1: Streptomyces sp. | 2.3±0.8c | 96.9±10.6a |
| Act2: A. humicola | 0.7±0.1d | 99.1±7.6a |
| Act3: S. atratus | 8.3±0.9b | 89.1±6.4ab |
| Cont. +ve (F) | 76.4±9.5a | 0.0±0.0c |
| Cont. -ve (H2O) | 0.0±0.0e | 100.0±0.0a |
Where: Cont. +ve (F): control positive (treated plants fungal pathogens); Cont. -ve (H2O): control negative (healthy plants). DI% = disease-index; CE% = control-effect. Values followed by different letters in each column are significantly different at p < 0.05 according to SPSS software. Values are mean of 3 replicates ± SDs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



