
Submitted:
09 September 2024
Posted:
09 September 2024
You are already at the latest version
Abstract
Porcine circovirus-associated diseases, caused by porcine circovirus type 2 (PCV2), are widespread and result in significant economic losses to the global swine industry. It is generally believed that PCV2 can be divided into nine genotypes (PCV2a to PCV2i), with the dominant being the PCV2d genotype. In this study, 2675 samples from pig farms in 13 cities in Jiangsu Province, China, were collected between 2014 and 2021 and subjected to polymerase chain reaction analysis to investigate the prevalence and genetic diversity of PCV2. The results showed that 41.42% (1108/2675) of samples tested positive for PCV2. The researchers further analyzed the genetic characteristics of 251 PCV2 strains and found that they belonged to the following four genotypes: PCV2a, PCV2b, PCV2d, and PCV2i. The dominant genotype was PCV2d, with a prevalence of 49.80% (125/251). The detection rate of PCV2b was significantly higher than those of PCV2a and PCV2i, at 35.46% (89/251), 7.57% (19/251), and 7.17% (18/251), respectively. We have further revealed the fingerprint of PCV2i genomic nucleotides for the first time. In conclusion, this study illustrates the high prevalence and evolutionary features of PCV2 in Jiangsu Province over the past few years.
Keywords:
porcine circovirus type 2
; Jiangsu Province
; genetic analysis
; genotype
Introduction
Porcine circoviruses (PCVs) are the smallest, circular, single-stranded DNA viruses belonging to the genus Circovirus of the family Circoviridae. The following four species of PCVs with similar structures have been identified: porcine circovirus type 1 (PCV1), PCV2, PCV3, and PCV4 [1,2,3,4,5]. In addition, other species of PCVs, including porcine circovirus-like viruses and porcine circovirus-like mini agents, have been reported [6]. PCV2 has been considered the primary causative agent of porcine circovirus-associated diseases (PCVADs) since its identification in Canada in the 1990s. Post-weaning multisystemic wasting syndrome (PMWS) is the most common PCVAD in clinical practice, characterized by slow growth in fattening pigs, which causes serious economic losses to the pig industry worldwide [7,8,9].
The genome of PCV2 is approximately 1767 nucleotides (nt) in length and mainly comprises two open reading frames (ORFs). Replication-related proteins (Rep and Rep’) are encoded by ORF1, whereas the capsid protein (Cap) is encoded by ORF2 [10,11,12]. Compared with other ORFs, the ORF2 gene can be used for genetic evolution analysis due to its high mutation rate. A nucleotide diversity (p-distance) cutoff value of 3.5% (for ORF2) or 2.0% (for the complete genome) was used to distinguish between different PCV2 genotypes [13]. Nine genotypes (PCV2a–PCV2i) have been identified [14,15]. PCV2 strains circulating in pigs before 2008 belonged to three genotypes (PCV2a–PCV2c). PCV2d was identified in 2010 and has become prevalent worldwide since 2012 [16]. Other novel PCV2 genotypes (PCV2e–PCV2i) have been identified in recent years [14,15,17,18,19]. Generally, there are two changes in the dominant genotype of PCV2: one from PCV2a to PCV2b and another from PCV2b to PCV2d [20,21].
Previous studies have examined the occurrence of PCV in Jiangsu Province [22,23]. However, there is a lack of comprehensive data on the genetic variety and prevalence of PCV2 in Jiangsu pig farms. Therefore, an investigation was conducted from 2014 to 2021 using polymerase chain reaction (PCR) to determine the epidemiological characteristics of PCV2 in Jiangsu Province. In addition, the complete genomes of 251 PCV2 strains from various periods and cities were analyzed.
Materials and Methods
Clinical Sample Collection
From April 2014 to September 2021, 2675 clinical samples (lung, serum, diarrheal stool, and lymph node) were collected from pig farms in the following 13 cities in Jiangsu Province: Nanjing, Suzhou, Wuxi, Changzhou, Zhenjiang, Nantong, Huaian, Yangzhou, Taizhou, Yancheng, Suqian, Lianyungang, and Xuzhou. Most of the pigs from these farms showed clinical signs of PMWS (Table 1).
DNA Extraction and PCV2 Detection
Commercial kits (TIANDZ, China) were used to extract viral DNA from the samples according to the manufacturer’s instructions. Subsequently, the presence of PCV2 nucleic acid was detected using PCR with the primers (F: 5’-TAGACGGATATTGTAGTCC-3’ and R: 5’-TTCCGCAGAAGAAGACAC-3’). PCR conditions were as follows: 95°C for 5 min; 35 cycles of 94°C for 30 s, 58°C for 30 s, and 72°C for 30 s; and a final extension for 10 min at 72°C. The PCR products were analyzed using 1.5% agarose gel electrophoresis, in which the samples with the expected 630 bp DNA band were recognized as PCV2-positive samples.
Complete Genome Sequencing of PCV2
According to the collected cities and years of the positive samples, 251 PCV2-positive samples were randomly selected to amplify the complete genome of the virus, as described previously [24]. PCR amplification was performed using the following steps: 95°C for 5 min; 35 cycles of 94°C for 30 s, 58°C for 30 s, and 72°C for 90 s, followed by 72°C for 10 min. The PCR products were subsequently purified using a Gel Extraction Kit (AXYGEN, China), cloned into the pMD18-T vector (TaKaRa Biotechnology Co. Ltd. China), and sequenced by Genscript Biotech Co. Ltd. (Nanjing, China).
The sequences of the 251 novel PCV2 strains were submitted to GenBank (OR533426–OR533483, OR537631–OR537693, OR542932–OR542954, OR553617–OR553646, and OR567166–567242).
Bioinformatics Analyses
The complete genomes of the following eight reference PCV2 strains were downloaded from GenBank: PCV2a (AF381175), PCV2b (KJ511872), PCV2c (EU148503), PCV2d (JQ181600), PCV2e (KT795290), PCV2f (MF278779), PCV2g (JX099786), and PCV2h (MH465473). The Lasergene DNAStar software was used to analyze the nucleotide sequences (complete, ORF1, and ORF2) and their corresponding amino acid (aa) sequences (Cap and Rep) of 251 novel PCV2 strains and reference strains. Phylogenetic trees were generated based on the complete genome sequences using the neighbor-joining method with maximum composite likelihood model in MEGA 7.0 software with 1000 bootstrap replications.
Prevalence Data Analyses
The prevalence data analysis of PCV2 in pigs in Jiangsu Province was performed using the chi-square test with Excel 2010, including infection rates by year and rates in different cities. Statistical significance was set at p < 0.05.
Results
Prevalence of PCV2 in Jiangsu Province from 2014 to 2021
In this study, 2675 samples from pig farms in 13 cities in Jiangsu Province were collected for PCV2 nucleic acid analysis using conventional PCR. The results showed that the positive rate of PCV2 was 41.42% (1108/2675), with positive rates varying from 0.00 to 100.00% in different cities and from 27.89 to 63.33% in different years. In 2021, the infection rate of PCV2 in Jiangsu Province was the highest at 63.33% (95/150), and the infection rate was the lowest in 2018 at 27.89% (94/337). From a regional perspective, the infection rate of PCV2 was the highest in Zhenjiang, at 63.64% (42/66), and the infection rate was the lowest in Changzhou, at 28.87% (28/97) (Table 2). There was an extremely significant difference in the infection rate of PCV2, both in terms of year and city (P < 0.01).
Genome Sequence Analysis
In this study, 251 PCV2 strains were randomly selected from different regions among the PCV2-positive samples to investigate the genetic features of recent PCV2 strains prevalent in Jiangsu Province. The complete genome sequences of all 251 PCV2 strains were amplified, sequenced, and analyzed and were found to be 1766–1768 nt in length. The ORF1 gene encoding Rep and ORF2 gene encoding Cap had lengths of 945 nt and 702–705 nt, respectively. The 251 PCV2 strains shared 94.1–100.0% (complete genome), 96.1–100.0% (ORF1), and 89.5–100.0% (ORF2) identity at the nt level and 96.5–100.0% (Rep protein) and 85.9–100.0% (Cap protein) identity at the aa level. Interestingly, a nucleotide deletion at site 1039 nt was unique to the JS890 strain. In contrast, sites 1357 (nucleotide deletion) and 1390 (nucleotide insertion) were unique to the JS953 strain, and site 1004 (nucleotide deletion) was unique to the JS6418 strain.
Phylogenetic Analysis
Phylogenetic trees were generated based on the complete genome sequences of 251 PCV2 strains identified in this study. The 251 PCV2 strains were divided into the following four genotypes: PCV2a, PCV2b, PCV2d, and PCV2i. Phylogenetic tree analysis further revealed that PCV2a and PCV2i strains had a closer phylogenetic relationship than PCV2b and PCV2d. Among the 251 strains, 19 (7.57%) belonged to genotype 2a, 18 (7.17%) to genotype 2i, 89 (35.46%) to genotype 2b, and nearly half of the strains (125) to genotype 2d (Figure 1).
Although the prevalence of PCV2d fluctuated, PCV2d was generally the dominant genotype that circulated between 2014 and 2021, followed by PCV2b, whereas the prevalence of PCV2i was lower than that of PCV2b. Notably, PCV2a still has a low prevalence (Figure 2).
The sequence similarity range of the novel PCV2a strains was 98.9–100.0% (complete genome), 99.2–100.0% (ORF1), and 98.3–100.0% (ORF2) at the nt level and 99.4–100.0% (Rep protein) and 98.3–100.0% (Cap protein) at the aa level.
In contrast, sequence similarities among the novel PCV2b strains varied between 98.0 and 100.0% (complete genome), 98.1 and 100.0% (ORF1), and 97.5 and 100.0% (ORF2) at the nt level and 98.1 and 100.0% (Rep protein) and 95.6 and 100.0% (Cap protein) at the aa level.
The sequence homology range among the novel PCV2d strains was 96.2–100.0% (complete genome), 96.5–100.0% (ORF1), and 95.3–100.0% (ORF2) at the nt level and the variation at the aa level was 97.1–100.0% (Rep protein) and 94.7–100.0% (Cap protein).
Finally, pairwise sequence comparisons among the novel PCV2i strains varied between 98.5 and 100.0% (complete genome), 98.7 and 100.0% (ORF1), and 98.1 and 100.0% (ORF2) at the nt level and 98.1 and 100.0% (Rep protein) and 97.9 and 100.0% (Cap protein) variation at the aa level.
Analysis of Genomic Sequences of PCV2i Strains
In a recent study, a new genotype called PCV2i was proposed through phylogenetic analysis, but not much information is available about this genotype [15]. To investigate the sequence characteristics of PCV2i obtained in this study, a sequence alignment was performed among the 251 PCV2 strains. The complete genome of all 18 PCV2i strains was 1768 nt in length, and the length of the Cap sequence was 233 aa.
Based on the complete genome sequence, the nucleotide sequence homology between PCV2i and PCV2a was 96.0–97.3%, 94.8–96.0% (PCV2b), and 93.8–95.6% (PCV2d), respectively, and 93.3–94.7% (PCV2a), 91.1–92.5% (PCV2b), and 91.6–92.9% (PCV2d), respectively, based on the ORF2 nucleotide sequence.
For the 251 PCV2 strains and eight reference strains in this study, nucleotides at positions 617 (T), 1164 (T), 1596 (C), and 1604 (A) were exclusively present in PCV2i strains. Additionally, aa residues 47 (S) and 191 (K) in the Cap of PCV2i strains were identified as potential signature markers for PCV2i (Table 3).
In addition, the nucleotide sites 1175, 1177, 1226, 1313, 1343, and 1673 of all PCV2i strains were A, T, G, G, A, and T, respectively, which are currently only observed in PCV2f with A at position 1175, in PCV2a with T at 1177, in PCV2f with G at position 1226, in PCV2f with G at position 1313, in PCV2f with A at position 1343, and in PCV2e with T at position 1673.
Discussion
The disease caused by PCV2, known as PCVAD, is a major threat to the pig industry worldwide as it is considered a global epidemic [25,26,27,28]. The first report of PCV2 infection in China was published in 2000; and since then, it has become widespread throughout the country [29].
To study the epidemiological characteristics of PCV2 in Jiangsu Province, southern China, 2675 pig samples were collected. Previous research has shown that the infection rates of PCV2 in breeding farms in southern China are higher than those in northern China [30]. However, the PCV2-positive rate among the samples collected in Jiangsu Province was 42.58%, which is similar to that in Shandong Province (36.98%, 490/1325) [31] in northern China but lower than that in Henan (62.4%, 73/117) in northern China [32].
PCV2 has a higher evolutionary rate than other DNA viruses, with a rate of 10-3–10-4 substitutions/site/year, which is similar to that of RNA viruses [33]. In the past, the PCR–RFLP method was employed for PCV2 genotyping, which used nucleotide sequence amplification and endonuclease digestion. Research conducted in Canada, Japan, and China using this method showed that there were five RFLP types (A–E) of PCV2 in Canada, at least five RFLP types in Japan, and nine different genotypes (A–I) in China [34,35,36].
To distinguish between different PCV2 genotypes, pairwise sequence comparison (PASC) analysis, which is based on the proportion of nucleotide sites at which two sequences differ (p-distance), is widely used. A diversity cutoff of 2.0% for the complete genome and 3.5% for ORF2 has been proposed [13].
A new PCV2 genotyping methodology based on three criteria was proposed by Franzo and Segalés in 2018. The first criterion was a maximum intragenotype p-distance of 13%, which was calculated for the ORF2 gene. The second criterion was bootstrap support at a corresponding internal node higher than 70%, and the third criterion was at least 15 available sequences. Based on these criteria, PCV2 has been divided into eight genotypes ranging from PCV-2a to PCV-2h [14]. Subsequently, a new genotype, PCV2i, was proposed through phylogenetic analysis [15].
PCV2 has undergone two significant global genotypic shifts. The dominant PCV2a genotype was replaced with PCV2b from 2002 to 2008. Since 2009, PCV2d has replaced PCV2b with the dominant genotype worldwide [37,38]. PCV2d is considered to be more virulent than PCV2a and PCV2b, with severe clinical signs, viremia, and lesions [39]. However, there are also reports suggesting that the virulence of PCV2d is similar to that of PCV2a and PCV2b [40,41]. Notably, the coprevalence of multiple PCV2 genotypes or with other circoviruses, such as PCV3, PCV4, and PCV-like virus P1, often appears in a pig farm or even in a sample [42,43,44].
Only the following four genotypes were identified among the 251 PCV2 strains obtained in this study: PCV2a, PCV2b, PCV2d, and PCV2i, with proportions of 7.57, 35.46, 49.80, and 7.17%, respectively. This indicates that PCV2b and PCV2d are the predominant genotypes in Jiangsu Province. Specifically, PCV2b and PCV2d were the dominant genotypes circulating in the Jiangsu Province from 2015 to 2017. Although the detection rates of PCV2a and PCV2i were similar, PCV2a was not detected before 2017, whereas PCV2i was scattered in samples from 2014 to 2021.
This study provides evidence supporting the existence of PCV2i as a new genotype via PASC analysis, and Franzo and Segalés proposed a new genotyping methodology or phylogenetic analysis. However, its genomic characteristics have not been elucidated since it has been recommended as a new genotype. This study revealed four characteristic nucleotide changes in the PCV2i genome at sites 617 (T), 1164 (T), 1596 (C), and 1604 (A), which resulted in characteristic changes in aa residues 47 (S) and 191 (K) of the viral capsid protein compared with those of other genotypes.
PCV2 Cap is the only structural protein involved in multiple biological processes, including viral invasion, replication, and immune response [45]. Studies have shown that certain aa residues in the protein are critical for viral nuclear localization and epitope recognition. Specifically, residues 12-18 and 34-41 are important for nuclear localization [46], while residues 47-63, 165 -200, and the last four aa residues at the C-terminus are crucial for epitope recognition [47,48]. Therefore, the two aa mutations in the Cap of PCV2i may determine the biological characteristics of the virus. Further research is needed to determine the virulence of PCV2i and the effectiveness of current vaccines against it. Interestingly, mutations in specific nucleotides of PCV2i have led to new endonuclease digestion sites that can be used for PCR-RFLP genotyping of PCV2.
Conclusions
This study sheds light on the epidemiology and genetic characteristics of PCV2 that circulated between 2014 and 2021 in Jiangsu Province, China. These findings reveal a high prevalence of multiple PCV2 genotypes in the region and provide insights into the genomic characteristics of PCV2i.
Author Contributions
Conceptualization, K.H.; methodology, Q.X.; software: L.W.; validation, M.Q., J.X., C.Z., A.M., Y.S., W.Q., J.Z., J.G., D.L. and J.N.; formal analysis, L.W.; investigation, Q.X.; data curation, Q.X. and K.H.; writing—original draft preparation, L.W.; writing—review and editing, K.H.; funding acquisition, K.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant Nos. 31972679).
Institutional Review Board Statement
The animal study was approved by the Committee on Ethics of Animal Experiments of the Institute of Veterinary Medicine, Jiangsu Academy of Agricultural Sciences (JAAS No 20100604).
Data Availability Statement
All data, including the names of the PCV2 strains and accession number(s) presented in this study are available in the GenBank repository.
Acknowledgments
We would like to thank all the staff who collected the samples. We thank LetPub (www.letpub.com.cn) for its linguistic assistance during the preparation of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tischer, I.; Gelderblom, H.; Vettermann, W.; Koch, M.A. A very small porcine virus with circular single-stranded DNA. Nature 1982, 295, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Meehan, B.M.; McNeilly, F.; Todd, D.; Kennedy, S.; Jewhurst, V.A.; Ellis, J.A.; Hassard, L.E; Clark, E.G.; Haines, D.M.; Allan, G.M. Characterization of novel circovirus DNAs associated with wasting syndromes in pigs. J. Gen. Virol. 1998, 79 Pt 9, 2171–2179. [Google Scholar] [CrossRef] [PubMed]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2016, 91, e01879–16. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Giannitti, F.; Rossow, S.; Marthaler, D.; Knutson, T.P.; Li, L.; Deng, X.; Resende, T.; Vannucci, F.; Delwart, E. Detection of a novel circovirus PCV3 in pigs with cardiac and multi-systemic inflammation. Virol. J. 2016, 13, 184. [Google Scholar] [CrossRef]
- Zhang, H.H.; Hu, W.Q.; Li, J.Y.; Liu, T.N.; Zhou, J.Y.; Opriessnig, T.; Xiao, C.T. Novel circovirus species identified in farmed pigs designated as Porcine circovirus 4, Hunan province, China. Transbound. Emerg. Dis. 2020, 67, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.B.; He, K.W. Genomic rearrangement and recombination of porcine circovirus type 2 and porcine circovirus-like virus P1 in China. Front. Vet. Sci. 2021, 8, 736366. [Google Scholar] [CrossRef]
- Allan, G.M.; Ellis, J.A. Porcine circoviruses: a review. J. Vet. Diagn. Invest. 2000, 12, 3–14. [Google Scholar] [CrossRef]
- Chae, C. A review of porcine circovirus 2-associated syndromes and diseases. Vet. J. 2005, 169, 326–336. [Google Scholar] [CrossRef]
- Opriessnig, T.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 associated disease: update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J. Vet. Diagn. Invest. 2007, 19, 591–615. [Google Scholar] [CrossRef]
- Cheung, A.K. Transcriptional analysis of porcine circovirus type 2. Virology. 2003, 305, 168–180. [Google Scholar] [CrossRef]
- Mankertz, A.; Hillenbrand, B. Replication of porcine circovirus type 1 requires two proteins encoded by the viral rep gene. Virology 2001, 279, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Nawagitgul, P.; Morozov, I.; Bolin, S.R.; Harms, P.A.; Sorden, S.D.; Paul, P.S. Open reading frame 2 of porcine circovirus type 2 encodes a major capsid protein. J. Gen. Virol. 2000, 81, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Grau-Roma, L.; Crisci, E.; Sibila, M.; López-Soria, S.; Nofrarias, M.; Cortey, M.; Fraile, L.; Olvera, A.; Segalés, J. A proposal on porcine circovirus type 2 (PCV2) genotype definition and their relation with postweaning multisystemic wasting syndrome (PMWS) occurrence. Vet. Microbiol. 2008, 128, 23–35. [Google Scholar] [CrossRef]
- Franzo, G.; Segalés, J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. PLoS ONE 2018, 13, e0208585. [Google Scholar] [CrossRef]
- Wang, Y.; Noll, L.; Lu, N.; Porter, E.; Stoy, C.; Zheng, W.; Liu, X.; Peddireddi, L.; Niederwerder, M.; Bai, J. Genetic diversity and prevalence of porcine circovirus type 3 (PCV3) and type 2 (PCV2) in the Midwest of the USA during 2016-2018. Transbound. Emerg. Dis. 2020, 67, 1284–1294. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.J.; Lu, Y.H.; Wei, Y.W.; Huang, L.P.; Liu, C.M. Porcine circovirus type 2 (PCV2): genetic variation and newly emerging genotypes in China. Virol. J. 2010, 7, 273. [Google Scholar] [CrossRef]
- Jantafong, T.; Boonsoongnern, A.; Poolperm, P.; Urairong, K.; Lekcharoensuk, C.; Lekcharoensuk, P. Genetic characterization of porcine circovirus type 2 in piglets from PMWS-affected and -negative farms in Thailand. Virol. J. 2011, 8, 88. [Google Scholar] [CrossRef]
- Bao, F.; Mi, S.; Luo, Q.; Guo, H.; Tu, C.; Zhu, G.; Gong, W. Retrospective study of porcine circovirus type 2 infection reveals a novel genotype PCV2f. Transbound. Emerg. Dis. 2018, 65, 432–440. [Google Scholar] [CrossRef]
- Weissenbacher-Lang, C.; Kristen, T.; Mendel, V.; Brunthaler, R.; Schwarz, L.; Weissenböck, H. Porcine circovirus type 2 (PCV2) genotyping in Austrian pigs in the years 2002 to 2017. BMC Vet. Res. 2020, 16, 198. [Google Scholar] [CrossRef]
- Trible, B.R.; Rowland, R.R. Genetic variation of porcine circovirus type 2 (PCV2) and its relevance to vaccination, pathogenesis and diagnosis. Virus Res. 2012, 164, 68–77. [Google Scholar] [CrossRef]
- Kwon, T.; Lee, D.U.; Yoo, S.J.; Je, S.H.; Shin, J.Y.; Lyoo, Y.S. Genotypic diversity of porcine circovirus type 2 (PCV2) and genotype shift to PCV2d in Korean pig population. Virus Res. 2017, 228, 24–29. [Google Scholar] [CrossRef]
- Zou, J.; Liu, H.; Chen, J.; Zhang, J.; Li, X.; Long, Y.; Jiang, Y.; Li, W.; Zhou, B. Development of a TaqMan-Probe-Based Multiplex Real-Time PCR for the Simultaneous Detection of Porcine Circovirus 2, 3, and 4 in East China from 2020 to 2022. Vet. Sci. 2022, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; He, K.; Yang, H.; Yu, Z.; Mao, A.; Zhong, S.; Ni, Y.; Zhang, X.; Li, B.; Wang, X.; Zhou, J.; Guo, R.; Lv, L.; Jiang, J. Prevalence of porcine circovirus-like agent P1 in Jiangsu, China. Virol. J. 2011, 8, 543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; He, K.; Wen, L.; Fan, H. Genetic and phylogenetic analysis of a new porcine circovirus type 2 (PCV2) strain in China. Arch. Virol. 2015, 160, 3149–3151. [Google Scholar] [CrossRef]
- Allan, G.M.; Ellis, J.A. Porcine circoviruses: a review. J. Vet. Diagn. Invest. 2000, 12, 3–14. [Google Scholar] [CrossRef]
- Segalés, J.; Domingo, M. Postweaning multisystemic wasting syndrome (PMWS) in pigs. A review. Vet. Q. 2002, 24, 109–124. [Google Scholar] [CrossRef]
- Chae, C. Postweaning multisystemic wasting syndrome: a review of aetiology, diagnosis and pathology. Vet. J. 2004, 168, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Fehér, E.; Jakab, F.; Bányai, K. Mechanisms of circovirus immunosuppression and pathogenesis with a focus on porcine circovirus 2: a review. Vet. Q. 2023, 43, 1–18. [Google Scholar] [CrossRef]
- Lang, H.W.; Zhang, G.C.; Wu, F.Q.; Zhang, C.G. Detection of serum antibody against postweaning multisystemic wasting syndrome in pigs. Chin. J. Vet. Sci. Technol. (in Chinese). 2000, 3, 3–5. [Google Scholar]
- Fan, M.; Bian, L.; Tian, X.; Hu, Z.; Wu, W.; Sun, L.; Yuan, G.; Li, S.; Yue, L.; Wang, Y.; Wu, L.; Wang, Y.; Yan, Z.; Ren, J.; Li, X. Infection characteristics of porcine circovirus type 2 in different herds from intensive farms in China, 2022. Front. Vet. Sci. 2023, 10, 1187753. [Google Scholar] [CrossRef]
- Ma, Z.C.; Liu, M.D.; Liu, Z.H.; Meng, F.L.; Wang, H.Y.; Cao, L.L.; Li, Y.; Jiao, Q.L.; Han, Z.F.; Liu, S.D. Epidemiological investigation of porcine circovirus type 2 and its coinfection rate in Shandong Province in China from 2015 to 2018. BMC Vet. Res. 2021, 17, 17. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.M.; Lu, Q.X.; Wang, F.Y.; Xing, G.X.; Feng, H.; Jin, Q.Y.; Guo, Z.H.; Teng, M.; Hao, H.F.; Li, D.L.; Wei, X.; Zhang, Y.H.; Deng, R.G.; Zhang, G.P. Phylogenetic analysis of porcine circovirus type 2 (PCV2) between 2015 and 2018 in Henan Province, China. BMC Vet.Res. 2020, 16, 6. [Google Scholar] [CrossRef] [PubMed]
- Firth, C.; Charleston, M.A.; Duffy, S.; Shapiro, B.; Holmes, E.C. Insights into the evolutionary history of an emerging livestock pathogen: porcine circovirus 2. J. Virol. 2009, 83, 12813–12821. [Google Scholar] [CrossRef] [PubMed]
- Hamel, A.L.; Lin, L.L.; Sachvie, C.; Grudeski, E.; Nayar, G.P. PCR detection and characterization of type-2 porcine circovirus. Can. J. Vet. Res. 2000, 64, 44–52. [Google Scholar]
- Takahagi, Y.; Nishiyama, Y.; Toki, S.; Yonekita, T.; Morimatsu, F.; Murakami, H. Genotypic change of porcine circovirus type 2 on Japanese pig farms as revealed by restriction fragment length polymorphism analysis. J. Vet. Med. Sci. 2008, 70, 603–606. [Google Scholar] [CrossRef]
- Wen, L.; Guo, X.; Yang, H. Genotyping of porcine circovirus type 2 from a variety of clinical conditions in China. Vet. Microbiol. 2005, 110, 141–146. [Google Scholar] [CrossRef]
- Wang, H.J.; Gu, J.Y.; Xing, G.; Qiu, X.H.; An, S.T.; Wang, Y.X.; Zhang, C.; Liu, C.M.; Gong, W.J.; Tu, C.C.; Su, S.; Zhou, J.Y. Genetic diversity of porcine circovirus type 2 in China between 1999-2017. Transbound. Emerg. Dis. 2019, 66, 599–605. [Google Scholar] [CrossRef]
- Li, N.; Liu, J.; Qi, J.L.; Hao, F.; Xu, L.; Guo, K.K. Genetic Diversity and Prevalence of Porcine Circovirus Type 2 in China During 2000-2019. Front. Vet. Sci. 2021, 8, 788172. [Google Scholar]
- Guo, L.; Fu, Y.; Wang, Y.; Lu, Y.; Wei, Y.; Tang, Q.; Fan, P.; Liu, J.; Zhang, L.; Zhang, F.; Huang, L.; Liu, D.; Li, S.; Wu, H.; Liu, C. A porcine circovirus type 2 (PCV2) mutant with 234 amino acids in capsid protein showed more virulence in vivo, compared with classical PCV2a /b strain. PLoS ONE 2012, 7, e41463. [Google Scholar] [CrossRef]
- Suh, J.; Oh, T.; Park, K.; Yang, S.; Cho, H.; Chae, C. A comparison of virulence of three porcine circovirus type 2 (PCV2) genotypes (a, b, and d) in pigs singularly inoculated with PCV2 and dually inoculated with PCV2 and porcine reproductive and respiratory syndrome virus. Pathogens 2021, 10, 891. [Google Scholar] [CrossRef]
- Oh, T.; Suh, J.; Park, K.H.; Yang, S.; Cho, H.; Chae, C. A comparison of pathogenicity and virulence of three porcine circovirus type 2 (PCV2) genotypes (a, b, and d) in pigs singularly inoculated with PCV2 and dually inoculated with Mycoplasma hyopneumoniae and PCV2. Pathogens 2021, 10, 979. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhan, Y.; Wang, D.D.; Xie, X.H.; Liu, T.B.; Liu, W.; Wang, N.D.; Deng, Z.B.; Lei, H.Y.; Yang, Y.; Wang, A.B. Evidence of natural co-infection with PCV2b subtypes in vivo. Arch. Virol. 2017, 162, 2015–2020. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Zhang, X.W.; Liu, X.H.; Ren, L.Z. Co-infection of swine with porcine circovirus type 2 and other swine viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Yin, L.; Zhu, J.; Li, H.; Zhang, F.; Hu, Q.; Xiao, Q.; Xie, J.; He, K. Nearly 20 Years of Genetic Diversity and Evolution of Porcine Circovirus-like Virus P1 from China. Viruses 2022, 14, 696. [Google Scholar] [CrossRef]
- Zhan, Y.; Yu, W.; Cai, X.; Lei, X.; Lei, H.; Wang, A.; Sun, Y.; Wang, N.; Deng, Z.; Yang, Y. The carboxyl terminus of the porcine circovirus type 2 capsid protein is critical to virus-like particle assembly, cell entry, and propagation. J. Virol. 2020, 94, e00042–e00020. [Google Scholar] [CrossRef]
- Liu, Q.; Tikoo, S.K.; Babiuk, L.A. Nuclear localization of the ORF2 protein encoded by porcine circovirus type 2. Virology 2001, 285, 91–99. [Google Scholar] [CrossRef]
- Lekcharoensuk, P.; Morozov, I.; Paul, P.S.; Thangthumniyom, N.; Wajjawalku, W.; Meng, X.J. Epitope mapping of the major capsid protein of type 2 porcine circovirus (PCV2) by using chimeric PCV1 and PCV2. J. Virol. 2004, 78, 8135–8145. [Google Scholar] [CrossRef]
- Shang, S.B.; Jin, Y.L.; Jiang, X.T.; Zhou, J.Y.; Zhang, X.; Xing, G.; He, J.L.; Yan, Y. Fine mapping of antigenic epitopes on capsid proteins of porcine circovirus, and antigenic phenotype of porcine circovirus type 2. Mol. Immunol. 2009, 46, 327–334. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree based on 251 PCV2 complete sequences used in this study. The tree was constructed with MEGA7.0 software using the neighbor-joining (NJ) method with1000 bootstrap replications.
Figure 1.
Phylogenetic tree based on 251 PCV2 complete sequences used in this study. The tree was constructed with MEGA7.0 software using the neighbor-joining (NJ) method with1000 bootstrap replications.
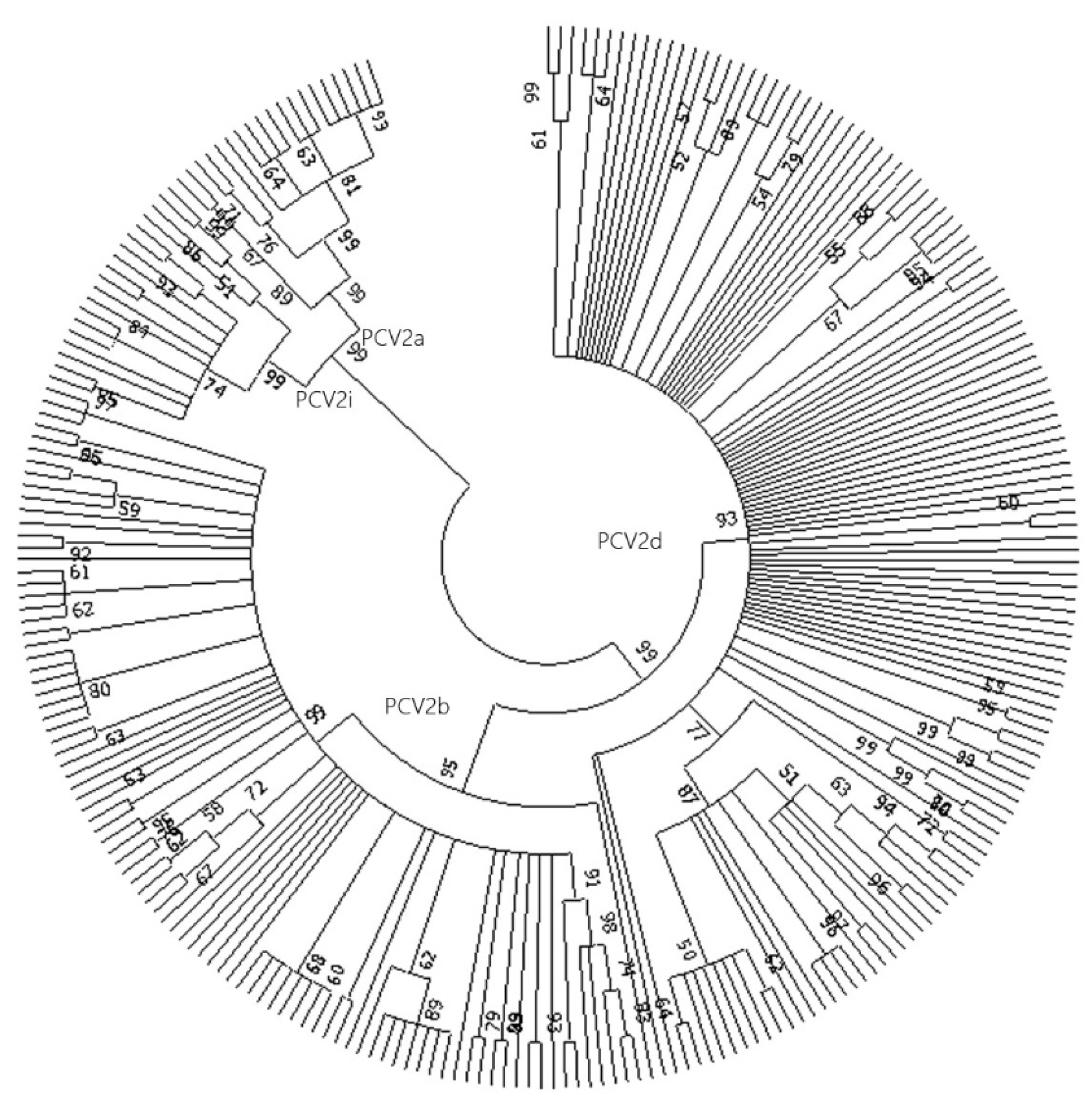
Figure 2.
Proportion of different PCV2 genotypes in 251 strains in Jiangsu from 2014 to 2021.
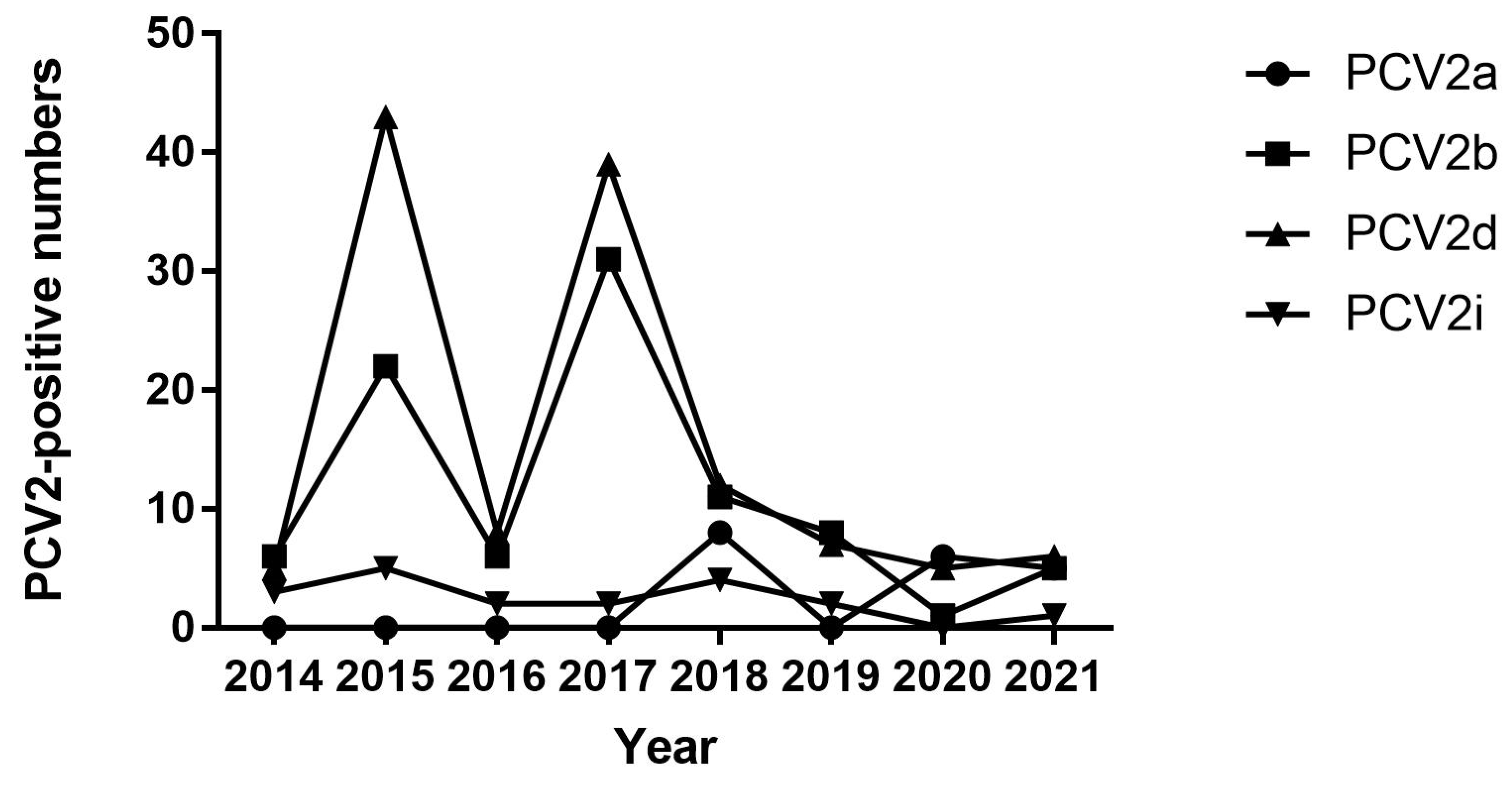

Table 1.
Information of samples obtained from 2014 to 2021 in Jiangsu Province, China.
| Collection Year | Sample type | Sample number | Clinical symptoms |
|---|---|---|---|
| 2014 | lung, serum, diarrheal stool, and lymph node | 408 | PMWS |
| 2015 | lung, diarrheal stool, and lymph node | 553 | PMWS |
| 2016 | lung, diarrheal stool, and lymph node | 459 | PMWS |
| 2017 | lung, serum, diarrheal stool, and lymph node | 463 | PMWS |
| 2018 | lung | 337 | PMWS |
| 2019 | lung, and lymph node | 170 | PMWS |
| 2020 | lung, diarrheal stool | 135 | PMWS |
| 2021 | lung | 150 | PMWS |
Table 2.
Infection rate of PCV2 in 13 cities in Jiangsu Province from 2014 to 2021.
| 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | |
|---|---|---|---|---|---|---|---|---|
| Changzhou | 0.00 | 24.39 | 47.37 | 0.00 | 42.86 | 44.44 | 0.00 | 50.00 |
| Huaian | 26.67 | 30.00 | 14.56 | 30.97 | 15.38 | 66.67 | 36.36 | 68.00 |
| Lianyungang | 58.33 | 65.85 | 32.73 | 48.89 | 22.58 | 71.43 | 47.37 | 62.50 |
| Nanjing | 62.50 | 30.77 | 47.06 | 30.00 | 75.00 | 60.00 | 57.14 | 66.67 |
| Nantong | 54.05 | 26.92 | 35.71 | 65.52 | 25.00 | 87.50 | 50.00 | 56.25 |
| Suzhou | 52.17 | 31.25 | 39.29 | 66.67 | 0.00 | 20.00 | 0.00 | 40.00 |
| Taizhou | 33.90 | 36.17 | 72.73 | 68.29 | 56.25 | 40.00 | 40.00 | 66.67 |
| Wuxi | 100.00 | 53.13 | 88.89 | 0.00 | 66.67 | 0.00 | 40.00 | 33.33 |
| Suqian | 72.22 | 40.28 | 18.95 | 45.45 | 38.89 | 91.67 | 62.50 | 80.00 |
| Xuzhou | 20.00 | 35.71 | 43.75 | 42.42 | 20.18 | 27.27 | 43.75 | 60.00 |
| Yancheng | 17.39 | 62.50 | 20.00 | 33.33 | 45.45 | 100.00 | 41.67 | 55.56 |
| Yangzhou | 76.19 | 66.67 | 56.52 | 20.00 | 33.33 | 100.00 | 85.71 | 80.00 |
| Zhenjiang | 63.16 | 47.62 | 100.00 | 100.00 | 27.89 | 0.00 | 50.00 | 50.00 |
Table 3.
Nucleotide sequence differences among 9 genotypes of PCV2. This study included 251 full-length sequences of PCV2 obtained in this study and eight complete sequences representing all eight PCV2 genotypes (a-h) retrieved from GenBank.
Table 3.
Nucleotide sequence differences among 9 genotypes of PCV2. This study included 251 full-length sequences of PCV2 obtained in this study and eight complete sequences representing all eight PCV2 genotypes (a-h) retrieved from GenBank.
| Nt position | PCV2a | PCV2b | PCV2c | PCV2d | PCV2e | PCV2f | PCV2g | PCV2h | PCV2i |
|---|---|---|---|---|---|---|---|---|---|
| 617 | C | C | C | C | C | C | C | C | T |
| 1164 | C | C | C | C | C | C | G | G | T |
| 1596 | G | G | G | G | G | G | G | G | C |
| 1604 | G | G or T | G | G | G | G | G | G | A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



