Submitted:
02 July 2024
Posted:
03 July 2024
You are already at the latest version
Abstract
ABSTRACTCaffeine is known to impair both growth and development in the model social amoeba Dictyostelium discoideum. However, little is known about the transcriptional regulation in response to caffeine treatment in Dictyostelium. In yeast cells, caffeine mimics the effect of rapamycin in inhibiting mTORC1 activity. Here, we investigated whether changes in growth and development of Dictyostelium in the presence of caffeine or rapamycin are mediated by mTORC1 and/or mTORC2. Results: Treatment of wild type cells with caffeine or rapamycin resulted in severely impaired growth and in the formation of small aggregates in comparison to untreated cells. rip3‾ cells (lacking a mTORC2 component) and lst8‾cells (lacking a component of both mTOR complexes) showed higher and slower growth rates, respectively, in comparison to parental AX3 cells. Parental AX3 and rip3‾ cells displayed similar sensitivity to rapamycin and caffeine, while lst8‾ cells were more sensitive to these compounds. When developed in the presence of caffeine or rapamycin, AX3 cells formed aggregates after a long delay. Caffeine did not alter the morphology of small rip3‾ mounds and lst8‾ cells continued to remain solitary. However, in the presence of 1µM rapamycin rip3‾cells failed to form mounds while AX3 cells streamed and formed aggregates. Complementation of lst8‾ cells with the wild type lst8 gene rescued the aggregation phenotype. cDNA microarray analysis to uncover the spectrum of differentially regulated genes in response to caffeine treatment revealed 104 and 193 differentially expressed genes at 3-and 6-hours of development, respectively, in comparison to control cells. After 6 hours of starvation in the presence of caffeine, 25 genes encoding ribosomal proteins and 4 encoding elongation factors were up regulated. Furthermore, we identified that upon caffeine treatment at the 3- and 6-hours’ time points, 12 (10 up and 2 down) and 21 (13 up and 8 down) genes, respectively, encoding proteins involved in early development. Conclusion: We conclude that caffeine and rapamycin have similar effects in decreasing the growth rate and in reducing the aggregate size of D. discoideum. Studies with lst8‾ and rip3‾ cells suggest that these compounds affect cell growth and aggregation by acting on both mTOR complexes, mTORC1 and mTORC2. Analysis of the differentially expressed genes after caffeine treatment suggests a key role of these genes in growth and aggregation.
Keywords:
Dictyostelium
; caffeine
; rapamycin
; mTORC1 and mTORC2
; cell growth
; microarray
Introduction
Dictyostelium discoideum is a single-celled protist that enters development on sensing a shortage of nutrients in its environment. In response to starvation, a large number of cells chemotax leading to the formation of a facultative multicellular organism [1]. As a simple system, Dictyostelium is subject to considerable genetic analysis to unravel major questions with respect to developmental fate and multicellular organization. Such a unique life cycle requires precise regulation of key signaling pathways, cell proliferation and differentiation.
The target of rapamycin (mTOR) is found in two conserved complexes, namely mTOR complex-1 (mTORC1) and mTOR complex-2 (mTORC2) in Dictyostelium. mTORC1 is composed of mTOR, Raptor and Lst8 (lethal with SEC13 protein 8) and mTORC2 includes mTOR, Rictor, Rip3 (ras-interacting protein3) and Lst8. mTORC1 and mTORC2 have different functions but mTOR and Lst8 are common components in these two complexes [2,3,4]. mTORC1, a protein kinase that regulates cell growth and cell mass in response to nutrient levels, stress, and hormones [4], is highly conserved in yeast, amoebae and humans [2,3,5,6]. Processes like ribosome biogenesis, cell proliferation and autophagy are regulated by mTORC1 [7,8,9,10] while mTORC2 regulates chemotaxis, cAMP signal relay, cytoskeleton organization and development [3,11]. cAMP relay signal is oscillatory which is also regulated through mTORC2 activity [12]. Mutations in mTORC1 are lethal [9]. lst8‾ cells fail to aggregate due to impaired chemotaxis [11]. Rip3 is required for both the synthesis and relay of cAMP as well as chemotaxis [13]. rip-3‾ cells form mounds that do not develop further [13].
Caffeine affects a wide variety of cellular processes in several organisms [14,15,16,17,18,19,20]. In Dictyostelium, caffeine severely impairs mechanisms regulating growth, cell size, cAMP signal relay, cell adhesion, cytosolic glucose levels and cell counting mechanism [16,19,20]. Rapamycin, an immuno-suppressant drug, binds to the FK506 binding protein FKBP12, and when the complex binds the FRB domain of mTOR, TOR kinase activity is inhibited. In yeast, caffeine and rapamycin have similar effects with respect to the regulation of the activity of mTORC1 [2,3,14,21].
In mammals, caffeine is known to inhibit mTOR activity (IC50 = 0.4 mM) by directly acting on the phosphatidylinositol 3-kinase domain (PI3Kc) of mTOR [22]. In yeast, mTOR is the major caffeine target. Single mutations in these domains are less resistant to caffeine than double mutations, one in FRB domain and another in PI3Kc kinase domain [21].
We examined whether Dictyostelium growth and development are regulated by caffeine and rapamycin in AX3 (wild type) and mutants of the mTOR complex subunits Lst8 and Rip3. We found that both caffeine and rapamycin severely impair growth and development. To uncover the set of genes with altered expression in response to caffeine treatment, gene expression profiling was carried out in the presence of caffeine for 3 and 6 hours of development along with untreated control cells. Treatment with caffeine for 3 hours resulted in 62 up-regulated and 42 down-regulated genes reflecting early aggregation stages. The corresponding numbers at 6 hours of development were 142 up-regulated and 59 down-regulated genes, respectively. Interestingly, the differential expression depends strongly on the time of caffeine exposure, for 6 hours of treatment, resulted in a larger number of differentially expressed genes than 3 hours incubation time.
Methods
Cell Culture
AX3 Dictyostelium cells were grown in Cosson HL5 media (14.3 g bactopeptone, 7.15 g yeast extract, 18 g maltose monohydrate, 3.6 mM Na2HPO4 and KH2PO4) with penicillin (100 units/ml) and streptomycin (100 μg/ml) at 220C either in petridishes or in shaking suspension (150 rpm; [as described in [16,23]. lst8‾ and rip3‾ cells were grown in Cosson HL5 media containing 5 μg/ml of blasticidin in the presence of penicillin and streptomycin (concentrations see above) to prevent bacterial contamination.
Growth Kinetics
Cell division measurements were carried out as described in Jaiswal et al., 2012 [16]. Briefly, 0.1x106 cells/ml were inoculated in test tubes containing 10 ml of Cosson HL5 media with appropriate concentrations of either rapamycin or caffeine and incubated at 220C with constant shaking (180 rpm). Cell growth was monitored by counting the number of cells with a haemocytometer at regular intervals. We carried out five independent experiments to assay the cell division kinetics and the values represent mean ± SEM.
Analysis of aggregation patterns
Axenically grown Dictyostelium cells were harvested, washed with KK2 buffer and plated at a density of 1x106 cells/cm2 on non-nutrient agar plates [KK2 buffer (2.25 g KH2PO4 and 0.67 g K2HPO4 per liter H2O, pH 6.4) containing 15 g agar per liter] containing the specified concentrations of caffeine. Aggregation patterns were monitored either with a NikonTM SMZ1000 or a NikonTM eclipse 80i microscope.
Transformation
Axenically grown Dictyostelium cells were washed twice with electroporation buffer (EP buffer: 10 mM K2HPO4, 10 mM KH2PO4 and 50 mM sucrose) and 2x107 cells were resuspended in 100 µl EP++ buffer (EP buffer + 1 mM MgSO4, 1 mM NaHCO3, 1 mM ATP and 1 µM CaCl2) containing 10 µg of vector DNA. lst8‾cells were complemented with appropriate vector carrying lst8 driven by constitutive actin15 promoter. The cell suspension with the DNA was transferred into a chilled 2-mm cuvette and electroporated at 300V, 2ms, 5 square wave pulses in 5-sec. intervals using a BTX ECM830 electroporator [23]. The transformants were selected in the presence of 10 µg G418 in Cosson HL-5 media.
RNA sample preparation for Microarray and qPCR
Axenically grown cells were harvested during exponential growth phase and traces of media were washed off with KK2 buffer by centrifugation at 1,000 rpm for 5 minutes. Caffeine treated or untreated cells were starved in Sorensen buffer at a density of 1x107 cells/ml in an Erlenmeyer flask at 220C with constant shaking at 150 rpm. Total RNA was isolated at either 3 or 6 hours of starvation using the QiagenRNAeasy mini preparation kit [24] with slight modification in the protocol. RNA integrity was checked by running a denaturing MOPS-formaldehyde gel and with the Agilent bioanalyzer.
Quantitative PCR (RT-PCR)
We validated the microarray data by performing qRT-PCR. The RNA sample was prepared as described above. cDNA was prepared using the Go ScriptTM Reverse Transcription system (Promega-USA). cDNA was mixed with SYBR green and qPCR master mix (Promega-USA), and qRT-PCR was performed using the Applied Biosystems 7500 Real Time PCR machine [25]. The list of primers used in this study is given in supplementary Table 1.
Microarray analysis
Microarray expression profiling and data analysis were performed as described [26]. The cDNA microarrays comprise of a non-redundant set of 5,423 EST clones, appropriate positive and negative controls as well as partial sequences of 450 selected genes. All the probes were spotted in duplicates. Details of the microarray design are accessible at the Gene Expression Omnibus (GEO; accession number GPL1972). For the time course experiments with caffeine, RNA was isolated from treated and untreated control cells, reverse transcribed, labelled and used for hybridization. We analyzed six microarrays for treated and untreated samples from each time point with labeled cDNAs that were derived from RNAs from three independent cultures. Dye swaps were used for RNA labeling from each isolation. Scanning was performed with the Scan Array® 4000 XL confocal laser scanner, signals were quantified with Scan Array Express 3.0 (Perkin Elmer Life Sciences, Wellesley, USA) and the fluorescence ratios were normalized by LOWESS-normalization using the software R 1.6.2. Differentially expressed genes were identified with the program Significance Analysis of Microarrays (SAM) [27]. SAM calculates the score for every gene with a t-statistic modified for the use with microarray data. The higher the score, the more reliable the differential expression of the reported gene.
Results
mTORC1 and mTORC2 modulate Dictyostelium cell growth
We assayed cell division kinetics for mutants impaired in components of TOR complexes, i.e. lst8‾ (mTORC1 and mTORC2) and rip3‾ (mTORC2 only), in nutrient-rich medium. We found that the growth rate of rip3‾ cells was significantly higher than that of parental AX3 cells, while for lst8‾ cells the growth rate was lower (Figure 1).
Rapamycin and caffeine inhibit cell growth.
Both rapamycin and caffeine modulate the expression of similar sets of genes in yeast [14] and are also known to inhibit cell growth. We first performed cell division kinetics of wild type AX3 cells in either the absence or presence of rapamycin or caffeine. In the presence of 50 nM rapamycin and 2 mM caffeine, the cell numbers were about 50% lower after 72 hours as compared to untreated controls (Figure 2A). Increasing rapamycin concentration to 500 nM resulted in further decrease in cell number to about 40% in comparison to untreated cells. Addition of 5 mM caffeine completely arrested cell growth (Figure 2A). In summary, both compounds, caffeine and rapamycin, reduced the growth rate of Dictyostelium AX3 cells.
Further, to examine whether the growth defect in the presence of caffeine or rapamycin is due to mTORC1, mTORC2 or both TOR complexes, we tested growth of lst8‾ and rip3‾ cells. lst8‾ cells were sensitive to rapamycin, while rip3‾ cells were resistant to rapamycin as compared to its parental AX3 strains (Figure 2B and C). This result suggests that rapamycin action on mTORC1/mTORC2 is independent of Lst8, while Rip3 of mTORC2 is needed to mediate the inhibitory effect of rapamycin on cell growth. Caffeine treatment resulted in a severe growth defect in both lst8‾ and rip3‾ cells in comparison to AX3 cells (Figure 2B and C). After 72 hours, the relative number of lst8‾ cells with 2 mM and 5 mM caffeine dropped drastically to ~20 and ~15 %, respectively and similarly, the relative cell number of rip3‾ cells was reduced to ~5 and ~3 % respectively, (Figure 2B and C). Impaired growth of lst8‾ and rip3‾ cells in the presence of caffeine suggests that caffeine’s action on mTORC1 and mTORC2 is independent of Lst8. Similarly, caffeine’s action on mTORC2 is independent of Rip3. Alternatively, caffeine may have one or more additional targets which mediate its effects on growth in lst8‾ and rip3‾ cells.
Caffeine and rapamycin inhibit aggregate formation
We treated vegetative AX3 cells with different concentrations of either caffeine or rapamycin and monitored development at different time points. In controls, the first sign of aggregation was seen after 6 hours (not shown). At 9 hours, fully developed aggregates were observed, whereas at 12 hours, migrating slugs were seen, and at 24 hours fully developed fruiting bodies were observed (Figure 3A, control). Increasing caffeine concentrations from 2 to 3 mM, resulted in a gradual delay in development and the aggregate size also progressively decreased (Figure 3A). At 5 mM caffeine, there was no aggregation and all the cells remained solitary (Figure 3A). Rapamycin also inhibited aggregate formation in a dose-dependent manner. Increasing the concentrations of rapamycin also resulted in progressive inhibition of aggregation (Figure 3B). In the presence of 1 µM rapamycin, all the cells managed to form small aggregates with streams after 24 hours. In the same time period, fruiting bodies were observed in controls with no rapamycin added (Figure 3B). At 3 and 5 µM concentrations of rapamycin, there was no aggregation and cells remained solitary, resembling the effect of 5 mM caffeine (Figure 3). This result showed that caffeine and rapamycin have similar effects on development.
Loss of activity of both TOR complexes results in small sized aggregates
Inhibition of aggregate formation in the presence of caffeine and rapamycin can be due to the action of these compounds on mTORC1 and/or mTORC2. To analyse whether caffeine and rapamycin affect development through mTORC1, mTORC2 or both mTOR complexes, we made further use of the lst8‾ and rip3‾ cells and examined their aggregation response in the presence of these compounds.
The Lst8 knockout shows impaired chemotaxis and aggregation. lst8‾ cells were developed for 9 hours in 2mM caffeine, and no change was found in their single-celled phenotype, while control cells formed aggregates under these conditions (Figure 4A). 5mM caffeine treatment of AX3 cells resulted in solitary cells not participating in aggregation, a phenotype similar to the one of lst8‾ cells (compare Figure 3A and 4A). Next, we performed complementation studies to demonstrate that the inability of lst8‾ cells to form aggregates strictly depends on the absence of Lst8. When lst8‾ cells were complemented with lst8-OE vector, they formed aggregates similar to wild type cells. We tested the effect of 2 mM caffeine on these cells (Δlst8+lst8) at two different densities (1x105 cells/cm2 and 1x106 cells/cm2) and observed tight streams and small sized aggregates at both cell densities in the presence of caffeine that resembled aggregation phenotypes of AX3 cells treated with caffeine (compare Figure 4A and 4B). This suggests that Lst8, as a component of mTORC1 or mTORC2 or both complexes, is essential for aggregation. To examine whether caffeine can affect development by targeting mTORC2, rip3‾ cells (component of mTORC2 only) were allowed to develop in the presence of caffeine or rapamycin and were monitored during aggregation. rip3‾ cells aggregated without streaming to form a mound, which did not progress to form slugs and fruiting bodies [13,28]. Caffeine had no effect on the aggregation response in rip3‾ cells (Figure 4C) but 1 µm rapamycin further inhibited aggregation in rip3‾ cells that remained solitary without entering further morphogenetic stages (Figure 4C). Since rapamycin exerted an additional effect on rip3‾ cells but not caffeine this result suggests that, during differentiation, rapamycin acts on mTORC1 while caffeine may target both mTORC1 and mTORC2.
Microarray analyses reveal differentially expressed genes in response to caffeine
Most of the genes involved in early development are known to be expressed in the first 6 hours after onset of starvation [29]. Therefore, we performed microarray analyses of cells starved for 3 and 6 hours in the presence and absence (control) of caffeine. For each time point, we hybridized six microarrays and obtained up to twelve measurements for each probe. The software “Significance Analysis of Microarrays” revealed a number of differentially regulated genes [27]. Since small changes in expression cannot be confirmed with alternative methods and the biological impact of such small changes is unclear, we used an additional threshold for the fold change of 1.4 for further analysis of microarray results. We randomly selected three of the significantly differentially regulated genes to validate the microarray results by real time PCR and confirmed their differential expression (Figure 5).
Using the above criteria we found 104 differentially regulated genes, of which, 42 were down-regulated and 62 up-regulated at 3 hours of starvation and 201 genes of which, 59 were down-regulated and 142 were up-regulated at 6 hours of starvation in response to caffeine treatment in comparison to starved untreated cells. At both time points, a larger number of genes were up-regulated than down-regulated and at the 6-hours’ time point, the time when aggregation starts, approximately twice as many genes were differentially regulated compared to the 3-hour time point. We next compared the differentially regulated genes between the 3- and 6-hour time points. A Venn diagram shows the common up- and down-regulated genes among the differentially expressed genes at 3 and 6 hours of development upon caffeine treatment (Figure 6). Surprisingly, of the entire set of differentially expressed genes at the two time points, there were no common down-regulated genes and but 21common up-regulated genes (Table 1). These genes encode proteins involved in signal transduction and development, cellular biogenesis and metabolism. In addition there were 8 genes that were up-regulated at t3 and down-regulated at t6 and 10 genes that were up-regulated at t6 and down-regulated at t3 (Supplementary table 2). All differentially expressed genes with a fold-change of 1.4 or higher are listed in the supplementary Tables 3 and 4.
Next, we classified all the differentially expressed genes based on their biological function (Figure 7). At 3 hours, the largest fractions of differentially expressed genes of which most were upregulated are involved in “signal transduction, cellular biogenesis and multicellular organization” (20 of 104 genes) (Figure 7A). At 6 hours of development, genes belonging to the same three functional categories are also predominantly up regulated (17 of 201 genes). An additional prominent category at this time point is “translation” (33 of 201 genes) and nearly all of these genes are up-regulated (Figure 7B). Another prominent category at both time points is “metabolism” (13 of 104 genes and 31 of 201 genes, respectively). Again, most of these genes are up regulated. In summary, caffeine appears to inhibit development by modulating the expression of a number of genes involved in signal transduction, translation, metabolism, cellular biogenesis and multicellular organization.
We next looked at the set of differentially regulated genes for candidates downstream of mTORC1. There were small changes in the expression levels of genes related to ribosome biogenesis and translation at 3 hours of development in the presence of caffeine. However, after 6 hours in the presence of caffeine, 29 genes were upregulated including 25 genes encoding ribosomal proteins (Table 2). In addition, we found 3 genes encoding elongation factors, namely efa1B, efa1G (elongation factor 1 beta and gamma) and efbA (elongation factor 2). Treatment of starving cells with caffeine for 6 hours also increased the expression of nacA (nascent polypeptide-associated complex alpha subunit) (Table 2). The up-regulation of these genes is surprising because caffeine actually slows down the growth of cells. Their up-regulation might cause the delay in aggregation and might be responsible for the developmental block at higher caffeine concentrations.
The function of target of rapamycin complex-2 (mTORC2) in Dictyostelium is in cAMP signal relay, chemotaxis and aggregation. Hence, we particularly looked for altered expression of genes involved in chemotaxis and cAMP signal relay, cell adhesion, and density sensing mechanism and/or are regulated by activity of mTORC1 or mTORC2. At 3 and 6 hours of caffeine treatment we found 12 (10 up and 2 down) and 21 (13 up and 8 down) genes, respectively, whose protein products are involved in signal transduction, cell density sensing and cell adhesion (Table 3). For example, there were 3 genes encoding cell adhesion molecules which were down-regulated after 6 hours of caffeine treatment, namely csbB (contact site-B), cad-A and cad2 (calcium-dependent cell adhesion molecule 1 and 2). We also found several genes essential for cAMP signalling, pdsA (extracellular cAMP phosphodiesterase), pdiA (cAMP phosphodiesterase inhibitor), cAR1 (cAMP receptor 1), cAR3 (cAMP receptor 3), gpaD (G-protein alpha 4), gpaI (G-protein alpha 9) and pkbA (activates TOR; protein kinase Akt1/PKB alpha). In addition, 5 genes involved in cell density sensing, ctnA (countin A), cfaD (countin associated factor), cf60 (component of countin factor complex) and smlA (small A) were differentially regulated. In conclusion, in response to caffeine treatment a substantial number of genes involved in differentiation and development (regulated by activity of mTORC2) of Dictyostelium were differentially regulated in comparison to untreated control cells.
Discussion
The mTORC1 complex plays a crucial role in the regulation of cell growth, cell size and Growth to Developmental Transition. mTORC1 defines gore gene sets for Developmental Switch [2,3,6]. In Dictyostelium it was shown that activation of mTORC1 was involved in controlling cell growth. TOR complex 1 and 2 integrate signals arising from nutrients and cellular energy state to regulate growth. The targets of mTORC1 phosphorylation include the protein translation regulators S6kinase (S6K) and 4E-BP1. The phosphorylated form of S6K activates ribosomal protein S6, which stimulates protein synthesis [2,3,6]. Also, inactivation of mTORC1 through raptor deletion suppresses growth [11]. mTOR kinase domain phosphorylation at S2159 and T2164, modulates the interactions of mTOR with raptor and PRAS40 leading to increased intrinsic mTORC1 kinase activity promoting biochemical signaling, cell growth, and cell cycle progression [30]. Increasing rapamycin and caffeine concentrations led to a progressive decrease in growth rates of wild-type cells. Our results with rapamycin are consistent with earlier observation of this drug on the growth of Dictyostelium [11,13,28]. It was suggested that rapamycin acts on cell growth by inhibiting mTORC1 [2,3,6,11]. To further investigate the possible target(s) of rapamycin and caffeine we made use of the lst8 and rip3 knock-out mutants. Lst8 is a component of both TOR complexes while Rip3 is only present in mTORC2. It was previously shown that cells deficient for Raptor, an essential subunit of mTORC1, have a reduced growth rate [11]. Interestingly, lst8‾ cells had a reduced growth rate in comparison to AX3 while the growth rate of rip3‾ cells was increased. The faster growth rate of rip3‾ cells suggests that mTORC2 somehow has an inhibitory effect on cell growth. Possibly, there is crosstalk between mTORC1 and mTORC2 in cells and mTORC2 is inhibitory to the activity of mTORC1. Alternatively, mTORC2 may inhibit cell growth independently ofmTORC1. mTORC1 is known to activate cell growth and lst8‾ cells show decreased growth rate, which may be due to impaired activity of both mTORC1 and mTORC2 complexes. We treated AX3 and the two mutants, lst8‾ and rip3‾ cells, with increasing concentrations of rapamycin or caffeine and found that the compounds slowed cell growth in a concentration-dependent manner (Figure 2A). Interestingly, lst8‾cells were more sensitive to both compounds than wild-type AX3 cells (Figure 2A and B). Since Lst8 is a component of both TOR complexes, this result suggests that either mTORC1 and/or mTORC2 partially regulate cell growth in the absence of Lst8 or alternatively, both compounds have one or more additional cellular targets. rip3‾ cells on the other hand were more resistant to rapamycin than AX3 and lst8‾ cells. This again suggests that mTORC2 has a negative effect on cell growth, possibly by acting onmTORC1. In contrast, the effect of caffeine on growth of rip3‾cells was similar to lst8‾ cells and even stronger than on AX3. Thus, at the used concentrations caffeine is either more effective in inhibiting mTORC1 and/or mTORC2, or as described above it may have one or multiple additional molecular targets such as an inhibition of PDE1 (cAMP phosphodiesterase 1) and GABAAR antagonism [15] to elicit these effects. Deletion of components shared between mTORC1 and mTORC2 (e.g. TOR1/2 and Lst8) displayed hypersensitivity at higher concentration of caffeine [21]. In Dictyostelium, TOR is common to both mTORC1 and mTORC2 complexes, but yeast cells have separate TOR components i.e. tor1p and tor2p for their TOR complexes mTORC1 and mTORC2. Yeast cells lacking tor1p are resistant to caffeine [21].
FKBP12 (FK506 binding protein-12), a component of mTORC1, is required for mTORC1 kinase activity [9,31] and FKBP12 mutant cells are insensitive to 500 nM rapamycin, confirming that, as in all other organisms, FKBP12 is essential for rapamycin action [2,11]. Our studies show reduced sensitivity of rip3‾ cells (a component of mTORC2) to rapamycin suggesting that mTORC2 may somehow affect cell proliferation by inhibiting mTORC1 activity.
We also show that, with increasing caffeine concentrations, the aggregate size became progressively smaller (Figure 3). Interestingly, when wild type cells were allowed to develop in the presence of rapamycin, we observed a similar phenotype to the one seen in the presence of caffeine. Rapamycin is known to slow down cell growth by inactivating mTORC1, and the inhibition of aggregate formation upon rapamycin treatment suggests the involvement of mTORC1 in regulating aggregate formation. Since the effects of caffeine and rapamycin were similar, it implies that caffeine may also target mTORC1, which might cause the defect in differentiation and development.
We have carried out a large scale cDNA microarray transcriptome profiling of Dictyostelium at 3 and 6 hours post starvation in the presence of caffeine and identified a number of genes involved in translation and cAMP signaling. After 6 hours treatment with caffeine, the up-regulated genes included 25 genes encoding ribosomal proteins and 4 genes encoding elongation factors, which are required for protein synthesis and may be downstream of mTORC1 pathway. Since aggregates formed in the presence of caffeine were small, the increased expression of ribosomal genes and elongation factors may be correlated with the processes regulating aggregate size (Table 3). The mTORC2 complex regulates a number of developmental events by controlling cAMP signal relay and chemotaxis [3,13,28]. This is reflected by the differentially expressed genes involved in the regulation of chemotaxis and cAMP signal relay (Table 3). Since, caffeine induces DNA damage, its role in innate immunity can be pursued, in future [32].
Conclusions
We show that caffeine and rapamycin have similar effects in decreasing the growth rate and in reducing the aggregate size of D. discoideum. Studies with lst8‾ and rip3‾ cells suggest that these compounds affect cell growth and aggregation by acting on both TOR complexes, mTORC1 and mTORC2. The DNA microarray analysis revealed that the differentially expressed genes are involved in signal transduction, translation, metabolism, cellular biogenesis and multicellular organization. The differential expression of a subpopulation of these genes is likely due to the inhibition of the TOR complexes.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
References
- Bonner, J.T.; Princeton University Press: Princeton, 1967.
- Jaiswal, P.; Kimmel, A.R. mTORC1/AMPK responses define a core gene set for developmental cell fate switching. BMC Biology 2019, 17, 58. [CrossRef]
- Jaiswal, P.; Majithia, A.; Rosel, D.; Liao, X.; Khurana, T.; Kimmel, A. Integrated actions of mTOR complexes 1 and 2 for growth and development of Dictyostelium. The International Journal of Developmental Biology 2019, 63, 521-527.
- Soulard, A.; Cohen, A.; Hall, M.N. TOR signaling in invertebrates. Curr Opin Cell Biol 2009, 21, 825-836. [CrossRef]
- De Virgilio, C.; Loewith, R. The TOR signalling network from yeast to man. Int J Biochem Cell Biol 2006, 38, 1476-1481. [CrossRef]
- Jaiswal, P.K.; Kimmel, A.R. Nutrient/Starvation sensing for Reciprocal mTORC1/AMPK response in Dictyostelium, at the junction between Growth and Development. The FASEB Journal 2018, 32, lb141-lb141. [CrossRef]
- Powers, T.; Dilova, I.; Chen, C.Y.; Wedaman, K. Yeast TOR signaling: a mechanism for metabolic regulation. Curr Top Microbiol Immunol 2004, 279, 39-51. [CrossRef]
- Mayer, C.; Grummt, I. Ribosome biogenesis and cell growth: mTOR coordinates transcription by all three classes of nuclear RNA polymerases. Oncogene 2006, 25, 6384-6391. [CrossRef]
- Kuranda, K.; Leberre, V.; Sokol, S.; Palamarczyk, G.; François, J. Investigating the caffeine effects in the yeast Saccharomyces cerevisiae brings new insights into the connection between TOR, PKC and Ras/cAMP signalling pathways. Molecular Microbiology 2006, 61, 1147-1166. [CrossRef]
- Saiki, S.; Sasazawa, Y.; Imamichi, Y.; Kawajiri, S.; Fujimaki, T.; Tanida, I.; Kobayashi, H.; Sato, F.; Sato, S.; Ishikawa, K.; et al. Caffeine induces apoptosis by enhancement of autophagy via PI3K/Akt/mTOR/p70S6K inhibition. Autophagy 2011, 7, 176-187. [CrossRef]
- Rosel, D.; Khurana, T.; Majithia, A.; Huang, X.; Bhandari, R.; Kimmel, A.R. TOR complex 2 (TORC2) in Dictyostelium suppresses phagocytic nutrient capture independently of TORC1-mediated nutrient sensing. J Cell Sci 2012, 125, 37-48. [CrossRef]
- Jaiswal, P.; Meena, N.P.; Chang, F.-S.; Liao, X.-H.; Kim, L.; Kimmel, A.R. An integrated, cross-regulation pathway model involving activating/adaptive and feed-forward/feed-back loops for directed oscillatory cAMP signal-relay/response during the development of Dictyostelium</i>. Frontiers in cell and developmental biology 2023, 11, 1263316. [CrossRef]
- Lee, S.; Comer, F.I.; Sasaki, A.; McLeod, I.X.; Duong, Y.; Okumura, K.; Yates, J.R., 3rd; Parent, C.A.; Firtel, R.A. TOR complex 2 integrates cell movement during chemotaxis and signal relay in Dictyostelium. Mol Biol Cell 2005, 16, 4572-4583. [CrossRef]
- Liu, L.; Parent, C.A. Review series: TOR kinase complexes and cell migration. J Cell Biol 2011, 194, 815-824. [CrossRef]
- Yu, L.; Coelho, J.E.; Zhang, X.; Fu, Y.; Tillman, A.; Karaoz, U.; Fredholm, B.B.; Weng, Z.; Chen, J.F. Uncovering multiple molecular targets for caffeine using a drug target validation strategy combining A 2A receptor knockout mice with microarray profiling. Physiol Genomics 2009, 37, 199-210. [CrossRef]
- Jaiswal, P.; Soldati, T.; Thewes, S.; Baskar, R. Regulation of aggregate size and pattern by adenosine and caffeine in cellular slime molds. BMC Developmental Biology 2012, 12, 5. [CrossRef]
- Jaiswal, P.; Singh, S.P.; Aiyar, P.; Akkali, R.; Baskar, R. Regulation of multiple tip formation by caffeine in cellular slime molds. BMC Developmental Biology 2012, 12, 26. [CrossRef]
- van Haastert, P.J.; De Wit, R.J.; Konijn, T.M. Antagonists of chemoattractants reveal separate receptors for cAMP, folic acid and pterin in Dictyostelium. Exp Cell Res 1982, 140, 453-456. [CrossRef]
- Gonzalez, C.; Klein, G.; Satre, M. Caffeine, an inhibitor of endocytosis in Dictyostelium discoideum amoebae. J Cell Physiol 1990, 144, 408-415. [CrossRef]
- Brenner, M.; Thoms, S.D. Caffeine blocks activation of cyclic AMP synthesis in Dictyostelium discoideum. Dev Biol 1984, 101, 136-146. [CrossRef]
- Reinke, A.; Chen, J.C.Y.; Aronova, S.; Powers, T. Caffeine Targets TOR Complex I and Provides Evidence for a Regulatory Link between the FRB and Kinase Domains of Tor1p*. Journal of Biological Chemistry 2006, 281, 31616-31626. [CrossRef]
- Foukas, L.C.; Daniele, N.; Ktori, C.; Anderson, K.E.; Jensen, J.; Shepherd, P.R. Direct Effects of Caffeine and Theophylline on p110δ and Other Phosphoinositide 3-Kinases: DIFFERENTIAL EFFECTS ON LIPID KINASE AND PROTEIN KINASE ACTIVITIES*. Journal of Biological Chemistry 2002, 277, 37124-37130. [CrossRef]
- Singh, S.P.; Dhakshinamoorthy, R.; Jaiswal, P.; Schmidt, S.; Thewes, S.; Baskar, R. The thyroxine inactivating gene, type III deiodinase, suppresses multiple signaling centers in Dictyostelium discoideum. Developmental Biology 2014, 396, 256-268. [CrossRef]
- Meena, N.P.; Jaiswal, P.; Chang, F.-S.; Brzostowski, J.; Kimmel, A.R. DPF is a cell-density sensing factor, with cell-autonomous and non-autonomous functions during Dictyostelium growth and development. BMC Biology 2019, 17, 97. [CrossRef]
- Mehto, S.; Jena, K.K.; Yadav, R.; Priyadarsini, S.; Samal, P.; Krishna, S.; Dhar, K.; Jain, A.; Chauhan, N.R.; Murmu, K.C.; et al. Selective autophagy of RIPosomes maintains innate immune homeostasis during bacterial infection. The EMBO Journal 2022, 41, e111289. [CrossRef]
- Farbrother, P.; Wagner, C.; Na, J.; Tunggal, B.; Morio, T.; Urushihara, H.; Tanaka, Y.; Schleicher, M.; Steinert, M.; Eichinger, L. Dictyostelium transcriptional host cell response upon infection with Legionella. Cell Microbiol 2006, 8, 438-456. [CrossRef]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc Natl Acad Sci U S A 2001, 98, 5116-5121. [CrossRef]
- Lee, S.; Parent, C.A.; Insall, R.; Firtel, R.A. A novel Ras-interacting protein required for chemotaxis and cyclic adenosine monophosphate signal relay in Dictyostelium. Mol Biol Cell 1999, 10, 2829-2845. [CrossRef]
- Wu, L.; Hansen, D.; Franke, J.; Kessin, R.H.; Podgorski, G.J. Regulation of Dictyostelium early development genes in signal transduction mutants. Dev Biol 1995, 171, 149-158. [CrossRef]
- Ekim, B.; Magnuson, B.; Acosta-Jaquez, H.A.; Keller, J.A.; Feener, E.P.; Fingar, D.C. mTOR kinase domain phosphorylation promotes mTORC1 signaling, cell growth, and cell cycle progression. Mol Cell Biol 2011, 31, 2787-2801. [CrossRef]
- Vilella-Bach, M.; Nuzzi, P.; Fang, Y.; Chen, J. The FKBP12-rapamycin-binding domain is required for FKBP12-rapamycin-associated protein kinase activity and G1 progression. J Biol Chem 1999, 274, 4266-4272. [CrossRef]
- Chauhan, S.; Jena, K.K.; Mehto, S.; Chauhan, N.R.; Sahu, R.; Dhar, K.; Yadav, R.; Krishna, S.; Jaiswal, P.; Chauhan, S. Innate immunity and inflammophagy: balancing the defence and immune homeostasis. The FEBS Journal 2022, 289, 4112-4131. [CrossRef]
- Urushihara, H.; Morio, T.; Saito, T.; Kohara, Y.; Koriki, E.; Ochiai, H.; Maeda, M.; Williams, J.G.; Takeuchi, I.; Tanaka, Y. Analyses of cDNAs from growth and slug stages of Dictyostelium discoideum. Nucleic Acids Res 2004, 32, 1647-1653. [CrossRef]
Figure 1.
Growth rate of AX3, rip3‾ and lst8‾cells. 0.1x106cells/ml were inoculated in 10 ml HL5 media and cell growth was monitored for 72 hours. Aliquots were taken every twelve hours in the first 48 hours and a final sample after 72 hours. The values represent the mean ± SEM from 5 independent experiments.
Figure 1.
Growth rate of AX3, rip3‾ and lst8‾cells. 0.1x106cells/ml were inoculated in 10 ml HL5 media and cell growth was monitored for 72 hours. Aliquots were taken every twelve hours in the first 48 hours and a final sample after 72 hours. The values represent the mean ± SEM from 5 independent experiments.
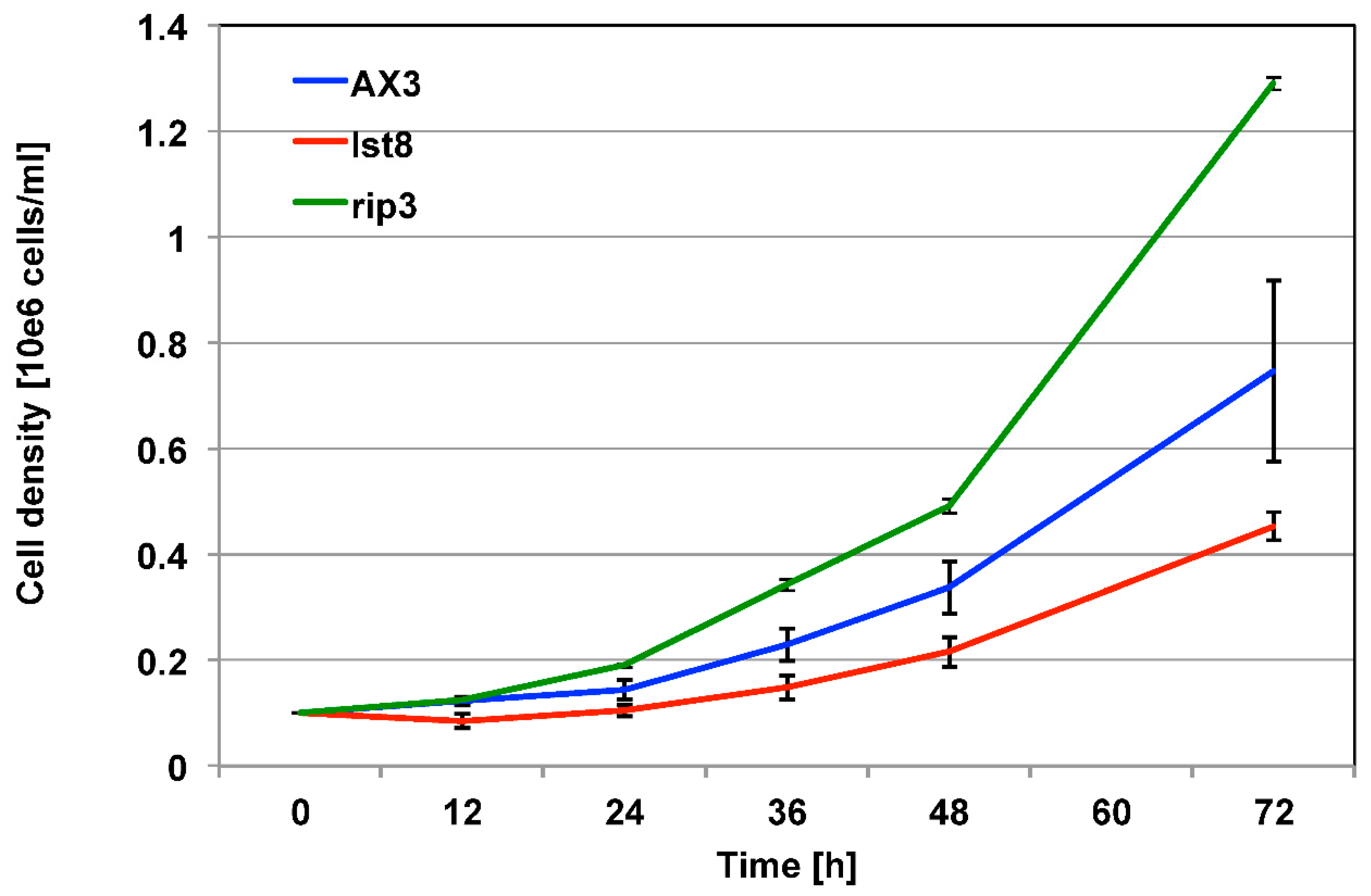
Figure 2.
The effect of caffeine and rapamycin on AX3 (A), rip3‾ (B) and lst8‾(C) cells. Cells were inoculated and cell growth was monitored for 72 hours. The highest cell number in each experiment after 72 hours was set to 100% and the other values were calculated accordingly. The values represent the mean ± SEM from 5 independent experiments.
Figure 2.
The effect of caffeine and rapamycin on AX3 (A), rip3‾ (B) and lst8‾(C) cells. Cells were inoculated and cell growth was monitored for 72 hours. The highest cell number in each experiment after 72 hours was set to 100% and the other values were calculated accordingly. The values represent the mean ± SEM from 5 independent experiments.
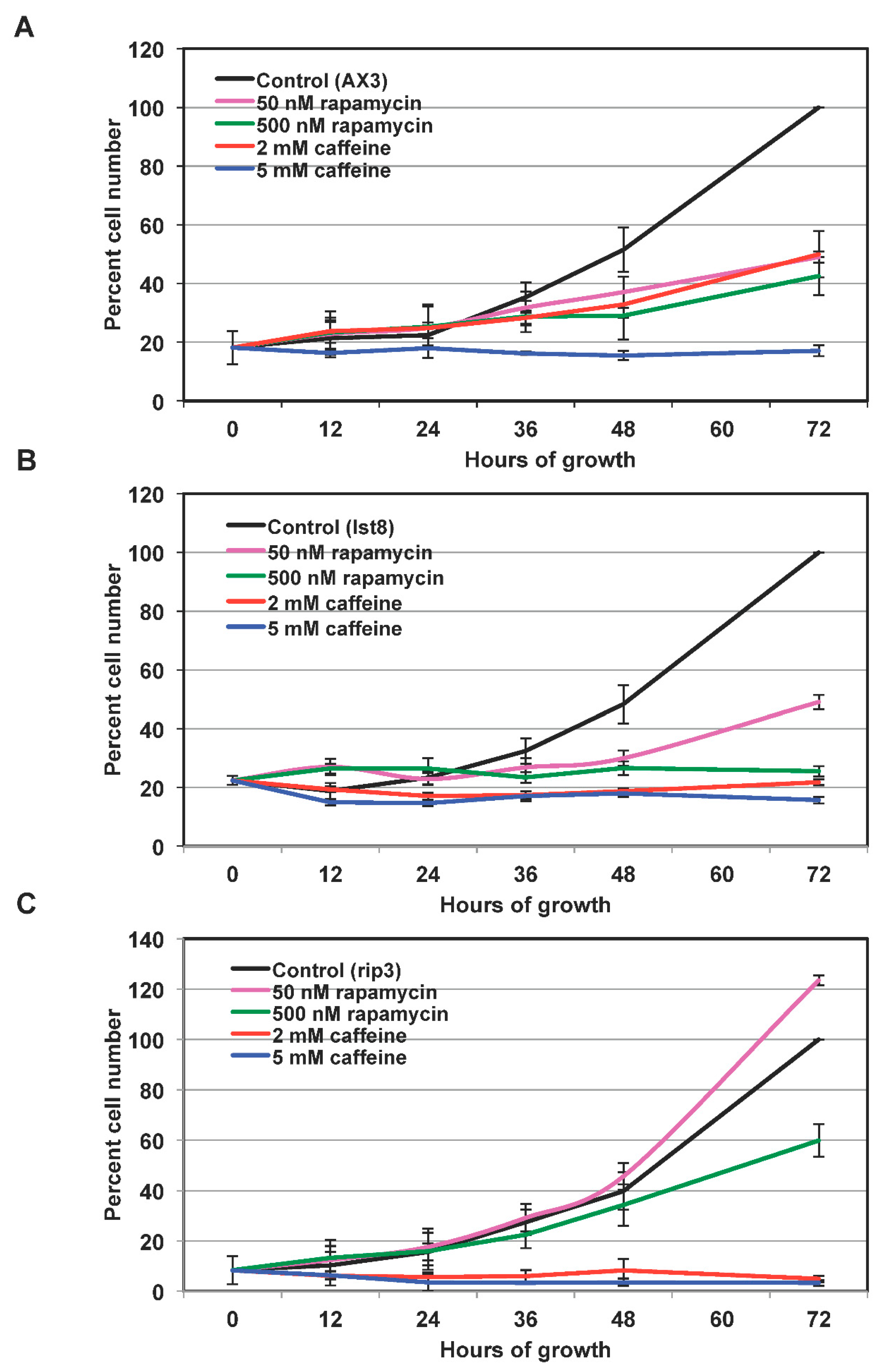
Figure 3.
Development of AX3 cells in the presence of caffeine or rapamycin. A) Time course of development ofAX3 cells either in the absence of caffeine (control) or in the presence of 2, 3 or 5 mM caffeine. B) Development of AX3 cells either in the absence of rapamycin (control) or in the presence of 1, 3 or 5 µM rapamycin. Cells were developed on non-nutrient agar plates at a density of 2x106 cells/cm2. Scale bar corresponds to 1000 μm.
Figure 3.
Development of AX3 cells in the presence of caffeine or rapamycin. A) Time course of development ofAX3 cells either in the absence of caffeine (control) or in the presence of 2, 3 or 5 mM caffeine. B) Development of AX3 cells either in the absence of rapamycin (control) or in the presence of 1, 3 or 5 µM rapamycin. Cells were developed on non-nutrient agar plates at a density of 2x106 cells/cm2. Scale bar corresponds to 1000 μm.
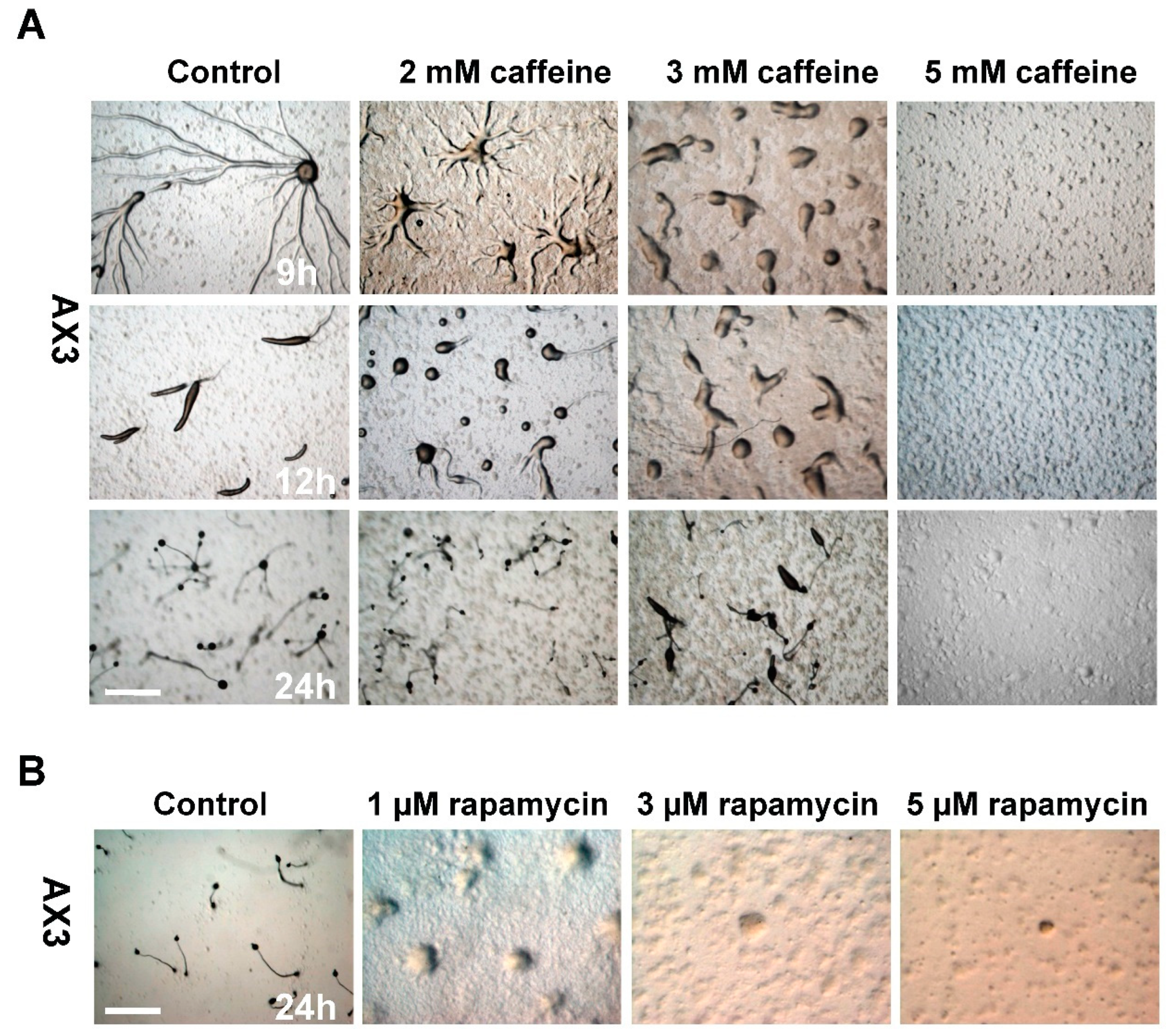
Figure 4.
Complementation of lst8‾cells rescues the aggregation defect. A) Aggregation pattern of wild type AX3 and lst8‾ cells. B) Aggregation pattern of lst8‾ cells over-expressing Lst8 at two different cell densities. The pictures of the aggregates were taken after 9h of development. C) Analysis of aggregation of rip3‾ cells. Aggregation of rip3‾ cells in response to either 2 mM caffeine or 1 μM rapamycin treatment analyzed after 24h of development. Scale bar corresponds to 1000 μm. .
Figure 4.
Complementation of lst8‾cells rescues the aggregation defect. A) Aggregation pattern of wild type AX3 and lst8‾ cells. B) Aggregation pattern of lst8‾ cells over-expressing Lst8 at two different cell densities. The pictures of the aggregates were taken after 9h of development. C) Analysis of aggregation of rip3‾ cells. Aggregation of rip3‾ cells in response to either 2 mM caffeine or 1 μM rapamycin treatment analyzed after 24h of development. Scale bar corresponds to 1000 μm. .

Figure 5.
Validation of microarray data. The differential regulation of DDB_G0285995 (pdsA), DDB_G0275741 (dio3), and DDB_G0286057 (adk1) genes were validated by performing qPCR.
Figure 5.
Validation of microarray data. The differential regulation of DDB_G0285995 (pdsA), DDB_G0275741 (dio3), and DDB_G0286057 (adk1) genes were validated by performing qPCR.
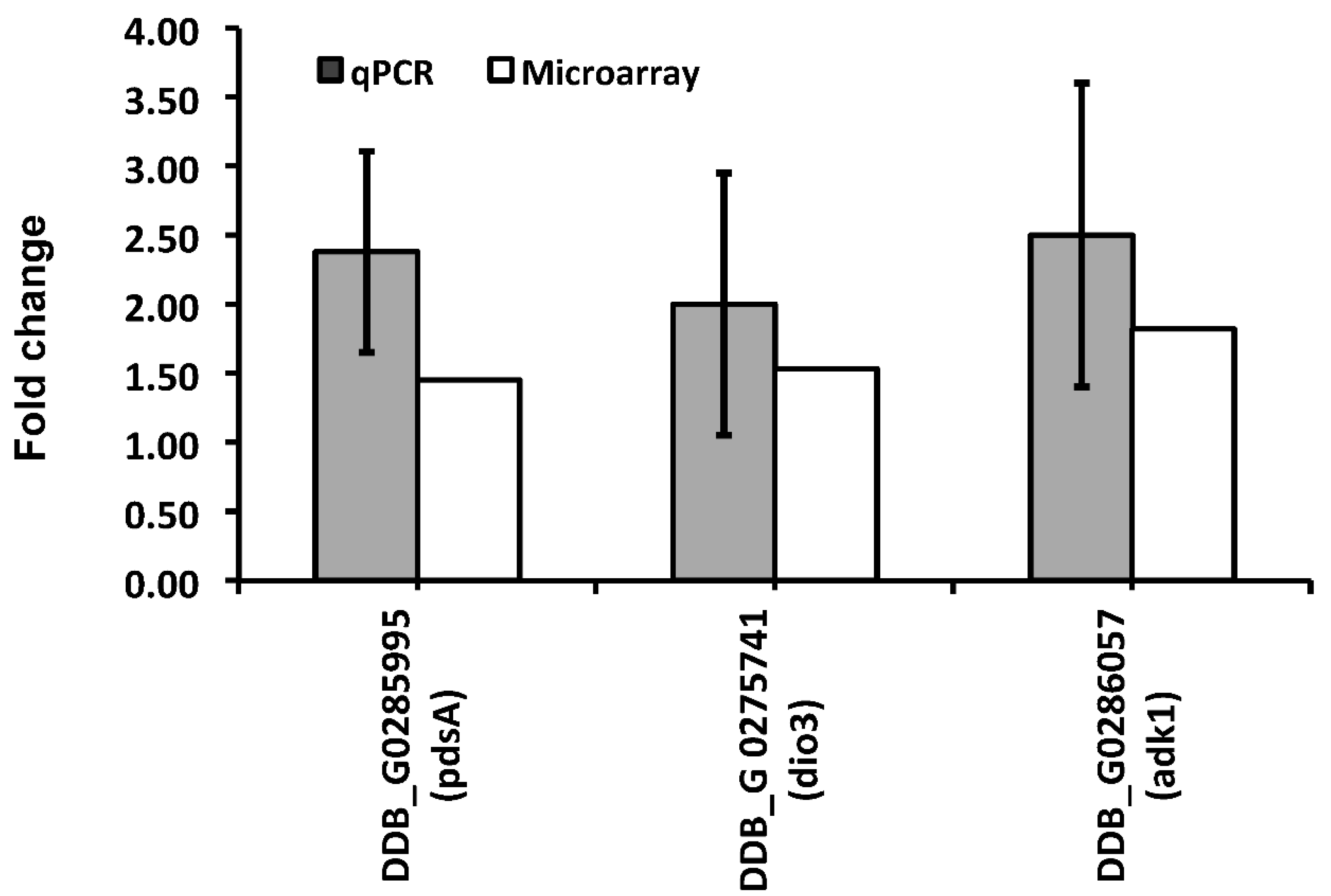
Figure 6.
Venn diagram of differentially regulated genes after 3 and 6 hours of caffeine treatment. The differentially regulated genes of caffeine treated versus untreated cells for 3 and 6 hours of development were determined. The numbers of common and separate up- and down-regulated genes at both time points are displayed in a Venn diagram.
Figure 6.
Venn diagram of differentially regulated genes after 3 and 6 hours of caffeine treatment. The differentially regulated genes of caffeine treated versus untreated cells for 3 and 6 hours of development were determined. The numbers of common and separate up- and down-regulated genes at both time points are displayed in a Venn diagram.

Figure 7.
Functional classification of the differentially expressed genes. The differentially regulated genes in the presence of caffeine as compared to untreated cells at 3 (A) and 6 hours (B) of development were placed into12 functional categories according to the yeast classification and adapted to D. discoideum are given [33]. The red and blue values in brackets indicate the percentage of up- and down-regulated genes, respectively, in the given category. The numbers in the outer circle indicates the total number of genes found differentially regulated in the respective processes.
Figure 7.
Functional classification of the differentially expressed genes. The differentially regulated genes in the presence of caffeine as compared to untreated cells at 3 (A) and 6 hours (B) of development were placed into12 functional categories according to the yeast classification and adapted to D. discoideum are given [33]. The red and blue values in brackets indicate the percentage of up- and down-regulated genes, respectively, in the given category. The numbers in the outer circle indicates the total number of genes found differentially regulated in the respective processes.
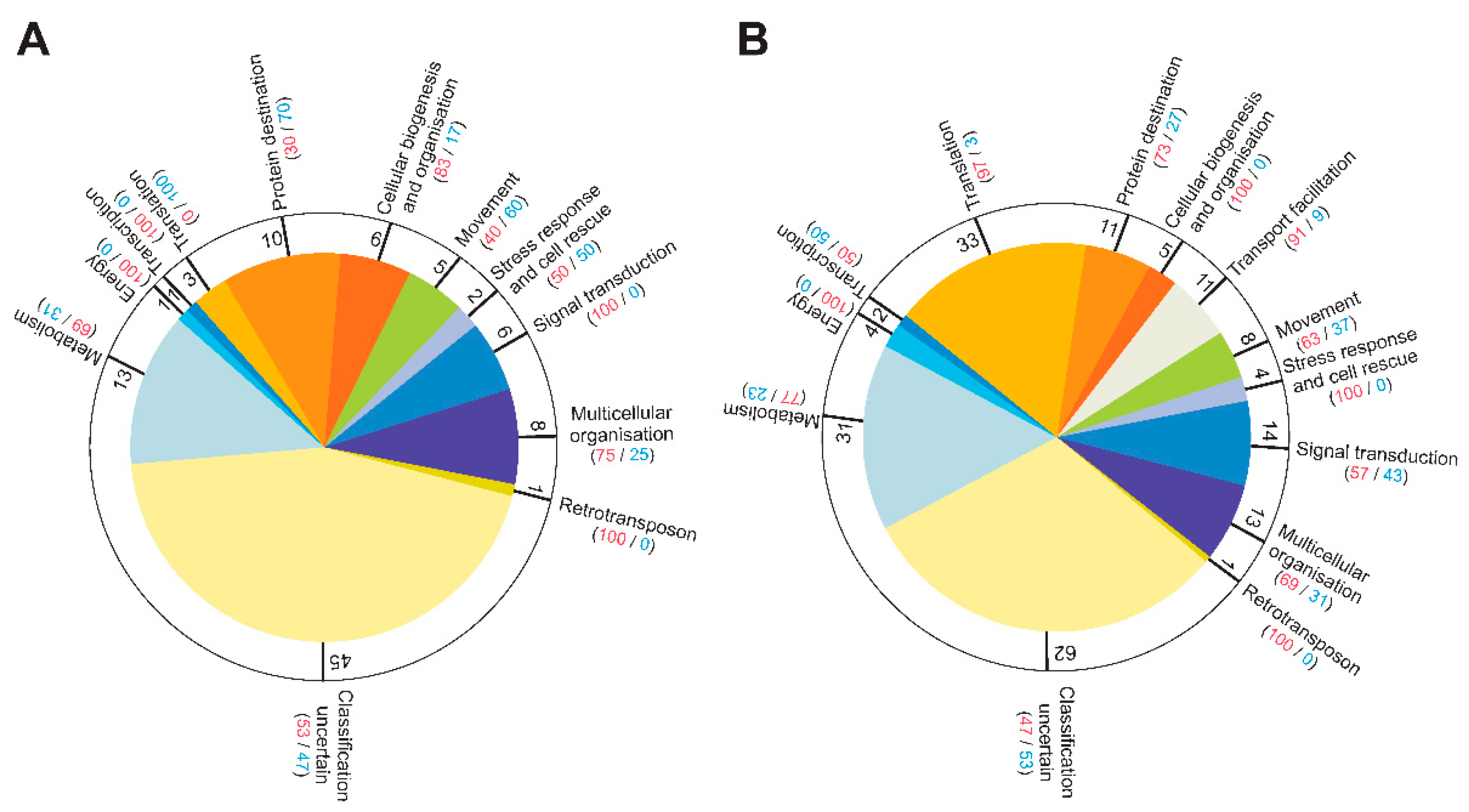

Table 1.
List of common up-regulated genes after 3 and 6 hours treatment with caffeine. Differentially regulated genes were identified using the SAM program. Listed are the genes with more than 1.4 fold differential regulation at both time points in comparison to untreated cells. DDB_G ID is the dictyBase (http://dictybase.org/) gene identification number. The gene name and the gene product description were obtained from dictyBase. FC: Fold Change.
Table 1.
List of common up-regulated genes after 3 and 6 hours treatment with caffeine. Differentially regulated genes were identified using the SAM program. Listed are the genes with more than 1.4 fold differential regulation at both time points in comparison to untreated cells. DDB_G ID is the dictyBase (http://dictybase.org/) gene identification number. The gene name and the gene product description were obtained from dictyBase. FC: Fold Change.
| DDB_G ID | FC 3h | FC 6h | Gene Name | Gene Product |
|---|---|---|---|---|
| DDB_G0281843 | 2,36 | 2,69 | DDB_G0281843 | PAN-1 domain-containing protein |
| DDB_G0273397 | 1,72 | 1,61 | carA-2 | cAMP receptor 1 |
| DDB_G0289393 | 1,69 | 1,95 | psiH | PA14 domain-containing protein |
| DDB_G0287035 | 1,66 | 2,22 | lmpB | lysosomal integral membrane glycoprotein (LmpB) |
| DDB_G0273919 | 1,59 | 1,40 | dscA-2 | discoidin I, alpha chain |
| DDB_G0277863 | 1,57 | 1,91 | pdiA | cAMP phosphodiesterase inhibitor |
| DDB_G0285617 | 1,57 | 1,58 | DDB_G0285617 | agglutinin domain-containing protein |
| DDB_G0285505 | 1,56 | 2,01 | ino1 | inositol-3-phosphate synthase, myo-inositol-1-phosphate synthase |
| DDB_G0292566 | 1,56 | 1,64 | hibA | 3-hydroxyisobutyrate dehydrogenase |
| DDB_G0287281 | 1,55 | 1,67 | DDB_G0287281 | citrate synthase |
| DDB_G0277501 | 1,53 | 1,71 | capB | cAMP binding protein B |
| DDB_G0274093 | 1,53 | 1,40 | adrm1-2 | adhesion regulating molecule family protein |
| DDB_G0272668 | 1,49 | 1,64 | ppr2 | protein phosphatase 4 regulatory subunit 2 |
| DDB_G0282817 | 1,47 | 2,42 | patB | P-type ATPase |
| DDB_G0273805 | 1,46 | 2,10 | ndkC-2 | nucleosidediphosphate kinase, NDP kinase |
| DDB_G0282051 | 1,45 | 1,66 | DDB_G0282051_RTE | TRE5-A ORF1 |
| DDB_G0285995 | 1,45 | 1,47 | pdsA | cAMP/cGMP phosphodiesterase |
| DDB_G0272819 | 1,42 | 1,76 | hspC | heat shock protein |
| DDB_G0272969 | 1,42 | 1,42 | psmB1 | proteasome subunit beta type 1, 20S proteasome subunit beta-1 |
| DDB_G0272875 | 1,42 | 1,58 | cpnB-2 | copine B |
| DDB_G0269122 | 1,41 | 1,42 | cysA | cystathionine gamma-lyase |
Table 2.
Subset of differentially expressed genes in the presence of caffeine as compared to untreated cells. Listed are those genes that encode proteins involved in translation. Orange: 1.4<FC<2; Red: FC>2; Light blue: 0.66<FC<0.71. FC: Fold change.
Table 2.
Subset of differentially expressed genes in the presence of caffeine as compared to untreated cells. Listed are those genes that encode proteins involved in translation. Orange: 1.4<FC<2; Red: FC>2; Light blue: 0.66<FC<0.71. FC: Fold change.
| DDB_G ID | FC 3h | FC 6h | Gene Name | Gene Product |
|---|---|---|---|---|
| DDB_G0269136 | 0,97 | 1,59 | efaAII | elongation factor 1 alpha |
| DDB_G0284035 | 0,97 | 1,45 | efa1B | elongation factor 1 beta |
| DDB_G0282979 | 0,86 | 1,53 | efa1G | elongation factor 1 gamma |
| DDB_G0288373 | 1,05 | 2,13 | efbA | elongation factor 2 |
| DDB_G0281403 | 0,99 | 1,72 | nacA | putative nascent polypeptide-associated complex alpha subunit |
| DDB_G0270316 | 1,16 | 1,41 | rpsA | 40S ribosomal protein SA |
| DDB_G0293742 | 1,23 | 1,80 | rps2 | 40S ribosomal protein S2 |
| DDB_G0293000 | 0,92 | 1,57 | rps3 | 40S ribosomal protein S3 |
| DDB_G0277345 | 1,19 | 1,58 | rps3a | 40S ribosomal protein S3a |
| DDB_G0272825 | 1,27 | 1,49 | rps4 | 40S ribosomal protein S4 |
| DDB_G0286075 | 1,19 | 1,46 | rps5 | 40S ribosomal protein S5 |
| DDB_G0280823 | 1,04 | 1,54 | rps6 | 40S ribosomal protein S6 |
| DDB_G0291864 | 1,02 | 1,45 | rps8 | 40S ribosomal protein S8 |
| DDB_G0284533 | 1,14 | 1,60 | rps13 | 40S ribosomal protein S13 |
| DDB_G0273743 | 1,16 | 1,49 | rps30-2 | 40S ribosomal protein S30 |
| DDB_G0291862 | 1,11 | 1,94 | rpl3 | 60S ribosomal protein L3 |
| DDB_G0277803 | 1,29 | 1,66 | rpl4 | 60S ribosomal protein L4 |
| DDB_G0278539 | 1,38 | 2,27 | rpl5 | 60S ribosomal protein L5 |
| DDB_G0276441 | 1,24 | 1,50 | rpl7 | S60 ribosomal protein L7 |
| DDB_G0274113 | 1,15 | 1,75 | rpl8 | 60S ribosomal protein L8 |
| DDB_G0278959 | 1,15 | 1,41 | rpl9 | 60S ribosomal protein L9 |
| DDB_G0291870 | 1,21 | 1,62 | rpl13 | S60 ribosomal protein L13 |
| DDB_G0275881 | 1,08 | 1,78 | rpl13a | S60 ribosomal protein L13a |
| DDB_G0277975 | 0,99 | 1,46 | rpl14 | S60 ribosomal protein L14 |
| DDB_G0273983 | 1,31 | 2,14 | rpl15-2 | S60 ribosomal protein L15 |
| DDB_G0279997 | 1,01 | 1,45 | rpl18 | S60 ribosomal protein L18 |
| DDB_G0281565 | 1,17 | 1,68 | rpl19 | S60 ribosomal protein L19 |
| DDB_G0290315 | 0,91 | 1,62 | rpl23 | S60 ribosomal protein L23 |
| DDB_G0293502 | 1,08 | 1,55 | rpl23a | S60 ribosomal protein L23a |
| DDB_G0292388 | 1,21 | 1,53 | rpl27a | S60 ribosomal protein L27a |
| DDB_G0282853 | 1,17 | 1,61 | DDB_G0282853 | TCTP family protein 1 |
| DDB_G0276871 | 0,66 | 1,28 | rpl10a | S60 ribosomal protein L10a |
| DDB_G0277823 | 0,65 | 1,35 | alas | alanyl-tRNAsynthetase |
| DDB_G0285451 | 0,71 | 0,90 | leuS | leucyl-tRNAsynthetase |
| DDB_G0279113 | 0,88 | 0,68 | metS | methionyl-tRNAsynthetase |
Table 3.
Subset of differentially expressed genes in the presence of caffeine as compared to untreated cells. Listed are those genes that encode proteins involved in early development. Orange: 1.4<FC<2; Red: FC>2; Light blue: 0.66<FC<0.71 Dark blue: FC<0.66. FC: Fold change.
Table 3.
Subset of differentially expressed genes in the presence of caffeine as compared to untreated cells. Listed are those genes that encode proteins involved in early development. Orange: 1.4<FC<2; Red: FC>2; Light blue: 0.66<FC<0.71 Dark blue: FC<0.66. FC: Fold change.
| DDB_G ID | FC 3h | FC 6h | Gene Name | Gene Product |
|---|---|---|---|---|
| DDB_G0273397 | 1,72 | 1,61 | carA-2 | cAMP receptor 1 |
| DDB_G0277829 | 1,36 | 1,47 | carC | cAMP receptor 3 |
| DDB_G0274023 | 1,35 | 1,77 | capA-2 | cAMP-binding protein |
| DDB_G0277501 | 1,53 | 1,71 | capB | cAMP-binding protein |
| DDB_G0285995 | 1,45 | 1,47 | pdsA | cAMP/cGMP phosphodiesterase |
| DDB_G0277863 | 1,57 | 1,91 | pdiA | cAMP phosphodiesterase inhibitor |
| DDB_G0268620 | 1,30 | 1,67 | pkbA | AKT/PKB protein kinase PkbA |
| DDB_G0272875 | 1,42 | 1,58 | cpnB-2 | copine B |
| DDB_G0287587 | 1,69 | 1,37 | smlA | unknown |
| DDB_G0284363 | 1,30 | 1,65 | cf60 | component of the counting factor (CF) complex |
| DDB_G0274597 | 0,87 | 2,95 | ctnA | component of the counting factor (CF) complex, countin |
| DDB_G0285425 | 1,14 | 1,65 | gpaD | G-protein subunit alpha 4 |
| DDB_G0283419 | 1,43 | 0,93 | gpaI | G-protein subunit alpha 9 |
| DDB_G0275045 | 1,14 | 1,70 | gpbB | GTP-binding protein subunit beta-like protein |
| DDB_G0273817 | 1,40 | 0,81 | ptpA1-2 | protein-tyrosine phosphatase 1 |
| DDB_G0272833 | 2,62 | 0,72 | csbA | contact site B protein |
| DDB_G0276883 | 1,09 | 0,68 | canA | calcineurin A |
| DDB_G0285845 | 0,95 | 0,62 | phdA | PH domain-containing protein; Developmental Gene |
| DDB_G0291253 | 1,06 | 0,50 | dia2 | unknown |
| DDB_G0275439 | 1,19 | 0,50 | cad2 | putative adhesion molecule |
| DDB_G0285793 | 1,18 | 0,36 | cadA | calcium-dependent cell adhesion molecule-1 |
| DDB_G0281605 | 0,67 | 1,22 | cfaD | counting factor associated protein |
| DDB_G0290261 | 0,77 | 1,66 | cfrC | cell surface glycoprotein GP138 C |
| DDB_G0269202 | 0,43 | 2,07 | gdcA | gp64 and disintegrin-like, cysteine-rich protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



