Submitted:
10 July 2024
Posted:
11 July 2024
You are already at the latest version
Abstract
Due to the genotype-dependent effects of water deficit (WD) on coffee physiology, climate change raises the risk of coffee yield. The aim of the study was to assess the effect of WD on physiological and growth traits in arabica coffee (Coffea arabica L.). Water status, photosynthetic response to in-tercellular concentration of CO2 (A/Ci curves) and growth parameters were evaluated in seedlings of four genotypes (Catimor ECU 02, Cavimor ECU, red Caturra and Sarchimor 4260). Most of the physiological traits evaluated differed significantly among genotypes. Between control and WD plants, there were significant differences in A/Ci parameters, showing a wide range of values for photosynthetic rate, stomatal conductance, and water use efficiency, with decreases ranging from 4 to 74%. Maximum rate of carboxylation by RuBisCO, maximum electron transport rate through photosystem II, and triose phosphate utilization rate were all strongly reduced by WD 61 % (red Caturra and Sarchimor 4260), followed by Cavimor ECU (35 %) and 24 % in Catimor ECU 02. Differences in response to WD among genotypes suggest possible genotypic differences in toler-ance. The results indicated that Catimor ECU 02 and Cavimor ECU were less sensitive to WD, while red Caturra and Sarchimor 4260 were the most susceptible.
Keywords:
Coffea arabica
; growth
; stomatal conductance
; photosynthesis
; water use efficiency
1. Introduction
Native to northern Ethiopia, arabica coffee (Coffea arabica L., Rubiaceae) is a tropical woody crop with great genetic value that is one of the most traded crops worldwide [1,2]. Twenty-five million agricultural households, mostly smallholders, grow coffee in 80 tropical countries for the consumption of one-third of the world's population [3,4].
Coffee is the second most valuable commodity in the world after oil, with sales estimated to be worth US$102 billion globally [5]. World coffee production was between 158 and 178 million 60 kg bags per year for the years 2023-2024 [6]. In Ecuador, coffee is a product of great social, economic and environmental importance [7] and its production during 2019-2020 was 559,000 and 500,000 bags, respectively, with a percentage of change of -10.5% [5].
In Brazil coffee has been extensively investigated ecophysiologically [4,8,9,10,11,12,13], while in Ecuador such studies are scant [14]. Ecuador's genetic improvement programs lack physiological information, particularly on the physiological response to water deficit (WD), despite knowledge on production, pests, and diseases tolerance in various coffee genotypes [7,14,15]. In order to choose coffee genotypes with improved physiological performance in WD conditions in Ecuador's various agroecological zones, it is necessary to understand the potential traits.
Due to global climate change, projections for Ecuador in 2050 indicate temperatures could rise by up to 2.5 °C, along with less rainfall and longer, more unpredictable droughts [16]. These temperature changes, coupled with increased climate variability, are anticipated to pose a danger to global agricultural productivity and sustainability, having a significant impact on the quantity and quality of crops, such as coffee [17,18,19]; these situations emphasize how important it is to comprehend how tropical crops responses physiologically to WD [3,20].
The Coffea species is very sensitive to climate change, according to modeling studies based on predictions of rising temperatures and shifting patterns of precipitation [3]. Changes in microclimatic factors, particularly those associated with WD, have a significant impact on coffee output and bean quality [20,21].
The physiological responses of plants to WD are intricate due to the disruption of numerous metabolic systems. Reduced growth and yield are caused by a drop in net photosynthetic rate (A) resulting from either impaired metabolic processes or reduced CO2 diffusion to carboxylation sites [22]. A wide range of plant responses to drought can be attributed to the physiological, biochemical, morphological, and molecular mechanisms involved; they include dynamic soil water depletion and failure to meet the water demands of growing coffee plants at different phenological states [20].
Low water availability reduces coffee production by interfering with the roots' ability to extract moisture, the roots' distribution in space, the size of the canopy, and the growth of their fruit [9]. A key factor in the genotypes of arabica coffee's variable tolerance to low water availability could be attributed to physiological and morphological features that influence water uptake and loss, such as A, stomatal conductance (gs), water use efficiency (WUE) and root depth, which should be recommended as traits for the potential selection of coffee genotypes with higher yields subjected to WD [9,23,24]. Understanding how WD affects arabica coffee's physiology and metabolism is crucial for developing drought resistant varieties [20,25].
We assessed the impact of WD on leaf and soil water status, leaf gas exchange, and growth traits of four Coffea arabica genotypes: Sarchimor 4260, Red Caturra, Catimor ECU 02, and Cavimor ECU. Finding potential characteristics to choose coffee genotypes with improved physiological performance in low-water environments was the goal of this study. This information can offer understandings into whether differences in genetic variability among C. arabica genotypes explain photosynthetic differences, and if the physiological response to WD is different among genotypes. Under WD, we hypothesized that: 1) coffee genotypes will respond differently from one another due to varying physiological performance; and 2) coffee genotypes might have varying effects on WUE.
2. Materials and Methods
2.1. Study Area and Site Conditions
The investigation was carried out in 2017 at 20 m.a.s.l at the Mutile Experimental Station, Universidad Técnica Luis Vargas Torres, in the Esmeralda Province of Ecuador (0 ° 53' 40" N; 79 ° 37´23" W). The research area's parameters include mean air temperature (Ta) of 24 °C, relative humidity (RH) of 85%, heliophany of 1,200 hours of sunlight per year, and mean annual precipitation of 1,030 mm [26]. One-year-all arabica coffee seedlings were raised in a 400-m2 shed shielded with a neutral polythene sheet, whereafter named shade house. Utilizing a light meter (LI-250l, LI-COR, Lincoln, Nebraska, USA), the photosynthetic photon flux density (PPFD) was determined. Values for the leaf-to-air vapor pressure deficit (ΔW) were obtained using Ta and RH data, following Jones' [27] equation. Table 1 displays the values of the shade house's microclimatic parameters. Shading reduced PPFD incident at noon on the roof to 20 %.
2.2. Plant Material
The Coffea arabica genotypes Sarchimor 4260, Red Caturra, Catimor ECU 02, and Cavimor ECU, which are commonly used in Ecuador, were chosen for this study due to their high production and resistance to coffee rust (Table 2). In the most important coffee-producing regions of Esmeraldas Province, Ecuador, we gathered seeds. Ten-month-old seedlings were cultivated, under semi-controlled conditions in the shade house, in bags with 10 kg of sandy loam soil (pH = 6.0; organic matter = 2 %; total N = 0.1 %; total P = 21 mg kg-1; total K = 1.4 cmol kg-1) and watered every two days. In order to ensure seedling growth and establishment, we fertilized plants once a month (10 g month-1) with N:P:K 12:11:18, a dose that has been shown to yield the greatest outcomes.
2.3. Experimental Design
A complete random block design was utilized with four genotypes, each with 20 plants and three repeats, for a total of 60 plants per genotype and 240 plants in total. In the shade house, three blocks totaling 121 m2 were set up, with 20 plants of each genotype spaced one meter apart in each block. Every block was separated by two meters. To prevent potential block and edge effects, plants were shifted weekly within and across blocks within each treatment. During seven months (February to August 2017), all physiological parameters were measured on seedlings that were ten months old. For each genotype (n = 5), a sample was taken from the central region of a fully opened, healthy leaf, which was the third leaf from the top.
2.4. Water Deficit Experiment
Irrigation in 10 seedlings of each genotype per block (total of 30 plants) was completely stopped for 29 days and corresponded to the water deficit treatment (WD). The remaining 30 plants were maintained well-watered and corresponded to the control. We measured water status as soil and leaf water contents, gas exchange variables, and performed A/Ci curves on days 0 (the first without irrigation) and on the following days: 8, 15, 22, and 29. On day 29, we started irrigation again, and the following day (on day 30), we took measurements of the same set of physiological variables.
2.5. Water Status
To determine the soil water content (SWC), relative water content (RWC), and leaf water content (LWC), samples of soil and leaves were collected. After soil samples were taken at a depth of 15 cm, their fresh weight (FW) was calculated, they were dried for 48 hours at 70 °C to achieve a constant mass, and then they were weighed to obtain their dry weight (DW). Soil water content was calculated as [(FW - DW) / FW] × 100. Ten leaves (n = 10) were collected between 7:00 and 8:00 h, and the leaf water content (LWC) was calculated as (FW - DW)/DW. To calculate the turgid weight (TW), ten leaves (n = 10) were gathered at 07:00 h and floated on distilled water at 4 °C for one hour. Leaf relative water content was then calculated as RWC = (FW-DW)/(TW-DW). The specific leaf area (SLA), which is the ratio of leaf area to DW, was determined by measuring the fresh area and DW of leaves taken from six individuals per genotype.
2.6. Gas Exchange
A portable infrared gas analyzer (CIRAS-II, PP Systems, MA, USA) in open mode linked to a PLC(B) leaf chamber was used to measure the gas exchange variables (A, gs, and E) in intact leaves in five individuals of each genotype (n = 5 per genotype). With PPFD of 1200 ± 10 μmol m-2 s-1 delivered by an LED Based Light Unit from PP systems Inc., measurements were conducted at a CO2 concentration of 410 ± 10 μmol mol-1, a leaf temperature of 30.0 ± 0.5 °C, and a ΔW of 1.8-2.2 kPa. The WUE was calculated as the ration between A and E. Every genotype that was randomly sampled had measurements taken throughout the course of three days, from 8:00 to 12:00.
2.7. A/Ci Curves
We performed A/Ci curves (n = 4) by gradually raising Ca to 1.800 μmol mol-1 after first lowering Ca to zero. The A/Ci measurements were conducted in a leaf chamber with the following parameters: ΔW of 1.8 ± 0.02 kPa, PPFD of 1.200 ± 10 μmol m-2 s-1, and leaf temperature of 30.0 ± 0.5 °C. An empirical equation was used to fit curves [28] that yields the carboxylation efficiency (CE), as the slope of the initial part of the curve. Using the equations in [29] and [30], the values of the relative stomatal limitation (Ls) and mesophyll limitation (Lm) were determined. Finally, the model in [31] was used to compute the maximum rate of electron transport through photosystem II (Jmax), the maximum rate of carboxylation by RuBisCO (Vcmax), and the triose phosphates utilization rate (TPU).
2.8. Growth
By weighing paper copies of the leaves and one square centimeter of paper, as well as by figuring out the link between paper mass and area, leaf area was measured allometrically. After oven drying at 70 ºC to a constant mass, these leaves were weighed with an analytical weighing balance (Model HR200, Japan). Throughout the course of the experiment, non-destructive allometric growth measurements were performed every two weeks for six months on five plants (n = 5) per treatment. The following measurements were taken: plant total height, measured from ground level to the apical shoot; stem diameter at 10 cm from ground level; crown diameter at the lower part of each plant, on the longest lower branch of the plant – branch length of the intermediate branch of the plant is considered and measured from the insertion of the branch in the central stem to the terminal bud –; number of branches present on the stem of the plant; flowering. For flowering an arbitrary scale from 0 to 3, where 0 = no flowering, 1 = low, 2 = medium and 3 = high intensity of flowering was used.
2.9. Statistical Analysis
The data displayed in tables and figures are expressed as means (5 ≤ n ≤ 10) ± standard error (SE). Statistical package, STATISTICA v10 (StatSoft Inc., OKlahoma, USA), was used to perform one and two-way analysis of variance (ANOVA) and Tukey's post hoc test at the 5% level of significance. Water status, gas exchange, and A/Ci traits were compared across four genotypes and between control and WD within each genotype using ANOVAs. SigmaPlot 11 (Systat Soft-ware, California, USA) was used for all regressions, whether they were linear or curvilinear, and significance was assessed at p < 0.05.
3. Results
3.1. Genotype Physiological Characterization
Time course of changes under WD for the four genotypes are shown in Figure 1. The A values ranged from 5 μmol m-2 s-1 in Sarchimor 4260 to 15 μmol m-2 s-1 in Cavimor ECU and Catimor ECU 02. There were notable variations among genotypes (p=0.02), with Sarchimor 4260 showing the smallest A, and Cavimor ECU and Catimor ECU 02 the biggest. The genotype Cavimor ECU had the lowest average E value, whereas Catimor ECU 02 had the greatest E value. When compared to the other genotypes, Cavimor ECU had the lowest gs. The lowest WUE was found in Sarchimor, whereas high WUE was found in Cavimor ECU genotypes (p=0.004).
3.2. Water Deficit Experiment
Water deficit negatively affected SWC, LWC and RWC among the four genotypes (Figure 2). A decline of 70% (Cavimor ECU), 77% (red Caturra), and 80% (Sarchimor 4260 and Catimor ECU 02) was noted in SWC after 29 days under WD. The reduction in SWC induced a 17 % decrease in LWC in Cavimor ECU, whereas in Sarchimor 4260, red Caturra and Catimor ECU 02, the decrease was lower, only 3%. After 29 days of WD, RWC was substantially lower in all coffee genotypes by 10–25%. The SLA ranged between 161.4 ± 8.5 cm2 g-1 (Cavimor ECU) and 200.6 ± 9.1cm2 g-1 (Sarchimor 4260); a small increase of 8-10 % in red Caturra and Sarchimor 4260 and 18 % in Cavimor ECU due to WD was observed. This could mean that leaves that are thinner would be less resistant to CO2 diffusing to the carboxylation sites. In Catimor ECU 02, SLA was unchanged by WD.
After 29 d of WD, most of the leaf gas exchange variables decreased relative to day 0 of treatment (Figure 3). Values of A diminished by 74, 59, 50 and 36 %, and gs by 53, 48, 41 and 24 % in Sarchimor 4260, red Caturra, Cavimor ECU and Catimor, respectively, whereas E decreased by 30-36 % in Sarchimor 4260, red Caturra and Cavimor ECU, and 4 % in Catimor. The WUE was significantly reduced by 61 % in Sarchimor 4260, 42 % in red Caturra and 34 % in both Cavimor ECU and Catimor ECU 02. In Sarchimor 4260, Ci increased in plants summited to WD, whereas in the other genotypes Ci was unchanged. After one day of re-irrigation, all leaf gas exchange variables in the Cavimor ECU and Catimor ECU 02 genotypes promptly recovered, but not in the Sarchimor 4260 and red Caturra.
With gs accounting for 48.8% of the variation in A, the correlation between gs and A was curved, and the regression did not cross the zero (Figure 4). It is very informative that some cultivars were capable of presenting values of A not significantly different from those in control plants albeit at much lower gs. That would reflect their higher WUE.
The A/Ci curves and their parameters did not exhibit any statistically significant differences between the control seedlings of the four genotypes under investigation (Figure 5). The greatest A at saturating intercellular [CO2] (Asat) and CE occurred in the order Sarchimor 4260 > red Caturra > Catimor ECU 02 > Cavimor ECU. Values were differentially affected by WD in the four genotypes. Values of Asat and CE declined by 57, 53, 42 and 11 %, and 32, 38, 20 and 3 % in Sarchimor 4260, red Caturra, Cavimor ECU and Catimor ECU 02, respectively. The Ls was unchanged after 29 days of WD in all genotypes, whereas Lm increased by 60, 53, 38 and 11 % in Sarchimor 4260, red Caturra, Cavimor ECU and Catimor ECU 02 genotypes, respectively.
The biochemical parameters determined from the A/Ci curves (Figure 5) showed that due to water deficit Jmax was reduced up to 56-69 % in red Caturra and Sarchimor 4260 genotypes, respectively, and 15-29 % in Cavimor ECU and Catimor ECU 02 genotypes, respectively; Vcmax was reduced in all coffee genotypes by 45-61% and TPU by 63-61% in red Caturra, Sarchimor 4260 and 21-11% Cavimor ECU and Catimor ECU 02 (Figure 5 E, F, G). A positive linear association occurred during WD between Asat and Jmax (Asat= 0.204 × Jmax -2.17; r2 = 0.78) and between CE and Vcmax (CE= 0.0005 × Vcmax - 0.065; r2 = 0.30). Only 30% of the variation in CE was explained by the Vcmax, compared to 78% by the Jmax in Asat.
3.3. Growth
In all four genotypes, there was a gradual increase over time in all growth variables evaluated (Figure 6). From February to July an increase in height was observed in Catimor ECU 02 and Cavimor ECU of 18-19 % and 20-24 % in Sarchimor 4260 and red Caturra; plants of the first two genotypes were the shortest, WD significantly decreasing height by 8.5 % in Catimor ECU 02. All genotypes flowered at six months after the start of the experiment, Cavimor ECU showing the highest flowering index, and Sarchimor 4260 the lowest; WD had a positive effect on flowering in all genotypes except for Sarchimor.
A significant increase in stem diameter was found from 47 to 67 % in all genotypes; WD caused a significant decreased of 29 % (Catimor), 23% (Sarchimor), 21% (red Caturra) and 11 % (Cavimor ECU). Similarly, an increase in crown diameter de 2.2-2.1 times in Sarchimor 4260 and red Caturra and 2.7-3.3 in Cavimor ECU and Catimor ECU 02 took place. The number and length of branches increased over time by 84 and 68 % in the four genotypes; WD caused a significant reduction in branch length of 14-23 %, this variable remaining unaffected in red Caturra. The number of internodes per branch and internode distance increased over time by 16 and 84 %, respectively. The WD reduced internode distance by 12-39 % in three of the genotypes.
4. Discussion
The findings validated our hypothesis 1 by demonstrating differences in gas exchange and growth characteristics among four genotypes of healthy, well-irrigated arabica coffee plants. Under irrigated conditions, the genotypes Cavimor ECU and Catimor ECU 02 exhibited the greatest physiological achievement, i.e., great A and WUE. Since 15 to 29 days without irrigation resulted in a significant drop in soil and leaf water status, photosynthetic variables, and growth parameters, all four genotypes were susceptible to WD. We noticed variations in genotype response to WD, indicating variations in drought tolerance. The results indicated that Catimor ECU 02 and Cavimor ECU were more resistant to WD, while red Caturra and Sarchimor 4260 were the most susceptible. The results are consistent with our hypothesis 2, which states that different arabica coffee genotypes would experience different effects on WUE due to physiological plasticity. Specifically, WUE was more reduced during WD in the Sarchimor 4260 and red Caturra genotypes than in the Cavimor ECU and Catimor ECU 02 genotypes; however, after re-irrigation for one day, WUE values rapidly regained in the Cavimor ECU and Catimor ECU 02 genotypes, but not in the Sarchimor 4260 and red Caturra genotypes.
Significant difference in water status and gas exchange performance of plantlet of the four genotypes develop under in semi-controlled conditions suggested physiological adaptability amidst genotypes. The research showed values of A and gs that are greater than those observed in 31 varieties of C. arabica and C. canephora clones evaluated in Ecuador [14,33] and comparable to those found in C. arabica in Brazil, Colombia, and Ethiopia [4,25,32]. For arabica and conilon coffee, mean values of A of 8 μmol m-2 s-1 and gs of 148 mmol m-2 s-1 have been observed in Brazil [34].
The study's gs values, which ranged from 277 to 400 mmol m-2 s-1 [14], were higher than those found in prior studies on arabica and conilon coffee (which had gs values of 108 and 148 mmol m-2 s-1, respectively [34]). The WUE observed in the study was comparable to that reported for clones of robusta and arabica coffee (2.8 and 3.8 mmol mol-1, respectively), assessed in the field and in greenhouses [14,25,32]. As SWC decreased under WD, a reduction in LWC and RWC in all four genotypes studied suggested that these were susceptible to water shortage. Coffee does not tolerate dehydration, nevertheless, as it maintains a relatively high RWC in dry conditions, making it a species that conserves water [35].
The four C. arabica genotypes under evaluation showed varied responses to WD in terms of leaf gas exchange (A, gs, E, and WUE). Similar results have been reported in arabica coffee [25,32]. In contrast, in two C. canephora clones gas exchange was unaffected by WD, although their water status was negatively affected by WD [23]. In Ecuador, it was found that in twenty-one C. arabica cultivars under WD in field conditions A, gs and WUE differed significantly among cultivars [14]. Cultivar Cavimor ECU had the highest A and WUE, maintained the values of these parameters up to 22 days of WD and showed full recovery with irrigation, suggesting that this cultivar could be selected for growth in areas subject to WD; however, further research under natural conditions is urgently required.
Results of WUE were significantly higher in Cavimor ECU and Catimor ECU 02, while the lowest were found in red Caturra. We conclude that A and WUE could be indicators of the tolerance of coffee cultivars to WD. In Sarchimor 4260 and red Caturra genotypes (sensitive to WD), A and gs were significantly reduced by water deficit more than in Catimor ECU 02 and Cavimor ECU, making the latter two more tolerant to WD. These results suggest that during the seedling stage, arabica coffee has physiological adaptability to WD. Previous reports have shown comparable outcomes with WD for arabica coffee genotypes – the sensitive Ca754 and CaJ-19 genotypes, as well as the Castillo variety, and the comparatively tolerant Ca74110 and Ca74112 [25,32]. The gas exchange of 21 genotypes of C. arabica that were assessed in field conditions decreased due to drought [14].
Gas exchange and related variables showed bimodal responses to WD. At first, as RWC decreased, A and gs declined as well, but Ci remained constant, indicating that stomata were not significantly limiting A under WD. Nevertheless, Ci under WD remained constant with an average Ci/Ca ratio of 0.77, suggesting that metabolic limitation of A became increasingly significant even if the relationship between A and gs in this study was curvilinear. This interpretation of the data is supported by the observation that after one day of re-irrigation, A was fully restored in Cavimor ECU and Catimor ECU 02 but not in Sarchimor 4260 and red Caturra. In contrast, genotypes of C. arabica from Brazil and Ecuador have previously shown significant linear relationship between A and gs [14,34], but not in conilon [34] or in robusta, indicating that in C. canephora cv. robusta coffee clones A was independent of gs [33].
Reduces in RWC, A, and gs without alterations in WUE in Catimor ECU 02 and Cavimor ECU after 29 days of WD were a general pattern that indicated coffee genotypes optimizing water use. Similarly, WUE did not change in arabica coffee in response to drought [14]. Water limitation may result in a decrease in A due of stomatal limitation, because of reductions in gs-related stomatal closure and decreases in Ci and/or through damage of metabolic process [22]. With the exception of Cavimor ECU, the majority of all four genotypes closed their stomata in response to WD, resulting in a drop in gs and a consequent avoidance of excessive water loss and reductions in A. Since WD had no effect on Ci, we do not completely rule out the possibility that metabolic variables were crucial in the regulation of A [22]. With the exception of Cavimor ECU and Catimor ECU 02, where colimitation of A by stomatal and metabolic factors occurred, values of Ls remained constant during the 29 days of WD, while Lm increased gradually in all four cultivars. This suggests that, as WD increased, biochemical and photochemical regulation of A became more important than decline in gs (stomatal closure). This implies that the regulation of A by photochemical and biochemistry became increasingly significant as WD rose, rather than stomatal closure (i.e. the reduction of gs).
All four genotypes had comparable photosynthetic capacities, which were influenced by WD in distinct ways as seen by the A/Ci curves. Control plants showed the maximum photosynthetic capacity, determined under saturating PPFD (Asat) of 24-32 μmol m-2 s-1, similar to reported values up to 30 μmol m-2 s-1 [36,37]. In Sarchimor 4260, red Caturra and Cavimor ECU, the noted declines in Asat and CE with decreasing A discernible decrease in Vcmax and Jmax, respectively, suggests that WD had a significant impact on the activity and/or quantity of RubisCO and the availability of RuBP. In contrast, only a small reduction of CE and Asat was found in Catimor ECU 02, which suggested that no metabolic process was impaired. Our estimations of biochemical parameters in the four C. arabica cultivars studied support data reported before in red Catuai and arabica coffee by [38,39] and the values of Vcmax, Jmax and TPU were higher than the results reported in Catimor [2].
Both the Catimor ECU 02 and Cavimor ECU genotypes, which partially tolerant to WD, displayed a significantly rapid and better recovery from the negative effects of WD following a single day of re-irrigation. The four arabica coffee genotypes showed notable variations in the A/Ci curve parameters; Catimor ECU 02 and Cavimor ECU showed less variation than Sarchimor 4260 and red Caturra. In addition to affecting Asat, Jmax, CE, and Vcmax, water deficit also increased Lm by 56% on average in two genotypes that were sensitive, indicating that both stomatal closure (to a lesser extent) and metabolic, photochemical, and/or biochemical limitations during WD were responsible for the reduction of A. In contrast, Ls accounted for 40% of the constraints in red Catuai grown in both full sun and shade, whereas Lm made up 30% of the constraints in the same radiation conditions [39].
Water deficit caused a reduction in some growth variables, this effect being stronger in Sarchimor 4260 and red Caturra than in Catimor ECU 02 and Cavimor ECU. Similar results, except for stem height, shoot fresh mass and dry biomass, were reported in tolerant and sensitive arabica coffee genotypes under drought [25], supporting the observation that WD reduces crop yields by affecting the plant’s vegetative growth [9].
5. Conclusions
- 5.1.
- Our findings show that Coffea arabica genotypes differ in their physiological performance, growth traits, and response to water deficit. The photosynthetic capacity of all four genotypes of arabica coffee was high. Bigger values of A and WUE were found in Cavimor ECU and Catimor ECU 02 compared to the Sarchimor 4260 and red Caturra genotypes, indicating that situations where water availability is a major constraint may be more favorable for the former genotypes. The evidence suggests that there may be significant differences in tolerance among different genotypes in coffee.
- 5.2.
- In plants of arabica coffee genotypes submitted to water deficit the stress not only affected photosynthetic capacity but also increased metabolic limitations, while stomatal limitations remained constant in the two sensitive genotypes and the resistant Cavimor ECU, indicating that both causes are responsible for the decrease in A: to a lesser degree stomatal closure and in major degree biochemical or metabolic damage during WD. In contrast, in Catimor ECU 02, CE, Vcmax, Jmax and Asat were less affected, suggesting that metabolic processes were not impaired and colimitation of A by stomatal and non-stomatal factors occurred. Nevertheless, neither type of factor's regulation of photosynthesis was an effective enough mechanism to preserve WUE, which decreased.
- 5.3.
- Although the genotypes of Cavimor ECU and Catimor ECU 02 seedlings did not fully optimize their use of water, WD had less of an impact on their growth and physiological performance than it did on Sarchimor 4260 and red Caturra. The former two genotypes with water deficit tolerance potential could be beneficial for coffee programs for developing plants in drier habitats. Further studies on physiology are required to assess photosynthetic capacity in arabica coffee seedlings and plants in Ecuador, aiming to select drought-tolerant genotypes for efficient cultivation.
Author Contributions
Conceptualization, Methodology, Data Acquisition, formal analysis, Writing, Reviewing and Editing, Funding Acquisition, W.T.; Methodology, Data Acquisition, Writing and Editing, D.W.L.; Methodology, Data Acquisition, supervision, V.H.R.C.; Conceptualization, Writing-Reviewing and Editing, A.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Prometeo Project of the Ministry of Higher Education, Science, Technology and Innovation (SENESCYT) of the Republic of Ecuador for their sponsorship and funding in this work, specifically the fellowship program for W.T. and to the UTLVTE research department.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author/s.
Acknowledgments
We thank E. Avila-Lovera for critically reading the manuscript. The authors are grateful to the Prometeo Project SENESCYT of the Republic of Ecuador for their sponsorship and funding in this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Caporaso, N.; Whitworth, M.B.; Grebby, S., Fisk, I.D. Non-destructive analysis of sucrose, caffeine and trigonelline on single green coffee beans by hyperspectral imaging. Food Res. Internat. 2018, 106, 193–203. [CrossRef]
- Xiao, Z.; Ma, G.; Bai, X.; Li, J.; Zhao, M.; Su, L., Zhou, H. The influence of leaf anatomical traits on photosynthesis in Catimor type Arabica coffee. Bever. Plant Res 2024, 4, e002 . [CrossRef]
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Change 2019, 152, 167–178. [CrossRef]
- Semedo, J.N.; Rodrigues, A.P.; Lidon, F.C.; Pais, I.P.; Marques, I.; Gouveia, D.; Armengaud, J.; Silva, M.J.; Martins, S.; Semedo, M.C.; Dubberstein, D.; Partelli, F.L.; Reboredo, F.H.; Scotti-Campos, P.; Ribeiro-Barros, A.I.; DaMatta, F.M.; Ramalho, J.C.. Intrinsic non-stomatal resilience to drought of the photosynthetic apparatus in Coffea spp. is strengthened by elevated air [CO2]. Tree Physiol. 2021, 41, 708–727. [CrossRef]
- ICO, International Coffee Organization (2021) https://icocoffee.org/.
- ICO, International Coffee Organization (2024) https://icocoffee.org/.
- Duicela, L. Café robusta: Producción y poscosecha. In: Humus ESPAM MFL. Ecuador: Calceta-Manabí-Ecuador, 2017, p. 292. ISBN: 9789942859587. [In Spanish].
- Cavatte, P.C.; Rodríguez-López, N.F.; Martins, S.C.V.; Mattos, M.S.; Sanglard, L.M.; DaMatta, F.M. Functional analysis of the relative growth rate, chemical composition, construction and maintenance costs, and the payback time of Coffea arabica L. leaves in response to light and water availability. J. Exp. Bot. 2012, 63, 3071–3082. [CrossRef]
- DaMatta, F.M. Exploring drought tolerance in coffee: a physiological approach with some insights for plant breeding. Braz. J. Plant Physiol. 2004, 16, 1–6. [CrossRef]
- DaMatta, F.M.; Ramalho, J.D. Impacts of drought and temperature stress on coffee physiology and production: a review. Braz. J. Plant Physiol. 2006, 18, 55–81. [CrossRef]
- DaMatta, F.M.; Godoy, A.G.; Menezes-Silva, P.E.; Martins, S.C.V.; Sanglard, L.M.V.; Morais, L.E.; Torre-Neto, A.; Ghini, R. Sustained enhancement of photosynthesis in coffee trees grown under free-air CO2 enrichment conditions: disentangling the contributions of stomatal, mesophyll, and biochemical limitations. J. Exp. Bot. 2016, 167, 341–352. [CrossRef]
- Martins, M.Q.; Rodrigues, W.P.; Fortunato, A.S.; Leitão, A.E.; Rodrigues, A.P.; Pais, I.P.; Martins, L.D.; Silva, M.J.; Rebodero, F.M.; Partelli, F.L.; Campostrini, E.; Tomaz, M.A.; Campos, P.S.; Barros, A.I.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C. Protective response mechanisms to heat stress in interaction with high [CO2] Conditions in Coffea spp. Front. Plant Sci. 2016, 7, 1–18. [CrossRef]
- Rodríguez-López, N.F.; Martins, S.C.V.; Cavatte, P.C.; Silva, P.E.M.; Morais, L.E.; Pereira, L.F.; Reis, J.V.; Ávila, R.T.; Godoy, A.G.; Lavinski, A.O.; DaMatta, F.M. Morphological and physiological acclimations of coffee seedlings to growth over a range of fixed or changing light supplies. Environ. Exp. Bot. 2014, 102, 1–10. [CrossRef]
- Tezara, W.; Duicela, G.L.A.; Reynel Chila, V.H.; Nazareno Ortiz, R.; Bolaños Ortega, M.J. Seasonal changes in gas exchange and yield of 21 genotypes of Coffea arabica. Bot. Sci. 2022,100, 1000–1013. [CrossRef]
- Duicela, L. Productividad y estabilidad ambiental de clones de café robusta en distintas localidades cafetaleras del Ecuador. PhD. Thesis. 2021. Universidad del Zulia. [In Spanish]. [CrossRef]
- Gornall, J., Betts, R.; Burke, E.; Clark, R.; Camp, J.; Willet, K,; Wiltshire, A. Implications of climate change for agricultural productivity in the early twenty–first century. Philos. Transact, Royal Soc. 2010, 365, 2973–2989. [CrossRef]
- DaMatta, F.M.; Grandis, A.; Arenque, B.C.; Buckeridge, M.S. Impacts of climate changes on crop physiology and food quality. Food Res. Int. 2010, 43, 1814–1823. [CrossRef]
- Semedo, J.N.; Rodrigues, W.P.; Dubberstein, D.; Martins, M.Q.; Martins, L.D.; Pais, I.P.; Rodrigues, A.P.; Leitão, A.E.; Partelli, F.L.; Campostrini, E.; Tomaz, M.A.; Reboredo, F.H.; Scotti-Campos, P.; Ribeiro-Barros, A.I.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C. Coffee responses to drought, warming and high [CO2] in a context of future climate change scenarios. In: Alves, F., Leal Filho, W., Azeiteiro, U. (eds) Theory and Practice of Climate Adaptation. Climate Change Management. Springer, Cham. Clim. Change Manage. 2018, 465–477. [CrossRef]
- Rodrigues, A.P.; Pais, I.P.; Leitão, A.E.; Dubberstein, D.; Lidon, F.C.; Marques, I.; Semedo, J.N.; Rakocevic, M.; Scotti-Campos, P.; Campostrini, E.; Rodrigues, W.P.; Simões-Costa, M.C.; Reboredo, F.H.; Partelli, F.L.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Ramalho, J.C. Uncovering the wide protective responses in Coffea spp. leaves to single and superimposed exposure of warming and severe water deficit. Front. Plant Sci. 2024,14, 1320552. 10.3389/fpls.2023.1320552.
- Merga, D.; Beksisa, L. Mechanisms of drought tolerance in coffee (Coffea arabica L.): Implication for genetic improvement program: review. American J. BioSci. 2023, 11, 63–70. 10.11648/j.ajbio.20231103.12.
- Läderach, P.; Ramirez-Villegas, J.; Navarro-Racines, C.; Zelaya, C.; Martinez-Valle, A.; Jarvis, A. Climate change adaptation of coffee production in space and time. Clim. Change 2017, 141, 47–62. [CrossRef]
- Lawlor, D.W.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a critical evaluation and integration of mechanisms and processes. Ann. Bot. 2009, 103, 561–579. [CrossRef]
- DaMatta, F.M.; Chaves, A.R.M.; Pinheiro, H.A.; Ducatti, C.; Loureiro, M.E. Drought tolerance of two field-grown clones of Coffea canephora. Plant Sci. 2003. 164, 111–117. [CrossRef]
- Pinheiro, H.A. 2004. Physiological and morphological adaptations as associated with drought tolerance in robusta coffee (Coffea canephora Pierre var. kouillou). PhD. Thesis. Universidade Federal de Viçosa.
- Chekol, H.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dabrowska, G.B.; Degu, A. Drought stress responses in arabica coffee genotypes: physiological and metabolic insights. Plants 2024, 13(6), 828; [CrossRef]
- INAMHI, (Red de Estaciones Automáticas Hidrometeorológicas) (2016) http://186.42.174.236/InamhiEmas/.
- Jones, H.G. Plants and microclimate. A quantitative approach to environmental plant physiology. Cambridge University Press, Cambridge, 1992, pp. 465.
- Tezara, W.; Torres-Domínguez, T. S.; Loyaga, D.W.; Nazareno-Ortiz, R.; Reynel-Chila, V.H.; Bolaños Ortega, M.J. Photosynthetic activity of oil palm (Elaeis guineensis) and interspecific hybrid genotypes (Elaeis oleifera × Elaeis guineensis), and response of hybrids to water déficit. Sci. Hortic. 2021, 287, 110263, . [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu Rev. Plant Physiol. 1982, 33, 317–345. [CrossRef]
- Jacob, J.; Lawlor, D.W. Stomatal and mesophyll limitations of photosynthesis in phosphate deficient sunflower, maize and wheat plants. J. Exp. Bot. 1991, 42, 1003–1011. [CrossRef]
- Sharkey, T.D. What gas exchange data can tell us about photosynthesis? Plant Cell Environ. 2015, 39, 1161–1163. [CrossRef]
- Reyes-Herrera, D.F.; Sánchez-Reinoso, A.D.; Lombardini, L.; Restrepo-Díaz, H. Physiological responses of coffee (Coffea arabica L.) plants to biochar application under water deficit conditions. Not. Bot. Horti. Agrobo. 2023, 51(3), 12873. [CrossRef]
- Tezara Fernández, W.; Mendoza Cortez, P.J.; Loyaga Guerrero, W.D.; Reynel Chila, V.H.; Bolaños Ortega, M.J. Capacidad fotosintética de 15 clones de café robusta (Coffea canephora) en Esmeraldas, Ecuador. Rev. ESPAMCIENCIA 2020, 11, 19–27. [In Spanish]. [CrossRef]
- DaMatta, F.M.; Ronchi, C.P.; Maestri, M.; Barros, R.S. Ecophysiology of coffee growth and production. Braz. J. Plant Physiol. 2007, 19, 485–510. [CrossRef]
- DaMatta, F.M.; Maestri, M.; Barros, R.S.; Regazzi, A.J. Water relations of coffee leaves (Coffea arabica and C. canephora) in response to drought. J. Horticult. Sci. 1993, 68, 741–746. [CrossRef]
- Campostrini, E.; Maestri, M. Photosynthetic potential of five genotypes of Coffea canephora Pierre. Rev. Bras. Fisiol. Veg. 1998,10, 13–18.
- DaMatta, F.M.; Loos, R.A.; Rodrigues, R.; Barros, R.S. Actual and potential photosynthetic rates of tropical crop species. Braz. J. Plant. Physiol. 2001, 13, 24–32. [CrossRef]
- Martins, S.C.V.; Galmés, J.G.; Molins, A.; DaMatta, F.M. Improving the estimation of mesophyll conductance: on the role of electron transport rate correction and respiration. J. Exp. Bot. 2013, 64, 3285–3298. 10.1093/jxb/ert168. Epub 2013 Jul 5.
- Martins, S.C.V.; Galmés, J.; Cavatte, P.C.; Pereira, L.F.; Ventrella, M.C.; DaMatta, F.M. Understanding the low photosynthetic rates of sun and shade coffee leaves: bridging the gap on the relative roles of hydraulic, diffusive and biochemical constraints to photosynthesis. PLoS ONE 2014, 9, e95571. [CrossRef]
Figure 1.
Gas exchange parameters of four arabica coffee genotypes. A, E) Net photosynthetic rate; B, F) transpiration rate; C, G) stomatal conductance; D, H) instantaneous water use efficiency. The vertical bars represent the mean of five plants ± SE. Distinct letters indicate significant variations in genotypes and/or sampling time (p < 0.05). Every bar shows a distinct sampling in February (black), March (red), April (green) and May (yellow) 2017. Panels E, D, F and G show the means of all samples for each genotype.
Figure 1.
Gas exchange parameters of four arabica coffee genotypes. A, E) Net photosynthetic rate; B, F) transpiration rate; C, G) stomatal conductance; D, H) instantaneous water use efficiency. The vertical bars represent the mean of five plants ± SE. Distinct letters indicate significant variations in genotypes and/or sampling time (p < 0.05). Every bar shows a distinct sampling in February (black), March (red), April (green) and May (yellow) 2017. Panels E, D, F and G show the means of all samples for each genotype.
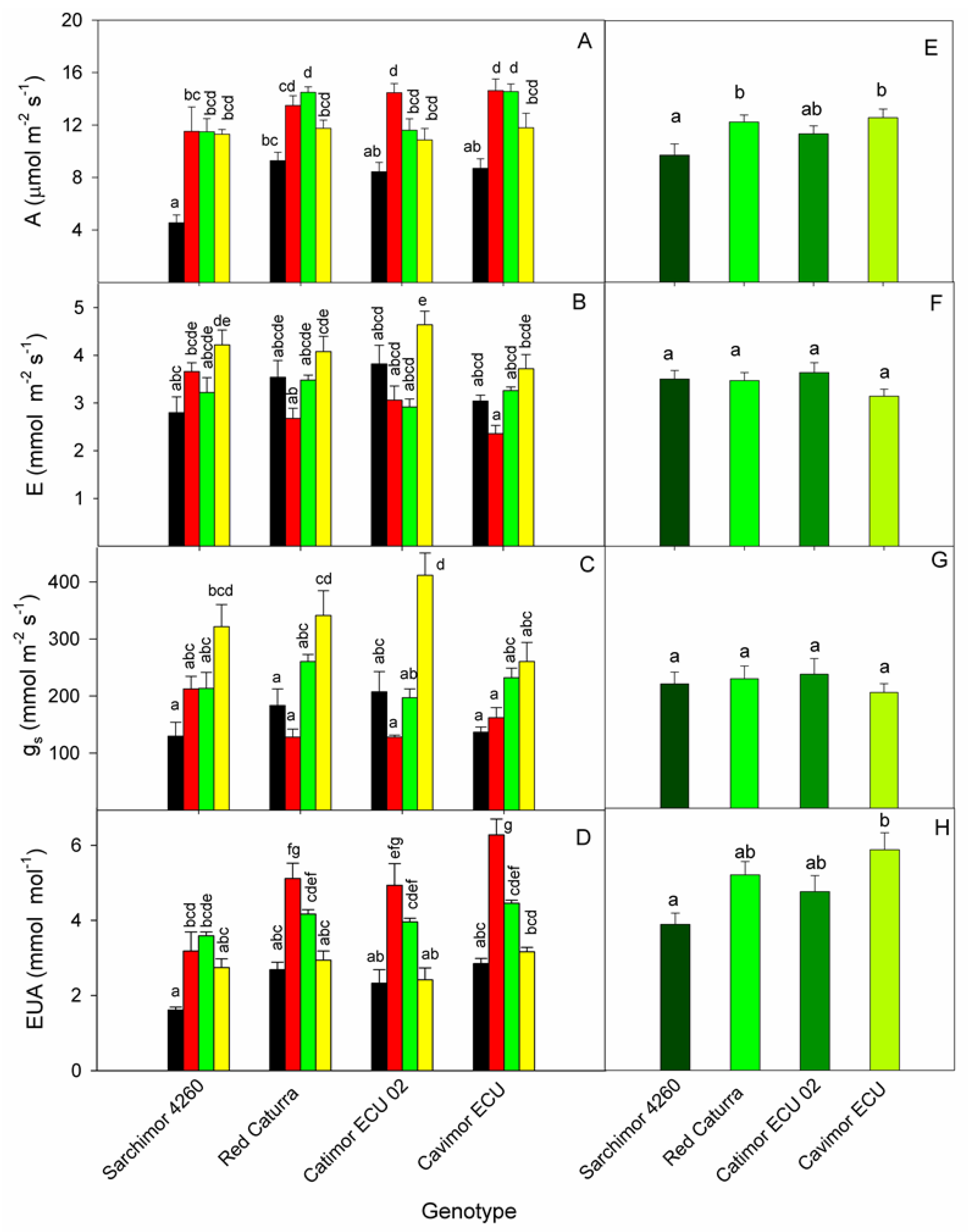
Figure 2.
Effects of water deficit duration on four coffee genotype plants' leaves: A-D, soil water content (SWC); B-H, leaf water content (LWC); I-L, relative water content (RWC); and M-P, specific leaf area (SLA), in both WD (open circles) and control (closed circles) plants. Genotype name is shown above the uppermost panels. The data represent means (6 ≤ n ≤ 10) ± SE. For each parameter at p < 0.05, distinct letters denote significant variations between WD and control seedlings. Measurements taken a day after re-irrigation are indicated by arrows.
Figure 2.
Effects of water deficit duration on four coffee genotype plants' leaves: A-D, soil water content (SWC); B-H, leaf water content (LWC); I-L, relative water content (RWC); and M-P, specific leaf area (SLA), in both WD (open circles) and control (closed circles) plants. Genotype name is shown above the uppermost panels. The data represent means (6 ≤ n ≤ 10) ± SE. For each parameter at p < 0.05, distinct letters denote significant variations between WD and control seedlings. Measurements taken a day after re-irrigation are indicated by arrows.
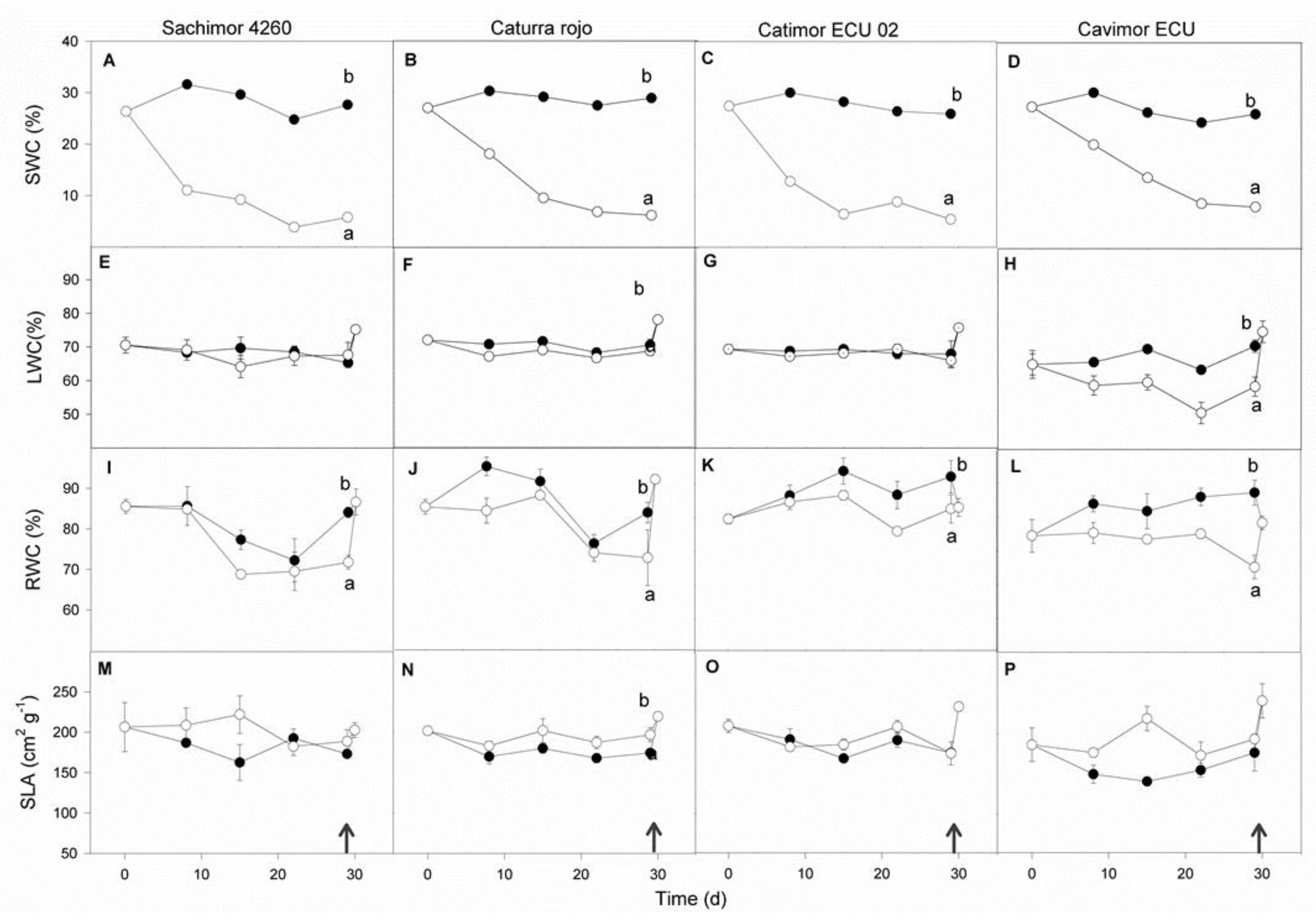
Figure 3.
Effects of water deficit duration on four coffee genotype plants' leaves: control (closed circles) and WD (open circles) plants. Net photosynthetic rate (A-D), transpiration rate (E-H), stomatal conductance (I-L), intercellular CO2 concentration (M-P), and water use efficiency (Q-T). Genotype name shown in the uppermost panels. The data are means (n = 5) ± SE. For each parameter at p < 0.05, distinct letters denote significant variations between WD and control seedlings. Measurements taken a day after re-irrigation are indicated by arrows.
Figure 3.
Effects of water deficit duration on four coffee genotype plants' leaves: control (closed circles) and WD (open circles) plants. Net photosynthetic rate (A-D), transpiration rate (E-H), stomatal conductance (I-L), intercellular CO2 concentration (M-P), and water use efficiency (Q-T). Genotype name shown in the uppermost panels. The data are means (n = 5) ± SE. For each parameter at p < 0.05, distinct letters denote significant variations between WD and control seedlings. Measurements taken a day after re-irrigation are indicated by arrows.
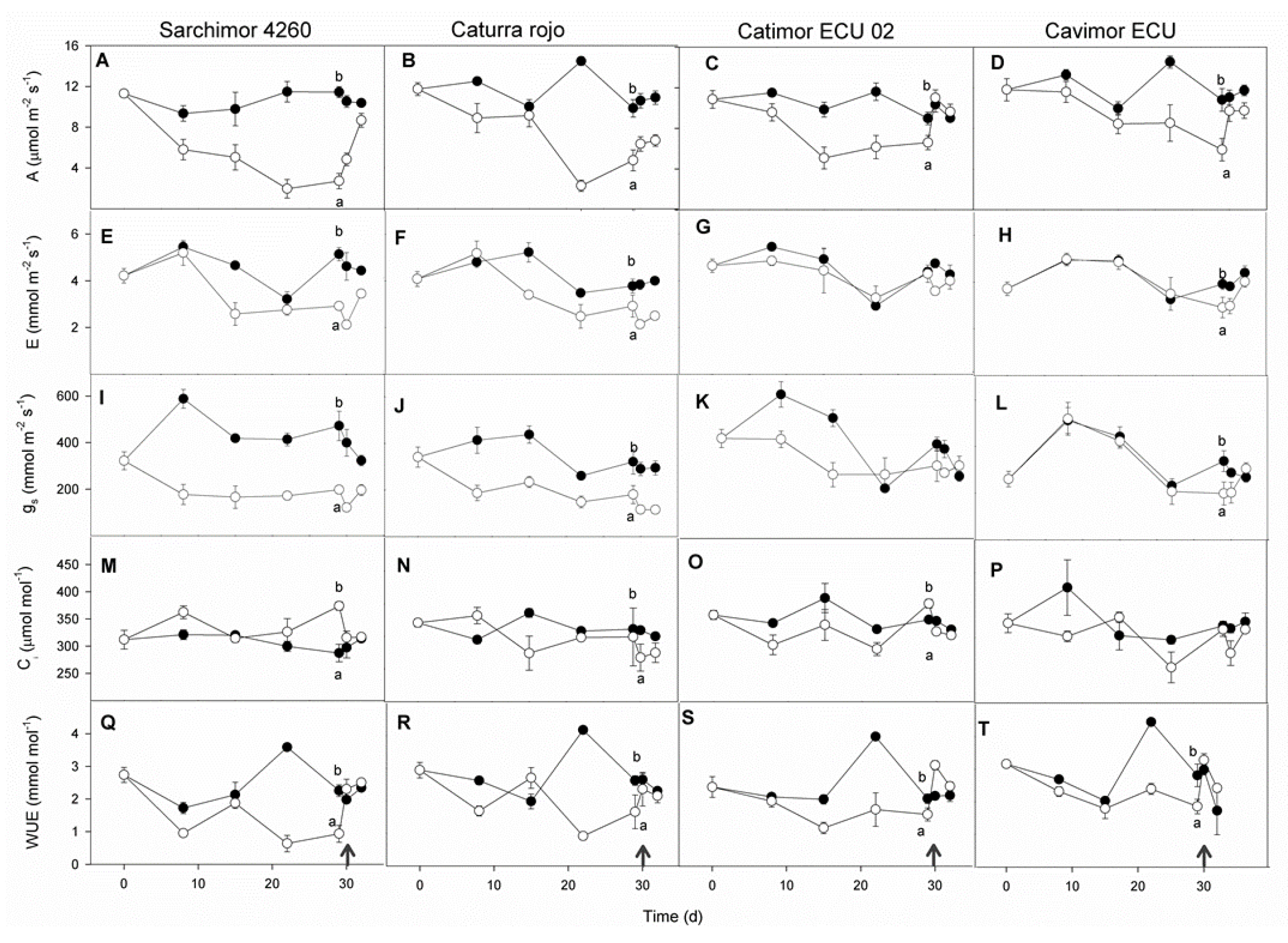
Figure 4.
The net photosynthetic rate (A) and stomata conductance (gs) of four genotypes of arabica coffee in the leaves of plants experiencing a water deficit (open circles) and control plants (closed circles) are correlated. The genotypes examined were Sarchimor 4260 (●), Red Caturra (■), Catimor ECU02 (▲) and Cavimor ECU (♦). The data are means (n = 5) ± SE. The regression was significant at p < 0.0001.
Figure 4.
The net photosynthetic rate (A) and stomata conductance (gs) of four genotypes of arabica coffee in the leaves of plants experiencing a water deficit (open circles) and control plants (closed circles) are correlated. The genotypes examined were Sarchimor 4260 (●), Red Caturra (■), Catimor ECU02 (▲) and Cavimor ECU (♦). The data are means (n = 5) ± SE. The regression was significant at p < 0.0001.
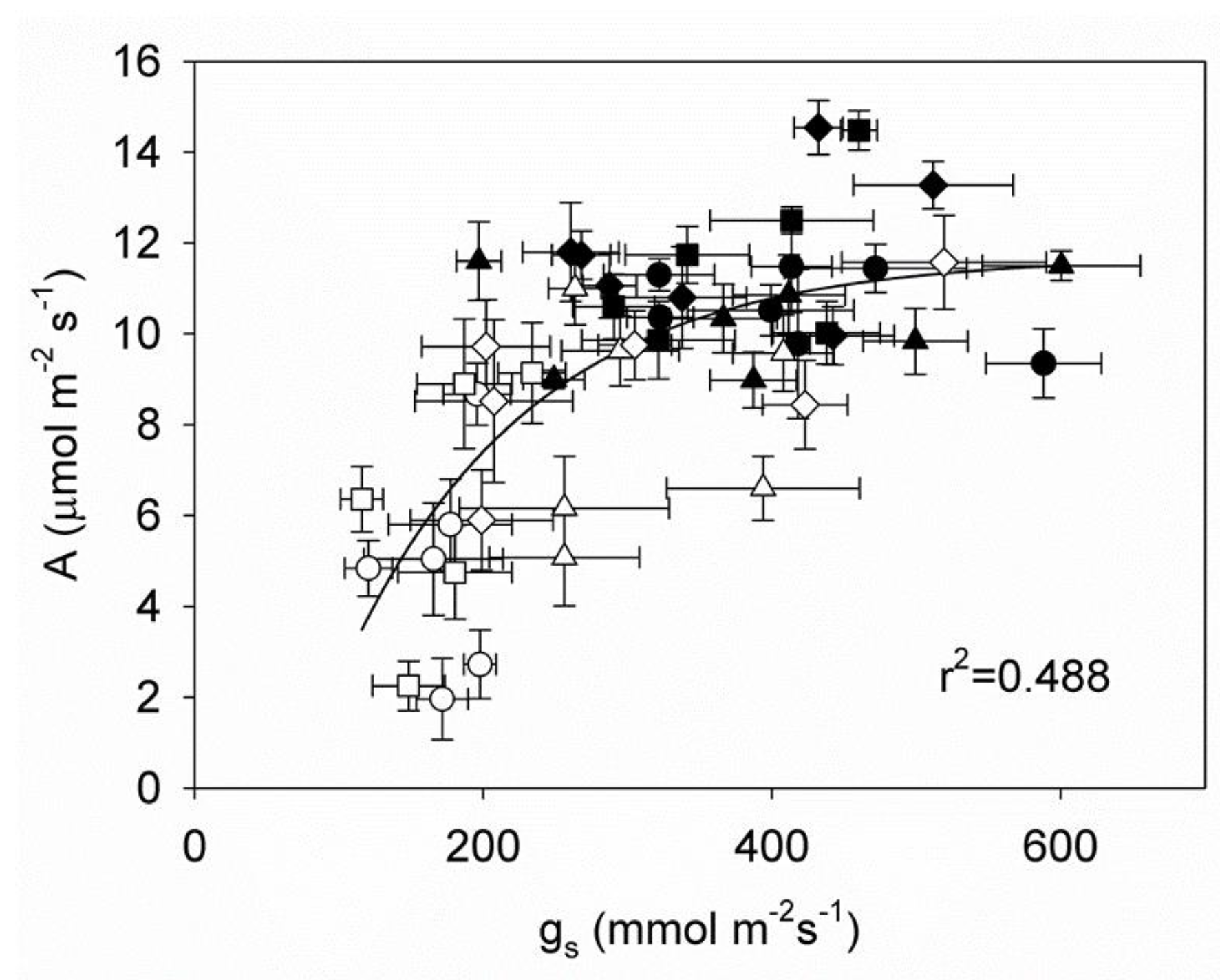
Figure 5.
Effects of water deficit duration in parameters calculated from A/Ci curves on four coffee genotype plants' leaves: A, Ci-saturated photosynthetic rate; B, carboxylation efficiency; C, relative stomatal limitation; D, relative mesophyll limitation; E, maximum rate of electron transport through PSII ; F, maximum rate of RuBisCO carboxylation and G, triose phosphates utilization rate. The genotypes examined were Sarchimor 4260 (●), Red Caturra (■), Catimor ECU02 (▲) and Cavimor ECU (♦). When a value exceeds the symbol size, standard errors are presented. The data are means (n = 4) ± SE.
Figure 5.
Effects of water deficit duration in parameters calculated from A/Ci curves on four coffee genotype plants' leaves: A, Ci-saturated photosynthetic rate; B, carboxylation efficiency; C, relative stomatal limitation; D, relative mesophyll limitation; E, maximum rate of electron transport through PSII ; F, maximum rate of RuBisCO carboxylation and G, triose phosphates utilization rate. The genotypes examined were Sarchimor 4260 (●), Red Caturra (■), Catimor ECU02 (▲) and Cavimor ECU (♦). When a value exceeds the symbol size, standard errors are presented. The data are means (n = 4) ± SE.
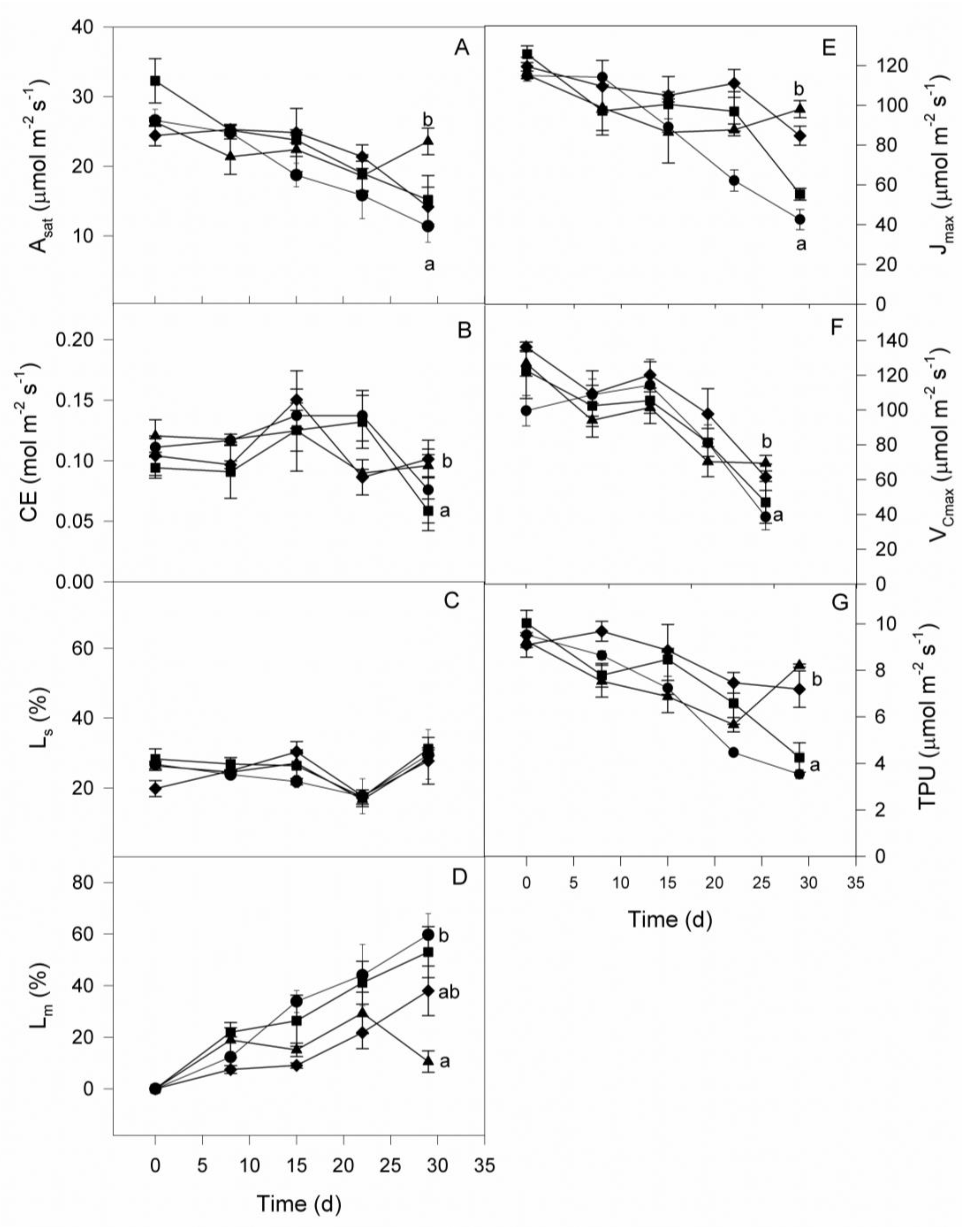
Figure 6.
Changes over time of growth variables in plants of four arabica coffee genotypes in: A) height; B) flowering index; C) stem diameter; D) crown diameter; E) number of branches; F) branch length; G) total number of nodes per branch, and H) distance between internodes. Control plants measured on February (black bars), March (red bars), April (green bars), May (yellow bars), June (blue bars) and July (pink bars); plants under water deficit for 15 d (turquoise) and 29 d (grey). Cultivar names are indicated on the abscissa. An asterisk indicates significant differences between control plants in July and those subjected to water deficit (p< 0.05).
Figure 6.
Changes over time of growth variables in plants of four arabica coffee genotypes in: A) height; B) flowering index; C) stem diameter; D) crown diameter; E) number of branches; F) branch length; G) total number of nodes per branch, and H) distance between internodes. Control plants measured on February (black bars), March (red bars), April (green bars), May (yellow bars), June (blue bars) and July (pink bars); plants under water deficit for 15 d (turquoise) and 29 d (grey). Cultivar names are indicated on the abscissa. An asterisk indicates significant differences between control plants in July and those subjected to water deficit (p< 0.05).
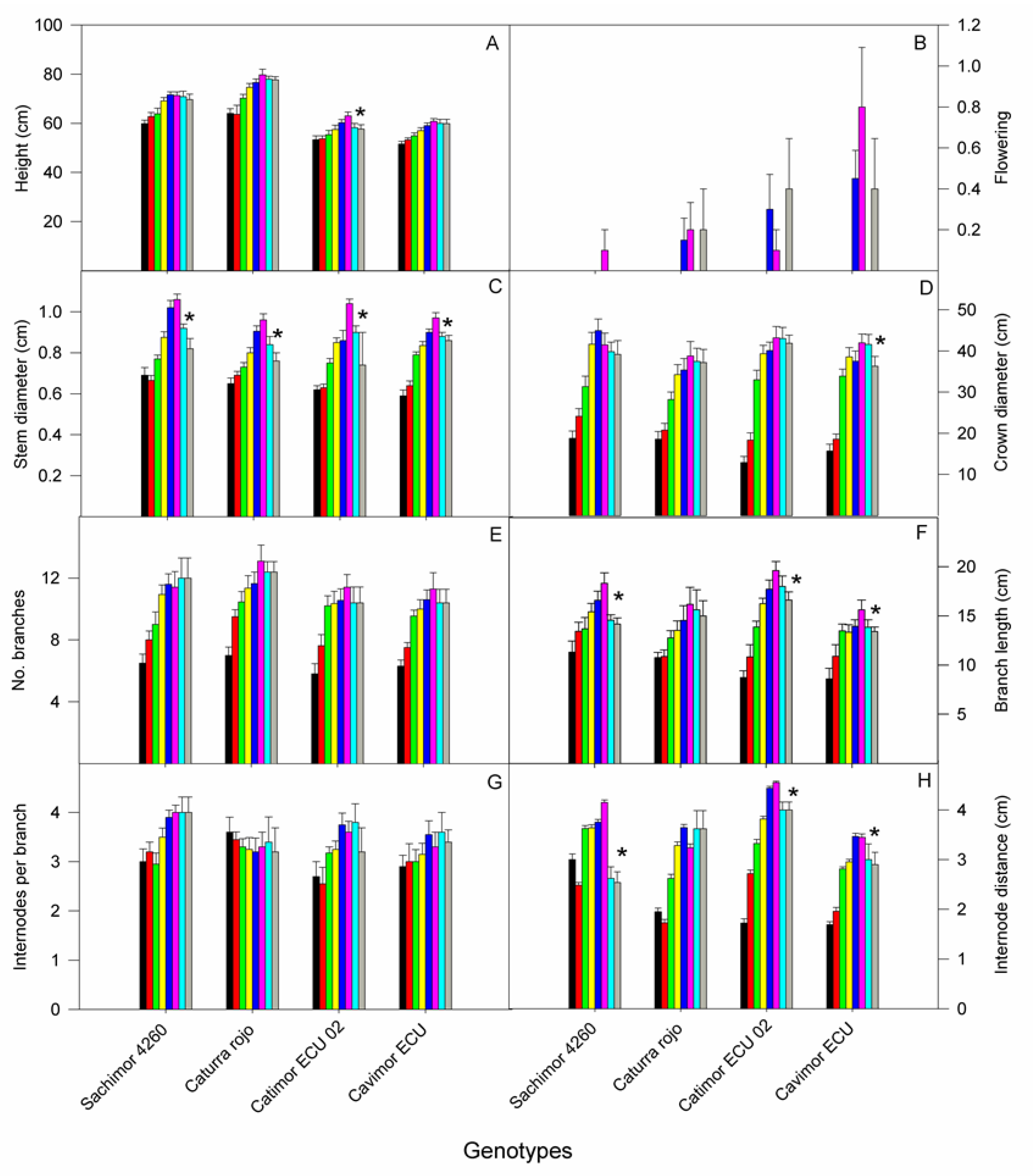

Table 1.
Variations in the daily values of the following microclimatic parameters, which were measured inside the shade house on ten separate sample days: ambient CO2 concentration (Ca), photosynthetic photon flux density (PPFD), air temperature (Ta), relative humidity (RH) and air vapor pressure deficit (ΔW). Values are mean ± SE (n = 10 days).
Table 1.
Variations in the daily values of the following microclimatic parameters, which were measured inside the shade house on ten separate sample days: ambient CO2 concentration (Ca), photosynthetic photon flux density (PPFD), air temperature (Ta), relative humidity (RH) and air vapor pressure deficit (ΔW). Values are mean ± SE (n = 10 days).
| Hour of day | Ca (µmol mol-1) |
PPFD (µmol m-2 s-1) |
Ta (°C) |
RH (%) |
Dw (kPa) |
|---|---|---|---|---|---|
| 08:00 | 510 ± 24 | 30 ± 3 | 24.8 ± 0.4 | 82.8 ± 0.4 | 0.53 ± 0.02 |
| 13:00 | 414 ± 7 | 260 ± 70 | 33.1 ± 0.7 | 53.7 ± 0.7 | 2.3 ± 0.08 |
| 17:00 | 418 ± 5 | 50 ± 6 | 28.4 ± 0.6 | 66.2 ± 0.6 | 1.3 ± 0.05 |
Table 2.
In Esmeraldas, Ecuador assessed the genotypes of C. arabica, including their names, attributes, genetic origins, and responses to coffee rust.
Table 2.
In Esmeraldas, Ecuador assessed the genotypes of C. arabica, including their names, attributes, genetic origins, and responses to coffee rust.
| Name | Traits and genetic origin |
| Sarchimor 4260 | Sarchimor from CIFC based on the cross of Hybrid Villa Sarchi × Timor Hybrid. Various genetic lines of Sarchimor are being evaluated in Ecuador, and to date have excellent agronomic, productive and resistance to coffee rust. |
| Red Caturra | Caturra are mutants of the coffee variety Bourbon, native to Brazil. This variety is considered to have a wide range of adaptability, high production, good agronomic and organoleptic characteristics, but susceptible to coffee rust. |
| Cavimor ECU | Developed at the Coffee Rust Research Center (CIFC, Oeiras, Portugal) based on the crossing of Hybrid Catuaí × Catimor. Several Cavimor genetic lines currently being evaluated showed resistant to coffee rust. |
| Catimor ECU 02 | The CIFC has developed the hybrid Catimor, the result of the cross between Caturra and Timor. This hybrid shows great genetic variability and resistance to coffee rust. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



