Submitted:
07 May 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
Panax notoginseng (Burk.) F. H. Chen, a valuable Chinese herb in Yunnan Province, is susceptible to leaf spot caused by Boeremia linicola. Nonetheless, effective control methods against leaf spot in P. notoginseng are lacking. We surveyed understorey P. notoginseng bases in Lincang, Lancang, and Xundian, with incidence rate reaching 80% and an average disease index of 25 in severe cases. We found that pathogenic bacteria overwinter in infected leaves and soil, with conidia and conidiophores serving as the primary infection source. The strain B. linicola LYB-2 showed the best growth at 20°C and pH 5.0. We also screened control agents using the plate and face-off culture methods. Tetramycin exhibited 38.56–74.67% inhibition with an EC50 of 0.60 mg/L. Copper abietate showed 3.10–34.76% inhibition with an EC50 of 3,120.05 mg/L. Trichoderma atroviride and T. harzianum exhibited inhibition rates of 52.48% and 44.19%, respectively, while Bacillus subtilis reached 78.45%. Field tests demonstrated significant control effects with 20% copper abresinate, B. subtilis, T. virenceae BH-10 preparation, and 0.3% tetramycin, achieving 82.57%, 72.56%, 71.3%, and 70.2% inhibition, respectively. These findings support their eco-friendly use for preventing and controlling leaf spot disease in P. notoginseng forests; we provide vital scientific evidence for disease prediction and green control strategies.
Keywords:
Panax notoginseng
; leaf spot disease
; occurrence regularity
; biological characteristics
; green control strategies
1. Introduction
Panax Notoginseng is a perennial herb belonging to the family Acanthaceae that has been artificially planted for over 400 years [1]. Moreover, it exhibits anti-inflammatory and other pharmacological properties and is widely used in the treatment of conditions such as coronary heart disease including angina pectoris [2,3,4]. In recent years, with people being more conscious about health care, the demand for high quality P. notoginseng has become more urgent [5]. However, obstacles related to continuous cropping have seriously restricted the sustainable development of P. notoginseng. Agricultural scientists in China have developed a new ecological cultivation model for underforest P. notoginseng by intercropping a model of P. notoginseng and pine forests named PPF, which prioritizes medicinal efficacy by leveraging mutual reinforcement among understory organisms and integrating the ecological requirements for P. notoginseng growth with the forest ecosystem. Thus, achieving standardized and large-scale production of P. notoginseng in underforest locations [8]. The PPF alleviate the land pressure caused by the obstacles of continuous P. notoginseng cultivation, save production costs, better solve the difficult problems of heavy metals and excessive agricultural residues in traditional cultivation, and improve the quality of P. notoginseng from the source [9]. By 2020, the PPF had exceeded 1,333 hm2, serving as a strong technical strategy for the high-quality production of the medicinal herbs cultivated in underforests based on the principle of habitat diversity for pest control [10].
The planting process of PPF includes avoiding the use of fertilizers or pesticides. However, the plant remains vulnerable to diseases, significantly impacting both yield and quality. Underground root rot is one of the main diseases affecting the growth and yield of P. notoginseng in forests, significantly reducing yields [8]. The incidence rate ranges from 5% to 20%, with severity reaching as high as 60–70%. Research indicates a complex array of pathogens, including Ilyonectria, Plectosphaerella, Clonostachys, Gibberella, Fusarium, and Chalara, contributing to P. notoginseng root rot [8,9]. Root rot exacerbates challenges associated with continuous cropping in P. notoginseng, worsening with each planting cycle. Aboveground, the plant is predominantly affected by black spot and round spot diseases. Black spot disease caused by Alternaria alternata, it can occur year-round, predominantly affecting leaves. Signs include the formation of round or irregular water-soaked brown spots on leaves, gradually expanding and leading to plant death [11]. The average annual incidence rate ranges from 20% to 35%, with severe cases exceeding 90% [12]. Round spot disease, caused by Mycocentrospora acerina (R. Hartig) Deighton, is a disease with devastating effects on the growth of P. notoginseng. This disease exhibits a sudden onset and rapid spread, affecting all parts of the plant. Initially, affected leaf spots measure 5–20 mm in diameter, appearing watery, and later develop into brown whorls with a pinpoint-sized center. A transparent point with a needle tip is visible within the lesion, accompanied by a yellow halo at the boundary between diseased and healthy tissue. In humid conditions, lesions turn reddish-brown and develops gray-white mildew on the surface in advanced stages. When multiple lesions occur on leaves, they merge, rot, and eventually detach. Round spot disease leads to annual losses of approximately 20–50%, with severe cases resulting in sterilization [13]. In 2021, a novel disease, B. linicola causing P. notoginseng leaf spots, emerged in the planting base of Lincang City, Yunnan Province, with an incidence rate of 31% and a disease index of 10–20% [14,15]. This new disease poses a significant threat to the healthy development of the P. notoginseng industry in forested areas. B. linicola is implicated in various plant diseases, including leaf spots in white clover, Siberian ginseng, and sage, resulting in leaf wilt and production losses [16,17,18]. Tomato stem and leaf rot are also reported due to B. linicola infection [19].
The PPF model requires that no chemical agents be used in agricultural production. Research into P. notoginseng disease control primarily focuses on agricultural, and biological control methods. Biological control offers multiple advantages, such as sufficient resources, low investment, and minimal pollution [20]. Therefore, the development of biogenic pesticides from plant sources and mineral fungicides has attracted considerable attention. Utilizing biocontrol bacteria such as Bacillus and Trichoderma to produce biological control agents shows promise for preventing and managing P. notoginseng diseases [21,22]. Tetramycin, a novel polyene antibiotic, has been isolated from Streptomyces culture broth, representing a potential advancement in disease management strategies [23]. It belongs to the group of tetraene macrolide antibiotics, which can inhibit mycelial growth and conidial germination of Fusarium graminis and Alternaria, thereby reducing occurrences of wheat scab and kiwifruit rot [24,25]. Wood vinegar (WV), comprising water and organic acids, serves as a plant fungicide and insecticide [26]. WV at a concentration of more than 2.25% can inhibit the mycelial growth and spore germination of P. ginseng black spot and gray mold bacteria belonging to the family Araliaceae [27]. Allelopathic substances secreted by pine play an important role in the organic cultivation of P. notoginseng under forest canopies. The main component of pine needle volatiles, α-pinene, can promote the growth of P. notoginseng and induce resistance, while α-terpineol, a specific antibacterial component of Pinus yunnanensis, can significantly inhibit the growth of pathogenic bacteria. At present, mineral fungicides, mainly Bordeaux liquid, stone sulfur mixture, copper sulfate, and copper pine ester, are widely used. Many copper preparations represent mineral-derived, multi-action site fungicides with outstanding bactericidal performance, low toxicity, low residue, and minimal pollution, thus finding widespread use in plant disease prevention and control [28].
Therefore, it is necessary to fully understand the occurrence and epidemic characteristics of P. notoginseng leaf spot in order to better provide effective prevention and biological control strategies. In this study, we aimed to explore the biological characteristics, pathogenesis, and biocontrol strategies of pathogenic bacteria causing P. notoginseng leaf spots. Our findings provide scientific insights to support effective prevention and control of P. notoginseng leaf spots in forest environments. It provides the basis for promoting the sustainable, stable, and high-quality development of the Chinese herbal medicine planting industry under the forest.
2. Materials and Methods
2.1. Prevalence and Incidence of B. linicola in Yunnan P. notoginseng Fields
We conducted a census of the understorey P. notoginseng bases in Lincang, Lancang, and Xundian to assess the occurrence of P. notoginseng leaf spot disease. At each investigation point, we documented the number of affected plants, disease severity, and total P. notoginseng plant count. Subsequently, we calculated the incidence rate and severity of the disease, categorizing severity based on spot number and leaf area occupation [29]. Additionally, we performed a dynamic survey on P. notoginseng leaf spot disease occurrence in organic P. notoginseng planted under the forest canopy in Boshang Town, Linxiang District, Lincang City, Yunnan Province (23.70°N, 100.12°E). Starting from the initial stage of leaf spot disease in July, we conducted monthly surveys using a five-point sampling method, with each point covering a 1 m2 area. We recorded and counted the incidence and severity of P. notoginseng leaf spot disease (Table 1). An automatic temperature and humidity recorder (HOBO MX2301A, USA) was installed in the P. notoginseng forest garden and set to record the temperature and air relative humidity data at hourly intervals. Data were collected once a month to calculate the average, minimum, and maximum monthly temperature and humidity. Two meteorological factors (temperature and relative humidity) affecting the occurrence of leaf spots in P. notoginseng in 2022 were analyzed using Pearson’s correlation analysis.
Incidence rate is calculated by the following formula:
The severity of the disease is expressed by the disease index, which is calculated according to the following formula:
2.2. Morphological Study
P. notoginseng leaf-spotted fungus LYB-2 (B. linicola), isolated from P. notoginseng under the forest of Lincang Xiaodaohe Forest Farm by the National Engineering Research Center of Biodiversity Application Technology, Yunnan Agricultural University, was verified using Koch’s rule.
Strain LYB-2 was activated on a PDA plate and placed in a constant temperature incubator with full illumination (EYELASLI-700, Tokyo, Japan) at 20℃. After 7 d, the morphology, color, and size of the colonies and mycelia were observed via microscopy (Olympus BX43F optical microscope, Japan). Subsequently, the morphology, color, and size of the colonies were examined under a microscope upon conidia and conidiophores production.
2.3. Growth Characteristics at Different Temperatures
The optimum temperatures for representative strains were evaluated. The representative strain on the PDA medium was incubated under varying temperatures of 8°C, 12°C, 16°C, 20°C, 22°C, 26°C, 28°C, 30°C, and 32°C in the dark. When the control colony diameter reached 2/3 of the plate diameter, the cross method was used to measure the colony diameter.
2.4. Growth Characteristics at Different pH Values
The optimum pH for representative strains was assessed. The pH of the medium was adjusted using 1.0 mol/L HCl or NaOH (Sinopharm Chemical Reagent Co., Ltd, Shanghai, China). The representative strain was inoculated on PDA medium with pH values of 4.0, 5.0, 6.0, 7.0, 8.0, 9.0, 10.0, and 11.0 for the growth assay and sporulation, and then incubated in the dark at 20℃. When the control colony diameter reached 2/3 of the plate diameter, the cross method was used to measure it.
2.5. Study on the Source of B. linicola Primary Infection
2.5.1. Investigation of Infection Sources
The rhizosphere soil from the affected area was collected and sealed in sterile polyethylene bags. The soil was returned to the laboratory for immediate microbial isolation and the detection of pathogenic spores. P. notoginseng leaf spot disease remnants, including P. notoginseng leaves, ground leaves (with obvious spots but no mold layer), stems, taproots, and fibrous roots, were also collected. These samples were placed in sterile paper bags, marked, and brought back to the laboratory. The tissue separation method was used to isolate pathogens and identify the isolates, with five isolates per dish and 10 dishes in total. The growth of pathogenic bacteria in the tissues of the diseased remains was observed, and the number of pathogenic bacterial colonies was recorded. Common plants around the leaf spots of P. notoginseng were collected, especially those with similar signs, and brought back to the laboratory for isolation and identification to determine whether they were intermediate hosts and overwintering sites for pathogenic bacteria.
2.5.2. Investigation into Transmission Mode
From August to November 2022, spore traps (Zhejiang Top Cloud-Agri Technology Co., Ltd TPBZ3 Portable spore catcher, Zhejiang, China) were installed at the planting bases of P. notoginseng in the forest to capture airborne pathogens. The spore catcher was placed on level ground (Figure 1a). Starting at 8:00 am every day, slides coated with a mixture of carbon tetrachloride and petroleum jelly (100 mL carbon tetrachloride dissolved in 10 g petroleum jelly) were assembled. The number of spores adsorbed on the slides with pathogenic spores of P. notoginseng leaf spot was determined by changing the slides every day. Full-slide detection was performed using a microscope. A natural collection method was used for auxiliary collection (Figure 1b). Slides coated with carbon tetrachloride petroleum jelly were placed directly on the ground to collect spores, with five slides being placed under diseased plants. The collection time ranged from 8:00 a.m. to 6:00 p.m., and the slides were changed daily. Additionally, 5 d per month, slides were viewed under a microscope.
2.6. Sensitivity Testing of B. linicola to Different Fungicides
2.6.1. Determination of Antibacterial Activity of Five Drugs
The antibacterial activities of tetramycin(LiaoningWkioc Bioengineering Co., Ltd, Liaoning, China), copper abietate(Jiangxi Heyi Chemical Co., Ltd, Jiangxi, China), α-terpineol(Shanghai Yien Chemical Technology Co., Ltd, Shanghai, China), α-pinene(Sanghai Macklin Biochemical Co., Ltd, Shanghai, China), and matrine WV(Shanxi Dewei Biochemical Co., Ltd, Shanxi, China) were determined using the mycelium growth rate method. The tested agents were added into PDA medium at a temperature of 40–50 ℃ and then poured into 90 mm diameter Petri dishes. Plates with varying concentrations were prepared, using the same amount of sterile water as the blank control. The concentrations of the tested agents are listed in Table 2, Table 3 and Table 4. The 5.0-mm diameter mushroom cake was transferred to the center of medicated PDA medium containing different concentration gradients, and unmedicated PDA medium was used as the control group. Each concentration was repeated three times. When the control colony grew to 2/3 of the Petri dish size, the cross method was used to measure the diameter of each treated colony.
2.6.2. Det
The bacteriostatic activities of T. harziensis and B. subtilis were determined using the plate confrontation method. Trichoderma and B. subtilis were incubated at 20℃ for three iterations in each treatment. When the control colony grew to 2/3 of the Petri dish size, the diameter was measured using the cross method. The inhibition rate was calculated using the following equation:
The concentration value (mg/L) of each drug was converted into a pair value (X), and the corresponding inhibition rate was converted into a probability value (Y). The regression equation for virulence was established using the least-squares method to calculate the half-maximal effective concentration (EC50) of each drug.
2.6.3. Field Trials
The experiment was conducted at the Xiasanqi Base of the Liangshan Forest, Boshang Town, Linxiang District, Lincang City, Yunnan Province (23.70°N, 100.12°E). The predominant forest species was Yunnan pine, while the P. notoginseng plants were of 2-year growth, exhibiting similar growth potential. The row spacing of P. notoginseng was maintained at 10–15 cm × 10–15 cm. Uniformity was ensured across experimental plots in terms of soil type, cultivation management, fertilizer application, and water conditions. The primary objective of the trial was to identify leaf spot occurrences in P. notoginseng within forest environments.
By combining the screening results of laboratory agents with the recommended dosage, seven agents including 0.3% tetramycin (LiaoningWkioc Bioengineering Co., Ltd, Liaoning, China), 300 million CFU/g Trichoderma kirhartz (BioWorks, Inc., USA), 10 billion CFU/g Bacillus quinquefolium (BioWorks, Inc., USA), 20% copper abietate (Jiangxi Heyi Chemical Co., Ltd, Jiangxi, China), 99% α-terpineol(Shanghai Yien Chemical Technology Co., Ltd, Shanghai, China), 97% α-pinene (Sanghai Macklin Biochemical Co., Ltd, Shanghai, China), and matrine WV (Shanxi Dewei Biochemical Co., Ltd, Shanxi, China), were selected and diluted to different concentrations. Table 3 lists the treatments administered for each drug.
The plots of the experimental agents and blank control were arranged according to random combinations; the plot area was 1.2 m × 5 m, each agent was applied three times, and each treatment was repeated three times. A knapsack sprayer was used to spray under still or breezy weather conditions, avoid applications under high temperature, humid, or rainy conditions to ensure even coverage of the liquid on the leaves of P. notoginseng. To verify the efficacy of the agent, application was scheduled before the peak occurrence of P. notoginseng leaf spot under the forest, specifically on October 25, with application occurring once. We selected 80–120 P. notoginseng plants in the plot to perform a fixed-point survey according to the proportion of the disease site to the whole plant. The total number of plants and the number of different disease levels were recorded. The grading criteria for leaf spots of P. notoginseng were consistent with those described in Section 2.1. The disease-based survey was conducted 1 d before application and repeated 15 d after application.
The incidence and disease index were calculated in the same manner as described in Section 2.1, using the following formulas:
The disease index growth rate and prevention and treatment effect are calculated according to the following formula:
where pt0, pt1, CK0, and CK1 represent the disease index before application to the treatment area, the disease index after application to the treatment area, the disease index before application to the control area, and the disease index after application to the control area, respectively.
2.7. Statistical Analysis
Statistical analyses were conducted using Excel 2019 and SPSS 19.0. Graphing was conducted using GraphPad 9.0. One-way analysis of variance and multiple comparisons were performed to evaluate the levels of significance level among the samples. DPS V9.5 was used to calculate the virulence regression equation and EC50 values. All tests were repeated thrice.
3. Results
3.1. Prevalence and Incidence of B. linicola in Yunnan P. notoginseng Fields
The occurrence of P. notoginseng leaf spot disease was studied in a P. notoginseng plantation located in the Xiaohe forest, Boshang Town, Linxiang District, Lincang City, Yunnan Province. The disease primarily affects P. notoginseng leaves, impacting plants of various growth stages (Figure 2a, b). Initially, brown water-stained spots develop on the leaves, gradually enlarging and forming black granular prominences, often encircled by yellow halos (Figure 2c). As the disease progresses, multiple lesions coalesce, causing the leaves to yellow and eventually drop (Figure 2b). In warmer temperatures, lesions dry up, crack, and perforate without distinct yellow halos (Figure 2d).
3.1.2. Epidemics of P. notoginseng Leaf Spot under Forest Conditions
In mid-August 2022, surveys conducted across multiple P. notoginseng underwood sites in Linxiang District, Lancang County, Puer City, Xundian City, and Kunming City revealed varying degrees of leaf spot occurrences. Specifically, within the underwood P. notoginseng plantation areas of Boshang Town, Linxiang District, and Lincang City, including Xiaohe, Liangshan, and Shuangyingpan, varying degrees of leaf spots were observed. The incidence rates of the Xiaohe and Liangshan bases were 7.061% and 7.221%, respectively, while the incidence rate of the Shuangying Base was lower (1.031%). The disease did not occur in the Datangzi, Donghui, or Xundian areas of Lancang (Figure 3).
Through a survey conducted from July 2022 to December 2022, we found that the disease began in mid-July, with an average incidence of 4.87%. The incidence and disease index increased in mid-August, and the incidence increased significantly in mid-September until the peak of the disease in mid-November, with an average incidence of 42.62%. The incidence rate was 80%, with an average disease index of 25 (Figure 4).
Pearson’s correlation coefficient analysis was conducted to examine the relationship between the leaf spot disease index of P. notoginseng and meteorological factors, as presented in Table 4. The findings revealed strong negative correlations between the disease index and monthly mean temperature (-0.93), monthly minimum temperature (-0.89), monthly mean relative humidity (-0.96), and monthly minimum relative humidity (-0.97). Air temperature and humidity were strongly correlated with the occurrence of leaf spots in P. notoginseng.
Analysis of the dynamic monitoring results on P. notoginseng leaf spot occurrences identified August 15 as a critical time point for the onset, while October 15 marked the rapid outbreak phase (Figure 5a, b). Subsequent examination of meteorological conditions before and after these key dates revealed notable trends. From August, the temperature began to decrease, and the disease index increased. In September and October, the average monthly temperature dropped to 15.6℃ and 13.7℃, respectively, while the disease index surged considerably leading to an epidemic of leaf spots. By mid-November, the disease worsened. These results indicate that temperature is a decisive factor affecting the occurrence of leaf spots in P. notoginseng.
3.2. Morphological Study
The mycelium was carefully extracted from the PDA plate and cultured under full light at 20℃ for 7 d to facilitate microscopic examination. Observations revealed distinctive characteristics of both the colony and mycelium: the colony displayed a grayish-brown center with a white margin on the front (Figure 6a), while the back exhibited a brown to black center with a white margin (Figure 6b). The mycelia were transparent, exhibiting colors ranging from colorless to brown, and featured septate, axonomically branched structures, predominantly with sharp-angle-bifurcated branches. As the mycelia aged, they turned brown, with enlarged lotus joints observed (Figure 6c). After 60 d of culture, the conidia were observed to be spherical to subspherical, generally devoid of hair or exhibiting minimal mycelial growth, and ranged from dark brown to black, measuring from 22.46 µm to 155.94 µm in length and from 18.20 µm to 130.5 µm in width (Figure 6f).
3.3. Growth Characteristics at Different Temperatures
As shown in Figure 7, temperature has a significant impact on the growth of the mycelia of pathogenic bacteria. Specifically, at 20°C, the colony diameter reached 6.2 cm after 7 days, significantly surpassing growth observed at other temperature conditions. Conversely, at 32°C, mycelial growth was markedly inhibited. The results suggest that the optimum temperature for pathogen growth lies at 20℃.
3.4. Growth Characteristics at Different pH Values
As can be seen from Figure 8, all pathogens grew in the pH range of 4.0–11.0. However, the most favorable pH range for growth was found to be 4.0–6.0, with pH 5.0 being the most conducive for pathogenic bacterial growth. After 7 d of culture, the colony diameter reached 7.75 cm, which was significantly higher than the colony diameter at pH 4.0 and 6.0. These results suggest that the pathogen thrives optimally in a weakly acidic environment, based on its preferred pH range.
3.5. Study on the Source of Primary Infection of B. linicola
3.5.1. Investigation into Infection Sources
Investigation into infection sources involved isolating and identifying culturable microorganisms from rhizosphere soil treated with P. notoginseng leaf spots, employing the dilution-coated plate method, with results depicted in Figure 9. Among the bacterial colonies growing in the soil bacterial suspensions at different concentrations, only one suspected pathogenic bacterium was identified. After purification, the results were consistent with the morphological characteristics of the colony of B. linicola on PDA. Figure. 9e showed that inoculation of the isolate on leaves resulted in the onset of P. notoginseng, and the signs were consistent with those of P. notoginseng leaf spots in the forest (Figure 9f). The tissue separation method was used to isolate pathogenic fungi from the diseased remains of P. notoginseng and the surrounding diseased vegetation with signs similar to P. notoginseng leaf spots, as shown in Figure 10 and Figure 11. These results show that P. notoginseng leaf spot bacteria can be isolated from two organs: P. notoginseng and the highest isolation rate from diseased leaves was 26%, followed by stems at 10%. However, pathogenic bacteria could not be isolated from the roots and vegetation around diseased P. notoginseng. These findings suggest soil-resident fungi as a potential primary infection source, thriving under suitable environmental conditions and infecting P. notoginseng. P. notoginseng leaf emerges as the primary overwintering site for leaf spot pathogens, representing the primary infection source.
3.5.2. Investigation of Transmission Mode
At the peak of disease occurrence (October to November), microscopic examination of the slides with adsorbed pathogens revealed detectable pathogen conidia gathered via the spore catcher (Figure 12a), with observed mycelial germination (Figure 12b). Microscopic examination of a slide coated with Vaseline directly attached to the affected part of the leaf revealed numerous conidia (Figure 12d). These findings emphasize the conidium as the main transmission mode for P. notoginseng, predominantly disseminated through airborne means.
3.6. Sensitivity Testing of B. linicola to Different Fungicides
The antibacterial activities of tetramycin, copper abietate, α-terpineol, α-pinene, matrine WV, Trichoderma, and B. subtilis were determined in the laboratory. The inhibitory effects of each agent on pathogenic mycelia are shown in Table 5. The inhibitory rate of tetramycin on pathogen growth ranged from 38.56 to 74.67%, with an EC50 value of 0.60 mg/L. The inhibition rate of the growth of pathogens was 3.10–34.76%, and the EC50 value was 3,120.05 mg/L. The inhibitory rate of α-terpineol on the growth of pathogens ranged from 24.46% to 72.40%, and the EC50 value was 48.14 μL/mL. In this concentration range, the inhibition rate of pathogen mycelial growth ranged from 1.62 to 78.82%, and the EC50 value was 27,385 mg/L. The EC50 value of α-pinene was not calculated because α-pinene had no significant inhibitory effect on the growth of pathogens. In addition, the inhibitory rate of Trichoderma was 44.19–52.48%, and the inhibitory rate of Trichoderma was 78.45%, while the inhibitory rate of B. subtilis was 78.45%. The antibacterial effects of each agent and colony growth are shown in Figure 13, Figure 14, Figure 15, Figure 16, Figure 17, Figure 18 and Figure 19.
In conclusion, in laboratory toxicity testing, tetramycin, α-terpineol, matrine WV, B. subtilis, and T. atroviride demonstrated the most effective inhibition of P. notoginseng mycelium growth. T. harzianum and cupric abresinate showed inhibition effects, while α-pinene showed no inhibitory effect.
3.6.1. Field Trials
The field control effects of the eight tested agents with different concentrations on P. notoginseng leaf spot are shown in Table 6. Among these agents, T2-1 (20% copper abietate diluted 500 times) exhibited the highest control effect at 82.57%. Additionally, T7-1 (10 billion CFU/g B. subtilis 0.15g/m2), T6-2 (300 million CFU/g T. atroviride BH-10 preparation 0.15g/m2), and T1-1 (0.3% tetramycin 500-times dilution) demonstrated control effects of 72.56%, 71.3%, and 70.2%, respectively. The remaining control effects, ranked from high to low, are as follows: T1-2 (0.3% tetramycin 1,000× dilution) > T5-3 (matrine WV 900× dilution) > T5-1 (matrine WV 300× dilution) > T5-2 (matrine WV 600× dilution) > T2-3 (20% copper resinate 2,000× dilution ) > T4-1 (α-pinene 10,000× dilution ) > T1-3 (0.3 tetramycin 2,000× dilution ) > T3-1 (α-pinene 10,000× dilution ) > T2-2 (20% copper ropinate 1,000× dilution ) > T3-3 (α-pinene 50,000× dilution ) > T4-2 (α-pinene 2,500× dilution ) > T3-2 (α-terpineol 25,000× dilution ) > T6-1 (Trichoderma BY10 preparation 0.15 g/m2). Among these, the lowest control effect was observed for T6-1 (300 million/g CFU Trichoderma BY10 0.15g/m2) at 40.4%. No harm was observed in P. notoginseng during the test, and no negative impacts were observed at the tested doses.
The results showed that a 500× dilution of 20% copper resinate (T2-1) effectively controlled leaf spot in P. notoginseng across different concentrations of the tested chemicals. B. subtilis 0.15g/m2 (T7-1), T. atroviride strain BH-10 preparation 0.15 g/m2 (T6-2), and 0.3% tetramycin 500× dilution (T1-1) also exhibited favorable control effects.
4. Discussion
Boeremia, formerly referred to as Phoma. comprises mostly pathogenic species affecting various host plants, including fruits, vegetables, and medicinal plants. These pathogens induce rot, necrosis, wilt, and ulcers in fruits, leaves, stems, and roots, significantly impeding plant growth and development. Notably, B. exigua causes leaf spots in Brazilian sweet potatoes, peas, Dumasia villosa, and ginseng [30,31,32,33], while B. lilacis induces ash necrosis [34]. B. strasseri causes root rot in mint [35], B. exigua var. Pseudolilacis causes black rot in vegetable thistle [36], and B. linicola primarily affects P. notoginseng during its late growth stage. The weakened growth potential of P. notoginseng during this period predisposes it to pathogenic bacterial colonization, leading to disease. Under favorable conditions, such as wet soil, the incidence of P. notoginseng leaf spot disease can reach 80%, posing a severe threat to its cultivation and production. Growers must pay close attention to the disease and implement timely prevention and control measures to reduce economic losses.
Wet soil conditions significantly favor the growth and survival of B. exigua [37]. Malcolmson et al. studied the relationship between soil moisture in eastern Scotland and the distribution of potato gangrene caused by this bacterium. They found that the incidence of this disease was the highest in the northeast region with the highest humidity. Similar considerations apply to low-temperature conditions in northeast China [38]. This finding is consistent with the results of this study, which found that the organic P. notoginseng planting base in Lincang forest has a high altitude, abundant rainfall, perennial humidity above 90%, and annual temperature below 20℃. We speculated that temperature and humidity are key meteorological factors affecting the occurrence and prevalence of P. notoginseng leaf spots in forests. In this study, the correlation between temperature and humidity and the occurrence of diseases was further analyzed, and the results showed that under high humidity conditions, temperature markedly affected the occurrence of leaf spots in the forest and was the decisive factor. Disease occurrence is the primary basis for disease prevention and control. By analyzing disease occurrences in previous years, it becomes feasible to predict future outbreaks, enabling the implementation of timely forecasting, prevention, and control measures. In our study, we investigated the occurrence of leaf spot disease in P. notoginseng within forest environments. Our findings revealed that the disease typically initiates in mid-July, with October representing a pivotal period and November marking the peak occurrence. To effectively manage the disease, control measures should be implemented before the peak period. Thus, October 15 was identified as the optimal date for preventive and control interventions in this study.
Trichoderma and Bacillus are widely used biological control agents. For example, B. subtilis BS06 can effectively control soybean root rot caused by Fusarium oxysporum [39]. Another strain, B. subtilis WZZ-6, exhibited a high control rate of 72% against P. notoginseng root rot, achieving a remarkable 100% control effect in both in vitro tuber and plant assays [40]. Additionally, species such as T. koningiopsis, T. atroviride, and T. harzianum have demonstrated inhibitory effects on the growth of P. notoginseng root rot caused by F. Solani, Ilyonectria destructans, and Phytophthora cactorum. Field treatments with Trichoderma and Bacillus have been shown to increase P. notoginseng biomass [41]. Trichoderma and Bacillus control the occurrence of diseases mainly through nutrition and space competition, the production of broad-spectrum antibiotics, and enzymes [41,42,43]. B. subtilis JK-14 enhances the resistance of peaches by both inhibiting the growth of postharvest crop disease pathogens such as Alternaria, Tenuis, and Botrytis cinerea and activating defense-related enzymes to reduce disease occurrence [44]. B. subtilis M4 produces iturin to reduce the damage of postharvest gray mold in apples and Pythium ultimum in soybean seedlings. Furthermore, tetramycin, an antibiotic fungicide, induces the activity of defense-related enzymes to improve disease resistance in plants [45]. Moreover, purified cell wall-degrading enzymes such as chitinase and glucanase from different T. harzianum strains can effectively inhibit spore germination and mycelial growth of pathogens such as Fusarium, Alternaria, and Botrytis [46].
Tetramycin can effectively inhibit mycelial growth, spore germination, and bud tube elongation in Colletotrichum scovillei, a pepper anthracnose pathogen, exerting its antifungal activity by destroying the cell membrane structure [47]. It also has a significant inhibitory effect on P. notoginseng root rot [48]. Additionally, α-terpinol inhibits the mycelial growth and spore germination of Aspergillus niger and disrupts cell membrane permeability, thereby reducing pathogen infection [49]. WV demonstrates significant control over Fusarium wilt in tomatoes, leading to increased tomato yields and reduced activities of MDA and H2O2 in tomato leaves. Moreover, WV enhances disease resistance in tomato leaves by increasing the activities of catalase, peroxidase, and superoxide dismutase [50]. Additionally, α-pinene in Zanthoxylum oil inhibits the growth of F. sulphureum mycelia, disrupts cell membrane integrity, and inhibits spore germination, effectively controlling potato dry rot [51]. Copper abresinate, with its main active component being copper ions, alters membrane permeability and interferes with enzymatic reactions and respiratory activities, thus inhibiting spore germination [52]. According to the relevant provisions of the organic product standard GB/T 19630-2019 [53], this study selected plant protection products suitable for use in the production of organic plants to determine their bacteriostatic effectiveness indoors and verify field control effectiveness. B. subtilis, tetramycin, α-terpinol, WV, and Trichoderma exhibited superior inhibition effects on the mycelial growth of P. notoginseng leaf spot pathogens. The field control results showed that each agent and biocontrol strain had a certain control effect on leaf spot in P. notoginseng, but there were differences between the agents and biocontrol bacteria. Among them, the control effect of 20% copper abresinate at 500× dilution was significantly higher than that of other products, reaching 82.57%. Moreover, the control effects of B. subtilis 0.15g/m2, T. atroviride BH-10 preparation 0.15 g/m2, and 0.3% tetramycin 500× dilution were above 70%. These three biological control measures effectively reduced the occurrence and spread of P. notoginseng leaf spot disease. In summary, P. notoginseng leaf spot can be effectively controlled by adding 20% copper abietate 500× dilution, 10 billion CFU/g B. subtilis at 0.15g/m2, 300 million CFU/g T. atroviride BH-10 preparation at 0.15 g/m2, and 0.3% tetramycin 500× dilution.
5. Conclusions
In summary, leaf spot disease of P. notoginseng grown under forest canopies mainly affects the leaves of P. notoginseng. The disease begins in mid-July and peaks in November, with the highest incidence of 80% and an average disease index of 25. Temperature, particularly under conditions of high humidity, plays a decisive role in the occurrence of leaf spots in P. notoginseng within forest environments. Pathogenic bacteria overwintered and accumulated in diseased leaves and soil with conidia and conidiophores, becoming the primary source of infection in the field. Presently, leaf spot disease in P. notoginseng grown under forest canopies is most prevalent in Lincang and poses serious harm. The biological characteristics of the pathogen B. linicola include optimal mycelium growth temperature of 20°C and an optimal pH value of 5.0. Tetramycin exhibits inhibitory rates ranging from 38.56% to 74.67%, with an EC50 of 0.60 mg/L, while B. subtilis in matrine WV solution demonstrates inhibitory rates ranging from 1.62% to 78.82%, with a consistent rate of 78.45%. In the field test, the control effects of 20% copper abresinate 500× dilution, B. subtilis at 0.15g/m2, T. atroviride BH-10 at 0.15 g/m2, and 0.3% tetramycin 500× dilution were significant, at 82.57%, 72.56%, 71.3%, and 70.2%, respectively. These findings support the utilization of these treatments as environmentally friendly options for preventing and controlling leaf spot disease in P. notoginseng forests. The results of this study enrich our knowledge of the specific characteristics of B. linicola and provide an important scientific basis for the effective control and management of P. notoginseng leaf spot disease.
Author Contributions
Methodology, Yulin He and KuanYang.; formal analysis, Yulin He.; Writing—original draft, Yuxuan Wang.; Writing—review and editing, Liwei Guo.; Funding acquisition, Xiahong He and Liwei guo. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Young Talent Program of High-level Talent Plan in Yunnan Province (Grant No. YNQR-QNRC-2020-073), Yunnan Xingdian Talent Support Program, Yunnan Agricultural Joint Special Project – Key Projects (202301 BD070001-138), China Agriculture Research System of MOF and MARA (CARS-21), and Major Science and Technology Project of Yunnan and Kunming (202102AE090042, 202204BI090003, 202205 AF150018, and 2021JH002).
Data Availability Statement
Not applicable.
Acknowledgments
We would like to express our sincere appreciation to all teachers, students, and instrument platforms in our research group for their invaluable assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tang, G.Z. Information Analysis Based on the Development of the Panax notoginseng Industry [Master’s Thesis]; Yunnan University: Kunming, China, 2018. [Google Scholar]
- Wang, T.; Guo, R.X.; Zhou, G.H.; Zhou, X.D.; Kou, Z.Z.; Sui, F.; Li, C.; Tang, L.Y.; Wang, Z.J. Traditional uses, botany, phytochemistry, pharmacology and toxicology of Panax notoginseng (Burk.) F.H. Chen: A review. J. Ethnopharmacol. 2016, 188, 234–258. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B. Pharmacological activity of Sanchi ginseng (Panax notoginseng). J. Pharm. Pharmacol. 2006, 58, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.M.; Huang, L.Q.; Guo, L.P.; Liu, D.H. Chinese Sanqi industry status and development countermeasures. Zhongguo Zhong Yao Za Zhi. 2014, 39, 553–557. [Google Scholar] [PubMed]
- Zhan, P.; Wei, M.T.; Xu, B.; Han, R.L. Research on the development status and countermeasures of China’s Panax notoginseng industry based on the industrial chain. North. Hortic. 2021, 14, 158–164. [Google Scholar]
- Ren, Z.Y.; Wang, T.; Shen, M. The path of science and technology innovation in poverty alleviation in Lancang Lohu autonomous county of Yunnan Province: A case in science and technology poverty alleviation of academician Zhu Youyongs’Team. J. Yunnan Agric. Univ. (Soc. Sci.). 2019, 13, 14–19. [Google Scholar]
- Yu, Z.F.; Zhang, R.J. Development advantages and key problems of organic Panax notoginseng industry under forest in Pu’er city. Forest Inventory Plan. 2019, 44, 177–181. [Google Scholar]
- Li, J.B.; Ai, M.T.; Hou, J.; Zhu, P.Q.; Cui, X.M.; Yang, Q. Plant–pathogen interaction with root rot of Panax notoginseng as a model: Insight into pathogen pathogenesis, plant defence response and biological control. Mol. Plant Pathol. 2024, 25. [Google Scholar] [CrossRef]
- Wang, P.P. Correlation Between Root Rot with Rhizosphere and Rootmicroorganisms and Mitigation Measures of Panax notoginseng [Master’s Thesis]; Kunming University of Science and Technology: Kunming, China, 2022. [Google Scholar]
- Yang, K.; Wang, H.L.; Luo, L.F.; Zhu, S.S.; Huang, H.P.; Wei, Z.X.; Zhu, Y.Y.; Guo, L.W.; He, X.H. Effects of different soil moisture on the growth, quality, and root rot disease of organic Panax notoginseng cultivated under pine forests. J. Environ. Manage 2022, 329, 117069. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.T.; Li, X.; Li, Y.Y.; Cui, X.M.; Liu, D.Q. First report of a Panax notoginseng leaf disease caused by Alternaria alternata in China. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Li, J.B.; Bao, Y.L.; Wang, Z.R.; Yang, Q.; Cui, X.M. Research progress in diseases of Panax notoginseng. Physiol. Mol. Plant Pathol. 2022, 121. [Google Scholar] [CrossRef]
- Lan, C.Z.; Lin, X. Control effects of seven kinds of fungicides and rain-shelter cultivation on round spot of Panax notoginseng planted under forest. Mod. Agric. Sci. Technol. 2023, 831, 67–70. [Google Scholar]
- Yang, K.; Wang, H.L.; Ye, C.; Wang, Z.H.; Ye, K.H.; Zhang, S.; Huang, H.P.; Wei, Z.X.; Zhu, S.S.; Zhu, Y.Y.; et al. Infection characteristics and physical prevention strategy of Panax notoginseng Round spot disease caused by Mycocentrospora acerina. Plant Dis. 2022, 106, 2607–2617. [Google Scholar] [CrossRef]
- Yang, K.; Zhuang, S.; Guo, L.W.; Jiang, C.H.; Wang, Z.H.; Zhu, Y.Y.; He, X.H. Occurrence of Understory Planted Panax notoginseng Round Spot in Lancang County and its Control Effect of Rain-sheltered Cultivation. Journal of Chinese Medicinal Materials 2020, 43, 2857–2863. [Google Scholar] [CrossRef]
- Qian, N.; Cheng, Y.P.; Zhang, L.; Feng, C.L.; Zhang, G.Z.; Lu, X.L.; Zhao, W.S. First report of leaf spot disease caused by Boeremia linicola on Trifolium repens in China. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Guan, Y.M.; Zhang, S.N.; Ma, Y.Y.; Zhang, Y. Leaf spot caused by Boeremia linicola on Siberian ginseng in China. Plant Dis. 2020, 105. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, A.; Gilardi, G.; Franco Ortega, S.; Gullino, M.L. First report of a leaf spot caused by Boeremia Exigua var. linicola on autumn sage (Salvia greggii) in Italy and worldwide. Plant Dis. 2016, 100, 1777. [Google Scholar] [CrossRef]
- Lee, G.B.; Oh, T.H.; Ryu, J.T.; Kim, W. Stem and leaf rot of tomato caused by Boeremia linicola. Res. Plant Dis. 2022, 28, 108–111. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms. 2022, 10, 596–596. [Google Scholar] [CrossRef]
- Chen, J.L.; Liu, K.; Miao, C.P.; Sun, S.Z.; Chen, Y.W.; Xu, L.H.; Guan, H.L.; Zhao, L.X. Salt tolerance of endophytic Trichoderma koningiopsis YIM PH30002 and its volatile organic compounds (VOCs) allelopathic activity against phytopathogens associated with Panax notoginseng. Ann. Microbiol. 2016, 66, 981–990. [Google Scholar] [CrossRef]
- Wang, C.X.; Zhao, X.L.; Wu, K.; Liang, C.; Liu, J.H.; Yang, H.; Wang, C.M.; Yang, B.; Yin, F.; Zhang, W.D. Isolation and characterization of Bacillus velezensis strain B19 for biocontrol of Panax notoginseng root rot. Biol. Control. 2023, 185. [Google Scholar] [CrossRef]
- Radics, L.; Incze, M.; Dornberger, K.; Thrum, H. ; Tetramycin B, a new polyene macrolide antibiotic. Tetrahedron. 1982, 38, 183–189. [Google Scholar] [CrossRef]
- Shi, W.Q.; Xiang, L.B.; Yu, D.Z.; Gong, S.J.; Yang, L.J. Impact of the biofungicide tetramycin on the development of Fusarium Head Blight, grain yield and deoxynivalenol accumulation in wheat. World Mycotoxin J. 2020, 13, 235–246. [Google Scholar] [CrossRef]
- Li, W.Z.; Long, Y.H.; Yin, X.H.; Wang, W.Z.; Zhang, R.Q.; Mo, F.X.; Zhang, Z.Z.; Chen, T.T.; Chen, J.; Wang, B.C.; et al. Antifungal activity and mechanism of tetramycin against Alternaria alternata, the soft rot causing fungi in kiwifruit. Pestic. Biochem. Physiol. 2023, 192, 105409. [Google Scholar] [CrossRef] [PubMed]
- Tiilikkala, K.; Fagernäs, L.; Tiilikkala, J. History and use of wood pyrolysis liquids as biocide and plant protection product. Open Agric. 2010, 4, 111–118. [Google Scholar] [CrossRef]
- Li, X.G.; Han, L.H.; Wu, S.Q.; Piao, R.Z.; Liu, H.F. Inhibition effects of wood vinegar on Alternaria panax and Botrytis cinerea. Zhong Yao Cai. 2014, 37, 1525–1528. [Google Scholar] [PubMed]
- Yu, Y.; Liu, H.F.; Xia, H.R.; Chu, Z.H. Double- or triple-tiered protection: Prospects for the sustainable application of copper-based antimicrobial compounds for another fourteen decades. Int. J. Mol. Sci. 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Zhu, F.R.; Bin, R.P.; Liao, Y.M.; Wei, H.L.; Li, Y. Determination of pathogenicity of pathogenic bacteria in different parts of mulberry tree. Guangxi Seric. 2016, 53, 12–15. [Google Scholar]
- Colmán, A.; Lima, M.I.; Costa, H.; Barreto, R.W. Boeremia exigua causing leaf spots on sweet potato in Brazil. Australasian plant disease notes. J. Australas. Plant Pathol. Soc. 2020, 15, 403–414. [Google Scholar]
- Li, Y.P.; You, M.P.; Finnegan, P.M.; Khan, T.N.; Lanoiselet, V.; Eyres, N.; Barbetti, M.J. First report of black spot caused by Boeremia exigua var. exigua on field pea in Australia. Plant Dis. 2012, 96, 148. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, J.; Song, Q.; Yin, C.; Zhang, Z.; Tan, L.; Weng, Q. First report of leaf blight on Dumasia villosa caused by Boeremia exigua in China. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Wang, H.L.; Yang, K.; Huang, H.P.; Wei, Z.X.; Zhu, Y.Y.; Guo, L.W.; He, X.H. First report of leaf spot caused by Boeremia exigua on panax ginseng in Liaoning, China. Plant Dis. 2024, 108. [Google Scholar] [CrossRef]
- Kowalski, T.; Kraj, W.; Bednarz, B.; Rossa, R. The association of Boeremia lilacis with necrotic lesions on shoots and leaf petioles and Its pathogenicity towards Fraxinus excelsior. Eur. J. Plant Pathol. 2019, 154, 961–974. [Google Scholar] [CrossRef]
- Zimowska, B.; Król, E.D. Effect of selected preparations on growth and development Boeremia strasseri, the causal agent of black stem and rhizomes rot of peppermint (Mentha piperita). Acta Sci.Pol.Hortorum Cultus. 2018, 17, 3–12. [Google Scholar] [CrossRef]
- Koike, S.T.; Groenewald, J.Z.; Crous, P.W. First report of black rot caused by Boeremia exigua var. pseudolilacis on artichoke in California. Plant Dis. 2016, 100, 524–525. [Google Scholar] [CrossRef]
- Malcolmson, J.F. Some factors affecting the occurrence and development in potatoes of gangrene caused by Phoma Solanicola Prill & Delacr. Ann. Appl. Biol. 1958, 46, 639–650. [Google Scholar] [CrossRef]
- Malcolmson, J.F.; Gray, E.G. Factors affecting the occurrence of gangrene (Phoma exigua) in potatoes. Ann. Appl. Biol. 1968, 62, 77–87. [Google Scholar] [CrossRef]
- Jia, Y.J.; Huang, J.X.; Qi, L.L.; Zhang, X.L.; Liu, J.H.; Guan, H.L.; Wang, C.J.; Tang, G.M.; Dou, X.L.; Lu, M. Bacillus subtilis strain BS06 protects soybean roots from Fusarium oxysporum infection. FEMS Microbiol. Lett. 2021, 368, fnab102. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.C.; Cao, B.B.; Hu, M.; Zhang, S.S.; Li, J.; Pu, Q.; Li, O.; Zeng, G.H.; Hu, X.F. Screening and activity analysis of antagonistic bacteria against root rot on Panax notoginseng. J. Zhejiang Sci-Tech Universit. 2019, 41, 806–811. [Google Scholar]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35 (Supplement), 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Guo, H.L.; Zhang, K.X.; Zhao, M.Y.; Ruan, J.J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Front. Microbiol. 2023, 14, 1160551. [Google Scholar] [CrossRef]
- Elad, Y. Biological control of foliar pathogens by means of Trichoderma harzianum and potential modes of action. Crop Prot. 2000, 19, 709–714. [Google Scholar] [CrossRef]
- Zhang, S.W.; Zheng, Q.; Xu, B.L.; Liu, J. Identification of the fungal pathogens of postharvest disease on peach fruits and the control mechanisms of Bacillus subtilis JK-14. Toxins. 2019, 11, 322–322. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.J.; Zhao, X.H.; Zhang, Q.H.; Xu, C.; Zhu, H.L. Studies on the rice resistance to rice blast induced by tetramycin. J. Anhui Agric. Sci. 2010, 38, 6263–6264. [Google Scholar]
- Monte, E. Understanding Trichoderma: Between biotechnology and microbial ecology. Int. Microbiol. 2001, 4, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.Y.; He, L.F.; Li, X.X.; Lin, J.; Mu, W.; Liu, F. Toxicity and biochemical action of the antibiotic fungicide tetramycin on Colletotrichum scovillei. Pestic. Biochem. Physiol. 2018, 147, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Guo, L.W.; Li, J.Z.; Yang, K.; Wang, H.L.; Zhang, S.; Su, S.; Zhu, Y.Y.; He, X.H. Effects of Treating Seeds with Three Microbial agents on Growth and seedling Quality of Panax notoginseng. J. China Agric. Univ. 2022, 27, 189–198. [Google Scholar]
- An, P.P.; Yang, X.B.; Yu, J.; Qi, J.R.; Ren, X.Y.; Kong, Q.J. α-terpineol and terpene-4-ol, the critical components of tea tree oil, exert antifungal activities in vitro and in vivo against Aspergillus niger in grapes by inducing morphous damage and metabolic changes of fungus. Food Control. 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Zhou, H.Y.; Fu, K.J.; Shen, Y.; Li, R.H.; Su, Y.B.; Deng, Y.S.; Xia, Y.S.; Zhang, N.M. Physiological and biochemical mechanisms of wood vinegar-induced stress response against tomato fusarium wilt disease. Plants (Basel). 2024, 13. [Google Scholar] [CrossRef] [PubMed]
- Xing-dong, L.; Hua-li, X. Antifungal activity of the essential oil of Zanthoxylum bungeanum and its major constituent on Fusarium sulphureum and dry rot of potato tubers. Phytoparasitica. 2014, 42, 509–517. [Google Scholar] [CrossRef]
- Anna, T.L.; Valeria, I.; Federica, C. Copper in plant protection: Current situation and prospects. Phytopathologia Mediterr. 2018, 57, 201–236. [Google Scholar]
- GB/T 19630–2019Standardization Administration of China. Organic Products—Requirements for Production, Processing, Labeling and Management System, 2019.
Figure 1.
Methods of spore capture. (a) Using a spore trap device; (b) Glass slide natural collection.
Figure 1.
Methods of spore capture. (a) Using a spore trap device; (b) Glass slide natural collection.

Figure 2.
Signs of P. notoginseng leaf spot disease across different growth years. (a) Three-year-old P. notoginseng; (b) Two-year-old P. notoginseng; (c, d). Leaf spot signs.
Figure 2.
Signs of P. notoginseng leaf spot disease across different growth years. (a) Three-year-old P. notoginseng; (b) Two-year-old P. notoginseng; (c, d). Leaf spot signs.

Figure 3.
Occurrence of P. notoginseng leaf spot disease across various planting bases. XD: Bo Shang Dao River; LS: Bo Shang Liang Mountain; SYP: Bo Shang Shuang Ying Pan; DTZ: Lan Cang Da Tang Zi; DH: Lan Cang Dong Hui; XD: Xun Dian. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 3.
Occurrence of P. notoginseng leaf spot disease across various planting bases. XD: Bo Shang Dao River; LS: Bo Shang Liang Mountain; SYP: Bo Shang Shuang Ying Pan; DTZ: Lan Cang Da Tang Zi; DH: Lan Cang Dong Hui; XD: Xun Dian. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
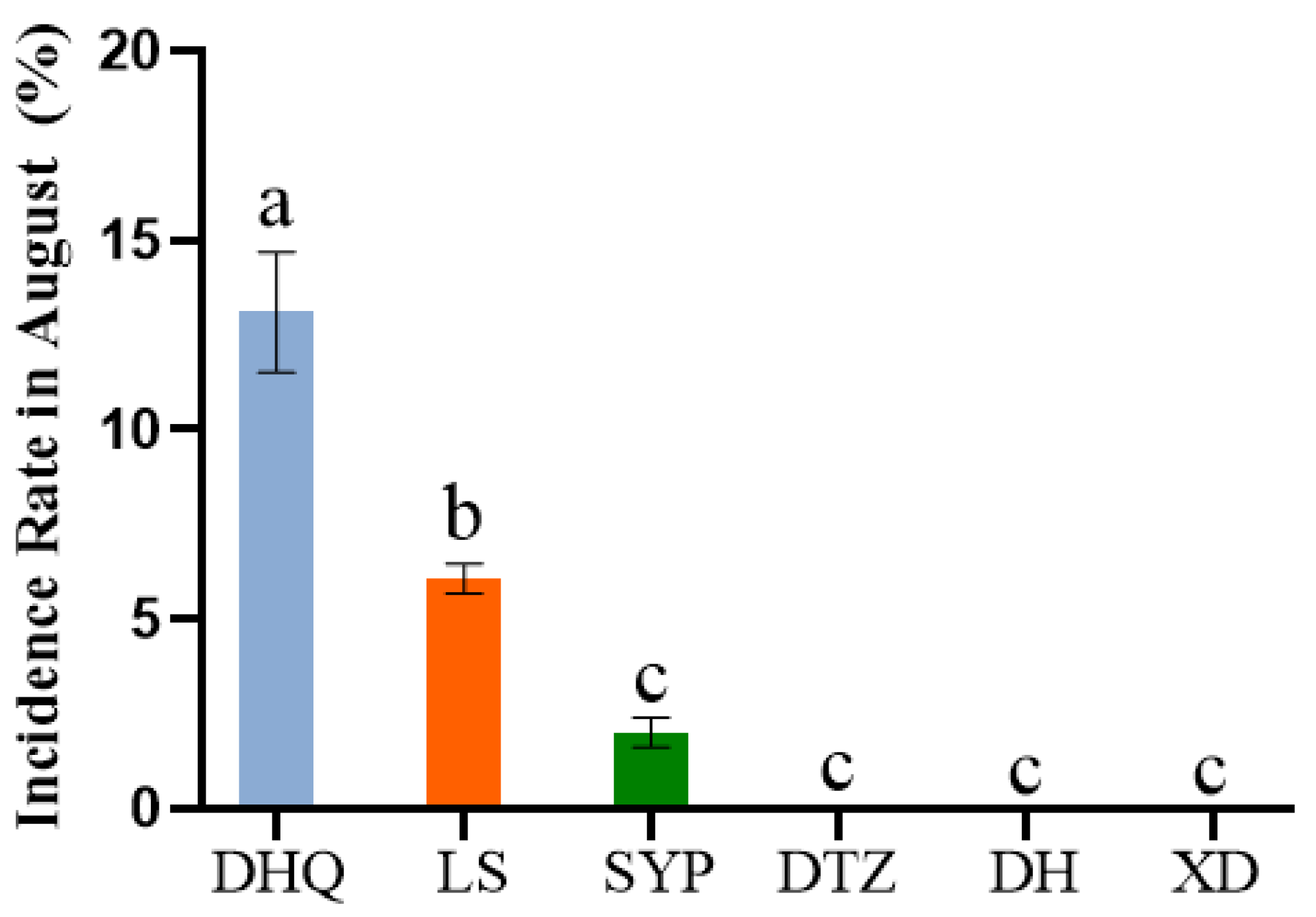
Figure 4.
Incidence of P. notoginseng leaf spot disease in forest environments. (a) Incidence rate; (b) Disease index. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 4.
Incidence of P. notoginseng leaf spot disease in forest environments. (a) Incidence rate; (b) Disease index. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
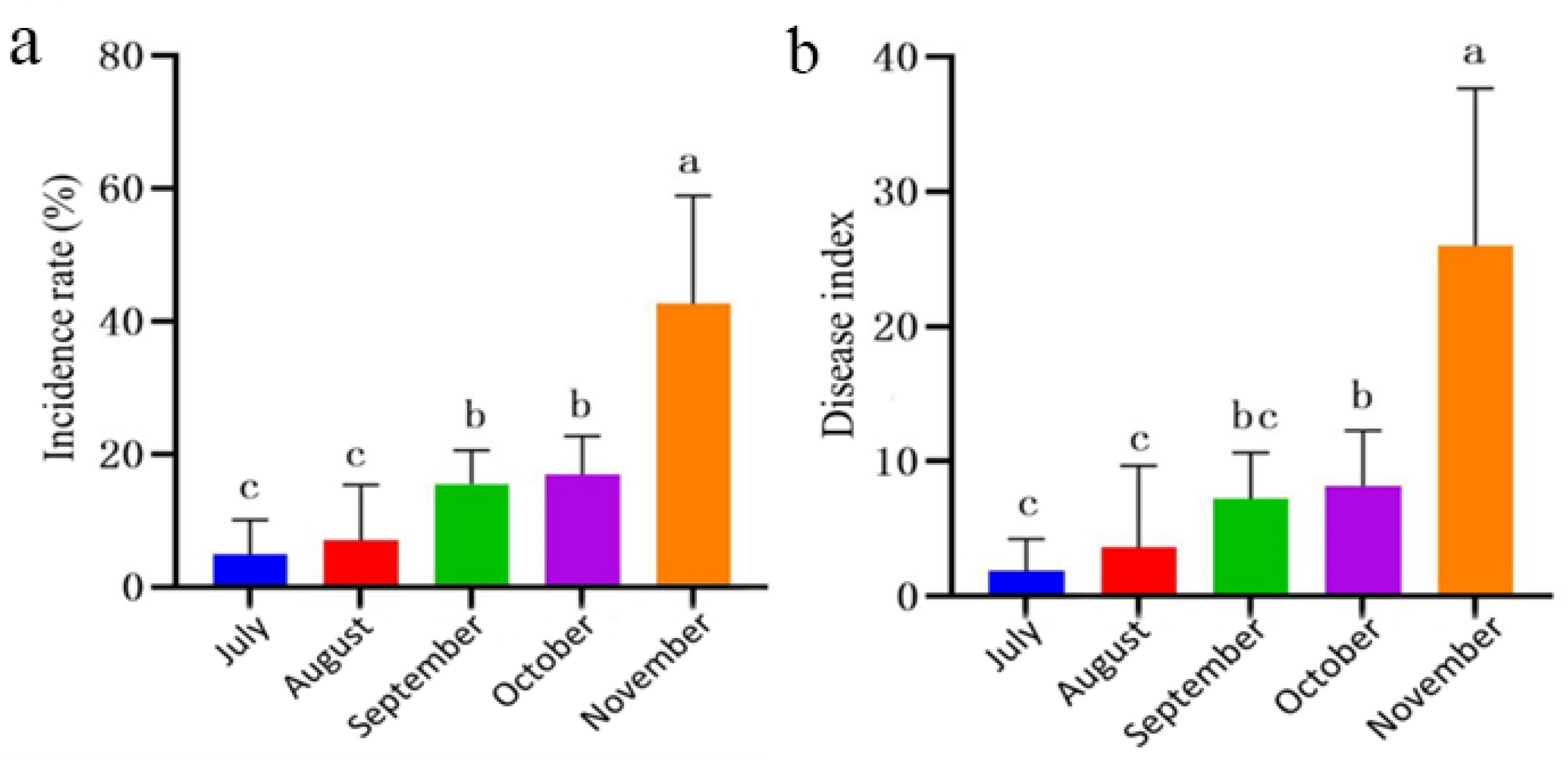
Figure 5.
The relationship between temperature, humidity, and disease index of P. notoginseng leaf spot.
Figure 5.
The relationship between temperature, humidity, and disease index of P. notoginseng leaf spot.
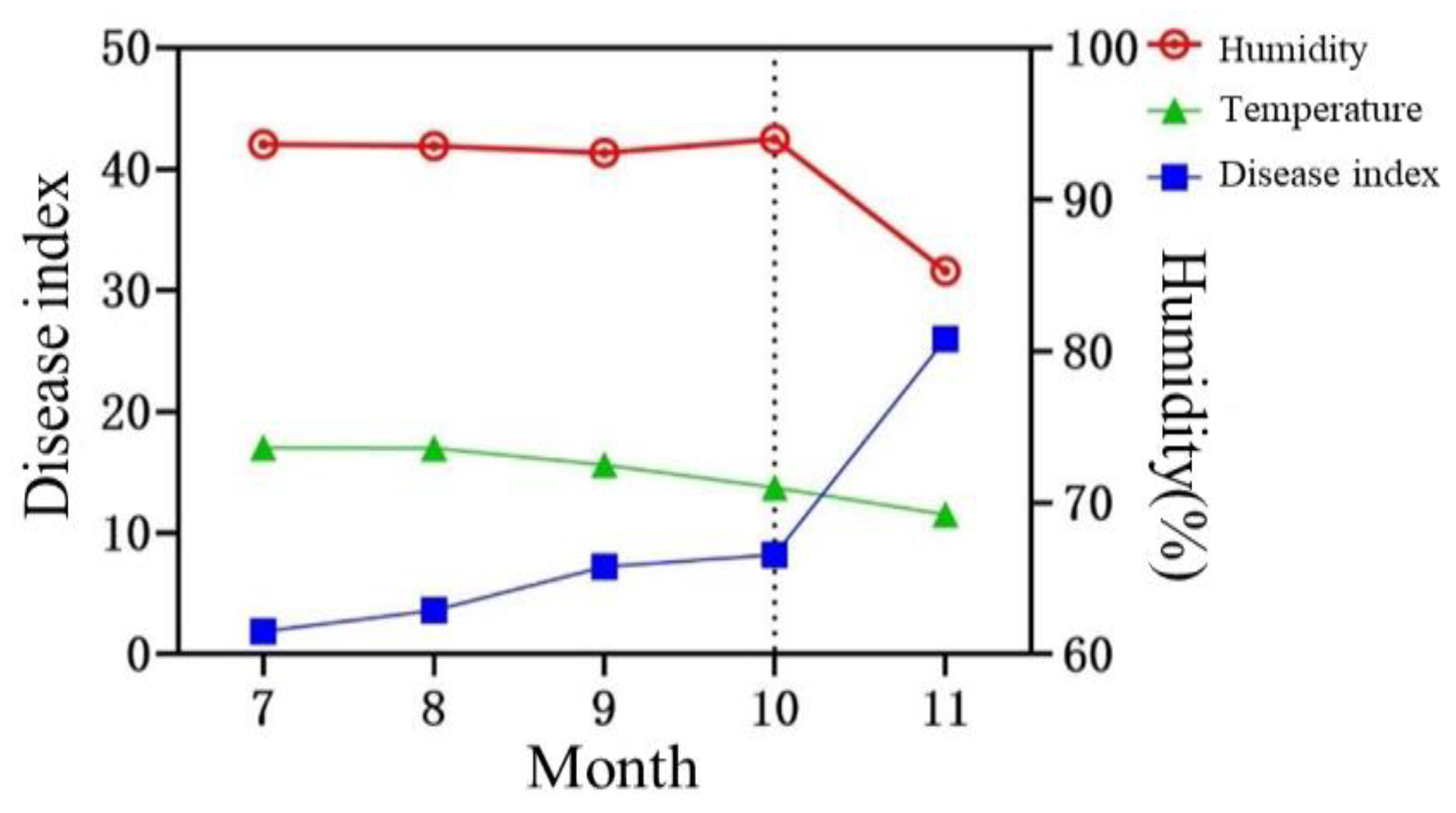
Figure 6.
Morphological features of LYB-2 grown on PDA medium for 7 d. (a) Top view of the medium; (b) back view of the medium; (c) mycelium; scale bar = 10 μm.(d, e) conidiophore; scale bar = 40 μm. (f) conidia; scale bar = 10 μm.
Figure 6.
Morphological features of LYB-2 grown on PDA medium for 7 d. (a) Top view of the medium; (b) back view of the medium; (c) mycelium; scale bar = 10 μm.(d, e) conidiophore; scale bar = 40 μm. (f) conidia; scale bar = 10 μm.
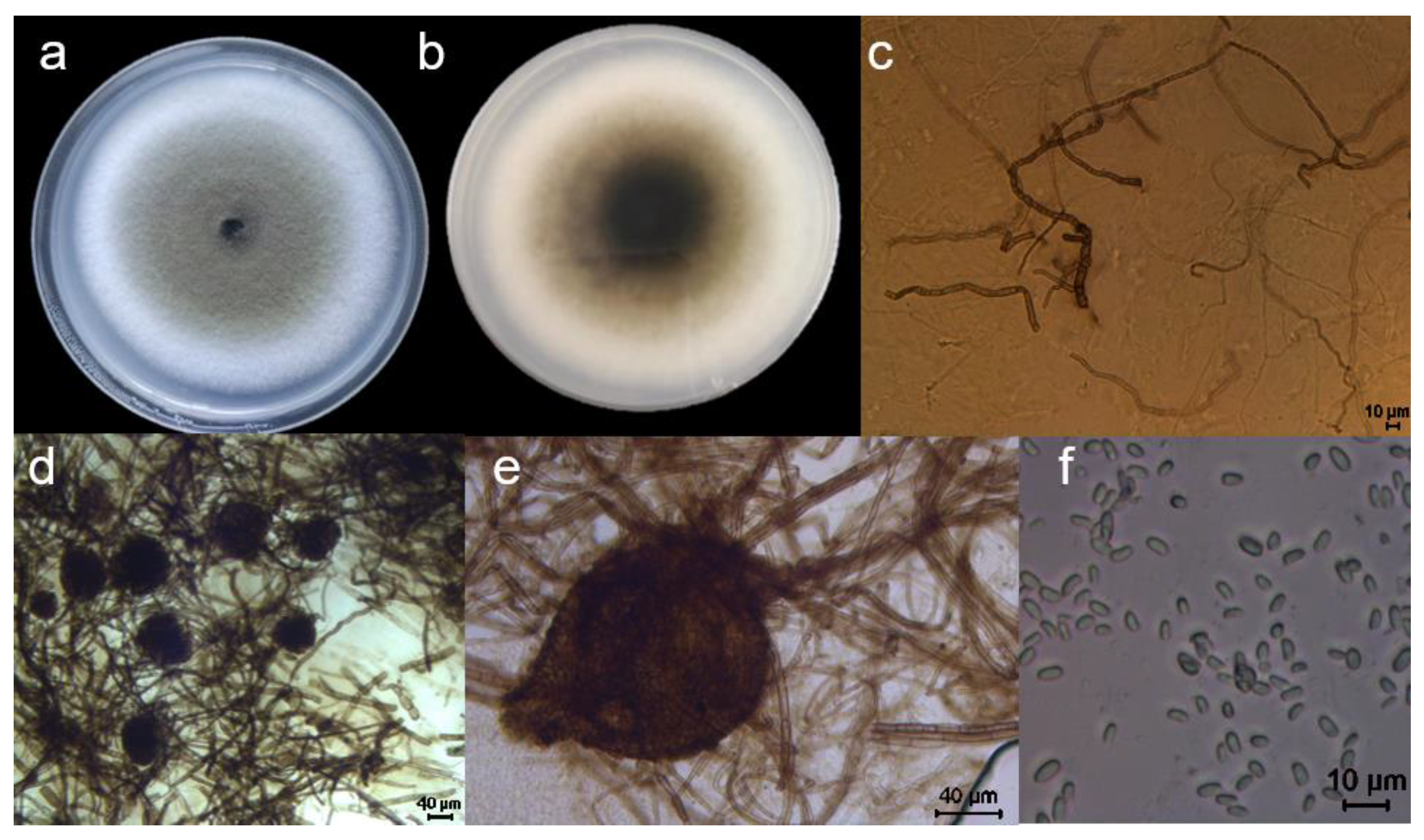
Figure 7.
Colony morphology and diameter of the representative strain LYB-2 at different temperatures. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 7.
Colony morphology and diameter of the representative strain LYB-2 at different temperatures. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
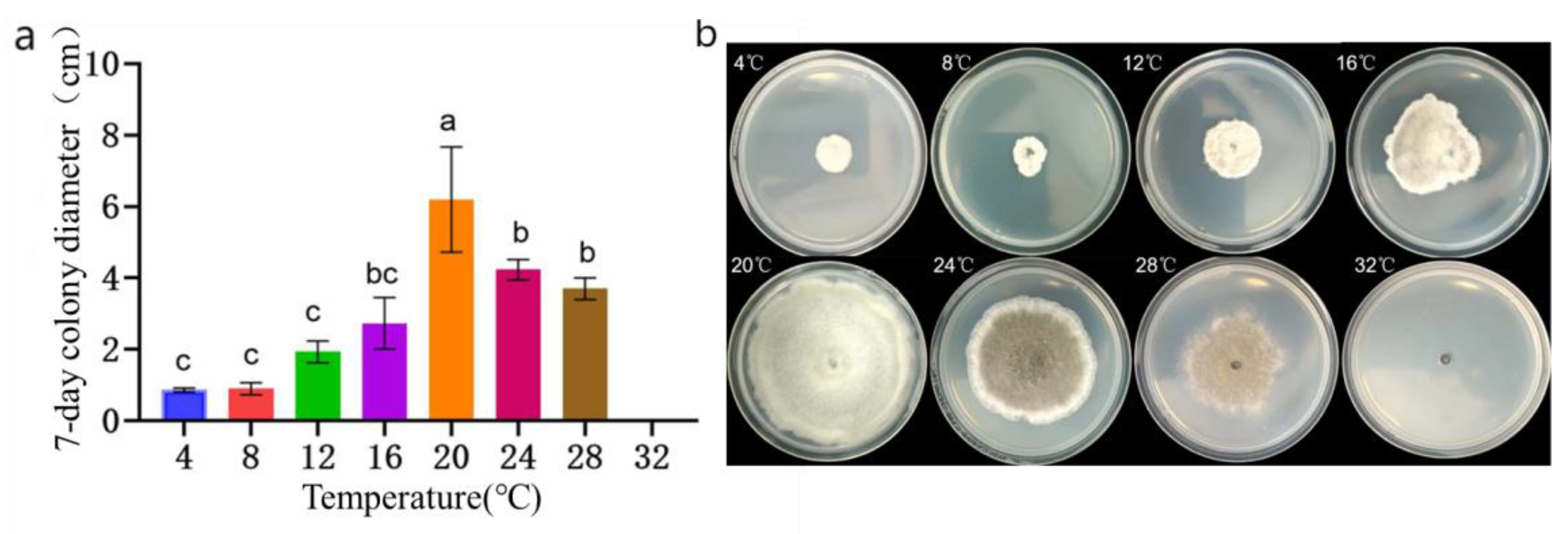
Figure 8.
Colony morphology, diameter of the representative strain LYB-2 at different pH values. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 8.
Colony morphology, diameter of the representative strain LYB-2 at different pH values. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
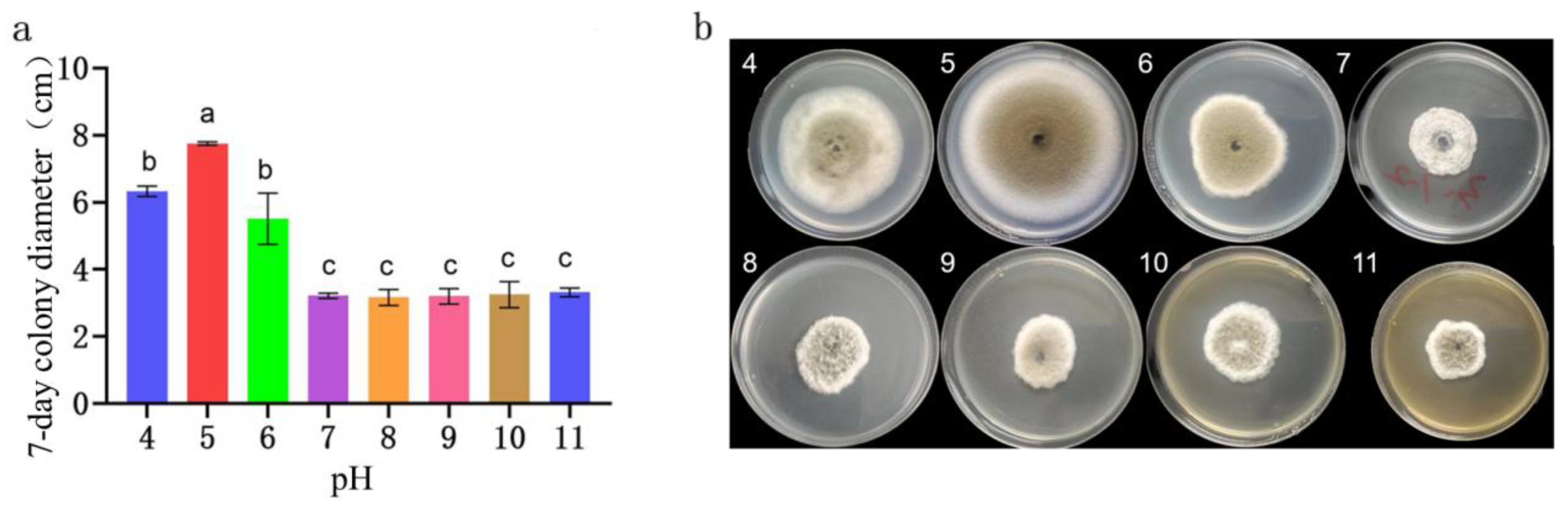
Figure 9.
Isolation of culturable fungi in the rhizosphere soil of diseased P. notoginseng. (a–d) Colonies formed at different dilution concentrations; (e) Purified pathogenic fungal colonies; (f) Pathogenicity determination of the isolate.
Figure 9.
Isolation of culturable fungi in the rhizosphere soil of diseased P. notoginseng. (a–d) Colonies formed at different dilution concentrations; (e) Purified pathogenic fungal colonies; (f) Pathogenicity determination of the isolate.

Figure 10.
Isolation rate of pathogenic fungi on different diseased tissues. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 10.
Isolation rate of pathogenic fungi on different diseased tissues. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
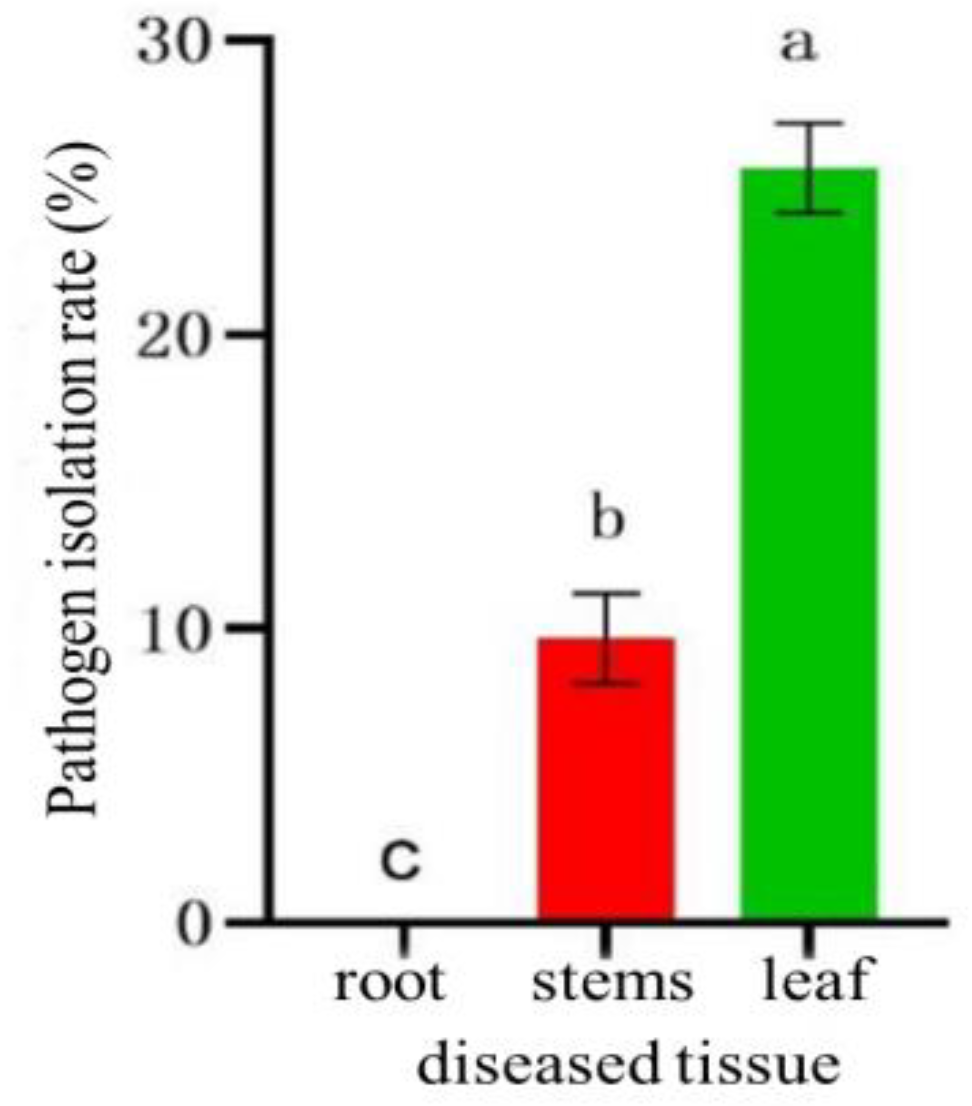
Figure 11.
Fungal isolation on different diseased plants. DY: Duoyi tree; QM: Spartina; PB1: loquat tree; SS: pine; ZJ: Eupatorium adenophorum; NN: Polygonum nepalense; JC: Jincai.
Figure 11.
Fungal isolation on different diseased plants. DY: Duoyi tree; QM: Spartina; PB1: loquat tree; SS: pine; ZJ: Eupatorium adenophorum; NN: Polygonum nepalense; JC: Jincai.
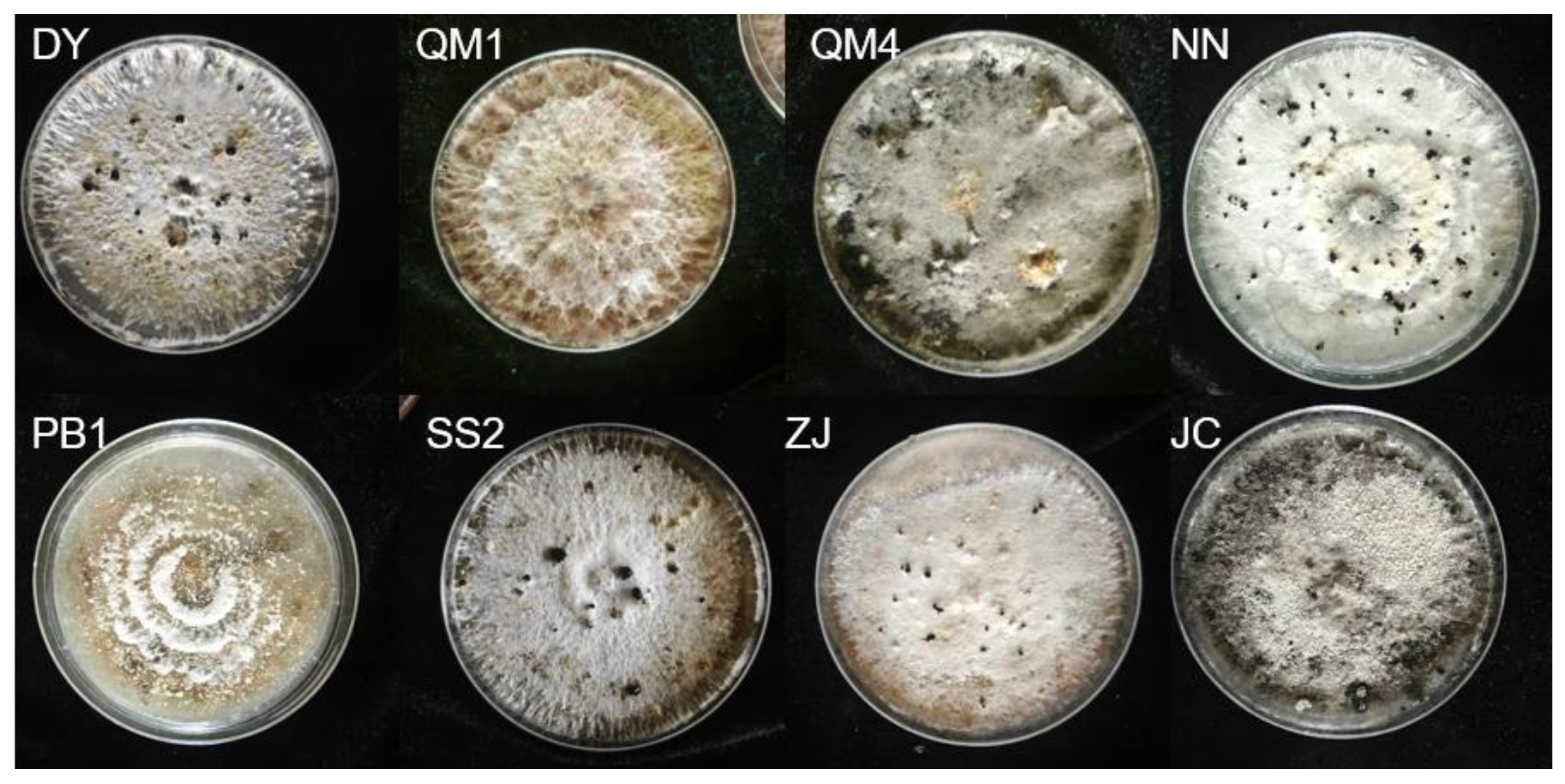
Figure 12.
Microscopic examination of sclerotium and conidia. (a, b) Conidiophore on slides, scale bar = 20 μm; (c) Conidiophore on the lesion, scale bar = 20 μm; (d) Conidia on slides, scale bar = 10 μm.
Figure 12.
Microscopic examination of sclerotium and conidia. (a, b) Conidiophore on slides, scale bar = 20 μm; (c) Conidiophore on the lesion, scale bar = 20 μm; (d) Conidia on slides, scale bar = 10 μm.

Figure 13.
Effect of tetramycin on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 13.
Effect of tetramycin on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
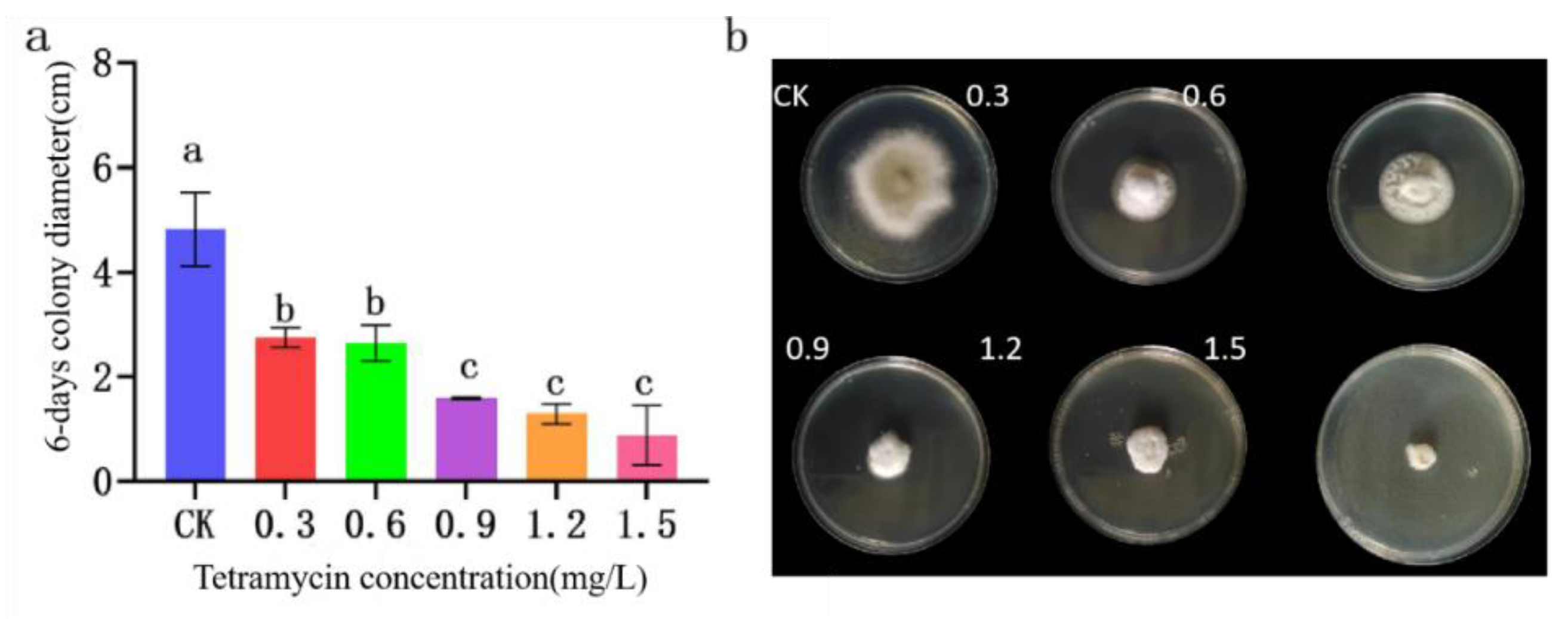
Figure 14.
Effect of α-terpineol on the colony growth of B. linicola LYB-2 (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 14.
Effect of α-terpineol on the colony growth of B. linicola LYB-2 (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
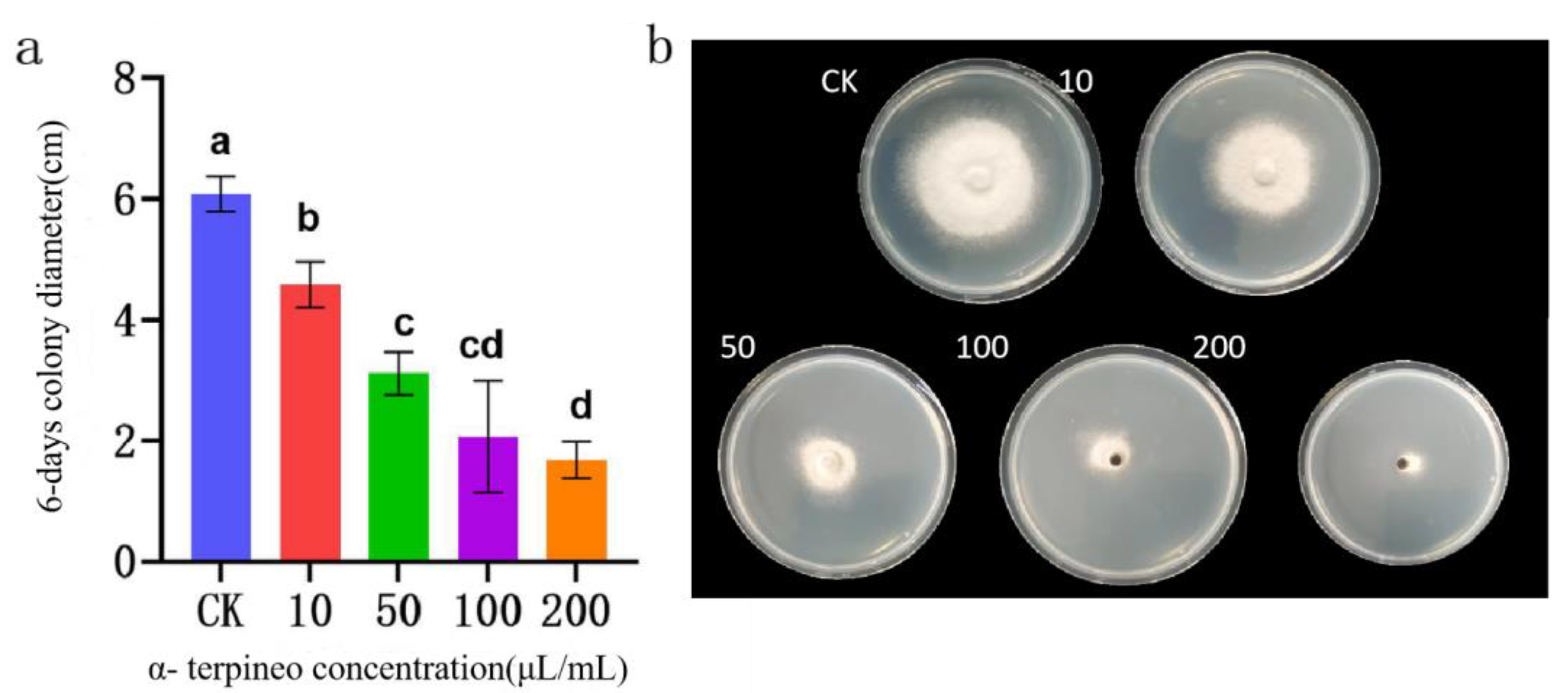
Figure 15.
Effect of α-pinene on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 15.
Effect of α-pinene on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
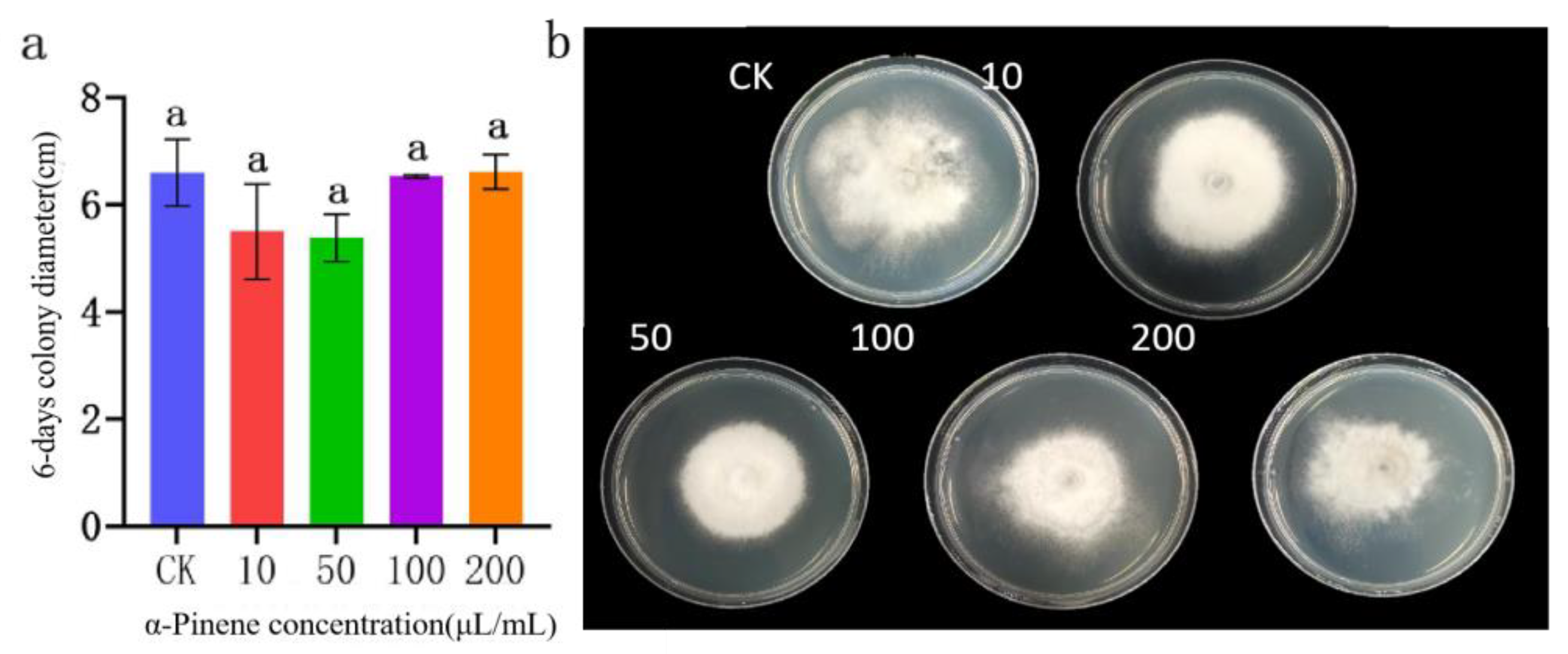
Figure 16.
Effect of copper abietate on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 16.
Effect of copper abietate on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
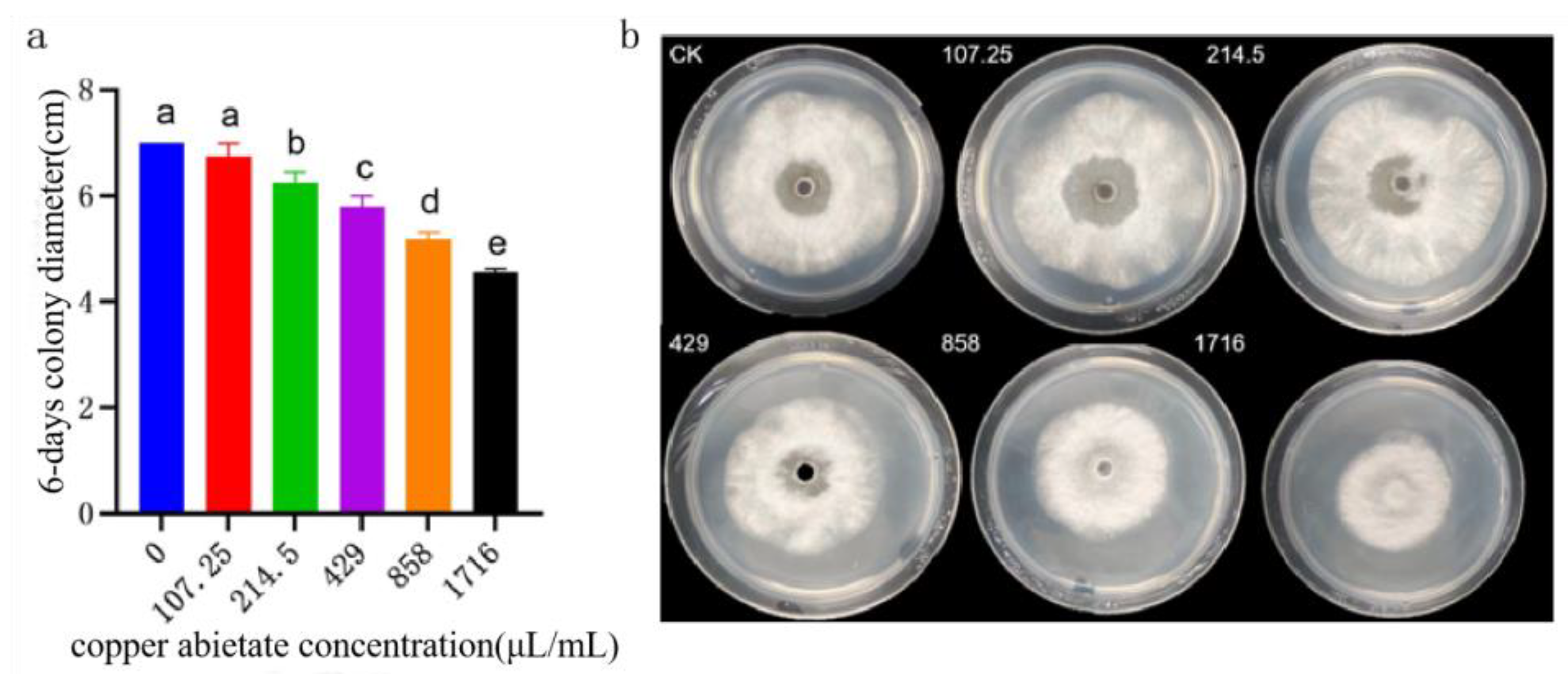
Figure 17.
Effect of WV on the colony growth of B. linicola LYB-2. (a) Colony diameter;(b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 17.
Effect of WV on the colony growth of B. linicola LYB-2. (a) Colony diameter;(b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
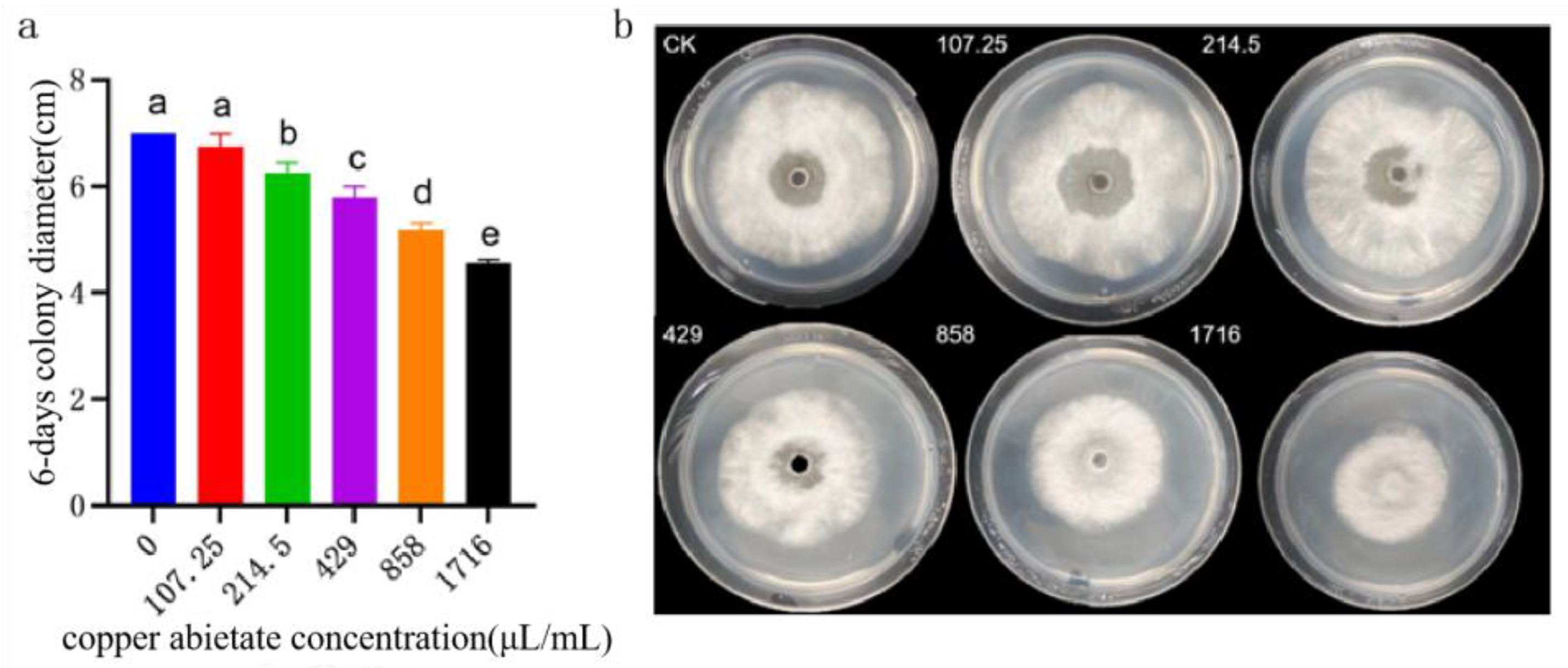
Figure 18.
Effect of Trichoderma on the colony growth of B. linicola LYB-2 under co-cultivation. (a) Colony diameter; (b) Colony morphology; M1: Trichoderma atroviride; M2: Trichoderma harzianum. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 18.
Effect of Trichoderma on the colony growth of B. linicola LYB-2 under co-cultivation. (a) Colony diameter; (b) Colony morphology; M1: Trichoderma atroviride; M2: Trichoderma harzianum. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
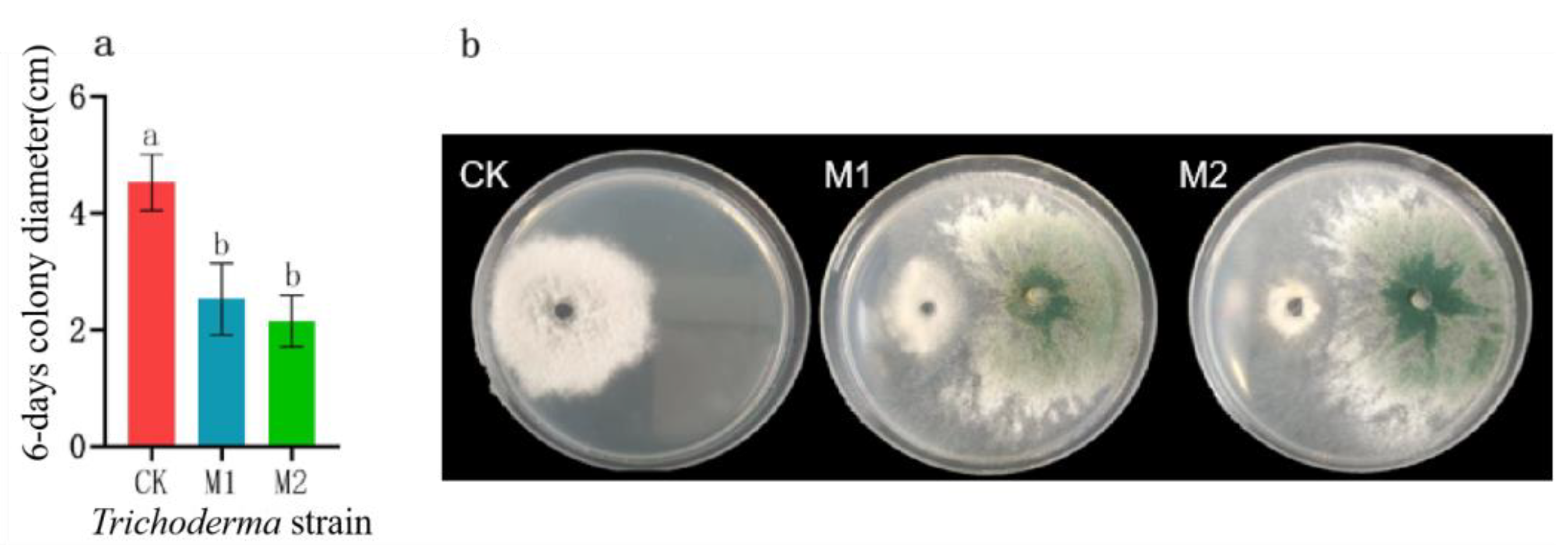
Figure 19.
Effect of copper abietate on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
Figure 19.
Effect of copper abietate on the colony growth of B. linicola LYB-2. (a) Colony diameter; (b) Colony morphology. Values of ±standard error are presented with error bars, and different letters indicate significant differences at p < 0.05.
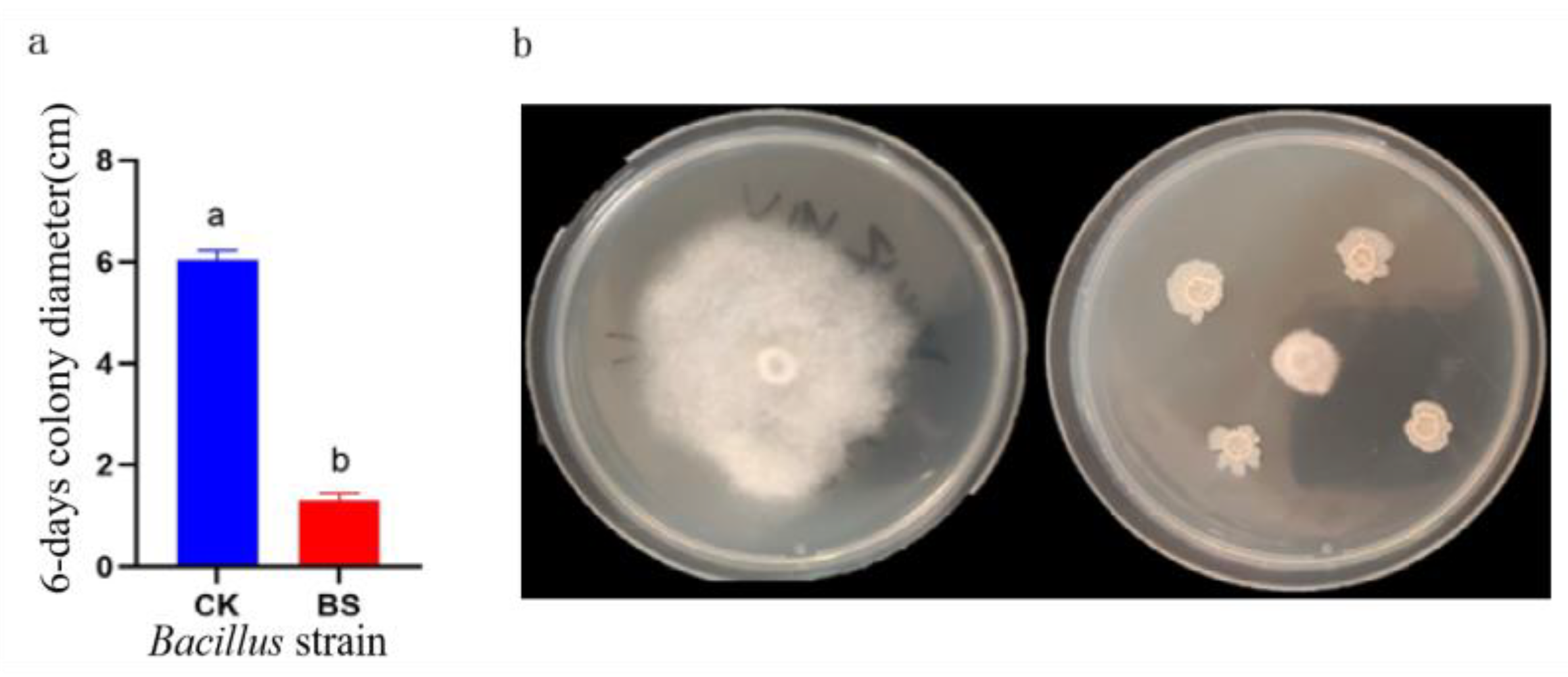

Table 1.
P. notoginseng leaf spot disease severity grading.
| Incidence level | Incidence degree | Representative value |
|---|---|---|
| 1 | No or virtually no spots | 0 |
| 2 | Spots are small, 1–2 spots per leaf, less than 5% of leaf area | 1 |
| 3 | More than 2 spots per leaf, 5–25% of leaf area | 2 |
| 4 | Spots cover 25–50% of leaf area, leaves do not fall off, or 1/3 of the stem is diseased | 3 |
| 5 | Spots occupy more than 50% of the leaf area, some or all of the leaves fall off, only the stem remains or the plant dies | 4 |
Table 2.
Concentration settings of the seven agents on the hyphal growth of P. notoginseng leaf spot pathogens.
Table 2.
Concentration settings of the seven agents on the hyphal growth of P. notoginseng leaf spot pathogens.
| Tested agent | Concentration gradient | Unit | ||||
| Tetramycin | 0.3 | 0.6 | 0.9 | 1.2 | 1.5 | mg/L |
| copper abietate | 107.25 | 214.5 | 429 | 858 | 1,716 | mg/L |
| α-terpineol | 10 | 50 | 100 | 200 | — | μL/mL |
| α-pinene | 10 | 50 | 100 | 200 | — | μL/mL |
| Sophora flavescens alt WV | 5,000 | 10,000 | 20,000 | 40,000 | — | mg/L |
Table 3.
Treatment concentration of tested agents.
| Treatment number | Test agent | Treatment concentration | Treatment method |
| TCK | Clear water control | — | Atomizing |
| T1-1 | 0.3% tetramycin | 500× | |
| T1-2 | 1,000× | ||
| T1-3 | 2,000× | ||
| T2-1 | 20% copper abietate | 500× | |
| T2-2 | 1,000× | ||
| T2-3 | 2,000× | ||
| T3-1 | α-terpineol | 10,000× | |
| T3-2 | 25,000× | ||
| T3-3 | 50,000× | ||
| T4-1 | α-pinene | 10,000× | |
| T4-2 | 25,000× | ||
| T4-3 | 50,000× | ||
| T5-1 | Sophora flavescens alt WV | 300× | |
| T5-2 | 600× | ||
| T5-3 | 900× | ||
| T6-1 | T. harzianum | 100 g/acres | |
| T6-2 | T. atroviride | 100 g/acres | |
| T7-1 | B. subtilis | 100 g/acres |
Table 4.
Correlation analysis between meteorological factors and the severity of P. notoginseng leaf spot disease.
Table 4.
Correlation analysis between meteorological factors and the severity of P. notoginseng leaf spot disease.
| Meteorological factor | Pearson correlation coefficient |
| Mean monthly temperature, ℃ | -0.93 |
| Monthly minimum temperature, ℃ | -0.89 |
| Monthly maximum temperature, ℃ | -0.31 |
| Average monthly relative humidity, % | -0.96 |
| Monthly minimum relative humidity, % | -0.97 |
| Maximum relative humidity in month, % | 0.70 |
Table 5.
Inhibitory effects of various agents on P. notoginseng leaf spot pathogens.
| Treatment | Inhibition rate range (%) | Virulence regression equation | EC50 | Correlation coefficient (R2) |
| Tetramycin | 38.56–74.67 | y=1.4x+5.31 | 0.60 (mg/L) | 0.9368 |
| copper abietate | 3.10–34.76 | y=1.17x+0.91 | 3,120.05 (mg/L) | 0.9766 |
| α-terpineol | 24.46–72.40 | y=1.02x+3.29 | 48.14 (μL /mL) | 0.9953 |
| WV | 1.62–78.82 | y=3.29x-9.95 | 27,385 (mg/L) | 0.9708 |
| α-pinene | 0–13.62 | — | — | — |
| T. atroviride | 52.48 | — | — | — |
| T. harzianum | 44.19 | — | — | — |
| B. subtilis | 78.45 | — | — | — |
Table 6.
The field control efficacy of eight fungicides against P. notoginseng leaf spot disease.
| Treatment | Test agent | Disease index before application | Disease index after application | Relative control efficiency |
| TCK | Clear water control | 0.57±0.12 | 2.9±0.35 | |
| T1-1 |
0.3% tetramycin |
2.45±0.41 | 3.92±1.03 | 70.2±3.24a |
| T1-2 | 0.7±0.12 | 0.89±0.16 | 69.95±9.6a | |
| T1-3 | 1.54±0.38 | 3.19±1.04 | 57.72±13.35a | |
| T2-1 | 20% copper rosinate | 1.94±0.25 | 2.65±0.48 | 82.57±5.42a |
| T2-2 | 1±0.26 | 1.47±0.09 | 55.05±27.7a | |
| T2-3 | 0.62±0.15 | 1.11±0.27 | 59.62±16.1a | |
| T3-1 | α-terpineol | 1.37±0.3 | 2.98±0.72 | 56.83±10.83a |
| T3-2 | 1.04±0.26 | 2.52±0.61 | 49.32±13.28a | |
| T3-3 | 1.27±0.32 | 2.43±0.19 | 51.92±22.66a | |
| T4-1 | α-pinene | 1.82±0.18 | 3.54±0.22 | 58.45±13.53a |
| T4-2 | 1.33±0.05 | 3.24±0.32 | 50.52±12.55a | |
| T4-3 | 1.89±0.57 | 3.59±0.61 | 51.74±19.62a | |
| T5-1 | WV | 1.16±0.14 | 1.95±0.31 | 65.06±9.12a |
| T5-2 | 1.38±0.34 | 2.35±0.47 | 64.68±6.86a | |
| T5-3 | 1.71±0.3 | 2.43±0.41 | 69.72±9.32a | |
| T6-1 | T. atroviride | 0.6±0.05 | 2.16±0.63 | 40.04±20.02a |
| T6-2 | T. harzianum | 0.56±0.06 | 0.75±0.16 | 71.3±9.41a |
| T7-1 | B. subtilis | 1.09±0.25 | 1.27±0.25 | 72.56±8.11a |
Note: T1-1: 0.3% Tetramycin 500× dilution; T1-2: 0.3% Tetramycin 1,000× dilution; T1-3: 0.3% Tetramycin 2,000× dilution; T2-1: 20% Copper abietate 500× dilution; T2-2: 20% Copper abietate 1,000× dilution; T2-3: 20% Copper abietate 2,000× dilution; T3-1: α-terpineol 10,000× dilution; T3-2: α-terpineol 25,000× dilution; T3-3: α-terpineol 50,000× dilution; T4-1: α-Pinene 10,000× dilution; T4-2: α-Pinene 25,000× dilution; T4-3: α-Pinene 50,000× dilution; T5-1: WV 300× dilution; T5-2: WV 600× dilution; T5-3: WV 900× dilution; T6-1: Trichoderma strain BY10 preparation 0.15g/m2; T6-1: Trichoderma strain BH-10 preparation 0.15g/m2; T7-1: B. subtilis 100 g/acre; TCK: Blank control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



