
Submitted:
29 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
The genus Beauveria include important entomopathogenic and endophytic fungi, among them, Beauveria bassiana is the most studied species. However, there are few knowledge regarding their antimicrobial activity. The current research has been conducted to evaluate in vitro antagonistic activity of B. bassiana and the antimicrobial efficacy of its Exo and Endo metabolites against Bacillus cereus, B. megaterium, Clavibacter michiganensis (gram positive bacteria, G+ve), Xanthomonas campestris, Pseudomonas aeruginosa and P. fluorescence (gram negative bacteria, G-ve). In addition, Solid-phase microextraction (SPME) was coupled to Gas Chromatography-Mass Spectrometry (GC/MS) to qualitatively measure the volatile organic compounds (VOCs) metabolic profile of the most efficient studied isolate of B. bassiana. The obtained results showed that, the isolate UniB2439-3 has promising antibacterial effect against most of studied target bacteria. SPME-GC/MS analysis of VOCs revealed the presence of ethanol; butanal,2-methyl; 2,4-dimethyl-1-heptene; octane, 4-methyl and β-elemene as the main dominant bioactive compounds. The outgoing results explicated that the efficient isolate of B. bassiana can be potentially used as a biocontrol agent against several bacteria especially G+ve ones.
Keywords:
biocontrol
; natural products
; phytopathogens
; microbial metabolites
; entomopathogens
; endophytic fungi.
1. Introduction
Genus Beauveria include entomopathogenic and endophytic fungi which are widely spread in different habitats [1,2,3]. Furthermore, many researchers reported that fungi in the genus Beauveria can produce enzymes for biotransformation and biodestructors [4,5]. On the other hand, the genus Beauveria is considered also a human pathogen where it causes opportunistic infections especially in patients with weak immune systems [6].
Some species of this genus, such as B. bassiana and B. brongniartii are able to produce mycoinsecticides [7]. B. bassiana is also a beneficial microorganism (BM) and endophytic fungi (EF) in several crops and is commonly known as biological control agent against a variety of agricultural pests [3,8,9,10]. The application of B. bassiana has many advantages such as being an eco-friendly management compared to chemical pesticides where it is harmless to human health [8,10,11,12]. For decades, several scientists have reported the importance of B. bassiana in reducing a range of nuisance insects, where it can induce direct insect mortality [2,13,14] and can also reach 90% reduction of life-time fecundity [15].
A recent study, conducted by Barra-Bucarei et al. [16] to evaluate the colonization ability of native endophyte of different strains of B. bassiana and their antifungal effect against Botrytis cinerea in tomato and chili pepper concluded that all studied strains had significant in vitro antagonism against B. cinerea. The same study reported that, the native strains of B. bassiana were able to colonize tomato and chili pepper tissues and provided important levels of antagonism [16].
Another research conducted by Sinno et al [10] has evaluated different isolates of B. bassiana as plant-growth promoting (PGP) and protective agent for tomato plants against B. cinerea, Alternaria alternata and the pest aphis, Macrosiphum euphorbiae. The results showed that some studied isolates were able to control the two phytopathogens, whereas one isolate was also able to promote plant growth [10]. The antibacterial activity of crude ethyl acetate extract of B. bassiana against some aerobic pathogenic bacteria been tested by Parine et al [17]. The results explicated that the extract of B. bassiana possess a strong inhibiting activity against many of tested species especially Bacillus megaterium, B. subtilis, B. sphaericus and Escherichia coli [17]. Whereas, it showed a moderate effect against Micrococcus luteus, Pseudomonas aeruginosa and low effect against Streptococcus pyogenes and Chromobacterium violaceum [17]. In another study, the application of conidia of B. bassiana has protected tomato seedlings from damping-off disease caused by the soil-borne pathogen Rhizoctonia solani [18].
Recently, there is a huge interest for discovering natural substances-based plant or microbe origins having herbicidal and/or pesticide effect [19,20]. However, the new discovered natural substances should be evaluated for safety to avoid any possible negative health impact [21,22]. In addition, the discovery of possible natural alternatives for reducing the excessive use of synthetic chemicals, decreasing the environmental hazards and avoiding the appearance of new resistance microbial strains to common microbicide compounds should be highly considered [18,23].
There is few information regarding the bioactive metabolites produced by B. bassiana either diffusible or volatiles and also their mechanism of action in the antimicrobial activity and plant growth promotion effect. A recent study conducted by Wang et al [24] reported that B. bassiana produces a variety of toxins such as beauvericin, bassianin, bassianolide, beauverolides, tenellin, oosporein and oxalic acid, which give the ability to B. bassiana to colonize, parasitize and kill the host tissues. Therefore, the precise chemical characterization and determination of the main bioactive single substances of B. bassiana will certainly aid in understanding its biological importance. In addition, the details of chemical constituents of B. bassiana will undoubtedly serve for various applications such as control plant diseases, taking in consideration the heavy reliance on chemicals that are extremely harmful to environment as well as plants, animals and human health. Furthermore, there are many studies have been carried out regarding the insecticidal effect of B. bassiana, but there are few studies undersigned their antifungal or antibacterial effect.
The main objective of this research is to study the chemical composition of the principal volatile organic compounds (VOCs) of B. bassiana. Hence, the chemical composition of B. bassiana metabolites will aid in the detection and differentiation of this species from others. The full identification of metabolite profile can help of its utilization in industrial, agricultural and pharmaceutical fields. In addition, in this research we will expand the possible benefits of B. bassiana against new non-reported target phytopathogens. In particular, the aims of the current research were to i) evaluate the antagonistic activity of five isolates of B. bassiana against some bacteria; ii) investigate in vitro antimicrobial activity of diffusible and volatile metabolites produced by the most efficient isolate; iii) chemically characterize of VOCs produced by the most efficient isolate using SPME-GC/MS analysis.
2. Results
2.1. Molecular identification of the studied isolates of Beauveria
The PCR amplification with Bt2a/Bt2b produced, for each gDNA extracted from the above five isolates (UniB2439-1, UniB2439-2, UniB2439-3, UniB2439-4 and UniB2439-5), amplicons with molecular weight about 330 bp. No amplification was observed in case of the negative control. The amplicons were directly sequenced (BMR Genomics, Padova, Italy) and the obtained sequences were compared with those available in GenBank nucleotide archive showing a high similarity percentage (97,29 %) with the following sequences AB829899, AB829898 and CP045886.1 available in the NCBI database using Basic Local Alignment Search Tool software BLAST (Bethesda, Rockville Pike, MD, USA) [25]. The five obtained sequences were deposited in the NCBI GeneBank with accession numbers FR989662 to FR989666.
2.2. Antagonistic activity of B. bassiana isolates
The preliminary results showed that, all tested isolates of B. bassiana, had antagonistic effect against most of tested bacterial strains as illustrated in Table (S1). Particularly, UniB2439-3 was the most efficient isolate. In fact, this isolate showed the highest significant effect against Bacillus cereus and Clavibacter michiganensis, moderate effect against B. megaterium and low effect against Xanthomonas campestris and Pseudomonas fluorescens. Whereas, this isolate did not show any activity against P. aeruginosa (Figure 1). Therefore, UniB2439-3 isolate was selected for further biological and chromatographic analyses.
2.3. Antimicrobial activity of Exo- and Endo- diffusible metabolites
The obtained results of the extracted metabolites from the selected isolate of B. bassiana UniB2439-3 showed that, the Extracellular (Exo-ME) was able to inhibit the growth of most tested bacterial strains higher than the Endocellular (Endo-ME) (Table 1). In particular, Exo-ME showed the highest significant activity against C. michiganensis. In addition, both extracts showed equal activity against X. campestris. Whereas, only Endo-ME showed antibacterial activity against P. aeruginosa. On the other hand, Endo-ME was not active against B. cereus and C. michiganensis. Although both extracts were lower efficient than the control (tetracycline), but they can be considered promising and optimistic antimicrobial agents, being natural biopesticides can be potential alternatives to chemical and synthetic antibiotics.
2.4. Antibacterial activity of volatiles metabolites
The in vitro antibacterial activity of eventually produced volatile metabolites from B. bassiana has been evaluated against either the grown-visible colonies (GVC) or aqueous suspension (AQS) of each tested bacterial strain. Results of in vitro bioactivity assay demonstrated that the studied isolate of B. bassiana (UniB2439-3) produced bioactive volatile metabolites which were able to significantly reduce the growth of tested bacterial strains compared to tetracycline (positive control). In particular, the efficacy of produced volatiles substances was highly active against AQS of all tested bacterial strains higher than GVC (Table 2). In addition, the highest antibacterial activity was observed in case of GVC against B. megaterium (G+ve) and P. fluorescens (G-ve) estimated by 77.5 and 52.5 %, respectively. On the other hand, the highest antibacterial activity in case of AQS was observed against B. megaterium (G+ve) and P. aeruginosa (G-ve) estimated by 92.0 and 87.5 %, respectively.
2.5. SPME-GC/MS analysis of VOCs
GC-MS analysis of the VOCs produced by B. bassiana UniB2439-3 was illustrated in Figure (S3). In particular, the dominant principal compounds, listed in Table (2), are: i) ethanol (Figure S4), ii) butanal,2-methyl (Figure S5), iii) 2,4-dimethyl-1-heptene (Figure S6), iv) octane, 4-methyl (Figure S7) and v) β-elemene (Figure S8) among the total 32 detected volatile compounds (Table S2). In particular, the most abundant constituents were ethanol and β-elemene with relative areas 4.69 and 6.98 %, respectively.
The eventual fragmentation of the acquired volatile metabolites, as described here, is also shown in Table (S2). Beauvericin was fragmented into diethyl phthalate with 90%, the major dominant constituent, carbon dioxide and nitrous oxide. Whereas, bassianolide was converted to butanal, 3-methyl with 81% or butanol, 3-methyl with 83%. Regarding bassianin, GC-MS analysis showed that this compound was converted into 2,4-Dimethyl-1-heptene with 90%. Whereas, beauveriolide was fragmented into butanal, 3-methyl with 81%, carbon dioxide and nitrous oxide. Regarding cyclosporine, results demonstrated that this compound was fragmented into butanal, 2-methyl- (90%), butanal, 3-methyl- (81%), 1-butanol, 3-methyl- (83%), carbon dioxide and nitrous oxide.

Table 2.
SPME-GC/MS analysis of VOCs extracted from B. bassiana UniB2439-3.
| RT (min) |
Area (%) |
Name | M.Wt (g/mol) |
Formula | CAS | Probability of identification (%) |
|---|---|---|---|---|---|---|
| 1.576 | 4.69 | Ethanol | 46,07 | C2H5OH | 000064-17-5 | 90 |
| 2.834 | 0.44 | Butanal, 2-methyl | 86 | C5H10O | 000096-17-3 | 90 |
| 5.372 | 00.63 | 2,4-Dimethyl-1-heptene | 126.24 | C9H18 | 019549-87-2 | 90 |
| 5.660 | 1.99 | Octane, 4-methyl | 128.25 | C9H20 | 002216-34-4 | 93 |
| 10.459 | 6.98 | β-elemenea | 204.35 | C15H24 | 000515-13-9 | 96 |
a. β-elemene: Cyclohexane, 1-ethenyl-1-methyl-2,4-bis(1-methylethenyl)-, [1S-(1.alpha.,2.beta.,4.beta.)]. For confirmation the obtained results, this experiment of GC-MS analysis of VOCs was carried out twice with three replicates (different injections). .
3. Discussion
Several research have been conducted recently to overcome the multi-drug resistant (MDR) microorganisms to different antibiotics and chemotherapeutic agents [26]. Hence, the search for new active and natural agents gets a great interest, particularly, for human health and environmental protection [27]. Beauveria, one of the most studied genera among entomopathogenic fungi, has various biological applications as growth promoting agent or insecticides [9,10,28,29]. The capacity of B. bassiana to produce several volatiles bioactive metabolites with promising antimicrobial properties is in agreement with the previous bibliographic research who investigate the antagonistic effect against several phytopathogens [9,17,30]. In fact, the bibliographic research revealed that the genus Beauveria produced some interesting metabolites such as oosporein, beauvericin, bassianolide, bassianin, beauveriolide, bassiacridin and cyclosporine which having notable insecticide and antimicrobial actions [31,32]. On the other hand, some of the produced metabolites from B. bassiana could contain interesting cell-wall hydrolytic enzymes which are able to degrade microbial cell wall and hence inhibition the growth of several phytopathogens [33].
On the other hand, Barra-Bucarei et al [16] studied the antifungal activity of ten native strains of B. bassiana, endophyte for tomato and chili pepper and observed that the majority of studied native strains were able to colonize tomato and chili pepper tissues and showed promising antagonistic effect against B. cinerea.
The obtained results of GC/MS analysis of VOCs metabolites extracted from B. bassiana UniB2439-3 are in agree with many bibliographic research which reported that oosporein, beauvericin, bassianolide, bassianin, beauveriolide, bassiacridin and cyclosporine are the most abundant metabolites produced by B. bassiana [34,35,36]. Several studies revealed that beauvericin and oosporein evidenced remarkable antibiotic and antifungal properties [37,38] which are probably involved in the microbial growth inhibition observed in the bioassay presented in this study. Furthermore, Wang and Xu [39] reported that beauvericin was one of the active constituents of B. bassiana and confirmed to have antimicrobial activity and anti-tumor effect especially against human leukemia. In another study, conducted by Manning and Wyatt [40], the results demonstrated that oosporein, extracted from the broth cultures of Beauveria and Chaetomium, has been identified as a toxic substance for plants and poultry.
Regarding the β-elemene, our obtained results detected the presence of an important sesquiterpene compound identified as β-elemene (cyclohexane, 1-ethenyl-1-methyl-2,4-bis(1-methylethenyl), 1S-(1.alpha.,2.beta.,4. beta.), among the detected VOCs substances from the studied Beauveria isolate. This compound (β-elemene) was identified for the first time in 1994 in the dry rhizome extract from Curcuma phaeocaulis, C. kwangsinensis and C. wenyujing [32]. In addition, β-elemene is also one of the common constituents of several aromatic essential oils extracted from Proteus vulgaris [41]. β-elemene was also found in wild hops from Lithuania at levels up to 14% [42] and in notable amounts in the medical cannabis cultivar 'bedropuur' [43]. The same compound has notable antimicrobial activity against different pathogens including Mycobacterium tuberculosis as reported by Sieniawska et al. [44].
Generally, the mechanism of the antimicrobial activity of several terpenes is highly related to their lipophilic property which enable them to dissolve in the phospholipid layers of microbial cell membrane [45]. Particularly, natural sesquiterpenes such as β-elemene originated from plant and microorganisms showed promising antimicrobial activity [46,47]. A recent study conducted by Monga and Sharma [48] reported that β-elemene and R-limonene playing an essential role in degrading the microbial cell wall altering the expressions of dprE1 and clgR genes, responsible for the cell wall synthesis and cell membrane preservation, respectively.
On the other hand, some recent studies reported the promising cytotoxic effect of β-elemene which can inhibit cell proliferation, arrest cell cycle and induce cell apoptosis or autophagy [49]. β-elemene is one of the most promising inhibitors of glycolysis rate-limiting enzyme especially (PKM2) through the interfering with tumor glycolysis which consider one of the most important recent strategies for treating tumors [50,51]. In fact, several research reported that the inhibition of tumor growth and proliferation can be achieved by down-regulating expression of PKM2 enzyme [52]. In addition, Pan et al. [53] pointed to the role of β-elemene in inhibiting the breast cancer cell migration by conversion the dimer and tetramer forms of PKM2 and inhibit the aerobic glycolysis and reduce the utilization of glucose and the production of lactic acid for tumor cell growth.
4. Materials and Methods
4.1. Isolation, culturing and identification
Five strains of Beauveria bassiana (UniB2439-1; UniB2439-2, UniB2439-3, UniB2439-4, UniB2439-5) were isolated from different rhizospheric soil samples of Actinidia spp. and identified based on their morphological features and molecular basis. For molecular identification, the total gDNA was extracted and a partial part of β-tubulin gene was amplified using the universal primer Bt2a (5′-GGTAACCAAATCGGTGCTGCTTTC) and Bt2b (5′-ACCCTCAGTGTAGTGACCCTTGGC) [54]. The obtained amplicons were sequenced and then analyzed using Basic Local Alignment Search Tool software (BLAST-USA). The studied isolates were maintained as lyophils at 4 °C in the fungal collection of School of Agricultural, Forestry, Food and Environmental Sciences (SAFE), University of Basilicata, Potenza, Italy. The subcultures were carried out on Sabouraud Dextrose Agar plus 1% Yeast-extract (SDAY) nutrient media [5] and incubated at 22±2 °C for 96 hrs [55].
4.2. Antagonistic activity
The antagonistic activity of the five studied isolates of B. bassiana was evaluated against some pathogenic bacteria. All tested isolates were obtained from the pure cultures conserved in the collection of SAFE and previously identified based on morphological and molecular methods. The tested bacteria strains are listed in Table (3).
The antibacterial assay was carried out as described by Elshafie et al [56]. About 0.5 cm fungal disc from the fresh PDA culture (96 hrs) of each studied isolates of B. bassiana were deposited in the center of KB Petri dish and incubated for 16 hrs at 22±2 °C. Successively, a suspension of soft-agar (0.7%) of each tested bacteria at 108 CFU/ml were sprayed over the plates using Eco-Spray Ecological Aerosol (Seidden Identificación, Madrid – Spain). All plates were incubated at 30 °C for 24 hrs. Two KB plates inoculated only with each tested bacteria were used as negative control. The experiment was run in triplicate and the diameter of inhibition zone was measured with a caliber and recorded as the mean ± SD (n = 3). The antagonistic bacterial activity percentage (ABP %) was calculated using the Equation (1):
ABP (%) = D.iz / D.ct × 100; (Equation 1)
where ABP: is antagonistic bacterial activity; D.iz: is the diameter of inhibition zones in cm; D.ct: is the diameter of control plates in cm.
Table 3.
The tested bacterial strains in the current study.
| Bacteria name | Author | Collection number | Gram type |
|---|---|---|---|
| B. cereus | Frankland & Frankland | UniB12421 | G+ve |
| B. megaterium | de Bary | UniB12421 | |
| C. michiganensis | (Smith) Davis | UniB3718 | |
| X. campestris | (Pammel) Dowson | UniB7718 | G-ve |
| P. aeruginosa | (Schröter) Migula | UniB02421 | |
| P. fluorescens | (Flügge) Migula | UniB05421 |
All tested bacteria, with a collection number for each strain, are conserved in the collection of SAFE, University of Basilicata, Potenza, Italy.
4.3. Extraction of secondary metabolites
On the base of the preliminary antagonistic assay, the most efficient isolate (UniB2439-3) of B. bassiana was selected for the successive studies. For this scope, 2 mL of the fungal suspension (106 spore/mL) of the above isolate, was used for inoculating 500 mL SDY broth nutrient media and then incubated for 7 days at 25 °C in agitation (180 rpm). Both, Exo-ME and Endo-ME have been extracted from the broth culture after the incubation period.
For Endo-ME, the incubated broth culture was centrifuged at 40,000 g for 15 min and the pellet (2 g) was collected, resuspended in 50 mL of Limonene (CAS 138-86-3- Aldrich, Steinheim, Germany), shaked for 2 hrs and the solvent was evaporated after that using the Rotary-evaporator (Heidolph WB2000, Schwabach, Germany). The residue was resuspended in 2 ml of sterile distilled water (SDW), extracted following Solid Phase Extraction (SPE) by using C-18 column (Thermo Scientific, Rockwood, USA) and recovered using 1 mL methanol to reach the final original concentration of (20 mg/mL) [23].
For Exo-ME, the supernatant (250 mL), obtained from the above centrifugation step, was filtered using 0.22 µm (Syringe filter - hydrophilic, Minisart, Goettingen, Germany) and extracted using a separator funnel containing 250 mL ethyl acetate/ethanol (70:30; v/v) and shacked for 15 min. The organic phase was filtered through a filter paper (Whatman, Ø. 25 mm, Merck KGaA, Darmstadt, Germany) and evaporated using the Rotary-evaporator. The dry residue (50 mg) was resuspended in 2 mL SDW, extracted through SPE using C-18 column and recovered using 1 mL methanol to reach the final original concentration of (16 mg/mL) [23].
4.4. Antibacterial activity of diffusible metabolites
The antibacterial activity of both metabolites extracts was carried out against the same bacteria strains used for the initial antagonistic assay, are listed in Table (3).
Disc diffusion assay. The antibacterial test of both metabolites’ extracts produced by the most bioactive isolate UniB2439-3 was carried out following the disc diffusion method as described by Elshafie el al [57] and Sofo et al. [58]. A bacterial suspension of each tested bacteria was prepared in sterile distilled water adjusted at 106 CFU/mL (OD ≈ 0.2 nm) using UV-Spectrophotometer (Amersham, Ultraspec 1100 pro/500 pro, UK). Four mL of bacterial suspension mixed with soft agar 0.7 % (9:1; v/v) was poured over each KB plate (Ø 9 cm). Blank discs of 6 mm (OXOID, Milan-Italy) were then placed over the plates and 15 µL from each tested metabolites extract (Exo-ME 16 mg/mL and Endo-ME 20 mg/mL) was carefully applied over discs. Tetracycline (1600 µg/mL) was used as a positive control. The experiment was performed in triplicates and the antibacterial activity was estimated by measuring the diameter of inhibition zone in mm ± SDs compared to the positive control ones.
4.5. Antibacterial activity of volatiles metabolites
The tested bacterial strains were initially sub-cultured on 14 ml KB medium in Petri dishes and incubated at 37°C for 24 hrs. The most efficient isolate of B. bassiana (UniB2439-3) was cultured on PDA media (14 mL Petri dishes) and incubated at 22°C for 96 h. The test was performed according to Wan, et al. [59] using a double-dish chamber containing target tested fungal isolate in one downward dish of KB ( Ø 90 mm) and the tested bacterial strains were singularly inoculated on the up-ward dish either by direct colonies-inoculation or spreading of 50 μL of aqueous suspension (107 CFU/mL). The chamber was sealed with Parafilm™ and incubated at 37°C in darkness for 48 h. The antibacterial activity of the eventually produced volatile metabolites was evaluated by measuring the inhibition percentage of each tested bacteria. The experiment was carried out two times with three replicates.
4.6. SPME-GC/MS of VOCs
The fresh culture (96 hrs) of the selected Beauveria isolate was inoculated in glass tube of 10 ml PDA nutrient media and incubated at 22 °C for 5 days under darkness for collecting the volatile organic compounds (VOCs) as described by Elshafie et al [60]. The eventually produced VOCs have been analyzed qualitatively using Solid Phase Micro Extraction method (SPME) as discussed below.
The SPME fiber coated with 100 μm of non-grafted poly (dimethylsiloxane) phase (Supelco 57300-U, mounted on a Supelco 57330 support- Merck KGaA, Darmstadt, Germania) was conditioned for 1 h at 250 °C in a stream of helium. A blank run was performed after each analysis in order to confirm that no residual compounds were polluting the fiber or the column. The fiber was later introduced into the injection port of a HP6890 plus gas chromatograph equipped with a Phenomenex Zebron ZB-5 MS capillary column (30 m x 0.25 mm ID x 0.25 μm film thickness). A HP 5973 mass selective (mass range: 15-800 mAU; scan rate: 1.9 scan/s; EM voltage: 1435) was used as detector, whereas helium at 0.8 mL/min was used as carrier gas. The injection port, equipped with a glass insert (internal diameter 0.75 mm) was splitted at 250 °C. The desorption time of 1.0 min was used. Detector was maintained at 230 °C. Oven was maintained at 80 °C for 3 min, then the temperature was increased until 250 °C (20 °C/min) for 10 min. All the analyses were performed in triplicate. The chromatograms obtained from the total ion current were integrated without any correction for coelutions and the results were expressed as percent of the total area of peaks. All peaks were identified from their mass spectra by comparison with those present in Wiley 6N and NIST11 libraries [60,61].
5. Conclusions
B. bassiana, apart from being a notable entomopathogenic fungi or biocontrol agent against some phytopathogenic fungi, itself or its bioactive metabolites could be also used efficiently to control several bacteria in agronomic field where it is forbidden to utilize antibiotic especially in organic farming. In addition, B. bassiana could be also useful biocontrol agent against MDR microorganisms to different antibiotics which are considered a dominant medical problem worldwide. The obtained results from the current research concluded that, B. bassiana UniB2439-3, was able to produce some interesting VOCs such as β-elemene which has been reported previously to have strong antimicrobial effect against several pathogenic microorganisms. The ability of B. bassiana to produce the above-mentioned metabolites can underling its antagonistic activity against several phytopathogens as reported previously in the bibliographic research. Future studies remain necessary for evaluation the in vivo antimicrobial activity of each single identified bioactive volatile metabolites from B. bassiana against serious phytopathogens taking in consideration that the antibiotics are forbidden in agriculture in many countries worldwide. Therefore, search for possible natural alternatives as efficient antimicrobial agents remain necessary.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Chromatogram of VOCs extracted from B. bassiana UniB2439-3; Figure S2: Mass spectra of ethanol; Figure S3: Mass spectra of Butanal, 2-methyl; Figure S4: Mass spectra of 2,4-Dimethyl-1-heptene; Figure S5: Mass spectra of Octane, 4-methyl; Figure S6: Mass spectra of β-elemene. Table S1: Antagonistic antibacterial activity of the five studied isolates of Beauveria sp.; Table S2: The whole list of SPME-GC/MS analysis of VOCs extracted from B. bassiana UniB2439-3.
Author Contributions
Conceptualization, Ippolito Camele and Hazem Elshafie; Data curation, Sadeek Sadeek and Rocco Racioppi; Formal analysis, Sadeek Sadeek, Rocco Racioppi and Hazem Elshafie; Investigation, Ippolito Camele and Hazem Elshafie; Methodology, Sadeek Sadeek and Rocco Racioppi; Supervision, Ippolito Camele and Sadeek Sadeek; Writing – original draft, Ippolito Camele and Hazem Elshafie; Writing – review & editing, Hazem Elshafie.
Funding
This study was carried out within the Agritech National Research Center and received funding from the European Union Next-GenerationEU (PIANO NAZIONALE DI RIPRESA E RESILIENZA (PNRR) – MISSIONE 4 COMPONENTE 2, INVESTIMENTO 1.4 – D.D. 1032 17/06/2022, CN00000022). This manuscript reflects only the authors’ views and opinions, neither the European Union nor the European Commission can be considered responsible for them.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monz´on, A.; Ownley, B.H.; Pell, J.K.; Rangel, D.E.N.; Roy, H.E. Fungal entomopathogens: new insights on their ecology. Fungal Ecol. 2009, 2, 149–159.
- Barbarin, A.M.; Jenkins, N.E.; Rajotte, E.G.; Thomas, M.B. A preliminary evaluation of the potential of Beauveria bassiana for bed bug control. J. Invertebrate Pathol. 2012, 111, 82–85. [CrossRef]
- S.P.Wraight, R.B.Lopes, and M.Faria. Chapter 16: Microbial Control of Mite and Insect Pests of Greenhouse Crops. In: Microbial Control of Insect and Mite Pests, 2017, 237-252.
- Yuan, W.; Wang, P.; Zhang, Z.; Li, S. Glycosylation of (–)-maackiain by Beauveria bassiana and Cunninghamella echinulata var. elegans. Biocatal. Biotransformation 2010, 28, 117-121.
- Berestetskiya, A.O.; Ivanovaa, A.N.; Petrovaa, M.O.; Prokof’evab, D.S.; Stepanychevaa, E.A.; Uspanovc, A.M.; Lednev, G R. Comparative analysis of the biological activity and chromatographic profiles of the extracts of Beauveria bassiana and B. pseudobassiana cultures grown on different nutrient substrates. Microbiology 2018, 87, 200–214.
- Begley, C.G.; Waggoner, P. Soft contact lens contamination by Beauveria bassiana. Int. Contact Lens Clinic. 1992, 19, 247‒251. [CrossRef]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: a review. Mycologia 2018, 110, 4–30. [CrossRef]
- Fabrice, D.H.; Elie, D.A.; Kobi, D.O.; Valerien, Z.A.; Thomas, H.A.; Joëlle, T.; Maurille, E.I.A.T.; Dénis, O.B.; Manuele, T. Toward the efficient use of Beauveria bassiana in integrated cotton insect pest management. J. Cotton Res. 2020, 3, 24.
- Wang, Y.; Tang, D.; Duan, D.; Wang, Y.; Yu, H. Morphology, molecular characterization, and virulence of Beauveria pseudobassiana isolated from different hosts. J. Inverteb. Pathol. 2020, 72, 107333. [CrossRef]
- Sinno, M.; Ranesi, M.; Di Lelio, I.; Iacomino, G.; Becchimanzi, A.; Barra, E.; Molisso, D.; Pennacchio, F.; Digilio, M.C.; Vitale, S.; et al. Selection of endophytic Beauveria bassiana as a dual biocontrol agent of tomato pathogens and pests. Pathogens 2021, 10, 1242. [CrossRef]
- Faria, M.R.; Wraight, S.P. Biological control of Bemisia tabaci with fungi. Crop Prot. 2001, 20, 767–778. [CrossRef]
- Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: a comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256.
- Fargues, J.; Remaudiere, G. Consideration on the specificity of entomopathogenic fungi. Mycopathologia 1977, 62, 31–37. [CrossRef]
- Hasaballah, A.I.; Fouda, M.A.; Hassan, M.I.; Omar, G.M. Pathogenicity of Beauveria bassiana and Metarhizium anisopliae on the adult housefly, Musca domestica L. Egypt. Acad. J. Biolog. Sci. (A. Entomology) 2017, 10, 79- 86. [CrossRef]
- Acharya, N.; Rajotte, E.G.; Jenkins, N.E.; Thomas, M.B. Potential for biocontrol of house flies, Musca domestica, using fungal biopesticides. Biocontrol Sci. Technol. 2015, 25, 513-524. [CrossRef]
- Barra-Bucarei, L.; Iglesias, A.F.; González, M.G.; Aguayo, G.S.; Carrasco-Fernández, J.; Castro, J.F.; Campos, J.O. Antifungal activity of Beauveria bassiana Endophyte against Botrytis cinerea in Two Solanaceae Crops. Microorganisms 2020, 8, 65. [CrossRef]
- Parine, N.R.; Pathan, A.K.; Sarayu, B.; Nishanth, V.S.; Bobbarala, V. Antibacterial e_cacy of secondary metabolites from entomopathogenic fungi Beauveria bassiana. Int. J. Chem. Anal. Sci. 2010, 1, 94–96.
- Ownley, B.H.; Bishop, D.G.; Pereira, R.M. Biocontrol of Rhizoctonia damping off of tomato with Beauveria bassiana. Phytopathol. 2000, 90, S58.
- Della Pepa, T.; Elshafie, H.S.; Capasso, R.; De Feo, V.; Camele, I.; Nazzaro, F.; Scognamiglio, M.R.; Caputo, L. Antimicrobial and phytotoxic activity of Origanum heracleoticum and O. majorana essential oils growing in Cilento (Southern Italy). Molecules 2019, 24, 2576.
- Gruľová, D., Caputo, L., Elshafie, H. S., Baranová, B., De Martino, L., Sedlák, V., Camele I.; De Feo, V. Thymol chemotype Origanum vulgare L. essential oil as a potential selective bio-based herbicide on monocot plant species. Molecules 2020, 25, 595. [CrossRef]
- Anyanwu, M. U., Okoye, R.C. Antimicrobial Activity of Nigerian Medicinal Plants. J. Intercult Ethnopharmacol 2017, 6, 240–259.
- Keifer, M.C.; Firestone, J. Neurotoxicity of pesticides. J. Agromed. 2007, 12, 17–25. [CrossRef]
- Camele I., Elshafie H.S., Caputo L., Sakr S.H., De Feo V. Bacillus mojavensis: Biofilm formation and biochemical investigation of its bioactive metabolites. J. Biol. Res. 2019, 92, 39-45. [CrossRef]
- Wang H., Peng H., Li W., Cheng P., Gong M. The toxins of Beauveria bassiana and the strategies to improve their virulence to insects. Front. Microbiol. 2021, 12, 705343.
- Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997, 1; 25, 3389-3402. [CrossRef]
- Yang, X.; Ye, W., Qi, Y., Ying, Y.; Xia, Z. Overcoming Multidrug Resistance in Bacteria Through Antibiotics Delivery in Surface-Engineered Nano-Cargos: Recent Developments for Future Nano-Antibiotics. Front. Bioeng. Biotechnol. 2021, 9, 696514. [CrossRef]
- Soothill, G.; Hu, Y.; Coates, A. Can we prevent antimicrobial resistance by csing antimicrobials better?. Pathogens 2013, 2, 422–435.
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596.
- Chen, W.H., Liu, M., Huang, Z.X., Yang, G.M., Han, Y.F., Liang, J.D., Liang, Z.Q. Beauveria majiangensis, a new entomopathogenic fungus from Guizhou, China. Phytotaxa 2018, 333, 243–250. [CrossRef]
- Ownley, B.H.; Pereira, R.M.; Klingeman,W.E.; Quigley, N.B.; Leckie, B.M. Beauveria bassiana, a dual purpose biological control with activity against insect pests and plant pathogens. Emerg. Concepts Plant Health Manag. 2004, 2004, 255–269.
- Suzuki, A.; Kanaoka, M.; Isogai, A.; Tamura, S.; Murakoshi, S.; Ichinoe, M. Bassianolide, a new insecticidal cyclodepsipeptide from Beauveria bassiana and Verticillium lecanii. Tetrahedron Lett. 1977, 18, 2167–2170. [CrossRef]
- Wang S., Zhao Z., Yun-Ting S., Zeng Z., Zhan X., Li C., Xie T. A review of medicinal plant species with elemene in China. Afr. J. Pharm. Pharmacol. 2012, 6, 3032–3040.
- El Kichaoui, A.; Elnabris, K.; Shafie, A.; Fayyad, N.; Arafa, M.; El Hindi, M. Development of Beauveria bassiana based bio-fungicide against Fusarium wilt pathogens for Capsicum annuum. IUG J. Nat. Stud. 2017, 183-190.
- Vining, L.C.; Kelleher, W.J.; Schwarting, A.E. Oosporein production by a strain of Beauveria bassiana originally identified as Amanita muscaria. Can. J. Microbiol. 1962, 8, 931–933. [CrossRef]
- Hamill, R.L.; Higgens, C.E.; Boaz, H.E.; Gorman, M. The structure of beauvericin, a new depsipeptide antibiotic toxic to Artemia salina. Tetrahedron Lett. 1969, 10, 4255–4258.
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553-596.
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin production by Fusarium species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [CrossRef]
- Nagaoka, T.; Nakata, K.; Kouno, K. Antifungal activity of oosporein from an antagonistic fungus against Phytophthora infestans. Z. Naturforschung C 2004, 59, 302–304. [CrossRef]
- Wang, Q.; Xu, L. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [CrossRef]
- Manning, R.O.; Wyatt, R.D. Comparative toxicity of Chaetomium contaminated corn and various chemical forms of oosporein in broiler chicks. Poultry Sci. 1984, 63, 251-259. [CrossRef]
- Golembiovska, O.; Tsurkan, A.; Vynogradov, B. Components of Prunella vulgaris L. Grown in Ukraine J. Pharmacognosy Phytochem. 2014, 2, 140-146.
- Bernotienë, G.; Nivinskienë O.; Butkienë, R.; Mockutë, D. Chemical composition of essential oils of hops (Humulus lupulus L.) growing wild in Aukštaitija. Chemija 2004, 15, 31-36.
- Fischedick, J.T.; Hazekamp, A.; Erkelens, T.; Choi, Y.H.; Verpoorte, R. Metabolic fingerprinting of Cannabis sativa L., cannabinoids and terpenoids for chemotaxonomic and drug standardization purposes. Phytochem. 2010, 71, 2058-2073. [CrossRef]
- Sieniawska, E.; Sawicki, R.; Golus, J.; Swatko-Ossor, M.; Ginalska, G.; Skalicka-Wozniak, K. Nigella damascena L. essential Oil—a valuable source of β-Elemene for antimicrobial testing. Molecules 2018, 23, 256. [CrossRef]
- Mendanha, S.A.; Alonso, A. Effects of terpenes on fluidity and lipid extraction in phospholipid membranes. Biophys. Chem. 2015, 198, 45–54. [CrossRef]
- Barrero, A.F.; Quilez del Moral, J.F.; Lara, A.; Herrador, M.M. Antimicrobial activity of sesquiterpenes from the essential oil of Juniperus thurifera Wood. Planta Med. 2005, 71, 67–71. [CrossRef]
- Drage, S.; Mitter, B.; Tröls, C.; Muchugi, A.; Jamnadass, R.H.; Sessitsch, A.; Hadacek, F. Antimicrobial drimane sesquiterpenes and their effect on endophyte communities in the medical tree Warburgiau gandensis. Front. Microbiol. 2014, 5, 13.
- Monga A.; Sharma A. 2020. Chapter 9 - Natural products encompassing antituberculosis activities. Studies in Natural Products Chemistry. 2020, 64, 263-301.
- Zhai, B.; Zhang, N.; Han, X.; Li, Q.; Zhang, M.; Chen, X.; Li, G.; Zhang, R.; Chen, P.; Wang, W.; Li, C.; Xiang, Y.; Liu, S.; Duan, T.; Lou, J.; Xie, T.; Sui, X. Molecular targets of β-elemene, a herbal extract used in traditional Chinese medicine, and its potential role in cancer therapy: A review. Biomed. Pharmacoth. 2019, 14, 108812. [CrossRef]
- T. Dong, Y. Yan, H. Chai, S. Chen, X. Xiong, D. Sun, Y. Yu, L. Deng, F. Cheng, Pyruvate kinase M2 affects liver cancer cell behavior through up-regulation of HIF-1α and Bcl-xL in culture. Biomed. Pharmacother. 69 (2015) 277–284.
- van Niekerk, G.; Engelbrecht, A.M. Role of PKM2 in directing the metabolic fate of glucose in cancer: a potential therapeutic target, Cell Oncol. (Dordr.) 2018, 41, 343–351.
- Sun, X.; Peng, Y.; Zhao, J.; Xie, Z.; Lei, X.; Tang, G. Discovery and development of tumor glycolysis rate-limiting enzyme inhibitors. Bioorganic Chem. 2021, 112, 104891. [CrossRef]
- Pan, Y.; Wang, W.; Huang, S.; Ni, W.; Wei, Z.; Cao, Y.; Yu, S.; Jia, Q.; Wu, Y.; Chai, C.; Zheng, Q., Zhang, L.; Wang, A., Sun, Z.; Huang, S.; Wang, S.; Chen, W.; Lu, Y. Beta-elemene inhibits breast cancer metastasis through blocking pyruvate kinase M2 dimerization and nuclear translocation. J. Cell Mol. Med., 2019, 23, 6846–6858. [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [CrossRef]
- Johnson, D.M.; White, R.L.; Pereira, R.M.; Geden, C.J. Beauveria bassiana culturing and harvesting for bioassays with house flies. J. Insect Sci. 2020, ,, 1–7. [CrossRef]
- Elshafie, H.S.; Racioppi, R.; Bufo, S.A.; Camele, I. In vitro study of biological activity of four strains of Burkholderia gladioli pv. agaricicola and identification of their bioactive metabolites using GC–MS. Saudia J. Biol Sci. 2017, 24, 295–301. [CrossRef]
- Elshafie, H.S.; Viggiani, L.; Mostafa, M.S.; El-Hashash, M.A.; Bufo, S.A.; Camele; I. Biological activity and chemical identification of ornithine lipid produced by Burkholderia gladioli pv. agaricicola ICMP 11096 using LC-MS and NMR analyses. J. Biol. Res. 2017, 90, 96-103. [CrossRef]
- Sofo, A.; Elshafie, H.S.; Scopa, A.; Mang, S.M.; Camele, I. Impact of airborne zinc pollution on the antimicrobial activity of olive oil and the microbial metabolic profiles of Zn-contaminated soils in an Italian olive orchard. J. Trace Elem. Med. Biol. 2018, 49, 276–284. [CrossRef]
- Wan, M.G.; Li, G.Q.; Zhang, J.B.; Jiang, D.H.; Huang, H.C. Effect of volatile substances of Streptomyces platensis F-1 on control of plant fungal diseases. Biol. Cont. 2008, 46, 552–559.
- Camele, I.; Grul’ová, D.; Elshafie, H.S. Chemical composition and antimicrobial properties of Mentha _ piperita cv. ‘Kristinka’ essential oil. Plants 2021, 10, 1567.
- Elshafie, H.S.; Camele, I.; Racioppi, R.; Scrano, L.; Iacobellis, N.S.; Bufo, S.A. In vitro antifungal activity of Burkholderia gladioli pv. agaricicola against some Phytopathogenic fungi. Int. J. Mol. Sci. 2012, 13, 16291-16302.
Figure 1.
Antagonistic activity of B. bassiana UniB2439-3 Bars with different letters are significantly different according to one-way ANOVA combined with Tukey B post hoc multiple comparison test at P < 0.05 using SPSS statistical analysis software. Data for each bar are expressed as the mean of three replicates ± SDs.
Figure 1.
Antagonistic activity of B. bassiana UniB2439-3 Bars with different letters are significantly different according to one-way ANOVA combined with Tukey B post hoc multiple comparison test at P < 0.05 using SPSS statistical analysis software. Data for each bar are expressed as the mean of three replicates ± SDs.
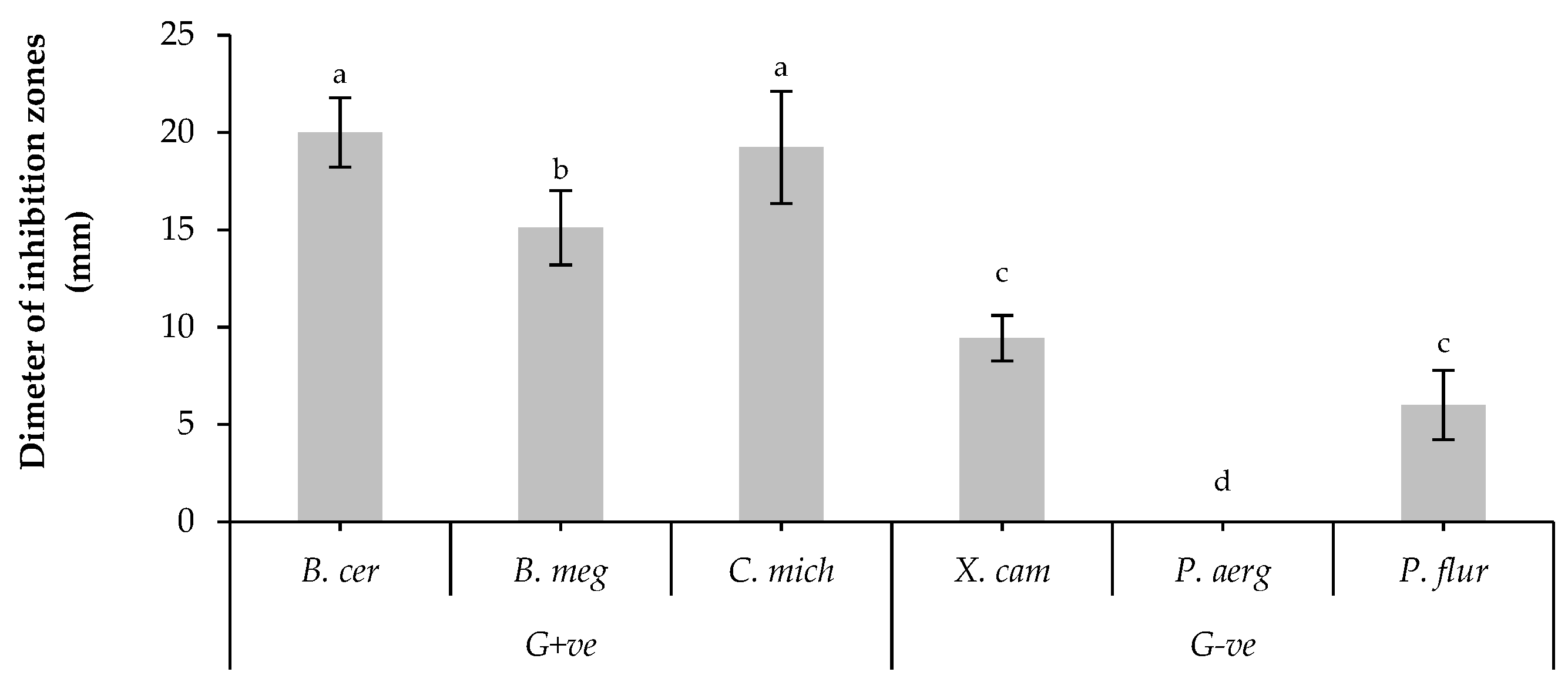
Table 1.
Antibacterial activity of diffusible metabolites from B. bassiana UniB2439-3.
| Tested bacteria | Diameter of inhibition zones (mm) | |||
|
Exo-ME 16 mg/mL |
Endo-ME 20 mg/mL |
Tetracycline 1600 µg/mL |
||
| G+ve | B. cereus | 8.5±1.0ab | 0.0±0.0c | 20.8±1.1b |
| B. megaterium | 10.0±1.9ab | 4.0±1.7b | 25.9±2.3ab | |
| C. michiganensis | 12.5±2.2a | 0.0±0.0c | 39.5±2.5a | |
| G-ve | X. campestris | 9.5±2.5ab | 9.0±1.9a | 23.5±1.7ab |
| P. aeruginosa | 0.0±0.0c | 6.5±2.8ab | 10.6±0.7c | |
| P. fluorescens | 6.5±1.5b | 4.5±1.7b | 12.3±0.9c | |
Values followed by different letters in each column for each tested extract against all tested bacteria are significantly different at P < 0.05 according to one-way ANOVA combined with Tukey B post hoc test by using SPPS program. Data are expressed as the mean of inhibition zone diameter (mm) for three replicates ± SDs compared to controls ± SDs.
Table 2.
Antibacterial activity of diffusible metabolites from B. bassiana UniB2439-3.
| Tested bacteria | Bacterial growth inhibition (%) | |||
| GVC | AQS |
Tetracycline 1600 µg/mL |
||
| G+ve | B. cereus | 35.0±5.8c | 60.0±5.8c | 20.8±1.1b |
| B. megaterium | 77.5±2.9a | 92.0±3.5a | 25.9±2.3ab | |
| C. michiganensis | 55.0±5.8b | 77.5±2.9b | 39.5±2.5a | |
| G-ve | X. campestris | 27.5±2.9d | 77.5±8.7b | 23.5±1.7ab |
| P. aeruginosa | 37.5±2.9c | 87.5±2.9a | 10.6±0.7c | |
| P. fluorescens | 52.5±2.9b | 72.5±2.9b | 12.3±0.9c | |
Where: (GVC) grown-visible colonies; (AQS) aqueous suspension. Values followed by different letters in each column for each tested extract against all tested bacteria are significantly different at P < 0.05 according to one-way ANOVA combined with Tukey B post hoc test by using SPPS program. Data are expressed as the mean of inhibition zone diameter (mm) for three replicates ± SDs compared to controls ± SDs.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



