
Submitted:
28 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
This study focuses on solving rural problems, sustaining environmental conditions, pastures improvement, and reduction of CO2 concentration, suggesting inoculated vegetation types found in the tropics as a nature-based solution for degraded agroecosystems. Noninoculated plants are intensively used for revegetation of degraded and mined lands, AMF taxonomic and plant-fungal ecological studies. Among those grasses, Urochloa brizantha is a tropical forage, having vigorous deep roots, commonly used for arbuscular mycorrhizae (AMF) multiplication in glasshouse and nursery, developing infective propagules to be conserved in germplasm banks. Other grass species, such as Urochloa decumbens, is also used for arbuscular mycorrhizal multiplication in addition to its cultivation for guaranteeing the sustainable of livestock systems. In the present study, a site of 100 m2 was selected to take soil samples in a plain area, with U. brizantha pasture, and the soil chemical characterization, microcharcoal content and AMF spore identification, were evaluated 3+6showing high amount of potassium (K), high microcharcoal content and the occurrence of 11.32 spores of AMF, most of Glomeraceae, in the rhizosphere of U. brizantha. Due to its economic importance for sustainable agricultural production and other uses besides its environmental services, as important promoters of soil carbon on grazing lands, more detailed research is needed on the biotic interactions and inoculant production in of the studied grasses.
Keywords:
grass
; arbuscular mycorrhizae
; Urochloa
; soil carbon
; grasses
1. Introduction
Due to the increasing interest on grasslands for forage cultivation (source of meat and milk) to obtain a better food production besides reducing environmental unfavorable emissions of greenhouse gases, researchers look for decrease nutrient and fertilizers leaching and employ of pesticides, enhancing carbon sequestration, thus improving soil sustainability [1]. However, The deleterious effects of chemical inputs on the soil of monocultures are well studied [2]. Fortunately, the effect of manures, wastes, residues, compost, and biochar amendments to the soil systems is increasingly studied worldwide, as organic agroecosystems are more procured. Thus, experiments on the implementation of natural soil conditioners by agriculture are rapidly increasing, and organic or regenerative agriculture adopted the addition of natural residues on horticultural plants and crops. Furthermore, the compatibility among biofertilizers and soil conditioners is also more investigated to support sustainable agricultural systems and to deal with the effects of global and especially, climatic change. Thus, the mycorrhizal symbiosis, application of compost of selected wastes [3], phosphate solubilizing microorganisms, microbial inoculants, and biochar, are increasingly explored. Of particular interest for research are perennial grassland plots of Urochloa brizantha (Figure 1), a tropical forage (source of meat and milk), commonly used also for Arbuscular mycorrhizae fungi (AMF) multiplication, as this plant species associate with this type of fungi, which highly colonize its roots [4] developing infective propagules. Thus, Urochloa decumbens is also used for arbuscular mycorrhizae multiplication, in addition to its cultivation for ensuring the sustainability of livestock systems. Moreover, these grass species is increasingly used in no-tillage ecosystems as cover crops [5], such as in coffee trees [6]. Some grass species actively propagate benefic microsymbionts after their roots are colonized by the soil microbiota. This is the case of Uroclhoa (Brachiaria) species, which are largely cultivated in pastures and cover crops. Urochloa species. are frequently selected for intercropping to improve land use and agricultural yields due to their high residue production, and slow decomposition[7]. The occurrence and life cycle of relevant symbionts such as the AMF, in grasslands were previously investigated, especially in Brazil [4]. This grass is utilized in pest management and have supported scientific experiments (most involving comparisons between inoculated vs. non-inoculated plants regarding plant growth. Additionally, this grass is intensively used for revegetation of degraded and mined lands, AMF taxonomic y studies and plant - fungi ecological studies. Reports (various on AMF spore inoculation, and two influential works on AMF root colonization [4,8]. indicated differences between the AMF species symbionts of Urochloa species. However, due to the economic importance of grass species, and due to the lack of complete data on this topic, more detailed research is needed to better understand the plant interactions with the soil microbiota. In the present study, with the purpose of examining the AMF associated to grasses in experimental farms, the AMF community structure was studied in a plot of U. brizantha grassland. Regarding spore numbers in the soil/ substrate, high values were previously observed in U. brizantha inoculated with A. longula (850 spores in 50 mL substrate), followed by plants inoculated with Acaulospora colombiana (755 spores) and Acaulospora morrowiae (228 spores, after 120 days of cultivation). In this sense, Barbosa et al. [8] highlighted the great multiplication potential of A. longula in Urochloa, thus indicating potential prolific species to be included as inoculants. In the present study, we reviewed, and examined the literature on the conclusions following the AMF associated to Urochloa species as well as the AMF-based inoculants in the field, including their benefits for plant cultivation. This paper examines the state of the art of Urochloa grasses in agroecosystems, with focus on its ecology including reports on their AMF symbioses. We searched the databases Scopus, and Google Scholar, with special attention to the most recent articles. In particular, the U. brizantha grassland plot at the experimental farm of EPAMIG was used for soil and fungal analyses.
2. Materials and Methods
2.1. Study Site
The U. brizantha grassland plot at the experimental farm of EPAMIG (19°42’51.5”S 44°53’37.0”W) in Brazil, South America, (estimated age of cultivation ~25 years, chopped up once a year, (Figure 1) was used to study the AMF community structure. A site of 100 m2 in the farm was selected to take soil samples in a plain area, with U. brizantha pasture, and the samples were collected between February (rainy period) and November (after the dry period) 2015.
2.2. Soil Sampling and Chemical Characterization
- Soil samples (three replicate plots (plot size, 5 by 20 m, composed by non-inoculated plants), obtained in February (rainy period) and November 2015 (after the dry period) at the site, were used for determination of chemical soil parameters (Embrapa, 1997) and for AMF spore extraction. For physical and chemical soil analysis a composite sample from soil of each sampled point at each season was prepared. In total 9 soil samples from grassland ecosystem weighing about 0.5–1 kg each were transported to the laboratory using sealed plastic bags, For determination of chemical soil parameters (Embrapa, 1997).
2.3. AMF Spore Isolation and Identification
AMF spores occurring in the soil samples (100g soil) were extracted by wet sieving, according to standard methods by Gerdemann and Nicolson [9], were counted and identified using specific descriptions of AMF species. The old and decaying spores with missing clear features were also counted as an approach for viability, according to Varga et al.[10], and the results expressed as percentage of unviable AMF spores.
2.4. Microcharcoal Content
The number of microscopic (>500 µm) charcoal fragments (adapted from [11] , was measured on the same Petri dishes used for spore isolation at 400 × magnification using a grid, and results were expressed as number of fragments in the subsamples (25g soil).
3. Results
The U. brizantha grassland presented a Typic Acrustox soil (Soil taxonomy- USDA, 2022 ), (Table 2). The region’s soil type was classified as sandy clay loam. The studied soil showed exclusive characteristics: high amount of K and higher charcoal content than samples of cultivated sites in the region (Table 3), studied previously by Pagano et al. [12]. The occurrence of AMF, most of Glomeraceae, was detected. Compared to other reports, Claroideoglomus etunicatum was dominant among the isolated species of Glomeromycota (Table 4). This study showed the AMF community associated to grasses, as well as potential for AMF inoculant formulation. Different AMF species (Acaulospora longula and Acaulospora colombiana) were previously reported by Barbosa et al. [8] as associated to U. brizantha in greenhouse, also in Minas Gerais, Brazil Table 1 shows limited available reports on Urochloa species and their associated microbiomes in Brazil.
In the present study, the AMF community structure was studied in a grassland in Brazil, The AMF community structure in the U. brizantha experimental plot showed dominance of Glomeraceae, compared to other reports (Table 1) where Acaulospora and Gigaspora were inoculated and, thus, theoretically occurred, such as in the reports by Baptistella et al.[6,7]. In the present work, five genera of Glomeromycota, four species and four unidentified morphotypes were detected in the U. brizantha grassland. The U. brizantha grassland showed exclusive characteristics: lower amount of K (Figure 2) and higher microcharcoal content (Table 2) than samples of cultivated sites in the region (Table 3).
This study showed the AMF communities associated to selected grasses at field conditions, as well as the potential for AMF inoculant formulation. Different AMF species (Acaulospora longula and Acaulospora colombiana) were previously associated with U. brizantha by Barbosa et al. [8]. (Table 1). However, to understand the U. brizantha ecology it is necessary to analyze the associated AMF species, the time of symbiosis establishment, root colonization and spore multiplication under different abiotic and biotic conditions. The hypothesis of this study is that U. brizantha associate with diverse AMF species, which have different behaviors when colonizing this host plant. We examined the arbuscular mycorrhizae associated to this grass in an experimental farm. The community structure was studied in a U. brizantha grassland in Brazil (Figure 3a), which showed a greater diversity of AMF compared to existing previous reports. Moreover, about 50 % of unviable AMF spores was estimated. Four genera of Glomeromycota, four species and four unidentified morphotypes were detected (Table 3). The grass showed exclusive characteristics: higher amount of soil organic matter and higher macroaggregates and charcoal content than samples of cultivated sites investigated in the region by Pagano et. al. [12] Most studies were carried out in glasshouse (pot experiments); however, the present study is the first to show in detail the AMF community associated to U. brizantha experimental plot (field site) besides previous reports on root colonization [4,12], and to point out the potential for AMF inoculant formulation to improve sustainable grass cultivation at field conditions.
AMF spore number
In the present study, a mean of 1.32 spores of AMF, most of Glomeraceae, were retrieved in the rhizosphere of U. brizantha
Table 4. AMF species associated with Urochloa brizantha at the experimental plot in the farm, Minas Gerais, Brazil.
In the report by Kanno et al.[13], mycorrhizal and soil pH treatments significantly affected shoot, root, and total dry weights of the tested grass species. Though inoculation with AMF increased plant growth, the responses varied among the plant species (U. humidicola did not responded to inoculation). Shoot, root, and total dry plant weight increased as pH increased; however, between the mycorrhizal treatment and soil pH, there was a significant interaction for root dry weight and total dry weight. AMF increased root dry weight significantly only at low pH, and total dry weight only in the low and high pH treatments. In previous studies, roots were highly colonized when inoculated, compared to than non-inoculated ones [8] (Table 5).
4. Discussion
Most previous reports on Urochloa come from greenhouse experiments, indicating differences between AMF species associated to U. brizantha, A. colombiana, showing the greatest percents of AMF root colonization in a short cultivation time, as evaluated by (Barbosa et al.[8] who found the highest number of AMF spores of Acaulospora longula and A. colombiana, exhibiting the highest mycorrhizal root colonization after 76 days, confirming that the inoculation favored the root growth of U. brizantha in greenhouse. The need of more information for a better understanding of the symbiotic relationships between different AMF species associated to U. brizantha was earlier pointed out and thus, we hypothesized that this plant species associates with a diverse community of AMF. including Acaulosporaceae, Gigasporaceae and Paraglomeraceae representants, as found in the AMF inoculation experiments and reports mentioned above. However, more investigation is needed regarding effectiveness of indigenous inocula and to compare it with commercial inocula formulation. In the present field study, the results indicated at least four AMF species (Figure 3), which can be propagated on farm, establishing sustainable Urochloa cultures in tropical agroecosystems Also, Sorghum cultures(Figure 2b) cultivated in pots with U. brizantha grassland soils can be employed. As AMF species vary in their performances for the different plant species, it is needed to compile the knowledge based on these grasses to optimize their management. In the present study, the AMF species were most from Glomeraceae, which are designated as r-strategists, and are commonly found in anthropized agroecosystems; however, some studies showed the effective potential of Acaulospora. Thus, it is necessary to study the different AMF species ( such as the great variety of Acaulospora) and propagules during the cultivation time to better understand the symbiosis in Urochloa species and other promising tropical forages. The production of biological inputs on farm (rural properties) is also increasingly implemented. The production of biological inputs such as inoculants and products for biological pest management is guided by the Ministry of Agriculture, Brazilian Agropecuary Company (EMBRAPA) for monitoring and regulate the quality and safety of the tested products.
4.1. Urochloa Species for Cover Crops
Perennial grasses have been increasingly employed as cover crops, and intercrops, besides their use for forage, and they were also utilized in pest management and for supporting scientific experiments. Among the Urochloa species, U. humidicola and U. decumbens and U. ruziziensis were investigated [4], presenting percents of AMF root colonization in their roots of 55, 78, and 66%, respectively. These high values indicate the relevant role of these plant species benefiting soil health through the fungal network provided by AMF colonizing the roots, which benefits plants and protect the soil. Furthermore, AMF secret a protein (glomalin), which favors soil aggregation, improving soil quality. In this sense, only one report showed the soil glomalin content in U. brizantha, which, varied from 3.30 to 3.74 mg. g soil −1 when inoculated with G. margarita and A. longula, respectively [6]. The low glomalin content when inoculated with Gigaspora sp. is expected as this AMF species grows slowly, compared to other ones, presenting a characteristic K-strategy lifestyle. Due to the relevant role of cover crops and intercrops in increasing diversity and sustainability of crops, they are commonly used by several reasons: the residues left after cutting protect the soil and provide nutrients to the next crop or intercropped culture. Additionally, Urochloa species have vigorous, abundant deep roots, faced to the superficial and scarce roots of crops. These grass traits facilitate carbon sequestration, soil organic matter stabilization and nutrient cycling. Urochloa roots also improve soil physical characteristics and influence soil nutrient dynamics, reducing nutrient losses and enhancing their cycling, which is benefic for nutrient use efficiency in sustainable agriculture. Furthermore, these cover crops are commonly employed in coffee trees [6] and in many orchards. Thus, meliorating the U. brizantha culture via benefits from AMF and increasing research on the symbionts and related microbiota, will favor sustainable grassy ecosystems destinated to forage, which can also provide sustainable plant systems to reproduce AMF propagules. Therefore, U. brizantha has showed high root colonization (35 %) when inoculated with A. longula; however, low values(16 %) were reported when inoculated with other species, such as G. margarita (16%) or Paraglomus occultum (27 %). The low root colonization when inoculated with G. margarita may be related to the characteristic slow root colonizer of this AMF species. The cultivation of Urochloa in agroecosystems have multiple benefits, such as reduced risk of soil erosion, better soil structure, higher levels of soil organic matter and biological activity [7].
4.2. Urochloa Species for AMF Studies
Urochloa brizantha is a tropical forage, having vigorous deep roots, commonly used for arbuscular mycorrhizae (AMF) multiplication (AMF-multiplier plant) in glasshouse and nursery, developing infective propagules for conservation in germplasm banks and, thus, to be used in taxonomy studies of AMF, providing information on the interactions of different AMF for mycorrhizal colonization and spore multiplication to be used in AMF collections and experiments. At the same time, most propagules provide information on the interactions of different AMF for mycorrhizal colonization and spore multiplication to be used in AMF collections. As AMF species used in the studies are identified by a deposit code in the AMF collection of each University or Research Institute, in Brazil. These AMF have unique value for inoculum to be applied in experimental research or in farms. Other grass species, such as U. decumbens, is also used for arbuscular mycorrhizae multiplication. Traditionally, a mix of dominant AMF species recruited from the local site can be employed. However, it was common to produce inoculum combining two different AMF species ( G. margarita and Glomus sp.). In other studies in greenhouse, Pedroso et al.[14] applied a mixture of spores (Glomus macrocarpum, P. occultum and Glomus sp. to inoculate U. brizantha in soil contaminated with heavy metals. They found greater root colonization in inoculated plants, showing beneficial effects of AMF on plant growth and alleviation of soil contaminants. As obligatory biotrophic, the multiplication of AMF is completed by axenic in vitro cultivation using transformed roots, or by its classical form, in cultivated trap plants according to Selvakumar et al. [15]. In vitro cultivation can enable the multiplication of cleaned (non-contaminated) spores in a shorter interval, being applicable for multiplication of fast-growing AMF species, such as those of Glomeraceae (mainly Glomus intraradices). It is also common to establish this symbiosis using G. margarita, isolated from introduced or autochthonous G. margarita isolates. For instance, AMF inoculum of G. margarita can be produced against species deposited in Glomeromycotan Germoplasm Banks, at the Universities, Institutes, or also in farms, isolated from introduced or autochthonous G. margarita spores, which can be multiplied in plastic vats containing ~20 kg of substrate( generally, a mixture of soil and sand 1:1 (v:v), having U. decumbens as host plant is utilized. During the period of growth, the plants are fertilized with nutritive solution (50 % P), but then, only water is used. Five months after planting, for drying the plants and substrate, irrigation is finished. Subsequently, the substrate is removed and sieved, and roots removed from soil, cut into fragments, as inoculum of more effective species for plants of economic interest. Thus, the knowledge of interactions between macro and microsymbionts, especially those positive, contributes to the introduction and targeted management of fungi in agroecosystems, selecting populations of more effective species for plants of economic interest. A great number of studies have utilized AMF spores multiplied using U. decumbens for propagation of spores used as inoculum.. Early reports by Kanno et al. [13] showed indigenous arbuscular mycorrhizas for 1 L inoculum suspensions, obtained against pasture soils, containing Gigaspora spp., Scutellospora spp., Glomus spp., Acaulospora spp. and Entrophospora spp., which was formulated based on microscopic observation. The grasses tested were Brachiaria brizantha cv. Marandu, B. decumbens cv. Basilisk , B. humidicola cv. Humidicola and Panicum maximum cv. Tanzania. That inoculum was used to inoculate the grasses in a glasshouse pot experiment where the percentage of root colonization by AMF per pot for each grass species and soil pH treatment (High, medium, low)was estimated. Average percentage of root colonization by AMF varied between 33% and 52% in the inoculated treatments (Table 4), contrasting to the uninoculated plants, which presented less percent ( 5.6 –15.6%) of roots colonized by AMF. To compare the advantages of non-AMF- and AMF-inoculated plants is a common practice for inoculum production. In this sense, most studies testing inoculants were achieved in greenhouse, microcosm, or nursery, employing sterilized soils or substrates. Among the reported AMF species belonging to the genus Gigaspora, G. margarita was initially one of the most utilized species, maybe due to its easily to distinguish characteristics and great spore size. Therefore, there are commonly used species, which can be included in farm inoculation of native AMF at field to improve grass cultivation.
Regarding Ph values for soil treatments, early reports firstly stated that AMF improved the growth and P uptake of U. decumbens at low and high pH levels, but not at an intermediate level. (5.7), concluding thus that the significance of AMF differs with values of soil pH and not only with the grass species.
4.3. Urochloa Species for Carbon Mitigation
Besides the contribution of U. brizantha plants fixing atmospheric CO2, the accumulation of glomalin (a glycoprotein produced by AMF hyphae and spores) in the soil is important due to its role protecting organic carbon, which contributes to the reduction of CO2 emissions. Thus, improving the growth of Urochloa plants results in a greater photosynthetic activity (increased fixation of atmospheric CO2), favoring the accumulation of organic carbon by roots and increasing the nutrient cycling in the soil/plant system. Glomalin, a glycoprotein found in the soil, produced on the surface the extra-radical hyphae (mycelium), intra-radical hyphae on roots in the rhizosphere, and by degradation of the AMF hyphae and spores, constitutes a stable form of carbon storage in the soil, according to Rillig and Lehmann et al. [17,18]. Thus, higher AMF hyphal length increases glomalin content, which contains 30-40% stable C, persistent in soils ( as evaluated by Hossain et al.[19]. In this sense, it is known that low glomalin content (0.007 mg per gram soil) was reported for Antarctic region, and higher contents (13.50 mg per gram soil) were common in tropical rainforest. On the other hand, in agricultural soils, glomalin content can vary between 0.30 and 0.70 favoring the accumulation of organic carbon by roots and increasing the nutrients flow and cycling in the plant-soil system. Therefore, The AMF increase plant growth, foliar biomass, and photosynthetic rate, and thus, fixation of atmospheric carbon dioxide (CO2), favoring the accumulation of organic carbon by roots, and the nutrient flow in the plant-soil system, as pointed by Wang et al. [20] and Lehmann et al. [18]. Furthermore, regarding carbon mitigation potential of tropical croplands, it is known that grazing lands occupy ~3.6 billion ha, accounting for one-fourth of possible C sequestration of worldwide soils. Based on global estimates, they can remove ~20% of the (CO2) released annually into the earth’s atmosphere from global deforestation and land-use changes. Atmospheric CO2 is incorporated by grazing lands through photosynthetic assimilation by grasses, and C sequestration by way of soil organic carbon (SOC) Therefore, grazing lands provide environmental benefits besides their economic returns. Among the environmental benefits provided by grasses, the maintenance of surrounding soil and water resources, air quality, and habitat for human and wildlife are mentioned. Thus, grasslands contribute to the economic well-being, exchange of products derived from them at local, regional, or national scales. Rates of SOC sequestration vary with climate, soil, and management. In this sense, several examples have showed the SOC sequestration attained by public policy efforts, as informed by Follett and Reed [21]. Additionally, it is known that 80% of the Total C saved in terrestrial ecosystems (approximately 3170 gigatons) is found in soils [22]. Soil respiration is one of the largest C fluxes from terrestrial ecosystems to atmosphere, as pointed by Luo and Zhou [23], minor changes in soil respiration can affect the C budget of soil. (as reported by Zhao et al. [24] . Soil carbon sequestration (process in which CO2 is removed from the atmosphere and stored in the soil, primarily mediated by plants, with carbon stored in the form of SOC, as pointed by (Lal [22] and (Ontl et al. [25]. Moreover, the tropics often have intermediate SOC levels due to high rates of primary productivity and decomposition due to the hot climate and profuse precipitations. It is known that promoting soil C management on grazing lands is recommended for reduction of emissions in compensation emission strategies. Soil carbon management has multiple societal benefits, including greenhouse gases (GHG) emission reduction in climate change mitigation programs. It is also known that grazing can affect the root C allocation between the synthesis of new tissue and the C loss caused by root exudation and respiration, as pointed by Wilson et al.[26]. Grazing changes the physicochemical properties of soil ( SOC mineralization, composition, and activity of microbial communities, according to Qu et al. [27], and by this means affects the C dynamics of soils, as verified by Hafner et al. [28] due to manure and urine released by livestock, as related by Vandandorj et al. [29].
4.4. Urochloa Species for Forage
this perennial monocotyledon is the principal source of diet for ruminant cattle in tropical and subtropical areas worldwide, such as the Americas, Africa, but especially in some countries of South America, such as Brazil, where animals are raised on pastures established with four significant Urochloa species (Urochloa brizantha, Urochloa decumbens, Urochloa humidicola, and Urochloa ruziziensis). Due to their economic relevance, these plant species were deeply studied, as Brazilian livestock production is concentrated in natural pastures› degraded planted pastures› pastures planted in good conditions [30,31]. Regarding plant differentiation, there were recognized two complexes of productive forages: 1)the brizantha complex (U. brizantha, U. decumbens and U. ruziziensis), and 2) the humidicola complex ( U. humidicola , Urochloa dictyoneura. Also, Urochloa maxima was previously transferred to Megathyrsus and Panicum, as reviewed by Tomaszewska [32]. Among tropical forages, Urochloa and Megathyrsus (syn. Panicum) present production potential in the tropical region of Brazil., due to their facility to adapt to different soil types and soil fertility, besides growing at different climatic situations, thus, Urochloa is one of the most planted grasses in Brazil, which livestock production is based on the use of pastures, ( ~ 158,622,704 hectares, ( 46,847,430 hectares of natural pastures, 11,821,006 of degraded planted pastures, and 99,954,268 hectares of pasture planted in great condition). Pastures of U. brizantha cv. Marandu are the most utilized forage in Brazil, responsible for ~ 40% of the sales of forage seeds cultivated in more than 70 million hectares [31], as verified by da Silva. [20], The Cerrado, a Brazilian savanna, which covers ~ 2 M km2 in the center of the country, is an important area for livestock production in the world. Since the 1970s, an estimated 0.5 M km2 has been occupied by tropical grasses of African origin such as Urochloa (Brachiaria) or Panicum maximum [32]. However, since the soils of the Brazilian savannas are highly acid, and low in phosphorus (P), the productivity and sustainability of the improved pastures are mainly affected by the ability of the established grass to extract P from the soil [33].
4.5. Urochloa for Revegetation
Urochloa is commonly used in revegetation and reclamation of degraded, contaminated, and mined soils. Early works have showed that revegetated mined soils using Brachiaria (Urochloa), and N2 fixing plants such as Mucuna and Crotalaria, commonly present in the seed cocktails used by mining companies, was a frequent approach. In riparian zones, Novais et al. [34]. used the practice of spread out a mixture of seeds rich in nitrogen-fixing plant species with sand and composted cotton, resulting in a rapid establishment of native plants plot. like the control reference site (Cerrado vegetation) thus, fortunately, U. brizantha was not dominant, and the low soil organic carbon indicated low plant diversity, showing that the competition caused by U. brizantha can affect the ecological diversity in areas experiencing ecological succession. Urochloa humidicola, Panicum maximum, and U. brizantha were tested in an experiment carried out throughout more than 10 years in a coal mining area in Candiota, Southern Brazil. Those species were compared to natural soil under native vegetation, which is considered a positive control or reference by Miguel et al. [35]. After that time those grasses improved soil physical conditions up to 10 cm depth. Moreover, as these sites are impacted by machine traffic events in the topsoil layer, they found an increase in the percentage of soil macroaggregates from zero traffic decreasing to twelve events of movement. Also, simultaneous reduction in the percentage of soil microaggregates was observed. In Previous studies of revegetation with perennial grasses also in Brazil (8.6-years) roots enhanced minesoil structure at 10 cm depth disrupting compacted aggregates caused by compression of dense machinery traffic, as pointed by Stumpf et al., 2016) [36]. Several works conducted for revegetation employed seeds of rough, aggressive U. decumbens and Melinis minutiflora for creation of a creeping layer, after soil correction and recover rapidly the desired areas. The seeds used to form the “cocktail of seeds” can be collected in adjacent areas of National Parks or used commercial ones. Sowing can be done manually, and the seeds can be disposed at a depth of 2 cm. Urochloa has been used especially in revegetation slopes of highways, as showed by Santos et al. [37].
4.6. Fertilization and Soil Amendments for Urochloa
4.6.1. Fertilization
Urochloa species can contain ~ 100 kg ha−1 of N, 130 kg ha−1 of K, 15 kg ha−1 of P, 40 kg ha−1 of Ca and 25 kg ha−1 of Mg, as pointed by Baptistella et al. [7]. Once correcting nutrient deficiencies, nitrogen availability controls plant production through assimilation of CO2 for growth. Among pastures of Urochloa brizantha cv. Marandu, maintained at a height of 25 cm, it was shown that weight gain per animal was similar between different doses of Nitrogen( 180 kg/ha of 4-14-8 fertilizer (urea, P2O5, and K2O) applied to grass, as reported by Barbero et al. [38]. Moreover, Kanno et al. [13]. added 54.4 mg/kg urea (24.5 mg/kg N), as a basic fertilizer in pot experiments seemed with Brachiaria brizantha. It is known that the substitution and removal of nutrients significantly affects the soil and plant conservation, and, thus, the conservation of pastures, therefore, soil fertilization is often carried out.
4.6.2. Soil Amendments
Regarding soil amendments, biochar-based fertilizers produced against poultry litter promoted 11% higher biomass production in U. brizantha cultivated pots under greenhouse conditions, as revealed by Lustosa Filho et al.[39]. Biochar is a promising material for improving plant and AMF performances, as it is insoluble in water and presents low short-term availability (pointed by those authors). However, it can increase over time due to the fertilizer dissolution when water distributes into the biochar pores, and at the same time, it can profit from discarded biomass promoting sustainable agriculture.
5. Genetics of Urochloa Species
Regarding the genetic structure of these plant species, there is great potential for more productive cultivars with high nutritional quality, resistance to pests and diseases, and tolerant to abiotic stresses more efficiently, and sustainably available in the future, as proposed by Ferreira et al. [40] However, the genetics and genomics of Urochloa pastures has been few investigated, existing a research gap, mainly due to their genomic complexity. [40], limited breeding programs, and few sequenced genomes of diploid Urochloa species currently available.
5.1. Inoculation of Urochloa Species
Rhizophagus (46%), Funneliformis (41%), Claroideoglomus (15%), Glomus (9%) and Gigaspora (2%) were frequently obtained in commercial inoculants. In this sense, Basiru and Hijri, [41] recently reviewed the application of commercial AMF inoculants and the ecological effects on the indigenous fungal communities under field conditions, highlighting the need to focus on selected species with a higher chance of success in the field, having minor side impacts. However, It is known that AMF commercial inoculants contain a few Glomeraceae species (Rhizophagus, Funneliformis, and Claroideoglomus) and that inoculants consisting of several AMF species can produce additive or synergistic effects on target crops. In the first studies, U. decumbens was inoculated with G. margarita and Glomus. clarum to study the recovering of degraded lands in greenhouse experiments, as showed by Santos et al.[42]. For mycorrhizal treatments, there are various possible procedures: 1) small holes can be prepared in the soil/ substrate surface of each pot, and the inoculum (~ 2.5 mL) placed in each hole: 2) Other procedure is to throw the spores into the roots of each plant( before transplanted) or pot. It is also more recently utilized to add (In the non-mycorrhizal treatments), 1,25 mL of the filtered leachate in each hole, to avoid the differences dues to other microorganisms(bacteria) living together with the fungal components; however it may cause contamination. Moreover, it was recently showed by Vather et al.[43]. that commercial AMF inoculants do not contain the species they declare. Thus, more research is needed on AMF inoculants at field conditions and their impacts on native communities, establishment in soil and roots, persistence, and influences on the native AMF community. In this sense, most studies indicate that introduced AMF can stay in the target field only a few months or some years decreasing abundance (60%) or with exclusion (30%). Further analysis shows that AMF inoculation exerts both positive and negative impacts on native AMF species, including suppression (33%), stimulation (38%), exclusion (19%), or neutral impacts (10% of examined cases). The factors influencing the ecology of AMF inoculants, such as the inherent properties of the inoculum, dosage, and frequency of inoculation, besides soil physical and biological factors, were discussed by Basiru and Hijri, 2022) [41]. In a different approach (Fertigation), Karti et al. [44] tested the inoculum of AMF acquired by hydroponic procedure, (spray, drip, and Nutrient Film in U. decumbens with different nutrients, as AMF promote adequate nutrition in soils.
6. Conclusions
This study has detailed the AM fungal community associated with Urochloa roots in a tropical grassland ecosystem using morphological methods for AM spore identification. Moreover, we compiled available reports on this plant species and its cultivation in tropical South America, expanding the topic with mention of major constrains and benefits of its cultivation and gaps to be solved in the future. Glomeraceae was the most prolific fungal family inhabiting the grassland; however, other AM fungal species were reported associated to this host along the period of more than twenty years. We confirmed four AMF species and four morphotypes associated to U. brizantha in a tropical managed soil This study contributes for the selection of highly-efficient inoculants for the mycorrhizal colonization of plant roots, especially, Urochloa species (U. brizantha), at field conditions. For instance, F. geosporus, D. heterogama and C. etunicatum can be indicated as potential inoculants for U. brizantha plots in this tropical region. Based on the findings of Vather et al.[43] who revealed few AMF species present in commercial inoculants., thus, the present study is valuable for grass cultivation, pointing out the need for well examining the AMF composition for a more precise inoculum available to the farmers and to the market, based on the evaluation of inoculum of AMF in Brachiaria decumbens [44]. As reviewed by Baptistella [7], it was highlighted that inoculation preferentially of Acaulospora species, favors the root growth, plant height, root and shoot dry matter of U. brizantha in tropical agroecosystems. However, in the present study, Claroideoglomus and Dentiscutata species are also indicated to be tested as inoculants. Thus, we recommend more detailed and long-term studies on the associated AMF species and their benefits for soil aggregation, as well as evaluation of glomalin content in more samples of Urochloa soils.
Declarations
Ethics approval and consent to participate: All authors approve and participate in the manuscript
Availability of data and material
Data available as requested.
Competing Interest
The authors declare that they have no conflict of interest.
Consent for publication
All authors approve the publication of the manuscript
Competing interests
No competing interests
Author Contributions
Conceptualization and writing, EJC MP, BY, NFD and A.A..; field and Lab work, MP.,EJC.
Funding
This research received no external funding.
Acknowledgements
We acknowledge the Institutions EPAMIG and IFMG, for samples and equipment provided, and the authors by their contributions.
References
- Lambers, H, Cong, W-F. 2022. Challenges Providing Multiple Ecosystem Benefits For Sustainable Managed Systems. Front. Agr. Sci. Eng., 9 (2) 170-176. [CrossRef]
- Pagano Kyriakides, M. , Kuyper, TW. 2023.Effects of Pesticides on the Arbuscular Mycorrhizal Symbiosis. Agrochemicals 2, 2. 10.3390/agrochemicals2020020.
- Goldan, E. ; Nedeff, V, Barsan, N.,Culea,M. Panainte-Lehadus, M, Mosnegutu,E.,Tomozei,C., Chitimus, Irimia, D. 2023. Assessment of Manure Compost Used as Soil Amendment—A Review. Processes, 11(4), 1167. [CrossRef]
- dos Santos Lucas, L. , Rubio Neto, M.A. de Moura, J.,B. Fernandes de Souza, R., Fernandes Santos, M E.; Fernandes de Moura, L, Gomes Xavier, E. J M dos Santos, J.M., Dutra Silva, R. N. S. 2022 Mycorrhizal fungi arbuscular in forage grasses cultivated in Cerrado soil.2022. Sci Reports, 12:3103. [CrossRef]
- Alves Teixeira, R. Gazel Soares, T. Rodrigues Fernandes, A. Martins de Souza Braz, A. 2014. Grasses and legumes as cover crop in no-tillage system in northeastern Pará, Brazil, Acta Amaz. 44 (4). [CrossRef]
- Baptistella, J.L.C.; Bettoni Teles, AP. Favarin J, Mazzafera, P. 2022 Phosphorus cycling by Urochloa decumbens intercropped with coffee. Experimental Agriculture, 58, 2022, e36.
- Baptistella, JLC; de Andrade SAL., Favarin,JL. Mazzafera,P..2020. Urochloa in Tropical Agroecosystems., Frontiers in Sustainable food systems, 4, 119.
- Barbosa, M. V. , Pedroso, D. de F.,F. Araujo Pinto, J. dos Santos V; Carbone Carneiro, M.A. 2019. Arbuscular mycorrhizal fungi and Urochloa brizantha: symbiosis and spore multiplication. Pesquisa Agropecuaria Tropical 49, 2019. [Google Scholar] [CrossRef]
- 9. Gerdemann JW, Nicolson TH (1963) Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans Br Mycol Soc, 46: 235–244.
- Varga, S. , Finozzi, C., Vestberg, M., K Arctic arbuscular mycorrhizal spore community and viability after storage in cold conditions. Mycorrhiza 25. 335–34.
- Stevenson, J. , Haberle, S., 2005. Macro Charcoal Analysis: a Modified Technique Used by the Department of Archaeology and Natural History. Palaeoworks Technical Papers, p. 5.108.
- Pagano, MC; Duarte, NF., Corrêa, EJA. 2020. Effect of crop and grassland management on mycorrhizal fungi and soil aggregation. Applied Soil Ecol, 147. 103385.
- Kanno, T. M. Saito M, Y. Ando, M.C.M. Macedo, Nakamura, T. Miranda C.H.B 2006. Importance of indigenous arbuscular mycorrhiza for growth and phosphorus uptake in tropical forage grasses growing on an acid, infertile soil from the Brazilian savannas. Tropical Grasslands, (2006. 40, 94–101.
- Pedroso, D., Barbosa, M.V., dos Santos, J.V. et al. (2018). Arbuscular Mycorrhizal Fungi favor the Initial Growth of Acacia mangium, Sorghum bicolor, and Urochloa brizantha in Soil Contaminated with Zn, Cu, Pb, and Cd. Bull Environ Contam Toxicol 101, 386–391. [CrossRef]
- Rillig 2004. Arbuscular mycorrhizae, glomalin, and soil aggregation. Canadian Journal of Soil Science. [CrossRef]
- Selvakumar, G. Shagol, C.C. Y. Kang, Y, Chung B.N., Han S.G., T.M. S. 2018. Arbuscular mycorrhizal fungi spore propagation using single spore as starter inoculum and a plant host. Journal of Applied Microbiology, 124, 6,2018, 1556–1565.
- Lehmann, A. , Leifheit, E.F. Rillig, M.C. 2017, Chapter 14–Mycorrhizas and Soil Aggregation. Mycorrhizal mediation of soil, In: Fertility, Structure, and Carbon Storage. pp. 241-262.
- Hossain, M.B. Contribution of Glomalin to Carbon Sequestration in Soil: A Review. Turkish Journal of Agriculture-Food Science and Technology, 9,1: 191-196, 2021.
- Wang, FEI Lu, X B Han, Z Ouyang, X Duan. Soil carbon sequestrations by nitrogen fertilizer application, straw return and no-tillage in China’s cropland. Global Change Biology, 2009 15, 2 p. 281-305.
- Pagano, MC, Correa, EJA, Lugo, MA, Duarte, NF.2022. Diversity and Benefits of Arbuscular Mycorrhizae in Restored Riparian Plantations. Diversity, 14(11), 938. [CrossRef]
- Follett, R.F. , Reed D.A. Soil Carbon Sequestration in Grazing Lands: Societal Benefits and Policy Implications. Rangeland Ecology & Management 63, 1, 2010, 4-15.
- Lal, 2008. Sequestration of atmospheric CO2 in global carbon pools Energy and environmental Science, 1, 86-100.
- Luo, Y. , ZHOU, X. Soil respiration and the environment. San Diego, Elsevier, 2006. 316p.
- ZhaoY., Yuqiang Tian, Y., Gao, Q., Xiaobing Li, X., Yong Zhang,, Ding, Y. Ouyang, S, Andrey Yurtaev, A., Kuzyakov, Y. Moderate grazing increases newly assimilated carbon allocation belowground. Rhizosphere, 22, 2022, 100547.
- Ontl, T. A. and Schulte, L. A. (2012) Soil Carbon Storage. Nature Education Knowledge, 3(10-35).
- Wilson, CH, Strickland, MS, Hutchings, JA. 2018. Grazing enhances belowground carbon allocation, microbial biomass, and soil carbon in a subtropical grassland, Global change Biol. 24(7):2997-3009.
- Qu T-b, Du W-c, Yuan X, Yang Z-m, Liu D-b, Wang D-l, et al. (2016) Impacts of Grazing Intensity and Plant Community Composition on Soil Bacterial Community Diversity in a Steppe Grassland. PLoS ONE 11(7): e0159680.
- Hafner, S. , Unteregelsbacher, S. Seeber, E.2012. Effect of grazing on carbon stocks and assimilate partitioning in a Tibetan montane pasture revealed by 13CO2 pulse labeling. Global Change Biology, 18, 2,528-538.
- Vandandorj, S.; Eldridge, D.J; Travers, S.; Oliver, I. Microsite and grazing intensity drive infiltration in a semi-arid woodland. Ecohydrology . 2016. [Google Scholar] [CrossRef]
- Feltran-Barbieri R, Féres JG. 2021. Degraded pastures in Brazil: improving livestock production and forest restoration. R. Soc.
- Open Sci. 8: 201854. [CrossRef]
- Tomaszewska, P. Vorontsova, M.S. Renvoize, S A. Ficinski, S.Z.Tohme, J. Schwarzacher, T. Castiblanco V., de Vega, J. J. Rowan A. C. Mitchell, Heslop-Harrison, J. S. Complex polyploid and hybrid species in an apomictic and sexual tropical forage grass group: genomic composition and evolution in Urochloa (Brachiaria) species. Annals of Botany, 131: 87–107, 2023.
- da Silva, LHX, · Okada, E.S.M, Soares, J.P. ·G. Oliveira, E.R. ·Gandra, J.R. · Costa Marques, O.F. · Neves, N. F., J.T.S. Gabriel, A.M A.. 2021. Organic management of Urochloa brizantha cv. Marandu intercropped with leguminous. Org. Agr. 11:539–552.
- Macedo, M.C.M. 1995. Pastagens no ecossistema Cerrados: Pesquisa para o desenvolvimento sustentavel. In: Simposio sobre pastagens nos ecossistemas Brasileiros: pesquisa para o desenvolvimento sustentavel. Anais. pp. 28–62. (Sociedade Brasileira de Zootecnia: Brasilia-DF, Brazil).
- Miguel, P. , Stumpf, L., Spinelli Pinto, L.F., Pauletto, E.A., Rodrigues, MF., Barboza,LS. Jéferson D., Leidemer, Duarte, B.T., M. A.B. Brito Pinto, B. G. Fernandez, M. Livia Oliveira Islabão, Silveira, LM., Peroba Rocha, J.V. Physical restoration of a minesoil after 10.6 years of revegetation. Soil and Tillage Research 227, 2023, 105599.
- Santos, A.L. De-Souza, F.A,. Berbara, R.L.L.; Guerra, J.G.M. 2000. Establishment and infective capacity of Gigaspora margarita Becker & Hall and Glomus clarum Nicol. & Gerd. in eroded soil. Acta bot. bras. 14(2): 127-139.
- Barbero, R.P., Ribeiro, A, C. , A. Moura , Zirondi V.L. , T. Almeida, T.F. Mattos A., MM. Barbero. 2021. Production potential of beef cattle in tropical pastures: a review. Animal Science. Ciênc. anim. bras. 22 . DOI: 10.1590/1809-6891v22e-69609.39. Lustosa Filho, J.F., Silva Carneiro, J.S. Barbosa, C.F, Pereira de Lima, K. , Amaral Leite, A. Azevedo Melo, LC. 2020. Aging of biochar-based fertilizers in soil: Effects on phosphorus pools and availability to Urochloa brizantha grass. Science of The Total Environment 709, 2020, 136028.
- Ferreira, R.C U., Moraes, ACL., Chiari, L., R. M. Simeão, Zanotto, BB. Vigna de Souza A,P. 2021. An Overview of the Genetics and Genomics of the Urochloa Species Most Commonly Used in Pastures Front. Plant Sci., 2021Sec. Plant Breeding 12–2021 |. [CrossRef]
- Basiru, Hijri, 2022. The Potential Applications of Commercial Arbuscular Mycorrhizal Fungal inoculants and Their Ecological Consequences. Microorganisms 10, 10 10.3390/microorganisms10101897.
- Vahter, T; Lillipuu, E.M. Oja, J. Öpik,M. Vasar, M. Hiiesalu, I. Do commercial arbuscular mycorrhizal inoculants contain the species that they claim? Mycorrhiza, 2023 33,3,:211-220. [CrossRef]
- Martins, C. R., Leite, L.L. Haridasan, M. 2001. The use of native grasses in the reclamation of an area degraded by gravel mining. R. Árvore, 25, 2, 157-|66.
- Karti, P D M H, Prihantoro I, Aryanto, A.T. Evaluation of inoculum arbuscular mycorrhizal fungi in Brachiaria decumbens. International e-Conference on Sustainable Agriculture and Farming System, IOP Conf. Series: Earth and Environmental Science 694 (2021) 012048, IOP Publishing. [CrossRef]
Figure 1.
grassland plot of Urochloa brizantha at the experimental farm.

Figure 2.
Soil elemental composition at different ecosystems in Pitangui experimental farm, grassland, cultivated site and adjacent forest.
Figure 2.
Soil elemental composition at different ecosystems in Pitangui experimental farm, grassland, cultivated site and adjacent forest.
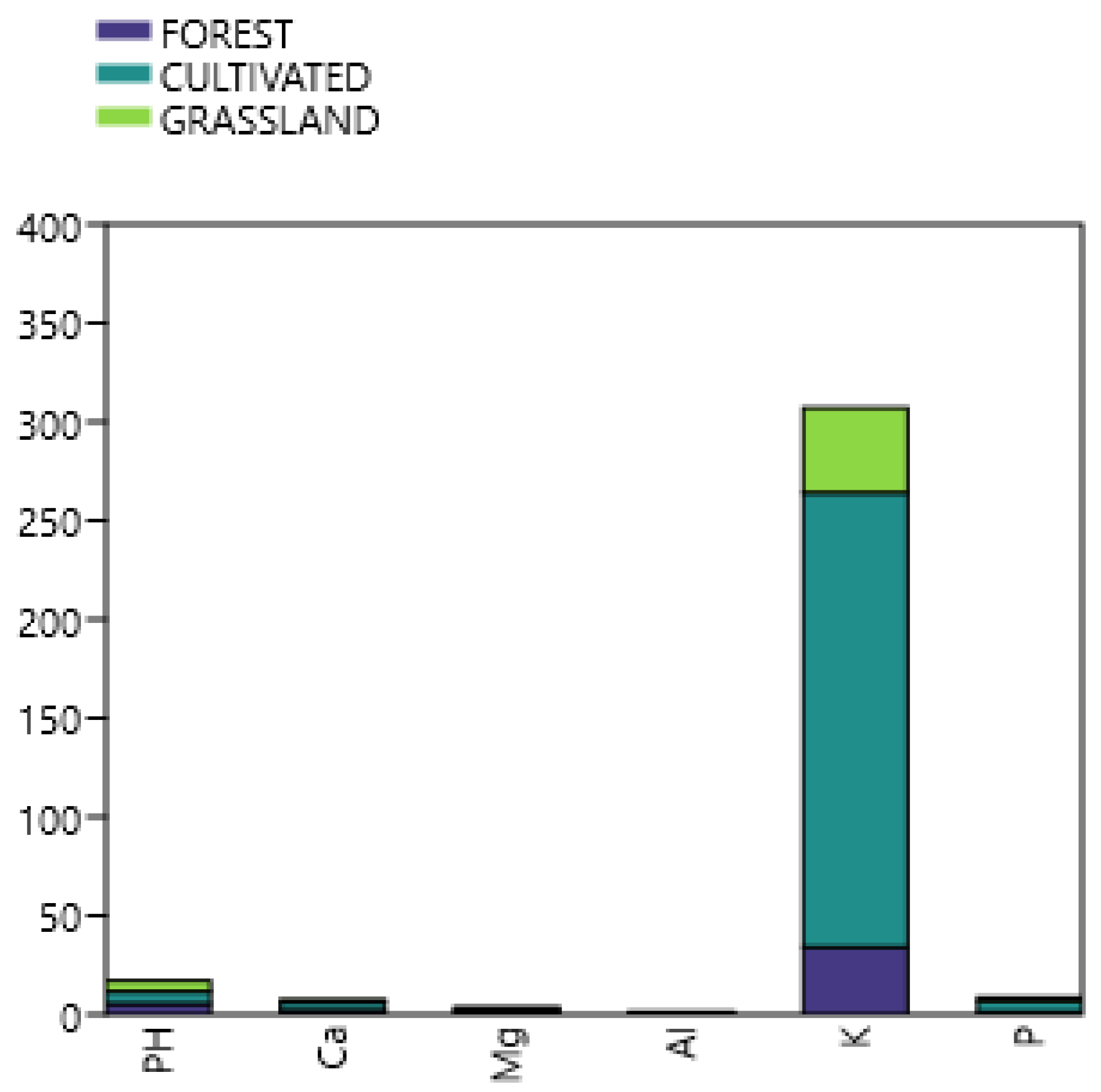
Figure 3.
U. brizantha grassland in Brazil, (A). Sorghum cultivated in pots (B) with U. brizantha soil for AMF multiplication.
Figure 3.
U. brizantha grassland in Brazil, (A). Sorghum cultivated in pots (B) with U. brizantha soil for AMF multiplication.

Figure 4.
AMF species found in the U. brizantha grassland. Claroideoglomus etunicatum, with hyaline subtending hyphae( sh) (A), Dentiscutata heterogama (B) with sporogenous cell (sc).
Figure 4.
AMF species found in the U. brizantha grassland. Claroideoglomus etunicatum, with hyaline subtending hyphae( sh) (A), Dentiscutata heterogama (B) with sporogenous cell (sc).
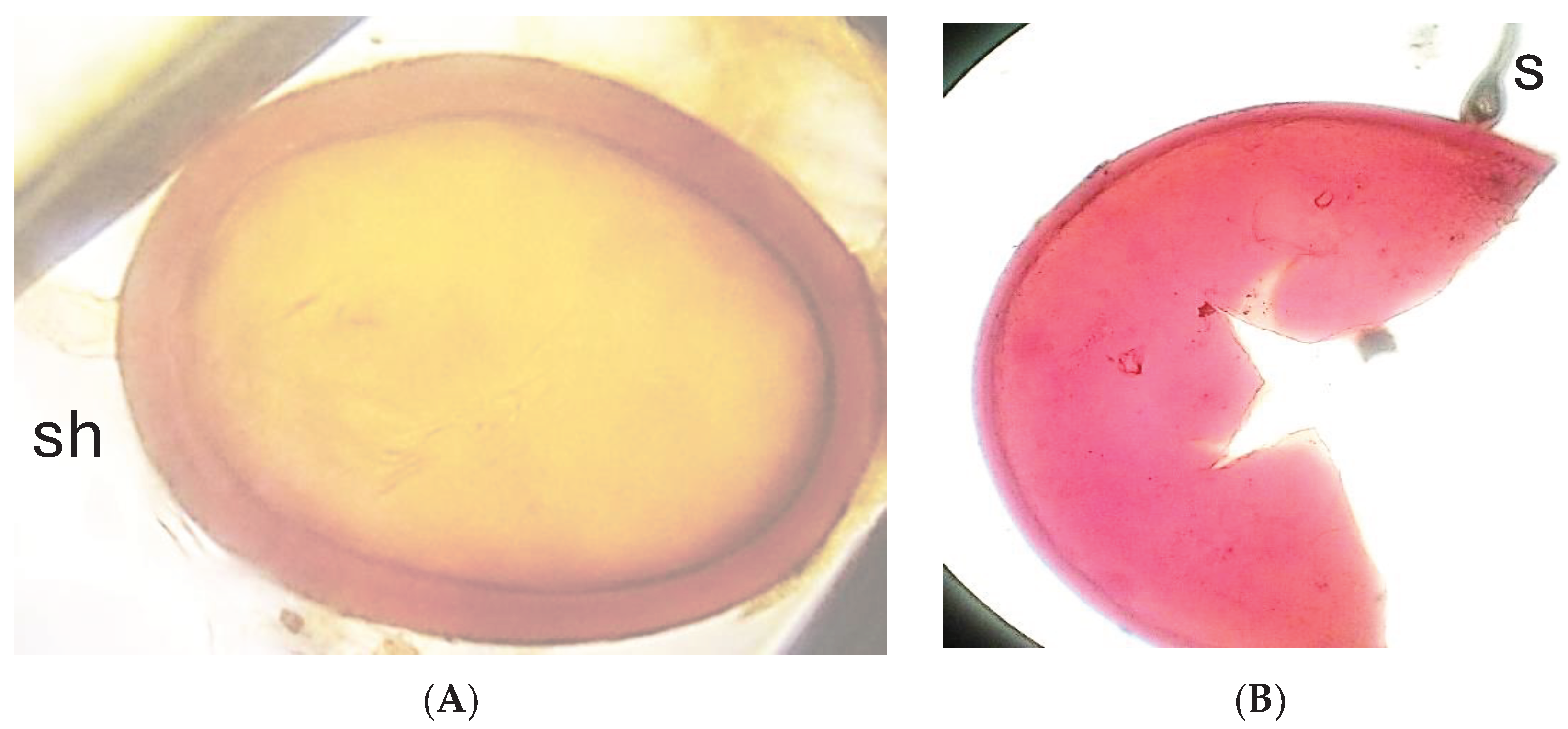

Table 1.
Reports on AMF in Urochloa species in Brazil.
| Vegetation / Field/ greenhouse | Plant species | Geographical coordinates | Altitude (m asl) | Estimated age of cultivation (year) | AMF species/Glomalin | State | Reference [8] |
|---|---|---|---|---|---|---|---|
| Field |
U. brizantha U. decumbens |
- - - |
Experimental Station of the Cerrado |
Diversispora sp., Scutellospora sp. Glomus sp. Gigaspora sp. | GO | [4] | |
| Greenhouse | U. brizantha | 19°42’51.5”S 44°53’37.0”W | 628,36 | >10 | A. colombiana, A. longula/ + | MG | [8] |
| U. brizantha | - | NA | SP | [7] * | |||
| U. brizantha | 48°26′W; 22°51′ S |
740 | NA | SP | [13] + |
NA: Not available. GO = Goiás; MG= Minas Gerais; SP = São Paulo state; + = Glomalin content evaluated (3.30 mg to 3.74g-1 soil).22 * = Review.
Table 2.
Physical-Chemical properties of soil at U. brizantha soil, in the experimental farm, Minas Gerais state, Brazil.
Table 2.
Physical-Chemical properties of soil at U. brizantha soil, in the experimental farm, Minas Gerais state, Brazil.
| pH (H2O) | 5.4 | ||
| OC (dag kg−1) | NA | ||
| Ca (cmolc kg−1) | 1.5 | ||
| Mg (cmolc kg−1) | 0.9 | ||
| Al (cmolc kg−1) | 0.7 | ||
| Available K mg⋅kg−1 | 43 | ||
| Available P mg⋅kg−1 | 1.8 | ||
| Soil texture | Clayey- loam | ||
| Charcoal content# | 19.66 |
NA: Not available. # Mean number of microcharcoal fragments in soil subsamples.
Table 3.
Chemical properties of soils† at different ecosystems in Pitangui experimental farm and adjacent forest, Minas Gerais state, Brazil.
Table 3.
Chemical properties of soils† at different ecosystems in Pitangui experimental farm and adjacent forest, Minas Gerais state, Brazil.
| Soil depth (0-20 cm) | |||||
| Forest | Cultivated | Grassland | |||
| pH (H2O) | 5.4ns | 6.2 | 5.4 | ||
| C (g kg-1) | - | - | - | ||
| Ca (cmolc kg-1) | 2b | 4.1a | 1.5 b | ||
| Mg (cmolc kg-1) | 0.8b | 2a | 0.9 b | ||
| Al (cmolc kg-1) | 0.5ns | 0.1 | 0.7 | ||
| K mg⋅kg-1 | 34b | 230a | 43b | ||
| P mg⋅kg-1 | 1.4c | 5.4a | 1.8 b | ||
Charcoal content 28.33 ab 8,33 bc 19.66 b three replicates plots (25 g soil). Tukey 5 % Values follow by the same lowercase letter in the same row do not differ ( p < 0.05), using Tukey test. ns= non-significant.
Table 4.
Distribution of AMF species associated with soil at Pitangui experimental farm, Minas Gerais state, Brazil.
Table 4.
Distribution of AMF species associated with soil at Pitangui experimental farm, Minas Gerais state, Brazil.
| AMF species | ||||||||||
| Dentiscutataceae | ||||||||||
| Dentiscutata heterogama | ||||||||||
| Claroideoglomeraceae | ||||||||||
| Claroideoglomus etunicatum | ||||||||||
| Racocetraceae | ||||||||||
| Racocetra fulgida | ||||||||||
| Glomeraceae | ||||||||||
| Funneliformes geosporus | ||||||||||
| Glomus sp. 1 | ||||||||||
| Glomus sp. 2 | ||||||||||
Data obtained at 0-20 cm soil depth. a (n = 3). .
Table 5.
Root colonization in inoculated Panicum and Urochloa species in Brazil (Previous studies). AMF inoculation (Indigenous AMF inoculum containing Gigaspora spp., Scutellospora spp., Glomus spp., Acaulospora spp. and Entrophospora spp.) Values are percentage of roots colonized by AMF for each grass species and soil pH treatment (High, medium, and low).
Table 5.
Root colonization in inoculated Panicum and Urochloa species in Brazil (Previous studies). AMF inoculation (Indigenous AMF inoculum containing Gigaspora spp., Scutellospora spp., Glomus spp., Acaulospora spp. and Entrophospora spp.) Values are percentage of roots colonized by AMF for each grass species and soil pH treatment (High, medium, and low).
| Type greenhouse |
Plant species | Soil Treatment (pH) | Non-inoculated | Inoculated | Reference | ||
|---|---|---|---|---|---|---|---|
| U. brizantha | - | 15.6 11.5 |
50.8 44.4 |
[8] [8] |
|||
| Field |
U. decumbens U. humidicola U. ruziziensis |
- | 78 5566 |
- | [4] [4] [4] |
||
| U. brizantha | - | 3 532 27 16 |
[4] [5] [5] [5] |
||||
|
U. decumbens U. humidicola P maximum |
10.4 - |
52.2 33.2 |
[13] [13] [13] |
||||
| Glasshouse | U. brizantha | High Medium Low |
D 49.7 00 31.4 | 54.3 49.7 31.4 |
[8] [8] [8] |
NA: Not available.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



