
Submitted:
28 August 2024
Posted:
29 August 2024
You are already at the latest version
Abstract
Somatic embryogenesis (SE) provides alternative methodologies for the propagation of grapevine (Vitis spp.) cultivars, conservation of its germplasm resources and for crop improvement. In this review, the current state of knowledge regarding grapevine SE as applied to these technologies is presented, with a focus on the benefits, challenges, and limitations of this method. The paper provides a comprehensive overview of the different steps involved in the grapevine SE process, including callus induction, maintenance of embryogenic cultures, and the production of plantlets. Additionally, the review explores development of high-health plant material through SE, the molecular and biochemical mechanisms underlying SE, including the regulation of gene expression, hormone signalling pathways, and metabolic pathways as well as its use in crop improvement programmes. The review concludes by highlighting the future directions for grapevine SE research, including the development of new and improved protocols, the integration of SE with other plant tissue culture techniques, and the application of SE for the production of elite grapevine cultivars, for the conservation of endangered grapevine species as well as for cultivars with unique traits that are valuable for breeding programs.
Keywords:
Germplasm
; Mutagenesis
; in vitro culture
; Tissue culture
; Micropropagation
; Conservation
; Somaclonal variation
; Transformation
; Chimera
; Virus
1. Introduction
The lack of motility and the resulting inability of plants to escape from predators, parasites and at the changes of surrounding environment has led to the development of very efficient defence strategies. Plants are generally highly plastic organisms compared with animals, being able to modulate their development depending on endogenous and environmental signals, even reprogramming the fate of somatic cells. At the basis of this ability is the mechanism of totipotency that is observed in plant cells [1]. Cell fate reprogramming is complex and frequently associated with significant changes in chromatin status. Chromatin change is characterized by DNA methylation and histone chemical modifications, mainly methylation or acetylation [2]. The early observations on the capacity that plants have to react to tissue injury by leading fully mature differentiated cells to change their function, thus favouring dedifferentiation and the subsequent regeneration of damaged tissue, led to the pioneering studies of plant tissue and cell culture in vitro. The first theoretical basis for plant tissue culture date back to the early 1900s when Gottlieb Haberlandt [3] observed that the cells of plant tissues cultured in vitro survived and increased in volume. However due to lack of adequate culture medium containing phytohormones such as 3-indole-acetic acid (isolated in 1885 by the chemist Salkowski) he failed to observe cell division under the experimental conditions used in his study [4]. Haberlandt hypothesized that a single cell is a living unit, an individual in itself that is to some extent independent of the whole organism [5]. This led to the idea that a single cell could be capable of giving origin to a complete and functional plant [6]. Direct evidence supporting this hypothesis has been lacking until 1958, when Steward et al. [7] showed that carrot segments of secondary phloem tissue cultured in vitro in appropriate conditions were able to regenerate whole plants.
The dedifferentiation of plant cells has long attracted interest as a key process for understanding the plasticity of plant development. These studies led to the hypothesis that many mature plant cells retain totipotency and related dedifferentiation to the initial step of the expression of totipotency. Considerable progress was achieved after discovery of the hormonal control of cell proliferation and organogenesis in vitro in the 1950s [8]. These studies have allowed to identify efficient in vitro regeneration protocols based on organogenesis and somatic embryogenesis (SE). If somatic cells are stimulated to generate cells with embryogenic potential, the new cells can give rise to structures capable of regenerating a complete plant [9].
SE is a process by which plants can produce bipolar structures from a single somatic cell without meiosis and fertilization, therefore the new plant derived from a somatic embryo is thus genetically identical to the mother plant. This complex process can follow two paths, called direct (from a single somatic cell) and indirect (from undifferentiated cells) embryogenesis. However, it is difficult to distinguish between the two routes which can sometimes occur simultaneously from the same explant. The most common route is indirect SE and begins through the typical formation of a callus, an apparently disorganized mass of cells showing varying degrees of compactness [10]. During this process of dedifferentiation and differentiation of plant cells, the explant responds to endogenous and exogenous stimuli, which trigger the induction of a signalling response and, consequently, can often profoundly modify the cell fate. For the understanding of this important plant regeneration model, the interactions between the different plant growth regulators (PGR), mainly auxins, cytokinins (CKs), ethylene and abscisic acid (ABA), during the induction of SE are of fundamental importance [11]. Usually a single somatic plant cell or a set of differentiated and specialized somatic cells with specific functions must receive a stimulus from a set of phytohormones, perceive it, and then trigger the transduction to the nucleus where the specific regulatory and structural genes will be transcribed and subsequently will be translated into proteins involved in the differentiation that ultimately will lead to the regeneration of a new somatic embryo [12]. SE allowed the development of an increasing number of practical and scientific applications. For example, it has the potential for the genetic and sanitary improvement of genotypes of commercial importance, as well as providing insights into the underlying mechanisms of biological processes [13]. Furthermore, the application of the most modern CRISPR-derived biotechnologies that have revolutionized the genetic engineering field, in many crops is limited by the lack of efficient in vitro plant regeneration protocols [14]. Finally, the preservation of germplasm through SE is an efficient method of conservation at reduced cost for those species which cannot be propagated through seeds [15]. Our purpose is to provide a comprehensive and updated overview of the application of SE in grapevine as well as critically discuss and highlight the future perspectives and challenges.
2. Explant Sources and Stages of Somatic Embryogenesis in Grapevine
One of the main constraints influencing the different applications of in vitro SE as a tool for plant regeneration is the low embryogenic potential of many crops and genotypes. A plant species/genotype, a tissue or a developmental phase of a plant is termed recalcitrant if commonly used protocols fail to regenerate somatic embryos in vitro [10]. This recalcitrance affects not only embryo differentiation but also the subsequent steps in the regeneration process from embryo germination to plantlet acclimatization in vivo. Although SE can be induced from a range of tissues, the correct choice of the type of explant is of fundamental importance [16]. In fact, it is important to determine first which part of the plant contains the most responsive tissues and at what stage of development and time of the year they must be collected. In grapevine, the best results are usually obtained with explants of floral origin such as whole flowers, anthers, filaments, stigmas/styles, ovaries and pistils (Figure 1 A-B). The age of the cells is also important in different species, usually younger cells have been reported as those in the most responsive state to induce embryogenic cultures [17,18,19].
The morphological and temporal development of somatic embryos proceed through a series of distinct stages, with globular, heart, torpedo, and cotyledon or plantlet stages for dicotyledons [20,21] and globular, elongated, scutellar, and coleoptilar stages for monocotyledons [22]. In grapevine, embryos go through distinct phases such as globular, heart, torpedo, and early cotyledon before germinating (Figure 1 E) [18].
Globular embryos usually appear on the surface of the embryogenic calli (Figure 1 C-D). The young embryo is circular or slightly oblong with small cells having thick walls and is in close contact with the callus from which it was generated [23]. When the embryo detaches from the callus, axial cells start to elongate marking the beginning of tissue differentiation process. The two apical meristems present by the end of the globular stage persist through the maturation step during which embryos pass from heart-shaped to the torpedo stage and cotyledons expand due to the deposition of storage materials [24,25]. Mature embryos having accumulated enough storage materials develop into normal plants passing from the torpedo stage to the germinated embryo [26]. Grapevine somatic embryos show radicle growth, tannin accumulation in the central cylinder and acquisition of an external suberin sheath [27,28].
3. Synchronisation of Somatic Embryo Production and Their Germination
Automation and scaling up are needed to improve cost-effectiveness in every process. To maximise the output of SE and lower unit costs through automation, it is imperative to synchronise the growth of somatic embryos in embryogenic suspension cultures. Synchronisation of somatic embryo production is highly desirable for applications in micropropagation, genetic transformation and for gene expression studies related to SE.
Jayasankar et al. [29] compared somatic embryo development on solid media with those cultured in liquid media. On agar-based medium, somatic embryos had large cotyledons, a negligible or absent suspensor structure, and a relatively undeveloped concave shoot apical meristem, whereas those growing in liquid medium showed a distinct suspensor, and a flat-to-convex shoot apical meristem enclosed in smaller cotyledons. Only the somatic embryos grown on solid media exhibited dormancy. According to their hypothesis, somatic embryos cultured in liquid media have a lasting suspensor that modifies development, leading to fast germination and a high rate of plant regeneration [29].
In an attempt to synchronise growth of somatic embryos, Jayasankar et al. [30] established suspension cultures of V. vinifera ‘Thompson Seedless’ and ‘Chardonnay’ incorporating 2,4-dichlorophenoxyacetic acid (2,4-D) and 6% sucrose. Cultures originated from anthers and ovaries of Chardonnay and leaves of ‘Thompson Seedless’. After two subcultures, the suspension consisted of differentiated somatic embryos, single cells and small cell aggregates composed both of embryogenic and non-embryogenic cells and started to turn brown. To synchronise the development of somatic embryos they decanted about 50% of the supernatant and the rest was swirled and quickly filtered through a 960 µm sieve. The fine fraction collected was re-filtered through a double-folded, Kimwipe™ in a funnel. About 100 mg of the fine fraction that adhered to the Kimwipe™ was resuspended in 40 ml fresh liquid medium and was subcultured at 14-day intervals. Sieving of proembryogenic masses (PEMs) and subculture resulted in the synchronization of embryo development and reduced browning and abnormalities such as fasciation or fusion during differentiation. After 4–6 weeks in liquid medium without 2,4-D, globular and early-heart stage somatic embryos appeared. After 8 weeks of culture of Chardonnay in medium devoid of 2,4-D resulted in somatic embryos with well-developed cotyledons. After 12–14 weeks in liquid medium without 2,4-D, ‘Chardonnay’ somatic embryos turned green and germinated, whereas ‘Thompson Seedless’ did not advance beyond the heart stage. Jayasankar et al. [30] demonstrated the normal development of ‘Chardonnay’ in two clones (01Ch and 02Ch). On the other hand, when 2-naphthoxyacetic acid was used in liquid culture, PEMs of ‘Chardonnay’ clone 76 showed poor competence for further development when the auxin was removed [31]. However, growth and development could be stimulated by daily subcultures and the authors attributed the arrest of development under standard subculture conditions to the accumulation of extracellular macromolecules of molecular weight > 10 kDa [31]. When embryogenic competent ‘41B’ (V. vinifera cv. Chasselas × V. berlandieri) cultures were compared with ‘Chardonnay’ clone 76 that show arrest at heart stage, the protein patterns in auxin enriched culture media were practically identical. When the auxin was removed, extracellular proteins of 38, 51 and 62 kDa were over accumulated in the CH76 cell culture compared to the 41B cell line, whereas 36- and 48-kDa proteins were excreted only by the 41B cell line. These differences were attributed to the differences in embryogenic competence in the two cell lines [32]. On the other hand, Zlenko et al. [33] successfully converted somatic embryos that had been grown on liquid induction media by subculturing them on liquid media that had either GA3 alone or a combination of BA and GA3 added. Solid media, either with or without BA, was successfully employed to promote plant development. Also Vasanth and Vivier [34] and Wang et al. [35,36] used liquid medium to produce synchronized somatic embryos for cryopreservation procedures.
Several factors influence rate of conversion of somatic embryos, and abnormalities can be due to genetic or epigenetic changes in the DNA. Stress factors such as high and low temperatures, drought, salt, and heavy metals, usage of mutagenic chemicals and plant growth regulators can affect modifications in DNA [37]. Among others, abscisic acid metabolism has been reported as crucial in the maturation of grapevine somatic embryos [38]. Despite these differences, a significant benefit is the capacity to sustain embryogenic cultures over an extended period of time without losing their embryogenic potential, as demonstrated by numerous research [39].
4. Factors Affecting Somatic Embryogenesis in Grapevine
SE in grapevine is influenced by several factors, both internal and external, which makes the application of the technique challenging. Embryogenesis processes are notably impacted by the selection of the appropriate explant, medium, phytohormones, genotype, carbohydrate, and gelling agent, among other factors including light regime, temperature, and humidity [15,40,41].
4.1. Genetic Control
SE competence is strongly genotype dependent. The embryogenic potential of cultivars varies considerably, and although multiple methods have been published, for certain cultivars the technique still needs further improvement [42,43]. Several authors have highlighted the different response to SE of grapevine according to genotype [15,39,43,44,45,46]. It is commonly recognised that genotypes used as rootstock have a stronger capacity for regeneration through both organogenesis and SE than V. vinifera hybrids and cultivars. For example, among three different Italian V. vinifera L. cultivars and four hybrid rootstocks, SE efficiency was higher for rootstocks irrespective of the medium and explant used [47]. In another Italian study, grapevine genotype was the key factor with a 50-fold difference in the percentage of somatic embryogenesis among eight Italian grapevine cultivars [15].
SE in grapevine is governed by complex gene signalling networks involving transcriptional regulation, protein signalling, and extracellular matrix interactions. The genes expressed during grapevine SEs have been studied, particularly the Somatic Embryogenesis Receptor Kinase (SERK) and Leafy Cotyledon (LEC and L1L) genes [48]. These genes play important roles in SE in various plant species. The expression of VvSERK1, VvSERK2, VvSERK3, and VvL1L genes has been analyzed during SE in grapevine. The results showed that these genes are involved in the regulation of SE in grapevine, with expression of VvSERK2 relatively stable during in vitro culture, VvSERK1, VvSERK3 and VvL1L are expressed more 4 to 6 weeks after transfer of the calli onto embryo induction medium before the appearance of the embryos on calli. After 8 weeks in the embryo induction media VvSERK1 is expressed in the calli and VvSERK3 in the embryos. Expression of VvL1L was low at this time [48]. Thus the differential expression of key genes, such as SERK and LEC1-like, is crucial for the embryogenic process, as these genes play pivotal roles in promoting somatic embryo formation through stress and developmental signaling pathways [49]. Additionally, lipid-transfer proteins (LTPs), secreted during somatic embryogenesis function as extracellular signaling molecules that are vital for proper cell to cell communication and membrane dynamics [50]. Overexpression of the VvLTP1 gene, however, disrupts normal embryo development, indicating that precise regulation of LTPs is essential for maintaining the balance of signaling necessary for embryogenesis [51]. Proteolytic regulation, facilitated by extracellular proteins and protease inhibitors, further influences the embryogenic process by modulating the extracellular matrix and thereby affecting gene signaling [52]. Moreover, distinct extracellular protein patterns observed in different embryogenic states underline the significance of the extracellular environment in shaping gene expression and subsequent developmental outcomes [32]. These studies collectively highlight the intricate interplay between intracellular signaling and extracellular factors in regulating somatic embryo formation in grapevine.
4.2. Other Factors Controlling Somatic Embryogenesis
The response to somatic embryo induction can also vary based on the organ/tissue types. Anthers, ovaries, leaves, petioles, tendrils, and nodal sections are the explants that are most frequently used for SE induction in grapevine [13]. A remarkable number of genotypes have been regenerated through anther culture [20] with a high success rate. Nevertheless, some authors report that, depending on the stage of growth of reproductive organs, the percentage of SE can change. For instance, according to Vidal et al. [53], the regeneration from the ovaries were around two times greater than that from the anthers when ovaries were cultured in later stages of development. According to a recent study by San Pedro et al. [54] mature seeds can also be used as explants for SE induction. However, the study indicated that the only way to obtain somatic embryos is to cut seeds and hold them for five months in media supplemented with thidiazuron (TDZ). However, seed-derived somatic embryos are not useful for clonal propagation. Embryogenic callus induction has also been achieved using nodal segments, leaf discs [55] petioles, stem nodal explants[56,57] and whole flowers [58] even if less commonly used.
It is widely known that the developmental stage of the explant affects the effectiveness of SE induction, and the physiological stage of the primary explant also has considerable influence on the success of the protocol [53,59]). The first factor to consider while choosing anthers at a particular developmental stage is the size of the floral buds. It has been reported that buds 1.55 cm long on average have anthers enclosing uninucleate microspores, which are the most responsive to SE in Vitis. Moreover, in proportion to their stage of differentiation, explants' ability to alter their evolutionary trajectory diminishes [60] and this seems to be the case both for carpels and for stamens [59]. Similar conclusions have been drawn by other authors [2,15,44]. Vidal et al. [61] showed that earlier flower developmental stages are more conducive for embryogenic culture induction from anthers, while later stages did so from ovaries. Three flower developmental stages were classified by Prado et al. [41]. R1 and R2 are equivalent to stages V and VI according to Gribaudo et al. [45], while R3 is late binucleate microspore stage. The authors reported that two cultivars — Mencía and Brancellao — were best utilised at the R3 stage, whereas four cultivars Albaríio, Treixadura, Torrontés, and Merenzao recorded the best results at the R2 stage [41]. Similarly, two V. vinifera cultivars ‘Chardonnay’ and ‘’Barbera’ responded better when anthers of early stage microsporogenesis were cultured, whereas for the rootstock ‘110 Richter’ (V. berlandieri x V. rupestris) more embryogenic cultures could be established using explants in the later stages of maturation [45].
The composition of the culture medium has a significant role in success of plant regeneration because it supplies essential nutrients for the growth of explants at various developmental stages. Usually the media adopted for inducing SE in vitro are based on MS [62] or NN [63] salts. However, media formulations differ among different laboratories and many different types of basal culture media have been tested such as LS [64], WPM [65], C2D [66] and DKW [67]. Focused research on the effects of micro and macronutrients is rarely reported. Nevertheless, it is known that ammonium promotes SE induction in some media [68]. The only carbohydrate supply used for embryogenic culture, SE induction, and development is sucrose at 10 to 180 g L–1 and being most widely used at 30 – 60 g L–1. Sucrose plays an important role also as osmoticum for SE germination and plant regeneration because dehydration of grapevine SE may increase plant development [69]. To improve grapevine regeneration process, some other protocols suggest supplementing media with amino acids such as glycine, phenylalanine, and L-glutamine [70,71].
The production of an embryogenic callus has been shown to be significantly influenced also by the type and concentration of plant growth regulators (PGRs) such as 2,4-D, N-(2-chloro-4-pyridyl)-N´-phenylurea (4-CPPU), 6-benzylaminopurine (BAP), gibberellic acid (GA3 ), indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), naphthalene acetic acid (NAA), 2-naphthoxyacetic acid (NOA), and thidiazuron (TDZ) [72]. Usually, a combination of auxins, mainly 2,4-D or NOA, and cytokinins, mainly BAP, added at different concentrations based on the type of explant and genotype is used to initiate embryogenic cultures. Some combinations that have been used to induce SE successfully include IAA combined with GA3 for fertilized ovules and urea derivatives like TDZ or 4-CPPU in combination with auxins in the induction phase in anther culture[73]. Continuous presence of PGRs is not suitable for somatic embryo development. For this reason, after embryogenic callus has been induced, in some cases auxins are removed, decreased or substituted with other PGRs. A comprehensive list of PGR combinations used for SE induction in grapevine can be found in Carimi et al. [74].
Also, physical culture conditions could significantly influence SE induction and regeneration frequency. Some authors indicate that a two-week culture period in the dark is useful for improving regeneration percentages [75]. Additional treatments aimed at enhancing regeneration efficiency include the use of activated charcoal [76], pre-treatment with chilling [77], cotyledon removal [78,79] and adjusting pH levels [80]. The kind of culture—liquid or solid—also can affect the outcome; in the initial induction phase, liquid cultures are preferable, but organised embryogenic callus develops more readily on solid medium [81].
Liquid suspension cultures are generally more efficient than solid media because plant cells are better exposed to the medium components and the uptake and consumption of nutrients are faster. Liquid cultures allow for a higher cell growth rate and are more effective in regenerating somatic embryos. However, they are considered more complex when compared to cell cultures on solid media because they require shorter subculture intervals (tend to senescence earlier) and are more susceptible to bacterial and fungal contamination. Liquid suspension cultures of grapevine are generally started from about 200-400 mg of PEM incubated in 50 ml of liquid culture medium (Figure 2 A-B) and after about two months new somatic embryos are regenerated (Figure 2 C). The embryos are separated from the undifferentiated cells by filtration of culture using a nylon mesh filter and then incubated for about 30 days on liquid medium without plant growth regulators (Figure 2 D) and subsequently placed to germinate on solidified medium (Figure 2 E). Within 4-6 months from the culture initiation of PEMs it is possible to have acclimatized plants (Figure 2 F).
5. Applications of Somatic Embryogenesis and Embryogenic Cultures in Vitis
SE possess a wide array of potential applications in micropropagation, germplasm conservation, sanitary and genetic improvement, including the most modern genetic engineering techniques. Herein, we summarize the state of the art and the impact that SE may have on grapevine propagation, conservation and crop improvement.
5.1. Somatic Embryogenesis for Germplasm Management
As a clonally propagated heterozygous species, it is not possible to conserve grapevine clones used in wine making through seed banking. This same principle applies to rootstocks as these have also been selected for performance. For example, in France there are 15 certified clones of Richter 100 rootstock [82]. Long-term conservation of vegetative tissue is not as easy as seeds and therefore grapevine germplasm is maintained as field collections in many countries [83,84,85] leading to the erosion of valuable germplasm resources [86,87,88]. As an alternative, grapevine clones are maintained in slow-growth media in tissue culture [89,90,91], including rootstocks [92] and hybrid material used in breeding [93]. With SE as a better option due to its easier handling and storage, prolonging the lifespan of somatic embryos is important, and some of the outcomes from traditional shoot-based tissue cultures may apply to somatic embryos as well. It is important to reduce growth rate in tissue cultures used for conservation to prolong the period between subcultures, as frequent subculture has higher risk of contamination, mislabelling as well as higher labour and material input. Long-term storage has been facilitated by increasing sucrose levels to 7.5%, eliminating plant growth regulators from media and storage under low temperatures of up to 2°C [94,95,96], but Pedro et al. [90] report slowing the growth rates by halving the sucrose concentration in media. Gradual decrease of temperature over a few days facilitates better survival compared to material transferred to low temperature abruptly [96]. Addition of 1.5 g L-1 of sorbitol or mannitol also reduced the growth of cultures [97]. However, Hassan et al. [98] report reducing sucrose concentration from the standard 30 g L-1 to 20 g L-1 and including 10 gL-1 sorbitol. Depending on the cultivar, they have successfully used up to 50 gL-1 sorbitol enabling maintenance for one year without subculture. Compared to shoot cultures, longer periods of storage can be achieved using somatic embryos as the conservation propagule. Jayasankar et al. [81] demonstrated that by drying suspension culture-derived somatic embryos to 25% of their initial weight over a laminar hood and storage in tightly sealed Petri dishes at 4 °C can extend the storage time to 42 months. In contrast, in another experiment, plant recovery rate was only 32 % after 21 days of dehydration of somatic embryos under 70 % humidity [99]. As SE is so far the only pathway for genetic transformation in grapevine [100],our ability to maintain long-term viability of somatic embryos is important not only for conservation purposes but also for grapevine transformation providing an uninterrupted supply of plant material. The difficulty of storage of somatic embryos, unlike true seeds, is because the somatic embryos lack desiccation tolerance. By mimicking the process of acquisition of desiccation tolerance during sexual seed development, Senaratne et al. [101] were able to produce alfalfa somatic embryos which can be dried to 8 – 15 % moisture without losing viability. For this, they incorporated abscisic acid (ABA) in the media during the cotyledonary stage of development in a synchronised system. Hence, to achieve this goal of extending the shelf-life of grapevine somatic embryos, research on synchronisation of SE process should go alongside the research on acquisition of desiccation tolerance. Faure et al. [102] showed that V. vinifera ‘Grenache noir’ does not have a peak of ABA mid-embryogenesis. This cultivar shows precocious germination. They hypothesised that the switch from mid- to late-embryogenesis is not triggered because of low endogenous levels of ABA and suggested exogenous application of ABA to prevent precocious germination and trigger late-embryogenesis. Later, Goebel-Tourand et al. [103] showed that exogenous application of ABA can improve the maturation process of grapevine somatic embryos reducing precocious germination. Gene expression studies in maturing grapevine somatic embryos has demonstrated the involvement of ABA biosynthesis on precocious germination vs proper maturation [38].
Cryopreservation is considered the best method of storing germplasm efficiently and safely for long-term, particularly for the conservation of vegetatively propagated species [104,105]. Two decades ago two-step cooling procedures were successfully used to cryopreserve grape embryogenic cell suspensions, using both encapsulation vitrification [36,106] and encapsulation dehydration [35,106,107,108] methods. For example, using a two-step freezing procedure (-0.5 °C min-1 to -40 °C followed by immersion in liquid nitrogen) for embryogenic cells pretreated for 1 h with 0.25 M maltose and 5% dimethyl sulfoxide at 0 °C Dussert et al. [109] achieved a 60% survival rate of cryopreserved somatic embryos. Different modifications for encapsulation vitrification and encapsulation dehydration were also studied during this period. Gonzalez-Benito et al. [110] achieved 45-60% viability by cryopreserving embryogenic cells encapsulated in alginate beads and cultured in liquid media with increasing sucrose concentration (0.25, 0.5, 0.75, 1 M, one day on each step) followed by desiccation in the air flow of a laminar flow cabinet for 2-4 h. Similar procedure with an additional two days of incubation in 1 M sucrose resulting in desiccation of cells to 20.6% moisture resulted in about 78 % viability [35]. Solid media were better than liquid for post-thaw regeneration [35]. In encapsulation vitrification, the dehydration of cells in laminar flow cabinet or using silica gel in encapsulation dehydration is replaced using a vitrification solution. After preculture step in 0.75 M sucrose, Wang et al. [36] encapsulated the embryogenic cells and used Plant Vitrification Solution 2 (PVS2) developed by Sakai et al. [111] to dehydrate the alginate beads. They achieved 42-76% regrowth when the beads were treated with PVS2 solution for 270 min at 0 °C.
With the development of vitrification methods, particularly droplet and cryo-plate methods, cryopreservation of cells and other tissues has become easier and applicable to many species including grapevine [112,113,114]. Droplet vitrification is a simpler method applicable to a wide range of species. Recent studies with several grapevine genotypes for cryopreservation by droplet vitrification revealed that somatic embryos are more amenable to cryopreservation than shoot tips and axillary buds from in vitro grown plantlets [112]. Furthermore, Carimi et al. [115] were able to induce somatic embryos from pistils and anthers of the progenitor of cultivated grapevine, V. vinifera ssp Sylvestris and used axillary buds from germinated somatic embryos for cryopreservation by droplet vitrification with a success rate of up to 44%. Although somatic embryos have not been used for cryopreservation for germplasm conservation of Vitis, shoot tip cryopreservation experiments indicate similar outcomes from droplet and cryoplate vitrification methods. However, as cryoplate-based techniques use alginate beads, plant regeneration takes longer than shoot tips cryopreserved using droplet vitrification [116]. Widely observed differential response among genotypes [15,46,68,117] and interactions of genotype with explant and media [43,68,118] can pose challenges in the use of somatic embryos as the explant source for conservation of large collections. Nevertheless, according to some focused studies, media optimisation for multiple genotypes is possible [119]. Another barrier to the use of somatic embryos as a propagule for conservation of vegetatively propagated species such as grapes is the possibility of separation of chimeras in cultivars that have cell layers of different genetic background [120,121,122] as well as somaclonal variation [13,123,124]. Yet, other studies show genetic integrity of somatic embryo-derived plants using molecular markers [15,125]. Further studies on somaclonal variation during SE in grapevine are needed to understand the reasons, particularly the effect of genotype, plant growth regulators used and the duration of cultures.
5.2. Somatic Embryogenesis as a Tool for Sanitation
The vegetative propagation and exchange of budwood among grapevine growing regions and countries contribute to the spread of grapevine pathogens. Perennial life cycle results in the spread of these diseases within vineyards. With 86 different virus species known to infect Vitis spp., grapevines host the most viruses among cultivated species [126]. Among these, fanleaf and leafroll diseases are the most damaging and widespread [127]. It has been estimated that fan leaf disease caused by a nepovirus (Grapevine fanleaf virus) causes economic losses amounting to US$ 16,600 per ha, and in France where about 2/3rd of the vineyards are affected, economic impact of at least US $1.5 billion per year [127]. Among the five serologically distinct Closteroviridae viruses known to cause leafroll disease, Grapevine leafroll-associated virus 3 (GLRaV 3) is the most devastating. Leafroll disease is estimated to cause losses from US $25,000 to US $ 226,000 per ha over a 25-year vineyard lifespan depending on the location and cultivar [127]. Therefore, establishment of vineyards free of damaging grapevine viruses is an important control measure. Towards this many countries have sanitary selection programmes and certification of clonal stocks. However, once the stock is infected, it is important to have robust methods to eliminate the infecting viruses.
Several methods have been applied to eliminate viruses from infected grapevine clones. Traditionally heat therapy has been used to reduce the viral load, but some viruses are heat stable [128]. Although heat therapy is useful in reducing the incidence of the disease, when used alone it is often not useful for clean stock programmes. Therefore, combination of heat therapy with microshoot culture is often used in grapevine virus eradication programmes [128], particularly for nepoviruses such as GFLV that readily infect even the meristem [129]. A grapevine microshoot consists of the meristem and 2-3 leaf primordia and is less than 0.5 mm [128]. However, for reasons such as persistence of virus particles (e.g. GLRaV 3) in lower parts of the apical dome in 0.5 mm microshoots [130], some viruses infecting even the apical dome [131,132], possible cross-contamination during excision combined with difficulty in precise excision of microshoots [133] and poor regeneration of microshoots in some cultivars [134] have led to the emergence of more precise in vitro-based methods for virus eradication in horticultural species, including grapevine. These include cryotherapy [130,133,135,136], electrotherapy [135], in vitro chemo [137,138] and thermotherapy [139,140] applied separately or in combination.
Similar to other in vitro-based therapies, regeneration from somatic embryos has also become an important tool to eliminate viruses from infected grapevine clones. High efficiency of virus eradication through SE can be explained by the single cell origin or from a few organised embryogenic cells that lack vascular connections to the maternal tissue during SE [29,141]. Secondary somatic embryos are generally attached to the root primordia of the parent embryo by a suspensor-like structure, again without any vascular connection with maternal tissue or with one another [142]. Nevertheless, Goussard et al. [143] were able to demonstrate only the elimination of leafroll associated viruses but not GFLV in somatic embryo-derived plantlets originally initiated from ovaries. When somatic embryos were produced at 35 °C (thermotherapy) in the dark, Goussard and Wiid [144] were able to remove GFLV in addition to leafroll viruses. Plantlets derived from somatic embryos produced at 25 °C were still infected with GFLV [144]. Nepoviruses such as GFLV can readily invade plant meristems [129]. Using three GFLV infected Italian cultivars, Gambino et al. [129] demonstrated the presence of the virus in all tested anthers and ovaries and the callus derived from both these explants. Nevertheless, only few somatic embryos of one cultivar and only one out of 63 plants tested during micropropagation of somatic embryo-derived plantlets tested positive while all the tested plants after one or two dormancy periods in the greenhouse were negative for the virus [129]. Similar results were reported for three V. vinifera cultivars infected with GLRaV-1, GLRaV-3, GVA and GRSPaV. After four months of culture initiation, higher infection rate was reported in ovary cultures compared to anther cultures, but after eight months of culture none of the tested cultures were positive for any of the viruses with similar results for individual somatic embryos tested. All regenerated plantlets (after 12 months of culture initiation) and .greenhouse plants (after 24 months of culture initiation) were free of the viruses [145]. High-throughput sequencing and RT-PCR have been used to compare the efficiency of SE and meristem culture for elimination of several viruses in grapevine [146]. The results showed that SE using anthers with filaments as explants was effective for eliminating various grapevine viruses, including grapevine rupestris vein feathering virus (GRVFV), grapevine Syrah virus 1 (GSyV-1), Grapevine virus T (GVT), and grapevine Pinot gris virus (GPGV) [146]. Rapidly proliferating cells and embryoids originating from these may escape infection [145] or it is possible that embryogenic callus may originate from virus-free cells of the explant [142].
In conclusion, plant regeneration through SE from different explants of floral origin can be used to establish healthy grapevine stocks, free from a number of grapevine viruses. Sanitation through SE is technically more difficult and time consuming than traditional sanitation protocols [39]. Nevertheless, this technique is highly successful in grapevine. In the case of chimeric cultivars, virus elimination may be achieved either by traditional meristem tip culture and thermotherapy or by cryotherapy, sometimes requiring combination of therapies [130,133,135,147].
5.3. Induced Mutagenesis for Grapevine Improvement
Many horticultural species including grapevine are maintained through vegetative propagation over multiple cycles. While this practice helps preserving superior agronomic traits in the cultivars, accumulation of somatic mutations results in phenotypic diversity. The mutants can be selected and propagated as new clones of the mother variety. This diversity in traditional cultivars is the basis for the selection of improved clones without losing varietal identity for the very traditional wine industry. A good example of such selection is the grape cultivar ‘Benitaka’ (red berries) that was selected from ‘Italia’ (green berries).The cv. ‘Brazil’ (black berries) was then selected from ‘Benitaka’ [148]. A sequence analysis of the promoter region and coding sequence of VvmybA1 revealed a base substitution between ‘Benitaka’ and ‘Brazil’ in the promoter region and a deletion of a large DNA fragment in the promoter region of ‘Italia’. Anthocyanin content and expression of the VvmybA1 and UFGT genes in ‘Brazil’ were higher than in ‘Benitaka’ and barely detectable in ‘Italia’ [148]. Economically important clones of ‘Pinot noir’, ‘Cabernet sauvignon’ and ‘Chardonnay’ have been the result of clonal selection. Pinot is one of the oldest grape cultivars and is a noble cultivar used in many countries in different continents including French Champagne and Bourgogne wines [149,150]. It displays extensive clonal diversity and, in France alone 64 different Pinot clones have been certified and marketed. Furthermore, approximately 95% of grapevine plants produced in French nurseries originate from clonal selection [149]. While naturally occurring mutations are able to produce agronomically valuable clones for selection, this process is slow [151] and not ideal for a breeding program.
Induced mutagenesis can increase the frequency of mutations in genomes [151,152]. Although more than 3300 induced mutants have been registered and published [151], the number of mutant cultivars in horticultural species is very limited [151,152]. Unlike in seed propagated species, mutant selection in vegetatively propagated crops is not straight-forward. Using either ionizing radiation or mutagenic chemicals it is possible to induce mutations in planting material, including tissue cultured plantlets. However, in vegetatively propagated plants, following mutagen treatment, several cycles of propagation are needed to obtain homo-histonts or to ‘dissolve’ chimeras, and to obtain ‘solid’ mutants [153]. This is because, the meristem is multicellular and the cell with desired mutation produces a sector, resulting in a chimera. The in vitro subculture of mutagen treated material through several generations can be achieved more rapidly than grafting or rooting of cuttings in classical vegetative propagation of grapevine. Even then, the resulting mutant is often a sectorial chimera [152,154,155]. Several researchers have used mutagen treatment of in vitro cultured shoots for mutation induction, followed by several subcultures to remove the chimeras. Khawale et al. [156] used two nodal microcuttings of V. vinifera ‘Pusa Seedless’ for mutation induction in ethyl methane sulfonate (EMS) and ethidium bromide (EB) supplemented media (ten concentrations from 0.01 – 0.1%) in culture medium. Based on in vitro survival of microcuttings and their subsequent in vitro growth response, the LD50 value for EMS was recorded as 0.04% and for EB it was 0.06%. Randomly Amplified Polymorphic DNA (RAPD) markers were used to detect mutant plants after three subcultures. Seven out of 30 RAPD primers used showed polymorphisms in the mutant population [156]. Munir et al. [157] also used RAPD analysis to identify mutants after irradiation of cultures from three cultivars with gamma-rays and report high yield of mutants, based on polymorphisms for some of the RAPD primers used.
Use of SE is the solution to chimerism in mutation breeding as somatic embryos arise from single cells. The existence of growth centres comprising 5 – 50 cells in embryogenic callus of grapevine was first reported by Krul and Worley [158]. Subsequent anatomical observations using scanning electron microscopy also failed to confirm single cell origin of grapevine somatic embryos due to the technical difficulties involved in observing the sequential development of single living embryogenic cells [141,142]. Gambino et al. [145] observed the differentiation and development of somatic embryos from fast growing callus. However, Faure et al. [159] were the first to report the single cell origin of grapevine somatic embryos and this has been demonstrated in other species as well [160,161,162]. Thus, SE based regeneration approach has great potential for isolating ‘solid’ mutants in vegetatively propagated species [153,163,164,165,166], however only few researchers have used the embryogenic cultures of grapevine for mutation induction. Low rate of somatic embryo induction in many cultivars could be the reason [44,103]. Kuksova et al. [167] tested the effect of five doses of gamma rays (5 – 500 Gy) and exposure to 0.025 mM colchicine over 3 days on embryogenic cultures of V. vinifera ‘Podarok Magaracha’, They observed polyploidisation with gamma rays (in 5 – 100 Gy treatments) but not with colchicine. Except for polyploids, only chlorophyll mutants were reported. The authors emphasised the value of the use of embryogenic cultures in mutagenesis as none of the polyploids displayed chimerism for the chromosome numbers [167]. Yang et al. [168] used colchicine treatment on globular stage somatic embryos derived from immature zygotic embryos of diploid V. vinifera ‘Sinsaut’. They were able to produce tetraploids when the cultures were treated with 20 mg L-1 colchicine for 1 – 3 days, with 1 day treatment producing the highest frequency of 4 % tetraploids among regenerated somatic embryos. They also reported uniformity of tetraploidy in the individual plants in repeated tests, confirming that SE-based mutagenesis can produce chimera-free mutants [168]. Polyploidization in grapevine may allow a greater fruit size and a delay in ripening time. Capriotti et al. [169] treated 2 mm slices of embryogenic masses of V. vinifera ‘Chardonnay’, ‘Melot’ and ‘Pinot Grigio’ with 0.05, 0.25 and 0.5 % EMS solution for 3 h and with 0.03 and 0.04 % sodium azide for 4 h and regenerated over 1400 plants which were screened for a natural infection of powdery mildew (Erisiphe necator). They identified 5 Pinot Grigio, 81 Merlot and 59 Chardonnay putative mutants showing low infection [169]. Pathirana and Carimi [43] optimised the EMS treatment of V. vinifera ‘Chardonnay’, ‘Sauvignon blanc’ and ‘Riesling’ for mutation induction and reported that treating somatic embryos with 0.1% EMS solution for one hour results in 50% survival, which they considered optimal for mutation induction experiments.
In Figure 3 we illustrate a scheme for mutation induction and selection using embryogenic cultures of grapevine, also applicable to any other crop. We suggest optimising treatment with mutagens using growth reduction curves as demonstrated in Figure 3 and in Pathirana and Carimi [43]. It is recommended that the mutant dose resulting in 50% growth reduction be used for inducing mutations in large populations of embryogenic cultures [152,155]. The regenerated embryos after mutagen treatment can be already challenged in vitro for many agronomic traits such as toxic chemicals, salinity, pH, drought, viruses etc. [13,152,155,163], or they can be tested under greenhouse or field conditions (Figure 3).
Another approach for grapevine improvement would be the generation and screening of mutant populations developed through transposon activation in the embryogenic tissues by exposure to stress treatments. Movement and insertion of transposons is an important source of variation and evolution of the plant kingdom [170]. Colour variation in maize kernels due to transposon insertion [171,172] is a classic example. In grapevine, red berry variants often encountered as mutants in white berry cultivars have been shown to be the result of recombination between long terminal repeats of the Gret1 retrotransposon, present in a homozygous state at the promoter of VvMybA1 in white grapevine cultivars [173,174]. Furthermore, new generation sequencing of phenotypically different ‘Pinot noir’ clones has revealed that insertion polymorphism generated by mobile elements displayed the highest number of mutational events with respect to clonal variation [149]. The publication of the complete grapevine genome has provided further evidence that mobile elements, in particular Class II elements, have contributed to the genomic variability of V. vinifera [175] and of the repetitive sequences representing 66.47% of the genome, largest portion comprised transposable elements (63.90%) [176]. With several research groups reporting induction of grapevine secondary somatic embryos [15,18,56,177] and their cryopreservation [34,35,36,106,107,109,110], it is now possible to use RNAseq tagging and recover mutant embryos with the same mutation. This proposed scheme is presented in Figure 4.
5.4. Genetic Engineering
Somatic embryogenesis is a valuable biotechnological tool that allows for the genetic manipulation of clonally propagated species, such as grapevines [43,151]. Various grapevine varieties have been successfully genetically transformed using embryogenic cultures to produce highly regenerative target material [178]. The grapevine genetic background influences somatic embryogenesis, which is the most commonly used regeneration method in genetic engineering protocols for this crop [44,179]. Somatic embryogenesis is the primary regenerative process utilized in grapevines for genetic transformation, providing a dependable method for clonal propagation, genetic enhancement and in functional genomic research following transformation [180,181].
Genetic engineering in grapevine has the potential to improve various traits relating to grapevine cultivation. It can lead to the development of stress-tolerant and disease-resistant varieties with increased productivity, efficiency, sustainability, and environmental friendliness [182]. However, the successful commercialization of genetically improved grapevine varieties faces several challenges. These include scientific, legal, and regulatory issues, intellectual property and patenting concerns, political and economic factors, and negative public perception of genetically modified products [182]. Overcoming these hurdles is crucial for the implementation and widespread adoption of genetically improved grapevine varieties.
Research has also been conducted on the characterization of tumorigenic strains of Agrobacterium spp. isolated from grapevine tumors [183]. These strains, including A. vitis, A. tumefaciens, and A. rhizogenes, were found to be tumorigenic on grapevines and exhibited different pathogenicity on other hosts [183]. The study also identified chromosomal and Ti plasmid genes that can be targeted for PCR amplifications to detect these Agrobacterium species in grapevine [183]. Biological control of crown gall, a disease caused by A. tumefaciens, has been investigated in grapevine using nonpathogenic strains of Rhizobium vitis [184]. These nonpathogenic strains, such as ARK-1, have been shown to reduce the incidence of crown gall in grapevine plants [184]. The use of biological control agents like ARK-1 can provide an alternative to chemical treatments for managing crown gall disease in grapevine. Anthocyanin acyltransferases play a crucial role in the production of acylated anthocyanins in grape skins [185]. The regulation of these enzymes by transcription factors like VvMYBA can influence the composition of anthocyanins in grapes [185]. Understanding the biosynthesis and regulation of these compounds is important for determining the aroma profiles and quality of grapes and wines.
Genetic engineering techniques, such as ZFN, TALEN, and CRISPR/Cas, have been employed in V. vinifera research to improve traits such as disease resistance and sugar accumulation [186,187,188,189,190]. These techniques offer targeted genome editing capabilities, allowing for precise modifications to the grapevine genome. The CRISPR/Cas9 system has been used in grapevine to edit specific genes of interest. Wang et al. [190] performed whole-genome sequencing of Cas9-edited grapevine plants and identified rare off-target mutations. Wang et al. [190] and Ren et al. [189] optimized the CRISPR/Cas9 system in grapevine by using grape promoters, which significantly increased the editing efficiency. Wan et al[191] reported the use of CRISPR/Cas9-mediated mutagenesis to enhance resistance to powdery mildew in grapevine.
In addition to CRISPR/Cas9, other genetic engineering techniques have been explored in grapevine. For example, Vidal et al. [192] demonstrated high-efficiency biolistic co-transformation and regeneration of grapevine plants containing antimicrobial peptide genes. When regenerated and acclimated plants were challenged in the greenhouse with either A. vitis strains (bacterial crown gall pathogen) or Uncinula necator (powdery mildew pathogen) for evaluation of disease resistance, a total of 6 mag2 (natural magainin-2) and 5 MSI99 (a synthetic derivative) lines expressing the antimicrobial genes exhibited significant reductions of crown gall symptoms as compared to non-transformed controls. However, only two mag2 lines showed measurable symptom reductions in response to U. necator, but not strong resistance. Their results suggest that the expression of magainin-type genes in grapevines may be more effective against bacteria than fungi [61].
Dhekney et al. [193] used cisgenic engineering to develop grapevines with improved fungal disease resistance by isolating and modifying the V. vinifera thaumatin-like protein gene. Bosco et al. [194] investigated the correlation between the expression of disease resistance in genetically modified grapevines and the contents of viral sequences in the T-DNA and global genome methylation. These genetic engineering techniques offer potential solutions for improving disease resistance in grapevine. Traditional breeding methods have limitations in identifying Vitis species with virus resistance, making genetic engineering an attractive alternative [187,195]. By targeting specific susceptibility genes, genome editing technologies like CRISPR/Cas9 can decrease susceptibility to fungal and oomycete diseases in grapevine [186]. Furthermore, the introduction of genes with antimicrobial activity from other plants or microorganisms has been used to enhance resistance to fungal and bacterial diseases in grapevine [194]. He et al. [196] isolated a gene encoding a pathogenesis-related thaumatin-like protein from a clone of downy mildew-resistant V. amurensis and transformed it into SE calli of V. vinifera 'Thompson Seedless' via Agrobacterium tumefaciens. The transgenic grapevines exhibited improved resistance against downy mildew, with significant inhibition of hyphae growth and asexual reproduction of the pathogen [196].
Transgenic grapevines have been the subject of several studies, exploring different aspects of grapevine biology, biochemistry and genetics. One study by Rinaldo et al. [185] investigated the role of a grapevine anthocyanin acyltransferase gene, VvMYBA, in the production of acylated anthocyanins in grape skins. They ectopically expressed the VlMYBA1 gene from V. labruscana in grapevine hairy root tissue and analyzed gene expression changes in the transcriptome of these roots. They found that VlMYBA1 regulated a narrow set of genes involved in anthocyanin biosynthesis and identified novel genes associated with anthocyanin transport [185]. In another study, Zou et al. [197] focused on the development of transferable DNA markers for grapevine breeding and genetics. They developed a marker strategy targeting the Vitis collinear core genome and developed 2,000 rhAmpSeq markers. They validated the marker panel in four biparental populations spanning the diversity of the Vitis genus, showing a transferability rate of 91.9% [197]. This marker development strategy has the potential to improve marker transferability in grapevine breeding.
Furthermore, the influence of transcription factors, on grapevine biology and disease resistance has been investigated using grapevine transformed through SE pathway. Guillaumie et al. [198] studied the role of the grapevine transcription factor VvWRKY2 in cell wall structure and lignin biosynthesis. Transgenic tobacco plants overexpressing VvWRKY2 exhibited alterations in lignin composition and expression of genes involved in lignin biosynthesis and cell wall formation [198].
In conclusion, research on transgenic grapevines involving Agrobacterium transformation, ZFN, TALEN, and CRISPR/Cas has explored various aspects of grapevine biology, including anthocyanin biosynthesis, sugar accumulation, disease resistance, and genetic factors influencing cell wall structure and lignin biosynthesis. Optimization of the CRISPR/Cas9 system using grape promoters has been shown to increase editing efficiency. Other genetic engineering approaches, such as biolistic co-transformation and cisgenic engineering, have also been explored in grapevine. These techniques offer targeted genome editing capabilities and have been used to edit specific genes of interest in grapevine providing valuable insights into the potential applications of genetic engineering and marker-assisted breeding in grapevine improvement. These solutions achieved for trait improvement using genetic engineering techniques in grapevine, would be challenging to achieve through traditional breeding methods.
6. Genetic Stability of Plants Regenerated from Somatic Embryos
SE is a method that offers possibility for clonal plant regeneration. However, in some cases, plantlets regenerated in vitro culture might develop altered characteristics and reveal a wide array of genetic variants. This variation can arise due to two phenomena: Somaclonal variation and separation of chimeric layers.
6.1. Somaclonal Variation
Some of the changes caused by somaclonal variation can be stable and therefore can be transmitted in seeds or grafted plants. Therefore, somaclonal variations may constitute a new source of genetic variability, a crucial element in any breeding program aimed at enhancing the agronomic traits of plants [44]. The most dominant hypothesis is that the genetic instability is caused by stress to which the explant cells and the new cells generated in vitro are subjected. SE is usually achieved in vitro by exposing plant explants to PGR treatments which induces the typical formation of a callus; an apparently disorganized mass of cells. These passages leading to callus formation often expose plant cells to stress and ultimately lead to unwanted alterations in the genome and epigenome [2].
Among the effects caused by in vitro culture stress are the occurrence of anomalous cytological events during callus formation, and the prevalence of polyploidization and chromosome reduction events [199]. The presence of 2,4-dichlorophenoxyacetic acid (2,4-D), one of the most used growth regulators in plant tissue culture, is considered the main agent responsible for this and other chromosomal abnormalities. Other factors, such as temperature variation or physical and chemical stresses, may contribute to chromosomal instability [200]. It has been observed that in explants of different plant species cultured in vitro, the initial events leading to SE appear to have undergone reprogramming of somatic cells to a gamete-like state, including chromosome segregation and the emergence of haploid gamete-like cell appearance [199,201,202].
Although the production of haploids was initially considered a negative effect induced by the physiological and morphological disorders to which cells grown in vitro are subjected, today it constitutes an opportunity for innovative breeding strategies aimed at promoting and improving sustainable agriculture. The practical values of haploids in plant breeding have been illustrated by several authors and therefore the in vitro switch from mitotic cell division to meiosis has aroused growing interest. Murray et al. [203] and De La Fuente et al. [204] introduced the concept of a cell-based in vitro breeding system (termed In Vitro Nurseries; IVNs). In IVNs, breeding cycle time could be substantially reduced by enabling rapid cell-level breeding cycles, without the need for flowering. The explants collected from mother plants could be cultured and brought to induce haploid cells after recombination without gametophyte development (artificial gametes), these cells can then be fused artificially in vitro [205]. This opportunity is of great interest especially for crops like grapevine that have a long juvenile phase. In addition, IVNs will significantly reduce field management costs and environmental risks related to biotic and abiotic stresses. However, to apply IVNs more widely requires overcoming several bottlenecks. Cook et al. [205] distinguish three distinct phases: i) in vitro production of haploid gamete-like cell inducing meiosis from somatic vegetative tissues; ii) identifying/isolating artificial gametes carrying favourable alleles; iii) producing cell lines from selected artificial gametes followed by the fusion of selected artificial gametes to generate diploid cells, as a starting point for the next generation in IVNs. Therefore, it is useful to develop efficient protocols to induce meiosis in vitro and regenerate haploid cell lines. Several substances added to culture media reduce chromosome number in cells maintained in vitro. Among these, chloramphenicol antibiotic treatment was shown to reduce chromosomes to a haploid state in root cells of barley seedlings [206]. Caffeine treatments have been used to induce somatic meiosis-like reductions in Vicia root tips [207]. The exogenous application of trichostatin A has also been used to induce the formation of haploid somatic embryos from male gametes of different species [208,209,210].
Somaclonal variation in grapevine can be phenotypically evaluated by observing morphological and physiological traits in ex vitro and in vivo grown plants, using the international standard descriptors (ampelographic and ampelometric) provided by the International Organization of Vine and Wine (OIV) [211]. However, the evaluation of the uniformity of morphological traits in field-grown plants is expensive and it is necessary to wait several years in order to overcome the juvenile phase and thus be able to evaluate the characteristic of the fruit. Moreover, some changes obtained after in vitro culture cannot be observed in planta, because differences that influence the biological activity may not affect the phenotype.
To complement the morphological characterization of regenerants in the field using the descriptors from the OIV, cytogenetic, biochemical, as well as DNA- and RNA-based technologies are sensitive tools, which can quickly provide information on genetic stability [13]. Cytogenetic studies on grapevine are often difficult, mainly due to the large number of small chromosomes and the difficulty of obtaining good chromosome preparations [212]. One of the most efficient techniques to detect different ploidy levels is based on flow cytometry. Different ploidy levels in grapevine regenerants via SE were detected by flow cytometry. Autotetraploid plants showed marked anatomical and morphological changes in shoots and mature leaves. Alterations have also been observed in stomata and chloroplast number which was higher in the tetraploids than in the diploid mother plants. On the contrary, the stomatal index was markedly decreased in leaves of tetraploid regenerants [13]. Capriotti et al. [169] tested over 2300 Ancellotta and Lambrusco Salamino plants regenerated through somatic embryogenesis for somaclonal variants for downy mildew (Plasmopara viticola) resistance after inoculation with a spore suspension. They identified 54 plants of Lambrusco Salamino and 22 plants of Ancellotta showing low level of infection.
Different molecular markers have been used for the assessment of genetic fidelity of regenerants, most of which are based on PCR technology. The most used DNA markers for the verification of grapevine genetic fidelity are: random amplified polymorphic DNA (RAPD), sequence characterized amplified regions (SCARs), simple sequence repeats (SSRs), inter-simple sequence repeats (ISSRs), amplified fragment length polymorphisms (AFLPs), single nucleotide polymorphisms (SNPs), expressed sequence tags (ESTs), and random amplified microsatellite polymorphisms (RAMPs) [15,124,213,214,215,216,217]. For an exhaustive and updated review of the molecular markers used for the assessment of genetic fidelity of in vitro regenerated plants, see Biswas and Kumar [218] and for grapevine, see Butiuc-Keul and Coste [212].
6.2. Chimerism in Grapevine and Segregation of Genotypes Through Somatic Embryogenesis
Chimerism refers to the presence of genetically distinct cell layers within a single plant. This phenomenon has been observed in grapevines and has implications for cultivar identity, ancestry, and genetic improvement [120]. Chimerism can arise through somatic mutations that occur in one of the three meristematic cell layers in the apical meristem, which then differentiate into various plant tissues [219]. The existence of chimerism in grapevines has been demonstrated through DNA profiling using microsatellite loci [120]. In some cases, more than two alleles have been observed at a locus, indicating the presence of chimerism [120].
Chimerism can manifest in various forms. Somatic chimerism occurs when different genetic lineages exist within different tissues of a grapevine plant. For example, a grapevine may have shoots or leaves with genetic characteristics true to the clone while its berries or flowers display characteristics different from the clone. This variation can arise due to genetic mutations or somatic hybridization events. Periclinal chimerism refers to the presence of different genetic lineages in distinct layers of tissue within a grapevine. It occurs when genetic mutations affect specific cell layers during plant development. This type of chimerism can result in variegated patterns of color or leaf morphology, where different tissue layers display different traits.
Chimerism in grapevines can have significant effects on phenotype and genetic diversity. Somatic mutations that give rise to chimeras can lead to morphological and agronomical differences, which can stabilize in grapevine plants and contribute to the genetic diversity of grapevine accessions [120,220]. This suggests that chimerism can modify phenotype and influence grapevine improvement through genetic transformation and conventional breeding strategies [120]. Chimerism significantly impacts grapevine clone stability by introducing genetic variability within a single plant. Periclinal chimeras, which consist of distinct cell layers (L1 and L2), can exhibit different phenotypes and genetic profiles, influencing both agronomic traits and cultivar identity. For instance, the study of the ‘Merlot’ cultivar revealed specific periclinal chimeras that could be propagated, suggesting potential for improved clonal selection [221]. However, somatic embryogenesis, which typically regenerates plants from the L1 layer, may limit the expression of phenotypic diversity, as seen in the ‘Nebbiolo’ [121] and ‘Chardonnay 96’ [222] clones, where regenerated plants showed no significant phenotypic alterations compared to their parent plants. This indicates that while chimerism can enhance genetic diversity, the method of propagation can influence the stability and expression of these variations, highlighting a complex interplay between genetic chimerism and clonal propagation strategies [223].
Understanding chimerism is important for genetic diversity assessment, population structure analysis, and the development of new grapevine cultivars. Molecular markers, such as microsatellite loci and SNPs, can be used to investigate patterns of genetic diversity in grapevine germplasm collections [224]. These markers can provide reliable tools for characterizing the genetic diversity and population structure of grapevine accessions, including domesticated grapevine (Vitis vinifera ssp. sativa), wild relative (Vitis vinifera ssp. sylvestris), interspecific hybrid cultivars, and rootstocks [224].
Chimerism in grapevines can have both positive and negative implications. As positive implications chimerism can introduce genetic diversity within a grapevine plant, potentially leading to the development of unique traits or characteristics, and chimeras displaying desirable traits can be identified and propagated through clonal selection, allowing for the production of new grape clones with specific traits or improved quality. As negative implications, chimerism can lead to unpredictability in grapevine propagation, as the desired traits might not be stably inherited in subsequent generations. This can complicate breeding programs and commercial cultivation, and chimeric grapevines may produce fruits with varying characteristics, making it challenging to achieve uniformity in terms of flavour, colour, or size.
7. Conclusions and Future Prospects
SE is a transformative biotechnological approach for grapevine improvement, germplasm conservation, and disease management. The ability of SE to regenerate whole plants from somatic cells offers substantial benefits, particularly in generating genetically uniform plants, preserving valuable grapevine germplasm, and enabling genetic transformation. Despite these benefits, SE's practical application is constrained by challenges such as genotype recalcitrance, somaclonal variation, and difficulties in synchronizing embryo production. Addressing these challenges will be essential to fully realize SE's potential in both research and commercial contexts. Nevertheless, SE remains an indispensable tool for grapevine research, providing solutions where traditional breeding methods fall short due to the grapevine’s long reproductive cycles and complex genetic makeup.
Future research efforts must focus on optimizing SE protocols to improve reproducibility across a broader range of grapevine genotypes. This involves refining the selection of explants, adjusting culture media compositions, identifying optimal concentrations of PGR and culture conditions. A deeper understanding of the mechanisms underlying somaclonal variation is critical to mitigating the genetic instability frequently observed in in vitro cultures. Furthermore, improving the synchronization of somatic embryo production is crucial for scaling up SE systems for commercial use. Further advances in the understanding of signaling pathways and molecular mechanisms that regulate embryogenesis could lead to significant improvements in this area.
The integration of SE with modern biotechnologies, such as genome editing tools (e.g., CRISPR/Cas9), RNA sequencing, and metabolomics, presents an exciting opportunity to enhance grapevine breeding programs. SE provides a foundational platform for precise genetic modifications, enabling the creation of grapevine cultivars with improved traits such as disease resistance, stress tolerance, and enhanced yield and quality.
Cryopreservation techniques also require further refinement to ensure high regeneration rates of conserved somatic embryos, particularly for the preservation of endangered species and elite cultivars. Additionally, the development of automated SE processes is essential for reducing costs and making SE more commercially viable. Automated systems for embryo culture, synchronization, and plantlet acclimatization will be instrumental in scaling up SE for large-scale applications.
While SE faces several challenges, ongoing research and refinement of SE techniques, coupled with the integration of emerging technologies, hold great promise for expanding its applications in grapevine research and production. By overcoming these current limitations, SE has the potential to significantly enhance the resilience, sustainability, and adaptability of grapevine cultivation in response to the demands of modern agriculture and changing climate.
Author Contributions
Conceptualization, R.P. and F.C.; writing-original draft preparation, A.W., A.C., R.P. and F.C.; Writing-review and editing, R.P. and F.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Su, Y.H.; Tang, L.P.; Zhao, X.Y.; Zhang, X.S. Plant cell totipotency: Insights into cellular reprogramming. Journal of Integrative Plant Biology 2021, 63, 228–243. [Google Scholar] [CrossRef]
- Salaün, C.; Lepiniec, L.; Dubreucq, B. Genetic and molecular control of somatic embryogenesis. Plants 2021, 10, 1467. [Google Scholar] [CrossRef] [PubMed]
- Haberlandt, G. Culturversuche mit isolierten Pflanzenzellen. In Plant Tissue Culture: 100 Years Since Gottlieb Haberlandt; Springer: 2003; pp. 1–24.
- Gautheret, R. Plant Tissue Culture: 100 Years since Gottlieb Haberlandt. New York: SpringWien 2003, 205-214.
- Haberlandt, G. Zelle und Elementarorgan. BioI. Zentralbl. 1925, 45, 257–272. [Google Scholar]
- Höxtermann, E. Cellular ‘elementary organisms’ in vitro. The early vision of Gottlieb Haberlandt and its realization. Physiologia Plantarum 1997, 100, 716–728. [Google Scholar] [CrossRef]
- Steward, F.; Mapes, M.O.; Mears, K. Growth and organized development of cultured cells. II. Organization in cultures grown from freely suspended cells. American Journal of Botany 1958, 705–708. [Google Scholar] [CrossRef]
- Sugiyama, M. Historical review of research on plant cell dedifferentiation. Journal of Plant Research 2015, 128, 349–359. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef]
- Horstman, A.; Bemer, M.; Boutilier, K. A transcriptional view on somatic embryogenesis. Regeneration 2017, 4, 201–216. [Google Scholar] [CrossRef]
- Méndez-Hernández, H.A.; Ledezma-Rodríguez, M.; Avilez-Montalvo, R.N.; Juárez-Gómez, Y.L.; Skeete, A.; Avilez-Montalvo, J.; De-la-Peña, C.; Loyola-Vargas, V.M. Signaling overview of plant somatic embryogenesis. Frontiers in Plant Science 2019, 10, 77. [Google Scholar] [CrossRef]
- Loyola-Vargas, V.M.; Ochoa-Alejo, N. Somatic embryogenesis. An overview. In Somatic Embryogenesis. Fundamental Aspects and Applications, Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Springer: Berlin, 2016; pp. 1–10. [Google Scholar]
- Catalano, C.; Abbate, L.; Motisi, A.; Crucitti, D.; Cangelosi, V.; Pisciotta, A.; Di Lorenzo, R.; Carimi, F.; Carra, A. Autotetraploid emergence via somatic embryogenesis in Vitis vinifera induces marked morphological changes in shoots, mature leaves, and stomata. Cells 2021, 10. [Google Scholar] [CrossRef]
- Zhang, Y.; Iaffaldano, B.; Qi, Y. CRISPR ribonucleoprotein-mediated genetic engineering in plants. Plant communications 2021, 2. [Google Scholar] [CrossRef]
- Carra, A.; Sajeva, M.; Abbate, L.; Siragusa, M.; Pathirana, R.; Carimi, F. Factors affecting somatic embryogenesis in eight Italian grapevine cultivars and the genetic stability of embryo-derived regenerants as assessed by molecular markers. Scientia Horticulturae 2016, 204, 123–127. [Google Scholar] [CrossRef]
- Fiore, S.; De Pasquale, F.; Carimi, F.; Sajeva, M. Effect of 2, 4-D and 4-CPPU on somatic embryogenesis from stigma and style transverse thin cell layers of Citrus. Plant Cell, Tissue and Organ Culture 2002, 68, 57–63. [Google Scholar] [CrossRef]
- Bonga, J.M.; Klimaszewska, K.K.; von Aderkas, P. Recalcitrance in clonal propagation, in particular of conifers. Plant Cell Tiss Organ Cult 2009, 100, 241–254. [Google Scholar] [CrossRef]
- Carimi, F.; Barizza, E.; Gardiman, M.; Lo Schiavo, F. Somatic embryogenesis from stigmas and styles of grapevine. In Vitro Cellular & Developmental Biology-Plant 2005, 41, 249–252. [Google Scholar]
- Carimi, F.; De Pasquale, F. Micropropagation of Citrus. In Micropropagation of Woody Trees and Fruits; Forestry Sciences; 2003; pp. 589–619.
- Martinelli, L.; Gribaudo, I. Somatic embryogenesis in grapevine. In Molecular Biology and Biotechnology of the Grapevine, Roubelakis-Angelakis, K.A., Ed.; Kluwer Academic Publishers: Dordrecht-Boston-London, 2001; pp. 327–351. [Google Scholar]
- Zimmerman, J.L. Somatic embryogenesis: a model for early development in higher plants. The Plant Cell 1993, 5, 1411–1423. [Google Scholar] [CrossRef]
- Gupta, S.D.; Conger, B. Somatic embryogenesis and plant regeneration from suspension cultures of switchgrass. Crop science 1999, 39, 243–247. [Google Scholar] [CrossRef]
- Chapman, A.; Blervacq, A.-S.; Hendriks, T.; Slomianny, C.; Vasseur, J.; Hilbert, J.-L. Cell wall differentiation during early somatic embryogenesis in plants. II. Ultrastructural study and pectin immunolocalization on chicory embryos. Canadian Journal of Botany 2000, 78, 824–831. [Google Scholar]
- Yeung, E.C. Structural and developmental patterns in somatic embryogenesis. In Vitro Embryogenesis in Plants, Thorpe, T.A., Ed.; Kluwer Academic Publishers: Dordrecht, Boston, London, 1995; pp. 205–247. [Google Scholar]
- Thomas, T.L. Gene expression during plant embryogenesis and germination: an overview. Plant Cell 1993, 5, 1401–1410. [Google Scholar] [CrossRef]
- Joshi, R.; Kumar, P. Regulation of somatic embryogenesis in crops: a review. Agricultural Reviews 2013, 34, 1–20. [Google Scholar]
- Ammirato, P.V. Organizational events during somatic embryogenesis. In Proceedings of the VIth International Congress on Plant Tissue and Cell Culture, University of Minnesota, St. Paul, USA; 1986. [Google Scholar]
- Faure, O.; Aarrouf, J.; Nougarede, A. Ontogenesis, differentiation and precocious germination in anther-derived somatic embryos of grapevine (vitis vinifera L.): Embryonic organogenesis. Annals of Botany 1996, 78, 29–37. [Google Scholar] [CrossRef]
- Jayasankar, S.; Bondada, B.R.; Li, Z.J.; Gray, D.J. Comparative anatomy and morphology of Vitis vinifera (Vitaceae) somatic embryos from solid- and liquid-culture-derived proembryogenic masses. American Journal of Botany 2003, 90, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Jayasankar, S.; Gray, D.J.; Litz, R.E. High-efficiency somatic embryogenesis and plant regeneration from suspension cultures of grapevine. Plant Cell Reports 1999, 18, 533–537. [Google Scholar] [CrossRef]
- Coutos-Thevenot, P.; Goebel-Tourand, I.; Mauro, M.-C.; Jouanneau, J.-P.; Boulay, M.; Deloire, A.; Guern, J. Somatic embryogenesis from grapevine cells. I-Improvement of embryo development by changes in culture conditions. Plant Cell, Tissue and Organ Culture 1992, 29, 125–133. [Google Scholar] [CrossRef]
- Coutos-Thevenot, P.; Maes, O.; Jouenne, T.; Claude Mauro, M.; Boulay, M.; Deloire, A.; Guern, J. Extracellular protein patterns of grapevine cell suspensions in embryogenic and non-embryogenic situations. Plant Science 1992, 86, 137–145. [Google Scholar] [CrossRef]
- Zlenko, V.A.; Kotikov, I.V.; Troshin, L.P. Plant regeneration from somatic embryos of interspecific hybrids of grapevine formed in liquid medium. Journal of Horticultural Science & Biotechnology 2005, 80, 461–465. [Google Scholar]
- Vasanth, K.; Vivier, M.A. Improved cryopreservation procedure for long term storage of synchronised culture of grapevine. Biologia Plantarum 2011, 55, 365–369. [Google Scholar] [CrossRef]
- Wang, Q.C.; Gafny, R.; Sahar, N.; Sela, I.; Mawassi, M.; Tanne, E.; Perl, A. Cryopreservation of grapevine (Vitis vinifera L.) embryogenic cell suspensions by encapsulation-dehydration and subsequent plant regeneration. Plant Science 2002, 162, 551–558. [Google Scholar] [CrossRef]
- Wang, Q.C.; Mawassi, M.; Sahar, N.; Li, P.; Violeta, C.T.; Gafny, R.; Sela, I.; Tanne, E.; Perl, A. Cryopreservation of grapevine (Vitis spp.) embryogenic cell suspensions by encapsulation-vitrification. Plant Cell Tissue and Organ Culture 2004, 77, 267–275. [Google Scholar] [CrossRef]
- Garcia, C.; Furtado de Almeida, A.-A.; Costa, M.; Britto, D.; Valle, R.; Royaert, S.; Marelli, J.-P. Abnormalities in somatic embryogenesis caused by 2, 4-D: an overview. Plant Cell Tiss Organ Cult 2019, 137, 193–212. [Google Scholar] [CrossRef]
- Acanda, Y.; Martínez, Ó.; Prado, M.J.; González, M.V.; Rey, M. Changes in abscisic acid metabolism in relation to the maturation of grapevine (Vitis vinifera L., cv. Mencía) somatic embryos. BMC Plant Biology 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Martinelli, L.; Gribaudo, I. Strategies for effective somatic embryogenesis in grapevine: an appraisal. In Grapevine Molecular Physiology & Biotechnology, Roubelakis-Angelakis, K.A., Ed.; Springer Netherlands: Dordrecht, 2009; pp. 461–493. [Google Scholar]
- Bidabadi, S.S.; Jain, S.M. Cellular, Molecular, and Physiological Aspects of In Vitro Plant Regeneration. Plants (Basel) 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Prado, M.J.; Grueiro, M.P.; González, M.V.; Testillano, P.S.; Domínguez, C.; López, M.; Rey, M. Efficient plant regeneration through somatic embryogenesis from anthers and ovaries of six autochthonous grapevine cultivars from Galicia (Spain). Scientia Horticulturae 2010, 125, 342–352. [Google Scholar] [CrossRef]
- Gambino, G.; Minuto, M.; Boccacci, P.; Perrone, I.; Vallania, R.; Gribaudo, I. Characterization of expression dynamics of WOX homeodomain transcription factors during somatic embryogenesis in Vitis vinifera. Journal of Experimental Botany 2011, 62, 1089–1101. [Google Scholar] [CrossRef]
- Pathirana, R.; Carimi, F. Studies on improving the efficiency of somatic embryogenesis in grapevine (Vitis vinifera L.) and optimising ethyl methanesulfonate treatment for mutation induction. Plants 2023, 12, 4126. [Google Scholar] [CrossRef] [PubMed]
- Carimi, F.; Pathirana, R.; Carra, A. Somatic embryogenesis and Agrobacterium mediated genetic transformation in Vitis In Somatic Embryogenesis and Genetic Transformation in Plants Aslam, J., Srivastave, P.S., Sharma, M.P., Eds.; Narosa Publishing House: New Delhi, 2013; pp. 199–218. [Google Scholar]
- Gribaudo, I.; Gambino, G.; Vallania, R. Somatic embryogenesis from grapevine anthers: The optimal developmental stage for collecting explants. American Journal of Enology and Viticulture 2004, 55, 427–430. [Google Scholar] [CrossRef]
- Oláh, R.; Zok, A.; Pedryc, A.; Howard, S.; Kovács, L.G. Somatic embryogenesis in a broad spectrum of grape genotypes. Scientia Horticulturae 2009, 120, 134–137. [Google Scholar] [CrossRef]
- Capriotti, L.; Limera, C.; Mezzetti, B.; Ricci, A.; Sabbadini, S. From induction to embryo proliferation: improved somatic embryogenesis protocol in grapevine for Italian cultivars and hybrid Vitis rootstocks. Plant Cell Tiss Organ Cult 2022, 151, 221–233. [Google Scholar] [CrossRef]
- Schellenbaum, P.; Jacques, A.; Maillot, P.; Bertsch, C.; Mazet, F.; Farine, S.; Walter, B. Characterization of VvSERK1, VvSERK2, VvSERK3 and VvL1L genes and their expression during somatic embryogenesis of grapevine (Vitis vinifera L.). Plant cell reports 2008, 27, 1799–1809. [Google Scholar] [CrossRef]
- Maillot, P.; Lebel, S.; Schellenbaum, P.; Jacques, A.; Walter, B. Differential regulation of SERK, LEC1-Like and Pathogenesis-Related genes during indirect secondary somatic embryogenesis in grapevine. Plant Physiology and Biochemistry 2009, 47, 743–752. [Google Scholar] [CrossRef]
- Coutos-thevenot, P.; Jouenne, T.; Maes, O.; Guerbette, F.; Grosbois, M.; Le Caer, J.P.; Boulay, M.; Deloire, A.; Kader, J.C.; Guern, J. Four 9-kDa proteins excreted by somatic embryos of grapevine are isoforms of lipid-transfer proteins. European Journal of Biochemistry 1993, 217, 885–889. [Google Scholar] [CrossRef]
- Francois, J.; Lallemand, M.; Fleurat-Lessard, P.; Laquitaine, L.; Delrot, S.; Coutos-Thevenot, P.; Gomes, E. Overexpression of the VvLTP1 gene interferes with somatic embryo development in grapevine. Funct Plant Biol 2008, 35, 394–402. [Google Scholar] [CrossRef]
- Maës, O.; Coutos-Thévenot, P.; Jouenne, T.; Boulay, M.; Guern, J. Influence of extracellular proteins, proteases and protease inhibitors on grapevine somatic embryogenesis. Plant Cell, Tissue and Organ Culture 1997, 50, 97–105. [Google Scholar] [CrossRef]
- Vidal, J.; Rama, J.; Taboada, L.; Martin, C.; Ibanez, M.; Segura, A.; Gonzalez-Benito, M.E. Improved somatic embryogenesis of grapevine (Vitis vinifera) with focus on induction parameters and efficient plant regeneration. Plant Cell Tissue and Organ Culture 2009, 96, 85–94. [Google Scholar] [CrossRef]
- San Pedro, T.; Gammoudi, N.; Peiró, R.; Olmos, A.; Gisbert, C. Somatic embryogenesis from seeds in a broad range of Vitis vinifera L. varieties: rescue of true-to-type virus-free plants. BMC Plant Biology 2017, 17, 226. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Tripathi, M.; Tiwari, R.; Baghel, B.; Ahuja, A. Somatic embryogenesis and plantlet regeneration via embryogenic suspensions of grape (Vitis vinifera L.). Asian J. Microbiol. Biotechnol. Environ. Sci 2018, 20, S112–S125. [Google Scholar]
- Maillot, P.; Deglène-Benbrahim, L.; Walter, B. Efficient somatic embryogenesis from meristematic explants in grapevine (Vitis vinifera L.) cv. Chardonnay: an improved protocol. Trees 2016, 30, 1377–1387. [Google Scholar] [CrossRef]
- Maillot, P.; Kieffer, F.; Walter, B. Somatic embryogenesis from stem nodal sections of grapevine. Vitis 2006, 45, 185–189. [Google Scholar]
- Gambino, G.; Ruffa, P.; Vallania, R.; Gribaudo, I. Somatic embryogenesis from whole flowers, anthers and ovaries of grapevine (Vitis spp.). Plant Cell Tissue and Organ Culture 2007, 90, 79–83. [Google Scholar] [CrossRef]
- Cardoso, H.G.; Campos, M.C.; Pais, M.S.; Peixe, A. Use of morphometric parameters for tracking ovule and microspore evolution in grapevine (Vitis vinifera L., cv. “Aragonez”) and evaluation of their potential to improve in vitro somatic embryogenesis efficiency from gametophyte tissues. In Vitro Cellular & Developmental Biology - Plant 2010, 46, 499–508. [Google Scholar] [CrossRef]
- Emons, M.C. Somatic embryogenesis: cell biological aspects. Acta Botanica Neerlandica 1994, 43, 1–14. [Google Scholar] [CrossRef]
- Vidal, J.R.; Kikkert, J.R.; Malnoy, M.A.; Wallace, P.G.; Barnard, J.; Reisch, B.I. Evaluation of transgenic 'Chardonnay' (Vitis vinifera) containing magainin genes for resistance to crown gall and powdery mildew. Transgenic Research 2006, 15, 69–82. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Nitsch, J.; Nitsch, C. Haploid plants from pollen grains. Science 1969, 163, 85–87. [Google Scholar] [CrossRef] [PubMed]
- Linsmaier, E.M.; Skoog, F. Organic growth factor requirements of tobacco tissue cultures. Physiologia Plantarum 1965, 18, 100–106. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B.H. Commercially feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Proc. Int. Plant Prop. Soc. 1980, 30, 421–427. [Google Scholar]
- Chee, R.; Pool, R.; Bucher, D. A method for large scale in vitro propagation of Vitis. New York's Food and Life Sciences Bulletin 1984, 1-9.
- Driver, J.A.; Kuniyuki, A.H. In vitro propagation of Paradox walnut rootstock. HortScience 1984, 19, 507–509. [Google Scholar] [CrossRef]
- Perrin, M.; Martin, D.; Joly, D.; Demangeat, G.; This, P.; Masson, J.E. Medium-dependent response of grapevine somatic embryogenic cells. Plant Science 2001, 161, 107–116. [Google Scholar] [CrossRef]
- Elidemir, A.; Uzun, H.; Bayir, A. Effect of different medium and sucrose concentrations on germination of somatic embryos in grape. Acta Horticulturae 2007, 754, 117–121. [Google Scholar] [CrossRef]
- Carvalho, D.C.d.; Silva, A.L.L.d.; Schuck, M.R.; Purcino, M.; Tanno, G.N.; Biasi, L.A. Fox grape cv. Bordô (Vitis labrusca L.) and grapevine cv. Chardonnay (Vitis vinifera L.) cultivated in vitro under different carbohydrates, amino acids and 6-Benzylaminopurine levels. Brazilian Archives of Biology and Technology 2013, 56, 191–201. [Google Scholar] [CrossRef]
- Nookaraju, A.; Agrawal, D. Use of amino acids for a highly efficient somatic embryogenesis in grapevine 'Crimson Seedless'. Vitis 2013, 52, 137–140. [Google Scholar]
- Zhang, X.-M.; Wu, Y.-F.; Zhi, L.; Song, C.-B.; Wang, X.-P. Advancements in plant regeneration and genetic transformation of grapevine (Vitis spp.). Journal of Integrative Agriculture 2021, 20, 1407–1434. [Google Scholar] [CrossRef]
- Bouamama, B.; Salem-Fnayou, A.B.; Jouira, H.B.; Ghorbel, A.; Mliki, A. Influence of the flower stage and culture medium on the induction of somatic embryogenesis from anther culture in Tunisian grapevine cultivars. J. Int. Sci. Vigne Vin 2007, 41, 185–192. [Google Scholar] [CrossRef]
- Carimi, F.; Pathirana, R.; Carra, A. Biotechnologies for germplasm management and improvement. In Grapevines – Varieties, Cultivation and Management, Szabo, P.V., Shojania, J., Eds.; Nova Science Publishers: New York, 2012; pp. 199–249. [Google Scholar]
- Zhang, P.; Yu, Z.-Y.; Cheng, Z.-M.; Zhang, Z.; Tao, J.-M. In vitro explants regeneration of the grape ‘Wink’(Vitis vinifera L.‘Wink’). Journal of Plant Breeding and Crop Science 2011, 3, 276–282. [Google Scholar]
- López-Pérez, A.J.; Carreño, J.; Martínez-Cutillas, A.; Dabuza, M. High embryogenic ability and plant regeneration of table grapevine cultivars (Vitis vinifera L.) induced by activated charcoal. Vitis 2005, 44, 79–85. [Google Scholar]
- Li, H.; Li, F.-l.; Du, J.-c.; Lu, H.; He, Z.-q. Somatic embryogenesis and histological analysis from zygotic embryos in Vitis vinifera L.‘Moldova’. Forestry Studies in China 2008, 10, 253–258. [Google Scholar] [CrossRef]
- Li, Z.T.; Dhekney, S.A.; Dutt, M.; Gray, D.J. An improved protocol for Agrobacterium-mediated transformation of grapevine (Vitis vinifera L.). Plant Cell, Tissue and Organ Culture 2008, 93, 311–321. [Google Scholar] [CrossRef]
- Dhekney, S.A.; Li, Z.T.; Gray, D.J. Factors influencing induction and maintenance of Vitis rotundifolia Michx. embryogenic cultures. Plant Cell Tiss Organ Cult 2011, 105, 175–180. [Google Scholar] [CrossRef]
- Park, H.-J.; Lee, H.-R.; Pyee, J.; Cha, H.-C. Regeneration of grape (Vitis labruscana cv. Kyoho) by shoot-tip culture. Journal of Plant Biology 2001, 44, 185–192. [Google Scholar] [CrossRef]
- Jayasankar, S.; Van Aman, M.; Cordts, J.; Dhekney, S.; Li, Z.T.; Gray, D.J. Low temperature storage of suspension culture-derived grapevine somatic embryos and regeneration of plants. In Vitro Cellular & Developmental Biology-Plant 2005, 41, 752–756. [Google Scholar] [CrossRef]
- PlantGrape. 110 Richter. Available online: https://www.plantgrape.fr/en/varieties/rootstock-varieties/21 (accessed on 10 June 2024).
- Ortiz, J.M.; Martin, J.P.; Borrego, J.; Chavez, J.; Rodriguez, I.; Munoz, G.; Cabello, F. Management and characterization of a Vitis germplasm bank in Spain. Proceedings of the 8th International Conference on Grape Genetics and Breeding, Vols 1 and 2 2003, 619-625.
- Popescu, C.F.; Crespan, M. Combining Microsatellite Markers and Ampelography for Better Management of Romanian Grapevine Germplasm Collections. Notulae Scientia Biologicae 2018, 10, 193–198. [Google Scholar] [CrossRef]
- Kaya, H.B.; Dilli, Y.; Oncu-Oner, T.; Unal, A. Exploring genetic diversity and population structure of a large grapevine (Vitis vinifera L.) germplasm collection in Turkiye. Front Plant Sci 2023, 14, 1121811. [Google Scholar] [CrossRef]
- Maul, E.; Töpfer, R.; Carka, F.; Cornea, V.; Crespan, M.; Dallakyan, M.; de Andrés Domínguez, T.; De Lorenzis, G.; Dejeu, L.; Goryslavets, S. Identification and characterization of grapevine genetic resources maintained in Eastern European Collections. Vitis 2015, 54, 5–12. [Google Scholar]
- Margaryan, K.; Maul, E.; Muradyan, Z.; Hovhannisyan, A.; Devejyan, H.; Melyan, G.; Aroutiounian, R. Armenian national grapevine collection: Conservation, characterization and prospects. In Proceedings of the BIO Web of Conferences; 2019; p. 01002. [Google Scholar]
- Akhalkatsi, M.; Ekhvaia, J.; Mosulishvili, M.; Nakhutsrishvili, G.; Abdaladze, O.; Batsatsashvili, K. Reasons and processes leading to the erosion of crop genetic diversity in mountainous regions of Georgia. Mountain Research and Development 2010, 30, 304–310. [Google Scholar] [CrossRef]
- Silva, R.d.C.; Luis, Z.G.; Scherwinski-Pereira, J.E. Short-term storage in vitro and large-scale propagation of grapevine genotypes. Pesqui. Agropecu. Bras. 2012, 47, 344–350. [Google Scholar] [CrossRef]
- San Pedro, T.; Muñoz, P.; Peiró, R.; Jiménez, C.; Olmos, A.; Gisbert, C. Evaluation of conditions for in vitro storage of commercial and minor grapevine (Vitis vinifera L.) cultivars. The Journal of Horticultural Science and Biotechnology 2018, 93, 19–25. [Google Scholar] [CrossRef]
- Alleweldt, G.; Harstlangenbucher, M. The effect of growth-inhibitors on long-term storage of in vitro cultures of grapevine. Vitis 1987, 26, 57–64. [Google Scholar]
- Alzubi, H.; Yepes, L.M.; Fuchs, M. In vitro storage of micropropagated grapevine rootstocks at low temperature. In Vitro Cellular & Developmental Biology - Plant 2019, 55, 334–341. [Google Scholar] [CrossRef]
- Romadanova, N.V.; Aralbayeva, M.M.; Zemtsova, A.S.; Alexandrova, A.M.; Kazybayeva, S.Z.; Mikhailenko, N.V.; Kushnarenko, S.V.; Bettoni, J.C. In Vitro Collection for the Safe Storage of Grapevine Hybrids and Identification of the Presence of Plasmopara viticola Resistance Genes. Plants (Basel) 2024, 13, 1089. [Google Scholar] [CrossRef]
- Ruseva, R. Possibility for short-term in vitro conservation of grapevine explants by excluding growth regulators from the nutrient medium. Rasteniev"dni Nauki 1998, 35, 888–890. [Google Scholar]
- Ruseva, R. Decreasing the growth rate of in vitro grapevine plants by increasing the level of sucrose in the nutrient medium. Rasteniev"dni Nauki 1998, 35, 891–894. [Google Scholar]
- Ruseva, R. Study of the possibilities for in vitro storage of grapevine (Vitis vinifera L.) explants at low temperatures. Rasteniev"dni Nauki 2001, 38, 374–376. [Google Scholar]
- Tehrim, S.; SAJID, G.M. In vitro establishment, conservation and its implications for grape germplasm biodiversity. Romanian Biotechnological Letters 2011, 16, 6781–6789. [Google Scholar]
- Hassan, N.A.; Gomaa, A.H.; Shahin, M.A.; El Homosany, A.A. In vitro storage and cryopreservation of some grape varieties. Journal of Horticultural Science & Ornamental Plants 2013, 5, 183–193. [Google Scholar]
- Gray, D. Effects of dehydration and exogenous growth regulators on dormancy, quiescence and germination of grape somatic embryos. In Vitro Cellular & Developmental Biology 1989, 25, 1173–1178. [Google Scholar]
- Capriotti, L.; Ricci, A.; Molesini, B.; Mezzetti, B.; Pandolfini, T.; Piunti, I.; Sabbadini, S. Efficient protocol of de novo shoot organogenesis from somatic embryos for grapevine genetic transformation. Frontiers in Plant Science 2023, 14, 1172758. [Google Scholar] [CrossRef]
- Senaratna, T.; McKersie, B.D.; Bowley, S.R. Artificial seeds of alfalfa ( Medicago sativa L.). Induction of desiccation tolerance in somatic embryos. In Vitro Cellular & Developmental Biology 1990, 26, 85–90. [Google Scholar] [CrossRef]
- Faure, O.; Dewitte, W.; Nougarède, A.; Van Onckelen, H. Precociously germinating somatic embryos of Vitis vinifera have lower ABA and IAA levels than their germinating zygotic counterparts. Physiologia Plantarum 1998, 102, 591–595. [Google Scholar] [CrossRef]
- Goebel-Tourand, I.; Mauro, M.-C.; Sossountzov, L.; Miginiac, E.; Deloire, A. Arrest of somatic embryo development in grapevine: histological characterization and the effect of ABA, BAP and zeatin in stimulating plantlet development. Plant Cell, Tissue and Organ Culture 1993, 33, 91–103. [Google Scholar] [CrossRef]
- Wang, M.-R.; Lambardi, M.; Engelmann, F.; Pathirana, R.; Panis, B.; Volk, G.M.; Wang, Q.-C. Advances in cryopreservation of in vitro-derived propagules: Technologies and explant sources. Plant Cell Tiss Organ Cult 2021, 144, 7–20. [Google Scholar] [CrossRef]
- Ochatt, S.; Lambardi, M.; Panis, B.; Pathirana, R.; Revilla, M.A.; Wang, Q.-C. Cryopreservation and In Vitro banking: a cool subject – Preface from the editors. Plant Cell Tiss Organ Cult 2021, 144, 1–5. [Google Scholar] [CrossRef]
- Miaja, M.L.; Gambino, G.; Vallania, R.; Gribaudo, I. Cryopreservation of Vitis vinifera L. somatic embryos by vitrification or encapsulation-dehydration. Acta Horticulturae 2004, 663, 599–603. [Google Scholar] [CrossRef]
- Ben-Amar, A.; Daldoul, S.; Allel, D.; Reustle, G.; Mliki, A. Reliable encapsulation-based cryopreservation protocol for safe storage and recovery of grapevine embryogenic cell cultures. Scientia horticulturae 2013, 157, 32–38. [Google Scholar] [CrossRef]
- Wang, Q.C.; Tanne, E.; Arav, A.; Gafny, R. Cryopreservation of in vitro-grown shoot tips of grapevine by encapsulation-dehydration. Plant Cell Tissue and Organ Culture 2000, 63, 41–46. [Google Scholar] [CrossRef]
- Dussert, S.; Mauro, M.C.; Deloire, A.; Hamon, A.; Engelmann, F. Cryopreservation of grape embryogenic suspensions. 1. Influence of pretreatment, freezing and thawing conditions. Cryoletters 1991, 12, 287–298. [Google Scholar]
- Gonzalez-Benito, M.E.; Martin, C.; Vidal, J.R. Cryopreservation of embryogenic cell suspensions of the Spanish grapevine cultivars 'Albarino' and 'Tempranillo'. Vitis 2009, 48, 131–136. [Google Scholar]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Reports 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Pathirana, R. Cryopreserved grapevine: a new way to maintain high-health germplasm and cultivar imports with less rigorous quarantine; NZW 10-107; Plant and Food Research: New Zealand, June 2011 2011; p. 14. [Google Scholar]
- Pathirana, R.; Mathew, L.; Jibran, R.; Hunter, D.A.; Morgan, E.R. Cryopreservation and cryotherapy research on horticultural crops in New Zealand. Acta Horticulturae 2019, 1234, 29–36. [Google Scholar] [CrossRef]
- Pathirana, R.; McLachlan, A.; Hedderley, D.; Panis, B.; Carimi, F. Pre-treatment with salicylic acid improves plant regeneration after cryopreservation of grapevine (Vitis spp.) by droplet vitrification. Acta Physiologiae Plantarum 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Carimi, F.; Carra, A.; Panis, B.; Pathirana, R. Strategies for conservation of endangered wild grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi). Acta Horticulturae 2016, 1115, 81–86. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Bonnart, R.; Shepherd, A.N.; Kretzschmar, A.A.; Volk, G.M. Modifications to a Vitis shoot tip cryopreservation procedure: Effect of shoot tip size and use of cryoplates. Cryoletters 2019, 40, 103–112. [Google Scholar] [PubMed]
- Gribaudo, I.; Vallania, R.; Miaja, M.L.; Franks, T.; Thomas, M. Genotype influence on somatic embryogenesis in grapevine anther and leaf cultures. Acta Horticulturae 2000, 528, 407–410. [Google Scholar] [CrossRef]
- Kikkert, J.R.; Striem, M.J.; Vidal, J.R.; Wallace, P.G.; Barnard, J.; Reisch, B.I. Long-term study of somatic embryogenests from anthers and ovaries of 12 grapevine (Vitis sp.) genotypes. In Vitro Cellular & Developmental Biology-Plant 2005, 41, 232–239. [Google Scholar]
- Perrin, M.; Gertz, C.; Masson, J.E. High efficiency initiation of regenerable embryogenic callus from anther filaments of 19-grapevine genotypes grown worldwide. Plant Science 2004, 167, 1343–1349. [Google Scholar] [CrossRef]
- Franks, T.; Botta, R.; Thomas, M.R.; Franks, J. Chimerism in grapevines: implications for cultivar identity, ancestry and genetic improvement. TAG Theoretical and Applied Genetics 2002, 104, 192–199. [Google Scholar] [CrossRef]
- Gambino, G.; Moine, A.; Boccacci, P.; Perrone, I.; Pagliarani, C. Somatic embryogenesis is an effective strategy for dissecting chimerism phenomena in Vitis vinifera cv Nebbiolo. Plant Cell Reports 2021, 40, 205–211. [Google Scholar] [CrossRef]
- Riaz, S.; Garrison, K.E.; Dangl, G.S.; Boursiquot, J.-M.; Meredith, C.P. Genetic Divergence and Chimerism within Ancient Asexually Propagated Winegrape Cultivars. J. Amer. Soc. Hort. Sci. 2002, 127, 508–514. [Google Scholar] [CrossRef]
- Prado, M.J.; Rodriguez, E.; Rey, L.; Gonzalez, M.V.; Santos, C.; Rey, M. Detection of somaclonal variants in somatic embryogenesis-regenerated plants of Vitis vinifera by flow cytometry and microsatellite markers. Plant Cell Tissue and Organ Culture 2010, 103, 49–59. [Google Scholar] [CrossRef]
- Schneider, S.; Reustle, G.; Zyprian, E. Detection of somaclonal variation in grapevine regenerants from protoplasts by RAPD-PCR. Vitis 1996, 35, 99–100. [Google Scholar]
- Gardiman, M.; Carimi, F.; Meneghetti, S.; Barizza, E.; Schiavo, F.l. Micropropagation and genetic stability of Vitis vinifera, cv Aglianico, plants obtained by somatic embryo-genesis. Italus Hortus 2009, 16, 124–127. [Google Scholar]
- Martelli, G. Where grapevine virology is heading to. In Proceedings of the Proceedings of the 19th Congress of ICVG, Santiago, Chile, April 9-12, 2018, 2018; pp. 10–15.
- Fuchs, M.; Lemaire, O. Novel approaches for viral disease management. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishers: 2017; pp. 599–621.
- Golino, D.; Fuchs, M.; Sim, S.; Farrar, K.; Martelli, G. Improvement of grapevine planting stock through sanitary selection and pathogen elimination. In Grapevine Viruses: Molecular Biology, Diagnostics and Management, Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishers: 2017; pp. 561–579.
- Gambino, G.; Di Matteo, D.; Gribaudo, I. Elimination of Grapevine fanleaf virus from three Vitis vinifera cultivars by somatic embryogenesis. European Journal of Plant Pathology 2009, 123, 57–60. [Google Scholar] [CrossRef]
- Bi, W.-L.; Hao, X.-Y.; Cui, Z.-H.; Pathirana, R.; Volk, G.M.; Wang, Q.-C. Shoot tip cryotherapy for efficient eradication of grapevine leafroll-associated virus-3 from diseased grapevine in vitro plants. Annals of Applied Biology 2018, 173, 261–270. [Google Scholar] [CrossRef]
- Bradamante, G.; Mittelsten Scheid, O.; Incarbone, M. Under siege: virus control in plant meristems and progeny. The Plant Cell 2021, 33, 2523–2537. [Google Scholar] [CrossRef] [PubMed]
- Mathew, L.; Tiffin, H.; Erridge, Z.; McLachlan, A.; Hunter, D.; Pathirana, R. Efficiency of eradication of Raspberry bushy dwarf virus from infected raspberry (Rubus idaeus) by in vitro chemotherapy, thermotherapy and cryotherapy and their combinations. Plant Cell Tiss Organ Cult 2021, 144, 133–141. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Dalla Costa, M.; Gardin, J.P.P.; Kretzschmar, A.A.; Pathirana, R. Cryotherapy: a new technique to obtain grapevine plants free of viruses. Revista Brasileira De Fruticultura 2016, 38. [Google Scholar] [CrossRef]
- Kim, M.Y.; Cho, K.H.; Chun, J.A.; Park, S.J.; Kim, S.H.; Lee, H.C. Elimination ofGrapevine fleck virusfrom infected grapevines ‘Kyoho’ through meristem-tip culture of dormant buds. Journal of Plant Biotechnology 2017, 44, 401–408. [Google Scholar] [CrossRef]
- Bayati, S.; Shams-Bakhsh, M.; Moieni, A. Elimination of Grapevine Virus A (GVA) by Cryotherapy and Electrotherapy. Journal of Agricultural Science and Technology 2011, 13, 443–450. [Google Scholar]
- Markovic, Z.; Preiner, D.; Stupic, D.; Andabaka, Z.; Simon, S.; Voncina, D.; Maletic, E.; Kontic, J.K.; Chatelet, P.; Engelmann, F. Cryopreservation and cryotherapy of grapevine (Vitis vinifera L.). Vitis 2015, 54, 247–251. [Google Scholar]
- Guta, I.C.; Buciumeanu, E.-C. Grapevine chemotherapy for elimination of multiple virus infection. Romanian Biotechnological Letters 2011, 16, 6535–6539. [Google Scholar]
- Skiada, F.G.; Maliogka, V.I.; Katis, N.I.; Eleftheriou, E.P. Elimination of Grapevine rupestris stem pitting-associated virus (GRSPaV) from two Vitis vinifera cultivars by in vitro chemotherapy. European Journal of Plant Pathology 2013, 135, 407–414. [Google Scholar] [CrossRef]
- Infante, R.; Fiore, N. Combined effect of thermotherapy and in vitro shoot culture on the Grapevine leafroll associated virus 2 Red Globe strain affecting 'Red Globe' vines. Journal of Food Agriculture & Environment 2009, 7, 274–277. [Google Scholar]
- Maliogka, V.I.; Skiada, F.G.; Eleftheriou, E.P.; Katis, N.I. Elimination of a new ampelovirus (GLRaV-Pr) and Grapevine rupestris stem pitting associated virus (GRSPaV) from two Vitis vinifera cultivars combining in vitro thermotherapy with shoot tip culture. Scientia Horticulturae 2009, 123, 280–282. [Google Scholar] [CrossRef]
- Nakano, M.; Watanabe, Y.; Hoshino, Y. Histological examination of callogenesis and adventitious embryogenesis in immature ovary culture of grapevine (Vitis vinifera L.). The Journal of Horticultural Science and Biotechnology 2000, 75, 154–160. [Google Scholar] [CrossRef]
- Newton, D.; Goussard, P. The ontogeny of somatic embryos from in vitro cultured grapevine anthers. South African Journal of Enology & Viticulture 1990, 11, 70–75. [Google Scholar]
- Goussard, P.G.; Wiid, J.; Kasdon, G.G.F. The Effectiveness of in vitro somatic embryogenesis in eliminating fanleaf virus and leafroll associated viruses from grapevines. South African Journal of Enology & Viticulture 1991, 12, 77–81. [Google Scholar] [CrossRef]
- Goussard, P.; Wiid, J. The elimination of fanleaf virus from grapevines using in vitro somatic embryogenesis combined with heat therapy. South Afrrican Journal of Enolology and Viticulture 1992, 13, 81–83. [Google Scholar] [CrossRef]
- Gambino, G.; Bondaz, J.; Gribaudo, I. Detection and elimination of viruses in callus, somatic embryos and regenerated plantlets of grapevine. European Journal of Plant Pathology 2006, 114, 397–404. [Google Scholar] [CrossRef]
- Turcsan, M.; Demian, E.; Varga, T.; Jaksa-Czotter, N.; Szegedi, E.; Olah, R.; Varallyay, E. Hts-based monitoring of the efficiency of somatic embryogenesis and meristem cultures used for virus elimination in grapevine. Plants 2020, 9, 1782. [Google Scholar] [CrossRef]
- Wang, Q.C.; Panis, B.; Engelmann, F.; Lambardi, M.; Valkonen, J.P.T. Cryotherapy of shoot tips: a technique for pathogen eradication to produce healthy planting materials and prepare healthy plant genetic resources for cryopreservation. Annals of Applied Biology 2009, 154, 351–363. [Google Scholar] [CrossRef]
- Xu, Y.; Jiang, N.; Zhang, Y.; Wang, M.; Ren, J.; Tao, J. A SNP in the promoter region of theVvmybA1 gene is responsible for differences in grape berry color between two related bud sports of grape. Plant Growth Regulation 2017, 82, 457–465. [Google Scholar] [CrossRef]
- Carrier, G.; Le Cunff, L.; Dereeper, A.; Legrand, D.; Sabot, F.; Bouchez, O.; Audeguin, L.; Boursiquot, J.-M.; This, P. Transposable elements are a major cause of somatic polymorphism in Vitis vinifera L. PLOS ONE 2012, 7, e32973. [Google Scholar] [CrossRef]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. TRENDS in Genetics 2006, 22, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Pathirana, R. Mutations in plant evolution, crop domestication and breeding. Tropical Agricultural Research and Extension 2021, 24. [Google Scholar] [CrossRef]
- Ahloowalia, B.S.; Maluszynski, M. Induced mutations - A new paradigm in plant breeding. Euphytica 2001, 118, 167–173. [Google Scholar] [CrossRef]
- Pathirana, R.; Deroles, S.; Hoeata, K.; Montefiori, M.; Tyson, J.; Wang, T.; Datson, P.M.; Hellens, R.P. Fast-tracking kiwifruit breeding through mutagenesis. Acta Horticulturae 2016, 1127, 217–222. [Google Scholar] [CrossRef]
- Maluszynski, M.; Ahloowalia, B.S.; Sigurbjörnsson, B. Application of in vivo and in vitro mutation techniques for crop improvement. Euphytica 1995, 81, 303–315. [Google Scholar] [CrossRef]
- Pathirana, R. Plant mutation breeding in agriculture. CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources 2011, 6, 1–20.
- Khawale, R.N.; Yerramilli, V.; Singh, S.K. Molecular marker-assisted selection of in vitro chemical mutagen-induced grapevine mutants. Current Science 2007, 92, 1056–1060. [Google Scholar]
- Munir, N.; Safdar, I.; Naz, S. Effect of induced mutation for varietal improvement in some local grapevine cultivars. 2015.
- Krul, W.R.; Worley, J.F. Formation of adventitious embryos in callus cultures of 'Seyval', a French hybrid grape. J. Am. Soc. Hortic. Sci. 1977, 102, 360–363. [Google Scholar] [CrossRef]
- Faure, O.; Aarrouf, J.; Nougarede, A. Ontogenesis, differentiation and precocious germination in anther-derived somatic embryos of grapevine (Vitis vinifera L.): Proembryogenesis. Annals of Botany 1996, 78, 23–28. [Google Scholar] [CrossRef]
- Bhojwani, S.S.; Dantu, P.K. Somatic embryogenesis. In Plant Tissue Culture: An Introductory Text, Bhojwani, S.S., Dantu, P.K., Eds.; Springer India: India, 2013; pp. 75–92. [Google Scholar]
- Nagmani, R.; Becwar, M.; Wann, S. Single-cell origin and development of somatic embryos in Picea abies (L.) Karst.(Norway spruce) and P. glauca (Moench) Voss (white spruce). Plant Cell Reports 1987, 6, 157–159. [Google Scholar] [CrossRef]
- Toonen, M.A.; Hendriks, T.; Schmidt, E.D.; Verhoeven, H.A.; Van Kammen, A.; de Vries, S.C. Description of somatic-embryo-forming single cells in carrot suspension cultures employing video cell tracking. Planta 1994, 194, 565–572. [Google Scholar] [CrossRef]
- Masoabi, M.; Lloyd, J.; Kossmann, J.; van der Vyver, C. Ethyl methanesulfonate mutagenesis and in vitro polyethylene glycol selection for drought tolerance in sugarcane (Saccharum spp.). Sugar Tech 2018, 20, 50–59. [Google Scholar] [CrossRef]
- Nikam, A.A.; Devarumath, R.M.; Ahuja, A.; Babu, H.; Shitole, M.G.; Suprasanna, P. Radiation-induced in vitro mutagenesis system for salt tolerance and other agronomic characters in sugarcane (Saccharum officinarum L.). The Crop Journal 2015, 3, 46–56. [Google Scholar] [CrossRef]
- Patade, V.Y.; Suprasanna, P.; Bapat, V.A. Gamma irradiation of embryogenic callus cultures and in vitro selection for salt tolerance in sugarcane (Saccharum officinarum L.). Agricultural Sciences in China 2008, 7, 1147–1152. [Google Scholar] [CrossRef]
- Pathirana, R.; Carimi, F.; Carra, A.; Cheah, L.-H. Intergrating biotechnological advancements with induced mutagenesis: New opportunities for horticulture with special reference to Vitis vinifera. In Proceedings of the FAO/IAEA International Symposium on Induced Mutations in Plants., Vienna, Austria, 2008, 12-15 August 2008; p. 115. [Google Scholar]
- Kuksova, V.B.; Piven, N.M.; Gleba, Y.Y. Somaclonal variation and in vitro induced mutagenesis in grapevine. Plant Cell, Tissue and Organ Culture 1997, 49, 17–27. [Google Scholar] [CrossRef]
- Yang, X.M.; Cao, Z.Y.; An, L.Z.; Wang, Y.M.; Fang, X.W. In vitro tetraploid induction via colchicine treatment from diploid somatic embryos in grapevine (Vitis vinifera L.). Euphytica 2006, 152, 217. [Google Scholar] [CrossRef]
- Capriotti, L.; Sabbadini, S.; Limera, C.; Navacchi, O.; Mezzetti, B. Somaclonal variation and induced mutagenesis in several grapevine cultivars. Acta Horticulturae 2022, 1359, 131–138. [Google Scholar] [CrossRef]
- Baduel, P.; Colot, V. The epiallelic potential of transposable elements and its evolutionary significance in plants. Philosophical Transactions of the Royal Society B 2021, 376, 20200123. [Google Scholar] [CrossRef]
- McClintock, B. The significance of responses of the genome to challenge. Science 1984, 226, 792–801. [Google Scholar] [CrossRef]
- McClintock, B. Controlling elements and the gene. In Proceedings of the Cold Spring Harbor symposia on quantitative biology; 1956; pp. 197–216. [Google Scholar]
- Lijavetzky, D.; Ruiz-García, L.; Cabezas, J.A.; De Andrés, M.T.; Bravo, G.; Ibáñez, A.; Carreño, J.; Cabello, F.; Ibáñez, J.; Martínez-Zapater, J.M. Molecular genetics of berry colour variation in table grape. Molecular Genetics and Genomics 2006, 276, 427–435. [Google Scholar] [CrossRef]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982–982. [Google Scholar] [CrossRef] [PubMed]
- Benjak, A.; Forneck, A.; Casacuberta, J.M. Genome-Wide Analysis of the “Cut-and-Paste” Transposons of Grapevine. PLOS ONE 2008, 3, e3107. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Cao, S.; Wang, X.; Huang, S.; Wang, Y.; Liu, Z.; Liu, W.; Leng, X.; Peng, Y.; Wang, N.; et al. The complete reference genome for grapevine (Vitis vinifera L.) genetics and breeding. Horticulture Research 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, L.; Gribaudo, I.; Bertoldi, D.; Candioli, E.; Poletti, V. High efficiency somatic embryogenesis and plant germination in grapevine cultivars Chardonnay and Brachetto a grappolo lungo. Vitis 2001, 40, 111–115. [Google Scholar]
- Vivier, M.A.; Pretorius, I.S. Genetic improvement of grapevine: tailoring grape varieties for the third millennium - a review. South African Journal for Enology and Viticulture 2000, 21, 5–26. [Google Scholar] [CrossRef]
- Campos, G.; Chialva, C.; Miras, S.; Lijavetzky, D. New technologies and strategies for grapevine breeding through genetic transformation. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef]
- Boccacci, P.; Mela, A.; Pavez Mina, C.; Chitarra, W.; Perrone, I.; Gribaudo, I.; Gambino, G. Cultivar-specific gene modulation in Vitis vinifera: analysis of the promoters regulating the expression of WOX transcription factors. Scientific Reports 2017, 7, 45670. [Google Scholar] [CrossRef]
- Nuzzo, F.; Moine, A.; Nerva, L.; Pagliarani, C.; Perrone, I.; Boccacci, P.; Gribaudo, I.; Chitarra, W.; Gambino, G. Grapevine virome and production of healthy plants by somatic embryogenesis. Microbial Biotechnology 2022, 15, 1357–1373. [Google Scholar] [CrossRef]
- Ali, K.; Maltese, F.; Choi, Y.H.; Verpoorte, R. Metabolic constituents of grapevine and grape-derived products. Phytochem Rev 2010, 9, 357–378. [Google Scholar] [CrossRef]
- Palacio-Bielsa, A.; González-Abolafio, R.; Álvarez, B.; Lastra, B.; Cambra, M.A.; Salcedo, C.I.; López, M.M.; Penyalver, R. Chromosomal and Ti plasmid characterization of tumorigenic strains of three Agrobacterium species isolated from grapevine tumours. Plant Pathology 2009, 58, 584–593. [Google Scholar] [CrossRef]
- Kawaguchi, A. Biological control of crown gall on grapevine and root colonization by nonpathogenic Rhizobium vitis strain ARK-1. Microbes and environments 2013, 28, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, A.R.; Cavallini, E.; Jia, Y.; Moss, S.M.; McDavid, D.A.; Hooper, L.C.; Robinson, S.P.; Tornielli, G.B.; Zenoni, S.; Ford, C.M. A grapevine anthocyanin acyltransferase, transcriptionally regulated by VvMYBA, can produce most acylated anthocyanins present in grape skins. Plant Physiology 2015, 169, 1897–1916. [Google Scholar] [CrossRef] [PubMed]
- Capriotti, L.; Baraldi, E.; Mezzetti, B.; Limera, C.; Sabbadini, S. Biotechnological approaches: gene overexpression, gene silencing, and genome editing to control fungal and oomycete diseases in grapevine. International Journal of Molecular Sciences 2020, 21, 5701. [Google Scholar] [CrossRef]
- Dry, I.; Riaz, S.; Fuchs, M.; Sosnowski, M.; Thomas, M. Scion breeding for resistance to biotic stresses. In The Grape Genome, Cantu, D., Walker, M., Eds.; Springer, Cham.: 2019; pp. 319–347.
- Paul, N.C.; Park, S.-W.; Liu, H.; Choi, S.; Ma, J.; MacCready, J.S.; Chilvers, M.I.; Sang, H. Plant and fungal genome editing to enhance plant disease resistance using the CRISPR/Cas9 system. Frontiers in Plant Science 2021, 12, 700925. [Google Scholar] [CrossRef]
- Ren, C.; Liu, Y.; Guo, Y.; Duan, W.; Fan, P.; Li, S.; Liang, Z. Optimizing the CRISPR/Cas9 system for genome editing in grape by using grape promoters. Horticulture Research 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tu, M.; Wang, Y.; Yin, W.; Zhang, Y.; Wu, H.; Gu, Y.; Li, Z.; Xi, Z.; Wang, X. Whole-genome sequencing reveals rare off-target mutations in CRISPR/Cas9-edited grapevine. Horticulture research 2021, 8. [Google Scholar]
- Wan, D.-Y.; Guo, Y.; Cheng, Y.; Hu, Y.; Xiao, S.; Wang, Y.; Wen, Y.-Q. CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Horticulture research 2020, 7. [Google Scholar] [CrossRef]
- Vidal, J.; Kikkert, J.; Wallace, P.; Reisch, B. High-efficiency biolistic co-transformation and regeneration of 'Chardonnay' (Vitis vinifera L.) containing npt-II and antimicrobial peptide genes. Plant Cell Reports 2003, 22, 252–260. [Google Scholar] [CrossRef]
- Dhekney, S.A.; Li, Z.T.; Gray, D.J. Grapevines engineered to express cisgenic Vitis vinifera thaumatin-like protein exhibit fungal disease resistance. In Vitro Cellular & Developmental Biology-Plant 2011, 47, 458–466. [Google Scholar]
- Dal Bosco, D.; Sinski, I.; Ritschel, P.S.; Camargo, U.A.; Fajardo, T.V.; Harakava, R.; Quecini, V. Expression of disease resistance in genetically modified grapevines correlates with the contents of viral sequences in the T-DNA and global genome methylation. Transgenic Research 2018, 27, 379–396. [Google Scholar] [CrossRef]
- Hemmer, C.; Djennane, S.; Ackerer, L.; Hleibieh, K.; Marmonier, A.; Gersch, S.; Garcia, S.; Vigne, E.; Komar, V.; Perrin, M. Nanobody-mediated resistance to Grapevine fanleaf virus in plants. Plant Biotechnology Journal 2018, 16, 660–671. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Wu, J.; Zhang, Y.; Agüero, C.B.; Li, X.; Liu, S.; Wang, C.; Walker, M.A.; Lu, J. Overexpression of a thaumatin-like protein gene from Vitis amurensis improves downy mildew resistance in Vitis vinifera grapevine. Protoplasma 2017, 254, 1579–1589. [Google Scholar] [CrossRef]
- Zou, C.; Karn, A.; Reisch, B.; Nguyen, A.; Sun, Y.; Bao, Y.; Campbell, M.S.; Church, D.; Williams, S.; Xu, X.; et al. Haplotyping the Vitis collinear core genome with rhAmpSeq improves marker transferability in a diverse genus. Nat Commun 2020, 11, 413. [Google Scholar] [CrossRef] [PubMed]
- Guillaumie, S.; Mzid, R.; Méchin, V.; Léon, C.; Hichri, I.; Destrac-Irvine, A.; Trossat-Magnin, C.; Delrot, S.; Lauvergeat, V. The grapevine transcription factor WRKY2 influences the lignin pathway and xylem development in tobacco. Plant molecular biology 2010, 72, 215–234. [Google Scholar] [CrossRef]
- Giorgetti, L.; RUFFINI CASTIGLIONE, M.; TURRINI, A.; ronchi Vittoria, N.; Chiara, G. Cytogenetic and histological approach for early detection of “mantled” somaclonal variants of oil palm regenerated by somatic embryogenesis: first results on the characterization of regeneration system. Caryologia 2011, 64, 221–232. [Google Scholar]
- Ronchi, V.N. Cytogenetics of plant cell cultures. In Developments in Crop Science, Bhojwani, S.S., Ed.; Elsevier: 1990; Volume 19, pp. 276–300.
- Giorgetti, L.; Vergara, M.; Evangelista, M.; Lo Schiavo, F.; Terzi, M.; Nuti Ronchi, V. On the occurrence of somatic meiosis in embryogenic carrot cell cultures. Molecular and General Genetics MGG 1995, 246, 657–662. [Google Scholar] [CrossRef]
- Ronchi, V.N.; Giorgetti, L.; Tonelli, M.; Martini, G. Ploidy reduction and genome segregation in cultured carrot cell lines. I. Prophase chromosome reduction. Plant Cell, Tissue and Organ Culture 1992, 30, 107–114. [Google Scholar] [CrossRef]
- Murray, S.C.; Eckhoff, P.; Wood, L.; Paterson, A.H. A proposal to use gamete cycling in vitro to improve crops and livestock. Nature Biotechnology 2013, 31, 877–880. [Google Scholar] [CrossRef]
- De La Fuente, G.N.; Frei, U.K.; Lübberstedt, T. Accelerating plant breeding. Trends in Plant Science 2013, 18, 667–672. [Google Scholar] [CrossRef]
- Cook, T.M.; Isenegger, D.; Dutta, S.; Sahab, S.; Kay, P.; Aboobucker, S.I.; Biswas, E.; Heerschap, S.; Nikolau, B.J.; Dong, L. Overcoming roadblocks for in vitro nurseries in plants: induction of meiosis. Frontiers in Plant Science 2023, 14, 1204813. [Google Scholar] [CrossRef]
- Yoshida, H.; Yamaguchi, H. Arrangement and association of somatic chromosomes induced by chloramphenicol in barley. Chromosoma 1973, 43, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, L.; Zhou, Y.; Geng, Y.; Chen, Z. Inducing somatic meiosis-like reduction at high frequency by caffeine in root-tip cells of Vicia faba. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis 2000, 452, 67–72. [Google Scholar] [CrossRef]
- Jiang, F.; Ryabova, D.; Diedhiou, J.; Hucl, P.; Randhawa, H.; Marillia, E.; Foroud, N.; Eudes, F.; Kathiria, P. Trichostatin A increases embryo and green plant regeneration in wheat. Plant Cell Reports 2017, 36, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Soriano, M.; Cordewener, J.; Muiño, J.M.; Riksen, T.; Fukuoka, H.; Angenent, G.C.; Boutilier, K. The histone deacetylase inhibitor trichostatin a promotes totipotency in the male gametophyte. The Plant Cell 2014, 26, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Enns, J.L.; Nelson, K.L.; Brost, J.M.; Orr, T.D.; Ferrie, A.M.R. Improving the efficiency of wheat microspore culture methodology: evaluation of pretreatments, gradients, and epigenetic chemicals. Plant Cell, Tissue, and Organ Culture 2019, 139, 589–599. [Google Scholar] [CrossRef]
- Bouamama-Gzara, B.; Selmi, I.; Chebil, S.; Melki, I.; Mliki, A.; Ghorbel, A.; Carra, A.; Carimi, F.; Mahfoudhi, N. Elimination of Grapevine leafroll associated virus-3, Grapevine rupestris stem pitting associated virus and Grapevine virus A from a Tunisian cultivar by somatic embryogenesis and characterization of the somaclones using ampelographic descriptors. The Plant Pathology Journal 2017, 33, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Butiuc-Keul, A.; Coste, A. Biotechnologies and strategies for grapevine improvement. Horticulturae 2023, 9, 62. [Google Scholar] [CrossRef]
- Aljuaid, B.S.; Ismail, I.A.; Attia, A.O.; El Dessoky, S.D. Genetic stability of in vitro propagated grapevine (Vitis vinifera L.) cv. Al-Bayadi. Journal of Agriculture and Crops 2022, 8, 12–19. [Google Scholar] [CrossRef]
- Baránek, M.; Raddová, J.; Krizan, B.; Pidra, M. Genetic changes in grapevine genomes after stress induced by in vitro cultivation, thermotherapy and virus infection, as revealed by AFLP. Genetics and Molecular Biology 2009, 32, 834–839. [Google Scholar] [CrossRef]
- Crăciunaş, C.; Coste, A.; Oltean, B.; Farago, M.; Iliescu, M.; Iuoras, R.; Butiuc-Keul, A. Genetic stability of several cultivars of grapevine cultivated in vitro. Acta Horticulturae 2007, 812, 515–520. [Google Scholar] [CrossRef]
- Gribaudo, I.; Marinoni, D.T.; Gambino, G.; Mannini, F.; Akkak, A.; Botta, R. Assessment of genetic fidelity in regenerants from two Vitis vinifera cultivars. In Ix International Conference on Grape Genetics and Breeding, Peterlunger, E., DiGaspero, G., Cipriani, G., Eds.; Acta Horticulturae; 2009; Volume 827, pp. 131–135.
- Schellenbaum, P.; Mohler, V.; Wenzel, G.; Walter, B. Variation in DNA methylation patterns of grapevine somaclones (Vitis vinifera L.). BMC Plant Biology 2008, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.; Kumar, N. Application of molecular markers for the assessment of genetic fidelity of in vitro raised plants: current status and future prospects. In Molecular Marker Techniques: A Potential Approach of Crop Improvement, Kumar, N., Ed.; Springer: Singapore, 2023; pp. 233–256. [Google Scholar]
- Nybom, H.; Weising, K.; Rotter, B. DNA fingerprinting in botany: past, present, future. Investigative Genetics 2014, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Matus, J.T.; Pinto-Carnide, O.; Carrasco, D.; Arroyo-García, R.; Castro, I. Genetic analysis of a white-to-red berry skin color reversion and its transcriptomic and metabolic consequences in grapevine (Vitis vinifera cv.‘Moscatel Galego’). BMC Genomics 2019, 20, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sichel, V.; Sarah, G.; Girollet, N.; Laucou, V.; Roux, C.; Roques, M.; Mournet, P.; Cunff, L.L.; Bert, P.F.; This, P.; et al. Chimeras in Merlot grapevine revealed by phased assembly. BMC Genomics 2023, 24, 396. [Google Scholar] [CrossRef]
- Bertsch, C.; Kieffer, F.; Maillot, P.; Farine, S.; Butterlin, G.; Merdinoglu, D.; Walter, B. Genetic chimerism of Vitis vinifera cv. Chardonnay 96 is maintained through organogenesis but not somatic embryogenesis. BMC Plant Biology 2005, 5, 20. [Google Scholar] [CrossRef]
- Forneck, A. Plant breeding: clonality—a concept for stability and variability during vegetative propagation. In Progress in Botany: Genetics Physiology Systematics Ecology, Esser, K., Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin, Heidelberg, 2005; Volume 66, pp. 164–183. [Google Scholar]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC plant biology 2013, 13, 1–17. [Google Scholar] [CrossRef]
Figure 1.
Somatic embryogenesis and plant regeneration from immature flower tissues in Vitis vinifera. (A) Immature flowers are collected a few days before opening and stored at 4 °C. (B) Different floral tissues utilized to induce embryogenic cell lines (Anthers, Pistils, Stigmas/Styles, Ovary, Pistils, Whole flowers). (C) Callus generated from a pistil (arrow) after 3 months of culture on embryogenic medium. (D) Somatic embryos regenerated after 4-6 months of culture initiation at the surface of explant-derived callus. (E) Different developmental stages of somatic embryos regenerated in vitro.
Figure 1.
Somatic embryogenesis and plant regeneration from immature flower tissues in Vitis vinifera. (A) Immature flowers are collected a few days before opening and stored at 4 °C. (B) Different floral tissues utilized to induce embryogenic cell lines (Anthers, Pistils, Stigmas/Styles, Ovary, Pistils, Whole flowers). (C) Callus generated from a pistil (arrow) after 3 months of culture on embryogenic medium. (D) Somatic embryos regenerated after 4-6 months of culture initiation at the surface of explant-derived callus. (E) Different developmental stages of somatic embryos regenerated in vitro.

Figure 2.
Development of somatic embryos and plantlets from cell suspension cultures of grapevine. (A) Pro-embryogenic masses (200-400 mg) are used for culture initiation. (B) Liquid cultures are maintained in 250-ml Erlenmeyer flasks containing 50 ml of liquid culture medium. (C) Images of cells growing in liquid culture and somatic embryos (globule and heart-shaped stages) differentiated after 40 days of initiation of culture. (D) Somatic embryos, collected by filtration after 2-3 months from the start of culture using nylon mesh filter (2 mm), are incubated on growth regulator-free liquid medium. (E) Germination of the embryos occurs after approximately 30 days of culture on growth regulator free solid medium. (F) The plantlets are acclimated in Jiffy pots and reach about 15 cm in height in about 40-60 days.
Figure 2.
Development of somatic embryos and plantlets from cell suspension cultures of grapevine. (A) Pro-embryogenic masses (200-400 mg) are used for culture initiation. (B) Liquid cultures are maintained in 250-ml Erlenmeyer flasks containing 50 ml of liquid culture medium. (C) Images of cells growing in liquid culture and somatic embryos (globule and heart-shaped stages) differentiated after 40 days of initiation of culture. (D) Somatic embryos, collected by filtration after 2-3 months from the start of culture using nylon mesh filter (2 mm), are incubated on growth regulator-free liquid medium. (E) Germination of the embryos occurs after approximately 30 days of culture on growth regulator free solid medium. (F) The plantlets are acclimated in Jiffy pots and reach about 15 cm in height in about 40-60 days.

Figure 3.
A scheme for using embryogenic cultures for mutation induction and screening. a) Embryogenic culture establishment, b) Optimising mutagen dose through growth reduction studies [43], c) Development of somatic embryos after treatment with optimized mutagen dose, d) Initial germination and e) Screening the germinated embryos for the trait of interest in vitro, f) and g) Testing mutagenized population in greenhouse and field, respectively, for traits difficult to screen in vitro such as bunch architecture, vine growth, fruit quality in table grapes etc. .
Figure 3.
A scheme for using embryogenic cultures for mutation induction and screening. a) Embryogenic culture establishment, b) Optimising mutagen dose through growth reduction studies [43], c) Development of somatic embryos after treatment with optimized mutagen dose, d) Initial germination and e) Screening the germinated embryos for the trait of interest in vitro, f) and g) Testing mutagenized population in greenhouse and field, respectively, for traits difficult to screen in vitro such as bunch architecture, vine growth, fruit quality in table grapes etc. .
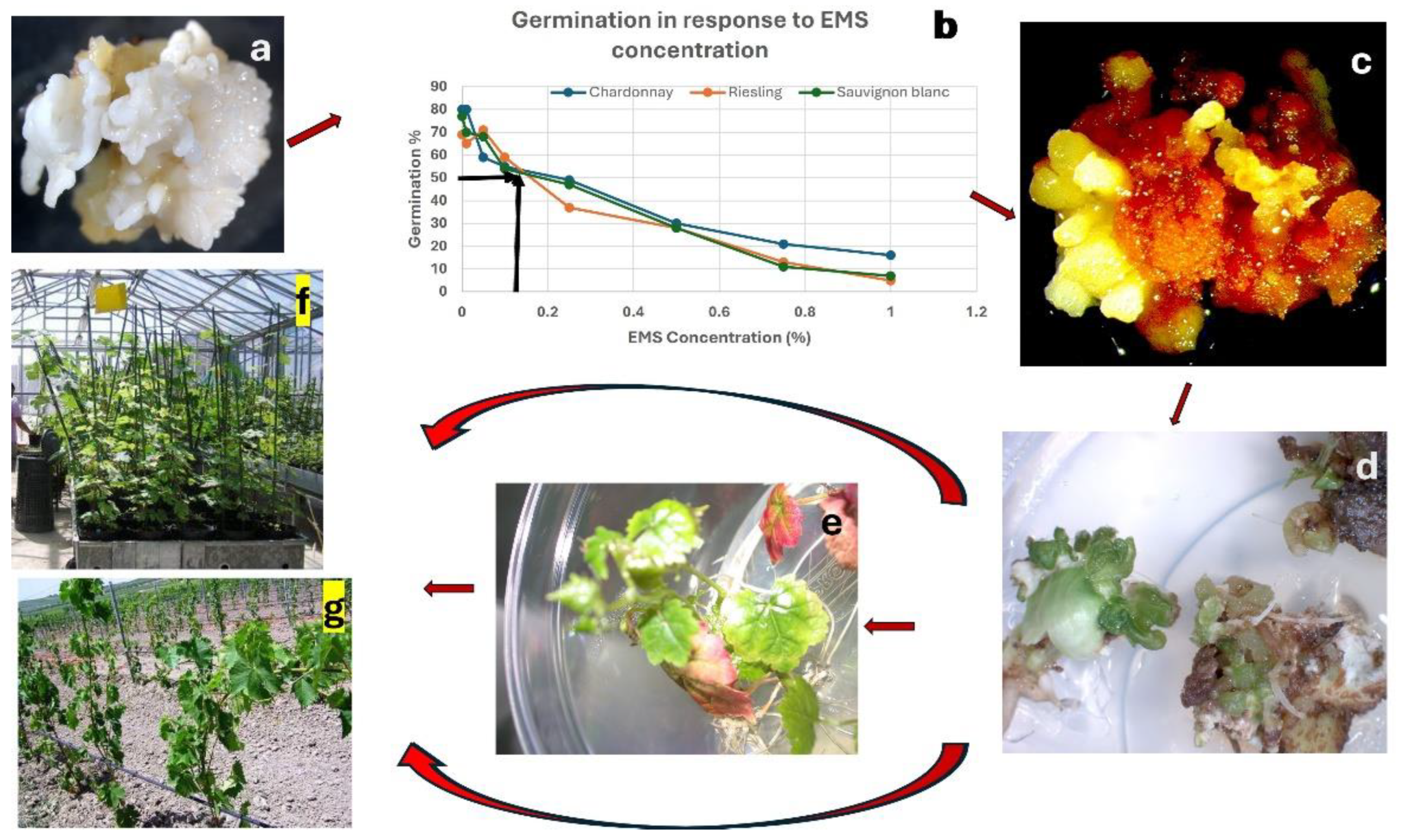
Figure 4.
A scheme for transposon activation, tagging mutants and their recovery using embryogenic cultures of grapevine. a) Cell culture established and subjected to stress for transposon activation, b) Cells subjected to stress, c) Somatic embryos (SEs) generated from stressed embryogenic cells, d) Secondary SEs induced from primary SEs and the clusters serially numbered, e) Part of the labelled secondary SEs cryopreserved, f) Other part of SEs subjected to RNAseq for transposon tagging, g) Identified mutants of interest recovered from cryopreservation and regenerated.
Figure 4.
A scheme for transposon activation, tagging mutants and their recovery using embryogenic cultures of grapevine. a) Cell culture established and subjected to stress for transposon activation, b) Cells subjected to stress, c) Somatic embryos (SEs) generated from stressed embryogenic cells, d) Secondary SEs induced from primary SEs and the clusters serially numbered, e) Part of the labelled secondary SEs cryopreserved, f) Other part of SEs subjected to RNAseq for transposon tagging, g) Identified mutants of interest recovered from cryopreservation and regenerated.
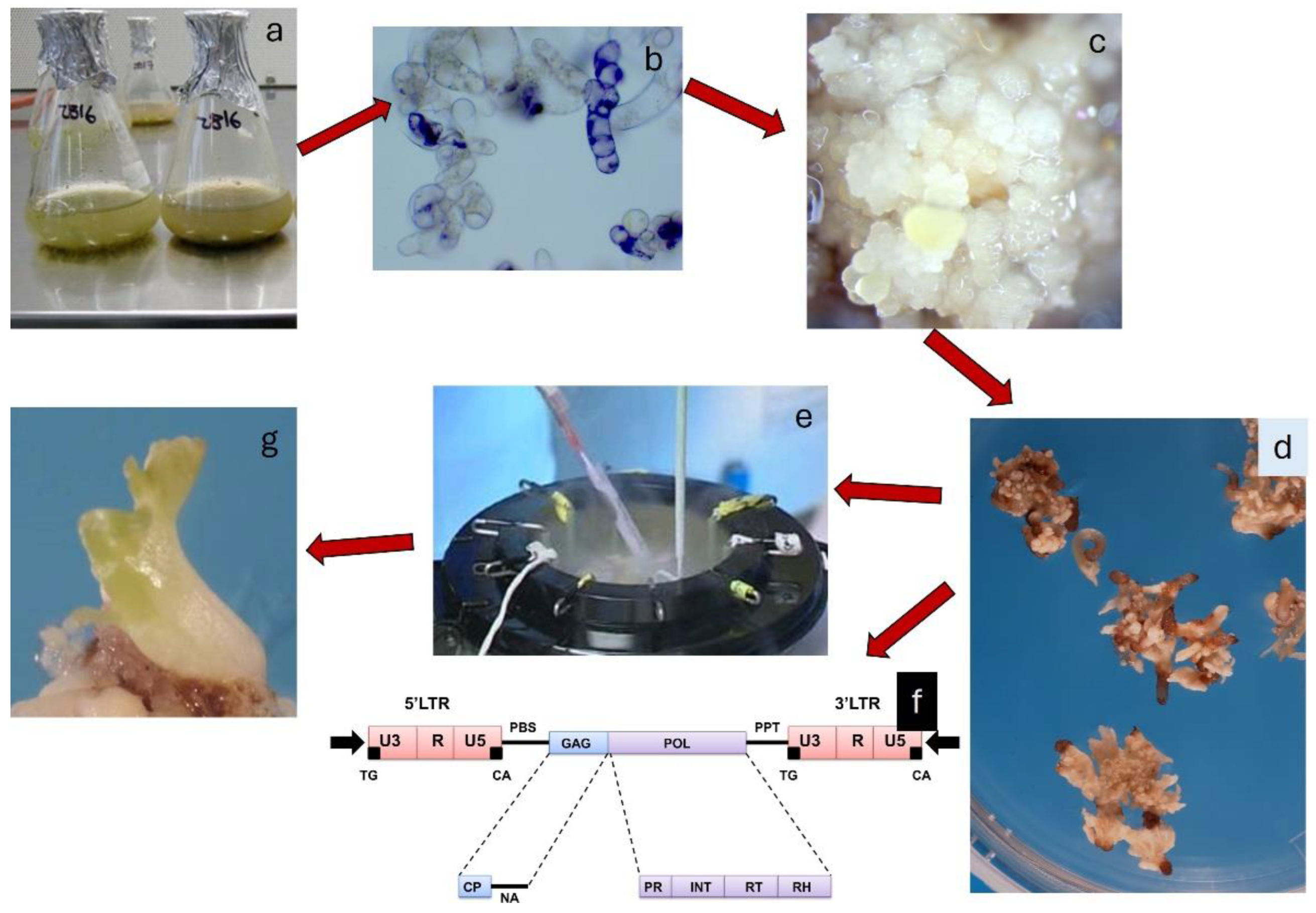
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



