
Submitted:
08 August 2024
Posted:
12 August 2024
You are already at the latest version
Abstract
Background; Cell-released extracellular vesicles (EVs) acting as 'metabolically and proteolytically active machines,' show potential in metabolomic and proteomic analysis of serum EVs. Despite diverse challenges, post-isolation omics characterization EVs offers crucial insights for effective analysis; (2) Methods: The research, involved children with Congenital Zika Syndrome, utilizing mass spectrometry for proteomics and GC-MS for metabolite identification. Vesicles were isolated using Izon qEV columns, quantified, and characterized by NTA and TEM. Data analysis employed Cytoescape/String and MetaboAnalyst, revealing variations in metabolic and proteomic profiles among groups through PCA and volcano plots. Proteins and Metabolite set enrichment analysis provided biologically meaningful patterns to enriched metabolites; (3) Results: Using molecular exclusion chromatography, the EVs were characterized, revealing size variations. Protein analysis identified 13 significantly altered proteins, including upregulated (e.g., AOM8Q6 - IGLC7) and downregulated (e.g., Q8TD86 - CALML6) ones. Metabolite analysis indicated involvement in the PI3K-AKT-mTOR pathway and suggested a role in Angiotensin inhibition in CZS+. Upstream of mTOR, Akt is the central signaling molecule in the PI3K pathway and plays critical roles in brain development as well as synaptic plasticity important for Zika Virus. The study provides insights into molecular mechanisms associated with CZS; (4) Conclusions: The study pinpointed valuable possible biomarkers, specifically proteins and metabolites, in Zika virus (ZIKV) infection. It stresses the necessity for broader investigations with advanced techniques to uncover molecular targets, potentially advancing pharmacological strategies.
Keywords:
Zika Infection
; Extracellular
; Vesicles
; Proteome
; Metabolome
; Network
1. Introduction
The infection of ZIKV (ZIKV) during gestation carries a risk of serious injury, including the loss of fetal tissue, microcephaly, and other congenital Zika-related brain malformations [1]. Additionally, severe neurological conditions like encephalopathies, meningoencephalitis, myelitis, uveitis, Guillain-Barré syndrome, and severe thrombocytopenia have been associated with ZIKV infection [2,3]. The causes of the severe forms of Zika are still unknown, as of this moment, there are no effective vaccines or specific treatments for the virus available. The increase in prevalence and range of ZIKV is attributed to the virulence of the circulating strains, the vulnerable populations, and the spread of its vectors.
The mosquitoes Aedes aegypti and Aedes albopictus are primarily responsible for transferring various Flaviviruses to humans, these viruses include Zika and dengue (DENV) [4,5]. The female mosquitoes become infected with the virus while consuming a host that is infected with it, this results in the virus's replication in the gut. Later, the virus was transferred to the salivary glands, where it was released into the saliva. The transmission to a new host is completed during the following feeding process [6]. Researchers described the documented increase in the viral infection of mammalian cells because of the exposure of these cells to the extract of mosquitoes's salivary glands. This was observed in the context of a DENV infection in humans. However, the specific mechanisms involved in the transmission of Flaviviruses from vector to host are still not fully understood [7].
EV are cell-secreted enveloped particles that are predominantly 50–200 nm in diameter and are released from the plasma membrane and endosomal compartments [8]. In human hosts, ZIKV primarily infects monocytes, macrophages, endothelial cells, and neurons. The process of cell differentiation or activation causes the release of EVs from the cell's exterior (also known as exosomes or microvesicles) [9]. Metabolites and proteins are both integral to biological importance and participate in various cellular processes [10,11]. The evaluation of metabolomes and proteomes becomes crucial, this is because it provides information about potential dysregulations, similar to the way EVs are used to investigate cellular behavior. EVs have the intriguing ability to function as metabolically and proteolytically active machines, as evidenced by the metabolomic analysis of serum samples in conjunction with hepatocytes' EVs [12]. While EVs have a bright future in metabolomics and proteomics, the field is still in the early stages of development, specifically about the diverse classes of EVs and their inherent diversity [13].
Mainly in the field of EV we note in the literature that PI3K inhibitors that target the stroma and focus on immune modulation, we refer the reader to Okkenhaug et al. 2016 [14] and Vanhaesebroeck et al. 2022 [14]. For more information on past and future PI3K inhibitors, we refer the reader to Castel et al. 2021 [14]. In addition, although our manuscript shows an overview of all upregulated proteins and metabolites, mostly all observed PI3K-driven precursors, for a specific perspective on the relevance of the PI3K pathway in ZIKV which have been extensively [15]. Furthermore, since this paper does not discuss in detail the role of the PI3K pathway in metabolism, we direct the reader to Vasan and Cantley 2022 [16] for this specific topic.
The characterization of omics following the isolation procedure via SEC has a significant impact on the scientific community as a whole and is of crucial importance to the research community specifically. As a result, we focus on the identity and intricate analysis we employed to facilitate additional advances in this field. Despite the inherent complexity, the metabolomic analysis of an isolated EV sample is a new approach. Employing established methods of metabolite/ protein extraction and analysis has been successful in producing large amounts of data and addressing these issues.
2. Materials and Methods
2.1. Ethical Aspects
The research adhered to the criteria outlined in the Helsinki conference. Before commencement, it underwent scrutiny and gained approval from the Research Ethics Committee at the Federal University of Maranhão Hospital and Medicine School of Jundiaí (CAAE: 86696618.7.0000.5467). All actions followed the acknowledgment and endorsement expressed through the informed consent form.
2.2. Study Design
The research involved a prospective cohort of children residing in the State of Maranhão, located in northeastern Brazil. It was a collaborative effort with the Reference Center for Neurodevelopment, Assistance, and Rehabilitation of Children (NINAR). Additional data for control children in this study were primarily collected at the Infectious Pediatric Laboratory of the Medicine School of Jundiaí in Jundiaí. A blood sample (5 mL) was obtained from each child. Microcephaly, in this study, is defined as a head-circumference measurement of less than two standard deviations (SDs) below the average [17,18]. The study included a total of 14 children with confirmed Congenital Zika Syndrome (CZS+) and 15 children with confirmed Congenital Zika Syndrome (CZS-). Serum from were fractionated by Izon qEV size-exclusion SEC resulting in five fractions of 500 µL. The F3 fractions from 14 children with confirmed Congenital Zika Syndrome (CZS+) and 15 children with confirmed Congenital Zika Syndrome (CZS-) were analyzed by mass spectrometry techniques and metabolomic analysis. We subsequently carried out the characterization of isolated EV fractions by nanoparticle tracking analysis (NTA) and transmission electron microscopy (TEM) was used. The protocols for isolation, characterization and spectrometry analyzes adhered to the methodology outlined by Pontes, et. al, 2020 [19]
2.3. Vesicle Isolation and Purification
To isolate vesicles, 500 μL of serum samples were placed onto Izon qEV size-exclusion chromatography columns that were pre-conditioned with PBS buffer (Izon Science, UK) [20,21]. The elution process was carried out using PBS buffer as per the manufacturer's guidelines. The columns were filled with a resin featuring a pore size of around 75 nm, a bed volume of 10 mL, an inner tube diameter of 15.6 mm, and a void volume of 3.0 ± 0.25 mL. Before use, the columns were pre-loaded with PBS containing 0.05% sodium azide. Following the exclusion of the column's void volume (3 mL), three fractions were collected from each sample, each consisting of 500 μL.
2.4. EV Sample Preparation for MS-Based Proteomics Analysis
EV (F3) suspended in phosphate-buffered saline (PBS) containing protease inhibitors (obtained from Sigma Aldrich, Brazil) and stored at −80°C for six months were reconstituted in a lysis buffer (comprising 7 M urea, 2 M thiourea, 10 mM dithiothreitol (DTT), and 0.01% Triton X-100) at room temperature for a duration of 2 hours. Subsequently, the lysis buffer was eliminated using Amicon® Ultra 0.5 3KD (Millipore), and protein quantification was confirmed using NanoDrop One (Thermo Scientific, United States). To assess protein banding patterns, a 10% SDS-Page gel analysis was conducted with 10 µg of bovine serum albumin (BSA) serving as the standard protein sample. The gel electrophoresis ran at 50 v for 4.5 hours. Following the run, the gel was detached from the glass plates and immersed in a fixative solution (composed of 50% methanol and 7% acetic acid) for one hour. Subsequently, it underwent a washing step with Milli-Q water and was left overnight in GelCode Blue stain.
For the EV samples (10 µg), incubation was carried out with RapiGest™ SF Surfactant (Waters, United States) for 15 minutes at 80°C, and 100 mM dithiothreitol (DTT) for 30 minutes. Alkylation followed, utilizing 300 mM iodoacetamide (IAA) for 30 minutes in the dark. The samples then underwent digestion with trypsin (at a ratio of 1:100 w/w trypsin: protein) overnight at room temperature. Finally, the peptides were acidified using 1% v/v formic acid to halt the trypsin digestion process and lyophilized before desalting. Resuspension of the samples took place in 0.1% v/v trifluoroacetic acid (TFA), and desalting was achieved using ZipTipC18 Pipette Tip with C18 Resin (Sigma Aldrich, Brazil) following the provided manufacturer’s instructions.
The processing parameters encompassed specific modifications, such as carbamidomethylation of cysteine, treated as a fixed amino acid modification. Variable modifications included methionine oxidation and N-terminal acetylation. Trypsin served as the proteolytic enzyme, allowing for a maximum of 2 potential cleavage errors. Peptide and fragment ion mass shift tolerances were set at 20 ppm and 0.05 Da, respectively. Peptide and protein identification adhered to a maximum false positive rate (FDR) of 1%, requiring at least one single peptide for protein identification. Adapted from the protocol Pontes, et. al, 2020 [19]
Bioinformatic analysis relied on information from UniProt and Cytoscape and String (https://cytoscape.org/ and https://string-db.org/). The mass spectrometry proteomics data (Table S1- raw proteomics data).
2.5. EV Identification of Metabolites with GC-MS
EV (F3) suspended in phosphate-buffered saline (PBS) of metabolites was carried out in accordance with the methodology outlined by Hoffman et al., incorporating minor adjustments [22]. Subsequently, extraction took place in microtubes, utilizing 200 mg of fungal macerate to which 1 mL of an ice-cold extraction solution comprising methanol:chloroform:water in a ratio of 6:2:2 was added. After rigorous vortexing of these extraction microtubes, they were subjected to ultrasonic low-temperature bath treatment at 20 Hz s-1 for 15 minutes. The ensuing step involved centrifugation (Eppendorf, Germany) at 4°C for 10 minutes at 14,000 rpm. The resulting supernatant underwent filtration using a 0.22 μm Whatman® filter (Merck, Germany) and was then transferred to a chromatographic vial for lyophilization (Thermo Fischer Scientific, MA, USA) until complete desiccation. The lyophilized samples were subsequently reconstituted in 200 μL of the extraction solution and portioned for utilization in GC-MS and LC-MS/MS analyses. Each extract aliquot received 10 μL of a 1 mg. mL-1 solution of the isotopically labeled compounds succinic acid (D4, 98%—DLM 584–5), myristic acid (1, 2, 3–13C3, 99%—CLM 3665– 0.5), and palmitic acid (1, 2, 3, 4 – 13C4) as external standards. The samples were once again lyophilized for subsequent derivatization using 30 μL of a 15 mg·mL-1 solution of methoxyamine and pyridine for 16 hours at room temperature. The silylation of the samples occurred promptly at room temperature for 1 hour by introducing 30 μL of MSTFA(N-methyl-trimethylsilyl-trifluoroacetamide) with 1% TMCS (trimethylchlorosilane). Finally, 30 μL of heptane containing 15 ng. g-1 of methylesterase was introduced.
Data derived from GC-MS underwent processing using ChromaTOF 4.32 software, involving baseline correction, deconvolution, retention index (RI), retention time correction (RT), identification, and alignment of peaks. Identification of metabolites employed the NIST library version 11, with only metabolites scoring 700 or above being considered. The intensity of each metabolite was normalized by the total ion count (TIC) of each sample. The analysis of samples using GC-MS adhered to the methodology outlined by Budzinski et al. [23], with minor modifications. At this stage, a series of n-alkanes (C12 –C40) was utilized to calculate the sample retention index [24]. One microliter of each derivatized sample was automatically injected in splitless mode by a CTC Combi Pal Xt Duo autosampler (CTC Analytics AG, Switzerland) into an Agilent 7890A gas chromatographer. The chromatographer featured two fused-silica capillary columns; one column with a 20 m x 0.18 mm chemically bonded with 0.18 μm DB-5 film (Agilent) stationary phase, and the other column with 0.9 m x 0.10 mm chemically bonded with 0.10 μm RX-T 17 film (Restek) stationary phase. The injection temperature was 280°C, with a purge flow of 20 mL min-1. The gas flow through the column was 1 mL min-1, and the column temperature was maintained at 70°C for 2 min, followed by an increase of 15°C min-1 until reaching 320°C, and then held at this temperature for 4 min. The column effluent was introduced into the GC x GC/TOF-MS Pegasus 4D ion source (Leco Corporation, St. Joseph, MI, USA) at a temperature of 250°C. Ions were generated by a 70 eV electron beam at an ionization current of 20 mA and 20 spectra·s-1, recorded in the range of 45–800 m/z. The detector voltage was set at 1500 V.
Statistical analyses were executed using the MetaboAnalyst 4.0 online software (accessible at http://www.metaboanalyst.ca/MetaboAnalyst/) [25]. A log2 transformation and Pareto scaling were implemented for data normalization. Analysis of variance (ANOVA) and principal component analysis (PCA) were applied to identify variations in the metabolic profiles among the different groups. Metabolite set enrichment analysis (MSEA) was conducted to explore and attribute biologically meaningful patterns to specific groups of significantly enriched metabolites.
3. Results
3.1. EV Characterization
To isolate EVs, the collected sera underwent fractionation using a molecular exclusion chromatography column, and the resulting isolates were preserved. Subsequently, quantification (Figure 1 - A), nanoparticle tracking (Figure 1 - B), and transmission electron microscopy analysis (Figure 1 - C) were performed on the isolates. The sizes of the isolated extra-cellular vesicles ranged from 154.9 nm for the control child pool (CZS-) to 185.4 nm for the affected child pool (CZS+) [26,27,28,29]
3.2. Proteomic Analysis
The differentially expressed proteins observed by mass spectrometry were compared to their obtained Fold change values (Figure 2) and interaction network (Figure 3). While, in the String software we were able to characterize the functionality and through its plugin for Cytoscape we were able to obtain a network for positive and negative proteins, compared to the control.
3.1. Proteomics and Metabolomics
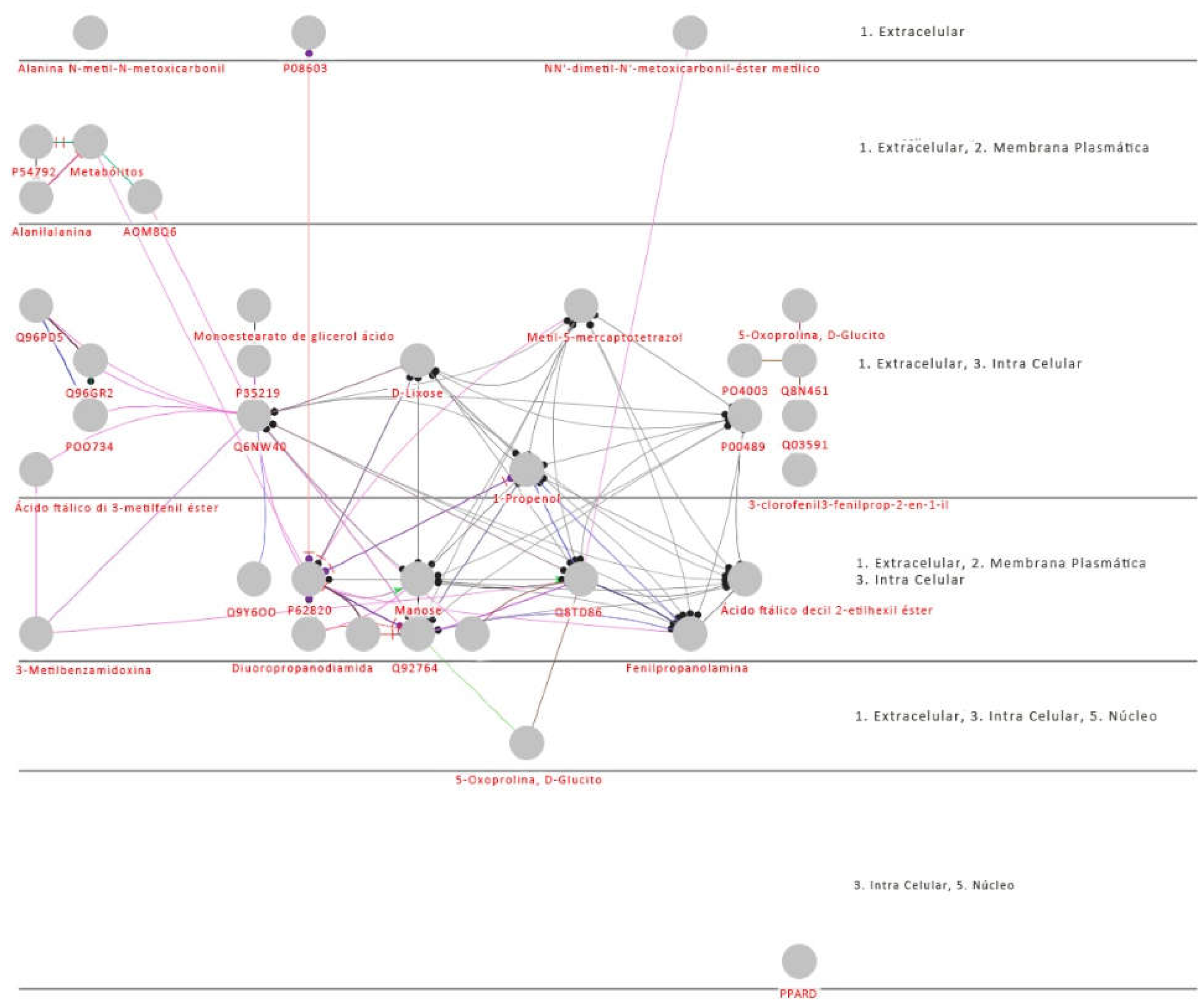
The Principal Component Analysis (PCA) data obtained from protein analyses (Figure 4 - A) reveal elevated values exclusively for the maximum variance. Conversely, when considering successive variances, the highest values are evident in the PCA derived from metabolite analyses (Figure 4 - B), utilizing the same samples of EVs obtained from both CZS- and CZS+ children. In our research, we identified 13 proteins (depicted as red circles) that exhibited at least a twofold alteration and statistical significance (p ≤ 0.05) in extracellular vesicle (EV) samples isolated from the serum of children with congenital Zika syndrome (CZS+) (Figure 5 - A). Notably, our investigation revealed an association between the up-regulation of extracellular vesicular proteins (AOM8Q6 - IGLC7, P35219 - CA8, Q6NW40 - RGMB, Q9Y6OO - CSAD, P62820 - RAB1A, and Q92764 - KRT35) and the down-regulation of others (Q8TD86 - CALML6, POO734, Q96GR2 - ACSBG1, Q8N461 - FBXL16), suggesting a crucial role of the complement cascade in children affected by CZS (Figure 5 - B) [30,31,32]. In light of this, we can highlight the presence of upregulated EVs metabolites, predominantly involved in the PI3K-AKT-mTOR pathway. These include Alanylalanine, NN'-dimethyl-N'-methoxycarbonyl-methyl ester, Phthalic acid di 3-methylphenyl ester, Phthalic acid decyl 2-ethylhexyl ester, D-Lyxose, Diuoropropanodiamide, and Manose, among others. Conversely, there are downregulated metabolites such as Phenylpropanolamine, Methyl-5-mercaptotetrazole, Glycerol monostearate succinic acid ester 3-chlorophenyl 3-phenylprop-2-en-1-yl, 5-Oxoproline, D-Glucitol, Phthalic acid decyl 2-ethylhexyl ester, 3-Methylbenzamidoxine, 1-Propenol, and Alanine N-methyl-N-methoxycarbonyl hexyl ester. These findings suggest a significant role in the inhibition of Angiotensin in CZS+ children. Studies on viral infections such as DENV and H5N1 have explored the inhibition of ACE [33,34,35,36] and kinases involved in the PI3K-AKT-mTOR pathway [37,38,39]. Literature data suggests that cellular locations may determine functions as follows [40,41,42]: 1) Extracellular - Cell shrinkage causes the cytosol to push vesicles towards the perinuclear region with "addressing labels" for swift transport; 2) Plasma Membrane – membrane ligand specificity and recognition of PAMPs; 3) Intracellular - leading to the formation of multivesicular bodies and actively participating in intracellular signaling pathways; and 4) Nucleus – intracellular signaling and modulation of the immune response (Figure 6).
4. Discussion
The cellular communication of EVs influences various physiological and pathological processes, including immune responses [43,44], cancer [45,46,47], and cardiovascular diseases [47]. Additionally, it plays a role in delivering bacterial virulence factors to the host [48,49,50] and modulating inflammatory and immune responses in fetal-maternal communication [51,52]. This communication occurs among different cell types, but when it involves the participation of immune system cells or molecules secreted by them, it gains greater prominence in the literature due to its immunomodulatory capacity. The discovery that EVs act as vehicles for the transfer of membrane, cytosolic proteins, lipids, and proteins between cells strongly suggests their contribution as communicators in cellular communication. When secreted by immune system cells, they exhibit immunomodulatory functions, inducing both suppression and activation of immune responses [53,54].
We have the PROS1 protein (positive regulation), which is described in the literature as vitamin K-dependent, serving as a cofactor for CRP-mediated cleavage (negative regulation) of factors Va and VIIIa on the surface of phospholipids in the presence of Ca²+ [55,56,57]. Additionally, the serpins SERPINC1, SERPING1, SERPINF2 (all with positive regulation), in turn, inhibit and block the activity of thrombin, fIXa, fXa, fXIIa, kallikrein, and plasmin [58]. Thus, compromising the activity of platelet procoagulant.
In summary, HPX (positive regulation) is produced in the brain [59,60,61] and has a higher affinity for binding to the heme group, potentially reducing the regulation of Angiotensin II Type 1 receptor [62]. The heme prosthetic group imparts a characteristic color to these proteins, consisting of an organic part and a ferrous iron atom in the ferrous state [63,64]. Its function is to capture proteins with the heme group released or lost during hemoglobin recycling, thereby protecting the body from oxidative damage that heme groups could induce [65]. The circulating level of HPX is associated with the prognosis in patients with septic shock [66,67,68].
The analyses of protein-protein interaction in EVs, as revealed by the String software, provided us with two network maps (Figure 3) and insights into the functionality of proteins obtained through mass spectrometry. In proteins exhibiting positive regulation, processes such as leukocyte migration, cytokine secretion, acute inflammatory response, and platelet degradation were observed [69]. On the other hand, proteins identified by mass spectrometry with negative regulation were associated with processes including regulation of protein activation cascade, reverse cholesterol transport, phagocytosis, negative regulation of lipid localization, B cell receptor signaling pathway, and adaptive immune response. Another crucial aspect in the functional evaluation of EVs is to consider their ability to activate the adaptive immune response [70].
Interestingly, similar findings have been reported previously in patients with an increased production of interferon alpha (IFN-α) by plasmacytoid dendritic cells [71]. This was observed through their interaction with EV originating from a hepatocarcinoma cell line infected with the hepatitis C virus (HCV) [72,73]. This result underscores a defense strategy of the organism, activating the innate immune response of the host [74,75]. Furthermore, the proteins with the codes Q03591 (CFHR1 abbreviation), P00489 (PYGM abbreviation), and PO4003 (C4BPA abbreviation) originating from the serum isolate of EV-affected children were also identified as positively regulated in HIV-1 [76] and SARS-CoV-2 [77,78] infections. EV derived from infected cells can transport viral regulatory proteins without being recognized by the immune system due to the lipid bilayer coating from host cells, functioning as a Trojan horse [79,80,81,82].
However, there are additional proteins identified in this study as potential biomarkers for serum EVs in children affected by SCZ that have already been described in the literature, exhibiting new functionalities such as immunomodulation and increased infectivity. Some of them include protein code P54792 (abbreviated as DVL-1), described with positive regulation in cellular susceptibility to HIV infection [83,84]; protein code P82970 (abbreviated as HMGN5), known for increasing apoptosis in CD4+ T lymphocytes, which may contribute to the depletion of these cells in the pathogenesis of Acquired Immunodeficiency Syndrome [85,86,87,88].
The protein with the code P08603 (CFH) enhances cellular lysis by B cell. The surface of viruses often has dense, repetitive structures that promote multivalent interactions with B-cell receptors and influence the functional characteristics of antibodies [89,90,91]. Early in pregnancy, B cells are stimulated to produce IL-10, which promotes the survival of mature B cells and reduces the frequency of naive B cells. This study examined the contribution of maternal plasma IgM to ZIKV neutralization, and the results suggest that IgM is effective within the first 3 months of infection [92].
On the other hand, this intravesicular protein has also been described as increased in the survival of the cytotoxic T cell lineage dependent on IL-2 [93,94,95]. Although, the extravesicular protein with the code Q96PD5 (abbreviated as PGLYRP2) plays an active role in stimulating the production of pro-inflammatory cytokines by cells through the activation of the NFkB pathway [96,97]. The NFkB pathway stands out in EVs due to its wide range of actions and the fact that various extracellular proteins are integrated into the dynamics of its activation [98,99]. In the analysis of metabolites, we observed six metabolites (red circles) with at least a two-fold alteration and a significant value (p ≤ 0.05) in samples of EVs isolated from the serum of CZS+ children (Figure 5 - B).
Furthermore, the most promising metabolites as potential biomarkers are Manose and Decyl 2-ethylhexyl phthalate acid ester, which are part of the activation of the Renin-Angiotensin System pathway. This pathway is directly involved in the uptake and excretion of sodium and potassium, possessing ions that promote vasoconstriction and regulation of blood pressure, respectively [100]. However, their importance in the viral infection process has only recently been observed, as demonstrated by a study in which rats infected with DENV were treated with losartan or enalapril. In this case, the treatment reduced DENV absorption by macrophages, indicating that the activation pathways of the RAS may be associated with the severity of the infection [101,102]. Therefore, the potential biomarkers elucidated in our study reveal that these species may also be linked to the control of the immune response to ZIKV infection.
The other four biomarkers found in serum-derived EVs from CZS+ children (Alanilalanine, NN'-dimethyl-N'-methoxycarbonylmethyl ester, Phthalic acid di 3-methylphenyl ester, D-Lyxose, and Diuoropropanodiamide) contribute to supporting the role of the immune response to infection [103]. These metabolites, when combined, are responsible for activating the PI3K-AKT-mTOR pathway [104,105]. Akt is upstream of mTOR and is fundamental in the PI3K pathway, playing important roles in brain development and synaptic plasticity. Non-functional Akt mutations cause microcephaly, while activating mutations cause megalencephaly. Expression of dominant negative Akt blocks neurogenesis in human foetuses in vitro. Neurogenesis, the process of forming neurons from neural stem cells or neural progenitor cells, is most active during prenatal development and is critical for brain growth. Genetic defects in neurogenesis and cell migration can lead to neurological disorders such as microcephaly [106]. Cell signalling pathways, especially the PI3K-Akt-mTOR pathway, are critical for neurogenesis, migration and neuronal maturation. Activating mutations in this pathway can lead to brain overdevelopment syndrome, while inhibition of this pathway results in microcephaly. Akt is a key molecule in the PI3K pathway, essential for brain development and synaptic plasticity. Mutations in Akt can cause microcephaly or megalencephaly [107]. The PI3K-Akt-mTOR pathway can be hijacked by human pathogens for replication, but a causal relationship between ZIKV infection and inhibition of this pathway has not been established to date [107,108]. However, despite the common outcomes of ZIKV infection and inhibition of the PI3K-Akt-mTOR pathway, no causal association between the two has been reported yet.
To conclude this data analysis, we will highlight the localization of both proteins and metabolites. Considering that the vesicle's location can shed light on its classification and transport function, we will perform this identification to distinguish between structural and non-structural proteins (Figure 6 ). While both the activation of SRA by metabolites and the activation and regulation of the complement by proteins contribute to higher concentrations of biomarkers, an additional factor appears to decrease selected biomarkers in the process of ZIKV infection by vesicles [109,110,111] (Melo et al., 2017).
After analyzing their location, we observed the presence of two non-structural proteins, CSAD and KRT35, which inhibit the AKT-mTOR signaling pathway [112,113,114]. This inhibition leads to the accumulation of intermediate metabolites and precursors involved in the PI3K-AKT-mTOR pathway signaling. As the virus inhibits the SRA signaling, other metabolites and proteins persist due to viral infection, maintaining positive modulation over PI3K [115,116,117]. Therefore, ZIKV infection induced alterations in different signaling pathways, resulting in the overexpression of certain metabolites. Our findings suggest that it is possible to study the mechanism of viral infection by directly analyzing serum EV from infected children.
5. Conclusions
Akt-mTOR signaling is essential for neurogenesis by human NSCs as well as the induction of autophagy. Specifically, phosphorylation of Akt on mTOR is required for its full kinase activity, and subsequently, Akt-mediated phosphorylation of mTOR on PI3K is essential for keeping autophagy in check. Our results show that ZIKV replication led to suppression of Akt phosphorylation, which subsequently led to reduced mTOR phosphorylation. Further screening of key proteins and metabolites in EV in individual ZIKV viruses revealed that PI3K/AKT/mTOR signaling pathway expression detectably reduced Akt phosphorylation under normal conditions. All possible biomarkers were carefully chosen and validated through rigorous statistical analysis, aligning with prior research that focused on proteins and metabolites implicated in the infectious process. Nevertheless, it is imperative to conduct further investigations involving a larger cohort of children, employing advanced sample extraction technologies and specific markers. This will enable a comprehensive understanding of the overexpression of specific metabolites. For instance, the introduction of quantitative NMR techniques has enhanced precision in quantifying minor components within a mixture. Thus, the proteomic and metabolomic knowledge regarding the EVs in the same samples, viewed uniquely in the context of human ZIKV infection, significantly contributes to our understanding of the disease's physiopathological aspects. It sheds light on molecular targets of the cellular immune response to infection and viral replication. This not only forms the basis for new developments in pharmacology for diverse therapies but also provides insights into the management of ZIKV infection.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The datasets generated by this study will be available upon reasonable request to the authors.
Author Contributions
Author Contributions: For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used Conceptualization, L.G.P. and A.C.N; methodology, L.A.B. and P.V.S.P; software, L.N.G and R.C.S; validation, R.C.S, S.M.S.N, L.N.G, and L.G.P; formal analysis, L.A.B. and R.C.S; investigation, S.M.S.N and S.D.P; resources, L.A.B and L.G.P ; data curation, R.C.S, S.M.S.N, L.N.G, A.C.N and L.G.P ; writing—original draft preparation, L.G.P and A.C.N; writing—review and editing, L.G.P., S.D.P, P.V.S.P, L.N.G, A.C.N and SDP; visualization, L.N.G, P.V.S.P, R.C.S, L.A.B. and L.G.P; supervision, L.G.P, A.C.N and SDP; project administration, A.C.N and SDP; funding acquisition, A.C.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Antonio Condino Neto, grant number 2016/22158-3 - Fundação de Amparo à Pesquisa do Estado de São Paulo, and Leticia Gomes de Pontes, grant number 2019/27294-0 - Fundação de Amparo à Pesquisa do Estado de São Paulo.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Ethics Committee of Universidade de São Paulo (protocol code CAAE: 86696618.7.0000.5467 and date of approval 02/14/2019).
Data Availability Statement
The datasets generated by this study will be available upon reasonable request to the authors.
Acknowledgments
We thank colleagues from the Department of Immunology, Institute of Biomedical Sciences, University of São Paulo, São Paulo, Brazil. We thank Core Facility in proteomics and metabolomics, Escola Superior de Agronomia Luiz de Queiroz – Esalq/USP, Piracicaba, São Paulo, Department of Pathology - Federal University of Maranhão and Infectious Pediatric Laboratory - Medicine School of Jundiaí for the use of its facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dick, G.W.A.; Kitchen, S.F.; Haddow, A.J. Zika Virus (I). Isolations and Serological Specificity. Trans R Soc Trop Med Hyg 1952, 46, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Oehler, E.; Watrin, L.; Larre, P.; Leparc-Goffart, I.; Lastère, S.; Valour, F.; Baudouin, L.; Mallet, H.P.; Musso, D.; Ghawche, F. Zika Virus Infection Complicated by Guillain-Barré Syndrome – Case Report, French Polynesia, December 2013. Eurosurveillance 2014, 19. [Google Scholar] [CrossRef] [PubMed]
- Rozé, B.; Najioullah, F.; Fergé, J.-L.; Apetse, K.; Brouste, Y.; Cesaire, R.; Fagour, C.; Fagour, L.; Hochedez, P.; Jeannin, S.; et al. Zika Virus Detection in Urine from Patients with Guillain-Barré Syndrome on Martinique, January 2016. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Harris, E. Dengue. The Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.N.; Marten, A.D.; Moore, G.A.; Tree, M.O.; McBrayer, S.P.; Conway, M.J. Extracellular Vesicles Restrict Dengue Virus Fusion in Aedes Aegypti Cells. Virology 2020, 541, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Calvet, G.; Aguiar, R.S.; Melo, A.S.O.; Sampaio, S.A.; de Filippis, I.; Fabri, A.; Araujo, E.S.M.; de Sequeira, P.C.; de Mendonça, M.C.L.; de Oliveira, L.; et al. Detection and Sequencing of Zika Virus from Amniotic Fluid of Fetuses with Microcephaly in Brazil: A Case Study. Lancet Infect Dis 2016, 16, 653–660. [Google Scholar] [CrossRef]
- Lee, M.F.; Voon, G.Z.; Lim, H.X.; Chua, M.L.; Poh, C.L. Innate and Adaptive Immune Evasion by Dengue Virus. Front Cell Infect Microbiol 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science (1979) 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Conzelmann, C.; Groß, R.; Zou, M.; Krüger, F.; Görgens, A.; Gustafsson, M.O.; El Andaloussi, S.; Münch, J.; Müller, J.A. Salivary Extracellular Vesicles Inhibit Zika Virus but Not SARS-CoV-2 Infection. J Extracell Vesicles 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rojas, P.P.; Quiroz-García, E.; Monroy-Martínez, V.; Agredano-Moreno, L.T.; Jiménez-García, L.F.; Ruiz-Ordaz, B.H. Participation of Extracellular Vesicles from Zika-Virus-Infected Mosquito Cells in the Modification of Naïve Cells’ Behavior by Mediating Cell-to-Cell Transmission of Viral Elements. Cells 2020, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.; Palviainen, M.; Reichardt, N.-C.; Siljander, P.R.-M.; Falcón-Pérez, J.M. Metabolomics Applied to the Study of Extracellular Vesicles. Metabolites 2019, 9, 276. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, W.; Guo, M.; Tan, Q.; Zhou, E.; Deng, J.; Li, M.; Chen, J.; Yang, Z.; Jin, Y. Metabolomics of Extracellular Vesicles: A Future Promise of Multiple Clinical Applications. Int J Nanomedicine 2022, Volume 17, 6113–6129. [Google Scholar] [CrossRef]
- Alseekh, S.; Aharoni, A.; Brotman, Y.; Contrepois, K.; D’Auria, J.; Ewald, J.; C. Ewald, J.; Fraser, P.D.; Giavalisco, P.; Hall, R.D.; et al. Mass Spectrometry-Based Metabolomics: A Guide for Annotation, Quantification and Best Reporting Practices. Nat Methods 2021, 18, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Okkenhaug, K.; Graupera, M.; Vanhaesebroeck, B. Targeting PI3K in Cancer: Impact on Tumor Cells, Their Protective Stroma, Angiogenesis, and Immunotherapy. Cancer Discov 2016, 6, 1090–1105. [Google Scholar] [CrossRef] [PubMed]
- Castel, P.; Toska, E.; Engelman, J.A.; Scaltriti, M. The Present and Future of PI3K Inhibitors for Cancer Therapy. Nat Cancer 2021, 2, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Vasan, N.; Cantley, L.C. At a Crossroads: How to Translate the Roles of PI3K in Oncogenic and Metabolic Signalling into Improvements in Cancer Therapy. Nat Rev Clin Oncol 2022, 19, 471–485. [Google Scholar] [CrossRef] [PubMed]
- França, G.V.A.; Schuler-Faccini, L.; Oliveira, W.K.; Henriques, C.M.P.; Carmo, E.H.; Pedi, V.D.; Nunes, M.L.; Castro, M.C.; Serruya, S.; Silveira, M.F.; et al. Congenital Zika Virus Syndrome in Brazil: A Case Series of the First 1501 Livebirths with Complete Investigation. The Lancet 2016, 388, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Moura da Silva, A.A.; Ganz, J.S.S.; Sousa, P. da S.; Doriqui, M.J.R.; Ribeiro, M.R.C.; Branco, M. dos R.F.C.; Queiroz, R.C. de S.; Pacheco, M. de J.T.; Vieira da Costa, F.R.; Silva, F. de S.; et al. Early Growth and Neurologic Outcomes of Infants with Probable Congenital Zika Virus Syndrome. Emerg Infect Dis 2016, 22, 1953–1956. [Google Scholar] [CrossRef] [PubMed]
- Pontes, L.G. de; Altei, W.F.; Galan, A.; Bilić, P.; Guillemin, N.; Kuleš, J.; Horvatić, A.; Ribeiro, L.N. de M.; Paula, E. de; Pereira, V.B.R.; et al. Extracellular Vesicles in Infectious Diseases Caused by Protozoan Parasites in Buffaloes. Journal of Venomous Animals and Toxins including Tropical Diseases 2020, 26. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.G.; Choi, Y.Y.; Mo, S.J.; Kim, T.H.; Ha, J.H.; Hong, D.K.; Lee, H.; Park, S.D.; Shim, J.-J.; Lee, J.-L.; et al. Effect of Gut Microbiome-Derived Metabolites and Extracellular Vesicles on Hepatocyte Functions in a Gut-Liver Axis Chip. Nano Converg 2023, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhai, J.; Ma, J.; Chen, P.; Lin, W.; Zhang, W.; Xiong, J.; Zhang, C.; Wei, H. Melatonin-Primed Mesenchymal Stem Cells-Derived Small Extracellular Vesicles Alleviated Neurogenic Erectile Dysfunction by Reversing Phenotypic Modulation. Adv Healthc Mater 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- HOFFMAN, D.E.; JONSSON, P.; BYLESJÖ, M.; TRYGG, J.; ANTTI, H.; ERIKSSON, M.E.; MORITZ, T. Changes in Diurnal Patterns within the Populus Transcriptome and Metabolome in Response to Photoperiod Variation. Plant Cell Environ 2010, 33, 1298–1313. [Google Scholar] [CrossRef] [PubMed]
- Budzinski, I.G.F.; Moon, D.H.; Morosini, J.S.; Lindén, P.; Bragatto, J.; Moritz, T.; Labate, C.A. Integrated Analysis of Gene Expression from Carbon Metabolism, Proteome and Metabolome, Reveals Altered Primary Metabolism in Eucalyptus Grandis Bark, in Response to Seasonal Variation. BMC Plant Biol 2016, 16, 149. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, P.; Johansson, A.I.; Gullberg, J.; Trygg, J.; A, J.; Grung, B.; Marklund, S.; Sjöström, M.; Antti, H.; Moritz, T. High-Throughput Data Analysis for Detecting and Identifying Differences between Samples in GC/MS-Based Metabolomic Analyses. Anal Chem 2005, 77, 5635–5642. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr Protoc Bioinformatics 2019, 68. [Google Scholar] [CrossRef] [PubMed]
- Jeannin, P.; Chaze, T.; Giai Gianetto, Q.; Matondo, M.; Gout, O.; Gessain, A.; Afonso, P. V. Proteomic Analysis of Plasma Extracellular Vesicles Reveals Mitochondrial Stress upon HTLV-1 Infection. Sci Rep 2018, 8, 5170. [Google Scholar] [CrossRef] [PubMed]
- Guha, D.; Lorenz, D.R.; Misra, V.; Chettimada, S.; Morgello, S.; Gabuzda, D. Proteomic Analysis of Cerebrospinal Fluid Extracellular Vesicles Reveals Synaptic Injury, Inflammation, and Stress Response Markers in HIV Patients with Cognitive Impairment. J Neuroinflammation 2019, 16, 254. [Google Scholar] [CrossRef] [PubMed]
- Urbanelli, L.; Buratta, S.; Tancini, B.; Sagini, K.; Delo, F.; Porcellati, S.; Emiliani, C. The Role of Extracellular Vesicles in Viral Infection and Transmission. Vaccines (Basel) 2019, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Simeone, P.; Bologna, G.; Lanuti, P.; Pierdomenico, L.; Guagnano, M.T.; Pieragostino, D.; Del Boccio, P.; Vergara, D.; Marchisio, M.; Miscia, S.; et al. Extracellular Vesicles as Signaling Mediators and Disease Biomarkers across Biological Barriers. Int J Mol Sci 2020, 21, 2514. [Google Scholar] [CrossRef] [PubMed]
- Askeland, A.; Borup, A.; Østergaard, O.; Olsen, J. V.; Lund, S.M.; Christiansen, G.; Kristensen, S.R.; Heegaard, N.H.H.; Pedersen, S. Mass-Spectrometry Based Proteome Comparison of Extracellular Vesicle Isolation Methods: Comparison of ME-Kit, Size-Exclusion Chromatography, and High-Speed Centrifugation. Biomedicines 2020, 8, 246. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Rojas, P.P.; Quiroz-García, E.; Monroy-Martínez, V.; Agredano-Moreno, L.T.; Jiménez-García, L.F.; Ruiz-Ordaz, B.H. Participation of Extracellular Vesicles from Zika-Virus-Infected Mosquito Cells in the Modification of Naïve Cells’ Behavior by Mediating Cell-to-Cell Transmission of Viral Elements. Cells 2020, 9, 123. [Google Scholar] [CrossRef] [PubMed]
- Martínez, L.E.; Garcia, G.; Contreras, D.; Gong, D.; Sun, R.; Arumugaswami, V. Zika Virus Mucosal Infection Provides Protective Immunity. J Virol 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Hallal, S.; Tűzesi, Á.; Grau, G.E.; Buckland, M.E.; Alexander, K.L. Understanding the Extracellular Vesicle Surface for Clinical Molecular Biology. J Extracell Vesicles 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Benedikter, B.J.; Bouwman, F.G.; Vajen, T.; Heinzmann, A.C.A.; Grauls, G.; Mariman, E.C.; Wouters, E.F.M.; Savelkoul, P.H.; Lopez-Iglesias, C.; Koenen, R.R.; et al. Ultrafiltration Combined with Size Exclusion Chromatography Efficiently Isolates Extracellular Vesicles from Cell Culture Media for Compositional and Functional Studies. Sci Rep 2017, 7, 15297. [Google Scholar] [CrossRef] [PubMed]
- Fialho, E.M.S.; Veras, E.M.; Jesus, C.M. de; Gomes, L.N.; Khouri, R.; Sousa, P.S.; Ribeiro, M.R.C.; Batista, R.F.L.; Costa, L.C.; Nascimento, F.R.F.; et al. Maternal Th17 Profile after Zika Virus Infection Is Involved in Congenital Zika Syndrome Development in Children. Viruses 2023, 15, 1320. [Google Scholar] [CrossRef] [PubMed]
- Oosthuizen, D.; Sturrock, E.D. Exploring the Impact of ACE Inhibition in Immunity and Disease. Journal of the Renin-Angiotensin-Aldosterone System 2022, 2022. [Google Scholar] [CrossRef] [PubMed]
- Glaviano, A.; Foo, A.S.C.; Lam, H.Y.; Yap, K.C.H.; Jacot, W.; Jones, R.H.; Eng, H.; Nair, M.G.; Makvandi, P.; Geoerger, B.; et al. PI3K/AKT/MTOR Signaling Transduction Pathway and Targeted Therapies in Cancer. Mol Cancer 2023, 22, 138. [Google Scholar] [CrossRef]
- Ersahin, T.; Tuncbag, N.; Cetin-Atalay, R. The PI3K/AKT/MTOR Interactive Pathway. Mol Biosyst 2015, 11, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Ersahin, T.; Tuncbag, N.; Cetin-Atalay, R. The PI3K/AKT/MTOR Interactive Pathway. Mol Biosyst 2015, 11, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, D.; Kim, E.-Y.; Rahman, A.H.; Raghunathan, R.; Kim-Schulze, S.; Che, Y.; Kalayci, S.; Gümüş, Z.H.; Kuan, G.; Balmaseda, A.; et al. Comprehensive Immunoprofiling of Pediatric Zika Reveals Key Role for Monocytes in the Acute Phase and No Effect of Prior Dengue Virus Infection. Cell Rep 2020, 31, 107569. [Google Scholar] [CrossRef]
- Srivastava, M.; Zhang, Y.; Chen, J.; Sirohi, D.; Miller, A.; Zhang, Y.; Chen, Z.; Lu, H.; Xu, J.; Kuhn, R.J.; et al. Chemical Proteomics Tracks Virus Entry and Uncovers NCAM1 as Zika Virus Receptor. Nat Commun 2020, 11, 3896. [Google Scholar] [CrossRef]
- Tatara, J.M.; Santi, L.; Beys-da-Silva, W.O. Proteome Alterations Promoted by Zika Virus Infection. In Zika Virus Biology, Transmission, and Pathology; Elsevier, 2021; pp. 307–317.
- Sun, X.; Hua, S.; Gao, C.; Blackmer, J.E.; Ouyang, Z.; Ard, K.; Ciaranello, A.; Yawetz, S.; Sax, P.E.; Rosenberg, E.S.; et al. Immune-Profiling of ZIKV-Infected Patients Identifies a Distinct Function of Plasmacytoid Dendritic Cells for Immune Cross-Regulation. Nat Commun 2020, 11, 2421. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, D.; Kim, E.-Y.; Rahman, A.H.; Raghunathan, R.; Kim-Schulze, S.; Che, Y.; Kalayci, S.; Gümüş, Z.H.; Kuan, G.; Balmaseda, A.; et al. Comprehensive Immunoprofiling of Pediatric Zika Reveals Key Role for Monocytes in the Acute Phase and No Effect of Prior Dengue Virus Infection. Cell Rep 2020, 31, 107569. [Google Scholar] [CrossRef]
- Giacobino, C.; Canta, M.; Fornaguera, C.; Borrós, S.; Cauda, V. Extracellular Vesicles and Their Current Role in Cancer Immunotherapy. Cancers (Basel) 2021, 13, 2280. [Google Scholar] [CrossRef]
- Chang, W.-H.; Cerione, R.A.; Antonyak, M.A. Extracellular Vesicles and Their Roles in Cancer Progression. In; 2021; pp. 143–170.
- Akhmerov, A.; Parimon, T. Extracellular Vesicles, Inflammation, and Cardiovascular Disease. Cells 2022, 11, 2229. [Google Scholar] [CrossRef]
- Du, Y.; Wu, L.; Wang, L.; Reiter, R.J.; Lip, G.Y.H.; Ren, J. Extracellular Vesicles in Cardiovascular Diseases: From Pathophysiology to Diagnosis and Therapy. Cytokine Growth Factor Rev 2023, 74, 40–55. [Google Scholar] [CrossRef]
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas Aeruginosa: Pathogenesis, Virulence Factors, Antibiotic Resistance, Interaction with Host, Technology Advances and Emerging Therapeutics. Signal Transduct Target Ther 2022, 7, 199. [Google Scholar] [CrossRef]
- Peregrino, E.S.; Castañeda-Casimiro, J.; Vázquez-Flores, L.; Estrada-Parra, S.; Wong-Baeza, C.; Serafín-López, J.; Wong-Baeza, I. The Role of Bacterial Extracellular Vesicles in the Immune Response to Pathogens, and Therapeutic Opportunities. Int J Mol Sci 2024, 25, 6210. [Google Scholar] [CrossRef]
- Buzas, E.I. The Roles of Extracellular Vesicles in the Immune System. Nat Rev Immunol 2023, 23, 236–250. [Google Scholar] [CrossRef]
- Garcia-Flores, V.; Romero, R.; Xu, Y.; Theis, K.R.; Arenas-Hernandez, M.; Miller, D.; Peyvandipour, A.; Bhatti, G.; Galaz, J.; Gershater, M.; et al. Maternal-Fetal Immune Responses in Pregnant Women Infected with SARS-CoV-2. Nat Commun 2022, 13, 320. [Google Scholar] [CrossRef]
- Heuberger, D.M.; Schuepbach, R.A. Protease-Activated Receptors (PARs): Mechanisms of Action and Potential Therapeutic Modulators in PAR-Driven Inflammatory Diseases. Thromb J 2019, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Tato, C.M.; Davis, M.M. How the Immune System Talks to Itself: The Varied Role of Synapses. Immunol Rev 2013, 251, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Dixson, A.C.; Dawson, T.R.; Di Vizio, D.; Weaver, A.M. Context-Specific Regulation of Extracellular Vesicle Biogenesis and Cargo Selection. Nat Rev Mol Cell Biol 2023, 24, 454–476. [Google Scholar] [CrossRef] [PubMed]
- Robbins, P.D.; Morelli, A.E. Regulation of Immune Responses by Extracellular Vesicles. Nat Rev Immunol 2014, 14, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Wright, S.S.; Rathinam, V.A. Role of Extracellular Vesicles in Immunity and Host Defense. Immunol Invest 2024, 53, 10–25. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Sun, X.; Wu, C.; Yao, L.; Zhang, Y.; Liu, S.; Jiang, Y.; Li, Y.; Wang, M.; Xu, Y. PROS1, a Clinical Prognostic Biomarker and Tumor Suppressor, Is Associated with Immune Cell Infiltration in Breast Cancer: A Bioinformatics Analysis Combined with Experimental Verification. Cell Signal 2023, 112, 110918. [Google Scholar] [CrossRef] [PubMed]
- Busatto, S.; Morad, G.; Guo, P.; Moses, M.A. The Role of Extracellular Vesicles in the Physiological and Pathological Regulation of the Blood–Brain Barrier. FASEB Bioadv 2021, 3, 665–675. [Google Scholar] [CrossRef]
- Gamage, T.K.J.B.; Fraser, M. The Role of Extracellular Vesicles in the Developing Brain: Current Perspective and Promising Source of Biomarkers and Therapy for Perinatal Brain Injury. Front Neurosci 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Cai, Y.; Lin, J.; Ma, L.; Han, H.; Li, F. Advancement in Modulation of Brain Extracellular Space and Unlocking Its Potential for Intervention of Neurological Diseases. Med-X 2024, 2, 6. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat Rev Mol Cell Biol 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Voltarelli, V.A.; Alves de Souza, R.W.; Miyauchi, K.; Hauser, C.J.; Otterbein, L.E. Heme: The Lord of the Iron Ring. Antioxidants 2023, 12, 1074. [Google Scholar] [CrossRef] [PubMed]
- Mense, S.M.; Zhang, L. Heme: A Versatile Signaling Molecule Controlling the Activities of Diverse Regulators Ranging from Transcription Factors to MAP Kinases. Cell Res 2006, 16, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Ellis-Guardiola, K.; Soule, J.; Clubb, R. Methods for the Extraction of Heme Prosthetic Groups from Hemoproteins. Bio Protoc 2021, 11. [Google Scholar] [CrossRef]
- Bandu, R.; Oh, J.W.; Kim, K.P. Mass Spectrometry-Based Proteome Profiling of Extracellular Vesicles and Their Roles in Cancer Biology. Exp Mol Med 2019, 51, 1–10. [Google Scholar] [CrossRef]
- Kwon, Y.; Oh, I.; Lee, N.; Lee, K.; Yoon, Y.J.; Lee, E.Y.; Kim, B.; Kim, D. Integrating Cell-free Biosyntheses of Heme Prosthetic Group and Apoenzyme for the Synthesis of Functional P450 Monooxygenase. Biotechnol Bioeng 2013, 110, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, M.C.; Sarkar, S.; Elliott, E.C.; Henry, H.R.; Powell, S.M.; Diaz Ludovico, I.; You, Y.; Huang, F.; Payne, S.H.; Ramanadham, S.; et al. A Proteomic Meta-Analysis Refinement of Plasma Extracellular Vesicles. Sci Data 2023, 10, 837. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-J.; Huang, Y.-N.; Lu, Y.-B.; Zhang, Y.; Wu, P.-H.; Huang, J.-S.; Yang, W.; Chiang, T.-Y.; Hsieh, H.-S.; Chung, W.-H.; et al. Proteomic Analysis of Serum Extracellular Vesicles from Biliary Tract Infection Patients to Identify Novel Biomarkers. Sci Rep 2024, 14, 5707. [Google Scholar] [CrossRef] [PubMed]
- Badosa, C.; Roldán, M.; Fernández-Irigoyen, J.; Santamaria, E.; Jimenez-Mallebrera, C. Proteomic and Functional Characterisation of Extracellular Vesicles from Collagen VI Deficient Human Fibroblasts Reveals a Role in Cell Motility. Sci Rep 2023, 13, 14622. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zhang, B.; Ocansey, D.K.W.; Xu, W.; Qian, H. Extracellular Vesicles: A Bright Star of Nanomedicine. Biomaterials 2021, 269, 120467. [Google Scholar] [CrossRef] [PubMed]
- Soloveva, N.; Novikova, S.; Farafonova, T.; Tikhonova, O.; Zgoda, V. Proteomic Signature of Extracellular Vesicles Associated with Colorectal Cancer. Molecules 2023, 28, 4227. [Google Scholar] [CrossRef] [PubMed]
- Barouch-Bentov, R.; Neveu, G.; Xiao, F.; Beer, M.; Bekerman, E.; Schor, S.; Campbell, J.; Boonyaratanakornkit, J.; Lindenbach, B.; Lu, A.; et al. Hepatitis C Virus Proteins Interact with the Endosomal Sorting Complex Required for Transport (ESCRT) Machinery via Ubiquitination To Facilitate Viral Envelopment. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, A.; Kumar, A.; Wencker, M.; Ricci, E.P. Shaping the Innate Immune Response Through Post-Transcriptional Regulation of Gene Expression Mediated by RNA-Binding Proteins. Front Immunol 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cao, X. Cellular and Molecular Regulation of Innate Inflammatory Responses. Cell Mol Immunol 2016, 13, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Medjeral-Thomas, N.R.; Lomax-Browne, H.J.; Beckwith, H.; Willicombe, M.; McLean, A.G.; Brookes, P.; Pusey, C.D.; Falchi, M.; Cook, H.T.; Pickering, M.C. Circulating Complement Factor H–Related Proteins 1 and 5 Correlate with Disease Activity in IgA Nephropathy. Kidney Int 2017, 92, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, W.; Tong, Y.; Sun, M.; Quan, S.; Zhu, J.; Zhang, Q.; Qin, Z.; Ni, Y.; Zhao, Y.; et al. Integrative Proteomics and Metabolomics Study Reveal Enhanced Immune Responses by COVID-19 Vaccine Booster Shot against Omicron SARS-CoV-2 Infection. J Med Virol 2023, 95. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.M.; Zhang, C.; Wang, Z.; Ni, Z.; Zhang, S.; Yang, S.; Huang, X.; Mo, L.; Li, J.; Lee, B.; et al. A Multi-Omics Investigation of the Composition and Function of Extracellular Vesicles along the Temporal Trajectory of COVID-19. Nat Metab 2021, 3, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yao, F.; Yin, Y.; Wu, C.; Xia, D.; Zhang, K.; Jin, Z.; Liu, X.; He, J.; Zhang, Z. Extracellular Vesicles Derived from Immune Cells: Role in Tumor Therapy. Int Immunopharmacol 2024, 133, 112150. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Xu, Y.; Liu, N.; Lv, D.; Chen, Y.; Liu, Z.; Jin, X.; Xiao, M.; Lavillette, D.; Zhong, J.; et al. Extracellular Vesicles from Zika Virus-infected Cells Display Viral E Protein That Binds ZIKV-neutralizing Antibodies to Prevent Infection Enhancement. EMBO J 2023, 42. [Google Scholar] [CrossRef] [PubMed]
- Fikatas, A.; Dehairs, J.; Noppen, S.; Doijen, J.; Vanderhoydonc, F.; Meyen, E.; Swinnen, J. V.; Pannecouque, C.; Schols, D. Deciphering the Role of Extracellular Vesicles Derived from ZIKV-Infected HcMEC/D3 Cells on the Blood–Brain Barrier System. Viruses 2021, 13, 2363. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kodidela, S.; Tadrous, E.; Cory, T.J.; Walker, C.M.; Smith, A.M.; Mukherjee, A.; Kumar, S. Extracellular Vesicles in Viral Replication and Pathogenesis and Their Potential Role in Therapeutic Intervention. Viruses 2020, 12, 887. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.A.; Narayanan, A.; Moustafa, I.M.; Bator, C.M.; Hafenstein, S.L.; Jose, J. Zika Virus M Protein Latches and Locks the E Protein from Transitioning to an Immature State after PrM Cleavage. npj Viruses 2023, 1, 4. [Google Scholar] [CrossRef]
- Macedo-da-Silva, J.; Santiago, V.F.; Rosa-Fernandes, L.; Marinho, C.R.F.; Palmisano, G. Protein Glycosylation in Extracellular Vesicles: Structural Characterization and Biological Functions. Mol Immunol 2021, 135, 226–246. [Google Scholar] [CrossRef] [PubMed]
- Smelik, M.; Zhao, Y.; Li, X.; Loscalzo, J.; Sysoev, O.; Mahmud, F.; Mansour Aly, D.; Benson, M. An Interactive Atlas of Genomic, Proteomic, and Metabolomic Biomarkers Promotes the Potential of Proteins to Predict Complex Diseases. Sci Rep 2024, 14, 12710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Takeuchi, T.; Takeda, A.; Mochizuki, H.; Nagai, Y. Comparison of Serum and Plasma as a Source of Blood Extracellular Vesicles: Increased Levels of Platelet-Derived Particles in Serum Extracellular Vesicle Fractions Alter Content Profiles from Plasma Extracellular Vesicle Fractions. PLoS One 2022, 17, e0270634. [Google Scholar] [CrossRef] [PubMed]
- Schumacher-Schuh, A.; Bieger, A.; Borelli, W. V.; Portley, M.K.; Awad, P.S.; Bandres-Ciga, S. Advances in Proteomic and Metabolomic Profiling of Neurodegenerative Diseases. Front Neurol 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Arakelyan, A.; Fitzgerald, W.; Zicari, S.; Vanpouille, C.; Margolis, L. Extracellular Vesicles Carry HIV Env and Facilitate Hiv Infection of Human Lymphoid Tissue. Sci Rep 2017, 7, 1695. [Google Scholar] [CrossRef] [PubMed]
- Cyster, J.G.; Allen, C.D.C. B Cell Responses: Cell Interaction Dynamics and Decisions. Cell 2019, 177, 524–540. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. The Emergence of Zika Virus and Its New Clinical Syndromes. Nature 2018, 560, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Giovanetti, M.; Pereira, L.A.; Adelino, T.É.R.; Fonseca, V.; Xavier, J.; de Araújo Fabri, A.; Slavov, S.N.; da Silva Lemos, P.; de Almeida Marques, W.; Kashima, S.; et al. A Retrospective Overview of Zika Virus Evolution in the Midwest of Brazil. Microbiol Spectr 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Hwang, K.-K.; Miller, A.S.; Jones, R.L.; Lopez, C.A.; Dulson, S.J.; Giuberti, C.; Gladden, M.A.; Miller, I.; Webster, H.S.; et al. A Zika Virus-Specific IgM Elicited in Pregnancy Exhibits Ultrapotent Neutralization. Cell 2022, 185, 4826–4840. [Google Scholar] [CrossRef]
- Yeung, C.-Y.C.; Dondelinger, F.; Schoof, E.M.; Georg, B.; Lu, Y.; Zheng, Z.; Zhang, J.; Hannibal, J.; Fahrenkrug, J.; Kjaer, M. Circadian Regulation of Protein Cargo in Extracellular Vesicles. Sci Adv 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Zebrowska, A.; Jelonek, K.; Mondal, S.; Gawin, M.; Mrowiec, K.; Widłak, P.; Whiteside, T.; Pietrowska, M. Proteomic and Metabolomic Profiles of T Cell-Derived Exosomes Isolated from Human Plasma. Cells 2022, 11, 1965. [Google Scholar] [CrossRef] [PubMed]
- Bludau, I.; Aebersold, R. Proteomic and Interactomic Insights into the Molecular Basis of Cell Functional Diversity. Nat Rev Mol Cell Biol 2020, 21, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Greening, D.W.; Xu, R.; Gopal, S.K.; Rai, A.; Simpson, R.J. Proteomic Insights into Extracellular Vesicle Biology – Defining Exosomes and Shed Microvesicles. Expert Rev Proteomics 2017, 14, 69–95. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, I.; Vo, T.; Paudel, K.; Wen, X.; Gupta, R.; Kesimer, M.; Patial, S.; Saini, Y. Vesicular and Extravesicular Protein Analyses from the Airspaces of Ozone-Exposed Mice Revealed Signatures Associated with Mucoinflammatory Lung Disease. Sci Rep 2021, 11, 23203. [Google Scholar] [CrossRef] [PubMed]
- Meier-Soelch, J.; Mayr-Buro, C.; Juli, J.; Leib, L.; Linne, U.; Dreute, J.; Papantonis, A.; Schmitz, M.L.; Kracht, M. Monitoring the Levels of Cellular NF-ΚB Activation States. Cancers (Basel) 2021, 13, 5351. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Jin, Y.; Chen, X.; Ye, X.; Shen, X.; Lin, M.; Zeng, C.; Zhou, T.; Zhang, J. NF-ΚB in Biology and Targeted Therapy: New Insights and Translational Implications. Signal Transduct Target Ther 2024, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Nix, C.; Fillet, M. New Insights into Extracellular Vesicles of Clinical and Therapeutic Interest Using Proteomic Mass Spectrometry Approaches. TrAC Trends in Analytical Chemistry 2024, 178, 117823. [Google Scholar] [CrossRef]
- Denolly, S.; Stukalov, A.; Barayeu, U.; Rosinski, A.N.; Kritsiligkou, P.; Joecks, S.; Dick, T.P.; Pichlmair, A.; Bartenschlager, R. Zika Virus Remodelled ER Membranes Contain Proviral Factors Involved in Redox and Methylation Pathways. Nat Commun 2023, 14, 8045. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.G.; Lim, X.N.; Ng, W.C.; Sim, A.Y.L.; Poh, H.X.; Shen, Y.; Lim, S.Y.; Sundstrom, K.B.; Sun, X.; Aw, J.G.; et al. Structure Mapping of Dengue and Zika Viruses Reveals Functional Long-Range Interactions. Nat Commun 2019, 10, 1408. [Google Scholar] [CrossRef] [PubMed]
- MIRZAA, G.M.; RIVIÈRE, J.; DOBYNS, W.B. Megalencephaly Syndromes and Activating Mutations in the PI3K-AKT Pathway: MPPH and MCAP. Am J Med Genet C Semin Med Genet 2013, 163, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y. Roles of MTOR Signaling in Brain Development. Exp Neurobiol 2015, 24, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Wahane, S.D.; Hellbach, N.; Prentzell, M.T.; Weise, S.C.; Vezzali, R.; Kreutz, C.; Timmer, J.; Krieglstein, K.; Thedieck, K.; Vogel, T. PI3K-p110-alpha-subtype Signalling Mediates Survival, Proliferation and Neurogenesis of Cortical Progenitor Cells via Activation of <scp>mTORC</Scp> 2. J Neurochem 2014, 130, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Dai, Q.; Yang, R.; Duan, Y.; Zhao, Q.; Haybaeck, J.; Yang, Z. A Review: PI3K/AKT/MTOR Signaling Pathway and Its Regulated Eukaryotic Translation Initiation Factors May Be a Potential Therapeutic Target in Esophageal Squamous Cell Carcinoma. Front Oncol 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, Y.; Ren, H.; Yao, Q.; Ba, J.; Luan, J.; Zhao, P.; Qin, Z.; Qi, Z. MTOR Signaling Regulates Zika Virus Replication Bidirectionally through Autophagy and Protein Translation. J Med Virol 2023, 95. [Google Scholar] [CrossRef] [PubMed]
- Franke, T.F. PI3K/Akt: Getting It Right Matters. Oncogene 2008, 27, 6473–6488. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Shin, O.S. Zika Virus Modulates Mitochondrial Dynamics, Mitophagy, and Mitochondria-Derived Vesicles to Facilitate Viral Replication in Trophoblast Cells. Front Immunol 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Sirohi, D.; Kuhn, R.J. Zika Virus Structure, Maturation, and Receptors. J Infect Dis 2017, 216, S935–S944. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.E.; Rossignol, E.D.; Chang, D.; Zaia, J.; Forrester, I.; Raja, K.; Winbigler, H.; Nicastro, D.; Jackson, W.T.; Bullitt, E. Complexity and Ultrastructure of Infectious Extracellular Vesicles from Cells Infected by Non-Enveloped Virus. Sci Rep 2020, 10, 7939. [Google Scholar] [CrossRef] [PubMed]
- Scaturro, P.; Stukalov, A.; Haas, D.A.; Cortese, M.; Draganova, K.; Płaszczyca, A.; Bartenschlager, R.; Götz, M.; Pichlmair, A. An Orthogonal Proteomic Survey Uncovers Novel Zika Virus Host Factors. Nature 2018, 561, 253–257. [Google Scholar] [CrossRef] [PubMed]
- York, S.B.; Sun, L.; Cone, A.S.; Duke, L.C.; Cheerathodi, M.R.; Meckes, D.G. Zika Virus Hijacks Extracellular Vesicle Tetraspanin Pathways for Cell-to-Cell Transmission. mSphere 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Hui, L.; Nie, Y.; Tefsen, B.; Wu, Y. ZIKV Viral Proteins and Their Roles in Virus-Host Interactions. Sci China Life Sci 2021, 64, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.Y.; Gustin, A.; Newhouse, D.; Gale, M. Viral Protein Accumulation of Zika Virus Variants Links with Regulation of Innate Immunity for Differential Control of Viral Replication, Spread, and Response to Interferon. J Virol 2023, 97. [Google Scholar] [CrossRef] [PubMed]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. The Journal of Immunology 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.J.; Komarasamy, T.V.; Adnan, N.A.A.; James, W.; RMT Balasubramaniam, V. Hide and Seek: The Interplay Between Zika Virus and the Host Immune Response. Front Immunol 2021, 12. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Analysis for EV Characterization: A) Results of quantification by NanoDrop One, B) Nanoparticle tracking analysis and C) transmission electron microscopy of extracellular vesicles isolated in the children's serum.
Figure 1.
Analysis for EV Characterization: A) Results of quantification by NanoDrop One, B) Nanoparticle tracking analysis and C) transmission electron microscopy of extracellular vesicles isolated in the children's serum.
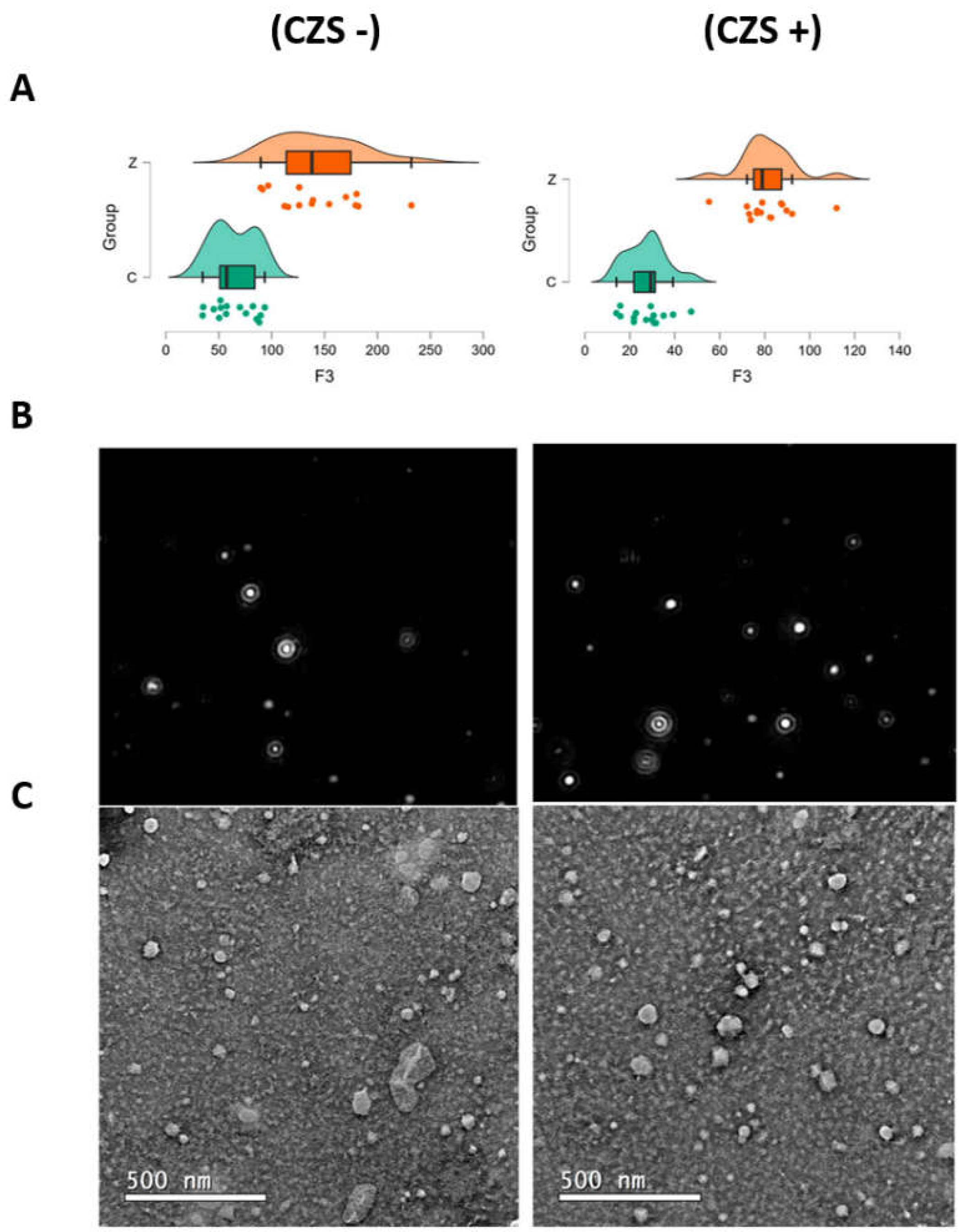
Figure 2.
Fold change of proteins identified by mass spectrometry that showed positive and negative regulation.
Figure 2.
Fold change of proteins identified by mass spectrometry that showed positive and negative regulation.

Figure 3.
Circles represent proteins, while straight lines represent interactions between different proteins. Stronger associations are represented by thicker lines. A) Interaction network between proteins identified by mass spectrometry that showed positive regulation, compared to the control. B) Interaction network between proteins identified by mass spectrometry that showed negative regulation, compared to the control. This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
Figure 3.
Circles represent proteins, while straight lines represent interactions between different proteins. Stronger associations are represented by thicker lines. A) Interaction network between proteins identified by mass spectrometry that showed positive regulation, compared to the control. B) Interaction network between proteins identified by mass spectrometry that showed negative regulation, compared to the control. This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
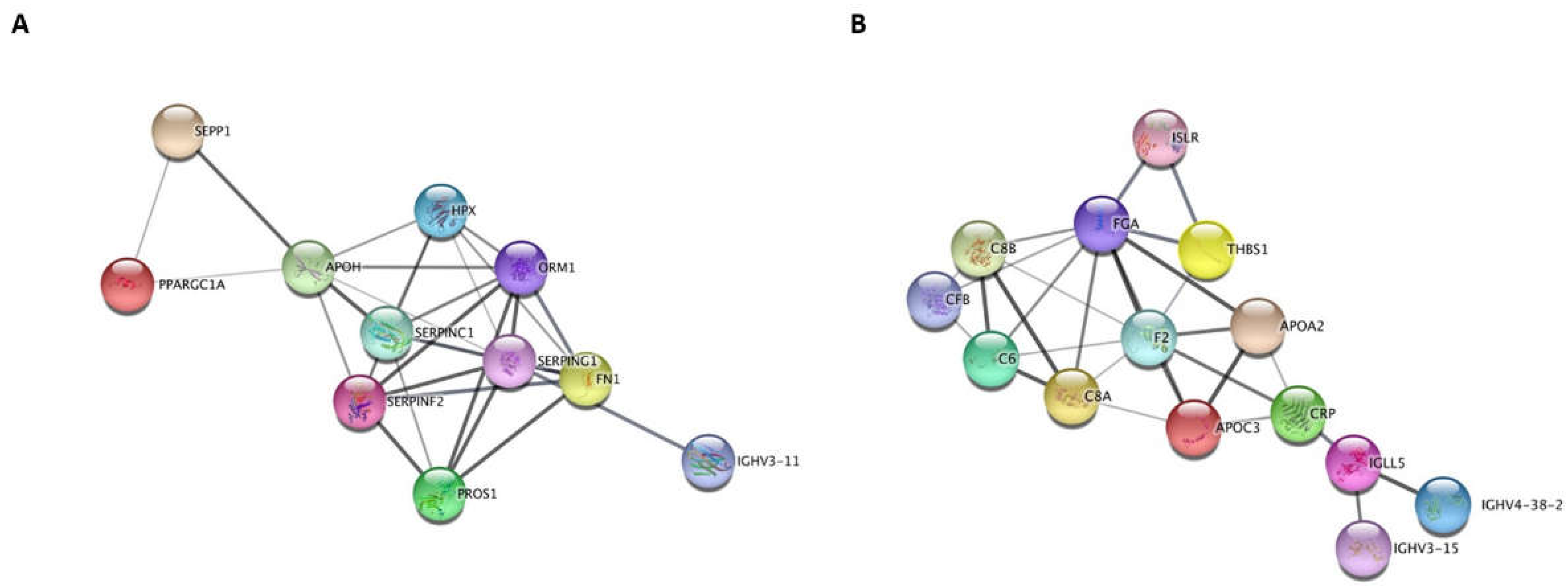
Figure 4.
Data analysis by volcano plot using the MetaboAnalyst 4.0 software. A) Analysis of main components of the analyzed data set obtained from proteins; B) Principal component analysis of the analyzed dataset obtained from metabolites. In both images, the red components represent the EV samples from CZS+ children and the green components represent the EV samples from CZS- children.
Figure 4.
Data analysis by volcano plot using the MetaboAnalyst 4.0 software. A) Analysis of main components of the analyzed data set obtained from proteins; B) Principal component analysis of the analyzed dataset obtained from metabolites. In both images, the red components represent the EV samples from CZS+ children and the green components represent the EV samples from CZS- children.
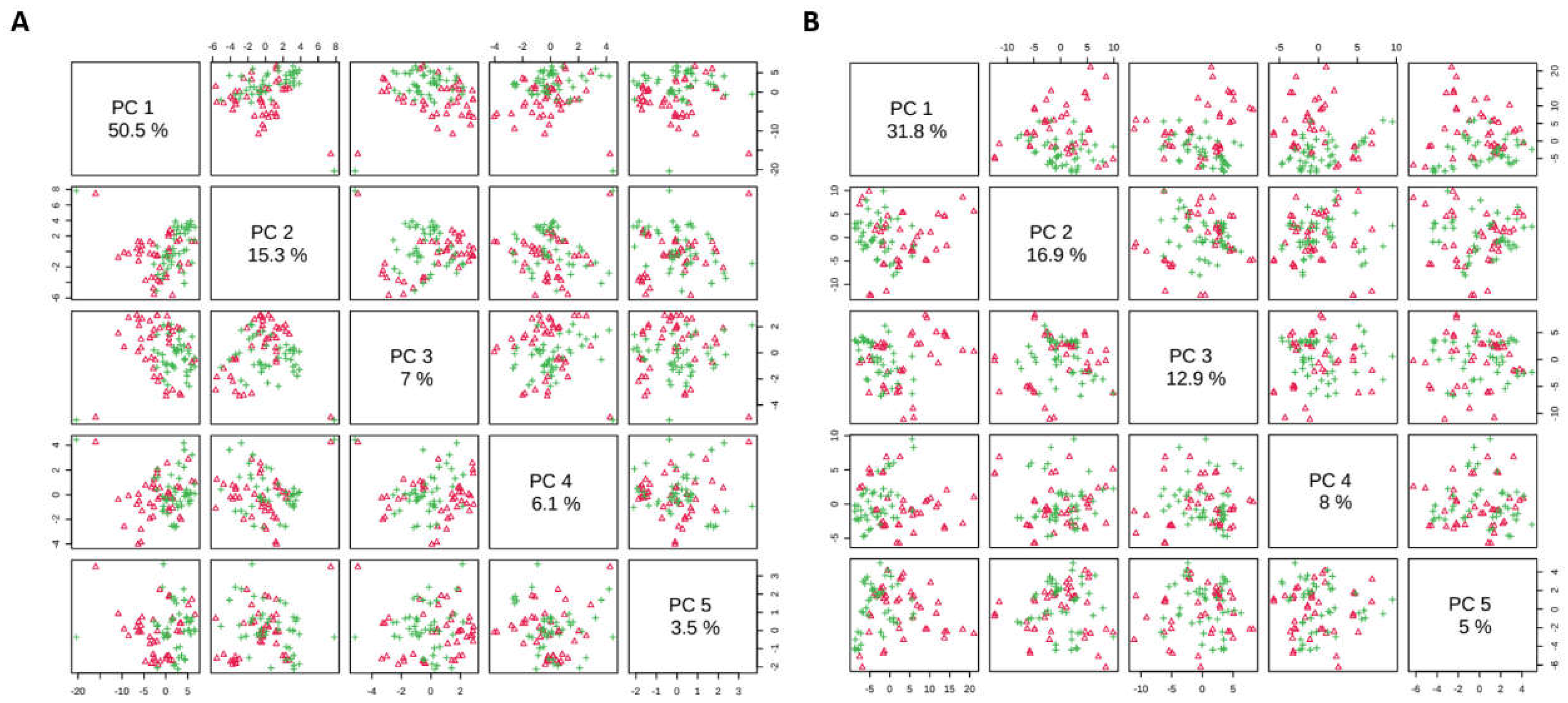
Figure 5.
Data analysis by volcano plot using the MetaboAnalyst 4.0 software. A) Volcano plot of the abundance of proteins identified by proteomics. B) Volcano plot of the abundance of metabolites identified by the metabolomics platform. In both graphs, the numbers and letters represent the code for each protein identified and analyzed. Red circles represent features above (UP) the threshold for fold change and t-test threshold (p ≤ 0.05) compared to control samples. Blue circles represent features below (DOWN) the fold change threshold and t-test threshold (p ≤ 0.05) compared to ZIKV samples. Both values are log transformed.
Figure 5.
Data analysis by volcano plot using the MetaboAnalyst 4.0 software. A) Volcano plot of the abundance of proteins identified by proteomics. B) Volcano plot of the abundance of metabolites identified by the metabolomics platform. In both graphs, the numbers and letters represent the code for each protein identified and analyzed. Red circles represent features above (UP) the threshold for fold change and t-test threshold (p ≤ 0.05) compared to control samples. Blue circles represent features below (DOWN) the fold change threshold and t-test threshold (p ≤ 0.05) compared to ZIKV samples. Both values are log transformed.
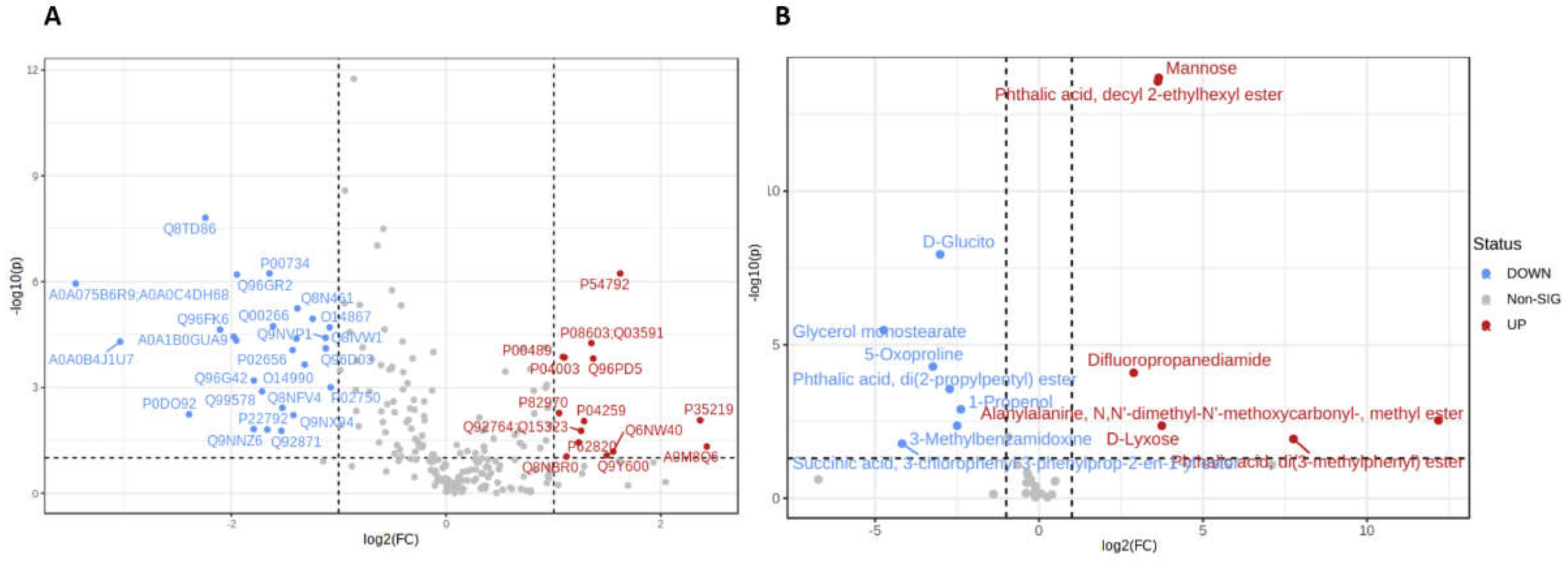
Figure 6.
Part of the image from the analysis of categories from the public domain database obtained in this study of serum EVs from CZS+ children.
Figure 6.
Part of the image from the analysis of categories from the public domain database obtained in this study of serum EVs from CZS+ children.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



