
Submitted:
03 June 2024
Posted:
03 June 2024
You are already at the latest version
Abstract
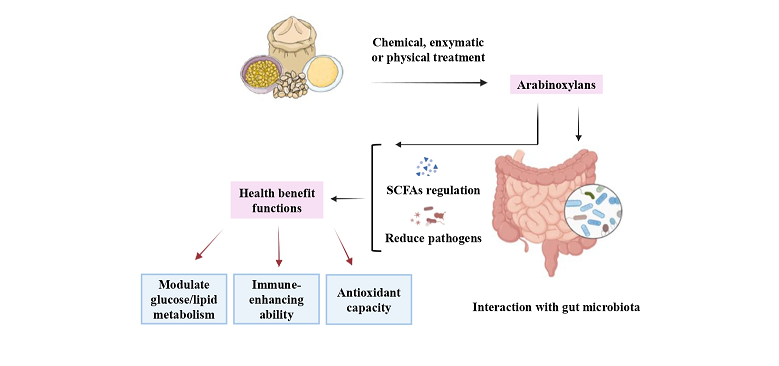
Arabinoxylans (AXs) are non-starch polysaccharides with complex structures naturally occurring in grains (i.e., barley, corn, and others) providing many health benefits, especially as prebiotics. AXs can be classified as water-extractable (WEAX) and water-unextractable (WUAX) based on their solubility, with properties influenced by grain sources and extraction methods. A large body of evidence shows that AXs exert an im-portant health impact, including glucose and lipid metabolism regulation and immune system enhancement, which is induced by the interactions between AXs and the gut microbiota. Recent research underscores the dependence of AX physiological effects on structure, advocating for a deeper understanding of struc-ture-activity relationships. While systematic studies on WEAX are prevalent, knowledge gaps persist re-garding WUAX, despite its higher grain abundance. Thus, this review reports recent data on WUAX struc-tural properties (chemical structure, branching, MW) in cereals under different treatments. It discusses WUAX applications in baking and its benefits deriving from gut fermentation.
Keywords:
water-unextractable arabinoxylans
; chemical properties human
; health properties
; gut microbiota
; fermentation
1. Introduction
The global market has seen a significant increase in the number of whole-grain products in recent decades [1]. These products have the potential to reduce the risk of cardiovascular disease, diabetes, and gastrointestinal cancers by lowering cholesterol and lipid accumulation, improving insulin resistance, and promoting a healthy gut microbiota ecosystem [2,3,4,5,6,7,8,9,10]. The dietary fiber found in cereals, specifically arabinoxylan (AX), plays a crucial role in reducing the risk of diet-related chronic diseases. AXs are a non-starch polysaccharide commonly found in cereals such as corn, wheat, barley, oat, and sorghum [11,12]. Typically, AXs consist of a linear backbone of 1,4-linked β-D-xylopyranosyl (1,4-β-D-xylp) residues and α-L-arabinofuranosyl (α-L-araf) units that can be substituted at the C(O)-2 and/or C(O)-3 positions [13,14]. In addition, the xylose (xyl) unit can carry methyl glucuronide and the arabinose unit can be bonded to ferulic acid residues [15]. Also, both xylose and arabinose can be acetylated. However, the structure of AX varies between different grains and even within the same grain [16].
The extraction of AX from grains can be performed using various methods, each resulting in unique structural features [17,18,19]. Nonetheless, the structure of AXs obtained from different raw materials or different extraction methods is not identical. These structural characteristics, such as the content of ferulic acid, molecular weight (MW), and substitution, influence the functional properties and the physiological activities of AXs [20].
There are two types of AXs based on solubility: water-extractable arabinoxylans (WEAX) and water-unextractable (WUAX). WEAX is a small fraction of total AXs and is loosely bound to the cell wall surface, while WUAX is the most abundant fraction and is connected to other cell wall components [21,22,23,24].
In recent years, AXs have found applications in the baking industry, as thus interact with other components of flour and dough to enhance dough and bread quality. AXs interact with gluten, which has been recognized as one of the main components controlling the quality of dough and bread [25]. It has been proven that the amount of WEAX and WUAX has the function of influencing the rheological behavior of the dough and the quality characteristics of the bread [26] WEAX and WUAX have different effects on baking quality. However, the presence of WUAX can harm baking quality [27].
The gut microbiota has received increased attention and is considered an eminent symbiotic partner in the maintenance of human health [28]. The gut microbiota can define and regulate host homeostasis by determining nutritional, immune, and neuroendocrine homeostasis [29]. AXs also play a crucial role in regulating the composition of gut microbiota and promoting intestinal health [30], with a variety of bioactivities such as immune and antioxidant enhancement, strengthening of the intestinal epithelial barrier, relief of constipation, and improvement of lipid and glucose metabolism [5,6,7,8,9,10,11,12]. AXs can modulate the profile and function of some of the beneficial bacteria in the human gut microbiota [31]. The structural properties of AXs determine their biological effects, fermentation process, and regulation of the intestinal microbiota. The MW of WEAX has been found to influence its prebiotic efficiency [32].
While most AXs in cereals are WUAX, these have not received as much attention as WEAX. Therefore, this paper aims to study the structure-activity relationship and treatment of WUAX in cereals, as well as its applications in baking and potential functional benefits for the human health.
2. Materials and Methods
This study consists of an up-to-date review of the literature, which contains data about structure feature, effects on baking products and human gut health of AXs. A search of the Web of Science database using the keyword "water non-extractable arabinoxylan" yielded 110 articles, and the addition of the keyword "cereals" yielded 69 articles. These articles were summarized and critically discussed to provide a coherent review.
3. Structural Features
3.1. Monosaccharide Compositions and Branching Degree
AX is composed of a xylose backbone with arabinose substitutions and ferulic acid, connected by various types of glycosidic bonds (Figure 1).
The monosaccharide composition and the ratio of arabinose to xylose (A/X) describe the degree of branching in AX. A lower A/X indicates less branching. Although of different origins, most AXs comprise arabinose, xylose, glucose and galactose [33]. While α-L-arabinofuranosyl is the main substitution on xylan backbones, it has been shown that small amounts of other sugar residues (i.e., glucuronic acid, 4-O-methylglucuronic acid (MeGlcA), or short oligomers consisting of L- arabinose, D-xylose, D-galactose, D-glucose and/or glucuronic acid) can act as substituents [34]. The monosaccharide composition of WUAX from spelt bran extraction was arabinose, xylose, mannose, glucose, and galactose, with xylose being the highest percentage [35]. The pattern and extent of arabinose substitution varies across plant species and tissue locations [36]. Arabinogalactan and glucomannan are potential sources of galactose or mannose [37]. The monosaccharide composition of AX in cereals generally has a higher proportion of arabinose and xylose. Based on data obtained by alkali extraction of WUAX from different cereals, wheat flour had 46.6% arabinose and 48.6% Xyl of AX [38], and rye bran had 36.53% arabinose and 63.31% Xyl of AX [39]. Research has shown that the proportion of Xyl in AX extracted from oats is as high as 78% [40]. WUAX has a higher polydispersity and lower A/X compared to WEAX. In general, the A/X ratio for AX in cereals ranges from 0.5 to 1.0, with sorghum and rice reaching 1.0, wheat close to 1, and the remaining cereals such as barley (0.23-0.58), maize (0.34-0.56) and rye (0.46-0.60) being smaller [41,42]. The A/X ratio of WUAX in spelt bran was 0.60-0.65, and the A/X of wheat was in most cases greater, meaning that the arabinose branch in spelt bran WUAX is relatively small. There are higher levels of galactose from rice and barley bran compared to other cereals with AX [43,44], and no detectable mannose in AX from wheat and oats. The A/X ratio of WUAX in 12 corn bran samples extracted using the alkaline method by Zhang et al. was also relatively low, with a range of 0.46 ~ 0.54 [45]. Different grains have varying branch chain and monosaccharide compositions, with WUAX generally having a higher branch chain degree and lower arabinose substitution, resulting in high MW and low solubility in water. The differences in monosaccharide composition are typically associated with grain type and genotype.
3.2. Glycosidic Linkages
Methylation analysis is commonly used to determine the glycosidic linkage of AX across different grain sources. In cereal AX, the 4-xylp backbone, composed of xylose sugar residues, is predominantly present with branching points at the 0-2 and/or 0-3 positions. Small amounts of txylp (terminal xylopyranosyl), 2-xylp, and 3-xylp can also be detected using this method [46,47]. Arabinose-based sugars, including t-araf and 3-araf, are detected in all cereal sources, while other arabinose residues, such as 2-araf, 4-araf, 5-araf, 2,5-araf, and 3,5-araf, are only occasionally detected in cereals. The linear backbone of AX consists of D-xylopyranose linked by β-1,4 glycosidic linkages. α-L-furan arabinose can be monosubstituted or disubstituted at the O-2 and O-3 positions, and ferulic acid and 4-O-methylglucuronide can also act as substituents [48]. Ferulic acid, when attached to arabinose residues via ester linkages at the C(O)-5 position, can have significant effects on the functional properties of AX. Xylp residues form the backbone through 1→4 linkages and occur in three forms: unsubstituted, monosubstituted, and disubstituted. Arabinofuranosyl molecules are mainly present as monosubstituted residues at O-3. A study by Zhang, et al. (2021) found that the side chain of the arabinose β-furanosyl residue in maize bran AX was mono-linked at the O-3 position. Xylopyranosyl groups with different substitution patterns have different A/X ratios, which carry important implications for the structural features and biological activities of AX [45]. The methylation analysis by Guo et al., (2021) showed that the xylose-based sugar residues in WUAX from alkali-extracted wheat bran contain T-xylp, 4-xylp, 3,4-xylop, and the arabinose-based sugar residues contain t-araf, 3-araf, and 2,3-araf. The xylan backbone contains 46.72% unsubstituted xylp, 27.42% monosubstituted xylp at the 0-3 position, and 25.88% doubly substituted xylp at the 0-2 and 0-3 positions [49]. Guo, et al., (2019) extracted a relatively low-branched AX from hulled barley, and of this, 71.19% of unsubstituted (1, 4-linked β-D-xylan), 14.78% of mono-substituted (1,3, 4-linked β-D-xylan) and 10.76% of doubly substituted (1,2,3, 4-linked β-D-xylp) xylose units were backbone components via β-(1→4) bonds [44]. However, it has also been shown that the AX of isolated barley and rye contained more AX molecules monosubstituted at O-2 [22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50]. Mono-substituted and di-substituted WUAX are present not only in hulled barley bran but also in wheat flour and rye bran [41]. These studies have confirmed that both monosubstituted and disubstituted WUAX can be present in cereals. This difference also affects the glycosidic bond composition of AX and implies that the structure of AX is more heterogeneous and different.
3.3. Combination of Phenolic Compounds
Grains contain high levels of phenolic compounds (e.g. ferulic, coumaric, p- coumaric, caffeic, chlorogenic, and eugenic acids) [51]. AXs can exhibit antioxidant activity by binding to these phenolic compounds. Xylan backbones with high substitution and low ferulic acid (FA) content are more water-soluble, while AX with low substitution and high ferulic acid content is water-insoluble [52]. Soluble dietary fibers, including β-1,3/1,4 glucan, water-soluble AX, and arabinogalactan protein, have low binding to phenolics (10%), whereas insoluble dietary fibers such as water-insoluble AX and cellulose-lignin complexes have high binding to phenolic acids (90%) [53]. The total phenolic acid content of WUAX obtained by Srinivasan et al., (2020 & 2019) from alkali extracts of four millet varieties was 104.3, 164.6, 345.5, and 1307.7 μg/g, respectively, all containing p-coumaric acid and ferulic acid, with ferulic acid being the most represented, suggesting that the phenolic acid content of AX is highly correlated with genotype in addition to structure and extraction method [54,55].
Ferulic acid is the most prevalent and abundant phenolic compound that binds to AX, but alkali-extracted WUAX contains less ferulic acid because the hydrolysis of alkali extraction breaks the covalent ester bond linking ferulic acid to the AX residue. Nevertheless, a sequential process using subcritical water, membrane filtration and selective enzyme treatment allowed some retention of ferulic acid in the AX, allowing the extraction of feruloyl AX and arabinoxylo-oligosaccharide (AXOS) from wheat bran. Bijalwan et al., (2016) determined the phenolic acids bound in hydroxycinnamic acid-bound arabinoxylan (HCA-AX) in five different millets by liquid chromatography-mass spectrometry (LC-MS) extraction, the results showed that ferulic acid was the highest across all millets [56]. In a study by Ayala-Soto et al., (2015), various phenolic compounds such as ferulic acid, p-coumaric acid, and other hydroxycinnamic acids were detected in the products obtained from alkaline extraction of WUAX from three different sorghums. The total hydroxycinnamic acid content ranged from 0.16-0.24 mg HCA/mg db and ferulic acid from 0.09-0.16 mg FA/mg db [57].
The binding of AX to phenolic acids significantly affects its functional activity, and different types of phenolic compounds have varying effects on its antioxidant capacity [58]. In a study by Li et al. (2016), the phenolic acid content was increased by binding ferulic acid more strongly to corn WUAX obtained by alkali extraction, and it was found that the structure of corn arabinoxylan (CAX) was not perturbed during the modification, with an A/X ratio of about 0.62 and an increase in the average MW accompanied by a significant increase in the antioxidant capacity [51]. Li et al. (2017) combined natural caffeic acid (CA) with maize WUAX obtained by alkali extraction through esterification to obtain corn bran arabinoxylan (CA-CAX) with varying degrees of substitution [59]. The results show that the antioxidant capacity of CA-CAX was significantly higher than that of CAX with increasing concentration and degree of substitution. Guo, et al. (2021) found that the MW of AX decreased from 694 kDa to 386 kDa after grafting catechin onto alkali-extracted wheat bran WUAX in the presence of hydroxyl radicals, indicating that AX was degraded but the linkage pattern was not significantly different from that of alkali-extracted AX [49]. Ayala-Soto et al. (2014) determined the phenolic acid content of AX in different fibers such as maize fiber (MF), insect-resistant maize pericarp (RP), and insect-susceptible maize pericarp (SP) and found that the antioxidant capacity of AX was related to the total phenolic, dehydrotriferulic acid (tri-FA) and p-coumaric acid (p-CA) concentrations, but differences in the A/X ratio of WUAX obtained from different fibre sources after alkali extraction were not significant [60].
4. Effects of Different Treatments on WUAX Structure in Cereals
4.1. Alkali Extraction
Aqueous treatment can easily extract WEAX, which is not strongly bound to the cell wall surface of cereals. However, WUAX, present in the cell wall and bound to other components like proteins, lignin, and cellulose through covalent and non-covalent bonds, requires the use of alkaline solutions to dissolve these compounds. Alkaline extraction breaks covalent and hydrogen bonds in the polysaccharide matrix, releasing various polysaccharides from the cell [61,62,63,64]. Since the carboxyl group in AX can become negatively charged in an alkaline environment, leading to repulsion between different molecules, AX is easier to extract (Hollmann and Lindhauer, 2004). Therefore, alkaline extraction methods (NaOH, Ba(OH)2) are commonly used to extract WUAX. This extraction process is shown in Figure 2.
The extraction of AX with different types of alkaline solutions also leads to different changes in the structure of AX. As the reaction during alkali extraction is very intense, it destroys functional groups such as ferulic acid [66]. Table 1 describes the effects of alkali extraction on the A/X ratio and MW of WUAX in different cereals.
The degree of disruption of the arabinose substituents during alkaline treatment is not high, resulting in an increase in the A/X ratio [75]. Gruppen, et al. (1993) found that the structure of AX obtained from wheat by alkali extraction (AE-AX) was almost identical to that of WEAX, with only minor differences in average MW, with AE-AX being higher than WEAX and with slightly higher A/X ratio due to a higher proportion of arabinose side branches [76]. The A/X ratio of WUAX extracted from three different sorghum brans by an alkali method was higher, namely, white sorghum bran (1.41), which was higher than that of red sorghum bran (1.08) and high tannin bran (1.14) [57]. In addition to the typical structure of WEAX in specific grains, such as the A/X of WEAX extracted from wheat which is 0.5 - 0.6, there are still great differences within different grains. For WUAX, in addition to differences between species, its structure is highly dependent on variables in the extraction process, such as the type of solvent used for alkali extraction and its concentration. WUAX obtained from wheat bran by alkali extraction had a MW of 694 kDa, and the A/X molar ratio of AX was 0.95, indicating a highly branched molecular structural formula [49]. Oat WUAX obtained by extraction with saturated Ba (OH)2 had very low substitution, whereas WUAX extracted with 1 or 6 M NaOH had small substitution branches and aggregated readily [40]. Furthermore, it was found that extraction with NaOH was also able to yield AX highly substituted with arabinose, with an A/X of 0.83 [67]. Guo, et al. (2019) used a modified alkali extraction method to extract WUAX from hull-less barley bran, yielding HBAX-25, a relatively low-branched AX with an average MW of 298.36 kDa and an A/X of 0.58 [44]. Buksa et al., (2016) The MWs of WEAX and WUAX were compared in wheat, and it was found that the MW of WUAX is higher than that of WEAX, with the trend of AX in rye being almost identical to that in wheat [77].
In summary, WUAX obtained by alkali treatment has a high MW, and the structural characteristics of AX (A /X, etc.) are still related to the type of grain. AX from the same grain and extracted with the same alkali solution, can yield different structures, which may be related to the concentration of the alkali solution and the extraction conditions, but the greatest influence does appear to be the variety and genotype of the grain.
4.2. Enzymatic Extraction
Enzymatic treatments are widely recognized for their ability to extract WUAX from cereals, breaking the link between AX and the cell wall and converting it to WEAX. AX can be degraded more accurately and efficiently using enzymatic extractions which are more environmentally friendly, providing the same results as chemical methods such as alkaline extraction [78,79,80,81]. Additionally, enzymatic treatment can help retain phenolic substituents such as ferulic acid. This extraction process is shown in Figure 2. Studies have shown that enzymatic extraction of WUAX using endo-xylanase is as effective as chemical methods [82]. Zhou et al. (2010) compared alkaline and enzymatic extraction of AXs from wheat bran and found that the enzymatically extracted AX had a lower average MW and polydispersity, lower substitution, and contained more protein and ferulic acid than the alkaline extracted AX, which may induce a higher immune response [67]. A comparison of the structures of WEAX and XEAX (xylanase-extracted arabinoxylan) obtained by enzymatic digestion in wheat endosperm showed that XEAX has a different degree of substitution compared to WEAX and that XEAX has more monosubstitution and less unsubstitution than WEAX [83]. Ma et al. (2020) investigated the effect of dual enzymes on the extraction rate and AX content of extracts and confirmed that combined enzymatic extraction of fresh maize fiber using xylanase and cellulase was more effective than chemical methods [84].
The structure of AX obtained by enzymatic treatments depends on the specific enzyme used [85]. The main enzyme families involved in the degradation of AXs are endo-β-(1,4)-xylanase, β-(1,4)-xylosidase, α-L-arabinofuranosidase, and ferulic esterase [86]. Endo-xylanases can attack the backbone of AX, changing its structure and affecting its function [87]. Enzymatic degradation of AX usually starts with the stripping of lignin from the cell wall, and since ferulic acid is the bridge between lignin and AX, the ester bond between arabinose and ferulic acid can be broken by ferulic acid esterase. Highly specific enzymes such as endo-xylanase and xylanase are used to convert WUAX to WEAX. However, endo-xylanases are not only able to convert WUAX to WEAX, but also further degrade it into its individual components (arabinose and xylose) [62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88]. To further degrade AX, arabinofuranosidase can remove side chains from the xylose backbone, and xylanase can degrade the xylose backbone.
Xylanases are the enzymes primarily involved in the degradation of AX and can be classified into two main families of glycoside hydrolases, glycoside hydrolase family 10 (GH10) and glycoside hydrolase family 11 (GH11). Most GH10 family xylanases can act on the xylosidic linkage at the non-reducing end of the substituted residue, resulting in a larger AXOS than that processed by GH10 xylanases due to the greater hindrance of the substituted group to GH11 xylanase [40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89]. This further suggests that GH11 has a higher substrate specificity than GH10. The polysaccharides in rice, wheat, and maize obtained by enzyme treatment were analyzed. It was found that the A/X ratio was less than 1, except for rice which had a high A/X ratio (3.6) [53]. The MW of AX (51.6-132 kDa) obtained after treatment of WUAX in maize bran with xylan endonuclease by Kale et al. (2018) was lower than that of natural AX (253 kDa) [90]. This may be due to the destruction of the β-1, 4-glucoside linkage of the xylose main chain by endo-xylanase while affecting arabinose substitution in the xylose chain. Table 2 describes the effects of two major enzymes on the A/X ratio and MW of WUAX in different cereals.
The method of enzyme preparation also has a specific effect on the structure of AX. Enzymatic extraction offers a promising alternative to traditional methods for extracting WUAX from cereals. The average MW of the alkali-extracted AXs was 3.5 × 102 kDa, which was approximately 10 times higher than that of the enzymatically extracted AX (32.52 kDa) [67]. In addition, the concentration of an enzyme can modify the MW of AX, for example in endo-xylanase the MW of AX decreased from 6.2 kDa to 2 kDa as the quantity of enzyme was increased [93].
4.3. Microbial Fermentation
Several studies on WUAX have indicated that the fermentation of grains can lead to structural changes and degradation. For example, during spontaneous fermentation of maize bran, the content of WEAX and free ferulic acid content increase, and the A/X ratio decreases [94]. An increase in insoluble polysaccharides (arabinose + 13%, xylose - 3%, and glucose + 53%) was observed in wheat bran after fermentation with Lactobacillus plantarum [95]. Fermentation of wheat bran by Bacillus spp. also leads to the appearance of different peaks of xylose and arabinose in the chromatogram of the extract [96].
Similarly, the degradation of WUAX can also occur during the production of grain products such as bread. Kiszonas et al. (2015) studied the changes in AX levels during bread-making and found that WEAX levels increased during dough fermentation regardless of whether whole wheat or refined flour was used [97]. This may be due to the hydrolysis of WUAX during this process. The chemical structure of AX changes during bread-making and dough fermentation. Analysis of samples taken at different stages of the bread making process revealed that WUAX is degraded during the first fermentation, increasing WEAX. Additionally, the A/X ratio also increases, possibly due to the presence of xylanase in flour [98].
Furthermore, the structural changes and degradation of WUAX were also found during the brewing process of grain. The abundance of WEAX (WEAX yield) in brewers’ spent grain (consisting of 60% barley and 40% rice) is related to the enzyme and L. plantarum strains involved in the saccharification and fermentation process. The average degree of polymerization (DP) of AX varied within a small range (4.7 -6.3), but the average DP of the sample without saccharification and fermentation was as high as 15.6. In addition to its role in enhancing enzyme activity, analysis of the L. plantarum plant genome revealed the presence of genes encoding for hydrolases and esterases that contribute to the breakdown or utilization of AX [99]. Solid-state fermentation of brewers’ spent grain with Fusarium oxysporum f. sp. lycopersici showed significant differences in WEAX content, indicating the ability of this fungus to degrade WUAX. However, the content of WEAX decreases with extended fermentation time, likely due to the microorganisms using these polysaccharides for metabolism and growth [100].
Microbial fermentation is a natural and potentially gentler treatment method for modifying WUAX. Some microorganisms possess enzymes that can biodegrade WUAX, leading to changes in its structure and increased solubility, resulting in higher WEAX content. However, the use of this method is currently limited, and further research is needed to classify and study the microorganisms capable of degrading WUAX. Additionally, identifying the genes of the enzymes carried by these microorganisms could provide further insights.
5. Technological Proprieties of Industrial Interest
5.1. Interaction with Gluten Protein
Gluten proteins have a decisive influence on the quality of flour products and are the main determinant of the viscoelastic properties of dough and final cereal products. The gluten network is formed after the hydration and mixing process of the dough and is stabilized through disulfide bonds and non-covalent bonds, such as hydrogen and ionic bonds [1]. Many studies have been carried out to understand the mechanisms of interaction between AX and gluten proteins. Wang et al. (2004) suggested that WUAX may act as a physical barrier, weakening the attraction between gluten molecules, directly interfering with the formation of gluten networks, and increasing the tensile strength of dough by altering the corresponding wheat gluten macromolecules [101]. Frederix et al. (2004) investigated the effects of WEAX and WUAX by adding their fractions with different MWs to wheat flour the study of the effect on gluten adhesion in dough, showing that WUAX was thought to harm gluten aggregates, reducing their numbers [13]. As discrete cell wall fragments, WUAX may interfere with gluten agglomeration by spatial dislocation or by creating physical barriers to interaction between gluten particles. Si et al. (2021) investigated the effect of interactions between glutenin and WUAX on the conformation of glutenin during heat treatment [102]. The results showed that the addition of WUAX reduced the viscoelasticity and thermal stability of gluten proteins, leading to a softening of the system. According to Wang et al. (2020), AX inhibits the formation of disulfide bonds in bread dough and interferes with protein aggregation. The low ferulic acid content of bran AX directly reduces dough extensibility as it is less cross-linked to gluten [1].
Given the negative effects of WUAX on gluten proteins in dough, many studies have been conducted to reduce these effects. Verjans et al. (2010) demonstrated that xylanase has the ability to degrade a certain amount of WUAX, with different types of xylanase presenting different abilities [103]. Jiang et al. (2018) added pentosidase (Pn) to the preparation of Chinese whole wheat steamed buns. The interaction of enzyme-interpreted AX with bran protein was investigated. The results showed that the addition of Pn reduced the MW of AX released from WUAX. The released AX increased the gluten content in the macromolecule and the increase in Pn made the gluten network more porous and dispersed, which helped produce good dough extensibility. This indicates that the right amount of Pn is beneficial to the formation of a homogeneous fine-crumb structure, resulting in larger and less stiff whole-grain buns [104]. Sun et al. (2022) used enzymatic hydrolysis of WUAX to investigate the effects of WUAX and its hydrolysis products on the aggregation and structure of gluten proteins. The results showed that WUAX disrupted the original homogeneous network structure of gluten proteins and formed a dispersed network structure [105]. However, enzymatic degradation significantly reduced the average MW of WUAX and induced the aggregation of gluten protein molecules, and the resulting small molecule AX contributed to the formation of the gluten network structure, thus improving the quality of whole wheat products.
5.2. Effect on Physical Properties of Dough
As the major non-starch polysaccharide in cereals, AX has various effects on many cereal-based products such as bread, cakes, and doughnuts. AX plays an important role in dough and bread quality by interacting with other components of the flour and dough system [25]. In general, WEAX is usually beneficial for the quality of dough and bread as it increases water absorption, improves dough mixing stability, and increases bread volume, while the presence of WUAX has negative effects [106]. WUAX tends to disrupt the bubble interface of fermented doughs during bread making, thereby disrupting the pores [102]. Espinosa-Ramírez et al. (2020) compared the effect of WUAX extracted from maize on gluten-free dough at different levels of addition: in the presence of 1% AX, the dough matrix was observed to be inhomogeneous and lacking in gluten networks. The addition of 3% AX led to a complete change in this structure, forming a more homogeneous network [107]. Arif et al. (2018) investigated the effects of pentosan on the processing properties of durum wheat flour doughs and found that both water-extractable and water-unextractable pentosan had positive effects on dough performance by improving the water absorption capacity of flour, delaying dough maturation and improving stability as well as the dough mixing tolerance index [106]. However, the positive effect on bread parameters of adding water-extractable pentosan was greater. The water holding capacity of WUAX in wheat bran was significantly higher than that of WEAX. A recent studiy have shown that bread with the addition of 10% AX retains more moisture after baking [108].
Based on the effect of WUAX on the properties of dough, it has been found that the addition of xylanase has a positive effect on dough quality. Yang et al. (2021) investigated the combined effect of xylanase (XYL) and glucose oxidase (GOX) in the production of whole wheat bread, which not only increased the gluten content of the dough (5.71% ~ 13.04%) and the alcoholic protein solubility (3.57% ~ 6.01%), but also reduced the viscoelasticity of the dough and resulted in a more open gluten skeleton structure [109]. The effects of XYL and GOX on wheat dough were highly dependent on the degree of WUAX hydrolysis. Xylanase likely converts part of the WUAX to WEAX. The molar mass distribution of either WEAX or WUAX in the wheat bran affects the strength of the dough, and this is related to the type of wheat [110,111]. The addition of high MW WUAX weakens the dough structure, whereas the low MW AX obtained after enzymatic digestion by xylanase eliminates this negative effect to some extent, since the low MW AX forms a better gluten network and improves the stability of the dough [112]. However, current studies come to different conclusions on whether the A/X ratio of AX in bran affects the rheological properties of dough, which requires further investigation [1,110,111]. Overall, since the AX structure in cereal is greatly affected by the type and genotype of cereal, further studies should be carried out on certain specific varieties of cereal to maintain greater feasibility.
5.3. Effect on Final Cereal Products
In cereals, WUAX is significantly more abundant than WEAX, and the MW and degree of substitution are relatively high. Therefore, WUAX is considered to be the predominant cause of the coarse texture and poor appearance of cereal products [113]. Crumb texture is one of the most important indicators of bread quality. WUAX has a high level of water absorption, which reduces the stability of the dough foam, ultimately reducing the volume of the bread and increasing the density of the bread matrix. As a result, the crumb hardness increases and the texture becomes rougher [108]. In a study by Espinosa-Ramírez et al. (2020), where WUAX from maize bran was added, there were no significant differences in crumb hardness between the breads from the pan-forming and the control breads, while the control breads had better elasticity, cohesion and resilience. For the drop dough model bread, the bread with WUAX had lower hardness and chewiness as well as higher 2D area, height, cell density, and surface area compared to the control bread, especially at higher AX levels. However, AX has a negative effect on elasticity, cohesiveness, and resilience [107].
Specific volume is also an important bread quality parameter, and this is influenced by crumb structure, moisture content, and dough gas retention [108]. The effect of AX on bread volume is also highly dependent on fiber type and amount added. In a study by Koegelenberg and Chimphango (2017a), it was found that regardless of purity, WUAX obtained by alkali extraction at a dose of 0.8% replaced 2.5% of the flour without affecting the physical properties of the bread other than color, maintaining the weight, height, and volume of the bread [42]. However, Zhang et al., (2019) observed increased volume at all levels of adding bran AX, up to 5% [108]. The increase in specific volume after the addition of bran AX is due to the increase in strength and elasticity of the gluten-starch network [114].
To improve the quality of cereal products, Pihlajaniemi, et al. (2020) added AX syrup, obtained by treating wheat bran with KOH, to bread, resulting in a softer texture [75]. Enzymes are widely used in the baking industry to improve the rheological and processing properties of doughs and the characteristics of the final product due to their greater efficacy and safety compared to chemical additives. Ghoshal et al. (2013) used partially purified xylanase in the production of whole-grain bread and investigated its effect on bread quality attributes at room temperature (25±2℃) and under refrigerated conditions (4±1℃). It reduced not only the water absorption of the dough but also the moisture loss of the bread during storage [115]. Arif et al. (2018) investigated the effect of pentosans on the quality of durum wheat bread and showed that the addition of aqueous pentosan increased bread volume and specific bread volume by 1-5%, whereas the addition of non-aqueous pentosan reduced bread volume by 30-44%. Both pentosans reduced crumb hardness by 5-20% [106]. Ghoshal et al. (2013) found that the addition of xylanase increased bread-specific volume, extended shelf life, reduced hardness during storage, reduced moisture loss, and improved bread color, and the addition of xylanase also improved the sensory properties of bread [115].
6. Biological Activities of AX from the Perspective of the Gut Health
6.1. Fermentability in the Human Gut
Due to its inability to be degraded by mammalian enzymes, AX can cross the small intestine and end up in the colon where they are degraded by the gut microbiota. There are numerous sources of AXs and the structure of AX obtained from different grains or different extraction methods varies, especially the carbohydrate structure of AX, in details including monosaccharide composition, branching, and average MW. These are important factors affecting its functional properties, and may influence the fermentability and fermentation rate of AX in the human gut [20]. The diagram of AX fermenting in the gut is shown in Figure 3.
The degree of fermentation of AX depends on its structure. Due to its complex structure, the fermentation rate in the human gut is slower, which means that WUAX stays in the human gut longer than WEAX. Unbranched AX has a higher fermentation ability than more branched AX [116]. Damen et al. (2011) studied the intestinal fermentation of WUAX, WEAX, and AXOS in rats. WUAX was the most resistant to catabolism, with only 27.2% fermenting in the cecum and 42.7% catabolizing in the colon. The average arabinose substitution rate of diets containing WUAX increased from approximately 0.50-0.70 in the diet to 0.80-0.90, indicating that WUAX becomes more branched during fermentation [117]. Rose et al. (2010) investigated the fermentation of WUAX from maize, rice, and wheat bran in human feces and showed that AXs with high MW and monosubstituted arabinose side chains are able to sustain fermentation in the human intestine to produce short-chain fatty acids, and that AXs can be degraded by different mechanisms depending on the diet [118]. Xu et al. (2022) used alkaline extraction to obtain CAX then the extract was partially purified and hydrolyzed to analyze its structure and study the human intestinal flora fermentation, showing that highly branched and complex low MW structures ferment more slowly in the human intestine, facilitating their transport to the colon and with fewer side effects [119]. Damen et al., ( 2011) have also shown that AX with a high MW or DP can have a positive effect on the rate of fermentation and the production of short-chain fatty acid (SCFA) [117].
In addition to the complex structure, the fermentation process and product of AX in the gut are also closely related to the type of grain and its genotype. Zhang et al. (2021) suggested that the gut microbiota is subjected to changes in the fine structure of fiber chemistry determined by plant genotypes, and is not significantly influenced by the growth environment. Since AX extracted from corn bran of different corn genotypes have different branching structures, different changes are induced when fermented by the human gut microbiota, resulting in different SCFA synthesis [45]. Chen et al. (2017) observed that sorghum bran AXs fermented faster than CAX and that the branching structures of the two AXs differed, with AXs from sorghum bran having relatively uniform arabinose substitutions in the xylose backbone and AXs from corn bran having more complex glycan and chemical linkages [120]. In the results of the study by Yao et al. (2023) the more highly branched red sorghum bran AX produced more SCFA, and several metabolites, during colonic fermentation compared to white sorghum bran AX [121].
In general, AXs with low relative MW, low degrees of substitution, low FA content, and relatively simple structures have higher fermentation rates and less SCFA production, but due to the differences in AX structure and extraction process, the minor structural differences of AXs also strongly influence the metabolic results of fermentation, so it is difficult to fully discover the pattern of AX degradation and fermentation by the intestinal microbiota.
6.2. Improvement of Gut Health
AXs are important member of the prebiotic family that targets beneficial microbial species in the gut, and its fermentation and degradation in the colon may affect SCFA production as well as the abundance and composition of the gut microbiota. Fermentation of AX significantly increased Bifidobacterium and Lactobacillus populations and decreased Enterobacteriaceae populations [122]. In addition to Lactobacillus and Bifidobacterium spp, the fermentation of AX in the gut can also increase other bacterial groups, including Bacteroides, Prevotella, Roseburia, and Streptococcaceae. Although these groups of bacteria are not considered to be the primary targets of prebiotics, they have also been shown to have a strong relationship with human health [123].
Li et al. (2023) used an in vitro fermentation model to simulate the fermentation of WUAX (RAX and CAX) and their deglycosylated counterparts (dRAX and dCAX) extracted from rice bran and corn bran in the human gut and found that all types of AX significantly increased the relative abundance of Bacteroides, and that the most predominant SCFAs in the fermentation products after 48 h of fermentation were acetic and propionic acids, followed by butyric acid [30]. Likewise, Damen et al. (2011) evaluated gut microbiota composition and SCFA production in rats fed WUAX, WEAX, AXOS and their combinations. Results showed that consumption of WUAX-enriched diets increased cecal and colonic butyrate content and production, but did not stimulate cecal Bifidobacterium growth. An increase in bacteria, such as Roseburia and Eubacterium rectale spp., was also been reported when rats were fed WUAX diets with higher DP compared to WEAX, resulting in an increase in butyric acid levels in the cecum of rats [117]. Neyrinck et al. (2011) treated mice with a control diet (CT), a high-fat diet (HF), or a high-fat diet supplemented with AX (10% w/w) for four weeks. The results showed that AX restored the number of bacteria reduced by HF feeding, i.e. species of the Bacteroides-Prevotella spp. and Roseburia spp., and that the addition of AX also significantly increased the content of Bifidobacterium in the cecum [124]. Nguyen, et al. (2020) conducted a parallel two-group exploratory randomized controlled trial in 31 overweight and class I obese adults to investigate the composition of their fecal gut flora and SCFAs in the presence of high-dose long-chain AX supplementation. The results showed that AXs promoted specific intestinal flora classifications, including those associated with Bifidobacterium longum, Blautia obeum, and Prevotella copri, and further increased fecal propionic acid concentrations [125]. Truchado et al. (2017) investigated the modulatory effects of long-chain arabinoxylan (LC-AX) on the luminal and mucosal microbiota; the addition of 6 g/L LC-AX significantly increased the relative abundance of Bifidobacterium as well as propionic acid among others, in the lumen and mucus, compared to the control group [126].
Differences in the structure and origin of WUAX not only affect their physicochemical properties but also have different effects on the gut microbiota. Overall, the fermentation of WUAX in the gut promotes the production of beneficial bacteria while increasing the production of SCFA. Also, due to its complex structure such as its high DP, it is more effective than WEAX in interacting with gut microorganisms in some situations. The potential effects of AXs on host gut microbiota and health benefits are shown in Figure 4.
6.3. Alleviation of Metabolic Syndrome by Enhancement of Gut Health
The gut microbiota is a complex microbial ecosystem, and bacteria in the gut can ferment carbohydrates to produce substances such as SCFAs. These metabolites can act as a source of energy for colon cells and can increase feelings of fullness. Additionally, they have been shown to reduce inflammation, carcinogenesis, oxidative stress, and improve intestinal barrier function, all of which have a significant impact on human health [127,128]. As a typical dietary fiber, AX not only regulates the health of the human intestinal flora but also influences glucose and lipid metabolism and the occurrence of inflammation in the body.
Neyrinck, et al., (2011) demonstrated that dietary supplementation with a water-extractable concentrate of high MW AX counteracted high-fat diet-induced dysbiosis of the gut ecology, and improved adiposity and lipid-lowering effects. A significant increase in cecal Bifidobacterium, particularly Bifidobacterium animalis, was observed, and this effect was accompanied by improved gut barrier function and lower circulating inflammatory markers. More interestingly, rumenic acid was increased in white adipose tissue as a result of the AX treatment, suggesting that intestinal bacterial metabolism has an effect on host tissues. Thus, they suggest that AX may exert beneficial health effects by regulating the gut microbiota [124]. In another study by Chen et al. (2021), 50 male ICR/KM mice were randomly divided into 5 groups: Control diet (CON) group, High-fat diet (HFD) group, High-fat diet with AX (HFAX) group, High-fat diet with AX and β-glucan (HFAB) group and High fat diet with AX and xyloglucan (HFAG) group; the results showed that compared to HFD, HFAB, and HFAG, mice had lower body weight, cholesterol, and triglyceride levels; reduced bile acids, increased microbial species diversity, increased numbers of beneficial bacteria and reduced numbers of conditionally pathogenic bacteria [130]. Furthermore, research by Tong et al. (2014) showed that AX increased the production of SCFAs in the colon, leading to the excretion of lipids, cholesterol, and bile acids, ultimately reducing plasma total cholesterol and low-density lipoprotein (LDL) concentrations.
In addition, Luo et al. (2022) showed that mice supplemented with AX from rice bran (RAX) demonstrated significantly ameliorated obesity as induced by a high-fat diet, restoring intestinal microbiota alpha-diversity, increasing the relative abundance of anti-inflammatory bacteria such as Bifidobacterium and Akkermansia, and decreasing pro-inflammatory bacteria. These changes in the gut not only lead to a reduction in serum lipopolysaccharide (LPS) and systemic inflammation but ultimately to improved metabolic parameters and weight loss. The researchers believe that RAX is most likely able to restore host health by regulating the gut microbiome and its metabolite, SCFA [131]. Experimental results from Li et al., (2019) also support the possibility that AX may alter the colonic microbial metabolism by, for example, regulating intestinal flora and thereby improving the colonic mucosal barrier. After feeding rats diets containing AX for 5 weeks, the number of E. coli in the colonic digestive fluid decreased, while the number of Lactobacillus, Bifidobacterium, and Bacteroides increased, and serum lipopolysaccharides decreased in rats. Furthermore, AX upregulated the levels of genes for proteins associated with tight junctions in the colonic mucosa [132].
In conclusion, both WUAX and WEAX have potential health benefits, but their functional activity is mainly influenced by their structural characteristics and the source of the grain. The structural characteristics of AX may alter the interaction of AX with the gut microbiome, thereby affecting SCFA production and its effects on the host. AX has been shown to regulate the gut microbiome and its metabolites, leading to improvements in human health. However, the exact mechanism of action and its effects on gene expression relating to glucose and lipid metabolism are still unclear. It can be assumed that some of the effects of AX on human health are indeed achieved by regulating the gut microbiome and its metabolites.
7. Conclusions and Perspective
AXs are a significant group of dietary polysaccharides found in various cereals. Their structure is influenced by factors such as source, maturity, grain location, extraction and purification methods, and specific processing techniques. While WEAX has been extensively studied and shown to possess numerous beneficial properties, its low content in cereals poses challenges for industrial production. On the other hand, WUAX has a high content and availability, offering great potential for exploration. The chemical structure of AX greatly impacts the physical properties of food. Modifying WUAX in various ways can positively influence the food matrix. Additionally, WUAX has potential health benefits including modulation of SCFA in the colon, improved antioxidant capacity, and reduced blood glucose response through various mechanisms. Therefore, future research must focus on understanding the structure-activity relationship of WUAX. This involves investigating the correlation between the structure and function of AXs, comparing their activities in the same test system, and establishing specific principles. To enhance the utilization of WUAX, it is essential to develop gentle and harmless methods, such as microbial fermentation, to obtain AXs with specific structures. This would result in the release of more WUAX. By utilizing WUAX more effectively in food production, both the quality of food products and human health can be improved.
Author Contributions
Writing - Original Draft, Resources, Investigation, Visualization: Manchun Huang. Writing - Review & Editing: Juan Bai. Visualization, Writing - Review & Editing: Daniele Giuseppe Buccato. Writing - Review & Editing: JiaYan Zhang. Writing - Review & Editing: Yufeng He. Writing - Review & Editing: Ying Zhu. Visualization: Zihan Yang. Supervision, Funding Acquisition: Xiang Xiao. Data curation, Supervision: Maria Daglia.
Funding
This review was supported by the National Natural Science Foundation of China (32072200), the Research Project of Lactobacillus plantarum to promote the release mechanism of bound phenols in barley and Improve Energy Metabolism, and the Key Research project of Jiangsu Province (BE2022353), and the research project of the Key technology of efficient solid-state fermentation to improve the processing quality of barley whole powder. English Language support was provided by Eris Scott-Perring.
Data Availability Statement
Not available.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Wang, J., Bai, J., Fan, M., Li, T., Li, Y., Qian, H., Wang, L., Zhang, H., Qi, X., & Rao, Z. (2020). Cereal-derived arabinoxylans: Structural features and structure–activity correlations. Trends in Food Science & Technology, 96(C), 157-165. [CrossRef]
- Bakken, T., Braaten, T., Olsen, A., Kyrø, C., Lund, E., & Skeie, G. (2016). Consumption of Whole-Grain Bread and Risk of Colorectal Cancer among Norwegian Women (the NOWAC Study). Nutrients, 8(1), 40. [CrossRef]
- Han, F., Wang, Y., Han, Y., Zhao, J., Han, F., Song, G., Jiang, P., & Miao, H. (2018). Effects of Whole-Grain Rice and Wheat on Composition of Gut Microbiota and Short-Chain Fatty Acids in Rats. Journal of agricultural and food chemistry, 66(25), 6326-6335. [CrossRef]
- Ullah H, Esposito C, Piccinocchi R, De Lellis LF, Santarcangelo C, Minno AD, Baldi A, Buccato DG, Khan A, Piccinocchi G, Sacchi R, Daglia M. Postprandial Glycemic and Insulinemic Response by a Brewer's Spent Grain Extract-Based Food Supplement in Subjects with Slightly Impaired Glucose Tolerance: A Monocentric, Randomized, Cross-Over, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients. 2022 Sep 21;14(19):3916. [CrossRef]
- Kim, H., Yu, K.-W., Hong, H.-D., & Shin, K.-S. (2017). Effect of arabinoxylan- and rhamnogalacturonan I-rich polysaccharides isolated from young barley leaf on intestinal immunostimulatory activity. Journal of Functional Foods, 35, 384-390. [CrossRef]
- Kyrø, C., Tjønneland, A., Overvad, K., Olsen, A., & Landberg, R. (2018). Higher Whole-Grain Intake Is Associated with Lower Risk of Type 2 Diabetes among Middle-Aged Men and Women: The Danish Diet, Cancer, and Health Cohort. The Journal of Nutrition, 148(9), 1434-1444. [CrossRef]
- Malin, S. K., Kullman, E. L., Scelsi, A. R., Godin, J.-P., Ross, A. B., & Kirwan, J. P. (2019). A Whole-Grain Diet Increases Glucose-Stimulated Insulin Secretion Independent of Gut Hormones in Adults at Risk for Type 2 Diabetes. Molecular nutrition & food research, 63(7), 1800967. [CrossRef]
- Wu, H., Flint, A. J., Qi, Q., Dam, R. M. v., Sampson, L. A., Rimm, E. B., Holmes, M. D., Willett, W. C., Hu, F. B., & Sun, Q. (2015). Association Between Dietary Whole Grain Intake and Risk of Mortality: Two Large Prospective Studies in US Men and Women. JAMA Internal Medicine, 175(3), 373-384. [CrossRef]
- Xu, Y., Wan, Q., Feng, J., Du, L., Li, K., & Zhou, Y. (2018). Whole grain diet reduces systemic inflammation: A meta-analysis of 9 randomized trials. Medicine, 97(43), e12995. [CrossRef]
- Xu, Y., Yang, J., Du, L., Li, K., & Zhou, Y. (2019). Association of whole grain, refined grain, and cereal consumption with gastric cancer risk: A meta-analysis of observational studies. Food science & nutrition, 7(1), 256-265. [CrossRef]
- Saulnier, L., Sado, P.-E., Branlard, G., Charmet, G., & Guillon, F. (2007). Wheat arabinoxylans: Exploiting variation in amount and composition to develop enhanced varieties. Journal of Cereal Science, 46(3), 261-281. [CrossRef]
- Wang, M., Zhao, Z., Niu, M., Zhao, S., Jia, C., & Wu, Y. (2020). Thermomechanical behaviors and protein polymerization in bread dough modified by bran components and transglutaminase LWT, 133, 109894. [CrossRef]
- Frederix, S. A., Van hoeymissen, K. E., Courtin, C. M., & Delcour, J. A. (2004). Water-extractable and water-unextractable arabinoxylans affect gluten agglomeration behavior during wheat flour gluten-starch separation. Journal of agricultural and food chemistry, 52(26), 7950-7956. [CrossRef]
- Rumpagaporn, P., Reuhs, B. L., Kaur, A., Patterson, J. A., Keshavarzian, A., & Hamaker, B. R. (2015). Structural features of soluble cereal arabinoxylan fibers associated with a slow rate of in vitro fermentation by human fecal microbiota. Carbohydrate Polymers, 130, 191-197. [CrossRef]
- Lin, S., Agger, J. W., Wilkens, C., & Meyer, A. S. (2021). Feruloylated Arabinoxylan and Oligosaccharides: Chemistry, Nutritional Functions, and Options for Enzymatic Modification. Annual review of food science and technology, 12, 331-354. [CrossRef]
- Döring, C., Hussein, M. A., Jekle, M., & Becker, T. (2017). On the assessments of arabinoxylan localization and enzymatic modifications for enhanced protein networking and its structural impact on rye dough and bread. Food Chemistry. [CrossRef]
- Fadel, A., Mahmoud, A. M., Ashworth, J. J., Li, W., Ng, Y. L., & Plunkett, A. (2018). Health-related effects and improving extractability of cereal arabinoxylans. International Journal of Biological Macromolecules, 109, 819-831. [CrossRef]
- Fry, S. C. (2004). Primary Cell Wall Metabolism: Tracking the Careers of Wall Polymers in Living Plant Cells. New Phytologist, 161(3), 641-675. [CrossRef]
- Yadav, M. P., Kale, M. S., Hicks, K. B., & Hanah, K. (2017). Isolation, characterization and the functional properties of cellulosic arabinoxylan fiber isolated from agricultural processing by-products, agricultural residues and energy crops. Food Hydrocolloids, 63, 545-551. [CrossRef]
- Chen, Z., Li, S., Fu, Y., Li, C., Chen, D., & Chen, H. (2019). Arabinoxylan structural characteristics, interaction with gut microbiota and potential health functions. Journal of Functional Foods, 54, 536-551. [CrossRef]
- Ganguli, N. K., & Turner, M. A. (2008). A simplified method for extracting water-extractable arabinoxylans from wheat flour. Journal of the Science of Food and Agriculture, 88(11), 1905-1910. [CrossRef]
- Izydorczyk, M. S., & Biliaderis, C. G. (1992). Influence of structure on the physicochemical properties of wheat arabinoxylan. Carbohydrate Polymers, 17(3), 237-247.
- Kurt, G., Emmie, D., Danuta, B., Anna, F., Wioletta, D., Zoltan, B., Mariann, R., A, D. J., & M, C. C. (2008). Variation in the content of dietary fiber and components thereof in wheats in the HEALTHGRAIN Diversity Screen. Journal of agricultural and food chemistry, 56(21), 9740-9749. [CrossRef]
- Zhang, Z., Smith, C., & Li, W. (2014). Extraction and modification technology of arabinoxylans from cereal by-products: A critical review. Food Research International, 65, 423-436. [CrossRef]
- Zhu, Y., Li, F., Wang, Y., Li, J., Teng, C., Wang, C., & Li, X. (2019). Effects of different MW water-extractable arabinoxylans on the physicochemical properties and structure of wheat gluten. Journal of food science and technology, 56(1), 340-349. [CrossRef]
- Izydorczyk, M. S., & Biliaderis, C. G. (1995). Cereal arabinoxylans: advances in structure and physicochemical properties. Carbohydrate Polymers, 28(1), 33-48. [CrossRef]
- Nishitsuji, Y., Whitney, K., Nakamura, K., Hayakawa, K., & Simsek, S. (2020b). Changes in structure and solubility of wheat arabinoxylan during the breadmaking process %J Food Hydrocolloids. 109(prepublish).
- Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K. S., Manichanh, C., Nielsen, T., Pons, N., Levenez, F., Yamada, T., Mende, D. R., Li, J., X, J., Li, S., Li, D., Cao, J., Wang, B., Liang, H., Zheng, H., Xie, Y., Tap, J., Lepag, P., Bertalan, M., Batto, J.-M., Consortium, M., & Wang, J. (2010). A human gut microbial gene catalogue established by metagenomic sequencing. Nature, 464(7285), 59-65. [CrossRef]
- Emiliano, S. (2014). Microbiome, holobiont and the net of life. Critical Reviews in Microbiology, 42(3), 485-494. [CrossRef]
- Li, Z., Zhang, H., He, L., Hou, Y., Che, Y., Liu, T., Xiong, S., Zhang, X., Luo, S., Liu, C., & Chen, T. (2023). Influence of structural features and feruloylation on fermentability and ability to modulate gut microbiota of arabinoxylan in in vitro fermentation. Frontiers in Microbiology, 13, 1113601. [CrossRef]
- Paesani, C., Sciarini, L. S., Moiraghi, M., Salvucci, E., Prado, S. B. R., & P, G. T. (2020). Human colonic in vitro fermentation of water-soluble arabinoxylans from hard and soft wheat alters Bifidobacterium abundance and short-chain fatty acids concentration. LWT, 134, 110253. [CrossRef]
- Valerie, V. C., Katrien, S., Emmie, D., Tom, V. d. W., Massimo, M., Willy, V., Yasmine, D., Okanlawon, O., Eddy, D., Johan, B., Bart, D. K., F, B. W., A, D. J., & M, C. C. (2008). Structurally different wheat-derived arabinoxylooligosaccharides have different prebiotic and fermentation properties in rats. The Journal of Nutrition, 138(12), 2348-2355. [CrossRef]
- Chateigner-Boutin. (2016). Developing Pericarp of Maize: A Model to Study Arabinoxylan Synthesis and Feruloylation. Frontiers in Plant Science, 7, 1476. [CrossRef]
- Broekaert, W. F., Courtin, C. M., Verbeke, K., Wiele, T. V. d., Verstraete, W., & Delcour, J. A. (2011). Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Critical Reviews in Food Science and Nutrition, 51(2), 178-194. [CrossRef]
- Escarnot, E., Aguedo, M., Agneessens, R., Wathelet, B., & Paquot, M. (2011). Extraction and characterization of water-extractable and water-unextractable arabinoxylans from spelt bran: Study of the hydrolysis conditions for monosaccharides analysis. Journal of Cereal Science, 53(1), 45-52. [CrossRef]
- Niño-Medina, G., Carvajal-Millán, E., Rascon-Chu, A., Marquez-Escalante, J. A., Guerrero, V., & Salas-Muñoz, E. (2010). Feruloylated arabinoxylans and arabinoxylan gels: structure, sources and applications. Phytochemistry Reviews, 9(1), 111-120. [CrossRef]
- Buksa, K., Nowotna, A., Ziobro, R., & Praznik, W. (2014). Molecular properties of arabinoxylan fractions isolated from rye grain of different quality. Journal of Cereal Science, 60(2), 368-373. [CrossRef]
- Revanappa, S. B., Nandini, C. D., & Salimath, P. V. (2015). Structural variations of arabinoxylans extracted from different wheat ( Triticum aestivum ) cultivars in relation to chapati -quality. Food Hydrocolloids, 43, 736-742. [CrossRef]
- Sun, Y., Cui, S. W., Gu, X., & Zhang, J. (2011). Isolation and structural characterization of water unextractable arabinoxylans from Chinese black-grained wheat bran. Carbohydrate Polymers, 85(3), 615-621. [CrossRef]
- Tian, L., Gruppen, H., & Schols, H. A. (2015). Characterization of (Glucurono)arabinoxylans from Oats Using Enzymatic Fingerprinting. Journal of agricultural and food chemistry, 63(50), 10822-10830. [CrossRef]
- Pang, J., Zhang, Y., Tong, X., Zhong, Y., Kong, F., Li, D., Liu, X., & Qiao, Y. (2023). Recent Developments in Molecular Characterization, Bioactivity, and Application of Arabinoxylans from Different Sources. Polymers, 15(1), 225. [CrossRef]
- Koegelenberg, D., & Chimphango, A. F. A. (2017). Effects of wheat-bran arabinoxylan as partial flour replacer on bread properties Food Chemistry, 221, 1606-1613. [CrossRef]
- Casas, G. A., Lærke, H. N., Knudsen, K. E. B., & Stein, H. H. (2018). Arabinoxylan is the main polysaccharide in fiber from rice coproducts, and increased concentration of fiber decreases in vitro digestibility of dry matter. Animal Feed Science and Technology, 247, 255-261. [CrossRef]
- Guo, R., Xu, Z., Wu, S., Li, X., Li, J., Hu, H., Wu, Y., & Ai, L. (2019). Molecular properties and structural characterization of an alkaline extractable arabinoxylan from hull-less barley bran. Carbohydrate Polymers, 218, 250-260. [CrossRef]
- Zhang, X., Xie, J., Chen, T., Ma, D., Yao, T., Gu, F., Lim, J., Tuinstra, M. R., & Hamaker, B. R. (2021). High Arabinoxylan Fine Structure Specificity to Gut Bacteria Driven by Corn Genotypes but not Environment. Carbohydrate Polymers, 257, 117667. [CrossRef]
- Han, J.-Y. (2000). Structural characteristics of arabinoxylan in barley, malt, and beer. Food Chemistry, 70(2), 131-138. [CrossRef]
- Rao, R. S. P., & Muralikrishna, G. (2007). Structural characteristics of water-soluble feruloyl arabinoxylans from rice ( Oryza sativa ) and ragi (finger millet, Eleusine coracana ): Variations upon malting. Food Chemistry, 104(3), 1160-1170. [CrossRef]
- Höije, A., Sandström, C., Roubroeks, J. P., Andersson, R., Gohil, S., & Gatenholm, P. (2006). Evidence of the presence of 2-O-β-d-xylopyranosyl-α-l-arabinofuranose side chains in barley husk arabinoxylan. Carbohydrate Research, 341(18), 2959-2966. [CrossRef]
- Guo, Q., Xiao, X., Li, C., Kang, J., Liu, G., Douglas, G. H., & Wang, C. (2021). Catechin-grafted arabinoxylan conjugate: Preparation, structural characterization and property investigation. International Journal of Biological Macromolecules, 182, 796-805. [CrossRef]
- Anna, E., Zdenka, H., Eva, P., & Miloš, H. (1990). Structural features of a water-soluble l-arabino-d-xylan from rye bran. Carbohydrate Research, 198(1), 57-66. [CrossRef]
- Li, Y., & Yang, C. (2016). Synthesis and properties of feruloyl corn bran arabinoxylan esters. International journal of cosmetic science, 38(3), 238-245. [CrossRef]
- Andreasen, M. F., Kroon, P. A., Williamson, G., & Garcia-Conesa, M. T. (2001). Esterase activity able to hydrolyze dietary antioxidant hydroxycinnamates is distributed along the intestine of mammals. Journal of agricultural and food chemistry, 49(11), 5679-5684. [CrossRef]
- Veenashri, B. R., & Muralikrishna, G. (2011a). In vitro anti-oxidant activity of xylo-oligosaccharides derived from cereal and millet brans – A comparative study. Food Chemistry, 126(3), 1475-1481. [CrossRef]
- Srinivasan, A., Aruldhas, J., Perumal, S. S., & Ekambaram, S. P. (2020). Phenolic acid bound arabinoxylans extracted from Little and Kodo millets modulate immune system mediators and pathways in RAW 264.7 cells. Journal of Food Biochemistry, 45(1), e13563. [CrossRef]
- Srinivasan, A., Ekambaram, S. P., Perumal, S. S., Aruldhas, J., & Erusappan, T. (2019). Chemical characterization and immunostimulatory activity of phenolic acid bound arabinoxylans derived from foxtail and barnyard millets. Journal of Food Biochemistry, 44(2), e13116. [CrossRef]
- Bijalwan, V., Ali, U., Kesarwani, A. K., Yadav, K., & Mazumder, K. (2016). Hydroxycinnamic acid bound arabinoxylans from millet brans-structural features and antioxidant activity. International Journal of Biological Macromolecules, 88, 296-305. [CrossRef]
- Ayala-Soto, F. E., Serna-Saldívar, S. O., Welti-Chanes, J., & Gutierrez-Uribe, J. A. (2015). Phenolic compounds, antioxidant capacity and gelling properties of glucoarabinoxylans from three types of sorghum brans. Journal of Cereal Science, 65, 277-284. [CrossRef]
- He, H.-J., Qiao, J., Liu, Y., Guo, Q., Ou, X., & Wang, X. (2021). Isolation, Structural, Functional, and Bioactive Properties of Cereal Arabinoxylan─A Critical Review. Journal of agricultural and food chemistry, 69(51), 15437-15457. [CrossRef]
- Li, Y., Zhu, Y., Liang, R., & Yang, C. (2017). Synthesis and antioxidant properties of caffeic acid corn bran arabinoxylan esters. International journal of cosmetic science, 39(4), 402-410. [CrossRef]
- Ayala-Soto, F. E., Serna-Saldívar, S. O., García-Lara, S., & Pérez-Carrillo, E. (2014). Hydroxycinnamic acids, sugar composition and antioxidant capacity of arabinoxylans extracted from different maize fiber sources. Food Hydrocolloids, 35, 471-475. [CrossRef]
- Anderson, C., & Simsek, S. (2019). A novel combination of methods for the extraction and purification of arabinoxylan from byproducts of the cereal industry Journal of Food Measurement and Characterization, 13(2), 1049-1057. [CrossRef]
- Petit-Benvegnen, M. D., Saulnier, L., & Rouau, X. (1998). Solubilization of Arabinoxylans from Isolated Water-Unextractable Pentosans and Wheat Flour Doughs by Cell-Wall-Degrading Enzymes. Cereal Chemistry, 75(4), 551-556. [CrossRef]
- Ruthes, A. C., Martínez-Abad, A., Tan, H.-T., Bulone, V., & Vilaplana, F. (2017). Sequential fractionation of feruloylated hemicelluloses and oligosaccharides from wheat bran using subcritical water and xylanolytic enzymes. Green Chemistry, 19(8), 1919-1931. [CrossRef]
- Yilmaz-Turan, S., Jiménez-Quero, A., Menzel, C., Carvalho, D. M. d., Lindström, M. E., Sevastyanova, O., Moriana, R., & Vilaplana, F. (2020). Bio-based films from wheat bran feruloylated arabinoxylan: Effect of extraction technique, acetylation and feruloylation Carbohydrate Polymers, 250, 116916. [CrossRef]
- Hollmann, J., & Lindhauer, M. G. (2004). Pilot-scale isolation of glucuronoarabinoxylans from wheat bran. Carbohydrate Polymers, 59(2), 225-230. [CrossRef]
- Ruthes, A. C., Rudjito, R. C., Rencoret, J., Gutiérrez, A., Río, J. C. d., Jiménez-Quero, A., & Vilaplana, F. (2020). Comparative Recalcitrance and Extractability of Cell Wall Polysaccharides from Cereal (Wheat, Rye, and Barley) Brans Using Subcritical Water ACS Sustainable Chemistry & Engineering, 8(18), 7192-7204. [CrossRef]
- Zhou, S., Liu, X., Guo, Y., Wang, Q., Peng, D., & Cao, L. (2010). Comparison of the immunological activities of arabinoxylans from wheat bran with alkali and xylanase-aided extraction. Carbohydrate Polymers, 81(4), 784-789. [CrossRef]
- Kale, M. S., Yadav, M. P., Hicks, K. B., & Hanah, K. (2015). Concentration and shear rate dependence of solution viscosity for arabinoxylans from different sources. Food Hydrocolloids, 47, 178-183. [CrossRef]
- Kumar, D. P., Maire, d. P. A., Anja, L., Anette, L., Anna, S., & Gunnar, W. (2021). Oxidation Level and Glycidyl Ether Structure Determine Thermal Processability and Thermomechanical Properties of Arabinoxylan-Derived Thermoplastics ACS APPLIED BIO MATERIALS, 4(4), 3133-3144. [CrossRef]
- Wang, X., Zheng, X., & Li, L. (2011). Molecular Characterization of Arabinoxylans from Hull-Less Barley Milling Fractions. Molecules, 16(4), 2743-2753. [CrossRef]
- Li, L. Y., Wang, Y. X., Zhang, T., Zhang, J. F., Pan, M., Huang, X. J., Yin, J. Y., & Nie, S. P. (2020). Structural characteristics and rheological properties of alkali-extracted arabinoxylan from dehulled barley kernel. Carbohydrate Polymers, 249, 116813. [CrossRef]
- Takoudjou, M. A.-P., Gudipati, M., Bargui, K. B., & Germain, K. (2021). Purification and structural characterization of calcium hydroxide isolated arabinoxylans derived from bran, spent grain and sorghum grains. Journal of Cereal Science, 100, 103266. [CrossRef]
- Yan, J., Jia, X., Feng, L., Yadav, M., Li, X., & Yin, L. (2019). Rheological and emulsifying properties of arabinoxylans from various cereal brans Journal of Cereal Science, 90(C), 102844. [CrossRef]
- Li, S., Chen, H., Cheng, W., Yang, K., Cai, L., He, L., Du, L., Liu, Y., Liu, A., Zeng, Z., & Li, C. (2021). Impact of arabinoxylan on characteristics, stability and lipid oxidation of oil-in-water emulsions: Arabinoxylan from wheat bran, corn bran, rice bran, and rye bran Food Chemistry, 358, 129813. [CrossRef]
- Pihlajaniemi, V., Mattila, O., Koitto, T., Nikinmaa, M., Heiniö, R.-L., Sorsamäki, L., Siika-aho, M., & Nordlund, E. (2020). Production of syrup rich in arabinoxylan oligomers and antioxidants from wheat bran by alkaline pretreatment and enzymatic hydrolysis, and applicability in baking Journal of Cereal Science, 95, 103043. [CrossRef]
- Gruppen, H., Kormelink, F. J. M., & Voragen, A. G. J. (1993). Water-unextractable Cell Wall Material from Wheat Flour. 3. A Structural Model for Arabinoxylans. Journal of Cereal Science, 18(2), 111-128. [CrossRef]
- Buksa, K., Nowotna, A., & Ziobro, R. (2016). Application of cross-linked and hydrolyzed arabinoxylans in baking of model rye bread Food Chemistry, 192, 991-996. [CrossRef]
- Aguedo, M., Fougnies, C., Dermience, M., & Richel, A. (2014). Extraction by three processes of arabinoxylans from wheat bran and characterization of the fractions obtained. Carbohydrate Polymers, 105, 317-324. [CrossRef]
- Denisse, B., Renata, N., Michaela, W., Sylvia, G., Matilde, B., Kitti, T., Sandor, T., Stefano, D. A., & Regine, S. (2017). Optimization of Arabinoxylan Isolation from Rye Bran by Adapting Extraction Solvent and Use of Enzymes. Journal of food science, 82(11), 2562-2568. [CrossRef]
- Mathew, S., Karlsson, E. N., & Adlercreutz, P. (2017). Extraction of soluble arabinoxylan from enzymatically pretreated wheat bran and production of short xylo-oligosaccharides and arabinoxylo-oligosaccharides from arabinoxylan by glycoside hydrolase family 10 and 11 endoxylanases. Journal of Biotechnology, 260, 53-61. [CrossRef]
- Zhang, S., Li, W., Smith, C. J., & Musa, H. (2015). Cereal-derived arabinoxylans as biological response modifiers: extraction, molecular features, and immune-stimulating properties. Critical Reviews in Food Science and Nutrition, 55(8), 1035-1052. [CrossRef]
- Wang, J., Sun, B., Liu, Y., & Zhang, H. (2014). Optimisation of ultrasound-assisted enzymatic extraction of arabinoxylan from wheat bran. Food Chemistry, 150, 482-488. [CrossRef]
- Ordaz-Ortiz, J. J., & Saulnier, L. (2005). Structural variability of arabinoxylans from wheat flour. Comparison of water-extractable and xylanase-extractable arabinoxylans. Journal of Cereal Science, 42(1), 119-125. [CrossRef]
- Ma, F., Li, X., Yin, J., Ma, L., & Li, D. (2020). Optimisation of double-enzymatic extraction of arabinoxylan from fresh corn fibre. Journal of food science and technology, 57(12), 4649-4659. [CrossRef]
- Clemens, D., Mario, J., & Thomas, B. (2016). Technological and Analytical Methods for Arabinoxylan Quantification from Cereals. Critical Reviews in Food Science and Nutrition, 56(6), 999-1011. [CrossRef]
- Benamrouche, S., Crônier, D., Debeire, P., & Chabbert, B. (2002). A Chemical and Histological Study on the Effect of (1→4)-β-endo-xylanase Treatment on Wheat Bran. Journal of Cereal Science, 36(2), 253-260. [CrossRef]
- Dornez, E., Gebruers, K., Delcour, J. A., & Courtin, C. M. (2009). Grain-associated xylanases: occurrence, variability, and implications for cereal processing. Trends in Food Science & Technology, 20(11), 495-510. [CrossRef]
- Courtin, C. M., & Delcour, J. A. (2002). Arabinoxylans and Endoxylanases in Wheat Flour Bread-making. Journal of Cereal Science, 35(3), 225-243. [CrossRef]
- Verwimp, T., Craeyveld, V. V., Courtin, C. M., & Delcour, J. A. (2007). Variability in the structure of rye flour alkali-extractable arabinoxylans. Journal of agricultural and food chemistry, 55(5), 1985-1992. [CrossRef]
- Kale, M. S., Yadav, M. P., Chau, H. K., & Hotchkiss, A. T. (2018). Molecular and functional properties of a xylanase hydrolysate of corn bran arabinoxylan. Carbohydrate Polymers, 181, 119-123. [CrossRef]
- Li, W., Zhang, S., & Smith, C. (2015). The molecular structure features-immune stimulatory activity of arabinoxylans derived from the pentosan faction of wheat flour. Journal of Cereal Science, 62, 81-86. [CrossRef]
- Yuwang, P., Sulaeva, I., Hell, J., Henniges, U., Böhmdorfer, S., Rosenau, T., Chitsomboon, B., & Tongta, S. (2017). Phenolic compounds and antioxidant properties of arabinoxylan hydrolysates from defatted rice bran. Journal of the Science of Food and Agriculture, 98(1), 140-146. [CrossRef]
- Maes, C., Vangeneugden, B., & Delcour, J. A. (2004). Relative activity of two endoxylanases towards water-unextractable arabinoxylans in wheat bran. Journal of Cereal Science, 39(2), 181-186. [CrossRef]
- Marilù, D., Mattia, Q., Giovanni, R., Grazia, F. M., Milena, B., Tiziana, S., Federica, M., Daniela, E., Franca, C., & Cristina, C. M. (2017). Evaluation of microbial consortia and chemical changes in spontaneous maize bran fermentation. . AMB Express, 7(1), 1-13. [CrossRef]
- Hell, J., Donaldson, L., Michlmayr, H., Kraler, M., Kneifel, W., Potthast, A., Rosenau, T., & Böhmdorfer, S. (2015). Effect of pretreatment on arabinoxylan distribution in wheat bran. Carbohydrate Polymers, 121, 18-26. [CrossRef]
- Jakovetić, T. S., Nataša, Š., Jelena, J., Ivana, G., Sanja, G., Nikola, Đ., Vukašinović, S. M., Jian, H., Nevena, L., & Zorica, K. J. (2020). Upgrading of valuable food component contents and anti-nutritional factors depletion by solid-state fermentation: A way to valorize wheat bran for nutrition Journal of Cereal Science, 99, 103159. [CrossRef]
- Kiszonas, A. M., Fuerst, E. P., Luthria, D., & Morris, C. F. (2015). Arabinoxylan content and characterisation throughout the bread-baking process. International Journal of Food Science & Technology, 50(8), 1911-1921. [CrossRef]
- Nishitsuji, Y., Whitney, K., Nakamura, K., Hayakawa, K., & Simsek, S. (2020a). Changes in structure and solubility of wheat arabinoxylan during the breadmaking process. Food Hydrocolloids, 109, 106129. [CrossRef]
- Lynch, K. M., Strain, C. R., Johnson, C., Patangia, D., Stanton, C., Koc, F., Gil-Martinez, J., O’Riordan, P., Sahin, A. W., Ross, R. P., & Arendt, E. K. (2021). Extraction and characterisation of arabinoxylan from brewers spent grain and investigation of microbiome modulation potential. . European journal of nutrition, 60(8), 4393-4411. [CrossRef]
- Cervantes-Ramirez, J. G., Vasquez-Lara, F., Vasquez-Lara, F., Scholar, S. o., Sanchez-Estrada, A., Troncoso-Rojas, R., Heredia-Olea, E., & Islas-Rubio, A. R. (2022). Arabinoxylans Release from Brewers’ Spent Grain Using Extrusion and Solid-State Fermentation with Fusarium oxysporum and the Antioxidant Capacity of the Extracts Foods, 11(10), 1415. [CrossRef]
- Wang, M., Vliet, T. v., & Hamer, R. J. (2004). How gluten properties are affected by pentosans. Journal of Cereal Science, 39(3), 395-402. [CrossRef]
- Si, X., Li, T., Zhang, Y., Zhang, W., Qian, H., Li, Y., Zhang, H., Qi, X., & Wang, L. (2021). Interactions between gluten and water-unextractable arabinoxylan during the thermal treatment. Food Chemistry, 345, 128785. [CrossRef]
- Verjans, P., Dornez, E., Delcour, J. A., & Courtin, C. M. (2010). Selectivity for water-unextractable arabinoxylan and inhibition sensitivity govern the strong bread improving potential of an acidophilic GH11 Aureobasidium pullulans xylanase. Food Chemistry, 123(2), 331-337. [CrossRef]
- Jiang, Z., Liu, L., Yang, W., Ding, L., Awais, M., Wang, L., & Zhou, S. (2018). Improving the physicochemical properties of whole wheat model dough by modifying the water-unextractable solids. Food Chemistry, 259, 18-24. [CrossRef]
- Sun, J., Si, X., Li, T., Zhao, J., Qian, H., Li, Y., Zhang, H., Qi, X., & Wang, L. (2022). The Influence of Water-Unextractable Arabinoxylan and Its Hydrolysates on the Aggregation and Structure of Gluten Proteins. Frontiers in Nutrition, 9, 877135. [CrossRef]
- Arif, S., Ahmed, M., Chaudhry, Q., & Hasnain, A. (2018). Effects of water extractable and unextractable pentosans on dough and bread properties of hard wheat cultivars. LWT, 97, 736-742. [CrossRef]
- Espinosa-Ramírez, J., Garzon, R., Serna-Saldivar, S. O., & Rosell, C. M. (2020). Exploring the potential of arabinoxylan as structuring agent in model systems for gluten-free yeast-leavened breads Journal of Cereal Science, 95, 103080. [CrossRef]
- Zhang, L., Boven, A. v., Mulder, J., Grandia, J., Chen, X. D., Boom, R. M., & Schutyser, M. A. I. (2019). Arabinoxylans-enriched fractions: From dry fractionation of wheat bran to the investigation on bread baking performance Journal of Cereal Science, 87, 1-8. [CrossRef]
- Yang, M., Yue, Y., Liu, L., Tong, L., Wang, L., Ashraf, J., Li, N., Zhou, X., & Zhou, S. (2021). Investigation of combined effects of xylanase and glucose oxidase in whole wheat buns making based on reconstituted model dough system. LWT, 135, 110261. [CrossRef]
- Kaur, A., Yadav, M. P., Singh, B., Bhinder, S., Simon, S., & Singh, N. (2019). Isolation and characterization of arabinoxylans from wheat bran and study of their contribution to wheat flour dough rheology Carbohydrate Polymers, 221, 166-173. [CrossRef]
- Pavlovich-Abril, A., Rouzaud-Sández, O., Carvajal-Millán, E., Navarro, R. E., Robles-Sánchez, R. M., & Barrón-Hoyos, J. M. (2016). Molecular characterization of water extractable arabinoxylans isolated from wheat fine bran and their effect on dough viscosity LWT - Food Science and Technology, 74, 484-492. [CrossRef]
- Guo, X. N., Yang, S., & Zhu, K. X. (2018). Impact of arabinoxylan with different MW on the thermo-mechanical, rheological, water mobility and microstructural characteristics of wheat dough International Journal of Food Science & Technology, 53(9), 2150-2158. [CrossRef]
- Pritchard, J. R., Lawrence, G. J., Larroque, O., Li, Z., Laidlaw, H. K., Morell, M. K., & Rahman, S. (2011). A survey of β-glucan and arabinoxylan content in wheat. Journal of the Science of Food and Agriculture, 91(7), 1298-1303. [CrossRef]
- Janssen, F., Wouters, A. G. B., Meeus, Y., Moldenaers, P., Vermant, J., & Delcour, J. A. (2020). The role of non-starch polysaccharides in determining the air-water interfacial properties of wheat, rye, and oat dough liquor constituents Food Hydrocolloids, 105(C), 105771. [CrossRef]
- Ghoshal, G., Shivhare, U. S., & Banerjee, U. C. (2013). Effect of Xylanase on Quality Attributes of Whole-wheat bread. Journal of Food Quality, 36(3), 172-180. [CrossRef]
- Mendis, M., Leclerc, E., & Simsek, S. (2016). Arabinoxylans, gut microbiota and immunity Carbohydrate Polymers, 139, 159-166. [CrossRef]
- Damen, B., Verspreet, J., Pollet, A., Broekaert, W. F., Delcour, J. A., & Courtin, C. M. (2011). Prebiotic effects and intestinal fermentation of cereal arabinoxylans and arabinoxylan oligosaccharides in rats depend strongly on their structural properties and joint presence. . Molecular nutrition & food research, 55(12), 1862-1874. [CrossRef]
- Rose, D. J., Patterson, J. A., & Hamaker, B. R. (2010). Structural differences among alkali-soluble arabinoxylans from maize (Zea mays), rice (Oryza sativa), and wheat (Triticum aestivum) brans influence human fecal fermentation profiles. . Journal of agricultural and food chemistry, 58(1), 493-499. [CrossRef]
- Xu, H., Reuhs, B. L., Cantu-Jungles, T. M., Tuncil, Y. E., Kaur, A., Terekhov, A., Martens, E. C., & Hamaker, B. R. (2022). Corn arabinoxylan has a repeating structure of subunits of high branch complexity with slow gut microbiota fermentation Carbohydrate Polymers, 289, 119435. [CrossRef]
- Chen, T., Long, W., Zhang, C., Liu, S., Zhao, L., & Hamaker, B. R. (2017). Fiber-utilizing capacity varies in Prevotella- versus Bacteroides-dominated gut microbiota. Scientific Reports, 7(1), 2594. [CrossRef]
- Yao, T., Deemer, D. G., Chen, M.-H., Reuhs, B. L., Hamaker, B. R., & Lindemann, S. R. (2023). Differences in fine arabinoxylan structures govern microbial selection and competition among human gut microbiota. Carbohydrate Polymers, 316, 121039. [CrossRef]
- Martínez-López, A. L., Carvajal-Millan, E., Canett-Romero, R., Prakash, S., Rascón-Chu, A., López-Franco, Y. L., Lizardi-Mendoza, J., & Micard, V. (2022). Arabinoxylans-Based Oral Insulin Delivery System Targeting the Colon: Simulation in a Human Intestinal Microbial Ecosystem and Evaluation in Diabetic Rats. Pharmaceuticals, 15(9), 1062. [CrossRef]
- Emily, S., Sok Cheon, P., Shaoyu, W., Peter, S. M., Thomas, J., Soo Liang, O., Terry, G., Garth, H., & Emad, E.-O. (2021). The effects and benefits of arabinoxylans on human gut microbiota – A narrative review. Food Bioscience, 101267. [CrossRef]
- Neyrinck, A. M., Possemiers, S., Druart, C., Wiele, T. V. d., Backer, F. D., Cani, P. D., Larondelle, Y., & Delzenne, N. M. (2011). Prebiotic effects of wheat arabinoxylan related to the increase in bifidobacteria, Roseburia and Bacteroides/Prevotella in diet-induced obese mice. . PloS one, 6(6), e20944. [CrossRef]
- Nguyen, N. K., Deehan, E. C., Zhang, Z., Jin, M., Baskota, N., Perez-Muñoz, M. E., Cole, J., Tuncil, Y. E., Seethaler, B., Wang, T., Laville, M., Delzenne, N. M., Bischoff, S. C., Hamaker, B. R., Martínez, I., Knights, D., Bakal, J. A., Prado, C. M., & Walter, J. (2020). Gut microbiota modulation with long-chain corn bran arabinoxylan in adults with overweight and obesity is linked to an individualized temporal increase in fecal propionate. . Microbiome, 8(1), 1-21. [CrossRef]
- Truchado, P., Hernandez-Sanabria, E., Salden, B. N., Abbeele, P. V. d., Vilchez-Vargas, R., Jauregui, R., Pieper, D. H., Possemiers, S., & Wiele, T. V. d. (2017). Long chain arabinoxylans shift the mucosa-associated microbiota in the proximal colon of the simulator of the human intestinal microbial ecosystem (M-SHIME) Journal of Functional Foods, 32, 226-237. [CrossRef]
- Zhao, L. (2013). The gut microbiota and obesity: from correlation to causality. . Nature reviews. Microbiology, 11(9), 639-647. [CrossRef]
- Zhao, L., Zhang, F., Ding, X., Wu, G., Lam, Y. Y., Wang, X., Fu, H., Xue, X., Lu, C., Ma, J., Yu, L., Xu, C., Ren, Z., Xu, Y., Xu, S., Shen, H., Zhu, X., Shi, Y., Shen, Q., Dong, W., Liu, R., Ling, Y., Zeng, Y., Wang, X., Zhang, Q., Wang, J., Wang, L., Wu, Y., Zeng, B., Wei, H., Zhang, M., Peng, Y., & Zhang, C. (2018). Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science, 359(6380), 1151-1156. [CrossRef]
- Chen, H., Cheng, J., Zhou, S., Chen, D., Qin, W., Li, C., Li, H., Lin, D., Zhang, Q., Liu, Y., Liu, A., & Luo, Y. (2021). Arabinoxylan combined with different glucans improve lipid metabolism disorder by regulating bile acid and gut microbiota in mice fed with high-fat diet International Journal of Biological Macromolecules, 168, 279-288. [CrossRef]
- Tong, L.-T., Zhong, K., Liu, L., Qiu, J., Guo, L., Zhou, X., Cao, L., & Zhou, S. (2014). Effects of dietary wheat bran arabinoxylans on cholesterol metabolism of hypercholesterolemic hamsters Carbohydrate Polymers, 112, 1-5. [CrossRef]
- Luo, S., He, L., Zhang, H., Li, Z., Liu, C., & Chen, T. (2022). Arabinoxylan from rice bran protects mice against high-fat diet-induced obesity and metabolic inflammation by modulating gut microbiota and short-chain fatty acids. . Food & function, 13(14), 7707-7719. [CrossRef]
- Li, S., Sun, Y., Hu, X., Qin, W., Li, C., Liu, Y., Liu, A., Zhao, Y., Wu, D., Lin, D., Zhang, Q., Chen, D., & Chen, H. (2019). Effect of arabinoxylan on colonic bacterial metabolites and mucosal barrier in high-fat diet-induced rats. Food science & nutrition, 7(9), 3052-3061. [CrossRef]
Figure 1.
Structure of arabinoxylan, showing the xylose backbone (black), arabinose substitutions (blue), ferulic acid (red). The main chain of xylose is connected by the β-1, 4-glucoside bond, arabinose can be linked to the main chain by the α-1, 2 or α-1, 3-glucoside bond, and ferulic acid is substituted at the O-5 position of arabinose by the ester bond.
Figure 1.
Structure of arabinoxylan, showing the xylose backbone (black), arabinose substitutions (blue), ferulic acid (red). The main chain of xylose is connected by the β-1, 4-glucoside bond, arabinose can be linked to the main chain by the α-1, 2 or α-1, 3-glucoside bond, and ferulic acid is substituted at the O-5 position of arabinose by the ester bond.
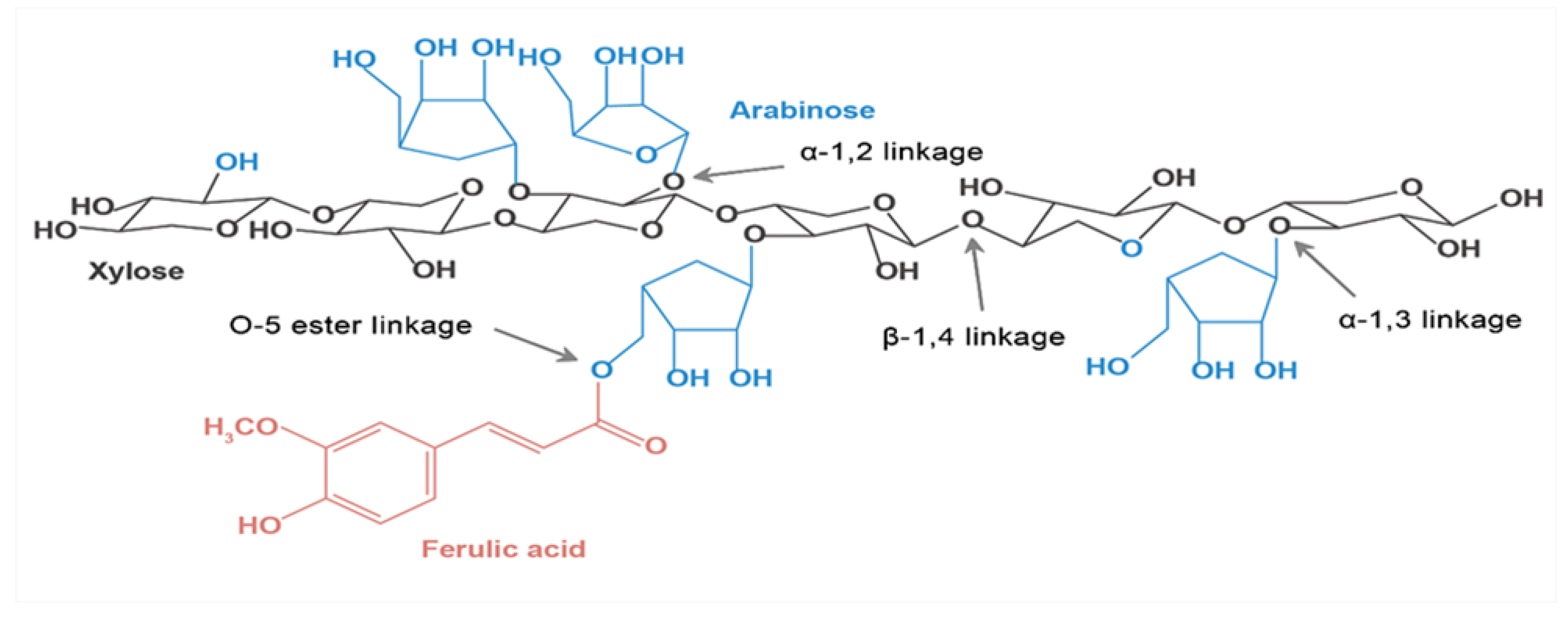
Figure 2.
Chemical treatments for WUAX extraction.
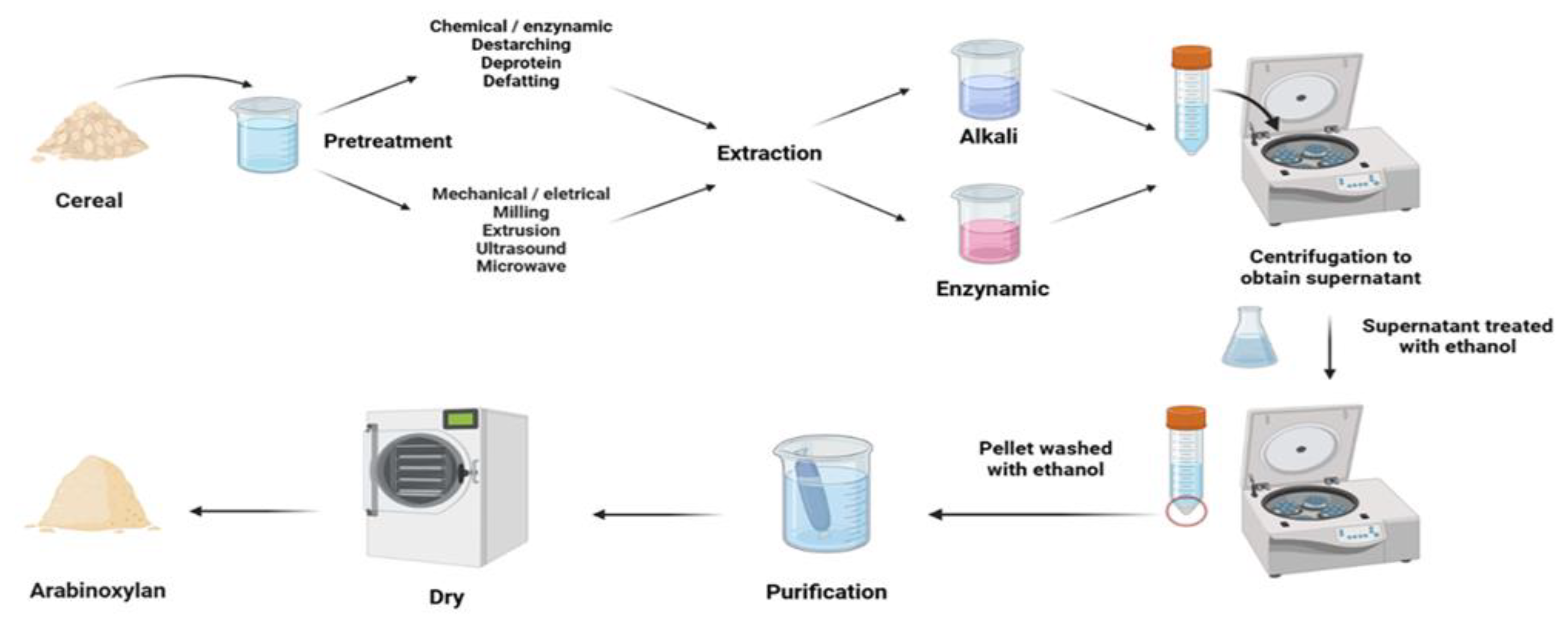
Figure 3.
Arabinoxylan fermented in the human gut.
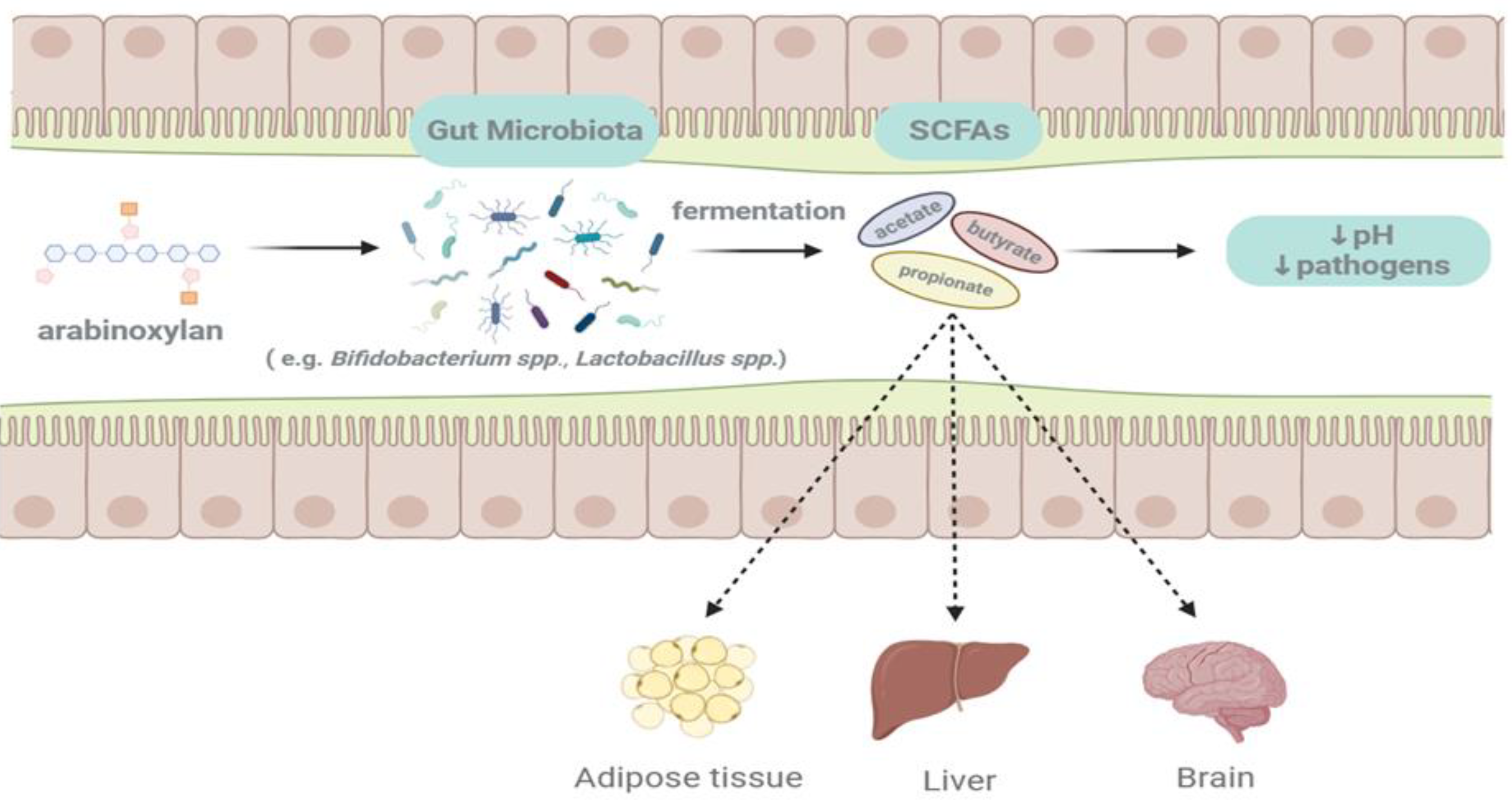
Figure 4.
Effect on the human gut microbiota and potential health benefits of arabinoxylan from different cereals.
Figure 4.
Effect on the human gut microbiota and potential health benefits of arabinoxylan from different cereals.
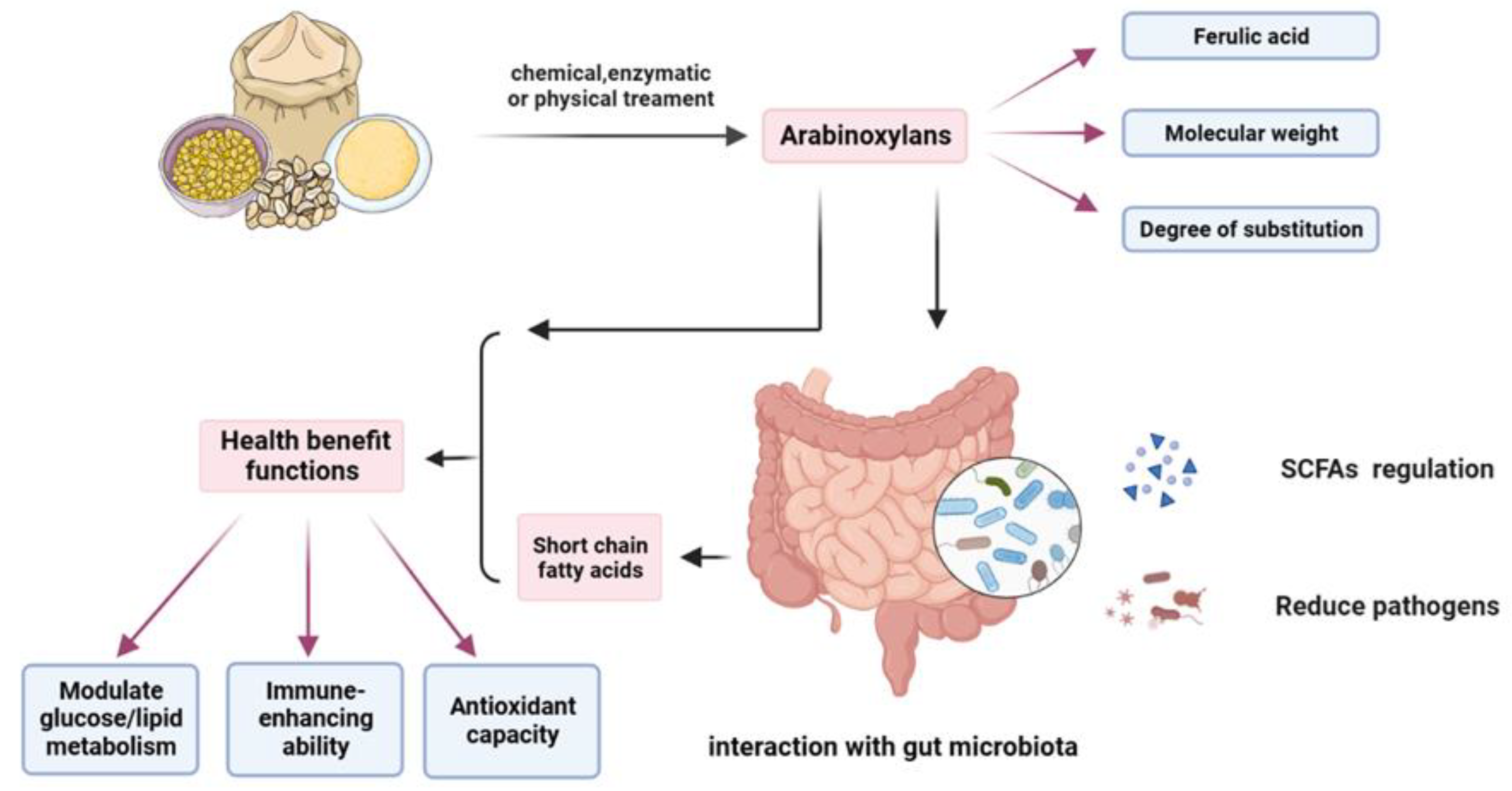

Table 1.
Effect of alkali extraction on the A/X ratio and MW of WUAX in different grains.
| Source | Extraction | MW/kDa | A/X | Reference |
| Wheat bran | NaOH | 351.7 | 0.83 | [67] |
| Wheat bran | NaOH and Ca (OH)2 | 437 | 1.14 | [68] |
| Wheat bran | NaOH | 694 | 0.95 | [49] |
| Wheat bran | NaOH | 770 | 0.51 | [61] |
| Wheat bran | NaOH | 163 | 0.77 | [69] |
| Barley bran | NaOH | 298.36 | 0.58 | [44] |
| Barley endosperm | NaOH | 877.1 | 0.76 | [70] |
| Peeled barley seeds | Ba (OH)2 | 1360 | 0.60 | [71] |
| Sorghum seeds | Ca (OH)2 | 223.9 | 1.09 | [72] |
| Red sorghum bran | NaOH | 356 | 1.1 | [73] |
| White sorghum bran | NaOH | 136.2 | 1.08 | [73] |
| Oat grain | NaOH | 100 | 0.11 | [40] |
| Corn bran | NaOH | 360 | 0.47 | [14] |
| Corn bran | NaOH | 590 | 0.51 | [61] |
| Corn bran | NaOH and Ca (OH)2 | 233.3 | 0.76 | [74] |
| Rye bran | Ba (OH)2 | 381 | 0.6 | [39] |
| Rye bran | NaOH and Ca (OH)2 | 234.9 | 0.72 | [74] |
Table 2.
Effect of enzyme extraction on the A/X ratio and MW of WUAX in different cereals.
| Source | Extraction | MW/kDa | A/X | Reference |
| Wheat bran | Xylanase | / | 0.44 | [80] |
| Wheat bran | Endo-xylanase | 1-25 (85.7%) 25-700(14.3%) |
0.81 | [91] |
| Wheat bran | Endo-xylanase | 12.5 | 0.73 | [78] |
| Wheat bran | Endo-xylanase | 420 | 1.14 | [14] |
| Corn bran | Xylanase | 51.6- 132 | / | [90] |
| Corn bran | Endo-xylanase | 43 | 0.54 | [14] |
| Rice bran | Endo-xylanase | 23 | 0.97 | [14] |
| Rice bran | Xylanase | 57.9 | 0.29 | [92] |
| Rice bran | Xylanase | 41.4 | 0.3 | [92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



