
Submitted:
06 May 2024
Posted:
08 May 2024
You are already at the latest version
Abstract
The broad global distribution of freshwater clams belonging to the genus Corbicula is driven by multiple hermaphroditic lineages. These lineages, characterized by shared morphological traits and phenotypic plasticity, pose challenges to morphological identification. Genetic markers, such as the mitochondrial COI gene, play a crucial role in delineating these lineages and their ranges. Morphotypes represent observed phenotypic variations, while lineages are defined based on genetic markers. Here, we comprehensively review Corbicula's distribution in Argentina, dis-criminate extant lineages based on both morphological and genetic (COI) data, and describe variation in internal and external morphology using 15 Argentine populations. Haplotype net-works identified two mitochondrial lineages: A/R morphotype (FW5 haplotype) and C/S mor-photype (FW17 haplotype). Strikingly, despite having similar vectors, origins, and invasive stages, Corbicula lineages exhibit virtually segregated distributions. However, intermediate morphotypes and both mitochondrial haplotypes are found in sympatry in northeastern Argen-tina, suggesting the presence of hybrids due to maternal genome retention. These findings con-tribute to the clarification of the identity and distribution of Corbicula lineages in Argentina, where the genus has been found for over half a century. Similar studies are needed in other areas to better understand the invasion patterns of a successful and plastic group.
Keywords:
Corbicula
; Morphology
; COI
; Argentina
; Distribution
1. Introduction
The freshwater bivalve genus Corbicula, first described by Megerle von Mühlfeld in 1811, is native to Africa, Asia, Australia, and the Middle East [1]. However, the fossil record indicates the presence of Corbicula in North America during the Eocene epoch, and multiple incursions into freshwater ecosystems in northwest Europe during Pleistocene interglacial periods, demonstrating the group’s significant dispersal capacity [2,3]. Consequently, it is not surprising that, given global interconnectedness and species movement across distant regions, Corbicula populations are now established in freshwater ecosystems throughout North and South America and Europe [4]. A further worldwide expansion of suitable habitats for invasive clams of the Corbicula genus, such as Corbicula fluminea (Müller 1774), has been forecasted in the context of climate change [5].
While in the native range sexual and asexual species coexist, recent proliferation of certain lineages has been facilitated by androgenesis, a particular reproductive strategy involving clonal asexual reproduction [6]. The widespread geographical and environmental distribution of these androgenetic lineages correlates with significant morphological diversity, particularly in taxonomic traits such as shell shape and color [7], leading to taxonomic complexity and confusion with numerous species described primarily based on shell characteristics [8]. As a result, the prevailing scientific consensus opts to use the term “forms” to denote the various morphological variations observed in Corbicula. In Europe, three forms are recognized: R, Rlc, and S [9,10], which correspond to morphotypes A, B, and C, respectively described in the Americas [11,12]. Recent molecular and genetic studies of the mitochondrial COI gene have shed light on the phylogeny of these forms in both their invasive and native ranges. These studies indicate that the A/R Form (commonly identified as C. fluminea) aligns with the native FW5 haplotype, prevalent in East Asia (Japan, Korea, Taiwan, China, Indonesia, and Myanmar; [13]. The C/S lineage is homologous to the FW17 haplotype [13], and its origin is cryptic but suggested in African populations [14]. There is a distinction within the Rlc/B form as well; while the North American form B corresponds to the native FW1, the European Rlc form is related to the FW4 haplotype, both of which are present in Korea [13]. Moreover, alterations in the external morphology in bivalves have been linked to internal variation in gill and palp areas [15,16]. Despite the critical roles these organs play in the reproduction and physiology of Corbicula species [17,18], their association with inter-specific differences or phenotypic plasticity remains largely unexplored.
Genetic characterization of these lineages does not fully resolve their taxonomy, partly because the same haplotype may occur in different morphotypes. This ambiguity could stem from phenotypic plasticity or androgenesis, a form of asexual reproduction prevalent among individuals in the invasive range [12]. These hermaphroditic individuals produce biflagellate diploid sperm that, during fertilization, expel the maternal nucleus, expressing solely the paternal genotype but retaining maternal mitochondrial DNA [19]. Furthermore, sperm from one lineage can fertilize oocytes from another, leading to “egg parasitism” and cytonuclear mismatches [12,20]. In some cases, this results in offspring with a hybrid genome due to incomplete extrusion of maternal nuclear DNA [21], displaying intermediate genotypes between the parents. Hence, reliance solely on mitochondrial DNA analysis may introduce bias, and an integrated taxonomic approach, incorporating examination of morphological and anatomical data, is recommended [22].
Recent research has comprehensively identified the invasive lineages of Corbicula spp. in South America using both morphological and genetic traits [23] although the geographical coverage remains uneven across the subcontinent. In Argentina, genetic studies have been limited to certain individuals from the Río de la Plata, upper Río Negro Basin, and Iguazú Falls, utilizing the mitochondrial gene COI [23] and the nuclear ribosomal gene 28S [12]. This constitutes a significant gap given the size of the country and widespread distribution of Corbicula spp. in it. The populations analyzed, morphologically assigned to C. largillierti (C/S lineage), C. fluminea (A/R lineage), and Corbicula sp., exhibited the mitochondrial COI haplotypes FW17, FW5, and FW1, respectively [12,23]. Notably, this latter mitochondrial haplotype (commonly found in individuals of form B) was detected in clams with nuclear haplotype C, in hybrid specimens between B and C, and was morphologically assigned to the C/S form. The extensive distribution observed in Argentina may be linked to a range of mitochondrial lineages that remain yet to be explored. Additionally, the presence of hybrids and morphogenetic inconsistencies underscores the likely complexity of combinations and the need for molecular tools to accurately identify populations.
However, despite various ecological, taxonomic, and genetic investigations, there remains a lack of comprehensive understanding of the current distribution, identity, and genetic relatedness of invasive Corbicula lineages in Argentina. In this study, we systematically analyzed the distribution of Corbicula spp. across a broad region of South America through a comprehensive examination of collections, literature, and our own field sampling. We used molecular markers to identify the populations and scrutinize their morphology. The principal objectives of this study were to: (1) update the distribution, (2) delineate the morphological and anatomical characteristics, (3) evaluate the genetic diversity and phylogeography of Corbicula spp. using mitochondrial DNA sequences, and (4) assess the correlation between morphological variation and molecular identity of the genus in Argentina.
2. Materials and Methods
Distribution of Corbicula in Argentina: Review and Sampling
The distribution of Corbicula in Argentina was compiled through an extensive review of malacological collections, literature sources, and expert consultations [24,25,26,27,28,29,30,31,32]. The primary collections consulted included the La Plata Museum (MLP), the Argentine Museum of Natural Sciences “Bernardino Rivadavia” (MACN), and the Miguel Lillo Foundation (FML). To gather current distribution data for Corbicula and collect specimens for subsequent analyses, a nationwide sampling initiative was conducted, covering the estuaries of the Negro and Río de La Plata rivers, the central and northwest basins, and the main plain rivers of the La Plata Basin from 2015 to 2017 (Table 1). The collected specimens were deposited in the MACN Invertebrate Collection for further analysis.
Morphological Assignment of Lineages
Despite the difficulties previously mentioned, the assignment of Corbicula morphotypes can be carried out when certain characteristics remain relatively stable and, especially when extreme states are observed. The distinction between the two previously reported forms in Argentina, C/S and A/R, relies on factors such as internal and external coloration, depth and number of ribs, the shape of the posterior margin and umbo, and thickness of the valves [29,33,34] (Figure 1). A conservative approach was adopted, whereby if any character exhibited ambiguities or contradicted others, it was not definitively assigned to either of the two forms and was designated as an “indeterminate” form (form I).
Shell Morphology
Shell length (L), width (W), and height (H) were measured for each individual using a caliper (precision: 0.01 mm) to estimate the maximum anteroposterior, lateral, and dorsoventral dimensions, respectively. Shell weight (SW) was measured after drying them at room temperature in a desiccator following [35]. The valves were scanned to capture their internal and external surfaces at the highest possible resolution (2400 dpi). Subsequently, the number of external concentric ribs per centimeter (Ribs, cm⁻¹) was quantified from the obtained images. Additionally, to incorporate the internal and external color of the valves as quantitative variables, RGB values were extracted through color queries conducted using Adobe® Photoshop® (2017). This process involved sampling three random sectors of the internal and external faces of the valves, each with a sample area of 101 pixels.
Internal Morphology
Gills and palps of each clam were carefully dissected and placed in a Petri dish with ethanol, before being photographed using a dissecting microscope (Leica S8APO; 10X magnification; Figure 2). The gill and palps areas were quantified by digitally outlining their contours using Digimizer version 5.3.5 (Ostend: MedCalc Software 2016; precision: 0.1 mm2) [36]. For the gills, both the internal and external demibranchs of one randomly chosen gill (left or right) were measured, and their combined areas were doubled to determine the total gill area (GA). Regarding the palps, all four were individually measured, including external and internal structures on both sides. Due to their curved shape, the palps were oriented with the concave side facing up, and their edges were extended to ensure they lay flat. Filament density (FD), defined as the average number of filaments per millimeter, was assessed in each clam (30 per population). Three randomly selected sections from a 50X magnification image of the gills (Figure 3D) were used to calculate FD, following the methodology outlined by [16].
Population Selection for Molecular Analysis: Geographic Diversity across Argentina
Genetic analyses were conducted utilizing samples obtained from the previously described campaigns. We selected individuals from 15 populations to ensure broad geographic representation, covering diverse forms of Corbicula in Argentina. These populations are drawn from five regions (refer to Table 1 for site codes and locations): the Río de la Plata estuary (PLA, BUE); Litoral (CON, APO, ASJ, SRO, ESP, RML); Central (COR); Northwest (DCC, MOL, RJU, SDE, CAT); and Patagonia (RNE).
DNA Extraction, PCR Amplification, and Sequencing
For a total of 225 individuals (15 specimens per sampling location), genomic DNA was obtained following glass fiber-based extraction protocol plates [37,38,39]. A 658 bp fragment near the 5’ end of the cytochrome c oxidase subunit I (COI) mitochondrial gene was amplified using BivF4_t1 (5’-TGTAAAACGACGGCCAGTGKTCWACWAATCATAARGATATTGG-3’) and BivR4_t1 (5’-CAGGAAACAGCTATGACTAMACCTCWGGRTGVCCRAARAACCA-3’) pair of primers. Thermocycling conditions consisted of 2 min at 94°C for 2 min;5 cycles of 30 s at 94°C, 40 s at 45°C and 1 min at 72°C 35 cycles of 30 s at 94 °C,40 s at 51°C and 1 min at 72°C for; final extension of 10 min at 72°C. Sequencing was performed bidirectionally at the Centre for Biodiversity Genomics (Guelph, Canda) for 36 individuals, and unidirectionally for 190 individuals at Macrogen (Seoul, Korea), with the same primers used for amplification. Sequences were edited using CODONCODE ALIGNER (CodonCode Corporation). We obtained good-quality sequences for 210 individuals, which can be found in GenBank under the accession numbers (XXX-XXX; Table S1).
Data Analyses
Phylogenetic and Phylogeographic Analyses
To increase our taxonomic and geographic coverage for the phylogenetic analyses, we mined from GenBank 58 COI public sequences representative of other native and invasive Corbicula species and mitochondrial lineages or haplogroups (see Table S2 - Supplementary Material). A gene tree based on 269 sequences was inferred through Bayesian methodology using MrBayes 3.2.2 [40]. The best-fit model nucleotide evolution for our COI data set was TPM3uf+G based on the Bayesian information criterion (BIC) as implemented in jModelTest 2.1.1 [41]. We specified in MrBayes the closest model available (HKY+G) and conducted two independent runs of 10 million generations under default priors and sampling trees every 100 generations. The average standard deviation of split frequencies between runs was < 0.01, indicating convergence. We verified that both runs reached stationarity and that we had a good sample of the posterior probability distribution with Tracer 1.7.2 [42]. We discarded the first 25% of sampled trees as burn-in and combined the remaining 75,000 topologies of each run to generate a 50% majority rule consensus tree. The tree was rooted with Neocorbicula limosa.
To further investigate the two major freshwater (FW) haplogroups found in Argentina, we generate an unrooted statistical parsimony network with TCS 1.21 [43] and a cut-off value of 95% [44]. We also include some specimens of C. sandai and other FW lineages for context. The obtained network was edited with tcsBU [45]. We used MEGA [46] to estimate mean uncorrected genetic distance (p-distance) between haplogroups, and the per-site nucleotide diversity (π) within haplogroups in DNASP 5.10 [47]. In both cases, we used the pairwise deletion option for missing data. Finally, we explored the correspondence between mitochondrial and morphological identities, and the geographic distribution of the different morphotypes and haplogroups found in Argentina. To do so, we first assessed how representatives of each of the three phenotypic forms (A/R, C/S, and I) are clustered within the two mitochondrial lineages and, then, how both lineages and morphotypes are distributed across sampling locations.
Morphological Clustering Analysis and Morphotype Reassignment
We assessed morphological variations among Corbicula populations through multivariate clustering analysis, standardizing morphological variables by size. Specifically, linear measurements of the valves were expressed as ratios between Height and Length (H:L), as well as Width and Length (W:L). Furthermore, we adjusted shell weight by dividing it by the general size, calculated as the product of the length of the three major body axes (L × H × W), a method known to reliably estimate shell thickness (ST, [17]). Gill area and palp area were also normalized relative to the general shell size of the valves (RGA and RPA, respectively). Additionally, the palp-to-gill ratio (PA:GA) was included in the analyses. Before incorporating color information, Principal Component Analysis (PCA) was performed on the six color variables obtained previously (Red, Green, and Blue composition for both internal and external shell faces), with retention of the first two principal axes (PC1_color and PC2_color). The scores of each clam in these axes were then incorporated as variables. Finally, we included the number of concentric ribs and filament density to create a matrix of ten morphometric variables.
We utilized fuzzy clustering to partition the data into clusters. Fuzzy clustering is a method employed to group data points into clusters, where membership in each cluster is not strictly binary, allowing for partial membership of data points in multiple clusters [48]. Unlike traditional “hard” clustering algorithms such as K-means, which assign each data point exclusively to a single cluster, fuzzy clustering enables more flexible assignments. We opted for this approach due to the potential presence of intermediate morphotypes within the Corbicula genus, attributed to cytonuclear inconsistencies and hybridization between lineages resulting from the androgenetic reproduction method. In fuzzy clustering, each data point is assigned a membership value for each cluster, indicating the degree of association with that cluster. These membership values typically range between 0 and 1, with higher values indicating stronger membership. The analysis was made by the fanny function from the ‘cluster’ package [49]. Before this, we determined the optimal number of clusters by the within-cluster sums of squares method, employing the fviz_nbclust() function from the ‘factoextra’ package [50]. The analyses consistently indicated two as the most probable number of clusters, coinciding with the recognized forms observed in the study area, therefore the number of clusters for fuzzy clustering was set to two. We obtained the membership probabilities for each cluster and established a cut-off threshold of 0.65 to determine membership in either cluster. Individuals failing to exceed this limit were classified as not belonging to any specific cluster, and categorized as “indeterminate”. These results were compared with the previously established assignments to assess the classification’s effectiveness and correspondence between both methods.
Lastly, we explored the underlying structure of the dataset and identified patterns among the variables that best captured the variability between forms and populations conducting a Principal Component Analysis (PCA). We identified the most relevant morphometric variables for distinguishing Corbicula morphotypes and examined the positioning of clusters derived from the preceding analysis, along with that of indeterminate individuals.
3. Results
Distribution of Corbicula in Argentina: Review and Sampling
From collections and bibliography, 157 Corbicula records in Argentina were compiled, predominantly belonging to the A/R form (C. fluminea, 93), with the remainder comprising C/S form (C. largillierti, 49), a hybrid form (4), and unidentified records (11) (Appendix, Table S3). Based on the data in hand, the temporal progression seems to indicate that between the 1980s and 1995, both Corbicula lineages rapidly spread northward along the Paraná River and lower Uruguay, primarily recorded in Santa Fe, Entre Ríos, Chaco, and Corrientes provinces. Subsequently, they underwent sudden geographical expansion into diverse regions, occupying basins unconnected to their original habitats (Appendix, Table S3). Finally, lineages exhibited distinct geographical distributions, with the A/R form concentrated in the east and southern main plain rivers, while the C/S form was found mainly in reservoirs and low-order rivers in the Central and Northwestern regions. Except for a few coexisting populations, lineages primarily inhabited allopatric areas. Notably, the Northeast featured populations with intermediate morphotypes (Appendix, Table S3). Our own surveys encompassed 26 populations, including previously unreported sites, with 11 corresponding to the A/R form, 8 to C/S, and 7 to the indeterminate form (Appendix, Table S4).
Phylogenetic and Phylogeographic Analyses
The Bayesian gene tree recovered the estuarine and freshwater (FW) Corbicula clades as reciprocally monophyletic with maximum node support (Figure 3; Supplementary Figure S1). All individuals sampled in Argentina for this study clustered within two of the four major freshwater (FW) mitochondrial lineages of the androgenetic, invasive Corbicula clams: FW5 and FW17 (Figure 3). Specimens were almost equally distributed between the two haplogroups, being FW5 slightly more frequent than FW17 in Argentina (Table 2). Both lineages were recovered as monophyletic with high (0.99) to maximum (1.0) support. The new, recently reported FWBRA lineage from Brazil [23], was recovered as an intermediate haplotype between FW5 and FW17, being slightly closer to the former in terms of genetic divergence (Figure 3). Mean, uncorrected genetic distance between FWBRA and FW5 was 1.25%, and 1.57% between the former and while between the former and FW17.
Figure 3.
Bayesian majority rule consensus tree obtained from the analysis of 268 sequences of the mitochondrial gene COI (683 bp), showing the relationships among estuarine and freshwater species and major haplogroups of Corbicula clams. Numbers above or below the branches indicate Bayesian posterior probability node support. Some clades were collapsed and a few support values were omitted for clarity and simplicity (see Figure S1 for the full topology with all tips shown). The tree was rooted with Neocorbicula limosa (not shown).
Figure 3.
Bayesian majority rule consensus tree obtained from the analysis of 268 sequences of the mitochondrial gene COI (683 bp), showing the relationships among estuarine and freshwater species and major haplogroups of Corbicula clams. Numbers above or below the branches indicate Bayesian posterior probability node support. Some clades were collapsed and a few support values were omitted for clarity and simplicity (see Figure S1 for the full topology with all tips shown). The tree was rooted with Neocorbicula limosa (not shown).
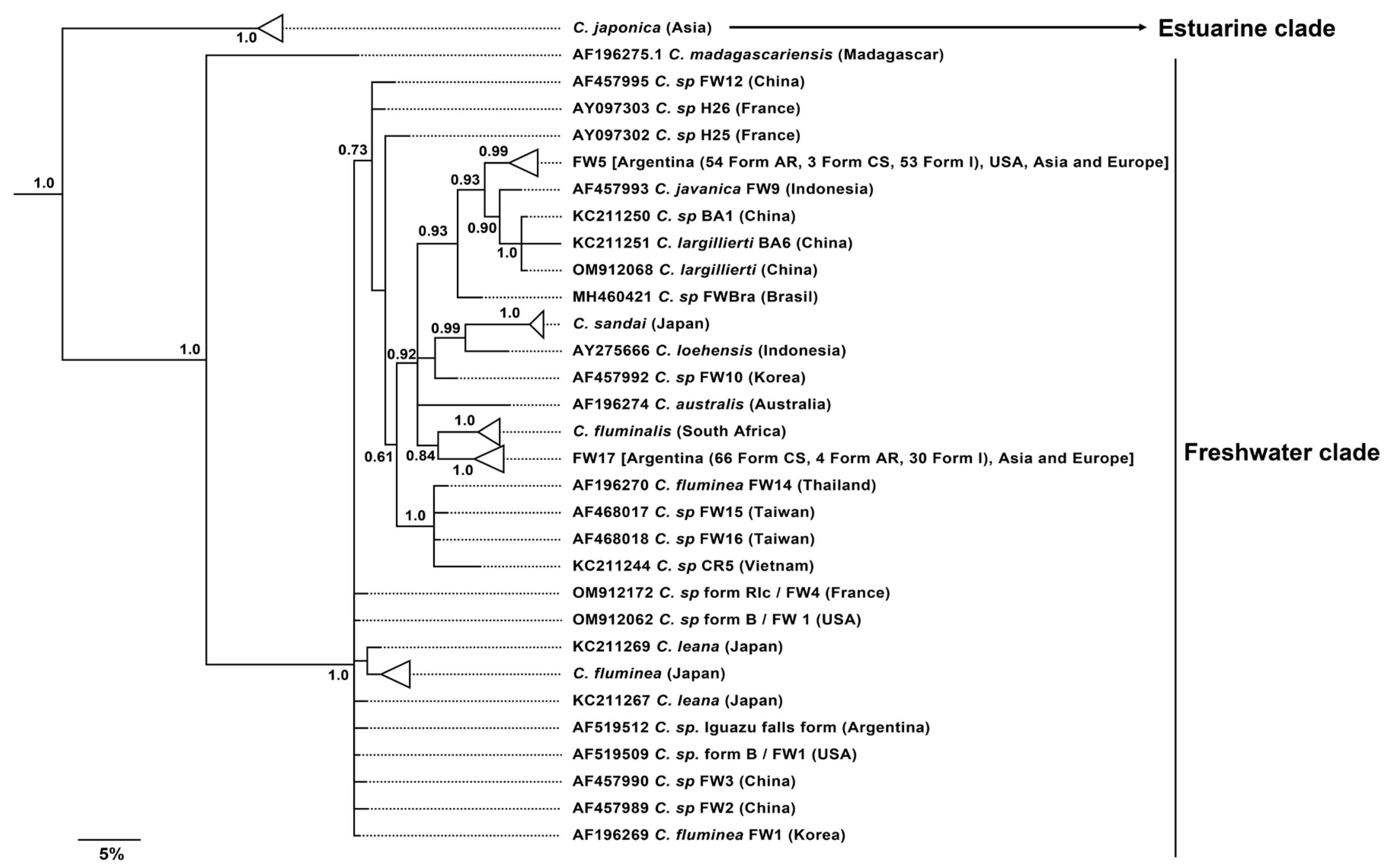
Individuals within the FW5 haplogroup corresponded to the phenotypic form A/R, while individuals categorized as C/S clustered within the FW17 lineage (Figure 3, Figure 4 and Figure S1; Table 2). The only exceptions were three specimens with a phenotype A/R from Buenos Aires province were grouped within the FW17, while three representatives of the C/S from Cordoba Province clustered within the FW5 haplogroup (Figure 4; Supplementary Figure S1). As for the specimens with intermediate forms, 53 individuals clustered inside the FW5 lineage and 30 within the FW17 haplogroup (Figure 4; Supplementary Figure S1, Table 2). Representatives of the form C/S were concentrated in northwestern Argentina, while specimens with an A/R phenotype were distributed to the east and south, and individuals with intermediate phenotype were found almost exclusively in northeastern Argentina. No sampling location held more than one phenotypic form (Figure 4A).
The haplogroup FW5 was found almost exclusively in eastern Argentina, from Misiones province in the north to northern Patagonia (Río Negro province) in the south (Figure 4A). In contrast, the FW17 lineage was chiefly distributed in central and northwestern Argentina. Haplogroups were found in sympatry in five sampling locations (Figure 4A), one in central Argentina (COR) and the remaining four to the east of the country (ASJ, ESP, PLA, RML). Finally, and as evidenced by the haplotype network, genetic diversity within both lineages was remarkably low, with a common central haplotype and a few, unique haplotypes closely related (one or two mutational steps) to it (Figure 4B; Table 2).
Morphological Clustering Analysis and Morphotype Reassignment
The cluster analysis exhibited strong agreement with our initial assignments, with 82.5% (245 out of 297) of individuals classified consistently into either the A/R, C/S forms, or indeterminate categories. Notably, discrepancies primarily emerged in Northeastern populations (Misiones, Formosa, and Chaco), where 32 individuals initially classified as indeterminate were identified with a greater than 65% probability of belonging to the A/R cluster. Similarly, at the DCC site, 7 out of 30 initially designated as C/S forms are now categorized as indeterminate.
The first three principal components (PCs) of the PCA collectively explained 71.19% (PC1 = 34.18%, PC2 = 19.69%, and PC3 = 17.32%) of the variance in the dataset, effectively summarizing its variability (Table 3). The PCA revealed distinct patterns in the morphometric variables of Corbicula populations: PC1 exhibited strong positive loadings for internal color and number of ribs, and strong negative loadings for ST and W:L ratio, distinguishing clusters associated with A/R and C/S forms (Figure 5). PC2 demonstrated significant positive loadings for RGA and negative loadings for H:L ratios, highlighting differences between populations. PC3 showed a notable correlation with the external color of valves, with indeterminate individuals characterized by positive values (Figure 5).
4. Discussion
This study presents the first review of Corbicula distribution in Argentina, encompassing a significant subcontinental region. Previous research on Corbicula distribution in Argentina, using morphological and genetic data, have been limited to a single basin or few specific environments [12,23,29,51]. Our review coupled an unprecedented reevaluation of taxonomic identification within malacological collections with a countrywide sampling effort for morphological and molecular analyses. Our findings reveal a prevalence of the A/R form, particularly in the east and southern main plain rivers, contrasting with the dominance of the C/S form mainly in reservoirs and low-order rivers in the Central and Northwestern regions. Surprisingly, despite similarities in vectors, geographical origins, and the temporal stages of invasion, distinct Corbicula lineages exhibit segregated distributions in Argentina. This divergence likely indicates other underlying factors such as morphological, physiological, or ecological disparities. Notably, the Northeast region displayed populations with intermediate morphotypes, suggesting potential hybridization or transitional characteristics.
Prior to our current study, mitochondrial identities had been established for three Argentinean populations: the FW5 and FW17 lineages coexisting in the Río de la Plata estuary, the FW17 lineage in the upper Río Negro basin in Northern Patagonia, and the FW1 lineage at the Iguazú Falls on the Argentine-Brazilian border [12,23]. Our phylogenetic analyses, covering 15 populations across the entire distribution range of Corbicula in Argentina, highlight the widespread prevalence of two primary freshwater mitochondrial lineages, FW5 and FW17. Most populations (10 out of 15) consist of a single mitochondrial lineage, revealing a clear pattern of geographic segregation among them. This discovery is particularly striking given the typical sympatric distribution of both lineages across Europe, North America [6,52], and Russia [53]. Furthermore, it’s noteworthy that in Argentina, the FW17 lineage exhibits a remarkable widespread distribution, establishing abundant populations that have persisted for over two decades in regions not colonized by the A/R form. This phenomenon is unique and contrasts with the lineage’s limited geographic range in Europe and North America [9,52,54,55]. Even in neighbouring Brazil, the predominant lineage is FW5, with FW17 found in sympatry only in a few locations [23].
In general, Argentine lineages FW5 and FW17 exhibited a high correspondence with forms A/R and C/S, respectively, mirroring observations in other locations worldwide [6,12]. However, there are exceptions to this pattern. Instances of discordance between mitochondrial lineages and morphotypes have been documented in several Corbicula populations belonging to different forms [6,9,12,21]. These discrepancies suggest processes of androgenetic egg parasitism [6,12,21]. In the present study, cytonuclear mismatches were identified in Buenos Aires, where three individuals displaying an A/R phenotype were classified under FW17, and in Cordoba, where three individuals with a C/S phenotype clustered within the FW5 haplogroup. Although currently only one form is prevalent in each of these locations [51,56], historical records indicate the past occurrence of both forms [29,57]. Thus, the current presence of both mitochondrial haplotypes likely reflects a previous coexistence. Another instance is illustrated by the intermediate morphotypes observed in Northeastern Argentina. These morphotypes may signify hybrid individuals resulting from sympatric invasive lineages, possibly due to androgenetic parasitism with incomplete expulsion of the maternal nuclear material [21].
The morphological variations observed in this study are consistent with previous research, which primarily relied on the ratios of linear shell measurements, a characteristic and conserved trait relevant to morph differentiation [9,23,54,58]. In contrast, relative gill area did not emerge as a significant variable in differentiating between morphs. This finding contrasts somewhat with previous research conducted in Argentina [17]. One reason for this disparity may be the different variables used to relativize gill area. While [17] used dry weight, a physiologically meaningful variable, in our study, shell volume was utilized solely for the purpose of morphometric descriptors. Another contributing factor may lie in the scope of the studies: [17] research was based on only a few populations from two sites, potentially failing to capture some of the intra-morph variation. In the present study, however, this index explained substantial intracluster interpopulation variability along PC2 of the PCA. Notably, individuals unable to be assigned to either form spanned a broad range of morphometric space. This significant phenotypic variability could be attributed to the presence of various combinations of nuclear genes among lineages, resulting in diverse phenotypes. Nevertheless, alternative hypotheses such as phenotypic plasticity cannot be dismissed without further studies involving nuclear genes.
Phylogenetic and cytological studies have shown the absence of reproductive barriers between lineages, challenging the utility of the biological species concept in this group [19,22]. Consequently, the scientific consensus has leaned towards classifying Corbicula as a “complex of polymorphic species” [6]. However, this classification does not imply ecological equivalence among different forms; in fact, there are lineage disparities not only in morphology but also in microhabitat preferences [59], individual growth rates [60,61], reproductive event frequency, offspring release numbers [62], and physiological traits such as filtration [63] and metabolic rates [17]. The pronounced segregation of forms and haplotypes discovered across Argentina bolsters this notion. It has been suggested that some of the variability in life-history traits may have a genetic basis [11], although phenotypic plasticity could also be a contributing factor, as observed in other freshwater invasive bivalves [16]. Further research is necessary to determine their relative significance. Future studies should delve into the potential adaptive advantages conferred by these differences in various environmental conditions, as well as their implications for distribution patterns.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Bayesian majority rule consensus tree obtained from the analysis of 268 sequences of the mitochondrial gene COI of Corbicula clams; Table S1: Genbank accession numbers of the sequences of the cytochrome oxidase I (COI) genes of 211 individuals of Corbicula collected in Argentina between 2015 and 2017;.Table S2: GenBank accession numbers of the mitochondrial sequences downloaded and used for the phylogenetic analysis of individuals from Corbicula populations in Argentina; Table S3: Records of the presence of Corbicula obtained from databases of the main malacological collections in Argentina; Table S4: Survey date location details for sites surveyed for Corbicula spp. clams during the sampling campaigns carried out in Argentina between 2015 and 2017
Author Contributions
Conceptualization, methodology, and investigation, F.S, E.M.P. and L.A.H.; formal analysis and validation, L.A.H and P.D.L; resources, F.S. and E.M.P.; data curation, L.A.H and P.D.L.; writing—original draft preparation, L.A.H. and P.D.L.; writing—review and editing, L.A.H, P.D.L, E.M.P, and F.S.; supervision, P.D.L. and F.S.; funding acquisition, F.S and E.M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by research grants from the Argentine Agencia Nacional de Promoción Científica y Tecnológica (PICT 2016–0631 to F.S. and PICT 2020-0629 to E.M.P.), the National Scientific and Technical Research Council (CONICET- PIP 2021-2023 to E.M.P) and the Consejo de Investigación, Universidad Nacional de Salta (CIUNSa 2621/0 to FS). We also acknowledge to the International Development Research Centre of Canada, Natural Sciences and Engineering Research Council of Canada y Fondo iBOL Argentina (CONICET).
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
Original data will be published in a open repository (GenBank and Zenodo)
Acknowledgments
We acknowledge Dora Davies, Lucía Nieva, Paola Reyna, Eugenia Salas, and Pablo Martin for the information provided to access sampling sites. To Gustavo Darrigran, Alejandro Tablado and Eugenia Salas for access to the collections of the MACN, MLP and FML museums, respectively. To Gustavo Darrigran, Pablo Martin and Liliana Salas for sending samples. To Eimi Font, Marta Leiva and Ezequiel Salomon, for information about the presence of Corbicula in new sites. To Elizabeth Vilacoba, Laura Barone and Darío Lijmaer of the MACN for their help in processing the genetic material.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- McMahon, R.F. The Occurence and Spread of the Introduced Asiatic Freshwater Clam, Corbicula fluminea (Muller) in North America: 1924-1982. Nautilus 1982, 96, 134–141. [Google Scholar]
- Araujo, R.; Moreno, D.; Ramos, M.A. The Asiatic Clam Corbicula fluminea (Müller, 1774) (Bivalvia: Corbiculidae) in Europe. Am. Malacol. Bull. 1993, 10, 39–49. [Google Scholar]
- Meijer, T.; Preece, R. A review of the occurrence of Corbicula in the Pleistocene of North-West Europe. Neth. J. Geosci. 2000, 79, 241–333. [Google Scholar] [CrossRef]
- Karatayev, A.Y.; Burlakova, L.E.; Padilla, D.K. Contrasting Distribution and Impacts of Two Freshwater Exotic Suspension Feeders, Dreissena polymorpha and Corbicula fluminea. In The Comparative Roles of Suspension-Feeders in Ecosystems. NATO Science Series IV: Earth and Environmental Series, vol 47; Dame, R.F., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 239–262. [Google Scholar]
- Gama, M.; Crespo, D.; Dolbeth, M.; Anastácio, P.M. Ensemble forecasting of Corbicula fluminea worldwide distribution: Projections of the impact of climate change. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 675–684. [Google Scholar] [CrossRef]
- Pigneur, L.-M.; Marescaux, J.; Roland, K.; Etoundi, E.; Descy, J.-P.; Van Doninck, K. Phylogeny and androgenesis in the invasive Corbicula clams (Bivalvia, Corbiculidae) in Western Europe. BMC Evol. Biol. 2011, 11, 147. [Google Scholar] [CrossRef]
- Bespalaya, Y.V.; Kropotin, A.V.; Kondakov, A.V.; Aksenova, O.V.; Gofarov, M.Y.; Kim, S.K.; Lee, J.H.; Travina, O.V.; Vikhrev, I.V.; Vinarski, M.V.; et al. A taxonomic reassessment of native and invasive species of Corbicula clams (Bivalvia: Cyrenidae) from the Russian Far East and Korea. Zoöl. J. Linn. Soc. 2022, 197, 104–126. [Google Scholar] [CrossRef]
- Morton, B. Corbicula in Asia - an Updated Synthesis. Am. Malacol. Bull. 1986, Spetial Ed, 113–124. [Google Scholar]
- Pfenninger, M.; Reinhardt, F.; Streit, B. Evidence for cryptic hybridization between different evolutionary lineages of the invasive clam genus Corbicula (Veneroida, Bivalvia). J. Evol. Biol. 2002, 15, 818–829. [Google Scholar] [CrossRef]
- Marescaux, J.; Pigneur, L.-M.; Van Doninck, K. New records of Corbicula clams in French rivers. Aquat. Invasions 2010, 5, S35–S39. [Google Scholar] [CrossRef]
- Siripattrawan, S.; Park, J.-K.; Foighil, D. . Two lineages of the introduced Asian freshwater clam Corbicula occur in North America. J. Molluscan Stud. 2000, 66, 423–429. [Google Scholar] [CrossRef]
- Lee, T.; Siripattrawan, S.; Ituarte, C.F.; Foighil, D.Ó. Invasion of the Clonal Clams: Corbicula Lineages in the New World. Am. Malacol. Bull. 2005, 20, 113–122. [Google Scholar]
- Park, J.; Kim, W. Two Corbicula (Corbiculidae: Bivalvia) mitochondrial lineages are widely distributed in Asian freshwater environment. Mol. Phylogenetics Evol. 2003, 29, 529–539. [Google Scholar] [CrossRef]
- Pigneur, L.; Etoundi, E.; Aldridge, D.C.; Marescaux, J.; Yasuda, N.; Van Doninck, K. Genetic uniformity and long-distance clonal dispersal in the invasive androgenetic Corbicula clams. Mol. Ecol. 2014, 23, 5102–5116. [Google Scholar] [CrossRef]
- Capelle, J.J.; Hartog, E.; Bogaart, L.v.D.; Jansen, H.M.; Wijsman, J.W. Adaptation of gill-palp ratio by mussels after transplantation to culture plots with different seston conditions. Aquaculture 2021, 541, 736794. [Google Scholar] [CrossRef]
- Paolucci, E.M.; Sardiña, P.; Sylvester, F.; Perepelizin, P.V.; Zhan, A.; Ghabooli, S.; Cristescu, M.E.; Oliveira, M.D.; MacIsaac, H.J. Morphological and genetic variability in an alien invasive mussel across an environmental gradient in South America. Limnol. Oceanogr. 2014, 59, 400–412. [Google Scholar] [CrossRef]
- Hünicken, L.A.; Sylvester, F.; Paolucci, E.M. Physiological and morphological assessments suggest opposite structural allocation strategies between closely related invasive clams. Hydrobiologia 2022, 849, 2859–2875. [Google Scholar] [CrossRef]
- Modesto, V.; Ilarri, M.; Labecka, A.M.; Ferreira-Rodríguez, N.; Coughlan, N.E.; Liu, X.; Sousa, R. What we know and do not know about the invasive Asian clam Corbicula fluminea. Hydrobiologia 2023, 1–32. [Google Scholar] [CrossRef]
- Komaru, A.; Kawagishi, T.; Konishi, K. Cytological evidence of spontaneous androgenesis in the freshwater clam Corbicula leana Prime. Dev. Genes Evol. 1998, 208, 46–50. [Google Scholar] [CrossRef]
- Pigneur, L.-M.; Hedtke, S.M.; Etoundi, E.; Van Doninck, K. Androgenesis: a review through the study of the selfish shellfish Corbicula spp. Heredity 2012, 108, 581–591. [Google Scholar] [CrossRef]
- Hedtke, S.M.; Stanger-Hall, K.; Baker, R.J.; Hillis, D.M. All-Male asexuality: origin and maintenance of androgenesis in the asian clam corbicula. Evolution 2008, 62, 1119–1136. [Google Scholar] [CrossRef]
- Kropotin, A.V.; Bespalaya, Y.V.; Aksenova, O.V.; Kondakov, A.V.; Aksenov, A.S.; Khrebtova, I.S.; Palatov, D.M.; Travina, O.V.; Bolotov, I.N. Genetic and Morphological Characterization of the Invasive Corbicula Lineages in European Russia. Water 2023, 15, 3226. [Google Scholar] [CrossRef]
- Ludwig, S.; Darrigran, G.; Boeger, W.A. Opening the black box of the invasion of Corbicula clams (Bivalvia, Corbiculidae) in South America: a genetic and morphological evaluation. Hydrobiologia 2023, 851, 1203–1217. [Google Scholar] [CrossRef]
- Cazzaniga, N.J. Asiatic Clam,Corbicula fluminea, Reaching Patagonia (Argentina). J. Freshw. Ecol. 1997, 12, 629–630. [Google Scholar] [CrossRef]
- Cazzaniga, N.J.; Perez, C. Asiatic Clam, Corbicula fluminea, in Northwestern Patagonia (Argentina). J. Freshw. Ecol. 1999, 14, 551–552. [Google Scholar] [CrossRef]
- Martín, P.R.; Estebenet, A.L. Spread of the Asiatic Clam Corbicula fluminea in Southern Pampas and Northern Patagonia, Argentina. J. Freshw. Ecol. 2002, 17, 331–333. [Google Scholar] [CrossRef]
- Semenas, L.; Flores, V. Presence ofCorbicula flumineain the Upper Negro River Basin (Patagonia, Argentina). J. Freshw. Ecol. 2005, 20, 615–616. [Google Scholar] [CrossRef]
- Martin, P.R.; Tiecher, M.J. Hallazgo de La Almeja Invasora Corbicula fluminea En El Río Sauce Grande (Provincia de Buenos Aires, Argentina). BioScriba 2009, 2, 115–120. [Google Scholar]
- Reyna, P.B.; Morán, A.G.; Tatián, M. Taxonomy, distribution and population structure of invasive Corbiculidae (Mollusca, Bivalvia) in the Suquía River basin, Córdoba, Argentina. Iheringia, Série Zool. 2013, 103, 77–84. [Google Scholar] [CrossRef]
- Torre, L.; Reyna, P. Bivalvia, Veneroidea, Corbiculidae, Corbicula largillierti (Philippi, 1844): New distribution record in the Del Valle Central basin, Catamarca Province, Argentina. Check List. 2013, 9, 165. [Google Scholar] [CrossRef]
- Molina, L.M.; Pereyra, P.J.; Carrizo, N.G.M.; Abrameto, M.A. Here come the clam: Southernmost record worldwide of the Asian clam Corbicula fluminea (Patagonia, Argentina). Russ. J. Biol. Invasions 2015, 6, 129–134. [Google Scholar] [CrossRef]
- Trovant, B.; Signorelli, J.H.; Battini, N. Invasive pest spreads beyond the last frontier: Corbicula clam in the Chubut River, Patagonia. Limnology 2023, 24, 1–8. [Google Scholar] [CrossRef]
- Ituarte, C.F. Corbicula and Neocorbicula (Bivalvia: Corbiculidae) in the Paraná, Uruguay, and Río de La Plata Basins. Nautilus 1994, 107, 129–135. [Google Scholar]
- Mansur, M.C.D.; Pereira, D. Bivalves límnicos da bacia do rio dos Sinos, Rio Grande do Sul, Brasil (Bivalvia, Unionoida, Veneroida e Mytiloida). Rev. Bras. de Zoöl. 2006, 23, 1123–1147. [Google Scholar] [CrossRef]
- Coughlan, N.E.; Cunningham, E.M.; Cuthbert, R.N.; Joyce, P.W.S.; Anastácio, P.; Banha, F.; Bonel, N.; Bradbeer, S.J.; Briski, E.; Butitta, V.L.; et al. Biometric conversion factors as a unifying platform for comparative assessment of invasive freshwater bivalves. J. Appl. Ecol. 2021, 58, 1945–1956. [Google Scholar] [CrossRef]
- Dutertre, M.; Barillé, L.; Beninger, P.G.; Rosa, P.; Gruet, Y. Variations in the pallial organ sizes of the invasive oyster, Crassostrea gigas, along an extreme turbidity gradient. Estuarine, Coast. Shelf Sci. 2009, 85, 431–436. [Google Scholar] [CrossRef]
- Ivanova, N.V; DeWard, J.; Hebert, P.D.N. An Inexpensive, Automation-Friendly Protocol for Recovering High-Quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Ivanova, N.V.; Fazekas, A.J.; Hebert, P.D.N. Semi-automated, Membrane-Based Protocol for DNA Isolation from Plants. Plant Mol. Biol. Rep. 2008, 26, 186–198. [Google Scholar] [CrossRef]
- Whitlock, R.; Hipperson, H.; Mannarelli, M.; Burke, T. A High Throughput Protocol for Extracting High-Purity Genomic DNA from Plants and Animals. Mol. Ecol. Resour. 2008, 8, 736–741. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Y. A Pattern Matching Algorithm for Double-Type Characters. Appl. Mech. Mater. 2014, 571-572, 461–464. [Google Scholar] [CrossRef]
- Hart, M.W.; Sunday, J. Things fall apart: biological species form unconnected parsimony networks. Biol. Lett. 2007, 3, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.M.; Cabezas, M.P.; I Tavares, A.; Xavier, R.; Branco, M. tcsBU: a tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Librado, P.; Sanchez-Delbarrio, J.; Messeguer, X.; Rozas, R. DnaSP 5. 10. 00 2009.
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Developments in Environmental Modelling; Elsevier: Amsterdam, The Netherlands; Oxford, UK, 2012; Volume 24, ISBN 978-0-444-53868-0. [Google Scholar]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Studer, M.; Gonzalez, J. Package ‘Cluster’ 2023.
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses, R Package v. 1.0. 7 2020.
- Reshaid, Y.; Cao, L.; Brea, F.; Blanche, M.O.; Torres, S.; Darrigran, G. Variation in the distribution of Corbicula species (Mollusca: Bivalvia: Corbiculidae) after 25 years of its introduction in the Río de la Plata, Argentina. Zoöl. 2017, 34, 1–6. [Google Scholar] [CrossRef]
- Peñarrubia, L.; Araguas, R.-M.; Vidal, O.; Pla, C.; Viñas, J.; Sanz, N. Genetic characterization of the Asian clam species complex (Corbicula) invasion in the Iberian Peninsula. Hydrobiologia 2016, 784, 349–365. [Google Scholar] [CrossRef]
- Bespalaya, Y.V.; Bolotov, I.N.; Aksenova, O.V.; Kondakov, A.V.; Gofarov, M.Y.; Laenko, T.M.; Sokolova, S.E.; Shevchenko, A.R.; Travina, O.V. Aliens are moving to the Arctic frontiers: an integrative approach reveals selective expansion of androgenic hybrid Corbicula lineages towards the North of Russia. Biol. Invasions 2018, 20, 2227–2243. [Google Scholar] [CrossRef]
- Renard, E.; Bachmann, V.; Cariou, M.L.; Moreteau, J.C. Morphological and molecular differentiation of invasive freshwater species of the genus Corbicula (Bivalvia,Corbiculidea) suggest the presence of three taxa in French rivers. Mol. Ecol. 2000, 9, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Sousa, R.; Freire, R.; Rufino, M.; Méndez, J.; Gaspar, M.; Antunes, C.; Guilhermino, L. Genetic and shell morphological variability of the invasive bivalve Corbicula fluminea (Müller, 1774) in two Portuguese estuaries. Estuarine, Coast. Shelf Sci. 2007, 74, 166–174. [Google Scholar] [CrossRef]
- Pereira, D.; Dreher Mansur, M.C.D.; Duarte, L.D.S.; de Oliveira, A.S.; Pimpão, D.M.; Callil, C.T.; Ituarte, C.; Parada, E.; Peredo, S.; Darrigran, G.; et al. Bivalve distribution in hydrographic regions in South America: historical overview and conservation. Hydrobiologia 2014, 735, 15–44. [Google Scholar] [CrossRef]
- Ituarte, C.F. Growth Dynamics in a Natural Population of Corbicula fluminea (Bivalvia Sphaeriacea) at Punta Atalaya, Rio de La Plata, Argentina. Stud. Neotrop. Fauna Environ. 1985, 20, 217–225. [Google Scholar] [CrossRef]
- Bódis, E.; Nosek, J.; Oertel, N.; Tóth, B.; Fehér, Z. A Comparative Study of Two Corbicula Morphs (Bivalvia, Corbiculidae) Inhabiting River Danube. Int. Rev. Hydrobiol. 2011, 96, 257–273. [Google Scholar] [CrossRef]
- Szarmach, D.; Wiśniewski, K.; Kobak, J.; Kakareko, T.; Labecka, A.M.; Sousa, R.; Poznańska-Kakareko, M. Differences in substratum preferences and behaviour within the invasive Corbicula species complex. Freshw. Biol. 2023, 68, 1489–1502. [Google Scholar] [CrossRef]
- Ituarte, C.F. Aspectos Biológicos de Las Poblaciones de Corbicula largillierti Phillippi (Mollusca Pelecypoda) En El Río de La Plata. Rev. del Mus. La Plata (Nueva Ser. 1984, Tomo XIII, 231–247.
- Darrigran, G.A.; Maroñas, M.E. Crecimiento de Las Poblaciones Naturales de Corbicula fluminea (Müller, 1774) y C. largillierti (Phillippi, 1844) (Bivalvia: Sphaeriacea) En El Litoral de Punta Blanca, Estuario Del Río de La Plata, República Argentina. Comun. la Soc. Malacológica del Uruguay 1989, VII, 139–157.
- Mansur, M.C.D.; Vanin, A.S.; Bergonci, P.E.A.; Oliveira, A.S. de Dinâmica Reprodutiva de Corbicula fluminea e Corbicula largillierti. In Moluscos límnicos invasores no Brasil: biologia, prevençâo e controle; Mansur, M.C.D., dos Santos, C.P., Pereira, D., Padula Paz, I.C., Zurita, M.L.L., Raya Rodriguez, M.T., Nehrke, M.V., Bergonci, P.E.A., Eds.; Redes Editora: Porto Alegre, 2012; pp. 119–124. [Google Scholar]
- Marescaux, J.; Falisse, E.; Lorquet, J.; Van Doninck, K.; Beisel, J.-N.; Descy, J.-P. Assessing filtration rates of exotic bivalves: dependence on algae concentration and seasonal factors. Hydrobiologia 2016, 777, 67–78. [Google Scholar] [CrossRef]
Figure 1.
Internal and external views of the left valve of Corbicula form A/R (C. fluminea sensu lato; Müller, 1774) and form C/S (C. largillierti sensu lato; Phillipi, 1844), along with the diagnostic characters utilized for morphological assignment of lineages in the present study.
Figure 1.
Internal and external views of the left valve of Corbicula form A/R (C. fluminea sensu lato; Müller, 1774) and form C/S (C. largillierti sensu lato; Phillipi, 1844), along with the diagnostic characters utilized for morphological assignment of lineages in the present study.
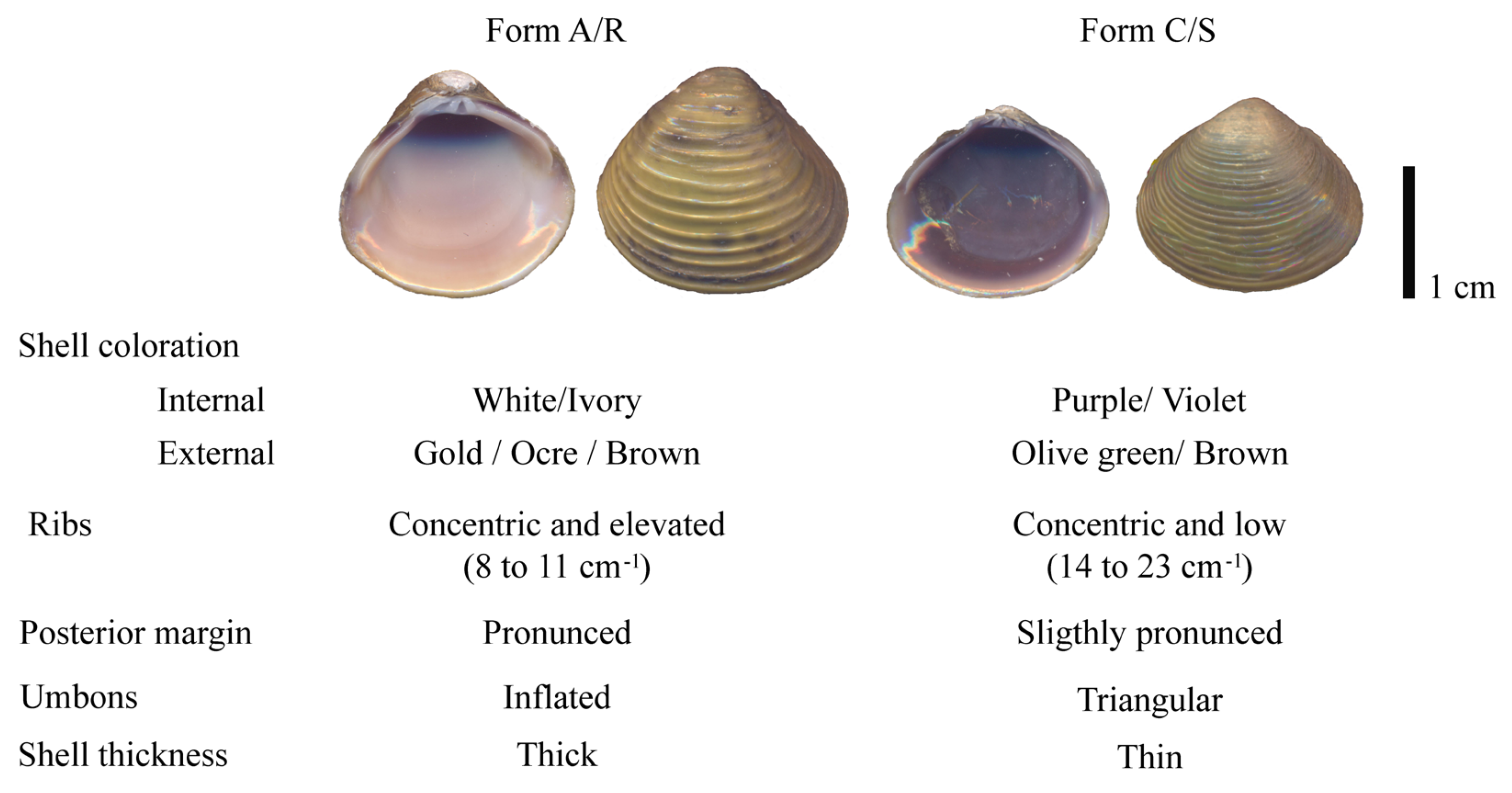
Figure 2.
Examples of images taken to measure the gill area: (a) external demibranch; and (b) internal demibranch), palp area (c), and filament density (d) in Corbicula specimens collected in Argentina between 2015 and 2017.
Figure 2.
Examples of images taken to measure the gill area: (a) external demibranch; and (b) internal demibranch), palp area (c), and filament density (d) in Corbicula specimens collected in Argentina between 2015 and 2017.
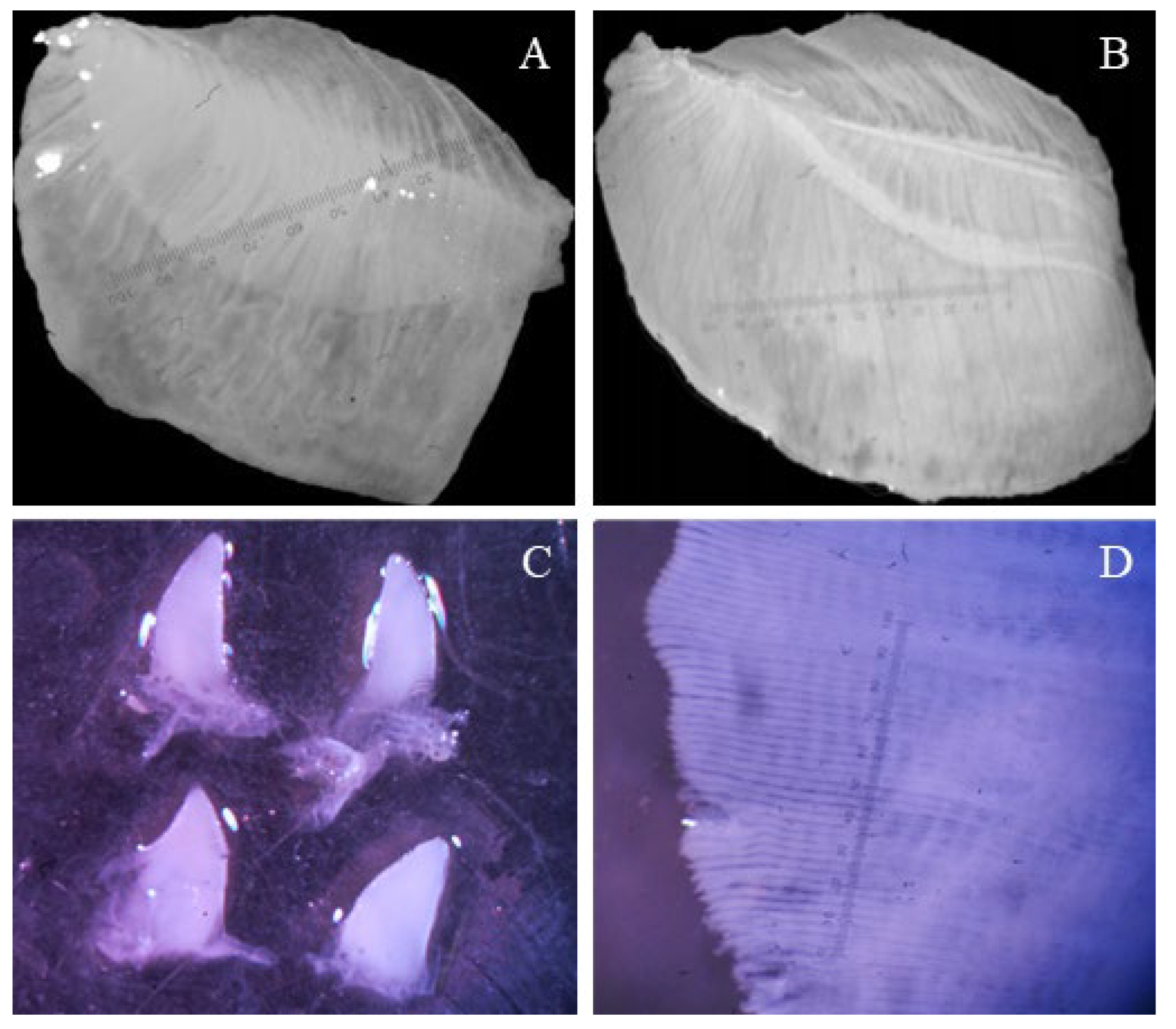
Figure 4.
(a) Sampling locations and the geographic distribution of the COI haplotypes for Corbicula clams collected at 15 sites between 2015 and 2017. Pie charts depict the proportion of haplotypes observed at each site corresponding to the two freshwater lineages found within the country, with FW5 in black and FW17 in white. The border color of each pie chart indicates the identified form (see codes in panel (b)). (b). Unrooted statistical parsimony network of Corbicula clams based on 658 bp of COI, focusing on the 210 specimens from Argentina sequenced in this study and representative of the freshwater (FW) haplogroups FW5 and FW17. Solid lines represent single mutational steps, empty circles represent intermediate, unsampled haplotypes, and circle sizes are proportional to the number of identical haplotypes. Haplotypes are color-coded based on the phenotypic form of each specimen, and major FW haplogroups are indicated with dashed arrows and circles over the network. Codes and details regarding sampling sites can be found in Table 1.
Figure 4.
(a) Sampling locations and the geographic distribution of the COI haplotypes for Corbicula clams collected at 15 sites between 2015 and 2017. Pie charts depict the proportion of haplotypes observed at each site corresponding to the two freshwater lineages found within the country, with FW5 in black and FW17 in white. The border color of each pie chart indicates the identified form (see codes in panel (b)). (b). Unrooted statistical parsimony network of Corbicula clams based on 658 bp of COI, focusing on the 210 specimens from Argentina sequenced in this study and representative of the freshwater (FW) haplogroups FW5 and FW17. Solid lines represent single mutational steps, empty circles represent intermediate, unsampled haplotypes, and circle sizes are proportional to the number of identical haplotypes. Haplotypes are color-coded based on the phenotypic form of each specimen, and major FW haplogroups are indicated with dashed arrows and circles over the network. Codes and details regarding sampling sites can be found in Table 1.
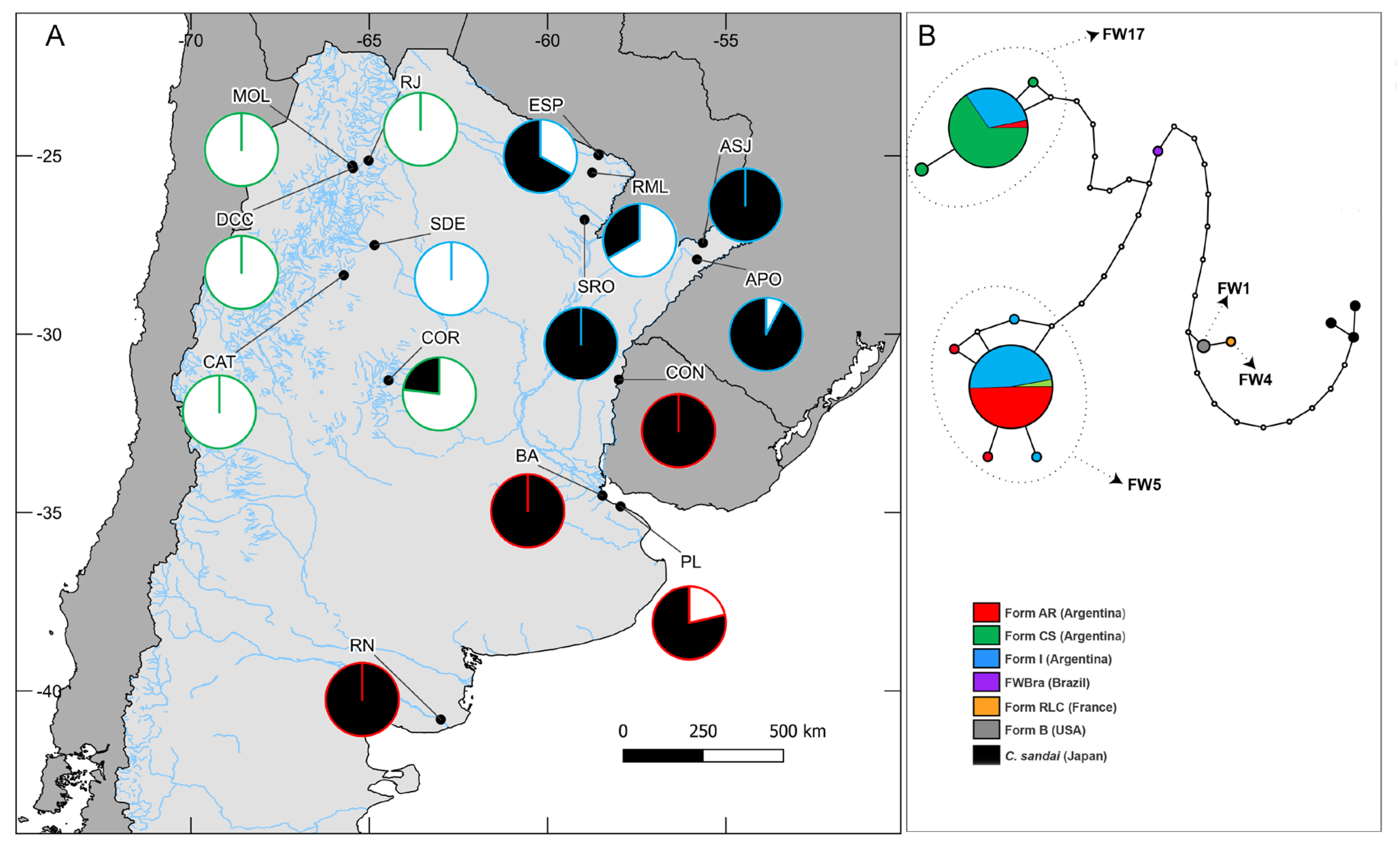
Figure 5.
Principal Component Analysis of morphometric variables of 225 Corbicula clams sampled from 15 populations in Argentina between 2015 and 2017. Each data point represents an individual, with colors indicating the probability of belonging to different clusters based on the fuzzy cluster analysis. Population centroids are indicated by triangles. Ellipses represent the morphospace containing 90% of individuals from each cluster (solid lines) and indeterminate individuals (dotted line) obtained from fuzzy cluster analyses. Variable codes correspond to those in Table 3, and population codes correspond to those in Table 1.
Figure 5.
Principal Component Analysis of morphometric variables of 225 Corbicula clams sampled from 15 populations in Argentina between 2015 and 2017. Each data point represents an individual, with colors indicating the probability of belonging to different clusters based on the fuzzy cluster analysis. Population centroids are indicated by triangles. Ellipses represent the morphospace containing 90% of individuals from each cluster (solid lines) and indeterminate individuals (dotted line) obtained from fuzzy cluster analyses. Variable codes correspond to those in Table 3, and population codes correspond to those in Table 1.
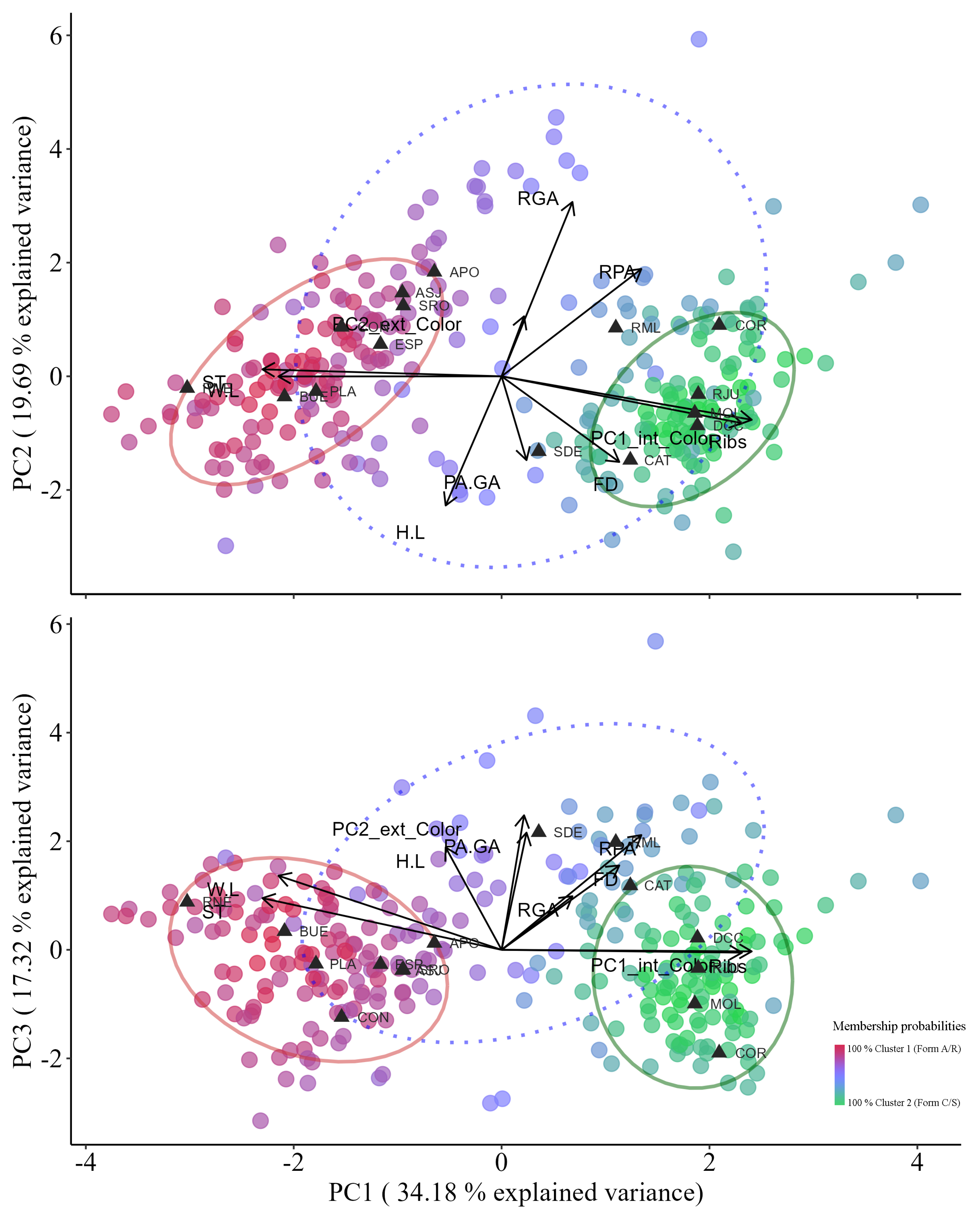

Table 1.
Identification of haplotypes of the mitochondrial cytochrome C oxidase subunit I (COI) gene in 15 Corbicula populations in Argentina, along with their correspondence with haplotypic groups and global forms as defined in Pigneur et al. (2014).
Table 1.
Identification of haplotypes of the mitochondrial cytochrome C oxidase subunit I (COI) gene in 15 Corbicula populations in Argentina, along with their correspondence with haplotypic groups and global forms as defined in Pigneur et al. (2014).
| ID | Region; Province | Water body; Basin | Lat. Lon. | N | Haplotypic group | Form |
| DCC | Northwest; Salta | Cabra Corral reservoir; Río de la Plata basin | -25,34;$$$-65,45 | 15 | FW17 | C/S |
| RJU | Northwest; Salta | Juramento river; Río de la Plata basin | -25,13;$$$-65,01 | 11 | FW17 | C/S |
| MOL | Northwest; Salta | Moldes irrigation ditch; Río de la Plata basin | -25,27;$$$-65,47 | 15 | FW17 | C/S |
| SDE | Northwest; Sgo. del Estero | Dulce river; Mar Chiquita basin | -27,50;$$$-64,85 | 15 | FW17 | C/S and Corbicula sp. |
| CAT | Northwest; Catamarca | del Valle river; Del Valle basin | -28,34;$$$-65,71 | 15 | FW17 | C/S |
| COR | Center; Córdoba | Cosquín river; Mar Chiquita basin | -31,29;$$$-64,46 | 13 | FW17(10); FW5(3) | C/S |
| PLA | Estuary; Buenos Aires | Río de la Plata estuary | -34,82;$$$-57,95 | 14 | FW5(11); FW17(3) | A/R |
| BUE | Estuary; Buenos Aires | Río de la Plata estuary | -34,52;$$$-58,45 | 14 | FW5 | A/R |
| RNE | Patagonia; Río Negro | Negro river | -40,80;$$$-62,99 | 15 | FW5 | A/R |
| CON | East plain river; Entre Ríos | Ayuí Grande stream; Río de la Plata basin | -31,27;$$$-58,00 | 15 | FW5 | A/R |
| APO | Northest; Misiones | Chirimay stream; Río de la Plata basin | -27,90;$$$-55,81 | 14 | FW5 | Corbicula sp. |
| ASJ | Northest; Misiones | San Juan stream; Río de la Plata Basin | -27,44;$$$-55,64 | 13 | FW5(12); FW17(1) | Corbicula sp. |
| SRO | Northest; Chaco | de Oro river; Río de la Plata basin | -26,78;$$$-58,96 | 14 | FW5 | Corbicula sp. |
| ESP | Northest; Formosa | Porteño river; Río de la Plata basin | -24,97;$$$-58,56 | 12 | FW5(8); FW17(4) | Corbicula sp. |
| RML | Northest; Formosa | Monte Lindo river; Río de la Plata basin | -25,47;$$$-58,75 | 15 | FW5(5); FW17(10) | Corbicula sp. |
Table 2.
Summary statistics for the two major haplogroups of Corbicula found in Argentina in this study. The table shows the sample size (n), nucleotide diversity (π), and number of specimens of each phenotypic form for each lineage, as well as the mean uncorrected genetic distance between the two haplogroups (p-distance, %).
Table 2.
Summary statistics for the two major haplogroups of Corbicula found in Argentina in this study. The table shows the sample size (n), nucleotide diversity (π), and number of specimens of each phenotypic form for each lineage, as well as the mean uncorrected genetic distance between the two haplogroups (p-distance, %).
| Haplogroup (Argentina) | ||
| FW5 | FW17 | |
| n | 111 | 99 |
| π | 0.00013 | 0.00012 |
| Form A/R | 55 | 3 |
| Form C/S | 3 | 66 |
| Form I | 53 | 30 |
| p-distance | 2.28% | |
Table 3.
Results of the principal component analysis (PCA). The table shows the scores obtained for each independent variable for each factor. Values higher than 0.6 are indicated in bold font.
Table 3.
Results of the principal component analysis (PCA). The table shows the scores obtained for each independent variable for each factor. Values higher than 0.6 are indicated in bold font.
| Variable | Principal Component | ||
| 1 | 2 | 3 | |
| PC1_int_Color | 0.89 | -0.21 | -0.01 |
| PC2_ext_Color | 0.08 | 0.30 | 0.65 |
| H:L | -0.20 | -0.64 | 0.50 |
| W:L | -0.79 | 0.00 | 0.36 |
| ST | -0.85 | 0.03 | 0.25 |
| RGA | 0.25 | 0.86 | 0.26 |
| Ribs | 0.86 | -0.23 | -0.01 |
| RPA | 0.50 | 0.53 | 0.56 |
| PA:GA | 0.09 | -0.41 | 0.57 |
| FD | 0.42 | -0.42 | 0.41 |
| Standard deviation | 1.85 | 1.40 | 1.32 |
| Proportion of Variance | 0.34 | 0.20 | 0.17 |
| Cumulative Proportion | 0.34 | 0.54 | 0.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



